







Bahasa
Halaman
Hukum


Chapter 6
Phylogenetic Relationships of Harpyionycterine Megabats(Chiroptera: Pteropodidae)
NORBERTO P. GIANNINI1,2, FRANCISCA CUNHA ALMEIDA1,3,
AND NANCY B. SIMMONS1
ABSTRACT
After almost 70 years of stability following publication of Andersen’s (1912) monograph onthe group, the systematics of megachiropteran bats (Chiroptera: Pteropodidae) was thrown intoflux with the advent of molecular phylogenetics in the 1980s—a state where it has remained eversince. One particularly problematic group has been the Austromalayan Harpyionycterinae,currently thought to include Dobsonia and Harpyionycteris, and probably also Aproteles. In thiscontribution we revisit the systematics of harpyionycterines. We examine historical hypothesesof relationships including the suggestion by O. Thomas (1896) that the rousettine Boneia bidensmay be related to Harpyionycteris, and report the results of a series of phylogenetic analysesbased on new as well as previously published sequence data from the genes RAG1, RAG2, vWF,c-mos, cytb, 12S, tVal, 16S, and ND2. Despite a striking lack of morphological synapomorphies,results of our combined analyses indicate that Boneia groups with Aproteles, Dobsonia, andHarpyionycteris in a well-supported, expanded Harpyionycterinae. While monophyly of thisgroup is well supported, topological changes within this clade across analyses of different datapartitions indicate conflicting phylogenetic signals in the mitochondrial partition. The positionof the harpyionycterine clade within the megachiropteran tree remains somewhat uncertain.Nevertheless, biogeographic patterns (vicariance-dispersal events) within Harpyionycterinaeappear clear and can be directly linked to major biogeographic boundaries of theAustromalayan region. The new phylogeny of Harpionycterinae also provides a new frameworkfor interpreting aspects of dental evolution in pteropodids (e.g., reduction in the incisordentition) and allows prediction of roosting habits for Harpyionycteris, whose habits areunknown.
INTRODUCTION
During the last decade, the traditionalclassification of Megachiroptera (Mammalia:Chiroptera) has been challenged by a numberof molecular phylogenetic studies that havecalled into question many of the systematicgroupings established by Andersen (1912)and revised recently by Bergmans (1997).One particularly controversial megachirop-teran taxon has been Harpyionycterinae
Miller, 1907, an Austromalayan group cur-rently thought to include HarpyionycterisThomas, 1896, Dobsonia Palmer, 1898, andalmost certainly Aproteles Menzies, 1977(Giannini et al., 2006). There has beendisagreement about the affinities of thenominate genus since its description, largelydue to a suite of unique craniodentalcharacter states seen in Harpyionycteris (seeAndersen, 1912). Thomas (1896) rathervaguely suggested placement of Harpyionyc-
Bulletin of the American Museum of Natural History amnb-582-00-06.3d 24/9/09 15:57:01 183 Cust # 582-6
1 Department of Mammalogy, American Museum of Natural History, Central Park West at 79th Street, New York, NY 10024-5192, USA ([email protected]).
2 Programa de Investigaciones de Biodiversidad Argentina, Facultad de Ciencias Naturales e Instituto Miguel Lillo, Miguel Lillo205, Universidad Nacional de Tucuman, Tucuman, CP 4000, Argentina.
3 Sackler Institute for Comparative Genomics, Molecular Systematics Laboratory, American Museum of Natural History, CentralPark West at 79th Street, New York, NY 10024, USA.

teris with Rousettus Gray, 1921, and BoneiaJentink, 1879. In contrast, Andersen (1912)linked Harpyionycteris with Dobsonia (seebelow), and Boneia with Rousettus. Boneiawas synonymized with Rousettus by Berg-mans and Rozendaal (1988) and this arrange-ment has been followed by many authors(Bergmans, 1994, 1997; Corbet and Hill,1992; Simmons, 2005).
Andersen (1912) formally followed Miller(1907) in recognizing Harpyionycterinae as adistinct group, but wrote in extenso about aclose relationship between Harpyionycterisand Dobsonia. He concluded that ‘‘So evidentis the phylogenetic connection between thesetwo genera that Harpyionycteris may be said,almost with certainty, to be the peculiarlymodified Philippine representative of theAustro-Malayan Dobsonia’’ (Andersen, 1912:803). Miller and Hollister (1921) subsequent-ly described Harpyionycteris celebensis fromSulawesi, providing a further biogeographiclink between Harpyionycteris and Dobsonia.
In spite of Andersen’s (1912) arguments,all subsequent authors (e.g., Corbet and Hill,1992; Koopman, 1993, 1994; Tate, 1951;Slaughter, 1970) followed Miller (1907) inconsidering Harpyionycteris as a peculiarpteropodid best placed in a subfamily of itsown. However, two recent studies providedevidence supporting Andersen’s (1912) hy-pothesis of a close relationship betweenHarypionycteris and Dobsonia. Support forthis grouping was found in Romagnoli andSpringer’s (2000) analysis of morphologicaldata, although caution must be exercised ininterpreting these results because of minimalclade support and the small number ofcharacters used in that study. More compel-lingly, separate and combined parsimony
analyses of two coding genes, one nuclear(exon 28 of the von Willebrand factor gene)and one mitochondrial (cytochrome b),recovered a highly supported clade groupingHarpyionycteris whiteheadi and four speciesof Dobsonia from three distinct speciesgroups (Giannini et al., 2006). As a conse-quence of these results, Giannini et al. (2006)expanded Harpyionycterinae Miller, 1907, toinclude Dobsonia. They also concurred withprevious authors in concluding that Aprotelesprobably also belongs in this subfamily basedboth on general morphological evidence(Flannery, 1995; Menzies, 1977; Gianniniand Simmons, 2005; Springer et al., 1995)and results of all previous phylogeneticanalyses in which Aproteles was included(Colgan and da Costa, 2002; Giannini andSimmons, 2003, 2005; Jones et al., 2002;Kirsch et al., 1995; Romagnoli and Springer,2000; Springer et al., 1995).
The harpyionycterines as currently under-stood occupy three major Austromalayansubregions, the Philippine, Wallacean, andPapuan (table 1). The distributions of a fewspecies marginally exceed the boundaries ofthose territories and enter the Australian andSundaic regions (see Byrnes, 2005; Corbet andHill, 1992; Koopman, 1993, 1994; Simmons,2005). The Austromalayan region is crossed bymajor biogeographic boundaries, including theWallace, Lyddeker, and Weber lines. Organ-isms distributed across these various boundar-ies, such as harpyionycterine bats, may con-tribute to the understanding of the complexconnections among the Philippine, Wallacean,and Papuan biogeographic regions and, moregenerally, between Asia and Australia.
In the present study, we explore further themembership and affinities of the expanded
Bulletin of the American Museum of Natural History amnb-582-00-06.3d 24/9/09 15:57:02 184 Cust # 582-6
TABLE 1Internal Primers for RAG1 and vWF Genes Designed for This Study
Gene Primer 59–39 sequence
RAG1 RAG1f2 GCCAAGCCCTTCATTGAGACA
RAG1r2 CTATGGAAGGGACTGTCTCAATG
vWF vWFf2 CTTYGTGSTCAGTGGTGTGGA
vWFr2 TCCACACCACTGASCACRAAG
vWFf3 CTATGYCCAGGGCCTGAAGAA
vWFr3 GCTCCTGTTGAAATTGGCCT
184 BULLETIN AMERICAN MUSEUM OF NATURAL HISTORY NO. 582

Harpyionycterinae (sensu Giannini et al.,2006) using new molecular data as well aspublished sequences of relevant taxa. We testthe relationship of Boneia and Rousettus toHarpyionycteris as well as the relationship ofAproteles to Dobsonia, and examine theproblem of placing Harpyionycterinae in themegachiropteran tree. Finally, we explore thebiogeographic patterns suggested by phylo-genetic relationships recovered in this study.
METHODS
TAXA
We selected megachiropteran taxa in orderto test monophyly of Harpyionycterinae ascurrently defined (Harpyionycteris, Dobsonia,and Aproteles) and investigate relationshipsof these taxa to other genera of megabats aspreviously proposed in the literature. Specif-ically, we sought to test (1) the association ofBoneia, Rousettus, and Harpyionycteris pro-posed in the description of the latter (Thomas,1896) versus the association of Boneia withRousettus (Andersen, 1912; Bergmans andRozendaal, 1988) and Harpyionycteris withDobsonia (Andersen, 1912; Giannini et al.,2006); (2) the sister relationship of Aprotelesand Dobsonia (Colgan and da Costa, 2002;Giannini and Simmons, 2003, 2005; Menzies,1977; Romagnoli and Springer, 2000); and (3)the inclusion of Dobsonia and their relatives ina pteropodine clade (Romagnoli and Spring-er, 2000) versus a rousettine clade (Andersen,1912; Bergmans, 1997).
Megachiropteran terminals were chosenfrom each main pteropodid clade followingGiannini et al. (2006): Nyctimene albiventerand N. vizcaccia (Nyctimeninae), Cynopterussphinx and Ptenochirus jagori (Cynopteri-nae), Boneia bidens, Eonycteris spelaea, Rou-settus amplexicaudatus, and R. aegyptiacus(Rousettinae), Myonycteris torquata andMegaloglossus woermanni (Epomophorinae,Myonycterini), Epomops franqueti and Epo-mophorus wahlbergi (Epomophorinae, Epo-mophorini), Macroglossus minimus and Syco-nycteris australis (Macroglossinae), Melony-cteris fardoulisi and Melonycteris melanops(Pteropodinae, Notopterini), and Pteropushypomelanus and P. tonganus (Pteropodinae,Pteropodini). Harpyionycterinae megabats
(sensu Giannini et al., 2006) included are:Aproteles bulmerae, Harpyionycteris white-headi, H. celebensis, and a member of eachof the four currently recognized Dobsoniaspecies groups (Simmons, 2005), namelyDobsonia minor (minor species group), inermis(viridis group), D. moluccensis (moluccensisgroup), and D. peronii (peronii group). HereD. magna is considered a junior synonym ofD. moluccensis following Helgen (2007; seealso Byrnes, 2005; Koopman, 1994).
Trees were rooted using Rhinopoma hard-wickii (Rhinopomatidae) and Artibeus jamai-censis (Phyllostomidae). Rhinopomatids arecurrently thought to belong to Rhinolophoi-dea, which is the sister clade of megabats inmultigene phylogenetic analyses, while phyl-lostomids represent the other major clade ofecholocating bats, Yangochiroptera (Eick etal., 2005; Teeling et al., 2005; Miller-Butter-worth et al., 2007).
SEQUENCES
We used sequences of exon 28 of the vonWillebrand factor gene (vWF, 1231 bp) andcytochrome b (cytb, 1140 bp) from Gianniniet al. (2006), and generated new sequences forharpyionycterine species for this study. Wealso incorporated sequences of two othernuclear coding genes, the recombinationactivating gene 1 (RAG1, 1084 bp), and therecombination activating gene 2 (RAG2, 760bp) from Giannini et al. (2008) for the sametaxa, and similarly generated new sequencesfor harpyioncterines. In order to includerelevant species whose samples were notavailable to us, we used published sequencesof the mitochondrial 12S rDNA (12S, ca. 970bp) and Valine tDNA (tVal, ca. 70 bp) andcontributed new sequences for these genes.For additional analyses (see below), weincorporated published sequences of themitochondrial 16S rDNA (16S, ca. 1560 bpfor complete taxa) and NADH dehydroge-nase subunit 2 (ND2, 1044 bp) genes, and theoocyte maturation factor Mos proto-onco-gene (c-mos, ca. 480 bp). Composition of thedata set, accession numbers of all sequencesused, and voucher information for newsequences are given in appendix 1.
Total DNA was obtained from preservedtissue samples with the DNeasy tissue kit
Bulletin of the American Museum of Natural History amnb-582-00-06.3d 24/9/09 15:57:03 185 Cust # 582-6
2009 GIANNINI ET AL.: HARPYIONYCTERINE PHYLOGENY 185

