







Bahasa
Halaman
Hukum


Comparative Analysis of Alu Insertion Sequences in the APP 50 FlankingRegion in Humans and Other Primates
Jordi Clarimon, Aida M. Andres, Jaume Bertranpetit, David Comas
Unitat de Biologia Evolutiva, Facultat de Ciencies de la Salut i de la Vida, Universitat Pompeu Fabra, C/ Doctor Aiguader 80,
Barcelona, 08003, Catalonia, Spain
Received: 11 July 2003 / Accepted: 9 December 2003
Abstract. Overexpression of the amyloid precursorprotein gene (APP) may play a role in the neuropa-thology of Alzheimer’s disease. Therefore, elucidatingthe mechanisms involved in APP gene regulation is ofprimary importance, and various cis-acting regula-tory elements located in 50 distal regions are known toplay a main role. Some of them lie within Alu ele-ments, one of which (Alu1) is only found in humansand apes while the other (Alu2) has a much olderhistory and is also found in rhesus. These Alu inser-tions harbor sequence motifs that may act as cis-regulatory elements, which may cause differences inAPP regulation among primate species and whosefunctionality may be ascertained through theirconservation in a comparative analysis. We haveperformed a comparative analysis of the regioncomprising the two Alu elements of the APP pro-moter in several primates, including humans. Wehave found a significant decrease in nucleotide di-versity in the Alu2 element (inserted in all the speciesanalyzed) compared to the Alu1 (inserted only inapes). This finding can be interpreted as a constric-tion in the Alu2 sequence variation as a consequenceof a functional role of this element in the APP ex-pression. The present results suggest a wider exten-sion of the regulatory elements than the known shortconsensus regulatory sequences. Moreover, thedifferent conservation of two highly similar andneighboring sequences suggests that, besides the
importance of the sequence motifs, their position inrelation to the gene suggests that they have played arole in being recruited as regulatory elements.
Key words: Alu element — APP gene — Alzhei-mer’s disease — Promoter — Regulatory region
Introduction
One of the hallmarks of Alzheimer’s disease (AD) isthe deposition of the 39- to 43- amino acid residueamyloid b-peptide (Ab) as a major constituent ofextracellular plaques. Ab is derived from the pro-teolytic processing of the b-amyloid precursor protein(bAPP), which is encoded by the APP gene, whoseoverexpression has been related to the etiology ofAD. Due to the evidence that a 50-upstream regula-tory region of around 500 bp is sufficient for APPgene expression in cultured cells, most of the studieson APP regulation have been focused on the analysisof this proximal 50-flanking region (La Fauci et al.1989; Lahiri and Robakis 1991; Pollwein et al. 1992;Quitschke and Goldgaber 1992). However, there isalso evidence that gene transcription can be greatlyinfluenced by several positive and negative regulatoryelements located upstream from this proximal region(Lahiri et al. 2000).
The comparison of rhesus monkey and humanAPP promoters showed a region of �270 bp (posi-tions )2432 to )2161) of the human sequence that
Correspondence to: Dr. David Comas; email: david.comas@upf.
edu
J Mol Evol (2004) 58:722–731DOI: 10.1007/s00239-004-2594-y

was absent in the rhesus (Song and Lahiri 1998).Interestingly, the presence of this sequence was re-lated to a reduced transcription activity. This phe-nomenon was attributed to a consensus sequence forNFjB/Rel transcription factor located within thisregion, from position )2250 to )2241 (Grilli et al.1995). In humans, this 270-bp region containing theNFjB/Rel regulatory element corresponds to an Alusequence which, in turn, is inserted 130 bp upstreamfrom another Alu-type repetitive element that spansfrom position )2027 to position )1727.Alu elements belong to the class of short inter-
spersed repeats (SINEs) that have retrotransposedthroughout primate genomes over the past 65 millionyears of primate evolution (Batzer and Deininger2002). In the human genome, there are more than 1million copies of Alu elements, which represent al-most 11% of the genome (Li et al. 2001). Clusters ofAlu elements from different subfamilies have beendescribed at several loci as a result of retropositionalevents which occurred in the same chromosomallocation during different periods of primate evolu-tion (Rowold and Herrera 2000). Some of the Alucopies were inserted in the common branch that ledto many primate groups, whereas others have ret-roposed so recently that their insertion at a specificlocation within the human genome remains poly-morphic (Stoneking et al. 1997; Comas et al. 2001).In all cases, the ancestral state corresponds to thelack of the insertion and thus the insertion can bedated according to the specific sequence. These Aluelements have been assumed to lack function andwere included in the group of ‘‘junk DNA’’sequences. Nevertheless, Alu sequences interact withtheir surrounding genomic environment (Malakow-ski 2003), making them genomically useful. In thissense, Alu elements might acquire (exapt by Brosiusand Gould 1992) regulatory functions that may alterexpression or processing of nearby genes (Brosius2003).
