







Bahasa
Halaman
Hukum


Journal of Photochemistry and Photobiology B: Biology 69 (2003) 107–120www.elsevier.com/ locate/ jphotobiol
C ellular uptake, localization and photodynamic effects of haematoporphyrinderivative in human glioma and squamous carcinoma cell lines
a a , b c*Seema Gupta , B.S. Dwarakanath , K. Muralidhar , V. JainaDepartment of Biocybernetics, Institute of Nuclear Medicine and Allied Sciences, Brig SK Mazumdar Road, Timarpur, Delhi 110054,India
bDepartment of Zoology, University of Delhi, Delhi 110007,IndiacWallace-Kettering Neuroscience Institute, Kettering Medical Center and Department of Emergency Medicine, Wright State University,
Dayton, OH 45429,USA
Received 22 February 2002; received in revised form 16 October 2002; accepted 20 November 2002
Abstract
Uptake, intracellular concentration, localization and photodynamic effects of a haematoporphyrin derivative (HpD, Photosan-3) werecompared in human glioma (BMG-1, wild-type p53) and squamous carcinoma (4451, mutated p53) cell lines. Concentration and timedependence of cellular uptake of HpD was assayed from methanol extracts and whole cell suspension spectroscopy, while localizationwas studied by fluorescence microscopy-based image analysis. Colony-forming ability, apoptosis, cell-cycle progression and cytogeneticdamage (micronuclei formation) were investigated as parameters of photodynamic response following irradiation with red light. BMG-1cells were more sensitive to the photodynamic treatment than 4451 cells, although the 4451 cells accumulated a higher amount of HpDand did not differ significantly from BMG-1 cells with respect to intracellular localization. Photodynamically-induced cytogenetic damageand apoptosis were considerably higher in BMG-1 cells as compared to 4451 cells. The present results strongly suggest that manifestationof the photodynamically-induced lesions in the form of cytogenetic damage and apoptosis are among the important determinants ofcellular sensitivity to HpD–PDT besides the photodynamic dose (intracellular concentration of the photosensitizer and the light dose). 2002 Elsevier Science B.V. All rights reserved.
Keywords: Haematoporphyrin derivative; Photodynamic therapy; Intracellular localization; Cytogenetic damage; Apoptosis; p53
1 . Introduction able commercially in a partially purified form (di-haematoporphyrinester /ether) under the trade name Photo-
Photodynamic diagnosis (PDD) and photodynamic frin has been approved for specific clinical applications intherapy (PDT) are promising new modalities shown to be several countries in Europe, America and Asia. Thougheffective in the early detection and treatment of several Photofrin, being a complex mixture containing monomers,types of malignant and non-malignant diseases [1]. Both dimers and oligomers, is not an ideal photosensitizer sincethese technologies depend on the preferential accumulation it may cause skin phototoxicity in some patients lastingof a light-activated dye (photosensitizer) in the pathologic several weeks, it is being used at present in the manage-lesion/cells. The fact that certain porphyrin-based com- ment of cancers of lung, esophagus, bladder and skin. Thepounds accumulate preferentially in tumors and can there- use of porphyrin-based photosensitizers in the diagnosisfore be used for the detection and therapy of tumors has and therapy of malignant tumors of the brain and head andbeen known for several decades [2,3]. In recent years, a neck region is under active investigation [4,5]. To promotelarge number of new photosensitizers (second generation) judicious and optimal clinical applications of PDD andhave been shown to preferentially accumulate in hyper- PDT, the complex mechanisms underlying these promisingproliferative and malignant lesions and are currently under techniques need to be better understood at various levels.investigation in basic and preclinical studies. A few are Upon irradiation with light matching its absorptionbeing evaluated in clinical studies. The haematoporphyrin characteristics, the photosensitizer is excited to a higherderivative (HpD), initially prepared by Lipson, now avail- energy singlet state. From the excited singlet energy state,
the return to ground state can occur by the emission oflight (fluorescence) or by dissipation of energy as heat. In*Corresponding author. Fax:191-11-391-9509.
E-mail address: [email protected](B.S. Dwarakanath). PDD, emission of fluorescence is utilized to image and
1011-1344/02/$ – see front matter 2002 Elsevier Science B.V. All rights reserved.doi:10.1016/S1011-1344(02)00408-6

108 S. Gupta et al. / Journal of Photochemistry and Photobiology B: Biology 69 (2003) 107–120
localize the tumor [2]. Clinically useful applications of and/or necrosis depending on the cell type, the target forPDD include intra-operative fluorescence guided surgical photosensitization, concentration and intracellular distribu-removal of malignant cells [6,7] besides early detection of tion of the sensitizer [31], the incubation conditions andpremalignant and malignant lesions in the bladder, breast, the light dose [19]. PDT has been shown to induce cell-gastro-intestinal tract, head and neck, lung and skin [8,9] cycle arrest and apoptosis in a variety of cells [32–34].using microscopic and endoscopic techniques. Tumor suppressor gene p53 is known to influence both
In contrast to PDD, photophysical and photochemical these processes and the treatment responses of tumors.processes involving interactions of the excited photosensit- Conflicting results have been, however, reported in recentizer with the surrounding molecules are responsible for studies designed to investigate the correlations between thePDT. In PDT, the excited singlet energy state of the p53 status and PDT-mediated apoptosis and treatmentphotosensitizer comes down to the longer lived triplet state responses in cancer cells [35–37]. The relative contribu-by intersystem crossing. Subsequent energy/charge trans- tions of the various factors in the manifestation of thefer processes from the triplet state to the surrounding photodynamic response remain yet to be clearly elucidatedmolecules can induce photosensitized chemical reactions. [29].In the presence of oxygen, reactive oxygen species (ROS) In the present study, we have investigated the relation-such as singlet oxygen and hydroxyl radicals are produced, ships between cellular uptake, localization and variouswhich can damage the biomolecules, cause alterations in photodynamic effects (on cell growth, colony formingfunctions of cell constituents and eventually lead to cell ability, cytogenetic damage and apoptosis) induced by adeath through a network of biochemical and metabolic haematoporphyrin derivative, HpD, in two human tumorpathways. The effects produced by photosensitized re- cell lines differing in their origin viz. a squamous cellactions involving oxygen have been termed as photo- carcinoma and a malignant glioma which also differ indynamic action [10]. The mechanisms underlying photo- their p53 status. Important results from these studies havedynamic action in cells and tissues are complex and been presented [38,39].depend upon multiple factors.
Uptake and intracellular distribution of the photosensit-izer in addition to its photophysical and photochemical 2 . Materials and methodscharacteristics are important in determining the efficacy ofboth PDD and PDT. Presence of the optimal amount of 2 .1. Tumor cell linessensitizer in the vicinity of the sensitive target in a cell isan important determinant of photodynamic efficacy since Human cerebral glioma cell line (BMG-1; DNA index5
singlet oxygen, the most important ROS generated by the 0.95; wild-type p53), established from a mixed glioma [40]interaction of sensitizer and light has a short life-time in and a human squamous carcinoma cell line (4451; DNAwhich it can travel only a short distance [11]. The uptake index51.50; mutated p53) established from an oral cavityand intracellular localization of the sensitizer are influ- lesion [41] were used in the present studies.enced by the physico-chemical properties of the sensitizer Monolayer BMG-1 cells were grown in DMEM with(charge distribution, lipophilicity, aggregation state etc.) as 5% fetal calf serum (FCS) and 4451 cells with DMEMwell as the cell type and its physiological state and containing 10% FCS, penicillin (100 units /ml), strep-microenvironment [12–16]. tomycin (50mg/ml) and nystatin (2mg/ml). Stock
Depending upon the photosensitizer used, photodynamic cultures were passaged every third day (BMG-1) or secondtreatment has been shown to induce lesions in membranes day (4451) after harvesting the cells with 0.05% trypsin
3 2[17–19], mitochondria [20–22], lysosomes [23–25] and/or and seeding 8310 cells /cm in tissue culture flasks toother essential biomolecules or organelles including DNA maintain the cells in the exponential phase. All experi-[26,27]. Cell nucleus, however, is a relatively less im- ments were carried out in exponentially growing cells.portant target for photodynamic cytotoxicity than mem-branes [28]. Even though specific damage to cellular 2 .2. Chemicalsstructures or functions has been observed at variousintracellular sites, a clear causal relationship between the The haematoporphyrin derivative (HpD) used in theselesions and cell death has not emerged so far [29]. studies was Photosan-3 (PS-3), a commercial preparation
The cellular sensitivity to PDT differs a great deal (SeeLab, Germany). Hank’s Balanced Salt Solutionbetween various cell types. Cells differing in their origin (HBSS), Dulbecco’s modified Phosphate Buffered Salineand radiosensitivity have been shown to respond different- (PBS), Dulbecco’s Modified Eagle’s Medium (DMEM),ly to PDT [28,30]. Cellular and tissue responses to PDT- fetal calf serum (FCS), (N-[2-hydroxyethyl] piperazine-N9-induced oxidative stress involve complex signal transduc- [2-ethanesulfonic acid]) (HEPES) buffer, propidium iodidetion pathways and alterations in gene expression leading (PI), 4,6-diamidino-2-phenyl indole (DAPI), Ribonu-either to repair of induced lesions, altered cell functions or clease-A (RNase-A), Hoechst-H33342 (bis benzimide (29-to cell death. Cell death may occur either by apoptosis [4-ethoxyphenyl]-5-[4-methyl-1-piperazinyl]-2,59-bi-1H-

S. Gupta et al. / Journal of Photochemistry and Photobiology B: Biology 69 (2003) 107–120 109
benzimidazole) trihydrochloride) and trypsin were obtained (Olympus, BX60, Japan) equipped with a monochrome¨from Sigma, USA. Calf thymus DNA was obtained from CCD camera (Grundig, FA87, Germany). The CCD ca-
E-Merck; Annexin-V-FITC from PharMingen, USA. All mera contained a monochrome frame grabber of 24 Bitother chemicals used in the present study were of ana- (Bit flow, USA) and the spectrum range was 400–1000 nmlytical grade from BDH, Glaxo laboratories (Qualigens), with an image sensor of 596 (V)3795 (H) total pixels.SRL, and E-Merck, India. Cells were grown on coverslips for these studies. After
incubation with HpD, coverslips were washed in PBS,2 .3. Estimation of intracellular concentration of the mounted on slides and examined under the fluorescencephotosensitizer microscope using a UV excitation filter (300–400 nm) and
emission recorded in the red region of the spectrum (400–Monolayer cells were incubated in HBSS containing 800 nm). Images were acquired and stored in a digital
different concentrations of the sensitizer (2.5–10mg/ml) pentium-based computer (166 MHz) and analyzed usingand incubated at 378C in dark for various time intervals the software provided by Optimas, USA.(0.5, 1, 2, 4 h). Cellular concentration of HpD was Cytoplasmic and nuclear localization of HpD wasestimated by methanol extraction essentially according to estimated by analyzing the images using area morphometryHilf et al. [42] and whole cell suspension spectroscopy. by marking the appropriate regions of interest (ROI).
