
Computation in spinal circuitry: Lessons from behaving primates
10
Behavioural Brain Research 194 (2008) 119–128 Contents lists available at ScienceDirect Behavioural Brain Research journal homepage: www.elsevier.com/locate/bbr Review Computation in spinal circuitry:Lessons from behaving primates Ran Harel a,b,c , Itay Asher a,b , Oren Cohen a,b , Zvi Israel d , Uri Shalit a,b , Yuval Yanai a,b , Nofya Zinger a,b , Yifat Prut a,b,∗ a The Hebrew University, Hadassah Medical School, P.O. Box 12272, Jerusalem 91120, Israel b The Interdisciplinary Center for Neural Computation, Israel c Department of Neurosurgery, Sheba Medical Center, Tel Aviv, Israel d Department of Neurosurgery, Hadassah Hospital, Jerusalem, Israel article info Article history: Received 29 May 2008 Accepted 8 July 2008 Available online 18 July 2008 Keywords: Motor cortex Spinal cord Corticospinal connectivity Motor control abstract Performing voluntary motor actions requires the translation of motor commands into a specific set of mus- cle activation. While it is assumed that this process is carried out via cooperative interactions between supraspinal and spinal neurons, the unique contribution of each of these areas to the process is still unknown. Many studies have focused on the neuronal representation of the motor command, mostly in the motor cortex. Nonetheless, to execute these commands there must be a mechanism that can translate this representation into a sustained drive to the spinal motoneurons (MNs). Here we review different candidate mechanisms for activating MNs and their possible role in voluntary movements. We discuss recent studies which directly estimate the contribution of segmental INs to the transmis- sion of cortical command to MNs, both in terms of functional connectivity and as a computational link. Finally, we suggest a conceptual framework in which the cortical motor command is processed simultaneously via MNs and INs. In this model, the motor cortex provides a transient signal which is important for initiating new patterns of recruited muscles, whereas the INs translate this command into a sustained, amplified and muscle-based signal which is necessary to maintain ongoing muscle activity. © 2008 Elsevier B.V. All rights reserved. Contents 1. Introduction ......................................................................................................................................... 120 1.1. Direct and indirect converging pathways onto forearm motoneurons ...................................................................... 120 1.2. What drives motoneurons to fire? ........................................................................................................... 120 1.3. Is IN relay of cortical commands to MNs predominantly inhibitory? ...................................................................... 120 1.4. Neuromodulatary effect on MNs ............................................................................................................ 122 2. Methodology: recording cortical and spinal activity in behaving primates ........................................................................ 122 3. Computational role of INs ........................................................................................................................... 123 3.1. Cortical input to INs is strong and has clear functional correlates .......................................................................... 123 3.2. Firing properties of INs ...................................................................................................................... 124 3.3. Dual role of cortical command on spinal flow of information .............................................................................. 124 3.4. Mixed representation of space and muscles exhibited by M1 neurons .................................................................... 124 4. The emerging picture about the role of spinal interneurons during voluntary movements: concluding remarks ................................ 127 Acknowledgments .................................................................................................................................. 127 References ........................................................................................................................................... 127 ∗ Corresponding author at: Department of Physiology, The Hebrew University- Hadassah Medical School, P.O. Box 12272, Jerusalem 91120, Israel. Tel.: +972 2 6757912; fax: +972 2 6439736. E-mail address: [email protected] (Y. Prut). 0166-4328/$ – see front matter © 2008 Elsevier B.V. All rights reserved. doi:10.1016/j.bbr.2008.07.013
Transcript of Computation in spinal circuitry: Lessons from behaving primates

Behavioural Brain Research 194 (2008) 119–128
Contents lists available at ScienceDirect
Behavioural Brain Research
journa l homepage: www.e lsev ier .com/ locate /bbr
Review
Computation in spinal circuitry:Lessons from behaving primates
Ran Harela,b,c, Itay Ashera,b, Oren Cohena,b, Zvi Israeld, Uri Shalit a,b, Yuval Yanaia,b,Nofya Zingera,b, Yifat Pruta,b,∗
a The Hebrew University, Hadassah Medical School, P.O. Box 12272, Jerusalem 91120, Israelb The Interdisciplinary Center for Neural Computation, Israelc Department of Neurosurgery, Sheba Medical Center, Tel Aviv, Israeld Department of Neurosurgery, Hadassah Hospital, Jerusalem, Israel
a r t i c l e i n f o
Article history:Received 29 May 2008Accepted 8 July 2008Available online 18 July 2008
Keywords:Motor cortexSpinal cordCorticospinal connectivityMotor control
a b s t r a c t
Performing voluntary motor actions requires the translation of motor commands into a specific set of mus-cle activation. While it is assumed that this process is carried out via cooperative interactions betweensupraspinal and spinal neurons, the unique contribution of each of these areas to the process is stillunknown. Many studies have focused on the neuronal representation of the motor command, mostlyin the motor cortex. Nonetheless, to execute these commands there must be a mechanism that cantranslate this representation into a sustained drive to the spinal motoneurons (MNs). Here we reviewdifferent candidate mechanisms for activating MNs and their possible role in voluntary movements.We discuss recent studies which directly estimate the contribution of segmental INs to the transmis-sion of cortical command to MNs, both in terms of functional connectivity and as a computationallink. Finally, we suggest a conceptual framework in which the cortical motor command is processedsimultaneously via MNs and INs. In this model, the motor cortex provides a transient signal which is
important for initiating new patterns of recruited muscles, whereas the INs translate this commandinto a sustained, amplified and muscle-based signal which is necessary to maintain ongoing muscleC
H6
0d
activity.© 2008 Elsevier B.V. All rights reserved.
ontents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1201.1. Direct and indirect converging pathways onto forearm motoneurons . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1201.2. What drives motoneurons to fire?. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1201.3. Is IN relay of cortical commands to MNs predominantly inhibitory? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1201.4. Neuromodulatary effect on MNs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 122
2. Methodology: recording cortical and spinal activity in behaving primates . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1223. Computational role of INs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 123
3.1. Cortical input to INs is strong and has clear functional correlates . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1233.2. Firing properties of INs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1243.3. Dual role of cortical command on spinal flow of information . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 124
3.4. Mixed representation of space and muscles exhibited by M1 neur4. The emerging picture about the role of spinal interneurons during volunAcknowledgments . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
∗ Corresponding author at: Department of Physiology, The Hebrew University-adassah Medical School, P.O. Box 12272, Jerusalem 91120, Israel. Tel.: +972 2757912; fax: +972 2 6439736.