(QIAGEN). PCR amplification was carriedout using previously published primers (12S-tVal: Springer et al., 1995; cytb: Bastian et al.,2002; RAG1 and RAG2: Teeling et al., 2000;vWF: Porter et al., 1996). To obtain bothforward and reverse sequences for each generegion, additional internal primers weredeveloped for the RAG1 and vWF genes(table 1). All sequences were obtained with anautomated ABI 3730XL sequencer. Sequenceediting and prealignment were done with theSequencher 4.2 software (Gene Codes). Ac-cession numbers for the new sequences areFJ218461-84. Although for some speciesmore than one sample was available, onlyone was included in each analysis. The aligneddataset is provided at ftp://ftp.amnh.org/pub/group/mammalogy/downloads/.
PHYLOGENETIC ANALYSES
Sequences of our coding and ribosomalgenes were submitted to parsimony analysis.In the present study, we used data for thegenes for which we contributed new sequences(i.e., 12S-tVal, cytb, RAG1, RAG2, and vWF)to conduct three principal sets of phylogeneticanalyses: (1) individual gene partitions; (2)nuclear and mitochondrial partitions; and (3)all genes combined. Here we report results ofthe main partitions and the combined set, andcomment on results from the individual geneswhere relevant. In additional analyses, weused genes not sequenced in our study (16S,ND2, and c-mos in individual and combinedsets; see details below).
Analyses involving only coding genes(individual analyses for c-mos, cytb, RAG1,RAG2, vWF, and the nuclear partition) wereperformed under conventional parsimony(static alignment). The tree search strategyconsisted of 500 replicates of random addi-tion sequences of taxa (RAS) each followedby tree bisection reconnection branch swap-ping (TBR). An additional round of TBRwas done on optimal trees so obtained. Cladesupport was assessed using Bremer or decayvalues (Bremer, 1994) and character resam-pling (Goloboff et al., 2003). We calculatedBremer values via incremental sampling ofsuboptimal trees (see Giannini and Bertelli,2004). Briefly, we saved up to 2000 subopti-mal trees one step longer than the previous
optimum in successive stages. That is, we firstsearched for suboptimal trees 1 step longerthan the optimal tree length, next savingsuboptimals at every step from 2 through 10steps longer than the optimal trees. Second,group frequency (based on unbiased sym-metric resampling; Goloboff et al., 2003) wascalculated on the basis of 5000 replications.These analyses were executed in TNT 1.0(Goloboff et al., 2004, 2008).
For all analyses involving ribosomal andtransfer RNA genes (i.e., individual 12S-tValpartition, the mitochondrial partition, thecombined set, and additional analyses using16S), we used direct optimization (DO;Wheeler, 1996). In this approach, alignmentis viewed as dynamic (i.e., varying acrosstopologies in tree space) and is thereforelinked to tree search. The transformation costof whole unaligned sequences on a topology iscalculated via optimization. Costs of eachtransformation type (indel or substitution)were established a priori—as in every align-ment procedure. According to these costs, thelength of the tree is calculated as the sum oftransformations on each internal node in thedownpass optimization, which minimizesindels and substitutions (steps) of wholesequences on the candidate tree. Minimum-length trees are chosen among those visited.In practice, this is done by way of treebuilding and branch swapping. We set equalcosts of indels and substitutions, and oursearch strategy consisted of replicated swap-ping + refinements (e.g., Giannini and Sim-mons, 2003, 2005). Briefly, 100 RAS + TBRbranch swapping were run, collecting alloptimal trees from each replication. Thosetrees were submitted to tree fusing (Goloboff,1999). In analyses combining ribosomal andcoding genes, the latter were treated asprealigned; i.e., no indels were allowed incoding genes (static alignment). All DOanalyses, including searches to calculateBremer support values, were performed usingPOY 4 (Varon et al., 2007).
Additional phylogenetic analyses were con-ducted to take advantage of additional pub-lished data. First, we added published 16S andc-mos sequences to our combined set toprovide a ‘‘total-evidence’’ analysis based onsequences available for the majority of ourtaxonomic samples. Second, we ran a specific
Bulletin of the American Museum of Natural History amnb-582-00-06.3d 24/9/09 15:57:03 186 Cust # 582-6
186 BULLETIN AMERICAN MUSEUM OF NATURAL HISTORY NO. 582

analysis of ND2 with the few sequences ofmegabats available in GenBank (which un-fortunately had low overlap with our taxo-nomic sampling) to provide a separate test ofthe affinities of Boneia. Data available includ-ed ND2 sequences of Dobsonia minor amongthe harpyionycterines and also Boneia bidens.This analysis was important because our dataon Boneia originated from a single individual(ZMA 23100) from which we extracted DNAand generated all sequences, whereas the ND2sequences originated from DNA that wasamplified from a different individual (Gen-Bank accession number AY504581).
In addition to the parsimony analyses, weperformed a maximum-likelihood (ML) treesearch for the combined nuclear data set usinga GTR + C + I partitioned model (i.e.,parameters were estimated from the data foreach gene separately) using the programRAxML (Stamatakis, 2006; Stamatakis et al,2008). Our search strategy consisted of execut-ing 100 rapid bootstrap inferences and there-after a thorough ML search. Statistical supportwas obtained with 100 bootstrap replications.
BIOGEOGRAPHIC ANALYSIS
Our biogeographic analysis consisted ofmapping onto the optimal tree obtained fromour total-evidence phylogenetic analysis asingle biogeographic character composed ofsix states, each of which consisted of knowndistribution areas of terminals (see below). Weperformed a partial (downpass-only) Fitch(1971) optimization in order to recover thepattern of minimal biogeographic events re-quired by the tree. In this case, we did notintend to provide wide coverage of Megachir-optera; our aim was instead to identify themain dispersal-vicariance events of our in-group, the Harpyionycterinae—those eventsthat led to the origin of currently recognizedgenera. This attempt is necessarily incompleteas we did not include all Dobsonia species.Therefore we restricted our analysis to thesampled harpyionycterines and their areas ofoccurrence, which were coded in a singlemultistate, unordered character whose statesconsisted of broad biogeographic subregionsof the eastern Austromalayan region asrecognized by Corbet and Hill (1992): Sula-wesi (SUL), the Philippines (PHI), Lesser
Sunda Islands (LSI), the Moluccas (MOL),New Guinea (NGU), Australia (AUS), andthe Solomon Islands (SOL). Taxa occurringin more than one area were scored aspolymorphic (for instance, Dobsonia minorwas scored NGU/SUL).
RESULTS
NUCLEAR PARTITION
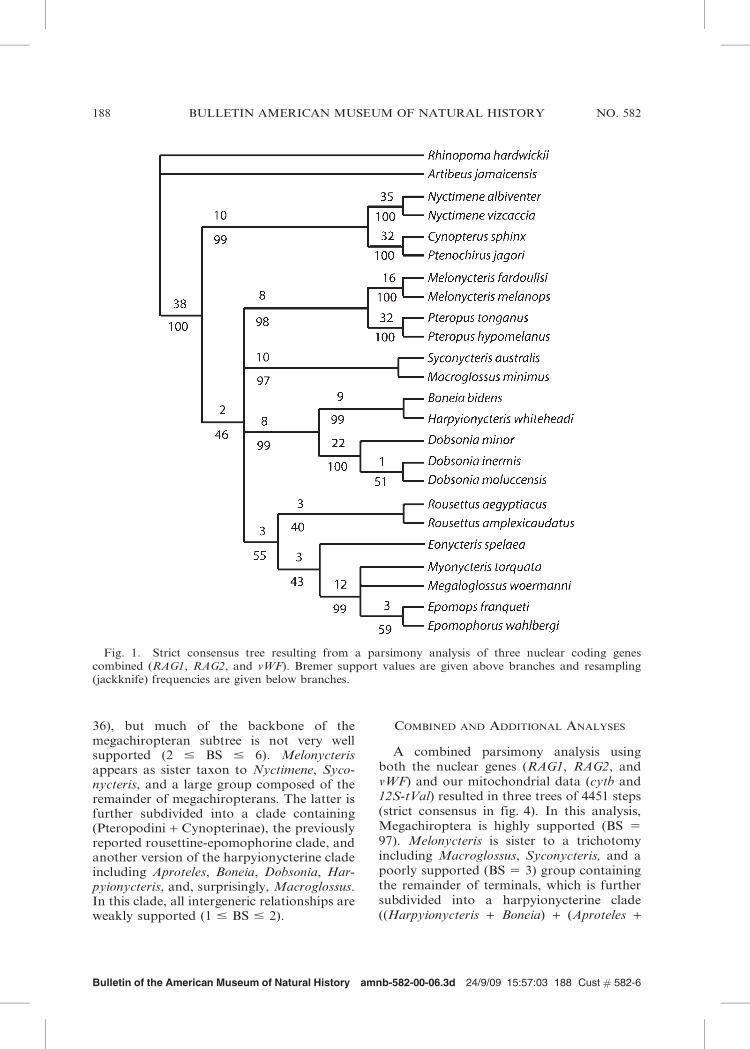
Parsimony analysis of nuclear data (RAG1,RAG2, and vWF sequences) resulted in 10most-parsimonious trees of 1922 steps (strictconsensus tree in fig. 1). In this tree, Mega-chiroptera is highly supported (Bremer sup-port value, BS 5 38). Mutually monophyleticNyctimeninae and Cynopterinae form a cladethat corresponds with Andersen’s (1912)Cynopterus section, and is sister to a poorlysupported multichotomy of four clades con-taining all other megachiropterans. The fourgroups are: a reduced macroglossine clade(Macroglossus + Syconycteris); a reducedpteropodine clade including Melonycterisand Pteropus (see Bergmans, 1997; Gianniniet al., 2008); a clade composed of rousettine(Rousettus and Eonycteris) and epomophor-ine megabats, including Myonycterini (Mega-loglossus and Myonycteris) and Epomophor-ini (Epomophorus and Epomops); and aharpyionycterine clade (sensu Giannini etal., 2006) that includes Boneia as sister toHarpyionycteris, and both sister to Dobsonia.Our ML analysis (fig. 2) recovered the samemajor groupings as the parsimony analysis(fig. 1), resolving the backbone polytomy ofthe parsimony result (cf. fig. 1) with varyingdegrees of support (bootstrap values 32–82).This topology is identical to one of theoptimal topologies obtained under parsimony(i.e., in this analysis, parsimony is conserva-tive in reflecting conflicts in the data).Regarding harpyionycterine megabats, thebranching pattern is the same in both MLand MP analyses (cf. figs. 1 and 2).
MITOCHONDRIAL PARTITION
The analysis of the 12S-tVal + cytbsequences under direct optimization pro-duced a single tree at 3220 steps (fig. 3).Megachiroptera is highly supported (BS 5
Bulletin of the American Museum of Natural History amnb-582-00-06.3d 24/9/09 15:57:03 187 Cust # 582-6
2009 GIANNINI ET AL.: HARPYIONYCTERINE PHYLOGENY 187
36), but much of the backbone of themegachiropteran subtree is not very wellsupported (2 # BS # 6). Melonycterisappears as sister taxon to Nyctimene, Syco-nycteris, and a large group composed of theremainder of megachiropterans. The latter isfurther subdivided into a clade containing(Pteropodini + Cynopterinae), the previouslyreported rousettine-epomophorine clade, andanother version of the harpyionycterine cladeincluding Aproteles, Boneia, Dobsonia, Har-pyionycteris, and, surprisingly, Macroglossus.In this clade, all intergeneric relationships areweakly supported (1 # BS # 2).
COMBINED AND ADDITIONAL ANALYSES
A combined parsimony analysis usingboth the nuclear genes (RAG1, RAG2, andvWF) and our mitochondrial data (cytb and12S-tVal) resulted in three trees of 4451 steps(strict consensus in fig. 4). In this analysis,Megachiroptera is highly supported (BS 5
97). Melonycteris is sister to a trichotomyincluding Macroglossus, Syconycteris, and apoorly supported (BS 5 3) group containingthe remainder of terminals, which is furthersubdivided into a harpyionycterine clade((Harpyionycteris + Boneia) + (Aproteles +
Bulletin of the American Museum of Natural History amnb-582-00-06.3d 24/9/09 15:57:03 188 Cust # 582-6
Fig. 1. Strict consensus tree resulting from a parsimony analysis of three nuclear coding genescombined (RAG1, RAG2, and vWF). Bremer support values are given above branches and resampling(jackknife) frequencies are given below branches.
188 BULLETIN AMERICAN MUSEUM OF NATURAL HISTORY NO. 582