2 .3.1. Methanol extraction 2 .5. Photodynamic treatmentAt the end of the incubation, HBSS was removed and
cells were washed twice with PBS. Methanol was added to Cells were incubated in HBSS at 378C for 4 h withthe attached cells and incubated for 1 h at 378C. Fluores- varying concentrations (1.25–20mg/ml) of HpD. After thecence spectra of the methanol extractable HpD was incubation, cells were washed with HBSS and exposed to
2obtained with an excitation wavelength of 405 nm and red light (power53 W/cm ) using an optical filter with aemission was collected in the range of 550–750 nm using cut-off at 610 nm from a high power (1000 W) xenon arca Jobin Yvon fluorimeter (Model JY3C, France). Quantita- lamp (Oriel, USA). Optical power at the cell surface wastion was carried out using the calibration curves generated measured using a radiometer (Model 1400 A, Internationalwith various concentrations of HpD in methanol by Radiometer, USA) having a detector head (SL021/FQ)exciting at 405 nm and measuring the fluorescence at 630 with a flat response in the spectral range 400–1000 nm.nm. Cells were incubated for another 2 h at 378C in HBSS
To determine the cell numbers in the culture, cells were before assay of cell response to treatment.scraped from flasks and homogenized using a handhomogenizer. The cell number was estimated from the 2 .6. Cellular response to photodynamic treatmentDNA content of the homogenate using an AT-specificDNA fluorochrome, bisbenzimidazole (Hoechst-33342) 2 .6.1. Clonogenic survival assayand a calibration curve generated with calf thymus DNA Nearly 150 cells were plated in growth medium[43]. (DMEM110% FCS) after the treatment (as described
above) and incubated in the dark under a humidified, 5%2 .3.2. Fluorescence spectroscopic measurements in cell CO atmosphere at 378C for 8–10 days to allow colony2
suspension formation. Colonies were fixed with methanol and stainedCells were trypsinized and incubated in the dark for with 1% crystal violet. Colonies having more than 50 cells
various time intervals with HpD in HBSS at 378C. were counted and plating efficiency (PE) and survivingFluorescence spectra (exc. 405 nm, em. 550–750 nm) of fraction (S.F.) were calculated.cells and supernatant before and after washing wereobtained. Cellular uptake was calculated using standard2 .6.2. Cell proliferation kineticscalibration curves of the photosensitizer in HBSS. After photodynamic treatment, cells were incubated in
Intracellular concentration of HpD was estimated from growth medium for varying intervals of time, harvested bythe average cell volume. The cell volume (v) was de- trypsinization and counted using a hemocytometertermined by measuring the cell diameter (d) with the help (adherent1floating). Flow cytometric measurements ofof a micrometer, assuming spherical cell shape and using cellular DNA content were performed with the ethanol
3the formulav 5pd /6. (70%) fixed cells using an intercalating DNA fluoro-chrome, propidium iodide (PI) as described earlier [44].
2 .4. Intracellular distribution of HpD using a Briefly, the cells (|0.5–1 million) were washed in PBSfluorescence image analysis system after removing ethanol and treated with Ribonuclease-A
(200mg/ml) for 30 min at 378C. Subsequently, cells wereIntracellular distribution of the sensitizer was studied by stained with PI (50mg/ml) in PBS. Measurements were
fluorescence microscopy using an image analysis system made with a laser-based (488 nm) flow cytometer (Facs
110 S. Gupta et al. / Journal of Photochemistry and Photobiology B: Biology 69 (2003) 107–120
Calibur; Becton and Dickinson, USA) and data acquired NaOH, pH 7.4; 140 mM NaCl; 2.5 mM CaCl ) and 5ml of2
using the Cell Quest software (Becton and Dickinson, Annexin-V-FITC and 10ml of PI (50 mg/ml) added. AfterUSA). Cell cycle analysis was performed using the Modfit 15 min at room temperature, 400ml of binding buffer wereprogram. added to each sample and analyzed by flow cytometry. The
percentages of Annexin1ve and 2ve cells were esti-mated by applying appropriate gates and using the regional2 .6.3. Micronuclei formationstatistics analysis facility provided in the Cell QuestAir-dried slides containing acetic acid–methanol (1:3,software (Becton and Dickinson, USA).v /v) fixed cells were stained with 2-aminophenylin-
doledihydrochloride (DAPI) (10mg/ml in citric acid (0.01M), disodium phosphate (0.45 M) buffer containing 0.05%
3 . ResultsTween-20 detergent) as described earlier [45]. Slides wereexamined under a fluorescence microscope using UV
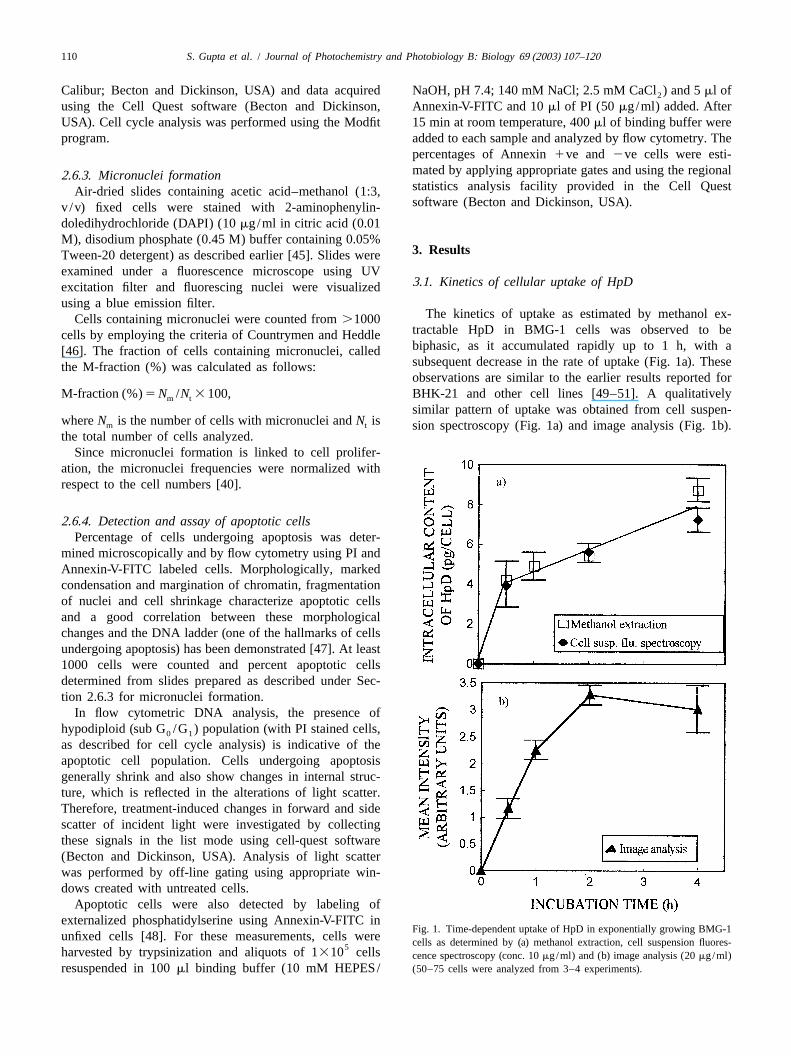
3 .1. Kinetics of cellular uptake of HpDexcitation filter and fluorescing nuclei were visualizedusing a blue emission filter.
The kinetics of uptake as estimated by methanol ex-Cells containing micronuclei were counted from.1000tractable HpD in BMG-1 cells was observed to becells by employing the criteria of Countrymen and Heddlebiphasic, as it accumulated rapidly up to 1 h, with a[46]. The fraction of cells containing micronuclei, calledsubsequent decrease in the rate of uptake (Fig. 1a). Thesethe M-fraction (%) was calculated as follows:observations are similar to the earlier results reported for
M-fraction (%)5N /N 3 100, BHK-21 and other cell lines [49–51]. A qualitativelym t
similar pattern of uptake was obtained from cell suspen-whereN is the number of cells with micronuclei andN ism t sion spectroscopy (Fig. 1a) and image analysis (Fig. 1b).the total number of cells analyzed.
Since micronuclei formation is linked to cell prolifer-ation, the micronuclei frequencies were normalized withrespect to the cell numbers [40].
2 .6.4. Detection and assay of apoptotic cellsPercentage of cells undergoing apoptosis was deter-
mined microscopically and by flow cytometry using PI andAnnexin-V-FITC labeled cells. Morphologically, markedcondensation and margination of chromatin, fragmentationof nuclei and cell shrinkage characterize apoptotic cellsand a good correlation between these morphologicalchanges and the DNA ladder (one of the hallmarks of cellsundergoing apoptosis) has been demonstrated [47]. At least1000 cells were counted and percent apoptotic cellsdetermined from slides prepared as described under Sec-tion 2.6.3 for micronuclei formation.
In flow cytometric DNA analysis, the presence ofhypodiploid (sub G /G ) population (with PI stained cells,0 1
as described for cell cycle analysis) is indicative of theapoptotic cell population. Cells undergoing apoptosisgenerally shrink and also show changes in internal struc-ture, which is reflected in the alterations of light scatter.Therefore, treatment-induced changes in forward and sidescatter of incident light were investigated by collectingthese signals in the list mode using cell-quest software(Becton and Dickinson, USA). Analysis of light scatterwas performed by off-line gating using appropriate win-dows created with untreated cells.