E-mail address: [email protected] (Y. Prut).
166-4328/$ – see front matter © 2008 Elsevier B.V. All rights reserved.oi:10.1016/j.bbr.2008.07.013
ons . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 124tary movements: concluding remarks . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 127. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 127. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 127

1 rain R
1
1m
ednwieatcisecvmiwatfisWamiwa
1
adhorpsnfis
slatmwiclctmeeosi
h[bpoewdsplratmp
1i
aaeMbsfapi
mIOiatbbtiAi[p
sfoSmitet[tbd
20 R. Harel et al. / Behavioural B
. Introduction
.1. Direct and indirect converging pathways onto forearmotoneurons
The way motor commands recruit muscles during control andxecution of voluntary movements is only partially understoodespite the extensive research devoted to this issue. In human andon-human primates, most studies have focused on the direct path-ay connecting the motor cortex with muscles [1]. The interest
n the direct route is motivated by the correlation between themergence of this pathway (appearing first in primates) and thebility to perform independent finger movements [2,3]. However,his focus may also reflect paradigmatic limitations. Motor corticalircuitry is more readily accessible to studies of neuronal activityn behaving animals. Recent attempts have shown that recordingpinal neuronal activity in behaving primates is feasible [4–8]. Thisxperimental tool allows researchers for the first time to directlyompare the direct (corticomotoneuornal) and indirect (relayedia spinal neurons) impact of the motor cortex on muscles duringotor behavior of primates. Furthermore, recording spinal activ-
ty provides a window to the way a motor command is processedhile it is being transmitted from the motor cortex to muscles. The
im of this review is to compare existing evidence on the contribu-ion of these two pathways to voluntary forearm movements. Werst review existing views about the way motor commands provideufficient and informative drive to relevant motoneurons (MNs).
e then describe some recent findings we obtained on the activitynd interneuronal interactions of spinal interneurons (INs) duringovement. Finally, we will suggest a possible role of IN circuitry
n planning and executing voluntary movement and the manner inhich MNs may integrate these two routes to yield coherent muscle
ctivation.
.2. What drives motoneurons to fire?
Motoneurons are distinct neurons in the nervous system. Theyre large cells with a widespread dendritic field that can extendeep into the intermediate zone and even into the base of the dorsalorn [9,10]. Each MN integrates multiple synaptic inputs (50–150 K)f which about 40% are excitatory and 60% inhibitory [11]. Thisatio may change in different compartments of the MN. For exam-le, the majority of somatic synapses are inhibitory, similar to theynaptic organization of cortical pyramidal neurons [11]. The largeumber of inhibitory synapses and their location is puzzling sincering these cells, given their size, may require (in the passive case)ubstantial driving current.
MNs integrate multiple synaptic inputs arising from diverseources, which may roughly be segregated based on their origin intoocal (segmental) and remote (descending pathways and peripheralfferents). Additionally, MNs are under powerful neuromodula-ory control which is mediated by serotonin and adrenalin. These
etabotropic substances can act via synaptic contacts or directly,ith no junctional specialization [11,12]. The relative proportion of
nputs provided by each of these sources is difficult to estimate, andan potentially vary in different animals and at different segmentalevels of the same animal (i.e., hind limb versus forelimb). Whenonsidering the forearm of primates, special attention is often paido the corticomotoneuronal (CM) system which transmits infor-
ation directly from motor cortex to MNs. While this system is
xpanded in primates compared to animal located lower on thevolutionary tree, a rough estimate suggests that no more than 10%f the synaptic contacts to MNs are CM in origin [13]. The number ofynaptic contacts made by each CM fiber with targeted MN is typ-cally small [14]. The strength of CM synapses is not exceedinglydtfdm
esearch 194 (2008) 119–128
igh compared to the synapses formed by cortical neurons on INs15,16] and compared to the synapses made by INs on MNs (this cane deduced by comparing functional studies in the spinal cord ofrimates [8] with data available from cortical studies [17]). Tracingf single corticospinal fibers [9,14] suggests that CM fibers are notxclusively dedicated to activating MNs; rather, they often synapseith segmental INs as well. The gross anatomy of corticospinalescending pathways [18] further reveals that most fibers that con-titute this pathway terminate in the intermediate zone which isredominantly populated by spinal INs and not MNs. These findings
ead to the conclusion that most of the direct cortical input to MNs iselayed via INs (it is still difficult to distinguish between segmentalnd propriospinal input to MNs). This may mean that INs can poten-ially participate in the process of translating motor commands into
uscle-based signals, yet the nature of their contribution to thisrocess is not fully understood.
.3. Is IN relay of cortical commands to MNs predominantlynhibitory?
A large body of work has been devoted to the question of thedded value of CM connectivity compared to the indirect route (forreview see Ref. [19]). Clearly the two systems need to act syn-
rgistically for movements to occur but their exact role is unclear.uch of this debate has revolved around the existence and contri-
ution of the higher cervical (C3–C4) propriospinal interneuronalystem to voluntary forearm movements in primates [20]. Here weocus on the role of segmental INs in voluntary movements, as theyre the main (though certainly not exclusive) spinal target of therimary motor cortex [21,22] and probably provide the bulk of IN
nput to MNs of the forearm.Spinal INs circuitry is diverse [23] and it is difficult to esti-
ate the actual way it integrates descending inputs to affect MNs.ndirect measures provide conflicting evidence about this process.n one hand, stimulating the pyramidal tract yielded primarily
nhibitory di-synaptic post-synaptic potentials in forearm MNs ofnaesthetized macaques [24]. As such, INs are more likely to shapehe descending excitatory volley than serve as a processing linketween motor cortex and muscles (though this assumption maye somewhat naive). One can explain such a result by assuminghat CS fibers terminate preferentially on inhibitory INs or thatnhibitory INs constitute the vast majority of segmental circuitry.lternatively, as di-synaptic excitatory potentials were discovered
n a similar experiment when glycenrgic inhibition was blocked25,26] local inhibition may mask the excitatory di-synaptic com-onent either generally or when using anesthesia.