Dobsonia)), and another group containingthe rousettine-epomophorine clade and acomposite clade (Pteropodini (Nyctimeninae+ Cynopterinae)).
Adding to this data set published sequenc-es of the mitochondrial 16S and the nuclearoncogene c-mos genes produces a single treeof 6119 steps (fig. 5). In this tree, Nyctimeneis sister to all other megachiropterans, andthis tree partition appeared highly supported(BS 5 19). Successive clades recovered were(Pteropodini + Cynopterinae), the rousettine-epomophorine clade, and a highly supportedharpyionycterine clade (BS 5 23) in which
Dobsonia was sister to Aproteles, Boneia, andHarpyionycteris. Several important dispersal-vicariance events are suggested by mappingdistribution of terminals onto the optimaltree of our combined phylogenetic analysis.These patterns are discussed below.
Finally, we obtained four trees of 1717steps in a separate analysis based on ND2sequences from pteropodid species availableon GenBank. The strict consensus tree(fig. 6) was generally poorly supported butplaced Boneia bidens and Dobsonia minor assister taxa. This result is compatible withthose of previous analyses that separately
Bulletin of the American Museum of Natural History amnb-582-00-06.3d 24/9/09 15:57:06 189 Cust # 582-6
Fig. 2. Maximum likelihood tree resulting from the analysis of the nuclear dataset (RAG1, RAG2, andvWF; gene partitions modeled using GTR + C + I; ML score -11,136.227759). Values near nodes representbootstrap estimates of clade support based on 100 replications.
2009 GIANNINI ET AL.: HARPYIONYCTERINE PHYLOGENY 189

support an association of Boneia with har-pyionycterine megabats—D. minor amongthe latter.
DISCUSSION
PHYLOGENETIC RELATIONSHIPS
AND CLASSIFICATION
The phylogenetic analyses of nuclear datarecovered a harpyionycterine clade composed
of Harpyionycteris, Aproteles, Dobsonia, and,rather unexpectedly, Boneia. Although Mac-roglossus was added to this group in theanalysis of the mitochondrial partition (withnegligible support, 1 # BS # 2; fig. 3), thegroup membership of the nuclear analysis wasrecovered in the total-evidence analysis withhigh support (BS 5 23; fig. 5). This representsstrong evidence that Boneia is not a synonymof Rousettus but a distinct genus that belongsin Harypionycterinae. We also confirmed the
Bulletin of the American Museum of Natural History amnb-582-00-06.3d 24/9/09 15:57:11 190 Cust # 582-6
Fig. 3. Single optimal tree resulting from a direct optimization, parsimony analysis of threemitochondrial genes combined (12S-tVal + cytb). Bremer support values are given below branches.
190 BULLETIN AMERICAN MUSEUM OF NATURAL HISTORY NO. 582

inclusion of Aproteles in this group (seediscussion in Giannini et al., 2006). Thus,we formally expand Harpyionycterinae Mil-ler, 1907, to include Boneia Jentink, 1879,Harpyionycteris Thomas, 1896, DobsoniaPalmer, 1898, and Aproteles Menzies, 1977.
Resolving the composition of Harpyionyc-terinae has proven difficult throughout thehistory of megachiropteran systematics.Thomas (1896) recognized the difficulties offinding allies of Harpyionycteris among the
megachiropterans known at the time of itsdescription. Thomas (1896: 243) attributedthe similarities in canine morphology withHarpyia (5 Nyctimene) to either homoplasy(‘‘accident’’ in his words) or plesiomorphy(i.e., ‘‘common descent from the (presum-ably) cuspidate-toothed ancestor of the Pter-opodidae’’). Thomas (1896: 243) suggestedthat ‘‘On the whole … [Harpyionycteris] maybe most conveniently placed near Xanthar-pyia [5 Rousettus] and Boneia, with which it
Bulletin of the American Museum of Natural History amnb-582-00-06.3d 24/9/09 15:57:16 191 Cust # 582-6
Fig. 4. Strict consensus tree of a parsimony analysis of RAG1, RAG2, vWF, cytb and 12S-tVal genescombined. Bremer support values are given below branches.
2009 GIANNINI ET AL.: HARPYIONYCTERINE PHYLOGENY 191

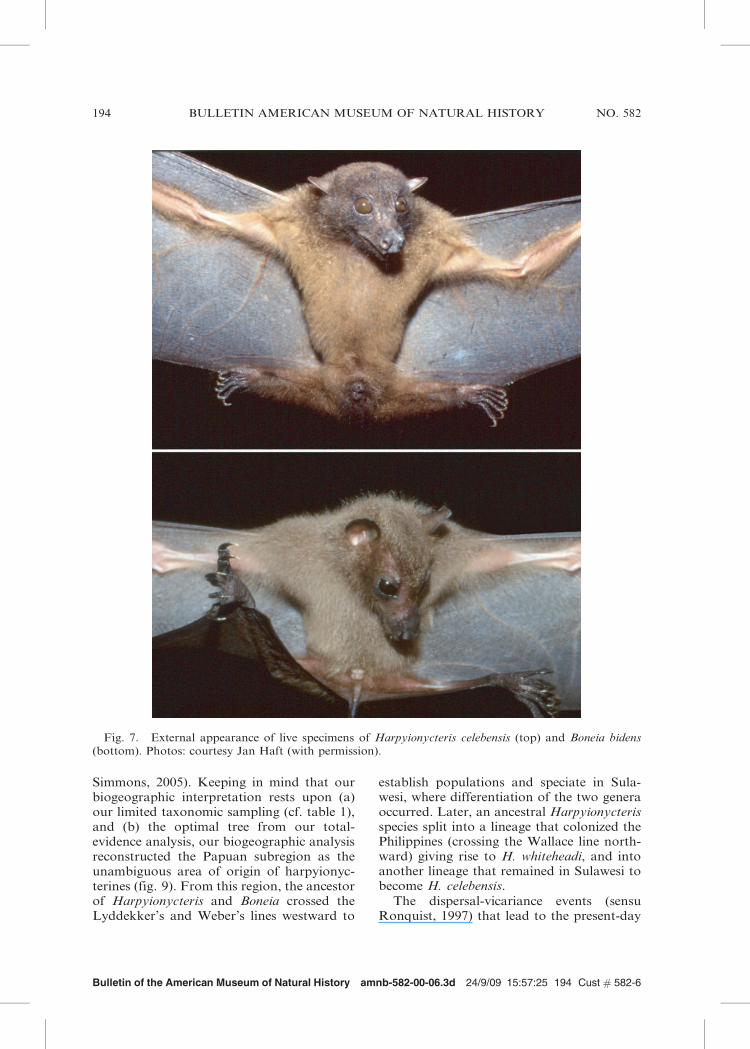
shares certain external characters, an indicalclaw, and the cheek-tooth formula of P. 3/3,M. 2/3.’’ The index finger claw and the cheek-tooth formula are likely plesiomorphic forMegachiroptera (see Giannini and Simmons,2005, 2007), and the external characters werenot specified (but see the external resem-blance of Harpyionycteris celebensis andBoneia bidens in fig. 7). Nevertheless, it seemsclear that Thomas (1896) did correctlyperceive a link between these forms.
Despite Thomas’s (1896) intuition and themolecular support discovered in the currentstudy, to our knowledge, no author subse-quent to Thomas considered a subfamily ortribal group inclusive of both Harpyionycterisand Boneia. This is reflected in a completelack of potential morphological synapomor-phies for such grouping in the literature. Inrecent systematic studies involving Boneia,discussion has centered on the nature of itsrelationship with Rousettus (e.g., Bergmans,
Bulletin of the American Museum of Natural History amnb-582-00-06.3d 24/9/09 15:57:20 192 Cust # 582-6
Fig. 5. Single optimal tree of a parsimony analysis of RAG1, RAG2, vWF, cytb, 12S-tVal, 16S, and c-mos genes combined. Bremer support values are given below branches.
192 BULLETIN AMERICAN MUSEUM OF NATURAL HISTORY NO. 582