Apoptotic cells were also detected by labeling ofexternalized phosphatidylserine using Annexin-V-FITC in
Fig. 1. Time-dependent uptake of HpD in exponentially growing BMG-1unfixed cells [48]. For these measurements, cells were cells as determined by (a) methanol extraction, cell suspension fluores-5harvested by trypsinization and aliquots of 1310 cells cence spectroscopy (conc. 10mg/ml) and (b) image analysis (20mg/ml)resuspended in 100ml binding buffer (10 mM HEPES/ (50–75 cells were analyzed from 3–4 experiments).
S. Gupta et al. / Journal of Photochemistry and Photobiology B: Biology 69 (2003) 107–120 111
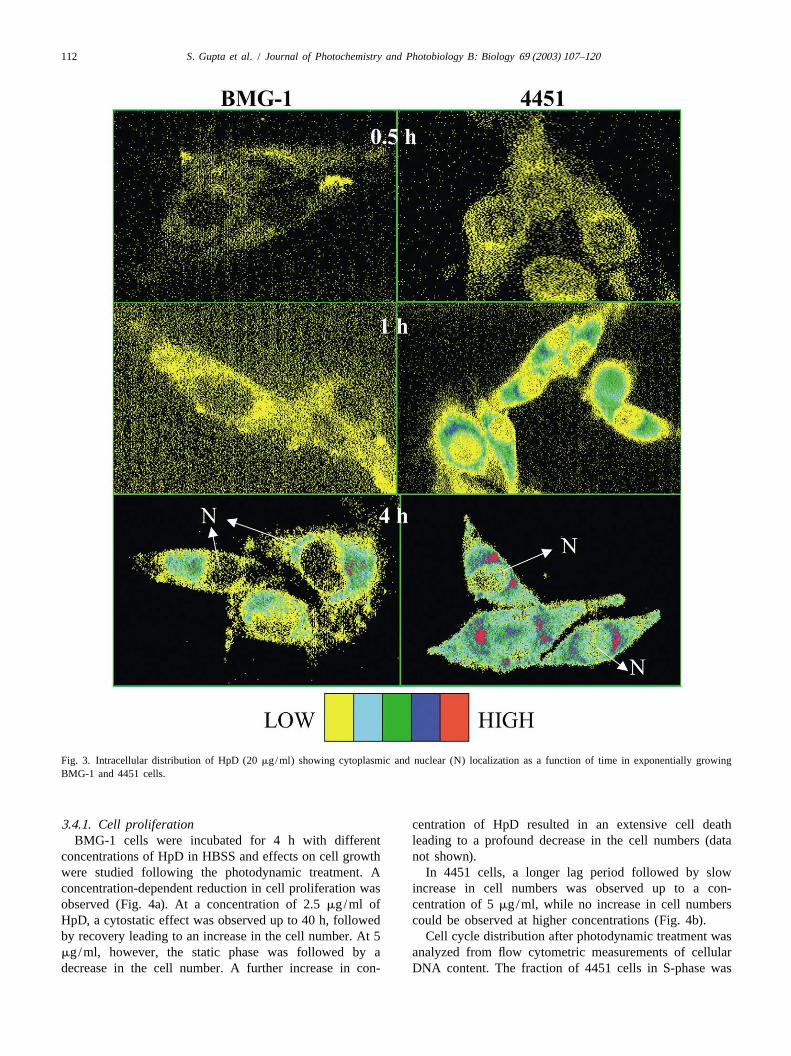
However, with the image analysis system, a saturation in cell membrane after 30 min of incubation; however, byuptake was observed at longer incubation times. Similar 1–2 h the red fluorescence increased in the cytoplasm anduptake kinetics of hypocrellin B (HB) in EMT6/Ed murine in the perinuclear region. A careful examination of thetumor cells has been reported using semiquantitative images revealed that at 4 h 75–90% of the sensitizer wasconfocal microscopy and radiolabeled HB [52]. localized in the cytoplasm of both cell lines (Fig. 3).
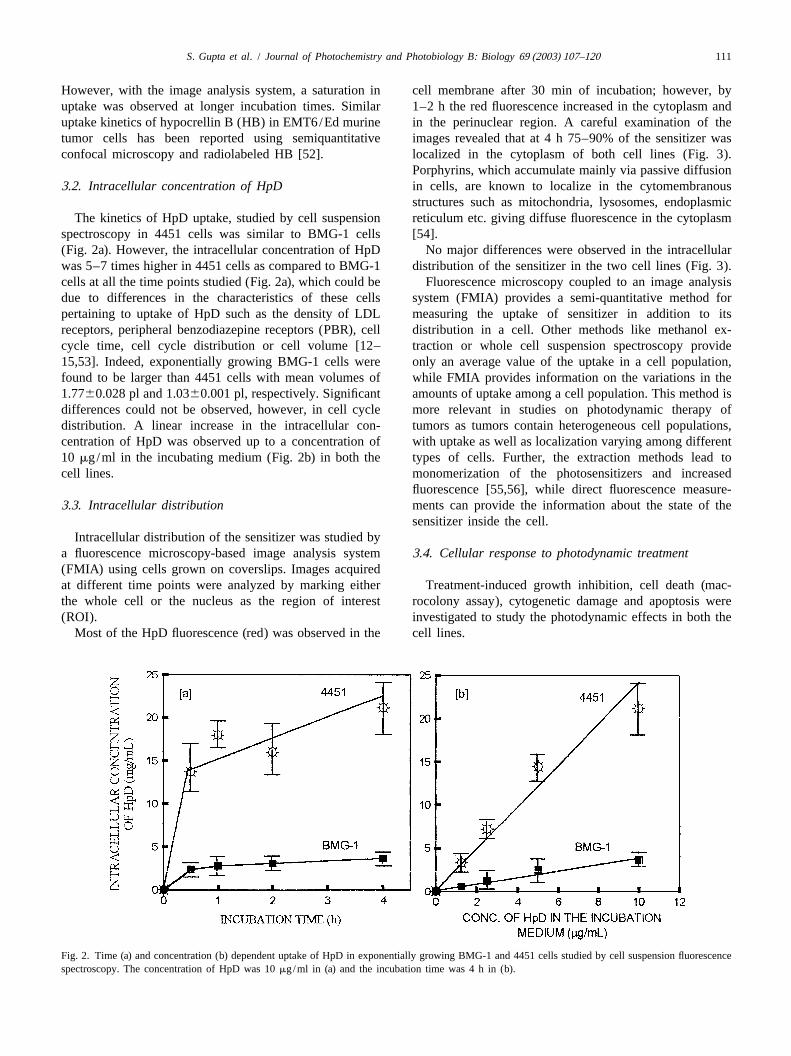
Porphyrins, which accumulate mainly via passive diffusion3 .2. Intracellular concentration of HpD in cells, are known to localize in the cytomembranous
structures such as mitochondria, lysosomes, endoplasmicThe kinetics of HpD uptake, studied by cell suspension reticulum etc. giving diffuse fluorescence in the cytoplasm
spectroscopy in 4451 cells was similar to BMG-1 cells [54].(Fig. 2a). However, the intracellular concentration of HpD No major differences were observed in the intracellularwas 5–7 times higher in 4451 cells as compared to BMG-1 distribution of the sensitizer in the two cell lines (Fig. 3).cells at all the time points studied (Fig. 2a), which could be Fluorescence microscopy coupled to an image analysisdue to differences in the characteristics of these cells system (FMIA) provides a semi-quantitative method forpertaining to uptake of HpD such as the density of LDL measuring the uptake of sensitizer in addition to itsreceptors, peripheral benzodiazepine receptors (PBR), cell distribution in a cell. Other methods like methanol ex-cycle time, cell cycle distribution or cell volume [12– traction or whole cell suspension spectroscopy provide15,53]. Indeed, exponentially growing BMG-1 cells were only an average value of the uptake in a cell population,found to be larger than 4451 cells with mean volumes of while FMIA provides information on the variations in the1.7760.028 pl and 1.0360.001 pl, respectively. Significant amounts of uptake among a cell population. This method isdifferences could not be observed, however, in cell cycle more relevant in studies on photodynamic therapy ofdistribution. A linear increase in the intracellular con- tumors as tumors contain heterogeneous cell populations,centration of HpD was observed up to a concentration of with uptake as well as localization varying among different10 mg/ml in the incubating medium (Fig. 2b) in both the types of cells. Further, the extraction methods lead tocell lines. monomerization of the photosensitizers and increased
fluorescence [55,56], while direct fluorescence measure-3 .3. Intracellular distribution ments can provide the information about the state of the
sensitizer inside the cell.Intracellular distribution of the sensitizer was studied by
a fluorescence microscopy-based image analysis system3 .4. Cellular response to photodynamic treatment(FMIA) using cells grown on coverslips. Images acquiredat different time points were analyzed by marking either Treatment-induced growth inhibition, cell death (mac-the whole cell or the nucleus as the region of interest rocolony assay), cytogenetic damage and apoptosis were(ROI). investigated to study the photodynamic effects in both the
Most of the HpD fluorescence (red) was observed in the cell lines.
Fig. 2. Time (a) and concentration (b) dependent uptake of HpD in exponentially growing BMG-1 and 4451 cells studied by cell suspension fluorescencespectroscopy. The concentration of HpD was 10mg/ml in (a) and the incubation time was 4 h in (b).
112 S. Gupta et al. / Journal of Photochemistry and Photobiology B: Biology 69 (2003) 107–120
Fig. 3. Intracellular distribution of HpD (20mg/ml) showing cytoplasmic and nuclear (N) localization as a function of time in exponentially growingBMG-1 and 4451 cells.