On the other hand, studying the functional interactions betweenpinal INs and muscles in behaving primates [7,8] has revealed a dif-erent picture. When averaging muscle activity around spike timesf spinal interneurons (i.e., computing spike-triggered averages,TAs) most of the post-spike effects at latency consistent with aonosynaptic connection were excitatory. A first concern was that
nhibitory INs were active in a reciprocal manner to the affectedarget, so that STAs could not reveal their inhibitory impact. How-ver, characterizing the response pattern of INs further revealedhat only a minority of INs have a reciprocal firing pattern (see Ref.8] and Fig. 2 here). Most INs exhibited background firing when noorque was applied, and then increased their rate in either one oroth directions. This response pattern was found consistently inifferent types of wrist tasks. The apparent conflict between the
educed synaptic organization of spinal INs (based on the distribu-ion of synaptic inputs on MNs) and their functional linkage withorearm muscles may stem from the fact that these studies involveifferent experimental preparations (anaesthetized versus awakeonkeys). Moreover, the functional connectivity observed in the
R. Harel et al. / Behavioural Brain Research 194 (2008) 119–128 121
Fig. 1. Recording configuration and behavioral paradigm. (a) Monkeys sat in front of a computer screen, and controlled an on-screen cursor by applying an isometric two-dimensional wrist torque in two axes: flexion–extension and radial-ulnar. The monkey’s hand was situated in either a pronation or suppination position. Single unit (SU)recordings were made from the motor cortex and cervical spinal cord simultaneously and EMG activity was recorded from up to 12 forearm muscles. (b) Sequence of eventscomposing a single trial. A trial was initiated by the appearance of a central target. The monkey positioned the cursor inside the target by generating zero torque for a restperiod (500–600 ms;). Then eight peripheral targets appeared, uniformly distributed around the center target at a fixed distance defining the onset of a delay period. Oneof these peripheral targets was presented in a distinct color for 500 ms (cue). The disappearance of the central target (850–1700 ms after cue onset) served as a “go” signal.T etricf pearer trial( or in t
iac
v
s
FTTcs
he monkey then had to acquire the previously filled target by generating an isomor an active-torque period (350–750 ms). Subsequently, the peripheral targets disapeceived a reward, after which the screen went blank for 1000–1500 ms and a newcue on) is marked by a magenta square. (For interpretation of the references to col
ntact animal encompasses many factors such as firing properties
nd network interactions in addition to the underlying anatomicalonnectivity.A more recent study [27] of spinal circuitry introduced an inno-ative view of the way segmental IN networks may operate. It
pamw
ig. 2. Spinal response to motor cortical stimulation. Single-pulse electrical stimulationshe unprocessed spinal recording is depicted around stimulus times (40 randomly selecteo isolate the multiunit spiking response the spinal recording was processed by band-paomponents. (c) The mean processed signal was computed separately for each set of sttimulation. (d) Similar effect shown for data collected from a second monkey.
torque in the appropriate direction, and to keep the cursor within the target boxd and the central target reappeared. The monkey returned to the rest position andstarted. Events and epochs are marked in the figure. Time at which target is filledhis figure legend, the reader is referred to the web version of the article.)
howed that when invoking a scratch reflex in an isolated spinal
reparation of a turtle, the input to MNs was composed of a bal-nced excitation–inhibition. This arrangement is different than theore traditionally assumed push–pull action INs exert on MNs, inhich excitation and inhibition alternate. While this mode of oper-were applied in the motor cortex while recording spinal population response. (a)d traces are shown). About 3 ms after stimulation there is a multiunit response. (b)ss filtering (1000–6000 Hz) and signal rectification, thus removing low-frequencyimulation intensities. Spinal response was evident already at the lowest level of
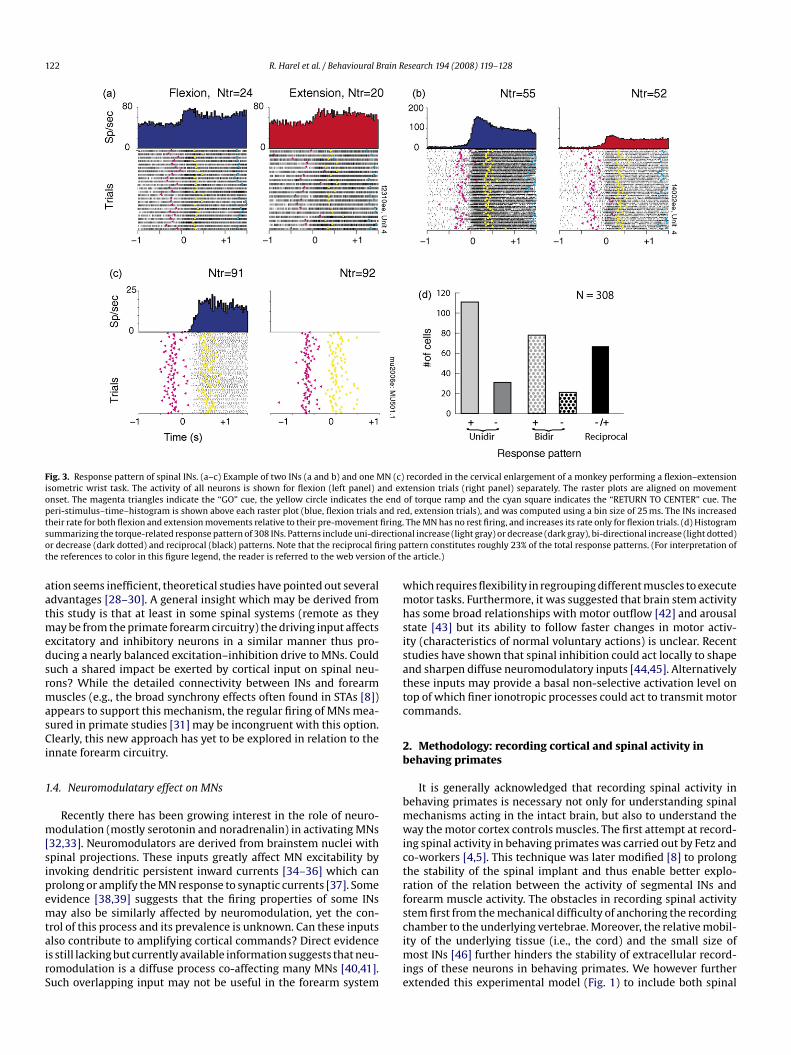
122 R. Harel et al. / Behavioural Brain Research 194 (2008) 119–128
Fig. 3. Response pattern of spinal INs. (a–c) Example of two INs (a and b) and one MN (c) recorded in the cervical enlargement of a monkey performing a flexion–extensionisometric wrist task. The activity of all neurons is shown for flexion (left panel) and extension trials (right panel) separately. The raster plots are aligned on movementonset. The magenta triangles indicate the “GO” cue, the yellow circle indicates the end of torque ramp and the cyan square indicates the “RETURN TO CENTER” cue. Theperi-stimulus–time–histogram is shown above each raster plot (blue, flexion trials and red, extension trials), and was computed using a bin size of 25 ms. The INs increasedt firing.s ectiono ring pt n of th
aatmedsrmasCi
1
m[sipemtairS
wmhsisattc
2b
bmwictrfs
heir rate for both flexion and extension movements relative to their pre-movementummarizing the torque-related response pattern of 308 INs. Patterns include uni-dirr decrease (dark dotted) and reciprocal (black) patterns. Note that the reciprocal fihe references to color in this figure legend, the reader is referred to the web versio
tion seems inefficient, theoretical studies have pointed out severaldvantages [28–30]. A general insight which may be derived fromhis study is that at least in some spinal systems (remote as they
ay be from the primate forearm circuitry) the driving input affectsxcitatory and inhibitory neurons in a similar manner thus pro-ucing a nearly balanced excitation–inhibition drive to MNs. Coulduch a shared impact be exerted by cortical input on spinal neu-ons? While the detailed connectivity between INs and forearmuscles (e.g., the broad synchrony effects often found in STAs [8])
ppears to support this mechanism, the regular firing of MNs mea-ured in primate studies [31] may be incongruent with this option.learly, this new approach has yet to be explored in relation to the
nnate forearm circuitry.
