
Ecological limitations on aquatic mosquito predator colonization in the urban environment

Zooarchaeological Evidence for Early Neolithic Colonization at Franchthi Cave (Peloponnese,Greece)Author(s): Natalie D. Munro and Mary C. StinerSource: Current Anthropology, Vol. 56, No. 4 (August 2015), pp. 596-603Published by: The University of Chicago Press on behalf of Wenner-Gren Foundation forAnthropological ResearchStable URL: http://www.jstor.org/stable/10.1086/682326 .
Accessed: 27/08/2015 06:49
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
The University of Chicago Press and Wenner-Gren Foundation for Anthropological Research are collaboratingwith JSTOR to digitize, preserve and extend access to Current Anthropology.
http://www.jstor.org
This content downloaded from 137.99.31.134 on Thu, 27 Aug 2015 06:49:45 AMAll use subject to JSTOR Terms and Conditions

Zooarchaeological Evidence for Early colonization in a few others (Colledge et al. 2013; Connolly
596 Current Anthropology Volume 56, Number 4, August 2015
Neolithic Colonization at Franchthi Cave(Peloponnese, Greece)
et al. 2011; Zeder 2008). Mapping the details of this complexprocess requires high-resolution data sets from many sites sothat the timing, context, and intensity of “neolithization” may
Franchthi Cave is a very large limestone cave on the south-
Natalie D. Munro and Mary C. Stiner
Department of Anthropology, Unit 1176, 354 MansfieldRoad, University of Connecticut, Storrs, Connecticut 06269-1176, USA ([email protected])/School of Anthro-pology, PO Box 210030, University of Arizona, Tucson,Arizona 85721-0030, USA. This paper was submitted 24 VI14, accepted 29 VIII 14, and electronically published 24 VII15.
CA1 Online-Only Material: Supplement A
Franchthi Cave is a cornerstone for research on the mecha-nisms of the forager-producer transition in the southern Bal-kans region. Publications on this site have documented thegeological, artifactual, and macrobotanical records, but de-tailed information on the faunas is lacking. This zooarchae-ological study focuses on the Final Mesolithic and Initial Neo-lithic periods and the question of whether livestock wereadopted as isolated components by late Mesolithic foragersor the site was colonized by people who possessed a fully ag-ricultural lifestyle. Because minor stratigraphic mixing mayunderlie earlier perceptions of a gradual inclusion of domesticanimals into the diet during the Initial Neolithic occupation,we examine this question with the help of zooarchaeologicaland taphonomic data. Changes in taxonomic abundance, con-trasting patterns of burning damage, and caprine (sheep andgoat) demographic and body-size data together reveal an abruptshift from a broad spectrum diet during the Final Mesolithicperiod to a fully fledged domestic economy in the Initial Neo-lithic that centered on caprines, especially sheep. The caprinestransported to Franchthi were small in size, and these ani-mals were intensively managed to optimize meat production.The evidence indicates a wholesale replacement of Mesolithiceconomies by a Neolithic package at Franchthi Cave soon after7000 cal BC.
Few themes have held the attention of archaeologists longeror more intensely than the Neolithic transformation of Eu-rope. Researchers have disputed whether the Neolithic expan-sion from the Middle East was a matter of outright coloniza-tion, adoption of isolated cultural elements (ceramics, domesticplants, or animals) by indigenous foragers, or some combi-nation of the two (e.g., Ammerman and Cavalli-Sforza 1973;Chapman 1994; Halstead 1996; Perlès 2001). Recent consensusfavors a mosaic scenario for many European regions and true
q 2015 by The Wenner-Gren Foundation for Anthropological Research.All rights reserved. 0011-3204/2015/5604-0008$10.00. DOI: 10.1086
/682326This content downloaded from 137.99.31.13All use subject to JSTOR T
be compared within and between regions.Southeastern Europe was among the first regions to be af-
fected by Neolithic expansion. Unfortunately, archaeologi-cal sites that preserve consecutive forager and early Neolithicoccupations are quite rare here and elsewhere, and those withgood biotic preservation are rarer still. The site of FranchthiCave on the Argolid Peninsula of Greece is a remarkable ex-ception. Because it contains an intact sequence of Final Meso-lithic and Initial Neolithic deposits, many archaeologists havelooked to Franchthi Cave to support their interpretations ofthe forager-producer transition in the Aegean area. Halstead(1996) and Séfériadès (2007) raised the possibility of a localdomestication of pig and cattle at Franchthi Cave, while Chap-man (1994) proposed piecemeal adoption of Near Eastern do-mesticates by local hunter-gatherer groups (see also Kotsakis2001). Perlès (2001) sees Franchthi as a strong case for colo-nization by Near Eastern farmers but holds out the possibilitythat local foragers took on a few domesticates during the briefinterval known as the “Initial” Neolithic, followed by a full-on colonization (Perlès, Quiles, and Valladas 2013). Halsteadand Isaakidou (2013) argue, moreover, that the wholesale re-placement of wild economies with domestic ones across theMesolithic-Neolithic boundary does not preclude the adoptionof agriculture by indigenous populations.
A detailed account of the Franchthi faunal sequence is cen-tral to resolving this debate, but little has been published onthe bones from the later periods (but see Payne 1975, 1982; Rose1995). Our study addresses this information gap using the fau-nal series from the FAN and FAS trenches inside the cave.We examine the taphonomic and dietary contexts of theforager-producer transition. Of greatest interest are the con-trasting patterns of burning damage between the two periods,the first appearance and relative importance of livestock, and thepopulation characteristics of and management practices for thecaprines.
Archaeological Background
western edge of the Argolid Peninsula (fig. 1). The cave wassituated up to 6 km inland during the Late Pleistocene, but thesea overtook the coastal plain in front of the cave by 7000 calBC, leaving a rugged landscape of open parkland and deciduous-oak forest behind it (van Andel and Sutton 1987). FranchthiCave was excavated between 1967 and 1979 under the direc-tion of T. W. Jacobsen of Indiana University, in collaborationwith the University of Pennsylvania and the American Schoolof Classical Studies at Athens. The sediments were removedin a series of fine vertical units that followed the natural stra-
4 on Thu, 27 Aug 2015 06:49:45 AMerms and Conditions

ting the water tab
time in the Final Mesolithic (X1), where nongeometric mi-croliths are nearly absent (Perlès 1999). Domestic two-rowed
e by unit indicates an abrupt
Munro and Stiner Early Neolithic Colonization at Franchthi Cave 597
history of human occupation from the Aurignacian to theFinal Neolithic (ca. 39,000–3,000 years ago; Farrand 2000;Douka et al. 2011).
The FAN and FAS areas of the FA trench (fig. 1) containthe most complete and least disturbed Mesolithic-Neolithicsequence (Farrand 2000; Perlès, Quiles, and Valladas 2013),and these sediments were screened systematically through finemesh (2.8 mm and 5 mm). The forager-producer transition isrepresented by the Final Mesolithic (FM) through Initial Neo-lithic (IN) deposits, two discrete strata called Stratum X1 andStratum X2 (Farrand 2000). Stratum X1 is a 40- to 60-cm-thickrocky layer that indicates a rapid deterioration of the cave ceiling.This stratum is separated from X2 by an unconformity and apronounced weathering episode that is visible in Stratum X2and upward into Stratum W. Stratum X2 is a distinctive grayclay layer 35–60 cm thick. Farrand initially proposed a hiatusbetween X1 and X2 of up to 300–600 years, but Perlès, Quiles,and Valladas (2013) estimate 100–200 years based on furtherradiocarbon dating. The new dates place Stratum X2 atca. 7000–6500 cal BC.