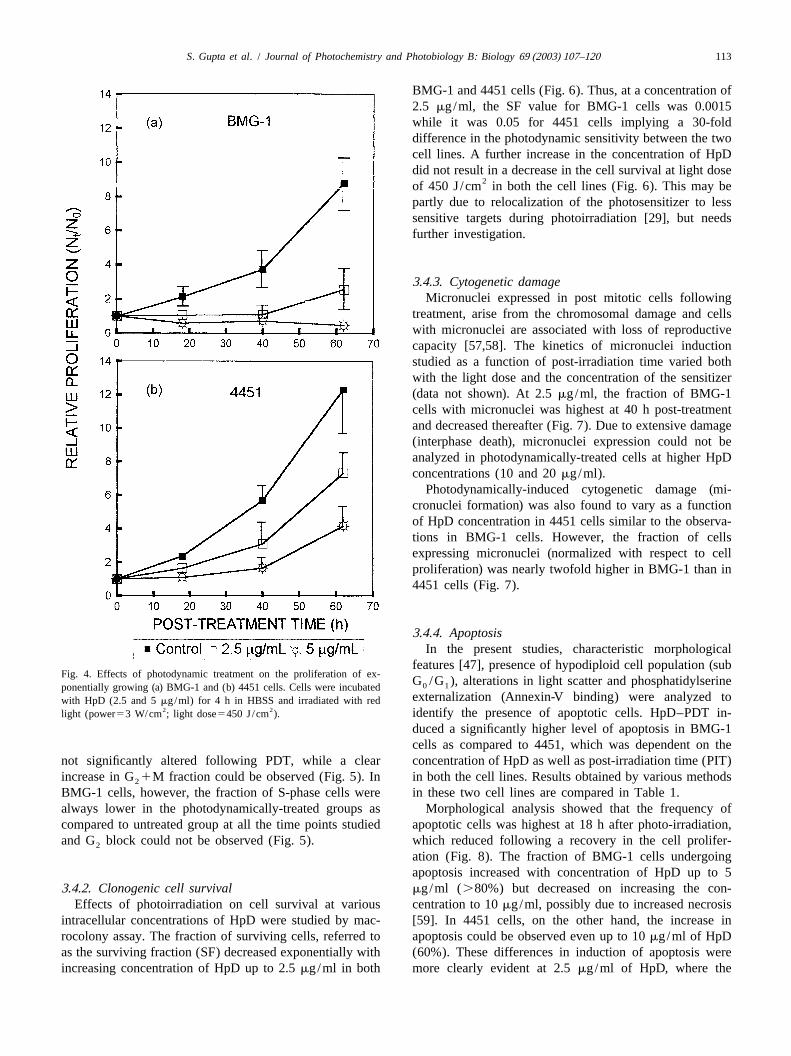
3 .4.1. Cell proliferation centration of HpD resulted in an extensive cell deathBMG-1 cells were incubated for 4 h with different leading to a profound decrease in the cell numbers (data
concentrations of HpD in HBSS and effects on cell growth not shown).were studied following the photodynamic treatment. A In 4451 cells, a longer lag period followed by slowconcentration-dependent reduction in cell proliferation was increase in cell numbers was observed up to a con-observed (Fig. 4a). At a concentration of 2.5mg/ml of centration of 5mg/ml, while no increase in cell numbersHpD, a cytostatic effect was observed up to 40 h, followed could be observed at higher concentrations (Fig. 4b).by recovery leading to an increase in the cell number. At 5 Cell cycle distribution after photodynamic treatment wasmg/ml, however, the static phase was followed by a analyzed from flow cytometric measurements of cellulardecrease in the cell number. A further increase in con- DNA content. The fraction of 4451 cells in S-phase was
S. Gupta et al. / Journal of Photochemistry and Photobiology B: Biology 69 (2003) 107–120 113
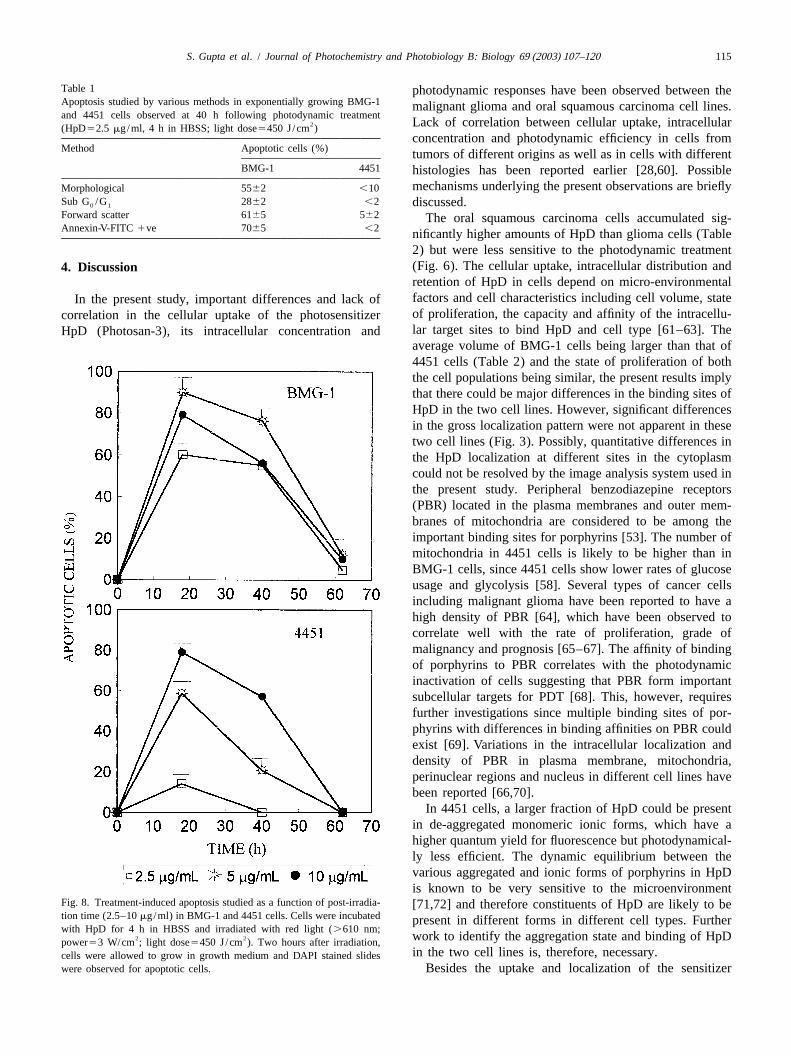
BMG-1 and 4451 cells (Fig. 6). Thus, at a concentration of2.5 mg/ml, the SF value for BMG-1 cells was 0.0015while it was 0.05 for 4451 cells implying a 30-folddifference in the photodynamic sensitivity between the twocell lines. A further increase in the concentration of HpDdid not result in a decrease in the cell survival at light dose
2of 450 J/cm in both the cell lines (Fig. 6). This may bepartly due to relocalization of the photosensitizer to lesssensitive targets during photoirradiation [29], but needsfurther investigation.
3 .4.3. Cytogenetic damageMicronuclei expressed in post mitotic cells following
treatment, arise from the chromosomal damage and cellswith micronuclei are associated with loss of reproductivecapacity [57,58]. The kinetics of micronuclei inductionstudied as a function of post-irradiation time varied bothwith the light dose and the concentration of the sensitizer(data not shown). At 2.5mg/ml, the fraction of BMG-1cells with micronuclei was highest at 40 h post-treatmentand decreased thereafter (Fig. 7). Due to extensive damage(interphase death), micronuclei expression could not beanalyzed in photodynamically-treated cells at higher HpDconcentrations (10 and 20mg/ml).
Photodynamically-induced cytogenetic damage (mi-cronuclei formation) was also found to vary as a functionof HpD concentration in 4451 cells similar to the observa-tions in BMG-1 cells. However, the fraction of cellsexpressing micronuclei (normalized with respect to cellproliferation) was nearly twofold higher in BMG-1 than in4451 cells (Fig. 7).
3 .4.4. ApoptosisIn the present studies, characteristic morphological
features [47], presence of hypodiploid cell population (subFig. 4. Effects of photodynamic treatment on the proliferation of ex-
G /G ), alterations in light scatter and phosphatidylserine0 1ponentially growing (a) BMG-1 and (b) 4451 cells. Cells were incubatedexternalization (Annexin-V binding) were analyzed towith HpD (2.5 and 5mg/ml) for 4 h in HBSS and irradiated with red
2 2 identify the presence of apoptotic cells. HpD–PDT in-light (power53 W/cm ; light dose5450 J/cm ).
duced a significantly higher level of apoptosis in BMG-1cells as compared to 4451, which was dependent on the
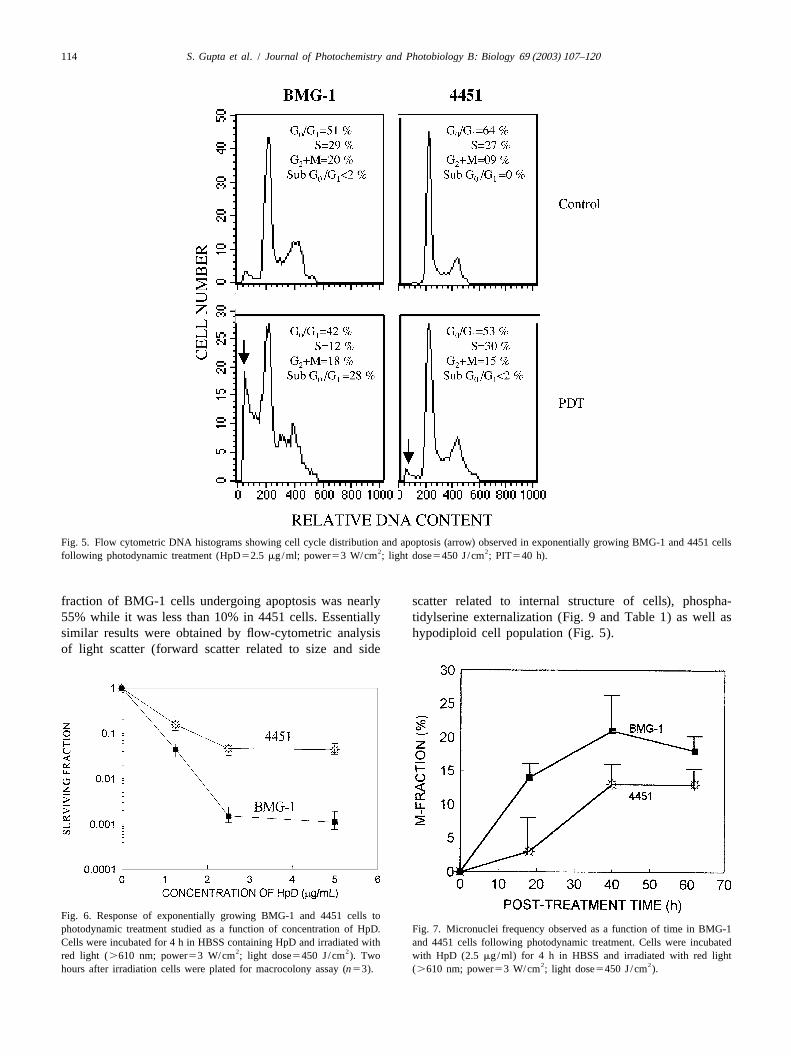
not significantly altered following PDT, while a clear concentration of HpD as well as post-irradiation time (PIT)increase in G1M fraction could be observed (Fig. 5). In in both the cell lines. Results obtained by various methods2
BMG-1 cells, however, the fraction of S-phase cells were in these two cell lines are compared in Table 1.always lower in the photodynamically-treated groups as Morphological analysis showed that the frequency ofcompared to untreated group at all the time points studied apoptotic cells was highest at 18 h after photo-irradiation,and G block could not be observed (Fig. 5). which reduced following a recovery in the cell prolifer-2
ation (Fig. 8). The fraction of BMG-1 cells undergoingapoptosis increased with concentration of HpD up to 5
3 .4.2. Clonogenic cell survival mg/ml (.80%) but decreased on increasing the con-Effects of photoirradiation on cell survival at various centration to 10mg/ml, possibly due to increased necrosis
intracellular concentrations of HpD were studied by mac- [59]. In 4451 cells, on the other hand, the increase inrocolony assay. The fraction of surviving cells, referred to apoptosis could be observed even up to 10mg/ml of HpDas the surviving fraction (SF) decreased exponentially with (60%). These differences in induction of apoptosis wereincreasing concentration of HpD up to 2.5mg/ml in both more clearly evident at 2.5mg/ml of HpD, where the
114 S. Gupta et al. / Journal of Photochemistry and Photobiology B: Biology 69 (2003) 107–120
Fig. 5. Flow cytometric DNA histograms showing cell cycle distribution and apoptosis (arrow) observed in exponentially growing BMG-1 and 4451 cells2 2following photodynamic treatment (HpD52.5 mg/ml; power53 W/cm ; light dose5450 J/cm ; PIT540 h).