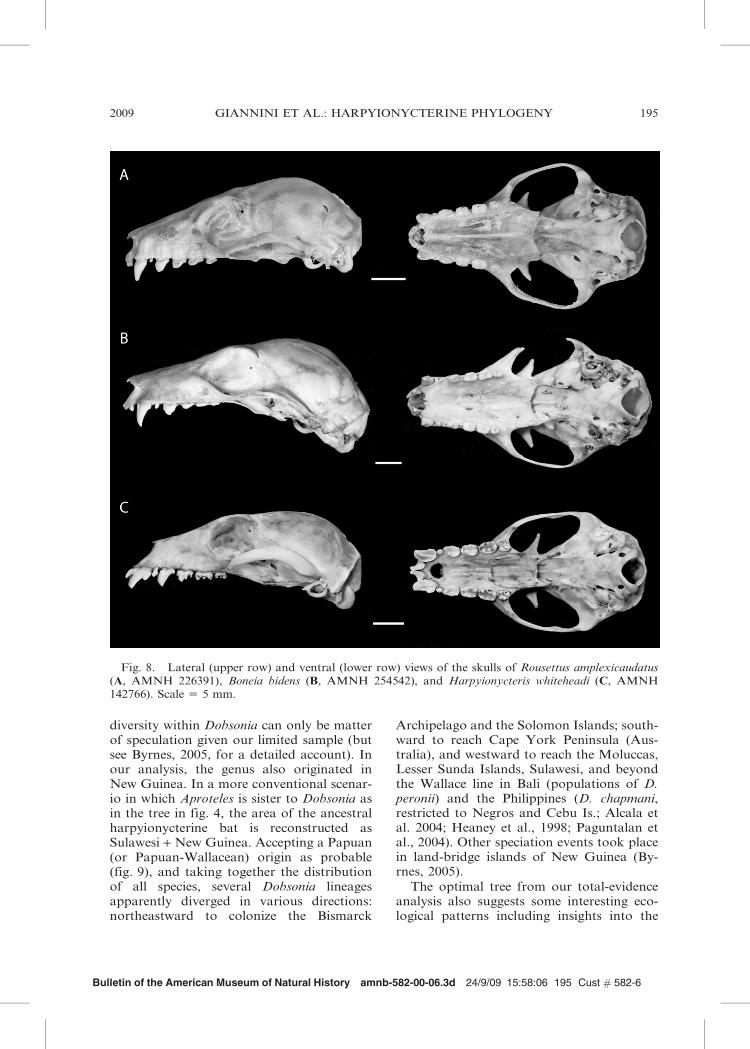
1994; Bergmans and Rozendaal, 1988). Thisis not surprising given the prominent rolethat Rousettus has played in megachiropteransystematics as a result of long being consid-ered close to the ancestral form of megabatsfollowing Andersen (1912). Although thisassessment has been refuted by all cladisticstudies since the early 1990s (e.g., Colgan andda Costa, 2002; Giannini and Simmons,2003, 2005; Kirsch et al., 1995; Romagnoliand Springer, 2000; Springer et al., 1995),most megachiropteran genera have beencompared with Rousettus on these grounds,and Boneia was no exception. Given the lackof unusual external features, unremarkableskull shape, dental formula close to thegeneralized condition of megabats, and thehabit of roosting gregariously in caves,Boneia was considered so close an ally ofRousettus as to be included in Rousettus as asubgenus (see Bergmans and Rozendaal,1988). The morphological evidence discussedby the authors in favor of this associationincludes both morphometric and qualitativedata and is rather robust, which makes ourfinding of a Boneia-Harpyionycteris clademore surprising. As shown in fig. 8, theskulls of Boneia and Harpyionycteris bearlittle mutual resemblance. In Boneia, the
rostrum is thin and strongly deflected relativeto the basicranial axis, the dentition issimplified but otherwise typically pteropodid,the zygomatic arch is weak, and in overallappearance it resembles Rousettus. By con-trast, Harpyionycteris shows a strong ‘‘dob-sonine’’ skull with the most peculiar rostrumand dentition seen in megabats (see com-ments in Giannini et al., 2006).
However difficult it may seem to reconcilethe current composition of the harpyionyc-terine clade with the morphological variationtraditionally considered in megabat system-atics, our phylogeny does suggest someinteresting scenarios of morphological andecological evolution. One particularly signif-icant example involves the dentition. Har-pyionycterines exhibit dental formulae thatvary across taxa, but are similar in the sensethat one formula can be parsimoniouslytransformed into any other in one-step lossesor gains. A most relevant case is the firstupper incisor, which is the sole tooth missingfrom the generalized pteropodid formula inBoneia (a variably deciduous tooth in thistaxon; Bergmans and Rozendaal, 1988). Thisis also the incisor demonstrably lacking inDobsonia and hypothesized to be absent inHarpyionycteris (see Giannini and Simmons,2007: table 2). Aproteles lacks both upperincisors. Our phylogeny, once it is acceptedas a supported hypothesis, allows us tosuggest that the loss or reduction of I1 iseffectively homologous across harpyionycter-ine taxa, and that the loss of incisors inAproteles likely happened sequentially (I1first lost first, followed by I2).
BIOGEOGRAPHIC AND ECOLOGICAL PATTERNS
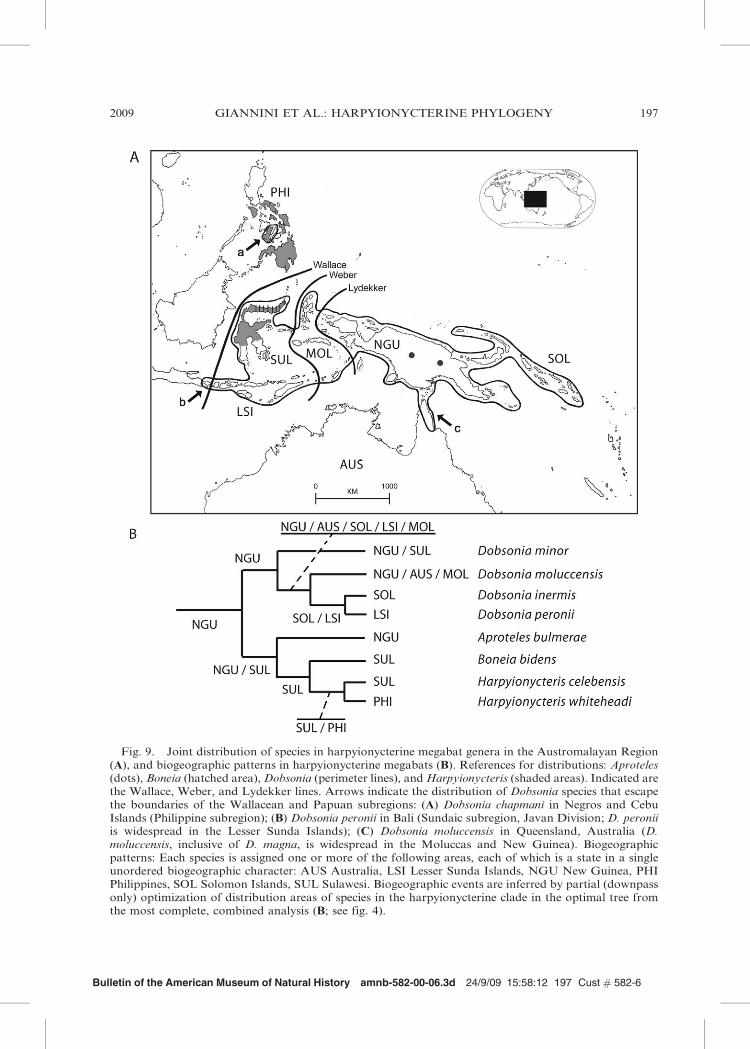
Harpyionycterinae sensu Giannini et al.(2006; i.e., composed of Harpyionycteris,Dobsonia, and Aproteles), expanded here toinclude Boneia, comprises 18 species whosejoint distribution encompasses the Philippine,Wallacean, and Papuan subregions of theAustromalayan area, plus two species thatnarrowly escape the biogeographic boundar-ies of those subregions (fig. 9; table 2). Thelatter two taxa are D. peronii, which alsooccurs in Bali (Sundaic subregion) and D.moluccensis, which also occurs in Queens-land, Australia (Corbet and Hill, 1992;
Bulletin of the American Museum of Natural History amnb-582-00-06.3d 24/9/09 15:57:24 193 Cust # 582-6
Fig. 6. Strict consensus tree of a parsimonyanalysis of the ND2 gene. Bremer and jackknifevalues (above and below branches, respectively)are reported for the harpyionycterine clade recov-ered in this analysis. Numbers in parenthesisindicate number of terminals included in thecynopterine and pteropodine clades recovered.
2009 GIANNINI ET AL.: HARPYIONYCTERINE PHYLOGENY 193
Simmons, 2005). Keeping in mind that ourbiogeographic interpretation rests upon (a)our limited taxonomic sampling (cf. table 1),and (b) the optimal tree from our total-evidence analysis, our biogeographic analysisreconstructed the Papuan subregion as theunambiguous area of origin of harpyionyc-terines (fig. 9). From this region, the ancestorof Harpyionycteris and Boneia crossed theLyddekker’s and Weber’s lines westward to
establish populations and speciate in Sula-wesi, where differentiation of the two generaoccurred. Later, an ancestral Harpyionycterisspecies split into a lineage that colonized thePhilippines (crossing the Wallace line north-ward) giving rise to H. whiteheadi, and intoanother lineage that remained in Sulawesi tobecome H. celebensis.
The dispersal-vicariance events (sensuRonquist, 1997) that lead to the present-day
Bulletin of the American Museum of Natural History amnb-582-00-06.3d 24/9/09 15:57:25 194 Cust # 582-6
Fig. 7. External appearance of live specimens of Harpyionycteris celebensis (top) and Boneia bidens(bottom). Photos: courtesy Jan Haft (with permission).
194 BULLETIN AMERICAN MUSEUM OF NATURAL HISTORY NO. 582
diversity within Dobsonia can only be matterof speculation given our limited sample (butsee Byrnes, 2005, for a detailed account). Inour analysis, the genus also originated inNew Guinea. In a more conventional scenar-io in which Aproteles is sister to Dobsonia asin the tree in fig. 4, the area of the ancestralharpyionycterine bat is reconstructed asSulawesi + New Guinea. Accepting a Papuan(or Papuan-Wallacean) origin as probable(fig. 9), and taking together the distributionof all species, several Dobsonia lineagesapparently diverged in various directions:northeastward to colonize the Bismarck
Archipelago and the Solomon Islands; south-ward to reach Cape York Peninsula (Aus-tralia), and westward to reach the Moluccas,Lesser Sunda Islands, Sulawesi, and beyondthe Wallace line in Bali (populations of D.peronii) and the Philippines (D. chapmani,restricted to Negros and Cebu Is.; Alcala etal. 2004; Heaney et al., 1998; Paguntalan etal., 2004). Other speciation events took placein land-bridge islands of New Guinea (By-rnes, 2005).
The optimal tree from our total-evidenceanalysis also suggests some interesting eco-logical patterns including insights into the
Bulletin of the American Museum of Natural History amnb-582-00-06.3d 24/9/09 15:58:06 195 Cust # 582-6
Fig. 8. Lateral (upper row) and ventral (lower row) views of the skulls of Rousettus amplexicaudatus(A, AMNH 226391), Boneia bidens (B, AMNH 254542), and Harpyionycteris whiteheadi (C, AMNH142766). Scale 5 5 mm.
2009 GIANNINI ET AL.: HARPYIONYCTERINE PHYLOGENY 195