.4. Neuromodulatary effect on MNs
Recently there has been growing interest in the role of neuro-odulation (mostly serotonin and noradrenalin) in activating MNs
32,33]. Neuromodulators are derived from brainstem nuclei withpinal projections. These inputs greatly affect MN excitability bynvoking dendritic persistent inward currents [34–36] which canrolong or amplify the MN response to synaptic currents [37]. Somevidence [38,39] suggests that the firing properties of some INsay also be similarly affected by neuromodulation, yet the con-
rol of this process and its prevalence is unknown. Can these inputslso contribute to amplifying cortical commands? Direct evidences still lacking but currently available information suggests that neu-omodulation is a diffuse process co-affecting many MNs [40,41].uch overlapping input may not be useful in the forearm system
cimie
The MN has no rest firing, and increases its rate only for flexion trials. (d) Histogramal increase (light gray) or decrease (dark gray), bi-directional increase (light dotted)
attern constitutes roughly 23% of the total response patterns. (For interpretation ofe article.)
hich requires flexibility in regrouping different muscles to executeotor tasks. Furthermore, it was suggested that brain stem activity
as some broad relationships with motor outflow [42] and arousaltate [43] but its ability to follow faster changes in motor activ-ty (characteristics of normal voluntary actions) is unclear. Recenttudies have shown that spinal inhibition could act locally to shapend sharpen diffuse neuromodulatory inputs [44,45]. Alternativelyhese inputs may provide a basal non-selective activation level onop of which finer ionotropic processes could act to transmit motorommands.
. Methodology: recording cortical and spinal activity inehaving primates
It is generally acknowledged that recording spinal activity inehaving primates is necessary not only for understanding spinalechanisms acting in the intact brain, but also to understand theay the motor cortex controls muscles. The first attempt at record-
ng spinal activity in behaving primates was carried out by Fetz ando-workers [4,5]. This technique was later modified [8] to prolonghe stability of the spinal implant and thus enable better explo-ation of the relation between the activity of segmental INs andorearm muscle activity. The obstacles in recording spinal activitytem first from the mechanical difficulty of anchoring the recording
hamber to the underlying vertebrae. Moreover, the relative mobil-ty of the underlying tissue (i.e., the cord) and the small size ofost INs [46] further hinders the stability of extracellular record-ngs of these neurons in behaving primates. We however furtherxtended this experimental model (Fig. 1) to include both spinal

R. Harel et al. / Behavioural Brain Research 194 (2008) 119–128 123
Fig. 4. Signal and noise correlation analysis of spinal and motor cortical neurons. The similarity in response pattern and response variability was measured using the signaland noise correlation. (a) An example of a computed signal (left) and noise (right) correlation values for a pair of INs. The signal correlation was computed as the correlationcoefficient between the two torque-dependent mean firing rates. The noise correlation was computed as the correlation coefficient between the normalized rate level(computed by subtracting the mean signal and dividing by the standard deviation). (b) The relation between signal (x-axis) and noise (y-axis) correlations computed for 204pairs of spinal interneurons. The marginal distributions of these parameters are shown as well. Note that the noise correlation values are symmetrically distributed aroundz . (c ani alysisw rrelatiN (ada
aumfiCtcistptcttu(mctw
3
wpm
ciwdp
3c
tmtircnTtoitm
ero and that they are independent of the corresponding signal correlation valuessometric wrist task as spinal neurons. Here we separated the noise correlation anas significantly biased towards positive values (t-test, p < 0.001) and the noise coonetheless, the effects were stronger for the rest period than for active hold period
nd motor cortical recordings [47,48]. This configuration allowss to address directly questions of transmission and processing ofotor-related information through the corticospinal system. The
rst advantage of simultaneous recordings is that we can identifyS connectivity either when it is evoked via electrical stimula-ions or ongoing, by computing various measures of functionalonnectivity. The second consideration is that motor performances inherently variable. Hence, comparing neurons recorded eithertrictly simultaneously, or in successive sessions could overcomehis behaviourally related variability to highlight unique responseroperties of each neuronal population. Our recording configura-ion is illustrated in Fig. 1. Monkeys were first implanted with aortical recording chamber located above the motor cortex. Wehen mapped the primary motor cortex and premotor cortex (ven-ral and dorsal). After completing the mapping procedure (lastingsually 3–4 weeks), monkeys were implanted with EMG electrodessubcutaneous or transcutaneous electrodes) in up to 14 forearm
uscles. After recovery, we implanted the monkey with a spinalhamber above the cervical enlargement (C4-T1). We recorded cor-ical activity (via one or two electrodes) in parallel to spinal activity,hile monkey performed a two-dimensional isometric wrist task.
. Computational role of INs
Based on the data available on the anatomy of descending path-ays and spinal functional connectivity with muscles in behavingrimates, our current working hypothesis is that during voluntaryovement spinal interneuronal circuitry participates in the pro-
a(syu
d d) The same analysis was applied to cortical neurons recorded during the sameinto rest period (c) and active hold period (d). In both cases, the noise correlationon was significantly related to the signal correlation (linear regression, p < 0.001).pted from Prut and Perlmutter [52]).
essing of cortical commands. The current challenge we are facings to understand what information is transmitted via this route,
hat role it plays in motor control, and how it interacts with theirect CM pathway. Next we review our current notions as to howrocessing takes place in the segmental level of the indirect route.