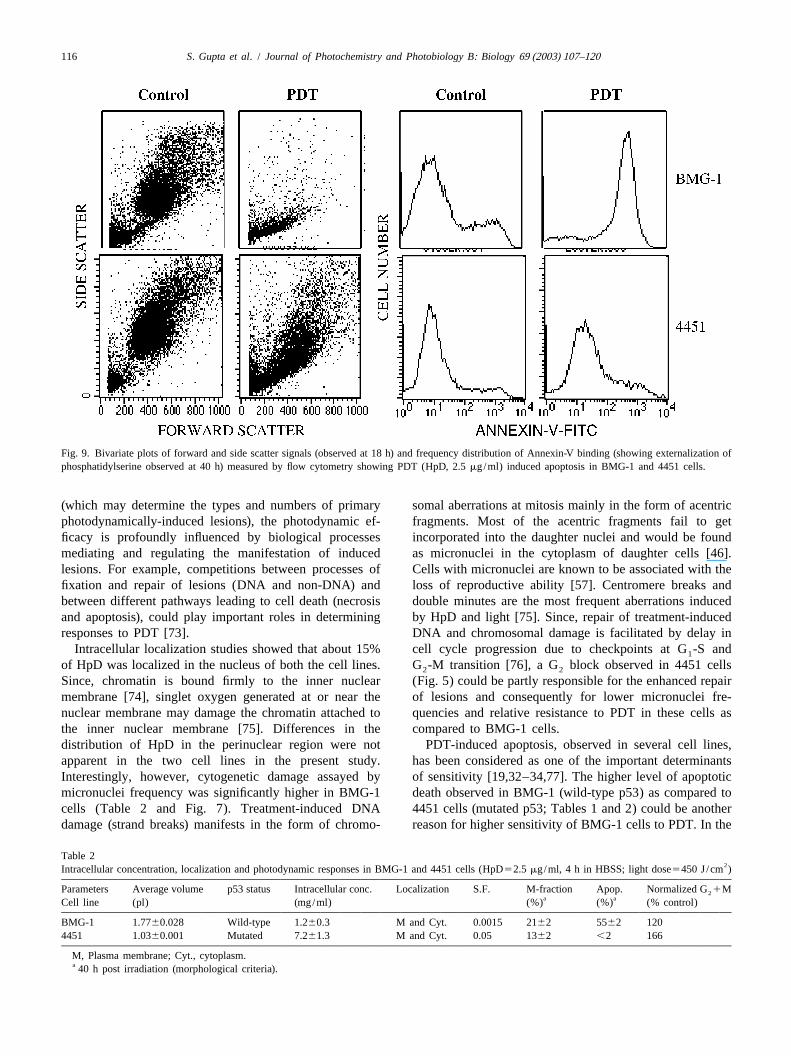
fraction of BMG-1 cells undergoing apoptosis was nearly scatter related to internal structure of cells), phospha-55% while it was less than 10% in 4451 cells. Essentially tidylserine externalization (Fig. 9 and Table 1) as well assimilar results were obtained by flow-cytometric analysis hypodiploid cell population (Fig. 5).of light scatter (forward scatter related to size and side
Fig. 6. Response of exponentially growing BMG-1 and 4451 cells tophotodynamic treatment studied as a function of concentration of HpD. Fig. 7. Micronuclei frequency observed as a function of time in BMG-1Cells were incubated for 4 h in HBSS containing HpD and irradiated with and 4451 cells following photodynamic treatment. Cells were incubated
2 2red light (.610 nm; power53 W/cm ; light dose5450 J/cm ). Two with HpD (2.5mg/ml) for 4 h in HBSS and irradiated with red light2 2hours after irradiation cells were plated for macrocolony assay (n53). (.610 nm; power53 W/cm ; light dose5450 J/cm ).
S. Gupta et al. / Journal of Photochemistry and Photobiology B: Biology 69 (2003) 107–120 115
Table 1 photodynamic responses have been observed between theApoptosis studied by various methods in exponentially growing BMG-1 malignant glioma and oral squamous carcinoma cell lines.and 4451 cells observed at 40 h following photodynamic treatment
2 Lack of correlation between cellular uptake, intracellular(HpD52.5 mg/ml, 4 h in HBSS; light dose5450 J/cm )concentration and photodynamic efficiency in cells from
Method Apoptotic cells (%) tumors of different origins as well as in cells with differentBMG-1 4451 histologies has been reported earlier [28,60]. Possible
mechanisms underlying the present observations are brieflyMorphological 5562 ,10Sub G /G 2862 ,2 discussed.0 1
Forward scatter 6165 562 The oral squamous carcinoma cells accumulated sig-Annexin-V-FITC1ve 7065 ,2 nificantly higher amounts of HpD than glioma cells (Table
2) but were less sensitive to the photodynamic treatment(Fig. 6). The cellular uptake, intracellular distribution and4 . Discussionretention of HpD in cells depend on micro-environmentalfactors and cell characteristics including cell volume, stateIn the present study, important differences and lack ofof proliferation, the capacity and affinity of the intracellu-correlation in the cellular uptake of the photosensitizerlar target sites to bind HpD and cell type [61–63]. TheHpD (Photosan-3), its intracellular concentration andaverage volume of BMG-1 cells being larger than that of4451 cells (Table 2) and the state of proliferation of boththe cell populations being similar, the present results implythat there could be major differences in the binding sites ofHpD in the two cell lines. However, significant differencesin the gross localization pattern were not apparent in thesetwo cell lines (Fig. 3). Possibly, quantitative differences inthe HpD localization at different sites in the cytoplasmcould not be resolved by the image analysis system used inthe present study. Peripheral benzodiazepine receptors(PBR) located in the plasma membranes and outer mem-branes of mitochondria are considered to be among theimportant binding sites for porphyrins [53]. The number ofmitochondria in 4451 cells is likely to be higher than inBMG-1 cells, since 4451 cells show lower rates of glucoseusage and glycolysis [58]. Several types of cancer cellsincluding malignant glioma have been reported to have ahigh density of PBR [64], which have been observed tocorrelate well with the rate of proliferation, grade ofmalignancy and prognosis [65–67]. The affinity of bindingof porphyrins to PBR correlates with the photodynamicinactivation of cells suggesting that PBR form importantsubcellular targets for PDT [68]. This, however, requiresfurther investigations since multiple binding sites of por-phyrins with differences in binding affinities on PBR couldexist [69]. Variations in the intracellular localization anddensity of PBR in plasma membrane, mitochondria,perinuclear regions and nucleus in different cell lines havebeen reported [66,70].
In 4451 cells, a larger fraction of HpD could be presentin de-aggregated monomeric ionic forms, which have ahigher quantum yield for fluorescence but photodynamical-ly less efficient. The dynamic equilibrium between thevarious aggregated and ionic forms of porphyrins in HpDis known to be very sensitive to the microenvironment
Fig. 8. Treatment-induced apoptosis studied as a function of post-irradia- [71,72] and therefore constituents of HpD are likely to betion time (2.5–10mg/ml) in BMG-1 and 4451 cells. Cells were incubated present in different forms in different cell types. Furtherwith HpD for 4 h in HBSS and irradiated with red light (.610 nm;
2 2 work to identify the aggregation state and binding of HpDpower53 W/cm ; light dose5450 J/cm ). Two hours after irradiation,in the two cell lines is, therefore, necessary.cells were allowed to grow in growth medium and DAPI stained slides
were observed for apoptotic cells. Besides the uptake and localization of the sensitizer
116 S. Gupta et al. / Journal of Photochemistry and Photobiology B: Biology 69 (2003) 107–120
Fig. 9. Bivariate plots of forward and side scatter signals (observed at 18 h) and frequency distribution of Annexin-V binding (showing externalization ofphosphatidylserine observed at 40 h) measured by flow cytometry showing PDT (HpD, 2.5mg/ml) induced apoptosis in BMG-1 and 4451 cells.