There has long been suggestive evidence that Aluinsertion elements may play a role in gene transcrip-tion modulation by either increasing or decreasingthe expression of a nearby gene, as a consequence ofthe presence of several cis-acting elements within theAlu sequence (Hambor et al. 1993; Brini et al. 1993;McHaffie and Ralston 1995; Norris et al. 1995;Vansant and Reynolds 1995; Piedrafita et al. 1996;Fornasari et al. 1997; Apoil et al. 2000). Recently, ithas been shown (Lev-Maor et al. 2003) that Alu ele-ments can be inserted into mature messenger RNAsvia a splicing-mediated process that turns intronicAlus into new exons. Furthermore, mutations withinAlu elements have been related to disease. A goodexample is acute myelocytic leukemia, which is oftencaused by a single A/G base transition within ahormone response element that is, indeed, located
within an Alu element preceding the MPO gene(Austin et al. 1993).
The variation due to the presence/absence of this270-bp Alu-related sequence described in two primatespecies (humans and rhesus monkeys) in the APPpromoter region poses some functional and evolu-tionary questions regarding the possible influence ofthis element in APP expression. In this sense, acomparative approach may shed light on the func-tional architecture of the APP gene, since new geneticvariants that appeared in past generations have beenevaluated through time (favored or eliminated) in realin vivo experiments, that is, real individuals. Therecognition of new functional elements in the genomethrough a comparative analysis has recently begun tobe employed and is expected to be of primary value inthe full understanding of genome regulation (Boffelliet al. 2003). In order to better understand this regu-latory region we have performed a comparativestudy of the Alu elements residing in the 50-flankingregion of the APP gene by analyzing about a 1-kbsequence of the APP promoter in several primates,including humans. We infer the possible role of thisregion in APP gene expression regulation in an at-tempt to use a comparative approach to understandthe functional implications of the variation that hasremained after the numerous trials in evolution,leading, after a fine molecular tuning, to the presentdivergence in closely related species. At the very end acomparative approach may help to elucidate in finedetail functional constraints difficult to recognizeotherwise.
Materials and Methods
Genomic DNA was isolated from whole blood of different spe-
cies—human (Homo sapiens), chimpanzee (Pan troglodytes), gorilla
(Gorilla gorilla), orangutan (Pongo pygmaeus), and drill (Mandrillus
leucophaeus)—or bulb hairs from patas monkey (Erythrocebus pa-
tas), using a standard phenol–chloroform method, after digestion
with proteinase K.
The amplification of the APP gene promoter region containing
the Alu sequences was carried out by PCR using the primers ALU-
F (50-ACCAGGGAATGTGTCAGTGTTG-30) and ALU-R (50-CGGGTCTGCATGAGCAAATAAG-30). The whole region was
amplified using specific primers out of the Alu elements, and the
flanking regions of the Alu sequences are homologous in all pri-
mates analyzed, which discards the possibility of paralogous se-
quences. According to the numbering system for humans of
Salbaum et al. (1988), the 30 ends of these primers were located at
positions )2549 and )1616, respectively. The PCR was performed
using 100 ng of DNA in a final volume of 25 ll. The reaction
contained 20 pmol of each primer, 0.4 mM dNTPs, 1.5 mMMgCl2,
20 mM Tris–HCl (pH 8.4), 50 mMKCl, and 2.5 U of PLATINUM
Taq DNA Polymerase (GIBCO BRL). PCR amplification condi-
tions comprised an initial denaturation step of 1 min at 94�C;followed by 35 cycles of 35 s at 94�C, 30 s at 60�C, and 1 min at
72�C; and a final elongation step of 3 min at 72�C. In order to
visualize the results, PCR products were run in a 2% agarose gel.
PCR-amplified products were purified using Gene Clean (BIO
101). The purified fragments were sequenced separately on both
723

strands using the same PCR primers and the DNA Sequencing Kit,
Dye Terminator Cycle-Sequencing with Ampli Taq DNA Polym-
erase (Applied Biosystems), and the product was subsequently run
in an ABI PRISM 377 (Applied Biosystems) automatic sequencer.
Some of the sequenced fragments containing two Alu repeats
(�975 bp in total length) were difficult to read due to the long
poly(A) tails presented by the Alu insertions. Therefore, internal
primers APP1 (50-TGAGACGGAGTCTCGCTCTG-30) and APP2
(50-AAGATGGCAATGGCAGAGCG-30) (30 end positions )2401and )2063, respectively) were designed in order to overcome this
problem. In these cases, the purified 975-bp amplified products
were diluted 1:100 and used as a template for a nested PCR using
the primers ALU-F and the internal primer APP2 in a PCR reac-
tion performed under the same conditions as described above. The
446-bp PCR product was purified using Gene Clean (BIO 101) and
the sequence reaction was performed using the internal primer
APP1.
For the analyses, we used the DNA stretch from position )2520to position )1623 (according to Salbaum et al. 1988). One macaque
sequence (Macaca mulatta; GenBank number AF067971) was also
included in the analysis. Sequences were aligned using CLU-
STALW and data were analyzed by standard packages, such as
PHYLIP 3.5c (Felsenstein 1989), MEGA version 2.1 (Kumar et al.
2001), and DnaSP software (DNA sequence polymorphism version
3.50 [Rozas and Rozas 1999]).