evolution of roosting habits (see also discus-sion in Byrnes, 2005). Roost availability is oneof the limiting factors of bat populations(Kunz and Pierson, 1994). Megachiropteransare either cavity dwellers (occupying struc-tures from tree holes to large caves) or foliagedwellers (a habit that varies from takingsolitary refuge on leaves or vines to living inlarger groups in actively defoliated trees;Kunz and Pierson, 1994). The roosting habitsof Harpyionycteris are unknown, but thisgenus is nested within successive sister cladesthat include many typical cave-dwelling bats:Boneia (Bergmans and Rozendaal, 1988),Aproteles (Flannery, 1995; Menzies, 1977),and several Dobsonia species including D.moluccensis (Bonaccorso, 1998; Flannery,1995; Helgen, 2007), and probably also D.peronii (Chiroptera Specialist Group, 1996)and D. inermis. By contrast, D. minor hasbeen reported to roost both in foliage(Flannery, 1995) and in caves (Boeadi andBergmans, 1987). The prediction we drawfrom reconstructing roosting habits (foliage
vs. cavities) in the optimal tree of our mostcomplete analysis is that Harpyionycteriscould be also a cavity dweller, as are manymembers of the harpyionycterine clade(fig. 10). It is interesting that the harpyionyc-terine clade appeared in our analysis as sisterto the rousettine-epomophorine clade (withhigh support in our analysis, BS 5 21). Takentogether, these two major megabat cladesshare a common ancestor that is also recon-structed as a cavity dweller (fig. 10). Onceagain, limited taxonomic sampling preventsfurther speculation, but we risk the hypothesisthat cavity dwelling may have had a majorimpact in the diversification of megabats.
CONCLUSIONS
Our results strongly confirm monophyly ofa harpyionycterine clade inclusive of Har-pyionycteris, Dobsonia, Aproteles, and Boneia(the latter previously included in Rousettus asa subgenus). Accordingly, we recognizeBoneia as a distinct genus and recommend
Bulletin of the American Museum of Natural History amnb-582-00-06.3d 24/9/09 15:58:11 196 Cust # 582-6
TABLE 2Distribution of Each of the 18 Species of Harpyionycterinae Miller, 1907
Biogeographic regions follow Corbet and Hill (1992).
Species Distribution Biogeographic regions
Aproteles bulmerae Papua New Guinea (highlands) Papuan
Boneia bidens Northern Sulawesi Wallacean (Sulawesi Division)
Dobsonia anderseni Bismarck Arch., Admiralty Isls. Papuan
Dobsonia beauforti Waigeo, Batanta, Salawati, Gebe,
Gag, and Biak Isls.
Papuan, Wallacean (Moluccan Division)
Dobsonia chapmani Cebu and Negros Isls. Philippines
Dobsonia crenulata N Moluccas, Togian, Sangihe,
Talaud, and Peleng Isls., Sulawesi
Wallacean (Sulawesi and Moluccan Divisions)
Dobsonia emersa Biak, Numfoor, and Owii Isls. Papuan
Dobsonia exoleta Sulawesi, Muna, Togian, Sula Isls. Wallacean (Sulawesi Division)
Dobsonia inermis Solomon Islands Solomons
Dobsonia minor Lowlands of New Guinea and
adjacent Isls., Sulawesi
Papuan, Wallacean (Sulawesi Division)
Dobsonia moluccensis Moluccas, New Guinea and
land-bridge islands, to Queensland
Papuan, Wallacean (Moluccan Division),
Australian
Dobsonia pannietensis Louisiade Arch., D’Entrecasteaux
Isls., Trobriand Isls.
Papuan
Dobsonia peronii Lesser Sunda Islands (Bali East to
Babar Isls.)
Sundaic (Javan Division) and Wallacean
(Lesser Sunda and Moluccan Divisions)
Dobsonia praedatrix Bismarck Arch. Papuan
Dobsonia viridis C and S Moluccas, Banda, and
Kai Isls.
Wallacean (Moluccan Division)
Harpyionycteris celebensis Sulawesi Wallacean (Sulawesi Division)
Harpyionycteris whiteheadi Philippines Philippine
196 BULLETIN AMERICAN MUSEUM OF NATURAL HISTORY NO. 582
Bulletin of the American Museum of Natural History amnb-582-00-06.3d 24/9/09 15:58:12 197 Cust # 582-6
Fig. 9. Joint distribution of species in harpyionycterine megabat genera in the Austromalayan Region(A), and biogeographic patterns in harpyionycterine megabats (B). References for distributions: Aproteles(dots), Boneia (hatched area), Dobsonia (perimeter lines), and Harpyionycteris (shaded areas). Indicated arethe Wallace, Weber, and Lydekker lines. Arrows indicate the distribution of Dobsonia species that escapethe boundaries of the Wallacean and Papuan subregions: (A) Dobsonia chapmani in Negros and CebuIslands (Philippine subregion); (B) Dobsonia peronii in Bali (Sundaic subregion, Javan Division; D. peroniiis widespread in the Lesser Sunda Islands); (C) Dobsonia moluccensis in Queensland, Australia (D.moluccensis, inclusive of D. magna, is widespread in the Moluccas and New Guinea). Biogeographicpatterns: Each species is assigned one or more of the following areas, each of which is a state in a singleunordered biogeographic character: AUS Australia, LSI Lesser Sunda Islands, NGU New Guinea, PHIPhilippines, SOL Solomon Islands, SUL Sulawesi. Biogeographic events are inferred by partial (downpassonly) optimization of distribution areas of species in the harpyionycterine clade in the optimal tree fromthe most complete, combined analysis (B; see fig. 4).
2009 GIANNINI ET AL.: HARPYIONYCTERINE PHYLOGENY 197

expanding Harpyionycterinae Miller, 1907,to include these four genera. Although nomorphological synapomorphies are apparentfor this grouping, some morphological andecological patterns are interesting, for in-stance, dental-formula patterns and theprediction that Harpyionycteris may be acavity-dwelling bat. Also inferred from our
phylogenetic hypothesis, we showed thatmajor biogeographic boundaries within theAustromalayan region play a key role inexplaining patterns of dispersal-vicarianceevents in harpyionycterine bats. These resultsand preliminary interpretations encouragethe undertaking of a full-scale phylogeny ofMegachiroptera.
Bulletin of the American Museum of Natural History amnb-582-00-06.3d 24/9/09 15:58:15 198 Cust # 582-6
Fig. 10. Reconstruction of roosting habits (cavity versus foliage) in the most parsimonious tree fromthe total dataset of this study (fig. 4). Hatched lines indicate ambiguity (in this case, terminals for whichboth roosting habits were documented). Cavity dwelling is indicated in thick branches. Question marksindicate lack of data for Harpyionycteris. Note that this genus is nested in a clade of cavity dwellers(see text).
198 BULLETIN AMERICAN MUSEUM OF NATURAL HISTORY NO. 582