(which may determine the types and numbers of primary somal aberrations at mitosis mainly in the form of acentricphotodynamically-induced lesions), the photodynamic ef- fragments. Most of the acentric fragments fail to getficacy is profoundly influenced by biological processes incorporated into the daughter nuclei and would be foundmediating and regulating the manifestation of induced as micronuclei in the cytoplasm of daughter cells [46].lesions. For example, competitions between processes of Cells with micronuclei are known to be associated with thefixation and repair of lesions (DNA and non-DNA) and loss of reproductive ability [57]. Centromere breaks andbetween different pathways leading to cell death (necrosis double minutes are the most frequent aberrations inducedand apoptosis), could play important roles in determining by HpD and light [75]. Since, repair of treatment-inducedresponses to PDT [73]. DNA and chromosomal damage is facilitated by delay in
Intracellular localization studies showed that about 15% cell cycle progression due to checkpoints at G -S and1
of HpD was localized in the nucleus of both the cell lines. G -M transition [76], a G block observed in 4451 cells2 2
Since, chromatin is bound firmly to the inner nuclear (Fig. 5) could be partly responsible for the enhanced repairmembrane [74], singlet oxygen generated at or near the of lesions and consequently for lower micronuclei fre-nuclear membrane may damage the chromatin attached to quencies and relative resistance to PDT in these cells asthe inner nuclear membrane [75]. Differences in the compared to BMG-1 cells.distribution of HpD in the perinuclear region were not PDT-induced apoptosis, observed in several cell lines,apparent in the two cell lines in the present study. has been considered as one of the important determinantsInterestingly, however, cytogenetic damage assayed by of sensitivity [19,32–34,77]. The higher level of apoptoticmicronuclei frequency was significantly higher in BMG-1 death observed in BMG-1 (wild-type p53) as compared tocells (Table 2 and Fig. 7). Treatment-induced DNA 4451 cells (mutated p53; Tables 1 and 2) could be anotherdamage (strand breaks) manifests in the form of chromo- reason for higher sensitivity of BMG-1 cells to PDT. In the
Table 22Intracellular concentration, localization and photodynamic responses in BMG-1 and 4451 cells (HpD52.5 mg/ml, 4 h in HBSS; light dose5450 J/cm )
Parameters Average volume p53 status Intracellular conc. Localization S.F. M-fraction Apop. Normalized G1M2a aCell line (pl) (mg/ml) (%) (%) (% control)
BMG-1 1.7760.028 Wild-type 1.260.3 M and Cyt. 0.0015 2162 5562 1204451 1.0360.001 Mutated 7.261.3 M and Cyt. 0.05 1362 ,2 166
M, Plasma membrane; Cyt., cytoplasm.a 40 h post irradiation (morphological criteria).

S. Gupta et al. / Journal of Photochemistry and Photobiology B: Biology 69 (2003) 107–120 117
present work, four different assays/parameters were used following apoptotic stimulation inducing chromosomefor detection and quantitative estimate of the apoptotic condensation and DNA fragmentation. AMID-inducedindex. All the methods used indicated a higher frequency apoptosis is however different as compared to the classicof apoptotic cells in BMG-1 cells as compared to 4451 apoptotic pathways as it is delayed and not inhibited bycells (Table 1). The percentage of apoptotic cells estimated Bcl-2. Further, association of AMID with the outer mem-from hypodiploid DNA content was, however, considera- brane of mitochondria appears to be critical although thebly lower in both the cell lines as compared to estimates of exact mechanisms involved are not known. These path-apoptotic cells by cell volume (forward scatter), PS ways could also contribute significantly to the HpD–PDTexternalization (Annexin-V binding) and morphological induced apoptosis, particularly in 4451 cells with mutatedfeatures (Table 1). Absence of DNA ladder in cells p53 and warrant further investigations.undergoing apoptosis revealed by different techniques has Taken together, the present studies suggest that differ-been reported earlier [78]. Differences in the estimates of ences in the accumulation and distribution of HpD betweenapoptotic fractions measured by methods based on differ- different cells could arise from the variations in theent parameters could be associated with different phases of frequency and affinity of HpD binding sites (mainly PBRthe apoptotic process. However, such differences could located in mitochondria) as well as the aggregation andalso arise due to a number of other reasons viz. negligible ionic states of intracellular HpD. Cellular responses tocleavage of DNA at internucleosomal regions with cleav- PDT crucially depend also on the processing of the PDT-age at higher levels of chromatin organization, or the induced lesions by competing repair /fixation pathways anddifferences in the preparatory method used for flow therefore on the cellular context viz. the genotype andcytometric DNA analysis, where an extraction buffer was phenotypic /metabolic characteristics of cells besides thenot used which facilitates resolution of sub G /G phase uptake and localization of the sensitizer. Since cell death0 1
from G cells [79]. following PDT can be induced by necrosis and multiple1
Apoptotic cell death can be initiated by a variety of apoptotic pathways (p53 dependent as well as indepen-metabolic and genetic stress signals. Several pathways that dent) regulated in a complex way by interactions ofmediate apoptotic cell death are regulated in a complex numerous genes and gene products, the differences in themanner by numerous genes and their products; p53 and PDT responses of various cell types cannot be predictedBcl family of proteins appear to be the most important only on the basis of the uptake/ intracellular concentrationones [35–37,80–83]. Since, photodynamic treatment can of the photosensitizer or status of a single gene such asinduce cell membrane, mitochondrial, cytoskeletal and p53.DNA damage, multiple independent as well as interdepen-dent pathways could be involved in photodynamically-induced apoptosis. Further, the expression of death re-A cknowledgementsceptors like CD95/Fas/Apo-1, TNF and TRAIL [84–87]and the release of apoptogenic factors such as cytochrome Thanks are due to Dr. N.K. Chaudhury, Mr. J.S.c, Smac/DIABLO through mitochondrial membrane per- Adhikari and Dr. Sudhir Chandna for help in spectro-meability transition (MMPT) as well as the expression of scopic, flow cytometric and image analysis studies. We areBcl family of genes are regulated by p53, although the grateful to Maj. Gen. T. Ravindranath, Director, INMASexact mechanisms remain yet to be completely elucidatedand Dr. T. Lazar Mathew, former Director for their interest[88–90]. and support for this work. S.G. was a recipient of a
Since, significant differences in the levels of anti-apo- research fellowship from the University Grants Commis-ptotic (Bcl-2, Bcl ) and pro-apoptotic (Bax) proteinsXL sion and CSIR, Govt. of India.could not be observed in these two cell lines (data notshown), it appears that apoptosis can indeed be inducedboth in wild type and mutated p53 gene carrying cell types
R eferencesalbeit at different photodynamic doses. It is pertinent tonote that although a positive correlation between the p53
[1] T.J. Dougherty, C.J. Gomer, B.W. Henderson, G. Jori, D. Kessel, M.status and photodynamic sensitivity has been found inKorbelik, J. Moan, Q. Peng, Photodynamic therapy, J. Natl. Cancerseveral studies through transfection of wild type p53 geneInst. 90 (1998) 889–905.
in p53 mutant cell lines [37,91], recent studies have [2] R.L. Lipson, E.J. Baldes, A.M. Olsen, Hematoporphyrin derivative:however shown that apoptosis can occur also through a new aid for endoscopic detection of malignant disease, J. Thorac.caspase and p53 independent pathways, for example, Cardiovasc. Surg. 42 (1961) 623.
[3] T.J. Dougherty, G.B. Grindey, R. Field, K.R. Weishaupt, D.G.through the apoptosis-inducing factor (AIF), a mitochon-Boyle, Photoradiation therapy II, cure of animal tumors withdrial flavoprotein [92] and its recently discovered homolo-hematoporphyrin and light, J. Natl. Cancer Inst. 55 (1975) 115–121.
gous molecule designated as AMID (AIF-homologous [4] P.J. Muller, B.C. Wilson, Photodynamic therapy for recurrentmitochondrion-associated inducer of death) [93]. AIF has supratentorial gliomas, Semin. Surg. Oncol. 11 (1995) 346–354.been shown to translocate from mitochondria to nucleus [5] S. Krishnamurthy, S.K. Powers, P. Witmer, T. Brown, Optimal light

118 S. Gupta et al. / Journal of Photochemistry and Photobiology B: Biology 69 (2003) 107–120
dose for interstitial photodynamic therapy in treatment for malignant [25] C.W. Lin, J.R. Shulock, S.D. Kirley, L. Cincotta, J.W. Foley,Photodynamic destruction of lysosomes mediated by Nile Bluebrain tumors, Lasers Surg. Med. 27 (2000) 224–234.photosensitizers, Photochem. Photobiol. 58 (1993) 81–91.[6] W. Stummer, A. Novotny, H. Stepp, C. Goetz, K. Bise, H.J. Reulen,
[26] J. Moan, H. Waksvik, T. Christensen, DNA single strand breaks andFluorescence-guided resection of glioblastoma multiforme by usingsister chromatid exchanges induced by treatment with hemato-5-aminolevulinic acid-induced porphyrins: a prospective study in 52porphyrin and light or by X-rays in human NHIK 3025 cells, Cancerconsecutive patients, J. Neurosurg. 93 (2000) 1003–1013.Res. 40 (1980) 2915–2918.[7] A. Zimmermann, M. Ritsch-Marte, H. Kostron, mTHPC-mediated
[27] L. Hong, D.S. Shin, Y.E. Lee, D.C. Nguyen, T.C. Trang, A.H. Pan,photodynamic diagnosis of malignant brain tumors, Photochem.S.L.J. Huang, D.H. Chong, M.W. Berns, Subcellular phototoxicity ofPhotobiol. 74 (2001) 611–616.5-aminolevulinic acid (ALA), Lasers Surg. Med. 22 (1998) 14–24.[8] R. Baumgartner, H. Fisslinger, D. Jocham, H. Lenz, L. Ruprecht, H.
[28] D.J. Gomer, N. Ruckter, A.L. Murphree, Differential cell photo-Stepp, E. Unsold, A fluorescence imaging device for endoscopicsensitivity following porphyrin photodynamic therapy, Cancer Res.detection of early stage cancer—instrumental and experimental48 (1988) 4539–4542.studies, Photochem. Photobiol. 46 (1987) 759–763.