.1. Cortical input to INs is strong and has clear functionalorrelates
In reduced preparations the existence of cortical drive is oftenested using spatial and/or temporal summation of inputs. This
ode of activation indicates that cortical stimulation (applied onhe cortical surface [49] or the pyramidal tract [50]) can invokentracellular monosynatpic potentials but rarely fires spinal neu-ons. Hence INs were considered to be only weakly affected byortical drive alone. Instead, it was assumed that driving theseeurons required co-activation of descending and afferent volleys.his presumed activation pattern was interpreted as an indica-ion that these neurons serve to incorporate motor commands intongoing motor states [51] and for low-level maintenance of ongo-ng motor actions in terms of joint stiffness and muscle-yield. Weested this hypothesis [47] directly and found that in behaving pri-
ates intracortical single-pulse stimulations can readily invoke
spinal spiking response (Fig. 2). We found that M1 sites often∼30% of the cases) induced such a response. Spinal responseslightly increased when the intensity of stimulation was increased,et responses were already observed at the lowest level of stim-lation (Fig. 2c and d). On the other hand, non-responsive sites

1 rain R
rTstovcbctt
3
napdapttvcnctr[stcr
3
bIaeipptrmereammai
iraitfom
mptipaiofid
ittisr
outotim
3n
cttccttmubwflspibticratocs(
wINs) is required for the final steps of translating motor commands
24 R. Harel et al. / Behavioural B
emained as such even at the highest levels of stimulation (250 �A).he theoretical likelihood of finding connected cortical and spinalites (given the fact that sites for cortical and spinal electroderacks were switched independently on a daily bases and thus thebserved CS connectivity pattern was unbiased) appeared at firstery remote. Therefore, the actual finding of multiple connectedortical and spinal sites consistently in three monkeys suggests aroad divergent/convergent connectivity pattern between motorortex and spinal cord. Such an extensive connectivity may implyhe involvement of segmental circuitry in the transmission of cor-ical command to muscles.
.2. Firing properties of INs
Spinal interneurons operate at a high firing level [7,31]. Theseeurons fire vigorously at rest (namely, when no torque is exerted)nd often increase their rate in both the preferred and non-referred torque direction (Fig. 3). This response pattern is veryifferent from the firing of MNs (Fig. 3c), which are silent at restnd fire only when an action is performed roughly within theirreferred direction. The increased IN excitability may seem at firsto simply add noise that could deteriorate the signal these neuronsransmit, but the elevated rate might be necessary in order to acti-ate spinal MNs. Furthermore, we have shown [52] that the noiseorrelation (i.e., trial-to-trial covariability of rate) among simulta-eously recorded spinal INs is lower compared to values found forortical neurons (Fig. 4) This attribute may be useful in representinghe cortical command in a non-redundant manner. Removing theedundancy which is known to exist among motor cortical neurons53] may be needed when transmitting information from the large-cale cortical network into much smaller populations of MNs. It ishus possible that spinal IN circuitry serves to translate the corticalommand into a non-redundant and amplified version as might beequired for rapid initiation of muscle activity.
.3. Dual role of cortical command on spinal flow of information
Pre-synaptic inhibition of afferent information was shown toe mediated in the spinal cord by a specific group of GABAergic
Ns [54]. These neurons synapse directly on incoming afferentsnd thus can block the flow of information without affecting thexcitability of the post-synaptic cell. Human studies have shownndirectly that such an attenuation of afferent information takeslace just before movement onset [55,56]. Direct IN recordings inrimates [57] have shown explicitly that during movement onsethe evoked spinal response to cutaneous stimulation is greatlyeduced, while the excitability of the cell is in fact enhanced. Such aechanism can change the balance among descending and periph-
ral inputs converging upon spinal neurons. Effectively, this processeduces the impact of a predicted self-generated feedback, thusnhancing the feedforward flow of information. The time-windowt which pre-synaptic inhibition takes place may suggest that theotor system switches between a feedforward mode of control ofovement onset, in which the motor cortex dictates motor output,
nd a feedback mode of control through which a constant positions maintained.
We tested the functional implications of such a model by study-ng the similarity in tuning between cortical and spinal neuronsecorded from functionally connected sites [47]. Our underlyingssumption was that if indeed during movement onset cortical
nput acts in parallel to activate INs and attenuate sensory flowhen cortical drive should be enhanced during this period. Theunctional correlate of this enhancement was a transient alignmentf tuning between cortical and spinal neurons (Fig. 5). This align-ent may indicate that cortical neurons recruit spinal neurons withieurc
esearch 194 (2008) 119–128
atching tuning properties. Furthermore, tuning was not a stableroperty of either cortical or spinal neurons. It is also conceivablehat cortical drive is the source that induces the transiently match-ng tuning among connected spinal neurons. Clearly this was aopulation-based phenomenon, which was not readily observed onsingle-neuronal level. Nonetheless, the increased tuning similar-
ty was accompanied by an increase in the trial-to-trial variabilityf firing. This increased variability (observed after correcting forring rate modulation) was prominent in time lags where corticalrive preceded the spinal response.
The robustness of the CS route was further demonstrated implic-tly by the fact that unconnected sites expressed a decrease inuning similarity. Such a decrease is expected if the connectivityo a single spinal site is derived from a broad region of cortical areancluding all sites with a preferred direction (PD) near the targetedpinal PD. In this case, we expect that unconnected sites will not beandomly related in their PDs but will tend to have dissimilar PDs.
Our results thus provide complementary evidence to the workf Seki et al. [57], indicating the dual role of descending commandpon spinal circuitry. Furthermore, our findings show that the func-ional implication of this process is a parametric representationf the motor command at the spinal level in terms of directionalorque-tuning (or its derivatives). As such, spinal INs could partic-pate in the processing of motor commands as a link between the
otor cortex and the spinal cord.