C. Perlès defined the cultural phases of the FAN and FASstratigraphies from multiple archaeological data sets (Perlès,Quiles, and Valladas 2013). Her phase assignments largelyagree with Farrand’s (2000) lithostratigraphy, with minor mod-ifications (table 1). Transverse arrowheads appear for the first
Table 1. Stratigraphic data for Initial Neolithic and Final Me
This content downloaded from 137.99.31.13All use subject to JSTOR T
barley (Hordeum distichum) and emmer wheat (Triticum di-coccum) first appear in Stratum X2 (Hansen 1991) when theceramic and lithic assemblages confirm an Initial Neolithicoccupation (Perlès 1999; Vitelli 1993). Like those of the FM,the IN lithics belong to a crude flake–based industry, but withthe addition of pressure-flaked blades and elongated trapezesmanufactured from high-quality raw material (Perlès 1990).Stratum X2 formed slowly (∼8 cm per 100 years) and is in-terpreted as a period of light humanoccupation (Farrand 2000).Neolithic occupations of the Paralia in front of the cave post-date the IN.Cut marks, frequent spiral fractures, and burning and break-
age patterns indicate anthropogenic origins for the faunal as-semblages. The samples from the FAN (2,628 number of iden-tified specimens [NISP]) and FAS (3,127 NISP) trenches are ofsimilar size, but the FAN sequence contains more units at-tributed to the Final Mesolithic period and a higher resolutionfor the transition overall. We therefore focus on the FAN se-quence in graphic representations and qualify these results byreference to the FAS data.
Results
shift from a wild to a domestic animal economy across theFM and the IN. The replacement of red deer with caprinescoincides with a rapid decline in small game use (especiallyfishing), as illustrated for the FAN trench (units 161–164) infigure 2. These changes are also apparent in the FAS trench,including a notable but milder decline in fishing in the IN(CA1 online supplement A, specifically tables A1 and A2).
Red deer (Cervus elaphus) constitute 93% of ungulates as-signed to taxonomic category (NISP) in the FM but drop to!2% in the IN. In contrast, caprines are extremely rare in theFM (!2%), but sheep and goats constitute 95% of ungulates inthe IN, indicating near total replacement of one major meatsource with another. Sheep, a nonnative species, dominate theIN caprine assemblage. Other ungulates in the FM-IN fau-nas include cattle (Bos primigenius) and pig (Sus scrofa), wildpopulations of which existed in the area throughout the Pa-leolithic andMesolithic periods (Payne 1982; Stiner andMunro
lithic layers
7000–6500
tratigraphic163 in FAN
n and Farrand 1987). The excavations pene-10 m below the modern surface before hit-le, and they exposed a rich, discontinuous
Variation in species abundanc
so
Excavation units
it Cultural period FAS FAN
Initial Neolithic 136–145 150–160Hiatus na naFinal Mesolithic 149 164–174
ation units and radiocarbon dates adapted from Perlès, Quiles, and Valladas (2013); lithocombine sediments from Stratum X1 and Stratum X2 (units 146–148 in FAS and units 161tion of this hiatus is 100–200 years (Perlès, Quiles, and Valladas 2013).
ation of Franchthi Cave on the Argolid Penin-nese. Right, plan of the cave interior showinge FAN and FAS trenches.
4e
onrm
Ts a
hu,nd
27Co
And
ug itio
20ns
15
06: 49–
sLithostratigraphic un Estimated years cal BC
Stratum X2Hiatusa
:45 AM
. . .
Stratum X1 7500–7000Note. Relevant excav
data are from Farrand (2000). Mixed unitsa
). na p not applicable.
Estimated duratigraphy (Jacobsetrated as deep as
Figure 1. Left, locsula of the Peloponthe locations of th

598 Current Anthropology Volume 56, Number 4, August 2015
drops from 5% in the FM to 3% in the IN. Small prey animalsconstitute 50% of the FM assemblage in contrast to 14% in theIN. Hares (Lepus europaeus) were important in both periods,but fishing all but drops out with the IN.
Determining the degree of stratigraphic mixing is centralto our investigation, because mixing of noncontemporane-ous materials could mimic the behavioral signature of grad-ual adoption of domesticates. Burning damage (following Stineret al. 1995) is especially useful at Franchthi Cave for distin-guishing the Mesolithic components from earlier and later cul-tural phases, because of the unusually high density of Me-solithic combustion features and burned waste (Stiner andMunro 2011). Unidentified bone specimens (number of un-identified specimens [NUSP]) are prevalent and particularlydiagnostic of burning trends in the FM and IN units. Those in
This content downloaded from 137.99.31.13All use subject to JSTOR T
tified specimens from the Initial Neolithic units are generallylarger and have crisp edges. The overall rate of burning dam-age drops sharply between units 165 and 164 in FAN (fig. 2),from about 69% in the FM to about 40% in the IN; the fre-quency of calcined bone drops from 4% to 1%. This trend isechoed in the identified bones (from 16% in the FM to 4% inthe IN). The incidence of burning damage is substantially andabruptly lower in the IN as a rule.
The mortality profiles for all IN caprines and the mor-phometric data for sheep reveal the domestic status of theseanimals. Survivorship based on bone fusion data (followingZeder 2006) declines steadily from 6 months to 4 years of age(fig. 3; table A3). About 80% of the caprines died before48 months of age. Somewhat different standards must be usedto construct the mortality profile from dental eruption and
e in both the FM (!1%) and the IN (1%) butthe IN deposits, while the frequency of pigs
the secure Mesolithic units havto intensive fragmentation, bu
f faunal variation by excavation unit for the Final Mesolithic to Initial Neolithof ungulates, carnivores, and small game (including fish) in the total assems (NISP; a); relation between red deer and caprine abundance (data in table A1 iof unburned, carbonized, and calcined bones among the unidentified bone fragmens) indicates units that lack unidentified bones. The west and east sections of ued (dashed brackets). Unit 151 also contains some Early Neolithic (FCP1) potithic deposit. Unit 168 is omitted since it represents an episode of profile clean
4e
onrm
Ts a
hu,nd
27Co
Aundi
g tio
201ns
5 0
6: 49:e
nt
i
2011). Cattle are rarincrease slightly in
a rubble-like appearance duerning, and abrasion. Uniden-
Figure 2. Patterns o c sequence in the FAN trench:relative abundance blage as counts of number ofidentified specimen n the CA1 supplement; b); andrelative proportions ents (c). “0 NUSP” (number ofunidentified specim its 161–163 crosscut Levels X1and X2 and are mix ery and therefore is not a truly“clean” Initial Neol ing.