ACKNOWLEDGMENTS
We are indebted with Rob DeSalle (Sack-ler Institute for Comparative Genomics,American Museum of Natural History) forproviding laboratory space and reagents forthe molecular part of this work. We thankLawrence Heaney (Field Museum of NaturalHistory, Chicago), Jim Patton and CarlaCicero (Museum of Vertebrate Zoology,Berkeley), John Wible and Suzanne McLaren(Carnegie Museum, Pittsburgh), DenisO’Meally (Australian Museum), Jeremy Ja-cobs, Louise Emmons, and James Mead(Smithsonian Institution, National Museumof Natural History, Washington DC), andWim Bergmans (ZMA Zoologisch Museum,Amsterdam) for access to tissue samples thatmade possible this contribution. We alsothank Jan Haft for photographs an addition-al information. Finally, our appreciation toDeanna P. Byrnes for her generosity inproviding two unpublished sequences foruse in this study. Funding for this reportwas provided by the National Science Foun-dation (research grant DEB-9873663 toN.B.S.), Coleman and Vernay postdoctoralfellowships at the AMNH to N.P.G., aHenry MacCracken doctoral fellowship atNew York University to F.C.A., and aVernay postdoctoral fellowship at theAMNH to F.C.A.
REFERENCES
Alcala, E.L., R.B. Paalan, L.T. Averia, and A.C.
Alcala. 2004. Rediscovery of the Philippine
bare-backed fruit bat (Dobsonia chapmani Ra-
bor) on southwestern Negros Island, the Phil-
ippines. Silliman Journal 45(2): 126–136.
Andersen, K. 1912. Catalogue of the Chiroptera in
the collection of the British Museum. London:
Megachiroptera British Museum (Natural His-
tory).
Bastian, S.T., Jr., K. Tanaka, R.V. Anunciado,
N.G. Natural, A.C. Sumalde, and T. Nami-
kawa. 2002. Evolutionary relationships of flying
foxes (genus Pteropus) in the Philippines in-
ferred from DNA sequences of cytochrome b
gene. Biochemical Genetics 40: 101–116.
Bergmans, W. 1994. Taxonomy and biogeography
of African fruit bats (Mammalia, Megachirop-
tera). 4. The genus Rousettus Gray, 1821.
Beaufortia 44(4): 79–126.
Bergmans, W. 1997. Taxonomy and biogeography
of African fruit bats (Mammalia, Megachirop-
tera). 5. The genera Lissonycteris Andersen,
1912, Myonycteris Matschie, 1899, and Mega-
loglossus Pagenstecher, 1885. General remarks
and conclusions. Annex: key to all species.
Beaufortia 47: 11–90.
Bergmans, W., and F.G. Rozendaal. 1988. Notes
on collections of fruit bats from Sulawesi and
some off-lying islands (Mammalia, Megachir-
optera). Zoologische Verhandelingen 248: 1–74.
Boeadi, B., and W. Bergmans. 1987. First record
of Dobsonia minor (Dobson, 1879) from Sula-
wesi, Indonesia (Mammalia, Megachiroptera).
Bulletin Zoologisch Museum Universiteit van
Amsterdam 11: 69–74.
Bonaccorso, F.J. 1998. Bats of Papua New
Guinea. Washington, DC: Conservation Inter-
national.
Bremer, K. 1994. Branch support and tree
stability. Cladistics 10: 295–304.
Byrnes, D.G.P. 2005. Evolution among the bare-
backed fruit bats, Dobsonia (Chiroptera, Pter-
opodidae). Unpublished Ph.D. dissertation.
University of Wisconsin, Madison.
Chiroptera Specialist Group, Dobsonia peroni. In
International Union for Conservation of Nature
and Natural Resources 2007. 2007 IUCN red
list of threatened species (downloaded 21
August 2008 from http://www.iucnredlist.org/).
Colgan, D.J., and P. da Costa. 2002. Megachir-
opteran evolution studied with 12S rDNA and
c-mos DNA sequences. Journal of Mammalian
Evolution 9: 3–22.
Corbet, G.B., and J.E. Hill. 1992. Mammals of the
Indomalayan region: a systematic review. Lon-
don: Oxford University Press. 2 vols, 488 pp.
Eick, G.N., D.S. Jacobs, and C.A. Matthee. 2005.
A nuclear DNA phylogenetic perspective on the
evolution of echolocation and historical bioge-
ography of extant bats (Chiroptera). Molecular
Biology and Evolution 22: 1869–1886.
Flannery, T.F. 1995. Mammals of Papua New
Guinea. Sydney: Robert Brown and Associa-
tion, 439 pp.
Fitch, W.M. 1971. Toward defining the course of
evolution: minimum change for a specific tree
topology. Systematic Zoology 20: 406–416.
Giannini, N.P., F.C. Almeida, N.B. Simmons, and
R. DeSalle. 2006. Phylogenetic relationships of
the enigmatic Harpy Fruit Bat, Harpyionycteris
(Mammalia: Chiroptera: Pteropodidae). Amer-
ican Museum Novitates 3533: 1–12.
Giannini, N.P., F.C. Almeida, N.B. Simmons, and
K.M. Helgen. 2008. The systematic position of
Pteropus leucopterus and its bearing on the
monophyly and relationships of Pteropus (Chi-
Bulletin of the American Museum of Natural History amnb-582-00-06.3d 24/9/09 15:58:20 199 Cust # 582-6
2009 GIANNINI ET AL.: HARPYIONYCTERINE PHYLOGENY 199

roptera: Pteropodidae). Acta Chiropterologica10: 11–20.
Giannini, N.P., and S. Bertelli. 2004. A phylogenyof extant penguins based on integumentarycharacters. Auk 121: 422–434.
Giannini, N.P., and N.B. Simmons. 2003. Aphylogeny of megachiropteran bats (Mammalia:Chiroptera: Pteropodidae) based on directoptimization analysis of one nuclear and fourmitochondrial genes. Cladistics 19: 496–511.
Giannini, N.P., and N.B. Simmons. 2005. Conflictand congruence in a combined DNA-morphol-ogy analysis of megachiropteran bat relation-ships (Mammalia: Chiroptera: Pteropodidae).Cladistics 21: 411–437.
Giannini, N.P., and N.B. Simmons. 2007. ElementHomology and the Evolution of Dental For-mulae in Megachiropteran Bats (Mammalia:Chiroptera: Pteropodidae). American MuseumNovitates 3559: 1–27.
Goloboff, P.A. 1999. Analyzing large data sets inreasonable times: solutions for composite opti-ma. Cladistics 15: 415–428.
Goloboff, P.A., J.S. Farris, M. Kallersjo, B.Oxelman, M. Ramırez, and C. Szumik. 2003.Improvements to resampling measures of groupsupport. Cladistics 19: 324–332.
Goloboff, P.A., J.S. Farris, and K. Nixon. 2004.T.N.T.: tree analysis using new technologies.Program and documentation (available at www.cladistics.org).
Goloboff, P.A., J.S. Farris, and K.C. Nixon. 2008.TNT, a free program for phylogenetic analysis.Cladistics 24(5): 774–786.
Heaney, L.R., D.S. Balete, M.L. Dolar, A.C.Alcala, A.T.L. Dans, P.C. Gonzales, N.R. Ingle,M.V. Lepiten, W.L.R. Oliver, P.S. Ong, E.A.Rickart, B.R. Tabaranza, Jr., and R.C.B.Utzurrum. 1998. A synopsis of the mammalianfauna of the Philippine Islands. FieldianaZoology New Series 88: 1–61.
Helgen, K.M. 2007. A taxonomic and geographicoverview of the mammals of Papua. In A.J.Marshall and B.M. Beehler (editors), Ecology ofIndonesia series. Vol. 6. Part 1, The ecology ofPapua: 689–749. Singapore: Periplus Editions.
Jones, K.E., A. Purvis, A. MacLarnon, O.R.P.Bininda-Emonds, and N.B. Simmons. 2002. Aphylogenetic supertree of the bats (Mammalia:Chiroptera). Biological Review 77: 223–259.
Kirsch, J.A., T.F. Flannery, M.S. Springer, andF.-J. Lapointe. 1995. Phylogeny of the Pter-opodidae (Mammalia: Chiroptera) based onDNA hybridization with evidence of bat mono-phyly. Australian Journal of Zoology 43:395–428.
Koopman, K.F. 1993. Order Chiroptera. In D.E.Wilson and D.M. Reeder (editors), Mammal
species of the world: a taxonomic and geo-graphic reference. 2nd ed.: 137–241. Washing-ton, DC: Smithsonian Institution Press.
Koopman, K.F. 1994. Chiroptera: systematics. InHandbook of zoology, vol. 8, part 60, Mam-malia: 1–217. Berlin: Walter de Gruyter.
Kunz, T.H., and E.D. Pierson. 1994. Bats of theworld: an introduction. In R.M. Nowak (edi-tor), Walker’s bats of the World: 1–46. Balti-more, MD: Johns Hopkins University Press.
Menzies, J.I. 1977. Fossil and subfossil bats fromthe mountains of New Guinea. AustralianJournal of Zoology 25: 329–336.
Miller, G.S., Jr. 1907. The families and genera ofbats. Bulletin of the United States NationalMuseum 57: 1–282.
Miller, G.S., Jr., and N. Hollister. 1921. Twentynew mammals collected by H. Raven in Celebes.Proceedings of the Biological Society of Wash-ington 34: 93–104.
Miller-Butterworth, C.M., W.J. Murphy, S.J.O’Brien, D.S. Jacobs, M.S. Springer, and E.C.Teeling. 2007. A family matter: conclusiveresolution of the taxonomic position of thelong-fingered bats, Miniopterus. Molecular Bi-ology and Evolution 24: 1553–1561.
Paguntalan, L.M.J., M.dG. Pedregosa, and M.J.C.Gadiana. 2004. The Philippine bare-backed fruitbat Dobsonia chapmani Rabor, 1952: rediscov-ery and conservation status on Cebu Island.Silliman Journal 45(2): 114–122.
Porter, C.A., M. Goodman, and M.J. Stanhope.1996. Evidence on mammalian phylogeny fromsequences of exon 28 of the von Willerbrandfactor gene. Molecular Phylogenetics and Evo-lution 5: 89–101.
Romagnoli, M.L., and M.S. Springer. 2000.Evolutionary relationships among Old Worldfruitbats (Megachiroptera: Pteropodidae) basedon 12S rRNA, tRNA valine and 16S rRNAgene sequences. Journal of Mammalian Evolu-tion 7: 259–284.
Ronquist, F. 1997. Dispersal–vicariance analysis: anew approach to the quantification of historicalbiogeography. Systematic Biology 46: 195–203.
Simmons, N.B. 2005. Order Chiroptera. In D.E.Wilson and D.M. Reeder (editors), Mammalspecies of the world: a taxonomic and geo-graphic reference, 3rd ed.: 312–529. Baltimore,MD: Johns Hopkins University Press.
Slaughter, B.H. 1970. Evolutionary trends inchiropteran dentitions. In B.H. Slaughter andW.D. Walton (editors), About bats: 51–83.Dallas, TX: Southern Methodist UniversityPress.
Springer, M.S., L.J. Hollar, and J.A. Kirsch. 1995.Phylogeny, molecules versus morphology andrates of character evolution among fruit bats
Bulletin of the American Museum of Natural History amnb-582-00-06.3d 24/9/09 15:58:20 200 Cust # 582-6
200 BULLETIN AMERICAN MUSEUM OF NATURAL HISTORY NO. 582