Substitution rate (r) was estimated as r = K/2T (Kimura 1980),
were K is the number of substitutions per site since the time of
divergence and T is the estimated time of divergence between the
two species. Divergence times for apes and humans have been
taken from recent estimations in molecular anthropology (Chen
and Li 2001). K and its standard error were calculated according to
Kimura’s (1980) two-parameter model.
Results
The APP upstream promoter region from position)2520 to position )1623 in humans contains two Aluinserted elements, whereas in rhesus monkey only oneis present (Salbaum et al. 1988). The most upstreamAlu element (named Alu1), which is absent in rhesusmonkey, spans from position )2432 to position)2177 in humans, with direct flanking repeats 17 bpin length. The other Alu element (named Alu2), pre-sent in both species, starts at position )2027 and endsat position )1727, with direct flanking repeats of16 bp. PCR amplification and sequencing of the re-gion containing the putative Alu insertions wereperformed in several primates (Homo sapiens, Pantroglodytes, Gorilla gorilla, Pongo pygmaeus, Erythro-cebus patas, and Mandrillus leucophaeus). The Alu2element was present in all primate species analyzedwhereas Alu1 was only inserted in apes and humans.Nucleotide changes and indels compared to the hu-man GenBank sequence are shown in Fig. 1. Asshown in the figure, the GenBank sequence presentssome incongruence with the present data set, mainlyan excess of deletions, which can be attributed toerrors in the previous sequence.
According to diagnostic base substitutions (Batzerand Deininger 2002), the Alu1 element was assignedto the young (Y) Alu subfamily, while Alu2 was
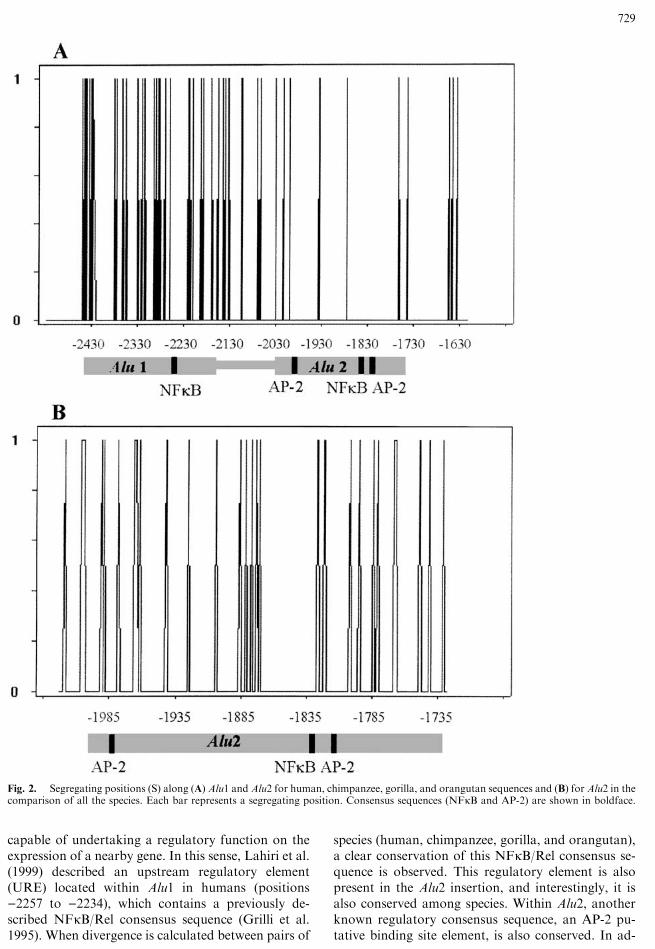
classified in the intermediate (Sx) subfamily. Numberof substitutions per site since time of divergence (K)was calculated for every pair of complete sequences(data not shown) and, also, for the Alu1, Alu2, andinter-Alu segments separately (values for human andapes are shown in Table 1) according to Kimura’s(1980) two-parameter model. As expected, humanspresented lower numbers of substitutions in relationto apes than to monkeys in all the comparisons per-formed, in agreement with multiple independentDNA sequence data sets (Ruvolo 1997).
In order to test any differences in evolutionary ratebetween both Alu elements, the number of nucleotidesubstitutions per site was compared. For all com-parisons, the Alu1 element presented higher K valuesthan Alu2 (Table 1) despite the more recent insertionof Alu1. However, these differences did not reachstatistical significance except for the comparisonsinvolving orangutans, which presented nonoverlap-ping confidence intervals, probably due to its longerperiod of divergence compared to the rest of apes.The divergence values found are similar to thosepresented in other 50-flanking regions between hu-mans and chimpanzees (Chen and Li 2001; Hellmannet al. 2003). Alu elements are the genomic regionswith higher divergence between humans and chim-panzees (Chen and Li 2001), with a mean K of 0.0200,which is similar to what it is found for the Alu1 ele-ment (0.0276) and higher than what it is found for theAlu2 element (0.0100), similar to other 50-UTRs(Hellmann et al. 2003), which agrees with the con-served pattern suggested in the present analysis.