45 AM
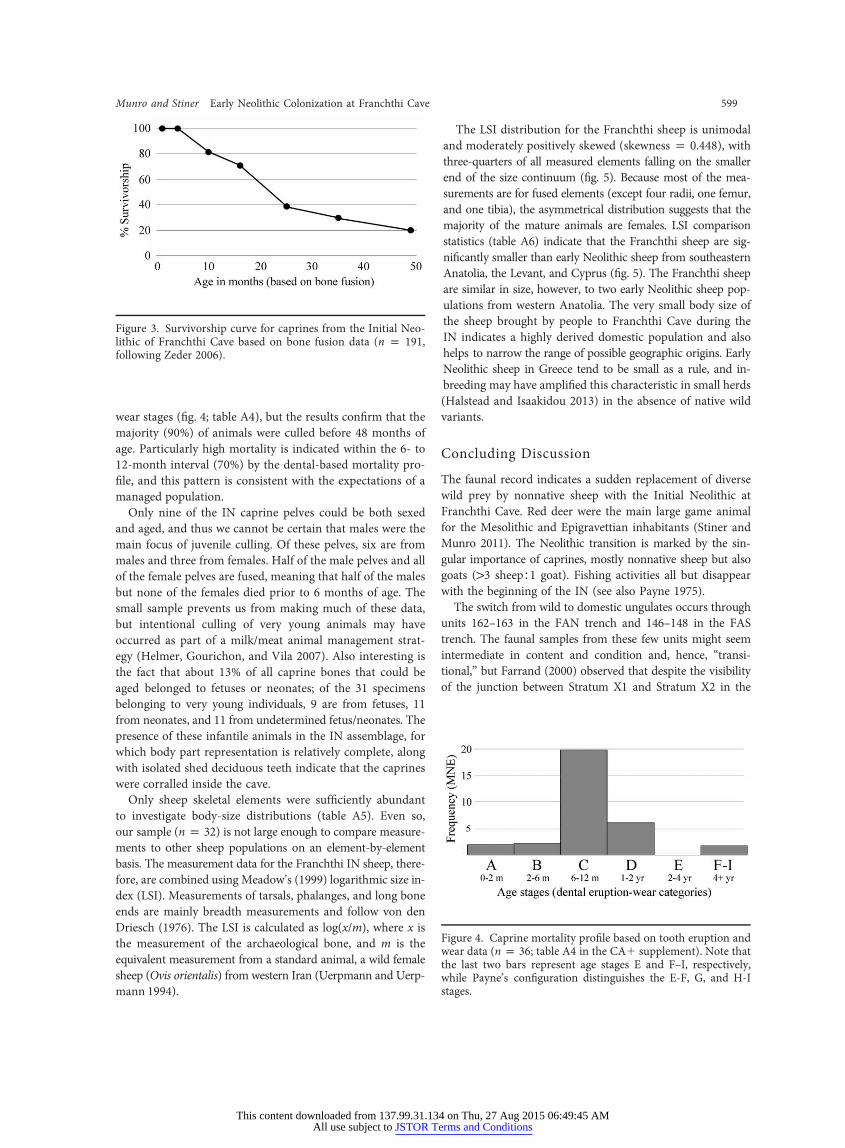
wear stages (fimajority (90%
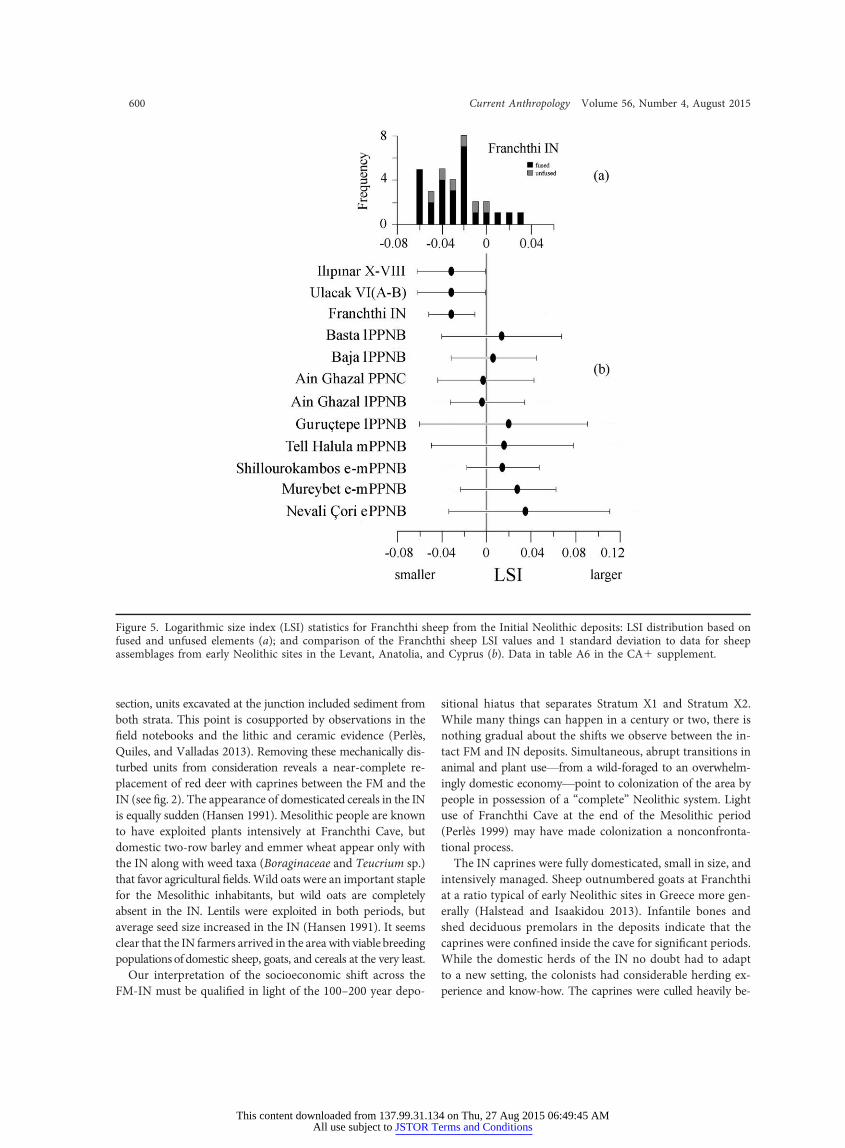
The LSI distribution for the Franchthi sheep is unimodaland moderately positively skewed (skewness p 0.448), with
Munro and Stiner Early Neolithic Colonization at Franchthi Cave 599
age. Particularly high mortality is indicated within the 6- to12-month interval (70%) by the dental-based mortality pro-file, and this pattern is consistent with the expectations of amanaged population.
Only nine of the IN caprine pelves could be both sexedand aged, and thus we cannot be certain that males were themain focus of juvenile culling. Of these pelves, six are frommales and three from females. Half of the male pelves and allof the female pelves are fused, meaning that half of the malesbut none of the females died prior to 6 months of age. Thesmall sample prevents us from making much of these data,but intentional culling of very young animals may haveoccurred as part of a milk/meat animal management strat-egy (Helmer, Gourichon, and Vila 2007). Also interesting isthe fact that about 13% of all caprine bones that could beaged belonged to fetuses or neonates; of the 31 specimensbelonging to very young individuals, 9 are from fetuses, 11from neonates, and 11 from undetermined fetus/neonates. Thepresence of these infantile animals in the IN assemblage, forwhich body part representation is relatively complete, alongwith isolated shed deciduous teeth indicate that the caprineswere corralled inside the cave.
Only sheep skeletal elements were sufficiently abundantto investigate body-size distributions (table A5). Even so,our sample (np 32) is not large enough to compare measure-ments to other sheep populations on an element-by-elementbasis. The measurement data for the Franchthi IN sheep, there-fore, are combined using Meadow’s (1999) logarithmic size in-dex (LSI). Measurements of tarsals, phalanges, and long boneends are mainly breadth measurements and follow von denDriesch (1976). The LSI is calculated as log(x/m), where x isthe measurement of the archaeological bone, and m is theequivalent measurement from a standard animal, a wild femalesheep (Ovis orientalis) from western Iran (Uerpmann and Uerp-mann 1994).
This content downloaded from 137.99.31.13All use subject to JSTOR T
three-quarters of all measured elements falling on the smallerend of the size continuum (fig. 5). Because most of the mea-surements are for fused elements (except four radii, one femur,and one tibia), the asymmetrical distribution suggests that themajority of the mature animals are females. LSI comparisonstatistics (table A6) indicate that the Franchthi sheep are sig-nificantly smaller than early Neolithic sheep from southeasternAnatolia, the Levant, and Cyprus (fig. 5). The Franchthi sheepare similar in size, however, to two early Neolithic sheep pop-ulations from western Anatolia. The very small body size ofthe sheep brought by people to Franchthi Cave during theIN indicates a highly derived domestic population and alsohelps to narrow the range of possible geographic origins. EarlyNeolithic sheep in Greece tend to be small as a rule, and in-breeding may have amplified this characteristic in small herds(Halstead and Isaakidou 2013) in the absence of native wildvariants.
Concluding Discussion
The faunal record indicates a sudden replacement of diversewild prey by nonnative sheep with the Initial Neolithic atFranchthi Cave. Red deer were the main large game animalfor the Mesolithic and Epigravettian inhabitants (Stiner andMunro 2011). The Neolithic transition is marked by the sin-gular importance of caprines, mostly nonnative sheep but alsogoats (13 sheep∶1 goat). Fishing activities all but disappearwith the beginning of the IN (see also Payne 1975).
The switch from wild to domestic ungulates occurs throughunits 162–163 in the FAN trench and 146–148 in the FAStrench. The faunal samples from these few units might seemintermediate in content and condition and, hence, “transi-tional,” but Farrand (2000) observed that despite the visibilityof the junction between Stratum X1 and Stratum X2 in the
g. 4; table A4), but the results confirm that the) of animals were culled before 48 months of
ivorship curve for caprines from the Initial Neo-hthi Cave based on bone fusion data (n p 191,er 2006).
Figure 4. Caprine mortality profile bwear data (n p 36; table A4 in the CAthe last two bars represent age stagwhile Payne’s configuration distingustages.