[29] J. Morgan, W.R. Potter, A.R. Oseroff, Comparison of photodynamic[9] K. Svanberg, I. Wang, S. Colleen, I. Idvall, C. Ingvar, R. Rydell, D.targets in a carcinoma cell line and its mitochondrial DNA-deficient
Jocham, H. Diddens, S. Bown, G. Gregory, S. Montan, S. Ander-derivative, Photochem. Photobiol. 71 (2000) 747–757.
sson-Engels, S. Svanberg, Clinical multi-colour fluorescence imag-[30] L.C. Penning, J.W.M. Lagerberg, J.H. VanDierendonck, C.J. Cor-
ing of malignant tumours—initial experience, Acta Radiol. 39nelisse, T.M.A.R. Dubbleman, J. VanSteveninck, The role of DNA
(1998) 2–9.damage and inhibition of poly (ADP-ribosyl) lation in loss of
[10] H.F. Blum, Photodynamic Action and Disease caused by Light, clonogenicity of murine L929 fibroblasts, caused by photodynami-Hofnov, New York, 1964. cally induced oxidative stress, Cancer Res. 54 (1994) 5561–5567.
[11] J. Moan, K. Berg, The photodegradation of porphyrins in cells can [31] D. Kessel, Y. Luo, Mitochondrial photodamage and PDT-inducedbe used to estimate the lifetime of singlet oxygen, Photochem. apoptosis, J. Photochem. Photobiol. B: Biol. 42 (1998) 89–95.Photobiol. 53 (1991) 549–553. [32] M.L. Agarwal, M.E. Clay, E.J. Harvey, H.H. Evans, A.R. Antunez,
[12] G. Oenbrink, P. Jurgenlimke, D. Gabel, Accumulation of porphyrin N.L. Oleinick, Photodynamic therapy induces rapid cell death byin cells: influence of hydrophobicity, aggregation and protein apoptosis in L5178Y mouse lymphoma cells, Cancer Res. 51 (1991)binding, Photochem. Photobiol. 48 (1988) 451–456. 5993–5996.
[13] K.W. Woodburn, N.J. Vardaxis, J.S. Hill, A.H. Kaye, D.R. Philips, [33] Y. Luo, C.K. Chang, D. Kessel, Rapid initiation of apoptosis bySubcellular localization of porphyrins using confocal laser scanning photodynamic therapy, Photochem. Photobiol. 63 (1996) 528–534.microscopy, Photochem. Photobiol. 54 (1991) 725–732. [34] V. Carre, C. Jayat, R. Granet, P. Krausz, M. Guilloton, Chronology
[14] K.W. Woodburn, N.J.Vardaxis, J.S. Hill, A.H. Kaye, J.A. Reiss, D.R. of apoptotic events induced in the K562 cell line by photodynamicPhillips, Evaluation of porphyrin characteristics required for photo- treatment with hematoporphyrin and monoglucosylporphyrin, Photo-dynamic therapy, Photochem. Photobiol. 55 (1992) 697–704. chem. Photobiol. 69 (1999) 55–60.
[15] F. Khanum, V. Jain, Studies concerning the role of energy metabo- [35] A.M.R. Fisher, N. Rucker, S. Wong, C.J. Gomer, Differentiallism in the photodynamic therapy, Assoc. Med. Phys. Bull. 12 photosensitivity in wild-type and mutant p53 human colon car-(1987) 113–124. cinoma cell lines, J. Photochem. Photobiol. B: Biol. 42 (1998)
[16] M. Soncin, L. Polo, E. Reddi, G. Jori, M.E. Kenney, G. Cheng, 104–107.M.A. Rodgers, Effect of axial ligation and delivery system on the [36] A.M.R. Fisher, A. Ferrario, N. Rucker, S. Zhang, C.J. Gomer,tumor-localizing and -photosensitizing properties of Ge(IV)-oc- Photodynamic therapy sensitivity is not altered in human tumor cellstabutoxy-phthalocyanines, Br. J. Cancer. 71 (1995) 727–732. after abrogation of p53 function, Cancer Res. 59 (1999) 331–335.
[17] D. Kessel, D. Woodburn, C.J. Gomer, N. Jagerovic, K.M. Smith, [37] Z. Tong, G. Singh, A.J. Rainbow, The role of p53 tumor suppressorPhotosensitization with derivatives of chlorin p , J. Photochem. in the response of human cells to photofrin-mediated photodynamic6
Photobiol. B: Biol. 28 (1995) 13–18. therapy, Photochem. Photobiol. 71 (2000) 201–210.[18] D.K. Gaffney, S.L. Schober, F. Sieber, Merocyanine 540-sensitized [38] S. Gupta, V. Jain, K. Muralidhar, Differences in photodynamic
photoinactivation of leukemia cells: role of oxygen and effects of effects of hematoporphyrin derivative in human squamous car-plasma membrane integrity and mitochondrial respiration, Exp. cinoma and glioma cell lines, in: M. Fahim (Ed.), Trends inHematol. 18 (1990) 23–26. Physiological Sciences: Cells to Systems, Serials Publications,
[19] M. Dellinger, Apoptosis or necrosis following photofrin photo- Delhi, 2002, pp. 225–247.sensitization: influence of the incubation protocol, Photochem. [39] S. Gupta, K. Muralidhar, V. Jain, Experimental studies to improvePhotobiol. 64 (1996) 182–187. photodynamic therapy of tumors, in: Radiobiology 2000-Internation-
[20] H. Dummin, T. Cernay, H.W. Zimmerman, Selective photosensitiza- al Conference on Radiation Biology, Trivandrum, India, Feb. 17–tion of mitochondria in HeLa cells by cationic Zn(II) phthalo- 19, 2000.cyanines with lipophilic side chains, J. Photochem. Photobiol. B: [40] B.S. Dwarakanath, V. Jain, Energy linked modifications of theBiol. 37 (1997) 219–229. radiation response in a human glioma cell line, Int. J. Radiat. Oncol.
[21] A.D. Munday, A. Sriratana, J.S. Hill, S.B. Kahl, P. Nagley, Biol. Phys. 17 (1989) 1033–1040.Mitochondria are the functional intracellular target for a photo- [41] F. Zolzer, S. Hillebrandt, C. Streffer, Radiation-induced G block1
sensitizing boronated porphyrin, Biochim. Biophys. Acta 1311 and p53 status in six human cell lines, Radiother. Oncol. 37 (1995)(1996) 1–4. 20–28.
[22] J.S. Modica-Napolitano, J.S. Joyal, G. Ara, A.R. Oseroff, J.R. [42] R. Hilf, P.B. Leakey, S.J. Sollot, S.L. Gibson, PhotodynamicAprille, Mitochondrial toxicity of cationic photosensitizes for photo- inactivation of R3230AC mammary carcinoma in vitro with hemato-chemotherapy, Cancer Res. 50 (1990) 7876–7881. porphyrin derivative: effects of dose, time and serum on uptake and
[23] K. Berg, J. Moan, Lysosomes and microtubules as targets for phototoxicity, Photochem. Photobiol. 37 (1983) 633–642.photochemotherapy of cancer, Photochem. Photobiol. 63 (1997) [43] C.F. Brunk, K.C. Jones, T.W. James, Assay for nanogram quantities403–409. of DNA in cellular homogenates, Anal. Biochem. 92 (1979) 497–
[24] M. Ambroz, A.J. MacRobert, J. Morgan, G. Rumbles, M.S.C. Foley, 500.D. Phillips, Time resolved fluorescence spectroscopy and intracellu- [44] B.S. Dwarakanath, J.S. Adhikari, V. Jain, Hematoporphyrin deriva-lar imaging of disulphonated aluminum phthalocyanine, J. Photo- tives potentiate the radiosensitizing effects of 2-deoxy-D-glucose inchem. Photobiol. B: Biol. 22 (1994) 105–117. cancer cells, Int. J. Radiat. Oncol. Biol. Phys. 43 (1999) 1125–1133.

S. Gupta et al. / Journal of Photochemistry and Photobiology B: Biology 69 (2003) 107–120 119
[45] B.S. Dwarakanath, V. Jain, Enhancement of the radiation-induced [65] H. Miettinen, J. Kononen, H. Haapasalo, P. Helen, P. Sallinen, T.damage by 2-deoxy-D-glucose in organ cultures of brain tumors, Harjuntausta, H. Helin, H. Alho, Expression of peripheral-typeIndian J. Med. Res. 82 (1985) 266–268. benzodiazepine receptor and diazepam binding inhibitor in human
astrocytomas: relationship to cell proliferation, Cancer Res. 55[46] P.I. Countryman, J.A. Heddle, Production of micronuclei from(1995) 2691–2695.chromosome aberrations in irradiated cultures of human lympho-
cytes, Mutat. Res. 41 (1976) 321–332. [66] M. Hardwick, D. Fertikh, M. Culty, H. Li, B. Vidic, V.Papadopoulos, Peripheral-type benzodiazepine receptor (PBR) in[47] K. Yanagihara, M. Nii, K. Numoto et al., Radiation-inducedhuman breast cancer: correlation of breast cancer cell aggressiveapoptotic cell death in human gastric epithelial tumor cells: correla-phenotype with PBR expression, nuclear localization, and PBR-tion between mitotic death and apoptosis, Int. J. Radiat. Biol. 77mediated cell proliferation and nuclear transport of cholesterol,(1995) 677–685.Cancer Res. 59 (1999) 831–842.[48] I. Vermes, C. Haanen, H. Steffens-Nakken, C. Reutelingsoerger, A
[67] A. Beinlich, R. Strohmeier, M. Kaufmann, H. Kuhl, Relation of cellnovel assay for apoptosis. Flow cytometric detection of phos-proliferation to expression of peripheral benzodiazepine receptors inphatidylserine expression on early apoptotic cells using fluoresceinhuman breast cancer cell lines, Biochem. Pharmacol. 60 (2000)labeled Annexin-V, J. Immunol. Methods 184 (1995) 39–51.397–402.[49] F. Khanum, V. Jain, Cellular accumulation and biological activity of
[68] M. Mesenholler, E.K. Matthews, A key role for the mitochondrialhematoporphyrin derivative (L) in comparison with photofrin II, Ind.benzodiazepine receptor in cellular photosensitization with delta-J. Exp. Biol. 35 (1997) 348–355.aminilevulinic acid, Eur. J. Pharmacol. 406 (2000) 171–180.[50] D. Kessel, Transport and binding of HpD and relative porphyrins by
[69] D. Kessel, M. Antolovich, K.M. Smith, The role of the peripheralmurine leukemia L1210 cells, Cancer Res. 41 (1981) 1318–1323.benzodiazepine receptor in the apoptotic response to photodynamic[51] R.M. Bohmer, G. Morstyn, Uptake of hematoporphyrin derivativetherapy, Photochem. Photobiol. 74 (2001) 346–349.by normal and malignant cells: effects of serum, pH, temperature
[70] C.H. Park, E. Carboni, P.L. Wood, K.W. Gee, Characterization ofand cell size, Cancer Res. 45 (1985) 5328–5334.peripheral benzodiazepine type sites in a cultured murine BV-2[52] G.G. Miller, K. Brown, R.B. Moore, Z.J. Diwu, J. Liu, L. Huang,microglial cell line, Glia 16 (1996) 65–70.J.W. Lown, D.A. Begg, V. Chlumecky, J. Tulip, M.S. McPhee,
[71] A. Kalia, V. Jain, Effects of some divalent metal ions on the agingUptake kinetics and intracellular localization of hypocrellin photo-phenomenon of hematoporphyrin and photofrin II, Ind. J. Biochem.sensitizers for photodynamic therapy: a confocal microscopy study,Biophys. 26 (1989) 213–218.Photochem. Photobiol. 61 (1995) 632–638.