Differences among the Alu elements and the inter-Alu region can also be analyzed by comparing theproportion of segregating sites among the three re-gions in human, chimpanzee, gorilla, and orangutan.Alu1 does not present any significant differences innumber of segregating sites compared to the inter-Aluregion (v2 = 0.58, p = 0.445). Nevertheless, Alu2presents a statistically significant lower number ofsegregating positions than Alu1 (v2 = 12.53, p <0.001) and the inter-Alu region (v2 = 6.04,p = 0.014). This lack of variation is indicative of aconstrain in the nucleotide diversity within Alu2,which may be attributed to some functional role ofthis Alu element.
In order to correct the number of substitutions forthe estimated age of divergence between each pair ofspecies, substitution rate values (r) were calculated(Table l). The Alu2 substitution rate is lower than therates presented by the Alu1 and the inter-Alu regions.The mean r values for each region analyzed point toa conserved substitution rate in Alu2 (0.54 · 10)9 ±0.06 · 10)9) compared to Alu1 (2.94 · 10)9 ± 0.54 ·10)9) and the inter-Alu (1.62 · 10)9 ± 0.34 · 10)9)regions. Again, Alu2 shows a clear evolutive slow-down in all lineages considered.
724

Because Alu elements contain a high presence ofCpG dinucleotides, which are known to mutate ap-proximately 10 times faster than any other dinucle-otide in the nuclear genome (Cooper and Krawczak1989; Martinez-Arias et al. 2001), substitution ratesexcluding CpG pairs were recalculated to avoid apossible mutational bias. The same general trend of
higher nucleotide substitution of Alu1 was also foundcompared to Alu2.
Consensus sequences for NFjB/Rel transcriptionfactor within each of these Alu elements have beendescribed (Grilli et al. 1995). One spans from posi-tion )2250 to position )2241 (within Alu1), and theother spans from )1837 to )1828 (within Alu2)
Fig. 1. Alignments of APP promoter sequences from all species analyzed together with the human GenBank sequence (accession numberX12751). Alu sequences are shown in boldface. The putative AP-2 and NFjB/Rel consensus sequences for control element binding sites areshown with line overhead.1 The consensus sequences for subfamilies Sx and Y (Batzer et al. 1996) are also shown.
725

(Fig. 1). These two sequences, which are specificallyrecognized by several transcription factors belongingto the NFjB/Rel family, are identical in all thespecies analyzed, suggesting a strong evolutionaryconservation. Furthermore, two AP-2 putativebinding sites have been described in the promoter ofthe rhesus monkey. These two sequences correspondto positions )1989 to )1981 and )1820 to )1814 inhumans, both within the Alu2 insertion (Song and
Lahiri 1998), and they are conserved among all thespecies analyzed except for a single substitution inthe upstream AP-2 element in the drill. Although itis tempting to interpret these results as the effect ofthe preservation of a functional activity, the shortlength of the elements and the relatively smalldivergence among the species analyzed could alsoexplain the conservation of these consensussequences.
Fig. 1. Continued.
726

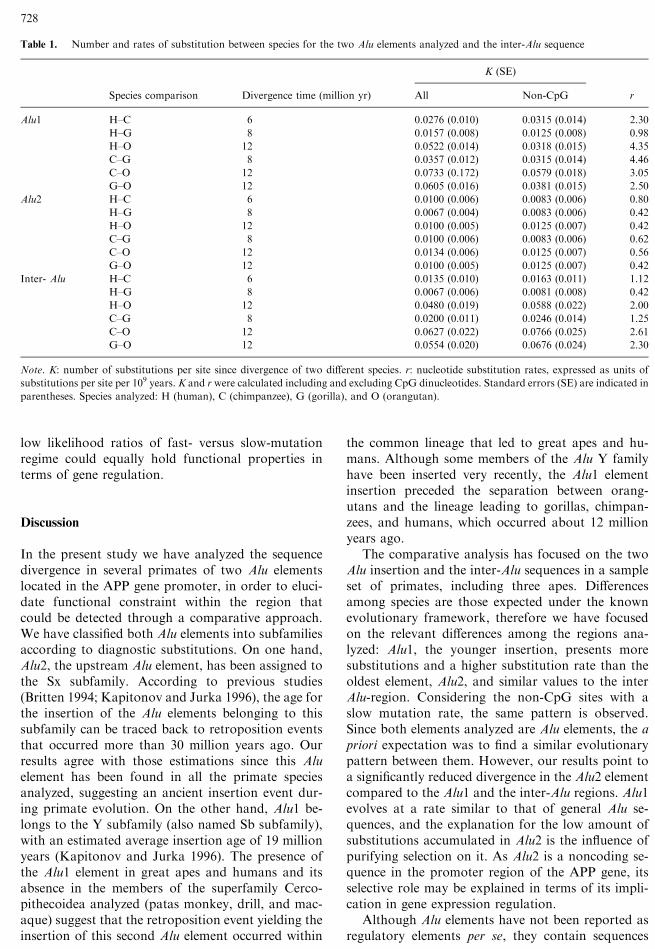
Segregating positions in humans, chimpanzees,gorillas, and orangutans along the entire sequence, aswell as for the Alu2 in the seven species analyzed, arerepresented in Fig. 2. The plot for both Alu elements(Fig. 2A) denotes a higher divergence along the Alu1element and the inter-Alu region compared to theAlu2.