4 oerm
n Ts
huand
, 2 C
7 Aon
ugditi
20on
15s
06
:4 9:4 5 Aa
e
Figure 3. Survlithic of Francfollowing Zed
sed on tooth eruption and1 supplement). Note thats E and F–I, respectively,ishes the E-F, G, and H-I
M
both or two, there is
600 Current Anthropology Volume 56, Number 4, August 2015
field notebooks and the lithic and ceramic evidence (Perlès,Quiles, and Valladas 2013). Removing these mechanically dis-turbed units from consideration reveals a near-complete re-placement of red deer with caprines between the FM and theIN (see fig. 2). The appearance of domesticated cereals in the INis equally sudden (Hansen 1991). Mesolithic people are knownto have exploited plants intensively at Franchthi Cave, butdomestic two-row barley and emmer wheat appear only withthe IN along with weed taxa (Boraginaceae and Teucrium sp.)that favor agricultural fields.Wild oats were an important staplefor the Mesolithic inhabitants, but wild oats are completelyabsent in the IN. Lentils were exploited in both periods, butaverage seed size increased in the IN (Hansen 1991). It seemsclear that the IN farmers arrived in the area with viable breedingpopulations of domestic sheep, goats, and cereals at the very least.
Our interpretation of the socioeconomic shift across theFM-IN must be qualified in light of the 100–200 year depo-
This content downloaded from 137.99.31.13All use subject to JSTOR T
nothing gradual about the shifts we observe between the in-tact FM and IN deposits. Simultaneous, abrupt transitions inanimal and plant use—from a wild-foraged to an overwhelm-ingly domestic economy—point to colonization of the area bypeople in possession of a “complete” Neolithic system. Lightuse of Franchthi Cave at the end of the Mesolithic period(Perlès 1999) may have made colonization a nonconfronta-tional process.
The IN caprines were fully domesticated, small in size, andintensively managed. Sheep outnumbered goats at Franchthiat a ratio typical of early Neolithic sites in Greece more gen-erally (Halstead and Isaakidou 2013). Infantile bones andshed deciduous premolars in the deposits indicate that thecaprines were confined inside the cave for significant periods.While the domestic herds of the IN no doubt had to adaptto a new setting, the colonists had considerable herding ex-perience and know-how. The caprines were culled heavily be-
on, units excavated at the junction included sediment fromstrata. This point is cosupported by observations in the
sitional hiatus that separates Stratum X1While many things can happen in a centur
re 5. Logarithmic size index (LSI) statistics for Franchthi sheep from the Initial Neolithic deposits: LSI distand unfused elements (a); and comparison of the Franchthi sheep LSI values and 1 standard deviationblages from early Neolithic sites in the Levant, Anatolia, and Cyprus (b). Data in table A6 in the CA1 s
4e
onrm
Ts a
hu,nd
27Co
And
ug itio
20ns
15
06: 49 :45 AMay
secti nd Stratum X2.
Figu ribution based onfused to data for sheepassem upplement.

tween 6 and 18 months of age, probably targeting young malesto a large degree, and few animals of either sex survived past
were present in the southern Levant by 8000 cal BC (Martin andEdwards 2013), and domestic sheep were transported by humans
Munro and Stiner Early Neolithic Colonization at Franchthi Cave 601
4 years of age. The off-take pattern fits a typical meat-focusedproduction model, with possible milking of some of the fe-males (type A; Helmer, Gourichon, and Vila 2007).
It is perhaps not surprising that the Franchthi IN sheep aresmaller than the sheep from Early Phase B (8050–7750 cal BC)at Shillourokambos on Cyprus (Vigne 2011) andmuch smallerthan the earliest managed sheep populations from the earlyPre-Pottery Neolithic B (PPNB; 8550 cal BC) of Nevali Çori,Dja’de, andCaferHöyük in upperMesopotamia (Helmer 2008;Vigne 2013) and the early PPNB sheep of Aswad in the Levant(Helmer and Gourichon 2008). These populations were in theearly stages of animal management and unlike the Franchthisheep had not undergone much, if any, body-size diminution.It is interesting that the IN sheep from Franchthi Cave are alsosmaller than later PPNB and Pre-Pottery Neolithic C sheepof the northern and southern Levant (Ain Ghazal, Baja, andBasta) and southeastern Anatolia (Guruçtepe) that had al-ready experienced some diminution and only slightly predatethe Franchthi sample (Martin and Edwards 2013; Peters et. al.1999). Sheep from Ulucak and Ilıpınar (6800 cal BC) in west-ern Anatolia are most similar in size to those from Franchthi,cases that are contemporaneous to or slightly postdate theFranchthi Cave IN (Cakırlar 2012).
The exceptionally small body size of the Franchthi sheep,especially in relation to previous managed populations in theearly stages of domestication, indicates that this populationhad already experienced intensive selection prior to its trans-port to the southern Argolid. Wild goats are absent from theFM assemblage at Franchthi, but they are present in smallnumbers in earlier Paleolithic layers (Stiner and Munro 2011).Few measurements are available for the Franchthi goats ofthe IN, but they are also quite small and likely arrived underthe same conditions as the sheep. The pig and cattle remainsfrom the IN deposit are more difficult to interpret, becausethey are uncommon in the assemblages and wild variants ex-isted in the region prior to the Neolithic period (Stiner andMunro 2011). The percentages of cattle and pig remains areless than 5% of the total ungulate assemblage in either the FMor the IN.
At least three pathways have been proposed for Neolithicexpansion into Greece: (1) an overland route originating fromwestern Anatolia and spreading into northern Greece via theBalkans (Thissen 2005; Weninger et al. 2006); (2) a maritimeroute originating fromwestern Anatolia and dispersing acrossthe Aegean; and (3) a distinct maritime route from southernAnatolia or the Levant to the northeastern Greek coast and thesoutheastern Peloponnese (Broodbank 1999; Çakırlar 2012;Özdoğan 2011; Perlès 2001). Wild sheep are not native toGreece, and thus their presence in archaeofaunas indicatesdeliberate introduction by humans. Sheep management practicesfirst appeared in Anatolia and the upper Euphrates region be-tween 8500 and 8200 cal BC (Hongo et al. 2009; Peters, von denDriesch, and Helmer 2005; Stiner et al. 2014). Domestic sheep
This content downloaded from 137.99.31.13All use subject to JSTOR T
to the island of Cyprus by 8000–7800 cal BC (Vigne 2013). TheIN occupation at Franchthi postdates the latter cases by roughly1,000 years (7000–6500 cal BC; Perlès, Quiles, and Valladas 2013),and thus the fully domestic status of the Franchthi sheep is un-surprising.
The Franchthi IN is generally coeval to the earliest Neo-lithic sites of western Anatolia and northern Greece (reviewedby Perlès, Quiles, and Valladas 2013) and to the earliest Neo-lithic occupations at Knossos on the island of Crete (7040–6770 cal BC; Efstratiou et al. 2004), a settlement that couldonly have been established by maritime voyagers. Coeval orslightly later populations of small-bodied sheep in westernAnatolia (Buitenhuis 1995; Çakırlar 2012) may mean that farm-ing entered western Anatolia and the Aegean as part of thesame process. Although the four primary animal domesticates(sheep, goat, pig, and cattle) may have arrived to Franchthi andwestern Anatolia as an integrated management package, pigand cattle had greater importance at early Neolithic sites inwestern Anatolian (Ulucak and Ilıpınar). This difference inpig and cattle importance may indicate different trajectories ofdispersal for the two regions or greater difficulties transportinglarge-bodied animals by sea.
Regardless of the route of maritime expansion, the some-what later settlement of the Greek Mainland ca. 6500 BC(Perlès, Quiles, and Valladas 2013; Reingruber and Thissen2009) and the general settlement history of the eastern Med-iterranean indicate that the IN settlement at Franchthi wasmade possible by maritime travel (Perlès 2001). There is aninteresting paradox in the Aegean area. Sophisticated seafaringtechnology first developed in the Aegean during the Mesolithicperiod, as demonstrated by the transport of obsidian from theisland of Melos to the Greek mainland and to Franchthi Cave,in particular (Perlès 2001). The presence of early Neolithic siteson Crete and Cyprus indicates that seafaring was also wellwithin the capacity of Neolithic colonists, but their economywas largely terrestrial in focus.