(Chiroptera: Megachiroptera). Australian Jour-nal of Zoology 43: 557–582.
Stamatakis, A. 2006. RAxML-VI-HPC: Maxi-mum likelihood based phylogenetic analyseswith thousands of taxa and mixed models.Bioinformatics 22: 2688–2690.
Stamatakis, A., P. Hoover, and J. Rougemont.2008. A rapid bootstrap algorithm for theRAxML web servers. Systematic Biology57(5): 758–771.
Tate, G.H.H. 1951. Harpyionycteris, a genus of rarefruit bats. American Museum Novitates 1522:1–9.
Teeling, E.C., M. Scally, D. Kao, M. Romagnoli,M.S. Springer, and M.J. Stanhope. 2000. Mo-lecular evidence regarding the origin of echolo-cation and flight in bats. Nature 403: 188–192.
Teeling, E.C., M.S. Springer, O. Madsen, P. Bates,S.J. O’Brien, and W.J.A. Murphy. 2005. Amolecular phylogeny for bats illuminates bioge-ography and the fossil record. Science 307:580–584.
Thomas, O. 1896. Mammals from Celebes, Bor-neo, and the Philippines recently received at theBritish Museum. Annals and Magazine ofNatural History 18(6): 241–250.
Varon, A., L.S. Vinh, I. Bomash, and W.C.Wheeler. 2007. POY 4.0 Beta 2398. New York:American Museum of Natural History (avail-able at http://research.amnh.org/scicomp/projects/poy.php)
Wheeler, W.C. 1996. Optimization alignment: theend of multiple sequence alignment in phyloge-netics? Cladistics 12: 1–9.
Bulletin of the American Museum of Natural History amnb-582-00-06.3d 24/9/09 15:58:20 201 Cust # 582-6
2009 GIANNINI ET AL.: HARPYIONYCTERINE PHYLOGENY 201
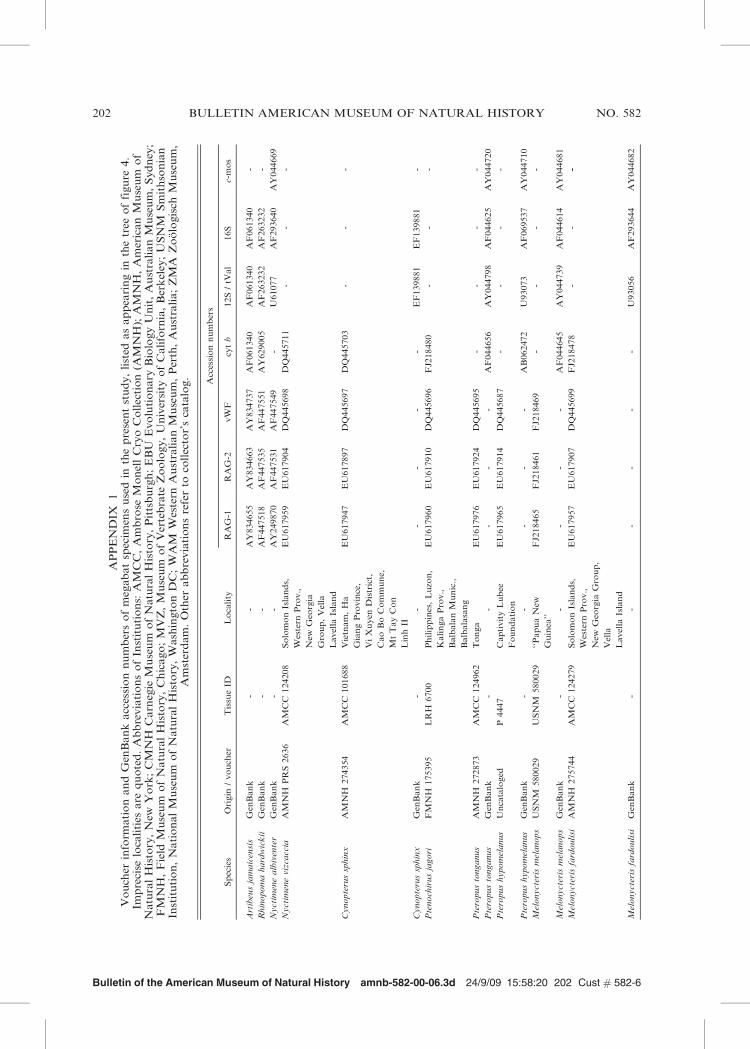
Bulletin of the American Museum of Natural History amnb-582-00-06.3d 24/9/09 15:58:20 202 Cust # 582-6
AP
PE
ND
IX1
Vo
uch
erin
form
ati
on
an
dG
enB
an
ka
cces
sio
nn
um
ber
so
fm
egab
at
spec
imen
su
sed
inth
ep
rese
nt
stu
dy
,li
sted
as
ap
pea
rin
gin
the
tree
of
figu
re4.
Imp
reci
selo
cali
ties
are
qu
ote
d.
Ab
bre
via
tio
ns
of
Inst
itu
tio
ns:
AM
CC
,A
mb
rose
Mo
nel
lC
ryo
Co
llec
tio
n(A
MN
H);
AM
NH
,A
mer
ican
Mu
seu
mo
fN
atu
ral
His
tory
,N
ewY
ork
;C
MN
HC
arn
egie
Mu
seu
mo
fN
atu
ral
His
tory
,P
itts
bu
rgh
;E
BU
Evo
luti
on
ary
Bio
logy
Un
it,
Au
stra
lia
nM
use
um
,S
yd
ney
;F
MN
H,
Fie
ldM
use
um
of
Na
tura
lH
isto
ry,
Ch
ica
go
;M
VZ
,M
use
um
of
Ver
teb
rate
Zo
olo
gy,
Un
iver
sity
of
Cali
forn
ia,
Ber
kel
ey;
US
NM
Sm
ith
son
ian
Inst
itu
tio
n,
Na
tio
nal
Mu
seu
mo
fN
atu
ral
His
tory
,W
ash
ingto
nD
C;
WA
MW
este
rnA
ust
rali
an
Mu
seu
m,
Per
th,
Au
stra
lia
;Z
MA
Zo
olo
gis
chM
use
um
,A
mst
erd
am
.O
ther
ab
bre
via
tio
ns
refe
rto
coll
ecto
r’s
cata
log.
Sp
ecie
sO
rig
in/
vo
uch
erT
issu
eID
Lo
cali
ty
Acc
essi
on
nu
mb
ers
RA
G-1
RA
G-2
vW
Fcy
tb
12
S/
tVal
16S
c-m
os
Art
ibeu
sja
ma
icensi
sG
enB
an
k-
-A
Y8
34
655
AY
83
46
63
AY
83
47
37
AF
06
134
0A
F06
13
40
AF
06
13
40
-
Rh
inop
om
ah
ard
wic
kii
Gen
Ba
nk
--
AF
447
518
AF
44
75
35
AF
44
75
51
AY
62
900
5A
F26
32
32
AF
26
32
32
-
Nycti
mene
alb
ivente
rG
enB
an
k-
-A
Y2
49
870
AF
44
75
31
AF
44
75
49
-U
61
07
7A
F29
36
40
AY
0446
69
Nycti
mene
viz
caccia
AM
NH
PR
S26
36
AM
CC
12
420
8S
olo
mo
nIs
lan
ds,
Wes
tern
Pro
v.,
New
Geo
rgia
Gro
up
,V
ella
Lav
ella
Isla
nd
EU
61
79
59
EU
61
79
04
DQ
44
56
98
DQ
445
71
1-
--
Cy
no
pte
rus
sph
inx
AM
NH
274
354
AM
CC
10
168
8V
ietn
am
,H
a
Gia
ng
Pro
vin
ce,
Vi
Xu
yen
Dis
tric
t,
Cao
Bo
Co
mm
un
e,
Mt
Tay
Co
n
Lin
hII
EU
61
79
47
EU
61
78
97
DQ
44
56
97
DQ
445
70
3-
--
Cy
no
pte
rus
sph
inx
Gen
Ba
nk
--
--
--
EF
13
98
81
EF
13
98
81
-
Pte
no
chir
us
jag
ori
FM
NH
175
395
LR
H6
700
Ph
ilip
pin
es,
Lu
zon
,
Kali
ng
aP
rov
.,
Balb
ala
nM
un
ic.,
Balb
ala
san
g
EU
61
79
60
EU
61
79
10
DQ
44
56
96
FJ2
18
48
0-
--
Pte
ropu
sto
ng
anu
sA
MN
H2
72
873
AM
CC
12
496
2T
on
ga
EU
61
79
76
EU
61
79
24
DQ
44
56
95
--
--
Pte
ropu
sto
ng
anu
sG
enB
an
k-
--
--
AF
04
465
6A
Y0
447
98
AF
04
46
25
AY
0447
20
Pte
ropu
shyp
om
ela
nus
Un
cata
log
edP
444
7C
ap
tiv
ity
Lu
bee
Fo
un
da
tio
n
EU
61
79
65
EU
61
79
14
DQ
44
56
87
--
--
Pte
ropu
shyp
om
ela
nus
Gen
Ba
nk
--
--
-A
B06
247
2U
93
07
3A
F06
95
37
AY
0447
10
Melo
ny
cte
ris
mela
no
ps
US
NM
58
002
9U
SN
M5
800
29
‘‘P
ap
ua
New
Gu
inea
’’
FJ2
184
65
FJ2
184
61
FJ2
184
69
--
--
Melo
ny
cte
ris
mela
no
ps
Gen
Ba
nk
--
--
-A
F04
464
5A
Y0
447
39
AF
04
46
14
AY
0446
81
Melo
ny
cte
ris
fard
ouli
siA
MN
H2
75
744
AM
CC
12
427
9S
olo
mo
nIs
lan