The analysis of the Alu2 element in the seven pri-mate species (Fig. 2B) allows us to define the mostconserved regions within this element, since func-tional blocks are expected to be maintained even fordistant species. Thus, the addition of sequences formore divergent species highlights regions highlyconserved and thus with higher constriction levels. Asexpected, a general increase in segregating positions isdetected when monkey sequences are included in theplot (Fig. 2B). However, several blocks were totallyconserved in all species, including the NFjB/Rbinding site, the adjacent AP-2 consensus sequence,and a 32-bp region of upstream of the NFjB/Rbinding site (positions )1870 to )1838).
Recently, phylogenetic shadowing has been dem-onstrated to be a powerful tool for the identificationof primate-specific gene-regulatory elements from a
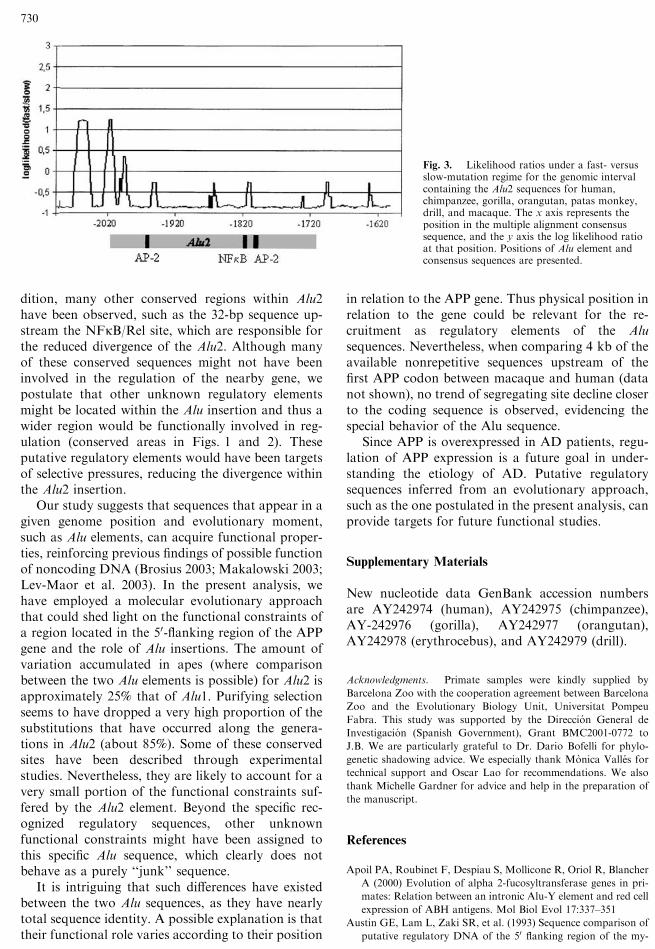
set of Old World monkey and hominoid sequences(Boffelli et al. 2003). In brief, this method determinesthe likelihood ratio under a fast- versus slow-muta-tion regime for each aligned nucleotide site across theDNA region, taking into account the phylogeneticrelationship between the species analyzed. We haveemployed this strategy to identify possible noncodingfunctional regions within the Alu2 segment. In theplot shown in Fig. 3, regions of varying conservationclearly appear. Several segments, varying in size be-tween 29 and 76 bp, show low levels of variation.Moreover, the NFjB/R and both AP-2 consensussequences have been totally fixed. Due to the fact thatthis method has a low sensitivity to sequences ofclosely related species, we have not included the Alu1element, which exists exclusively in apes, with only 12million years of divergence. Recently, similar resultswere found in the APOA promoter region (Boffelli etal. 2003) using the phylogenetic shadowing approach.In that case, the low variation accumulation ratewithin the APOA promoter was demonstrated to bethe result of functional constrains in regions involvedin transcriptional regulation activity. Therefore, itseems likely that the APP promoter regions with clear
Fig. 1. Continued.
727
low likelihood ratios of fast- versus slow-mutationregime could equally hold functional properties interms of gene regulation.
Discussion
In the present study we have analyzed the sequencedivergence in several primates of two Alu elementslocated in the APP gene promoter, in order to eluci-date functional constraint within the region thatcould be detected through a comparative approach.We have classified both Alu elements into subfamiliesaccording to diagnostic substitutions. On one hand,Alu2, the upstream Alu element, has been assigned tothe Sx subfamily. According to previous studies(Britten 1994; Kapitonov and Jurka 1996), the age forthe insertion of the Alu elements belonging to thissubfamily can be traced back to retroposition eventsthat occurred more than 30 million years ago. Ourresults agree with those estimations since this Aluelement has been found in all the primate speciesanalyzed, suggesting an ancient insertion event dur-ing primate evolution. On the other hand, Alu1 be-longs to the Y subfamily (also named Sb subfamily),with an estimated average insertion age of 19 millionyears (Kapitonov and Jurka 1996). The presence ofthe Alu1 element in great apes and humans and itsabsence in the members of the superfamily Cerco-pithecoidea analyzed (patas monkey, drill, and mac-aque) suggest that the retroposition event yielding theinsertion of this second Alu element occurred within
the common lineage that led to great apes and hu-mans. Although some members of the Alu Y familyhave been inserted very recently, the Alu1 elementinsertion preceded the separation between orang-utans and the lineage leading to gorillas, chimpan-zees, and humans, which occurred about 12 millionyears ago.