Although other Initial Neolithic settlements may still be dis-covered on the Argolid Peninsula, the low density of earlyNeolithic sites and rugged terrain of the southern Peloponnese(see fig. 1) suggest a situation of relative terrestrial isolation forthe first Neolithic settlement at Franchthi (Runnels and vanAndel 1987; van Andel and Sutton 1987). This IN communityseems to have been a kind of outpost, whose social networkwas maintained more by sea than by land (Thissen 2005).Oddly, the IN colonists were steadfast in their adherenceto a terrestrial economy and initially made little use of therich Aegean fishery, in contrast to the Mesolithic (Stiner andMunro 2011) and the later Neolithic occupants (Payne 1975;Rose 1995). The low density of cultural remains in the IN de-posits suggests a small community (Perlès, Quiles, and Valladas2013). It is not until the Early Neolithic phase that an archae-ologically discernible village was constructed on the talus(Paralia) in front of the cave (van Andel and Sutton 1987). If
4e
onrm
Ts a
hu,nd
27 Con
Aug 2015 06:49:45 AMditions

one may speculate, a relation between Mesolithic seafarersand expanding Neolithic populations could have existed where
Farrand, W. R. 2000. Depositional history of Franchthi Cave: stratigraphy,sedimentology, and chronology. In Excavations at Franchthi Cave, Greece,fasc. 7. Bloomington: Indiana University Press.
602 Current Anthropology Volume 56, Number 4, August 2015
travel was easier by sea than land (Broodbank 2006). Alterna-tively, some groups in both cultures may have been heavily in-vested in navigation technology. A completely independent in-vention of maritime travel seems unlikely, and hence maritimetravel could have been a main axis of forager-producer inter-action during this transitional period.
Acknowledgments
We gratef knowl htful a e
of Catherine Perlès and Karen Vitelli for this study. We alsoowe special thanks to W. R. (Bill) Farrand and MargaritaKoumouzelis for inspirational discussions in the planning ofthis work. We thank the staff of the Nafplio Museum, partic-ularly Angeliki Kossyva, who kindly accommodated our re-search over several years at the Apotheke in Nafplio. Jean-DenisVigne, Hijlke Buitenhuis, Çanan Çakırlar, Ben Arbuckle, andMark Rose provided valuable feedback and comparative data,some of it unpublished. Thanks to Cyprian Broodbank for help-ful comments. We also thank the American School of ClassicalStudies in Athens and especially Sherry Fox (formerly of theWeiner Lab) for their support. This research was made possibleby grants to N. D. Munro and M. C. Stiner from the Wenner-Gren Foundation (7975), to M. C. Stiner from the National Sci-ence Foundation (BCS-0410654), and to N. D. Munro from theUniversity of Connecticut Faculty Grant Program.References CitedAmmerman, A. J., and L. Cavalli-Sforza. 1973. A population model for the
diffusion of early farming in Europe. In The explanation of culture change.C. Renfrew, ed. Pp. 343–357. London: Duckworth.
Broodbank, C. 1999. Colonization and configuration in the insular Neolithicof the Aegean. In Neolithic society in Greece. P. Halstead, ed. Pp. 15–41.Sheffield: Sheffield Studies in Aegean Archaeology.
———. 2006. The origins and early development of Mediterranean mari-time activity. Journal of Mediterranean Archaeology 19:199–230.
Buitenhuis, H. 1995. The faunal remains. In The Ilıpınar Excavations 1: fiveseasons of fieldwork in NW Anatolia 1987–91. J. Roodenberg, ed. Pp. 151–157. Istanbul: Nederlands Historisch-Archaeologische Instituut.
Çakırlar, C. 2012. The evolution of animal husbandry in Neolithic central-west Anatolia: the zooarchaeological record from Ulucak Höyük (c. 7040–5660 cal. BC, Izmir, Turkey). Anatolian Studies 62:1–33.
Chapman, J. 1994. The origins of farming in southeast Europe. PréhistoireEuropéenne 6:133–156.
Colledge, S., J. Conolly, K. Dobney, K. Manning, and S. Shennan, eds. 2013.The origins and spread of domestic animals in southwest Asia and Europe.Walnut Creek, CA: Left Coast.
Conolly, J., S. Colledge, K. Dobney, J.-D. Vigne, J. Peters, B. Stopp, K. Manning,and S. Shennan. 2011. Meta-analysis of zooarchaeological data from SW Asiaand SE Europe provides insight into the origins and spread of animal hus-bandry. Journal of Archaeological Science 38(3):538–545.
Douka, K., C. Perlés, H. Valladas, M. Vanhaeren, and R. E. M. Hedges. 2011.Franchthi Cave revisited: the age of the Aurignaican in southeastern Europe.Antiquity 85:1131–1150.
Efstratiou, N., A. Karetsou, E. Banou, and D. Margomenou. 2004. The Neolithicsettlement of Knossos: new light on an old picture. In Knossos: palace, city,state, vol. 12 of British School at Athens Studies. G. Cadogan, E. Hatzaki, andE. Vasilakis, eds. Pp. 43–49. London: British School at Athens.
This content downloaded from 137.99.31.13All use subject to JSTOR T
Halstead, P. 1996. The development of agriculture and pastoralism in Greece:when, how, who and what? In The origins and spread of agriculture andpastoralism in Eurasia. D. R. Harris, ed. Pp. 296–309. London: UniversityCollege London Press.
Halstead, P., and V. Isaakidou. 2013. Early stock-keeping in Greece. In Theorigins and spread of domestic animals in Southwest Asia and Europe.S. Colledge, J. Conolly, K. Dobney, K. Manning, and S. Shennan, eds.Pp. 129–144. Walnut Creek, CA: Left Coast.
Hansen, J. M. 1991. The palaeoethnobotany of Franchthi Cave. In Excavationsat Franchthi Cave, Greece, fasc. 7. Bloomington: Indiana University Press.
Helmer, D. 2008. Révision de la faune de Cafer Höyük (Malatya, Turquie):apports des méthodes de l’analyse des mélanges et de l’analyse de Kernel àla mise en évidence de la domestication. In Archaeozoology of the Near EastVIII: proceedings of the 8th International Symposium on the archaeozoologyof southwestern Asia and adjacent areas. E. Vila, L. Gourichon, A. M.Choyke, and H. Buitenhuis, eds. Pp. 169–195. Lyon: Maison de l’Orient etde la Méditerranée.
Helmer, D., and L. Gourichon. 2008. Premières données sur les modalités desubsistance à Tell Aswad (Syrie, PPNB moyen et récent, Néolithique céra-mique ancien). In Archaeozoology of the Near East VIII: proceedings of the8th International Symposium on the archaeozoology of southwestern Asia andadjacent areas. E. Vila, L. Gourichon, A. M. Choyke, and H. Buitenhuis, eds.Pp. 120–151. Lyon: Maison de l’Orient et de la Méditerranée.
Helmer, D., L. Gourichon, and E. Vila. 2007. The development of the exploi-tation of products from Capra and Ovis (meat, milk, and fleece) from thePPNB to the Early Bronze Age in the northern Near East (8700–9200 calBP). Anthropozoologica 42(2):41–69.
Hongo, H., J. Pearson, B. Öksüz, and G. Ilgezdi. 2009. The process of ungulatedomestication at Cayönü, southeastern Turkey: a multidisciplinary approachfocusing on Bos sp. and Cervus elaphus. Anthropozoologica 44:63–78.
Jacobsen, T. W., and W. R. Farrand. 1987. Franchthi Cave and Paralia: maps,plans, and sections. In Excavations at Franchthi Cave, Greece, fasc. 1. Bloom-ington: Indiana University Press.
Kotsakis, K. 2001. Mesolithic to Neolithic in Greece: continuity, disconti-nuity or change of course? Documenta Praehistorica 28:63–73.
Martin, L., and Y. Edwards. 2013. Diverse strategies: evaluating the appearanceand spread of domestic caprines in the southern Levant. In The origins andspread of domestic animals in southwest Asia and Europe. S. Colledge, J.Conolly, K. Dobney, K. Manning, and S. Shennan, eds. Pp. 49–82. WalnutCreek, CA: Left Coast.
Meadow, R. H. 1999. The use of size index scaling techniques for research onarchaeozoological collections from the Middle East. In Historia animaliumex ossibus: Beiträge zur Paläoanatomie, Archäologie, Ägyptologie, Ethnologieund Geschichte der Tiermedizin. C. Becker, H. Manhart, J. Peters, and J.Schibler, eds. Pp. 285–300. Rahden: Marie Leidorf.