.4. Mixed representation of space and muscles exhibited by M1eurons
The question of a direct versus indirect pathway from the motorortex to muscles [1,26] is partially related to the question ofhe parametric representation of information in the motor cor-ex. If indeed M1 encodes motor information in muscle-basedoordinates than presumably there is no need for any further pro-essing between output units of M1 and working muscles. Onhe other hand, if M1 coding is more abstract, e.g., space-based,hen there may be a need for translating this command into a
uscle based representation [58,59]. We studied this question bysing a paradigm that was designed to dissociate extrinsic, muscle-ased and hand-based coordinate frames [60,61]. We found thathen using an isometric paradigm, M1 neurons operated in aexible frame of coordinates that gradually changed from extrin-ic (after cue onset) to near-intrinsic (during the sustained holderiod). At movement onset the majority of M1 neurons operated
n a coordinate frame which was neither extrinsic nor intrinsic,ut corresponded to a partial shift between the two systems. Onhe other hand, INs recorded during the same task expressed anntrinsic-based coding before movement onset and maintained thisoding scheme throughout sustained hold period. Several theo-etical studies [62,63] have suggested that a partial PD shift ischieved when assuming integration between external (target loca-ion) and internal (hand-position) information by a populationf neurons. Our results further suggest that this integration pro-ess is dynamic, such that the functional impact of these twoources is constantly changing according to the behavioral contextFig. 6).
Two cautionary remarks are relevant to this discussion. First,hile our results may suggest that spinal circuitry (MNs and/or
nto muscle activation the correlation between changes in PDs andxtrinsic or intrinsic representations are not necessarily trivial ornique [64]. Second, it is possible that the internal (muscle-based)epresentation emerges independently in INs and MNs with noausal link between these two representations.

R. Harel et al. / Behavioural Brain Research 194 (2008) 119–128 125
Fig. 5. Signal and noise correlates of corticospinal connectivity. (a) Example of torque-related activity of simultaneously recorded cortical and spinal neurons. Trials weresegregated according to the target reached (eight evenly distributed targets at equal distance from center position) and were aligned on torque onset. (b) Color maps presentingsignal (upper panels) and noise (lower panels) correlations computed for pairs recorded from connected (left) and unconnected (right) corticospinal sites. Each plot presentsthe average correlation matrix where the x-axis corresponds to time relative to torque onset and the y-axis corresponds to cortical-to-spinal time lags (positive lags indicatecortical signal preceding spinal signal). The Z-transformed correlation values are color coded so that the positive correlation is shown in red and the negative correlation isshown in blue. For connected sites, positive signal correlation values were found before torque onset and before steady torque. A positive noise correlation was found forthese sites before torque onset in time lags indicating cortical activity driving spinal activity. For unconnected sites, we found a negative signal correlation and zero noisecorrelation. (c) An illustration summarizing the events that occur during torque onset. The cortical signal is enhanced (due to synchronized firing of these cells) while theafferent flow of information is suppressed via cortically driven pre-synaptic inhibition. Thus, the balance of inputs affecting spinal INs at this epoch is biased in favor of thecortical command (adapted from Yanai et al. [47]). (For interpretation of the references to color in this figure legend, the reader is referred to the web version of the article.)

126 R. Harel et al. / Behavioural Brain Research 194 (2008) 119–128
Fig. 6. Coordinate frame of spinal and cortical activity. (a) Task-related activity of spinal neurons during trials performed with hand either pronated (upper plot) or suppinated(lower plot). Activity is aligned on cue onset (left) and torque onset (right). The gap between these two plots is due to the variable-delay policy used in this task. On the right,the PD (computed as the vector summation) of the neuron is shown for a time-window after torque onset. The obtained PDs were tested for significance and the differencebetween the PD levels (�PD) during the two hand postures was used to assign the appropriate coordinate frame. (b) �PDs were computed using a 300 ms sliding windowand a 50 ms step. The average �PD was computed in each window for cortical and spinal neurons that were tuned in the two postures. The percentage of cortical (red) andspinal (blue) neurons available for analysis at each point in time is color coded. As the trial progresses the representation of information gradually shifts from an extrinsic(zero shift) to an intrinsic (∼70◦ shift) representation of information. However, before movement onset, the spinal coordinate frame is already near-intrinsic while corticalneurons express a partial PD shift (∼30◦) at that time. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of the article.)

rain R
4ir
oit[IwTmtpms
(
(
(
(
A
5F
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
R. Harel et al. / Behavioural B
. The emerging picture about the role of spinalnterneurons during voluntary movements: concludingemarks
Our current understanding of spinal circuitry is primary basedn studies in reduced preparations. These studies have providednvaluable information about the organization of interneurons, andheir presumed role in integration of motor-related information23,50]. More recent studies have documented the activity of spinalNs during voluntary movements of primates and their relations
ith cortical activity on one hand and muscle activity on the other.his information is a first, yet critical, step in identifying the wayotor commands are translated into motor action. Taken together
hese results portray a picture in which spinal neurons serve as arocessing unit which matches up motor commands and workinguscles. This process includes several (simultaneously occurring)
teps:
a) Emergence of an enhanced and tuned “envelope” of activationat the segmental level which may operate to amplify the corticalinput as required for activating MNs.
b) Removal of space-based information from the cortical com-mand. This means translating the cortical command into amuscle-based representation. This process may occur simul-taneously in MNs and INs, but specific (muscle-based) INsactivation is required to bring MNs close to threshold.
(c) Subtraction of the predicted feedback when it is not useful.This is achieved through pre-synaptic inhibition of afferent flowwhile maintaining the increased excitability of INs.
d) Removing redundancy which may exist in the cortical com-mand and thus producing a more efficient representation ofconverging information. Note that this property may providethe spinal mechanism required to eliminate space-based infor-mation as mentioned above.
e) According to this view, a descending command affects bothsegmental interneurons and motoneurons. Cortico-segmentalinput provides the baseline activation pattern required forincreased excitability of motoneurons. Corticomotoneuronalinput operates in parallel to specifically recruit working motorunits. The direct CM route is insufficient to activate muscles,while the indirect pathways cannot provide the specific andindependent muscle activation which is a prerequisite for vol-untary action. Studies where the direct CM connection wasremoved provide evidence for the latter point [65,66]. This par-allel activation is not only a way to amplify cortical commandsbut also provides a link through which cortical commands caninteract with the flow of peripheral information depending onthe motor context. Thus, the emerging direct route exploits theexisting spinal machinery by fine-tuning it and hence generat-ing remarkable motor flexibility with minimal energetic costs(in terms of cortical processing). The existence of this spinallink also saves the motor cortex from representing movementin exclusive muscle-based coordinates: it is the corticospinaltermination pattern (probably on both INs and MNs) which han-dles this final step of coordinate transformation. In this scenario,the motor cortical circuitry transmits information downstreamin a transient manner and thus is continuously ready for anyrequired change in the motor state, while the spinal circuitry isresponsible for maintaining a pre-defined motor state.
cknowledgments
This work was supported by the Israel Science Foundation (ISF-55/01 and ISF-1355/05), the Rosetrees Foundation and the Baruchoundation.