The comparative analysis has focused on the twoAlu insertion and the inter-Alu sequences in a sampleset of primates, including three apes. Differencesamong species are those expected under the knownevolutionary framework, therefore we have focusedon the relevant differences among the regions ana-lyzed: Alu1, the younger insertion, presents moresubstitutions and a higher substitution rate than theoldest element, Alu2, and similar values to the interAlu-region. Considering the non-CpG sites with aslow mutation rate, the same pattern is observed.Since both elements analyzed are Alu elements, the apriori expectation was to find a similar evolutionarypattern between them. However, our results point toa significantly reduced divergence in the Alu2 elementcompared to the Alu1 and the inter-Alu regions. Alu1evolves at a rate similar to that of general Alu se-quences, and the explanation for the low amount ofsubstitutions accumulated in Alu2 is the influence ofpurifying selection on it. As Alu2 is a noncoding se-quence in the promoter region of the APP gene, itsselective role may be explained in terms of its impli-cation in gene expression regulation.
Although Alu elements have not been reported asregulatory elements per se, they contain sequences
Table 1. Number and rates of substitution between species for the two Alu elements analyzed and the inter-Alu sequence
K (SE)
Species comparison Divergence time (million yr) All Non-CpG r
Alu1 H–C 6 0.0276 (0.010) 0.0315 (0.014) 2.30
H–G 8 0.0157 (0.008) 0.0125 (0.008) 0.98
H–O 12 0.0522 (0.014) 0.0318 (0.015) 4.35
C–G 8 0.0357 (0.012) 0.0315 (0.014) 4.46
C–O 12 0.0733 (0.172) 0.0579 (0.018) 3.05
G–O 12 0.0605 (0.016) 0.0381 (0.015) 2.50
Alu2 H–C 6 0.0100 (0.006) 0.0083 (0.006) 0.80
H–G 8 0.0067 (0.004) 0.0083 (0.006) 0.42
H–O 12 0.0100 (0.005) 0.0125 (0.007) 0.42
C–G 8 0.0100 (0.006) 0.0083 (0.006) 0.62
C–O 12 0.0134 (0.006) 0.0125 (0.007) 0.56
G–O 12 0.0100 (0.005) 0.0125 (0.007) 0.42
Inter- Alu H–C 6 0.0135 (0.010) 0.0163 (0.011) 1.12
H–G 8 0.0067 (0.006) 0.0081 (0.008) 0.42
H–O 12 0.0480 (0.019) 0.0588 (0.022) 2.00
C–G 8 0.0200 (0.011) 0.0246 (0.014) 1.25
C–O 12 0.0627 (0.022) 0.0766 (0.025) 2.61
G–O 12 0.0554 (0.020) 0.0676 (0.024) 2.30
Note. K: number of substitutions per site since divergence of two different species. r: nucleotide substitution rates, expressed as units of
substitutions per site per 109 years. K and r were calculated including and excluding CpG dinucleotides. Standard errors (SE) are indicated in
parentheses. Species analyzed: H (human), C (chimpanzee), G (gorilla), and O (orangutan).
728
capable of undertaking a regulatory function on theexpression of a nearby gene. In this sense, Lahiri et al.(1999) described an upstream regulatory element(URE) located within Alu1 in humans (positions)2257 to )2234), which contains a previously de-scribed NFjB/Rel consensus sequence (Grilli et al.1995). When divergence is calculated between pairs of
species (human, chimpanzee, gorilla, and orangutan),a clear conservation of this NFjB/Rel consensus se-quence is observed. This regulatory element is alsopresent in the Alu2 insertion, and interestingly, it isalso conserved among species. Within Alu2, anotherknown regulatory consensus sequence, an AP-2 pu-tative binding site element, is also conserved. In ad-
Fig. 2. Segregating positions (S) along (A) Alu1 and Alu2 for human, chimpanzee, gorilla, and orangutan sequences and (B) for Alu2 in thecomparison of all the species. Each bar represents a segregating position. Consensus sequences (NFjB and AP-2) are shown in boldface.
729
dition, many other conserved regions within Alu2have been observed, such as the 32-bp sequence up-stream the NFjB/Rel site, which are responsible forthe reduced divergence of the Alu2. Although manyof these conserved sequences might not have beeninvolved in the regulation of the nearby gene, wepostulate that other unknown regulatory elementsmight be located within the Alu insertion and thus awider region would be functionally involved in reg-ulation (conserved areas in Figs. 1 and 2). Theseputative regulatory elements would have been targetsof selective pressures, reducing the divergence withinthe Alu2 insertion.