Özdoğan, M. 2011. Archaeological evidence on the westward expansion offarming communities from eastern Anatolia to the Aegean and the Balkans.Current Anthropology 52(S4):S415–S430.
Payne, S. 1975. Faunal change at Franchthi Cave from 20,000 B.C.–3000 B.C.In Archaeozoological studies. A. T. Clason, ed. Pp. 120–131. Amsterdam:North Holland.
———. 1982. Faunal evidence for environmental/climatic change at FranchthiCave (southern Argolid, Greece), 25,000 B.P.–5000 B.P. preliminary results. InPalaeoclimates, palaeoenvironments and human communities in the easternMediterranean region in later prehistory. J. L. Bintliff and W. van Zeist, eds.Pp. 133–137. Oxford: British Archaeological Reports.
Perlès, C. 1990. Les industries lithiques tailées de Franchithi (Argolide, Grèce).Tome II les industries duMésolithique et duNéolithique Initial. InExcavationsat Franchthi Cave, Greece, fasc. 7. Bloomington: Indiana University Press.
———. 1999. Long-term perspectives on the occupation of the FranchthiCave: continuity and discontinuity. The Palaeolithic archaeology of Greeceand adjacent areas. G. N. Bailey, E. Adam, E. Panagopoulou, C. Perlès, andK. Zachos, eds. Pp. 311–318. Proceedings of the ICOPAG Conference, Ioan-nina. Athens: British School at Athens.
———. 2001. The Early Neolithic in Greece: the first farming communities inEurope. Cambridge: Cambridge University Press.
Perlès, C., A. Quiles, and H. Valladas. 2013. Early seventh-millenium AMS datesfromdomestic seeds in the Initial Neolithic at Franchthi Cave (Argolid, Greece).Antiquity 87:1001–1015.
4 onerm
Thu, 27 Aug 2015 0s and Conditions
6:49:45 AM
ully ac
edge the tireless, thoug ssistanc
Peters, J., A. von den Driesch, and D. Helmer. 2005. The upper Euphrates-Tigrisbasin: cradle of agro-pastoralism. In First steps of animal domestication:new archaeozoological approaches. J.-D. Vigne, J. Peters, and D. Helmer, eds.
Uerpmann, M., and H.-P. Uerpmann. 1994. Animal bone finds from Exca-vation 520 at Qala’at al-Bahrain. In The northern city wall and the Islamicfortress, vol. 1 of Qala’at al-Bahrain. F. Holland and H. H. Anderson, eds.
Munro and Stiner Early Neolithic Colonization at Franchthi Cave 603
Pp. 96–134. Oxford: Oxbow.Peters, J., A. von den Driesch, D. Helmer, and M. Saña Segui. 1999. Early animal
husbandry in the Northern Levant. Paléorient 19:27–48.Reingruber, A., and L. Thissen. 2009. Depending on 14C data: chronological
frameworks in the Neolithic and Chalcolithic of southeastern Europe. Ra-diocarbon 51:751–770.
Rose, M. 1995. Fishing at Franchthi Cave, Greece: changing environmentsand patterns of exploitation. Old World Archaeology Newsletter 18:21–26.
Runnels, C. N., and T. H. Van Andel. 1987. The evolution of settlement in thesouthern Argolid, Greece: an economic explanation. Hesperia 56(3):303–334.
Séfériadès, M. 2007. Complexity of the processes of Neolithization: traditionand modernity of the Aegean world at the dawn of the Holocene period(11–9 kyr). Quaternary International 167–168:177–185.
Stiner, M. C., H. Buitenhuis, G. Duru, N. D. Munro, S. Mentzer, N. Pőllath,G. Tsartidou, and M. Özbaşaran. 2014. A forager-herder trade off, frombroad-spectrum hunting to sheep management at Aşıklı Höyük, Turkey.Proceedings of the National Academy of Sciences 111(23):8404–8409.
Stiner, M. C., and N. D. Munro. 2011. On the evolution of diet and landscapeduring the Upper Paleolithic through Mesolithic at Franchthi Cave(Peloponnese, Greece). Journal of Human Evolution 60:618–636.
Stiner, M. C., S.Weiner, O. Bar-Yosef, and S. L. Kuhn. 1995. Differential burning,fragmentation, and preservation of archaeological bone. Journal of Archaeo-logical Science 22:223–237.
Thissen, L. 2005. Coming to grips with the Aegean in prehistory: an outlineof the temporal framework. In How did farming reach Europe? Anatolian-European relations from the second half of the 7th through the first half ofthe 6th millennium cal BC. C. Licther, ed. Pp. 29–40. Byzas 2. Istanbul:Deutsches archäeologisches Institut.
This content downloaded from 137.99.31.13All use subject to JSTOR T
Pp. 417–444. Aarhus: Jutland Archaeological Society.van Andel, T. H., and S. B. Sutton. 1987. Landscape and people of the Franchthi
region. InExcavations at Franchthi Cave, Greece, fasc. 2. Bloomington: IndianaUniversity Press.
———. 2011. Annexes archéozoologiques: les données brutes des analyses.In Shillourokambos: un établissement néolithique pré-céramique à Chypre:les fouilles du Secteur 1. J. Guilaine, F. Briois, and J.-D. Vigne, eds. Pp. 1135–1157. Paris: Editions Errance/Ecole Française d’ Athènes.
Vigne, J.-D. 2013. Domestication process and domestic ungulates: new ob-servations from Cyprus. In The origins and spread of domestic animals insouthwest Asia and Europe. S. Colledge, J. Conolly, K. Dobney, K. Man-ning, and S. Shennan, eds. Pp. 115–128. Walnut Creek, CA: Left Coast.
Vitelli, K. D. 1993. Classification and ceramic phases 1 and 2, vol. 1 of FranchthiNeolithic pottery. Excavations at Franchthi Cave, Greece, fasc. 8. Bloomington:Indiana University Press.
Von den Driesch, A. 1976. A guide to the measurement of animal bones fromarchaeological sites. Peabody Museum Bulletin 1. Cambridge: PeabodyMuseum of Archaeology and Ethnology, Harvard University.
Weninger, B., E. Alram-Stern, E. Bauer, L. Clare, U. Danzeglocke, O. Joris, C.Kubatski, G. Rollefson, H. Todorova, and T. van Andel. 2006. Climate forcingdue to the 8200 cal yr BP event observed at Early Neolithic sites in the easternMediterranean. Quaternary Research 66:401–420.
Zeder, M. A. 2006. Reconciling rates of long bone fusion and tooth eruptionand wear in sheep (Ovis) and goat (Capra). In Recent advances in ageingand sexing animals. D. Ruscillo, ed. Pp. 87–118. Oxford: Oxbow.
———. 2008. Domestication and early agriculture in the MediterraneanBasin: origins, diffusion, and impact. Proceedings of the National Academyof Sciences 105:11597–11604.
4 oerm
n Thu, 27 As and Con
ug 2015 0ditions
6:49:45 AM

CA1 Supplement A from Munro and Stiner, “ZooarchaeologicalEvidence for Early Neolithic Colonization at Franchthi Cave(Peloponnese, Greece)”(Current Anthropology, vol. 56, no. 4, p. 000)
q 2015 by The Wenner-Gren Foundation for Anthropological Research. All rights reserved. DOI: 10.1086/682326
1

Tab
leA1.
Num
berof
identified
specim
ens(N
ISP)fortaxonomic
groups
intheFA
Ntrench
byexcavatio
nunit
FAN
unit
Testudo
sp.
Maurm
ys/
Emys
sp.
Chelonid
indet.
Sparidae
Dicentrarchus
labrax
Thunnus
thynnys
Sphyraena
sphyraena
Pisces
indet.
Medium
bird
Erinacaeus
europaeus
Lepus
europaeus
Small
mam
mal
150
00
00
00
00
10
32
151
00
00
00
00
11
712
152
00
00
00
00
10
43
153
00
00
00
00
01
01
154
00
00
00
00
12
02
155
00
00
00
00
00
07
156
00
00
00
00
00
23
157
00
00
00
00
00
00
158
00
00
00
01
00
11
159
00
00
00
00
01
35
160
00
00
00
00
011
24
161
00
00
00
00
02
132
162
00
00
00
00
11
256
163
01
10
00
00
30
5931
164
00
02
01
111
01
51
165
00
00
00
114
00
43
166
00
00
11
520
00
42
167
00
00
11
239
01
41
169
00
00
04
424
10
42
170
00
034
011
269
01
63
171
00
015
00
07
00
20
172
08
192
133
571
00
3417
173
00
00
020
00
00
30
174
01
038
059
46
20
165
TOTA
L0
102
181
3130
24262
1122
201
113
CA1 Supplement A from Munro and Stiner, Zooarchaeological Evidence for Early Neolithic Colonization at Franchthi Cave (Peloponnese, Greece)
2

FAN
unit
Cervidae
indet.