ds,
Wes
tern
Pro
v.,
New
Geo
rgia
Gro
up
,
Vel
la
Lav
ella
Isla
nd
EU
61
79
57
EU
61
79
07
DQ
44
56
99
FJ2
18
47
8-
--
Melo
ny
cte
ris
fard
ouli
siG
enB
an
k-
--
--
-U
93
05
6A
F29
36
44
AY
0446
82
202 BULLETIN AMERICAN MUSEUM OF NATURAL HISTORY NO. 582
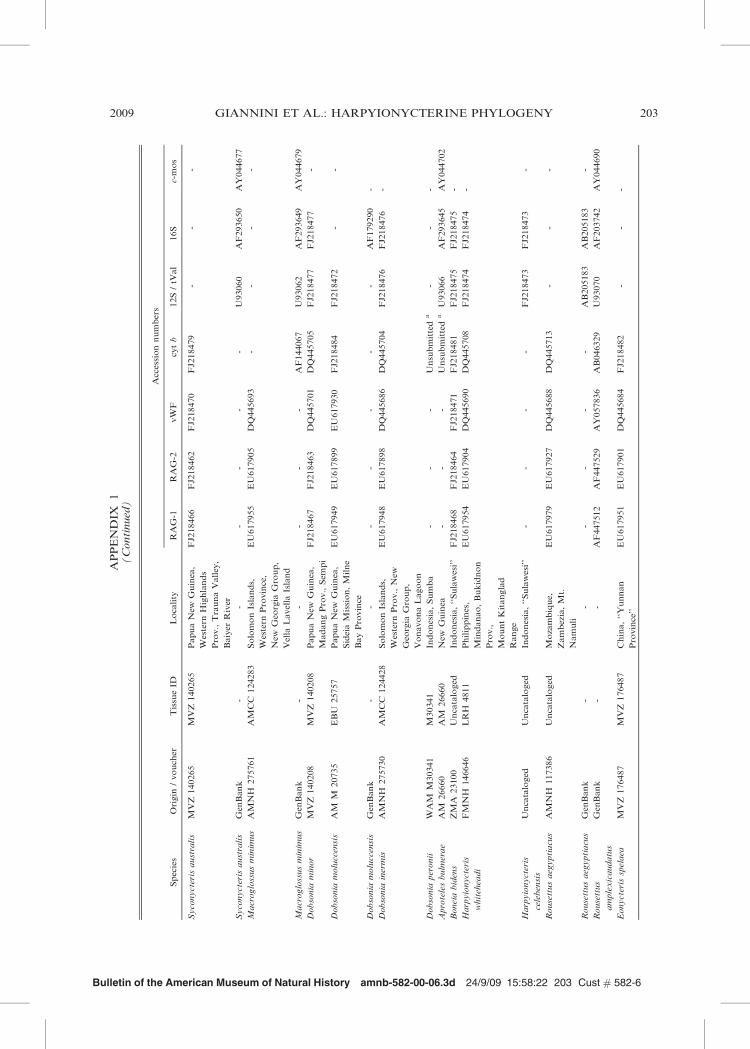
Bulletin of the American Museum of Natural History amnb-582-00-06.3d 24/9/09 15:58:22 203 Cust # 582-6
Sp
ecie
sO
rig
in/
vo
uch
erT
issu
eID
Lo
cali
ty
Acc
essi
on
nu
mb
ers
RA
G-1
RA
G-2
vW
Fcy
tb
12S
/tV
al
16S
c-m
os
Syco
nycte
ris
au
stra
lis
MV
Z14
02
65
MV
Z1
402
65
Pa
pu
aN
ewG
uin
ea,
Wes
tern
Hig
hla
nd
s
Pro
v.,
Tra
un
aV
all
ey,
Ba
iyer
Riv
er
FJ2
18
46
6F
J218
46
2F
J218
470
FJ2
18
479
--
-
Syco
nycte
ris
au
stra
lis
Gen
Ban
k-
--
--
-U
930
60
AF
2936
50
AY
044
677
Macro
glo
ssus
min
imus
AM
NH
27
576
1A
MC
C1
242
83
So
lom
on
Isla
nd
s,
Wes
tern
Pro
vin
ce,
New
Geo
rgia
Gro
up
,
Vel
laL
av
ella
Isla
nd
EU
61
79
55
EU
617
90
5D
Q4
45
693
--
--
Macro
glo
ssus
min
imus
Gen
Ban
k-
--
--
AF
14
40
67
U9
30
62
AF
2936
49
AY
044
679
Dob
son
iam
ino
rM
VZ
14
02
08
MV
Z1
402
08
Pa
pu
aN
ewG
uin
ea,
Mad
an
gP
rov.,
Sem
pi
FJ2
18
46
7F
J218
46
3D
Q4
45
701
DQ
445
705
FJ2
18
47
7F
J21
847
7-
Dob
son
iam
olu
ccensi
sA
MM
207
35
EB
U2
57
57
Pa
pu
aN
ewG
uin
ea,
Sid
eia
Mis
sio
n,
Mil
ne
Ba
yP
rov
ince
EU
61
79
49
EU
617
89
9E
U6
17
930
FJ2
18
484
FJ2
18
47
2-
-
Dob
son
iam
olu
ccensi
sG
enB
an
k-
--
--
--
AF
1792
90
-
Dob
son
iain
erm
isA
MN
H27
573
0A
MC
C1
244
28
So
lom
on
Isla
nd
s,
Wes
tern
Pro
v.,
New
Geo
rgia
Gro
up
,
Vo
nav
on
aL
ag
oo
n
EU
61
79
48
EU
617
89
8D
Q4
45
686
DQ
445
704
FJ2
18
47
6F
J21
847
6-
Dob
son
iap
ero
nii
WA
MM
30
34
1M
30
34
1In
do
nes
ia,
Su
mb
a-
--
Un
sub
mit
ted
a-
--
Ap
rote
les
bu
lmer
ae
AM
26
66
0A
M2
666
0N
ewG
uin
ea-
--
Un
sub
mit
ted
aU
930
66
AF
293645
AY
044702
Bon
eia
bid
ens
ZM
A2
310
0U
nca
talo
ged
Ind
on
esia
,‘‘
Su
law
esi’
’F
J218
46
8F
J218
46
4F
J218
471
FJ2
18
481
FJ2
18
47
5F
J21
847
5-
Harp
yio
ny
cte
ris
wh
itehead
i
FM
NH
14
66
46
LR
H4
81
1P
hil
ipp
ines
,
Min
da
nao
,B
uk
idn
on
Pro
v.,
Mo
un
tK
ita
ng
lad
Ran
ge
EU
61
79
54
EU
617
90
4D
Q4
45
690
DQ
445
708
FJ2
18
47
4F
J21
847
4-
Harp
yio
ny
cte
ris
cele
ben
sis
Un
cata
log
edU
nca
talo
ged
Ind
on
esia
,‘‘
Su
law
esi’
’-
--
-F
J21
847
3F
J21
847
3-
Ro
use
ttu
sa
egy
pti
acu
sA
MN
H11
738
6U
nca
talo
ged
Mo
zam
biq
ue,
Za
mb
ezia
,M
t.
Nam
uli
EU
61
79
79
EU
617
92
7D
Q4
45
688
DQ
445
713
--
-
Ro
use
ttu
sa
egy
pti
acu
sG
enB
an
k-
--
--
-A
B205
18
3A
B2051
83
-
Ro
use
ttu
s
am
ple
xic
au
da
tus
Gen
Ban
k-
-A
F44
751
2A
F44
752
9A
Y0
57
83
6A
B04
63
29
U9
30
70
AF
2037
42
AY
044
690
Eon
ycte
ris
spela
eaM
VZ
17
64
87
MV
Z1
764
87
Ch
ina
,‘‘
Yu
nn
an
Pro
vin
ce’’
EU
61
79
51
EU
617
90
1D
Q4
45
684
FJ2
18
482
--
-
AP
PE
ND
IX1
(C
on
tin
ued)
2009 GIANNINI ET AL.: HARPYIONYCTERINE PHYLOGENY 203
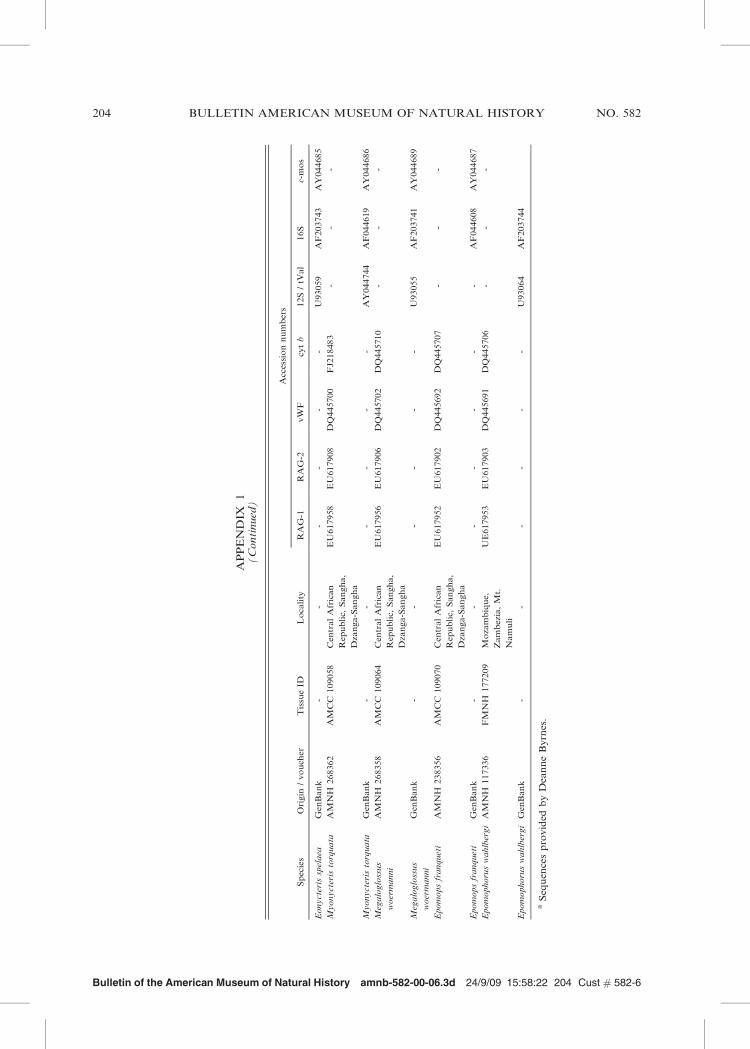
Bulletin of the American Museum of Natural History amnb-582-00-06.3d 24/9/09 15:58:22 204 Cust # 582-6
Sp
ecie
sO
rig
in/
vo
uch
erT
issu
eID
Lo
cali
ty
Acc
essi
on
nu
mb
ers
RA
G-1
RA
G-2
vW
Fcy
tb
12S
/tV
al
16
Sc-
mo
s
Eony
cte
ris
spel
aea
Gen
Ban
k-
--
--
-U
93059
AF
203743
AY
04468
5
My
ony
cte
ris
torq
ua
taA
MN
H2
68
36
2A
MC
C1
09
058
Cen
tral
Afr
ican
Rep
ub
lic,
Sa
ng
ha,
Dza
ng
a-S
an
gh
a
EU
617
958
EU
61
79
08
DQ
44
570
0F
J21
848
3-
--
My
ony
cte
ris
torq
ua
taG
enB
an
k-
--
--
-A
Y044744
AF
044619
AY
04468
6
Meg
alo
glo
ssus
woerm
anni
AM
NH
26
835
8A
MC
C1
09
064
Cen
tral
Afr
ican
Rep
ub
lic,
Sa
ng
ha,
Dza
ng
a-S
an
gh
a
EU
617
956
EU
61
79
06
DQ
44
570
2D
Q44
571
0-
--
Meg
alo
glo
ssus
woerm
anni
Gen
Ban
k-
--
--
-U
93055
AF
203741
AY
04468
9
Epom
ops
fran
queti
AM
NH
23
835
6A
MC
C1
09
070
Cen
tral
Afr
ican
Rep
ub
lic,
Sa
ng
ha,
Dza
ng
a-S
an
gh
a
EU
617
952
EU
61
79
02
DQ
44
569
2D
Q44
570
7-
--
Epom
ops
fran
queti
Gen
Ban
k-
--
--
--
AF
044608
AY
04468
7
Epom
oph
oru
sw
ah
lber
gi
AM
NH
11
733
6F
MN
H1
77
209
Mo
zam
biq
ue,
Zam
bez
ia,
Mt.
Nam
uli
UE
617
953
EU
61
79
03
DQ
44
569
1D
Q44
570
6-
--
Epom
oph
oru
sw
ah
lber
gi
Gen
Ban
k-
--
--
-U
93
06
4A
F20
37
44
aS
equ
ence
sp
rov
ided
by
Dea
nn
eB
yrn
es.
AP
PE
ND
IX1
(C
on
tin
ued)
204 BULLETIN AMERICAN MUSEUM OF NATURAL HISTORY NO. 582
Top Related
Copyright © 2022 FDOKUMEN