[
[
esearch 194 (2008) 119–128 127
eferences
[1] Lemon RN, Kirkwood PA, Maier MA, Nakajima K, Nathan P. Direct and indirectpathways for corticospinal control of upper limb motoneurons in the primate.Prog Brain Res 2004;143:263–79.
[2] Bortoff GA, Strick PL. Corticospinal terminations in two new-world pri-mates: further evidence that corticomotoneuronal connections provide partof the neural substrate for manual dexterity. J Neurosci 1993;13(12):5105–18.
[3] Bennett KM, Lemon RN. Corticomotoneuronal contribution to the fractiona-tion of muscle activity during precision grip in the monkey. J Neurophysiol1996;75(5):1826–42.
[4] Bromberg MB, Fetz EE. Responses of single units in cervical spinal cord of alertmonkeys. Exp Neurol 1977;55(2):469–82.
[5] Courtney KR, Fetz EE. Unit responses recorded from cervical spinal cord ofawake monkey. Brain Res 1973;53(2):445–50.
[6] Fetz EE, Perlmutter SI, Maier MA, Flament D, Fortier PA. Response patternsand postspike effects of premotor neurons in cervical spinal cord of behavingmonkeys. Can J Physiol Pharmacol 1996;74(4):531–46.
[7] Maier MA, Perlmutter SI, Fetz EE. Response patterns and force relations ofmonkey spinal interneurons during active wrist movement. J Neurophysiol1998;80(5):2495–513.
[8] Perlmutter SI, Maier MA, Fetz EE. Activity of spinal interneurons and theireffects on forearm muscles during voluntary wrist movements in the monkey.J Neurophysiol 1998;80(5):2475–94.
[9] Shinoda Y, Yokota J, Futami T. Divergent projection of individual corticospinalaxons to motoneurons of multiple muscles in the monkey. Neurosci Lett1981;23(1):7–12.
10] Scheibel ME, Scheibel AB. A structural analysis of spinal interneurons and Ren-shaw cells. UCLA Forum Med Sci 1969;11:159–208.
11] Ornung G, Ottersen OP, Cullheim S, Ulfhake B. Distribution of glutamate-,glycine- and GABA-immunoreactive nerve terminals on dendrites in the catspinal motor nucleus. Exp Brain Res 1998;118(4):517–32.
12] Umbriaco D, Garcia S, Beaulieu C, Descarries L. Relational features of acetyl-choline, noradrenaline, serotonin and GABA axon terminals in the stratumradiatum of adult rat hippocampus (CA1). Hippocampus 1995;5(6):605–20.
13] Porter R, Lemon RN. Corticospinal function and voluntary movement. Oxford:Calrendon Press; 1993.
14] Lawrence DG, Porter R, Redman SJ. Corticomotoneuronal synapses in the mon-key: light microscopic localization upon motoneurons of intrinsic muscles ofthe hand. J Comp Neurol 1985;232(4):499–510.
15] Jankowska E, Padel Y, Tanaka R. Projections of pyramidal tract cells toalpha-motoneurones innervating hind-limb muscles in the monkey. J Physiol1975;249(3):637–67.
16] Jankowska E, Padel Y, Tanaka R. Disynaptic inhibition of spinal motoneuronesfrom the motor cortex in the monkey. J Physiol 1976;258(2):467–87.
17] Fetz EE, Cheney PD. Postspike facilitation of forelimb muscle activity by primatecorticomotoneuronal cells. J Neurophysiol 1980;44(4):751–72.
18] Dum RP, Strick PL. Spinal cord terminations of the medial wall motor areas inmacaque monkeys. J Neurosci 1996;16(20):6513–25.
19] Lemon RN, Griffiths J. Comparing the function of the corticospinal system indifferent species: organizational differences for motor specialization? MuscleNerve 2005;32(3):261–79.
20] Alstermark B, Isa T, Pettersson LG, Sasaki S. The C3–C4 propriospinal systemin the cat and monkey: a spinal pre-motoneuronal centre for voluntary motorcontrol. Acta Physiol (Oxf) 2007;189(2):123–40.
21] Maier MA, et al. Differences in the corticospinal projection from primary motorcortex and supplementary motor area to macaque upper limb motoneurons: ananatomical and electrophysiological study. Cereb Cortex 2002;12(3):281–96.
22] Kuypers HG, Brinkman J. Precentral projections to different parts of the spinalintermediate zone in the rhesus monkey. Brain Res 1970;24(1):29–48.
23] Jankowska E. Interneuronal relay in spinal pathways from proprioceptors. ProgNeurobiol 1992;38(4):335–78.
24] Maier MA, Illert M, Kirkwood PA, Nielsen J, Lemon RN. Does a C3–C4propriospinal system transmit corticospinal excitation in the primate? Aninvestigation in the macaque monkey. J Physiol 1998;511(Pt 1):191–212.
25] Isa T, Ohki Y, Seki K, Alstermark B. Properties of propriospinal neurons inthe C3–C4 segments mediating disynaptic pyramidal excitation to forelimbmotoneurons in the macaque monkey. J Neurophysiol 2006;95(6):3674–85.
26] Isa T, Ohki Y, Alstermark B, Pettersson LG, Sasaki S. Direct and indirect cortico-motoneuronal pathways and control of hand/arm movements. Physiology(Bethesda) 2007;22:145–52.
27] Berg RW, Alaburda A, Hounsgaard J. Balanced inhibition and excitation drivespike activity in spinal half-centers. Science 2007;315(5810):390–3.
28] van Vreeswijk C, Sompolinsky H. Chaos in neuronal networks with balancedexcitatory and inhibitory activity. Science 1996;274(5293):1724–6.
29] Litvak V, Sompolinsky H, Segev I, Abeles M. On the transmission of rate codein long feedforward networks with excitatory–inhibitory balance. J Neurosci
2003;23(7):3006–15.30] Abbott LF, Chance FS. Drivers and modulators from push–pull and balancedsynaptic input. Prog Brain Res 2005;149:147–55.
31] Prut Y, Perlmutter SI. Firing properties of spinal interneurons duringvoluntary movement. I. State-dependent regularity of firing. J Neurosci2003;23(29):9600–10.

1 rain R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
28 R. Harel et al. / Behavioural B
32] Hounsgaard J, Hultborn H, Jespersen B, Kiehn O. Bistability of alpha-motoneurones in the decerebrate cat and in the acute spinal cat afterintravenous 5-hydroxytryptophan. J Physiol 1988;405:345–67.