Cervus
elaphus
Bos
primigenius
Sus
scrofa
Capra
hircus
Capra/
Ovis
Ovis
aries
Small
ungulate
Medium
ungulate
Large
ungulate
Very
large
ungulate
Medium/
large
ungulate
150
20
00
19
013
40
10
151
20
04
018
1951
63
00
152
00
08
241
340
30
30
153
00
00
03
19
51
01
154
10
10
041
1531
20
00
155
00
00
013
012
31
20
156
21
01
029
230
30
10
157
00
00
02
25
00
00
158
01
10
110
216
32
00
159
00
01
115
114
00
00
160
10
02
022
214
21
00
161
12
00
031
120
10
00
162
09
02
024
528
51
10
163
729
03
354
637
917
10
164
07
03
00
05
28
00
165
18
00
00
03
06
00
166
16
00
01
03
51
00
167
48
01
01
04
310
00
169
110
00
00
05
22
00
170
219
01
02
04
829
00
171
02
03
00
02
16
00
172
4106
15
00
014
22127
10
173
130
00
00
01
619
00
174
953
12
01
02
951
019
TOTA
L39
291
436
8317
59363
104
285
1020
Tab
leA1(Contin
ued)
CA1 Supplement A from Munro and Stiner, Zooarchaeological Evidence for Early Neolithic Colonization at Franchthi Cave (Peloponnese, Greece)
3

FAN
unit
Testudo
sp.
Maurm
ys/
Emys
sp.
Chelonid
indet.
Sparidae
Dicentrarchus
labrax
Thunnus
thynnys
Sphyraena
sphyraena
Pisces
indet.
Medium
bird
Erinacaeus
europaeus
Lepus
europaeus
Small
mam
mal
FAN
unit
Meles
meles
Felis
silvestris
Lynx
lynx
Vulpes
vulpes
Carnivora
indet.
Medium
mam
mal
Economic
taxa
total
150
00
01
01
38151
00
06
50
135
152
00
02
20
112
153
00
03
00
25154
00
07
00
103
155
00
00
12
41156
00
05
01
80157
00
00
00
9158
01
03
00
43159
00
00
00
41160
00
03
10
65161
00
02
00
75162
00
02
00
110
163
02
013
21
279
164
00
02
00
49165
00
02
00
42166
00
02
10
53167
00
02
00
82169
00
03
01
63170
00
17
30
202
171
00
01
00
39172
00
121
01
565
173
00
01
00
81174
10
115
10
296
TOTA
L1
33
103
167
2628
Note.
Unit168isom
itted
sinceitrepresentsan
episodeof
profi
lecleaning.Units161–
163aremixed
units
that
combine
sedimentsfrom
Stratum
X1andStratum
X2.
Tab
leA1(Contin
ued)
CA1 Supplement A from Munro and Stiner, Zooarchaeological Evidence for Early Neolithic Colonization at Franchthi Cave (Peloponnese, Greece)
4

Tab
leA2.
Num
berof
identified
specim
ensfortaxonomic
groups
inTrenchFA
Sby
excavatio
nunit
FAS
unit
Testudo
sp.
Maurm
ys/
Emys
sp.
Chelonid
indet.
Sparidae
Dicentrarchus
labrax
Thunnus
thynnys
Sphyraena
sphyraena
Pisces
indet.
Medium
bird
Erinacaeus
europaeus
Lepus
europaeus
Small
mam
mal
135
00
00
00
03
00
10
136
00
00
02
03
00
00
137
00
00
00
012
00
20
138
00
00
00
08
02
20
139
00
00
00
013
00
30
140
00
04
00
06
00
193
141
00
00
01
02
00
00
142
00
00
00
04
00
10
143
00
00
00
015
00
47
144
00
02
01
015
01
53
145
10
04
013
081
00
288
146
03
34
054
5498
326
7513
147
02
02
018
069
02
171
148
00
01
016
2122
13
196
149
00
01
00
033
10
110
TOTA
L1
53
180
105
7884
534
187
41
FAS
unit
Cervidae
indet.
Cervus
elaphus
Bos
primigenius
Sus
scrofa
Capra
hircus
Capra/
Ovis
Ovis
aries
Small
ungulate
Medium
ungulate
Large
ungulate
Verylarge
ungulate
Medium/large
ungulate
135
00
33
016
00
31
00
136
00
00
631
00
34
20
137
01
20
022
00
72
20
138
00
21
117
00
45
10
139
00
20
140
10
52
20
140
00
67
0226
20
23
30
141
00
00
014
00
20
00
142
00
00
118
30
40
00
143
01
04
1328
51
010
110
144
03
13
2104
80
133
10
145
05
03
1189
70
76
00
146
053
011
1089
142
8052
02
147
09
02
0126
60
58
00
148
125
05
06
01
1423
00
149
05
00
00
00
43
00
TOTA
L1
102
1639
231226
464
153
122
222
CA1 Supplement A from Munro and Stiner, Zooarchaeological Evidence for Early Neolithic Colonization at Franchthi Cave (Peloponnese, Greece)
5

FAS
unit
Testudo
sp.
Maurm
ys/
Emys
sp.
Chelonid
indet.
Sparidae
Dicentrarchus
labrax
Thunnus
thynnys
Sphyraena
sphyraena
Pisces
indet.
Medium
bird
Erinacaeus
europaeus
Lepus
europaeus
Small
mam
mal
FAS
unit
Meles
meles
Felis
silvestris
Lynx
lynx
Vulpes
vulpes
Carnivora
indet.
Medium
mam
mal
Economic
taxa
total
135
00
01
00
31136
00
00
00
51137
00
00
00
50138
00
01
00
44139
00
04
00
73140
00
02
01
281
141
00
00
00
19142
00
00
00
31143
00
06
06
392
144
00
01
10
164
145
00
06
03
354
146
00
825
40
1021
147
00
01
02
269
148
00
07
00
246
149
00
01
01
60TOTA
L0
08
555
133127
Note.
Units146–
148aremixed
units
that
combine
sedimentsfrom
Stratum
X1andStratum
X2.
Tab
leA2(Contin
ued)
CA1 Supplement A from Munro and Stiner, Zooarchaeological Evidence for Early Neolithic Colonization at Franchthi Cave (Peloponnese, Greece)
6

Table A3. Caprine fusion-based age stages and sample sizes
Agegroup Skeletal element
Fusionorder
Estimated age(months) N fused Total
A Radius, proximal 1 0–6 8 8B Humerus, distal 2 6–12 11 11B Pelvis 2 6–12 11 19B Scapula 2 6–12 14 14C Second phalanx, proximal 3 12–18 18 26C First phalanx, proximal 4 12–18 22 30D Tibia, distal 5 18–30 3 9D Metapodial, distal 6 18–30 8 19E Calcaneus 7 30–48 4 13E Femur, proximal 8 30–48 4 12E Femur, distal 8 30–48 4 11E Ulna, proximal 8 30–48 4 9E Radius, distal 9 30–48 3 14E Tibia, proximal 9 30–48 2 11F Humerus, proximal 10 481 1 5
Note. Adapted from Zeder’s (2006) scheme for sheep. The bone fusion data are used to produce survivorship curves based on sixage stages (A–F). Because our samples are moderate in size and only a fraction of the sheep and goat elements could be assigned tospecies, the survivorship curves are constructed for the combined sheep and goat assemblage. Estimated ages for goat bone fusion isthe same as for sheep (Zeder 2006).