Our study suggests that sequences that appear in agiven genome position and evolutionary moment,such as Alu elements, can acquire functional proper-ties, reinforcing previous findings of possible functionof noncoding DNA (Brosius 2003; Makalowski 2003;Lev-Maor et al. 2003). In the present analysis, wehave employed a molecular evolutionary approachthat could shed light on the functional constraints ofa region located in the 50-flanking region of the APPgene and the role of Alu insertions. The amount ofvariation accumulated in apes (where comparisonbetween the two Alu elements is possible) for Alu2 isapproximately 25% that of Alu1. Purifying selectionseems to have dropped a very high proportion of thesubstitutions that have occurred along the genera-tions in Alu2 (about 85%). Some of these conservedsites have been described through experimentalstudies. Nevertheless, they are likely to account for avery small portion of the functional constraints suf-fered by the Alu2 element. Beyond the specific rec-ognized regulatory sequences, other unknownfunctional constraints might have been assigned tothis specific Alu sequence, which clearly does notbehave as a purely ‘‘junk’’ sequence.
It is intriguing that such differences have existedbetween the two Alu sequences, as they have nearlytotal sequence identity. A possible explanation is thattheir functional role varies according to their position
in relation to the APP gene. Thus physical position inrelation to the gene could be relevant for the re-cruitment as regulatory elements of the Alusequences. Nevertheless, when comparing 4 kb of theavailable nonrepetitive sequences upstream of thefirst APP codon between macaque and human (datanot shown), no trend of segregating site decline closerto the coding sequence is observed, evidencing thespecial behavior of the Alu sequence.
Since APP is overexpressed in AD patients, regu-lation of APP expression is a future goal in under-standing the etiology of AD. Putative regulatorysequences inferred from an evolutionary approach,such as the one postulated in the present analysis, canprovide targets for future functional studies.
Supplementary Materials
New nucleotide data GenBank accession numbersare AY242974 (human), AY242975 (chimpanzee),AY-242976 (gorilla), AY242977 (orangutan),AY242978 (erythrocebus), and AY242979 (drill).
Acknowledgments. Primate samples were kindly supplied by
Barcelona Zoo with the cooperation agreement between Barcelona
Zoo and the Evolutionary Biology Unit, Universitat Pompeu
Fabra. This study was supported by the Direccion General de
Investigacion (Spanish Government), Grant BMC2001-0772 to
J.B. We are particularly grateful to Dr. Dario Bofelli for phylo-
genetic shadowing advice. We especially thank Monica Valles for
technical support and Oscar Lao for recommendations. We also
thank Michelle Gardner for advice and help in the preparation of
the manuscript.
References
Apoil PA, Roubinet F, Despiau S, Mollicone R, Oriol R, Blancher
A (2000) Evolution of alpha 2-fucosyltransferase genes in pri-
mates: Relation between an intronic Alu-Y element and red cell
expression of ABH antigens. Mol Biol Evol 17:337–351
Austin GE, Lam L, Zaki SR, et al. (1993) Sequence comparison of
putative regulatory DNA of the 50 flanking region of the my-
Fig. 3. Likelihood ratios under a fast- versusslow-mutation regime for the genomic intervalcontaining the Alu2 sequences for human,chimpanzee, gorilla, orangutan, patas monkey,drill, and macaque. The x axis represents theposition in the multiple alignment consensussequence, and the y axis the log likelihood ratioat that position. Positions of Alu element andconsensus sequences are presented.
730

eloperoxidase gene in normal and leukemic bone marrow cells.
Leukemia 7:1445–1450
Batzer MA, Deininger PL (2002) Alu repeats and human genomic
diversity. Nat Rev Genet 3:370–379
Batzer MA, Deininger PL, Hellmann-Blumberg U, Jurka J,
Labuda D, Rubin CM, Schmid CW, Zietkiewicz E, Zuckerk-
andl E (1996) Standardized nomenclature for Alu repeats.
J Mol Evol 42:3–6
Boffelli D, McAuliffe J, Ovcharenko D, Lewis KD, Ovcharenko I,
Pachter L, Rubin EM (2003) Phylogenetic shadowing of pri-
mate sequences to find functional regions of the human genome.
Science 299:1391–1394
Brini AT, Lee GM, Kinet JP (1993) Involvement of Alu sequences
in the cell-specific regulation of transcription of the gamma
chain of Fc and T cell receptors. J Biol Chem 268:1335–1361
Britten RJ (1994) Evidence that most human Alu sequences were
inserted in a process that ceased about 30 million years ago.