33] Heckman CJ, Lee RH, Brownstone RM. Hyperexcitable dendrites in motoneu-rons and their neuromodulatory control during motor behavior. TrendsNeurosci 2003;26(12):688–95.
34] Schwindt PC, Crill WE. Properties of a persistent inward current innormal and TEA-injected motoneurons. J Neurophysiol 1980;43(6):1700–24.
35] Schwindt P, Crill W. Role of a persistent inward current in motoneuron burstingduring spinal seizures. J Neurophysiol 1980;43(5):1296–318.
36] Lee RH, Heckman CJ. Influence of voltage-sensitive dendritic conductances onbistable firing and effective synaptic current in cat spinal motoneurons in vivo.J Neurophysiol 1996;76(3):2107–10.
37] Hultborn H, Denton ME, Wienecke J, Nielsen JB. Variable amplification of synap-tic input to cat spinal motoneurones by dendritic persistent inward current. JPhysiol 2003;552(Pt 3):945–52.
38] Kiehn O, Johnson BR, Raastad M. Plateau properties in mammalian spinalinterneurons during transmitter-induced locomotor activity. Neuroscience1996;75(1):263–73.
39] Derjean D, Bertrand S, Le Masson G, Landry M, Morisset V, Nagy F. Dynamicbalance of metabotropic inputs causes dorsal horn neurons to switch functionalstates. Nat Neurosci 2003;6(3):274–81.
40] Skagerberg G, Bjorklund A. Topographic principles in the spinal projections ofserotonergic and non-serotonergic brainstem neurons in the rat. Neuroscience1985;15(2):445–80.
41] Huisman AM, Kuypers HG, Verburgh CA. Quantitative differences in collater-alization of the descending spinal pathways from red nucleus and other brainstem cell groups in rat as demonstrated with the multiple fluorescent retro-grade tracer technique. Brain Res 1981;209(2):271–86.
42] Jacobs BL, Martin-Cora FJ, Fornal CA. Activity of medullary serotonergic neu-rons in freely moving animals. Brain Res Brain Res Rev 2002;40(1–3):45–52.
43] Aston-Jones G, Chen S, Zhu Y, Oshinsky ML. A neural circuit for circadian regu-lation of arousal. Nat Neurosci 2001;4(7):732–8.
44] Hyngstrom AS, Johnson MD, Miller JF, Heckman CJ. Intrinsic electricalproperties of spinal motoneurons vary with joint angle. Nat Neurosci2007;10(3):363–9.
45] Heckman CJ, Hyngstrom AS, Johnson MD. Active properties of motoneuronedendrites: diffuse descending neuromodulation, focused local inhibition. J
Physiol 2008;586(5):1225–31.46] Hunt CC, Kuno M. Properties of spinal interneurones. J Physiol1959;147:346–63.
47] Yanai Y, Adamit N, Harel R, Israel Z, Prut Y. Connected corticospinal sitesshow enhanced tuning similarity at the onset of voluntary action. J Neurosci2007;27(45):12349–57.
[
[
esearch 194 (2008) 119–128
48] Yanai Y, Adamit N, Israel Z, Harel R, Prut Y. Coordinate transforma-tion is first completed downstream of primary motor cortex. J Neurosci2008;28(7):1728–32.
49] Jankowska E, Padel Y, Tanaka R. The mode of activation of pyramidal tract cellsby intracortical stimuli. J Physiol 1975;249(3):617–36.
50] Baldissera F, Hultborn H, Illert M. Integration in spinal neuronal systems. In:Brookhart JM, Mountcastle VB editors. The nervous system. American Physio-logical Society: Bethesda; 1981, p. 509–595.
51] Todorov E. Direct cortical control of muscle activation in voluntary arm move-ments: a model. Nat Neurosci 2000;3(4):391–8.
52] Prut Y, Perlmutter SI. Firing properties of spinal interneurons during vol-untary movement. II. Interactions between spinal neurons. J Neurosci2003;23(29):9611–9.
53] Lee D, Port NL, Kruse W, Georgopoulos AP. Variability and correlated noise inthe discharge of neurons in motor and parietal areas of the primate cortex. JNeurosci 1998;18(3):1161–70.
54] Rudomin P, Schmidt RF. Presynaptic inhibition in the vertebrate spinal cordrevisited. Exp Brain Res 1999;129(1):1–37.
55] Voss M, Ingram JN, Haggard P, Wolpert DM. Sensorimotor attenuation bycentral motor command signals in the absence of movement. Nat Neurosci2006;9(1):26–7.
56] Haggard P, Whitford B. Supplementary motor area provides an efferent signalfor sensory suppression. Brain Res Cogn Brain Res 2004;19(1):52–8.
57] Seki K, Perlmutter SI, Fetz EE. Sensory input to primate spinal cord is presynapti-cally inhibited during voluntary movement. Nat Neurosci 2003;6(12):1309–16.
58] Bizzi E, Cheung VC, d′Avella A, Saltiel P, Tresch M. Combining modules formovement. Brain Res Rev 2008;57(1):125–33.
59] Alstermark B, Isa T. Premotoneuronal and direct corticomotoneuronal controlin the cat and macaque monkey. Adv Exp Med Biol 2002;508:281–97.
60] Kakei S, Hoffman DS, Strick PL. Direction of action is represented in the ventralpremotor cortex. Nat Neurosci 2001;4(10):1020–5.
61] Kakei S, Hoffman DS, Strick PL. Muscle and movement representations in theprimary motor cortex. Science 1999;285(5436):2136–9.
62] Buneo CA, Jarvis MR, Batista AP, Andersen RA. Direct visuomotor transforma-tions for reaching. Nature 2002;416(6881):632–6.
63] Deneve S, Duhamel JR, Pouget A. Optimal sensorimotor integration in recur-rent cortical networks: a neural implementation of Kalman filters. J Neurosci2007;27(21):5744–56.
64] Shah A, Fagg AH, Barto AG. Cortical involvement in the recruitment of wristmuscles. J Neurophysiol 2004;91(6):2445–56.
65] Pettersson LG, Alstermark B, Blagovechtchenski E, Isa T, Sasaski S. Skilled digitmovements in feline and primate—recovery after selective spinal cord lesions.Acta Physiol (Oxf) 2007;189(2):141–54.
66] Sasaki S, Isa T, Pettersson LG, Alstermark B, Naito K, Yoshimura K, Seki K, etal. Dexterous finger movements in primate without monosynaptic corticomo-toneuronal excitation. J Neurophysiol 2004;92(5):3142–7.




