[72] N.K. Chaudhury, V. Jain, Effects of microenvironment on aggrega-[53] A. Verma, S.H. Snyder, Peripheral type benzodiazepine receptors,tion and aging in homogeneous solutions of hematoporphyrin, in:Annu. Rev. Pharmacol. Toxicol. 29 (1989) 307–322.Fifth Congress of the European Society for Photobiology, Marburg,[54] R. Hilf, D.B. Small, R.S. Murant, P.B. Leaky, S.L. Gison, Hemato-Germany, 19–26 September, 1993.porphyrin derivative sensitivity of mitochondrial succinic dehydro-
genase and selected cytosolic enzymes of R3230AC mammary [73] V. Jain, Mechanisms and metabolic modulation of photosensitiza-adenocarcinoma of rats, Cancer Res. 44 (1984) 1483–1488. tion, in: V. Jain, H.C. Goel (Eds.), Selected Topics in Photobiology,
[55] R. Cubeddu, R. Ramponi, G. Bottiroli, Time resolved fluorescence Indian Photobiology Society, 1992, pp. 130–147.spectroscopy of hematoporphyrin in micelles, Chem. Phys. Lett. 128 [74] W.W. Franke, Structure, biochemistry, and functions of the nuclear(1986) 439–442. envelope, Int. Rev. Cytol. Suppl. 4 (1974) 71–236.
[56] R. Cubeddu, R. Ramponi, G. Bottiroli, Disaggregation effects on [75] J.F. Evensen, J. Moan, Photodynamic action and chromosomalhematoporphyrin derivative in the presence of surfactants at differ- damage: a comparison of hematoporphyrin derivative (HpD) andent concentrations: temperature dependence, SPIE 701 (1987) 316– light with X-irradiation, Br. J. Cancer 45 (1982) 456–465.319. [76] A. Maity, W.G. McKenna, R.J. Muschel, The molecular basis for
[57] J. Midander, L. Revesz, The frequency of micronuclei as a measure cell cycle delays following ionizing radiation: review, Radiother.of cell survival in irradiated cell populations, Int. J. Radiat. Biol. 38 Oncol. 31 (1994) 1–13.(1980) 237–242. [77] J. Dahle, H.B. Steen, J. Moan, The mode of cell death induced by
[58] B.S. Dwarakanath, F. Zolzer, S. Chandna, T. Bauch, J.S. Adhikari, photodynamic treatment depends on the cell density, Photochem.W. Muller, C. Streffer, V. Jain, Heterogeneity in 2-deoxy-D-glucose Photobiol. 70 (1999) 363–367.induced modification of radiation response in human tumor cell [78] S. Fournel, L. Genestier, J.-P. Rouault, G. Lizard, M. Flacher, O.lines, Int. J. Radiat. Oncol. Biol. Phys. 50 (2001) 1051–1061. Assossou, J.P. Revillard, Apoptosis without decrease of cell DNA
[59] B.B. Noodt, G.H. Rodal, M. Wainwright, Q. Peng, R. Horobin, J.M. content, FEBS Lett. 367 (1995) 188–192.Nesland, K. Berg, Apoptosis induction by different pathways by [79] S. Hara, H. Dorota Halicka, S. Bruno, J. Gong, F. Traganos, Z.methylene blue derivative and light from mitochondrial sites in V79 Darzynkiewicz, Effect of protease inhibitors on early events ofcells, Int. J. Cancer 75 (1998) 1–8. apoptosis, Exp. Cell Res. 223 (1996) 372–384.
[60] R.R. Perry, W. Matthew, J.B. Mitchell, A. Russo, S. Evans, H.I. [80] S.C. Denstman, L.E. Dillehay, J.R. Williams, Enhanced suscep-Pass, Sensitivity of different human lung cancer histologies to tibility to HPD sensitized phototoxicity and correlated resistance tophotodynamic therapy, Cancer Res. 50 (1990) 4272–4276. trypsin detachment in SV40 transformed IMR-90 cells, Photochem.
[61] T. Christensen, J. Moan, Binding of hematoporphyrin to synchron- Photobiol. 43 (1986) 145–147.ized cells from the line NHIK 3025, Cancer Lett. 9 (1980) 105–110. [81] J. He, M.L. Agarwal, H.E. Larkin, L.R. Friedman, L. Xue, N.L.
[62] V. Jain, F. Khanum, New horizons in the development of photo- Oleinick, The induction of partial resistance to photodynamicdynamic therapy of cancer, in: V. Jain, H. Goel, W. Pohlit (Eds.), therapy by the protooncogene Bcl-2, Photochem. Photobiol. 64Recent Advances in Radiation Oncology, Indian Council of Medical (1996) 845–852.Research, New Delhi, 1990, pp. 275–294. [82] Q. Yu, M. He, N.H. Lee, E.T. Liu, Identification of Myc-mediated
[63] A. Verma, S.L. Facchina, D.J. Hirsch, S.Y. Song, L.F. Dillahey, J.R. death response pathways by microarray analysis, J. Biol. Chem. 277Williams, S.H. Snyder, Photodynamic tumor therapy: mitochondrial (2002) 13059–13066.benzodiazepine receptors as a therapeutic target, Mol. Med. 4 [83] I.R. Radford, Review: initiation of ionizing radiation-induced apop-(1998) 40–45. tosis: DNA damage-mediated or does ceramide have a role?, Int. J.
[64] A. Beurdeley-Thomas, L. Miccoli, S. Oudard, B. Dutrillaux, M.F. Radiat. Biol. 75 (1999) 521–528.Poupon, The peripheral benzodiazepine receptors: a review, J. [84] D.W. Nicholson, N.A. Thornberry, Caspases: killer proteases, TrendsNeurooncol. 46 (2000) 45–56. Biochem. Sci. 22 (1997) 299–306.

120 S. Gupta et al. / Journal of Photochemistry and Photobiology B: Biology 69 (2003) 107–120
[85] L.B. Owen-Schaub, W. Zhang, J.C. Cusack, L.S. Angelo, S.M. M. Wie,V.M. Eng, D.M. Adelman, M.C. Simen, A. Ma, J.A. Golden,Santee, T. Fujiwara, J.A. Roth, A.B. Deisseroth, W.W. Zhang, E. G. Evans, S.J. Korsmayer, G.R. MacGregor, C.B. Thompson, TheKruzel, R. Radinsky, Wild-type human p53 and a temperature- combined functions of proapoptotic Bcl-2 family members bak andsensitive mutant induce Fas/APO-1 expression, Mol. Cell Biol. 15 bax are essential for normal development of multiple tissues, Mol.(1995) 3032–3040. Cell 6 (2000) 1389–1399.
[86] M. Bennet, K. MacDonald, S.-W. Chan, J.P. Luzio, R. Simari, P. [90] R.V. Sinov, Y. Haupt, The cellular response to p53: the decisionWeissberg, Cell surface trafficking of Fas: a rapid mechanism of between life and death, Oncogene 18 (1999) 6145–6157.p53-mediated apoptosis, Science 282 (1998) 290–293. [91] W.-G. Zhang, X.-W. Li, L.-P. Ma, S.-W. Wang, H.-Y. Yang, Z.-Y.
[87] G.S. Wu, T.F. Burns, E.R. McDonald, W. Jiang, R. Meng, I.D. Zhang, Wild-type p53 protein potentiates phototoxicity of 2-BA-2-Krantz, T. Kao, D.D. Gan, J.Y. Zhou, R. Mushcel, S.R. Hamilton, DMHA in HT29 cells expressing endogenous mutant p53, CancerN.B. Spinner, S. Markowitz, Y. Wu, W.S. Diery, Killer /DR5 is a Lett. 138 (1999) 189–195.DNA damage inducible p53 regulated death receptor gene, Nat. [92] H.K. Lorenzo, S.A. Susin, J. Penninger, G. Kroemer, ApoptosisGenet. 17 (1997) 141–143. inducing factor (AIF): a phylogenetically old, caspase-independent
[88] M.C. Wei, W.X. Zong, E.H.Y. Chery, T. Lindsten, V. Panoutsakop- effector of cell death, Cell Death Differ. 6 (1999) 516–524.poulou, A.J. Ross, K.A. Roth, G.R. MacGregor, C.B. Thompson, [93] M. Wu, L.-G. Xu, X. Li, Z. Zhai, H.-B. Shu, AMID, an apoptotic-S.J. Korsmayer, Proapoptotic Bax and Bak: a requisite gateway to inducing factor-homologous mitochondrion-associated protein, in-mitochondrial dysfunction and death, Science 292 (2001) 727–730. duces caspase-independent apoptosis, J. Biol. Chem. 277 (2002)
[89] T. Lindsten, A.J. Ross, A. King, W.X. Zong, J.L. Rathmell, H.A. 25617–25623.Shiels, E. Ulrich, K.G. Waymire, P. Mahar, K. Frauwirth, Y. Chen,
Top Related
Copyright © 2022 FDOKUMEN