Proc Natl Acad Sci USA 91:6148–6150
Brosius J (2003) How significant is 98.5% ‘junk’ in mammalian
genomes? Bioinformatics 19:ii35
Brosius J, Gould SJ (1992) On ‘‘nomenclature’’: A comprehensive
(and respectful) taxonomy for pseudogenes and other ‘‘junk
DNA.’’ Proc Natl Acad Sci USA 89:10706–10710
Chen FC, Li WH (2001) Genomic divergences between humans
and other hominoids and the effective population size of the
common ancestor of humans and chimpanzees. Am J Hum
Genet 68:444–456
Comas D, Plaza S, Calafell F, Sajantila A, Bertranpetit J (2001)
Recent insertion of an Alu element within a polymorphic hu-
man-specific Alu insertion. Mol Biol Evol 18:85–88
Cooper DN, Krawczak M (1989) Cytosine methylation and the
fate of CpG dinucleotides in vertebrate genomes. Hum Genet
83:181–188
Felsenstein J (1989) PHYLIP Phylogeny inference package (version
3.2). Cladistic 5:164–l66
Fornasari D, Battaglioli E, Flora A, Terzano S, Clementi F (1997)
Structural and functional characterization of the human alpha3
nicotinic subunit gene promoter. Mol Pharmacol 51:250–261
Grilli M, Ribola M, Alberici A, Valerio A, Memo M, Spano P
(1995) Identification and characterization of a kappa B/Rel
binding site in the regulatory region of the amyloid precursor
protein gene. J Biol Chem 270:26774–26777
Hambor JE, Mennone J, Coon ME, Hanke JH, Kavathas P (1993)
Identification and characterization of an Alu-containing, T-cell-
specific enhancer located in the last intron of the human CD8
alpha gene. Mol Cell Biol 13:7056–7070
Kapitonov V, Jurka J (1996) The age of Alu subfamilies. J Mol
Evol 42:59–65
Kimura M (1980) A simple method for estimating evolutionary
rates of base substitutions through comparative studies of nu-
cleotide sequences. J Mol Evol 16:111–120
Kumar S, Tamura K, Jakobsen I, Nei M (2001) MEGA2: Mo-
lecular Evolutionary Genetics Analysis software. Arizona State
University, Tempe
La Fauci G, Lahiri DK, Salton SR, Robakis NK (1989) Charac-
terization of the 50-end region and the first two exons of the
beta-protein precursor gene. Biochem Biophys Res Commun
159:297–304
Lahiri DK, Robakis NK (1991) The promoter activity of the gene
encoding Alzheimer beta-amyloid precursor protein (APP) is
regulated by two blocks of upstream sequences. Brain Res Mol
Brain Res 9:253–257
Lahiri DK, Nail C, Ge YW (1999) Promoter activity of the beta-
amyloid precursor protein gene is negatively modulated by an
upstream regulatory element. Brain Res Mol Brain Res 71:
32–41
Lahiri DK, Song W, Ge YW (2000) Analysis of the 50-flankingregion of the beta-amyloid precursor protein gene that con-
tributes to increased promoter activity in differentiated neur-
onal cells. Brain Res Mol Brain Res 77:185–198
Lev-Maor G, Sorek R, Shomron N, Ast G (2003) The birth of an
alternatively spliced exon: 30 splice-site selection in Alu exons.
Science 300:1288–1291
Li WH, Gu Z, Wang H, Nekrutenko A (2001) Evolutionary
analyses of the human genome. Nature 409:847–849
Makalowski W (2003) Not junk after all. Science 300:1246–1247
Martinez-Arias R, Calafell F, Mateu E, Comas D, Andres A,
Bertranpetit J (2001) Sequence variability of a human pseudo-
gene. Genome Res 11:1071–1085
McHaffie GS, Ralston SH (1995) Origin of a negative calcium re-
sponse element in an ALU-repeat: Implications for regulation
of gene expression by extracellular calcium. Bone 17:11–14
Norris J, Fan D, Aleman C, Marks JR, Futreal PA, Wiseman RW,
Iglehart JD, Deininger PL, McDonnell DP (1995) Identification
of a new subclass of Alu DNA repeats which can function as
estrogen receptor-dependent transcriptional enhancers. J Biol
Chem 270:22777–22782
Piedrafita FJ, Molander RB, Vansant G, Orlova EA, Pfahl M,
Reynolds WF (1996) An Alu element in the myeloperoxidase
promoter contains a composite SP1-thyroid hormone-retinoic
acid response element. J Biol Chem 271:14412–14420
Pollwein P, Masters CL, Beyreuther K (1992) The expression of the
amyloid precursor protein (APP) is regulated by two GC-ele-
ments in the promoter. Nucleic Acids Res 20:63–68
Quitschke WW, Goldgaber D (1992) The amyloid beta-protein
precursor promoter. A region essential for transcriptional ac-
tivity contains a nuclear factor binding domain. J Biol Chem
267:17362–17368
Rowold DJ, Herrera RJ (2000) Alu elements and the human ge-
nome. Genetica 108:57–72
Rozas J, Rozas R (1999) DnaSP version 3: An integrated program
for molecular population genetics and molecular evolution
analysis. Bioinformatics 15:174–175
Ruvolo M (1997) Molecular phylogeny of the hominoids: Infer-
ences from multiple independent DNA sequence data sets. Mol
Biol Evol 14:248–265
Salbaum JM, Weidemann A, Lemaire HG, Masters CL, Beyreu-
ther K (1988) The promoter of Alzheimer’s disease amyloid A4
precursor gene. EMBO J 7:2807–2813
Song W, Lahiri DK (1998) Molecular cloning of the promoter of
the gene encoding the Rhesus monkey beta-amyloid precursor
protein: Structural characterization and a comparative study
with other species. Gene 217:151–164
Stoneking M, Fontius JJ, Clifford SL, Soodyall H, Arcot SS, Saha
N, Jenkins T, Tahir MA, Deininger PL, Batzer MA (1997) Alu
insertion polymorphisms and human evolution: evidence for a
larger population size in Africa. Genome Res 7:1061–1071
Vansant G, Reynolds WF (1995) The consensus sequence of a
major Alu subfamily contains a functional retinoic acid re-
sponse element. Proc Natl Acad Sci USA 92:8229–8233
731
Top Related
Copyright © 2022 FDOKUMEN