Table A4. Raw and proportionally allocated (Payne 1973) counts by age stage for caprine dp4 and P4 dental elementsfrom the FAS and FAN trenches combined
MNE dP4/P4A
(0–2 months)B
(2–6 months)C
(6–12 months)D
(1–2 years)E
(2–4 years)F–I
(41 years) Total
Raw Count 2 1 8 5 0 2 18Proportionally allocated 0 1.2 11.6 1.2 0 0 14Final count 2 2.2 19.6 6.2 0 2 32
Note. Because the sheep and goat teeth usually could not be assigned to species, the mortality profile is constructed for the combined sheep and goat assemblage. To avoiddouble counting individuals, only the dp4-P4 sequence representing the caprine’s full life span is used, and dp4 elements were not counted if they appear to have been shed(significant root resorption; np 4) prior to the death of the individual. The majority of caprine mandibular teeth are isolated specimens that could not be aged via associationswith adjacent teeth. Isolated teeth were thus assigned initially to one or more mandibular wear stages following Payne (1973, fig. 11). Those specimens that could potentiallyspan two mandibular wear stages based on reference data (n p 17) were then reassigned to single wear stages following Payne’s (1973, table 2) correction method, inproportion to their occurrence in the raw (initial) counts. The only tooth with wear that could span more than two mandibular stages is from an animal older than 4 years, andthis specimen was combined with one other specimen to form a 41 years age category (stage F–I). MNE p minimum number of individuals.
Table A5. Sheep skeletal measurements for FAS and FAN assemblages combined at Franchthi Cave
Element Measure 1 2 3 4 5 6 7 8 9 10
Scapula SLC 16.5 . . . . . . . . . . . . . . . . . . . . . . . . . . .Scapula GLP 29.3 . . . . . . . . . . . . . . . . . . . . . . . . . . .Scapula LG 24.3 . . . . . . . . . . . . . . . . . . . . . . . . . . .Scapula BG 28.3 . . . . . . . . . . . . . . . . . . . . . . . . . . .Humerus Bd 28.7 30.7 . . . . . . . . . . . . . . . . . . . . . . . .Radius Bp 28.9 28.9 . . . . . . . . . . . . . . . . . . . . . . . .Radius Bd 30.7a 28.3a 27.5a 28.6a . . . . . . . . . . . . . . . . . .Radius BFp 27.4 26.5 . . . . . . . . . . . . . . . . . . . . . . . .Radius BFd 25.0a 25.5a 24.0a 25.0a . . . . . . . . . . . . . . . . . .Ulna BPC 17.1 . . . . . . . . . . . . . . . . . . . . . . . . . . .Ulna DPA 28.0 . . . . . . . . . . . . . . . . . . . . . . . . . . .Phalanx 1 Bp 11.6 11.2 11.5 12.5 11.6 11.9 12.4 . . . . . . . . .Phalanx 1 Bd 10.3 9.7 11.2 10.7 12.1 11.3 . . . . . . . . . . . .Phalanx 1 GL 36.2 24.2 36.4 37.4 38.2 39.4 36.2 . . . . . . . . .Femur Bd 41.5 37.2a . . . . . . . . . . . . . . . . . . . . . . . .Tibia Bd 27.1a . . . . . . . . . . . . . . . . . . . . . . . . . . .Tibia Dd 21.0a . . . . . . . . . . . . . . . . . . . . . . . . . . .Astragalus GLl 29.5 29.3 28.4 29.6 27.3 29.6 30.1 29.8 26.7 28.6
CA1 Supplement A from Munro and Stiner, Zooarchaeological Evidence for Early Neolithic Colonization at Franchthi Cave (Peloponnese, Greece)
7

Table A5 (Continued )
Element Measure 1 2 3 4 5 6 7 8 9 10
Astragalus GLm 27.7 27.1 27.5 26.6 27.9 26.3 29.0 28.2 27.5 26.9Astragalus Dl ∼16.9 17.1 16.8 15.3 16.5 15.9 16.4 17.2 15.8 16.1Calcaneus GL 61.7 . . . . . . . . . . . . . . . . . . . . . . . . . . .Calcaneus GB 20.6 . . . . . . . . . . . . . . . . . . . . . . . . . . .Metatarsus Bd 25.5 . . . . . . . . . . . . . . . . . . . . . . . . . . .
Note. Abbreviated measurement terms are from von den Driesch (1976). SLC: smallest length of the collum scapulae; GLP: greatest length of the processus articularis; LG:length of the glenoid cavity; BG: breadth of the glenoid cavity; Bd: greatest breadth of the distal end; Bp: greatest breadth of the proximal end; BFp: greatest breadth of thefacies articularis proximalis; BFd: greatest breadth of the facies articularis distalis; BPC: greatest breadth across the coronoid process; DPA: depth across the processusanconaeus; GL: greatest length; Dd: greatest depth of the distal end; GLl: greatest length of the lateral half; GLm: greatest length of the medial half; Dl: greatest depth of thelateral half; GB: greatest breadth. A tilde indicates an approximate measurement on slightly damaged element. Only selected measurements were used in logarithmic sizeindex (see text and table A6).a Measurement is for an unfused epiphysis.
Table A6. Mean logarithmic size index (LSI) values and standard deviations for sheep from Pre-Pottery Neolithicsites in the Levant, Anatolia, and eastern Aegean
Site Culture period Years cal BC N Lower range Mean Upper range Reference
Nevali Çori EPPNB 8500–8100 37 2.035 .035 .110 Peters et al. 1999Mureybet EPPNB/MPPNB (PPN IV) 8500–7250 24 2.024 .028 .061 Martin and Edwards 2013Shillourokambos EPPNB/MPPNB (Aa–Ac) 8300–7800 38 2.018 .014 .047 Vigne 2011Tell Halula MPPNB 8100–7250 23 .078 .015 2.050 Peters et al. 1999Guructepe LPPNB 7250–6700 89 .090 .020 2.060 Peters et al. 1999Ain Ghazal LPPNB 7250–6700 108 2.033 2.004 .034 Martin and Edwards 2013Ain Ghazal LPPNB/Early PPNC 7250–6250 142 2.042 2.004 .037 Martin and Edwards 2013Baja LPPNB 7250–6700 69 2.032 .006 .044 Martin and Edwards 2013Basta LPPNB 7250–6700 402 2.041 .013 .067 Martin and Edwards 2013Franchthi Initial Neolithic 7000–6500 32 2.057 2.034 2.010 This studyUlucak Höyük Early Neolithic (VIa-b) 7040–6470 44 2.065 2.034 2.003 Çakırlar 2012Ilıpınar Early Neolithic (X–VIII) 6400–5000 356 2.065 2.033 .000 Buitenhuis 1995
Note. LSI was calculated following Meadow (1999), using a standard for one individual sheep published by Uerpmann and Uerpmann (1994, table 12). Dates for southernLevantine sites are adapted from Martin and Edwards (2013). Upper and lower LSI ranges are based on 1 standard deviation. Summary data for Shillourokambos, FranchthiCave, Ulucak Höyük, and Ilıpınar are calculated directly from raw data; one measurement was included per element to avoid double counting. Breadth measurements werepreferentially used in all data sets. Measurements include scapula (LG), humerus (Bp, Bd), radius (Bp, Bd), ulna (Bpc), femur (DC, Bd), tibia (Bp, Bd), metapodials (Bop,Bd), astragalus (LI/GLI), calcaneus (GB), and phalanx 1 (Bp). EPPNB: Early Pre-Pottery Neolithic B; PPN: Pre-Pottery Neolithic; EPPNB/MPPNB: Early Pre-PotteryNeolithic B/Mid Pre-Pottery Neolithic B; MPPN: Mid Pre-Pottery Neolithic B; LPPNB: Late Pre-Pottery Neolithic B.
References Cited Only in Supplement APayne, S. 1973. Kill-off patterns in sheep and goats: the mandibles from Aşvan Kale. Anatolian Studies 23:281–303.Vigne, J.-D. 2011. Annexes archéozoologiques: les données brutes des analyses. In Shillourokambos. Un établissement néolithique pré-
céramique à Chypre. Les fouilles du Secteur 1. J. Guilaine, F. Briois, and J.-D. Vigne, eds. Pp. 1135–1157. Paris: Editions Errance/Ecole Française d’Athenes.
CA1 Supplement A from Munro and Stiner, Zooarchaeological Evidence for Early Neolithic Colonization at Franchthi Cave (Peloponnese, Greece)
8
Copyright © 2022 FDOKUMEN




















