Zonal and vertical distribution of radiolarians in the western and central Equatorial Pacific in...
40
Deep-Sea Research II 49 (2002) 2823–2862 Zonal and vertical distribution of radiolarians in the western and central Equatorial Pacific in January 1999 Hitoshi Yamashita, Kozo Takahashi*, Naoki Fujitani Department of Earth and Planetary Sciences, Graduate School of Sciences, Kyushu University, Hakozaki 6-1-10, Higashi Ku, Fukuoka 812-8581, Japan Received 26 July 2000; received in revised form 22 December 2000; accepted 25 July 2001 Abstract The zonal and vertical distribution of radiolarians was studied to document their relationship with their environment in the Equatorial Pacific in January 1999. Vertical plankton tows using a closing-type net with 63 mm mesh were made between the sea surface and 1000 m depth on board R/V Mirai at five stations located between 1401E and 1701W, i.e. in the western Pacific warm pool (WPWP) Region in the west and the Upwelling Region located in the east. The condition during the time of sampling was more or less La Ni * na, characterized by a zonal extension of the upwelling region at 1601E. Our survey yielded 261 radiolarian taxa including 114 Nassellaria, 120 Spumellaria and 27 Phaeodaria. Total radiolarian standing stocks increased from west to east, responding to the general increase in nutrients, chlorophyll-a, and diatom numbers. Three major radiolarian depth communities were identified: surface dwellers (0–120 m), subsurface dwellers (120–200 m), and intermediate-water dwellers (200–1000 m). When the vertical profiles were examined longitudinally, their abundance preference was characterized as follows: four taxa preferred the high- abundance water masses of the Upwelling Region: Lithomelissa setosa, Pseudocubus obeliscus, Stylodictya multispina, and Stylodictya validispina. On the other hand, Collosphaera tuberosa was most abundant in the WPWP. r 2002 Elsevier Science Ltd. All rights reserved. 1. Introduction Radiolarians are known to live at various depths in the epipelagic and mesopelagic zones (e.g., Renz, 1976; McMillen and Casey, 1978; Casey et al., 1979; Kling, 1979; Boltovskoy and Jankile- vich, 1985; Dworetzky and Morley, 1987; Kling and Boltovskoy, 1995; Welling et al., 1996; Abelmann and Gowing, 1997; Welling and Pisias, 1998) and are known to contribute to both the silica and organic carbon fluxes in the ocean. The equatorial region of the ocean displays a high radiolarian diversity (e.g., Takahashi, 1991) and primary production (Koblentz-Mishke et al., 1970; Berger et al., 1987; Barber and Chavez, 1991; Longhurst, 1998; Le Borgne et al., 2002) and their study can provide insights into the relationship between vertical radiolarian transport and the carbon cycle (Gowing, 1986, 1993). In addition, radiolarians comprise one of the dominant plank- ton and microfossil groups found in marine sediments. Therefore, a better knowledge of the ecology of living radiolarians can improve our understanding of the upper water circulation and *Corresponding author. Tel.: +81-92-642-2656; fax: +81- 92-642-2686. E-mail address: [email protected] (K. Takahashi). 0967-0645/02/$ - see front matter r 2002 Elsevier Science Ltd. All rights reserved. PII:S0967-0645(02)00060-7
Transcript of Zonal and vertical distribution of radiolarians in the western and central Equatorial Pacific in...

Deep-Sea Research II 49 (2002) 2823–2862
Zonal and vertical distribution of radiolarians in the westernand central Equatorial Pacific in January 1999
Hitoshi Yamashita, Kozo Takahashi*, Naoki Fujitani
Department of Earth and Planetary Sciences, Graduate School of Sciences, Kyushu University, Hakozaki 6-1-10, Higashi Ku,
Fukuoka 812-8581, Japan
Received 26 July 2000; received in revised form 22 December 2000; accepted 25 July 2001
Abstract
The zonal and vertical distribution of radiolarians was studied to document their relationship with their environment
in the Equatorial Pacific in January 1999. Vertical plankton tows using a closing-type net with 63 mm mesh were made
between the sea surface and 1000m depth on board R/V Mirai at five stations located between 1401E and 1701W, i.e. in
the western Pacific warm pool (WPWP) Region in the west and the Upwelling Region located in the east. The condition
during the time of sampling was more or less La Ni *na, characterized by a zonal extension of the upwelling region at
1601E. Our survey yielded 261 radiolarian taxa including 114 Nassellaria, 120 Spumellaria and 27 Phaeodaria. Total
radiolarian standing stocks increased from west to east, responding to the general increase in nutrients, chlorophyll-a,
and diatom numbers. Three major radiolarian depth communities were identified: surface dwellers (0–120m),
subsurface dwellers (120–200m), and intermediate-water dwellers (200–1000m). When the vertical profiles were
examined longitudinally, their abundance preference was characterized as follows: four taxa preferred the high-
abundance water masses of the Upwelling Region: Lithomelissa setosa, Pseudocubus obeliscus, Stylodictya multispina,
and Stylodictya validispina. On the other hand, Collosphaera tuberosa was most abundant in the WPWP. r 2002
Elsevier Science Ltd. All rights reserved.
1. Introduction
Radiolarians are known to live at various depthsin the epipelagic and mesopelagic zones (e.g.,Renz, 1976; McMillen and Casey, 1978; Caseyet al., 1979; Kling, 1979; Boltovskoy and Jankile-vich, 1985; Dworetzky and Morley, 1987; Klingand Boltovskoy, 1995; Welling et al., 1996;Abelmann and Gowing, 1997; Welling and Pisias,1998) and are known to contribute to both the
silica and organic carbon fluxes in the ocean. Theequatorial region of the ocean displays a highradiolarian diversity (e.g., Takahashi, 1991) andprimary production (Koblentz-Mishke et al., 1970;Berger et al., 1987; Barber and Chavez, 1991;Longhurst, 1998; Le Borgne et al., 2002) and theirstudy can provide insights into the relationshipbetween vertical radiolarian transport and thecarbon cycle (Gowing, 1986, 1993). In addition,radiolarians comprise one of the dominant plank-ton and microfossil groups found in marinesediments. Therefore, a better knowledge of theecology of living radiolarians can improve ourunderstanding of the upper water circulation and
*Corresponding author. Tel.: +81-92-642-2656; fax: +81-
92-642-2686.
E-mail address: [email protected] (K. Takahashi).
0967-0645/02/$ - see front matter r 2002 Elsevier Science Ltd. All rights reserved.
PII: S 0 9 6 7 - 0 6 4 5 ( 0 2 ) 0 0 0 6 0 - 7

past climate. Since radiolarian abundance is likelyinfluenced by various environmental factors (e.g.,hydrographic parameters, water mass movements,food availability), one of our objectives was toevaluate the relationship between the radiolarianspecific composition and the physico-chemicalproperties of the tropical region.Samples were collected as part of the Global
Carbon Cycle and Related Mapping based onSatellite Imagery Program (GCMAPS) in thewestern and central Equatorial Pacific. This regioncomprises the western Pacific warm pool (WPWP)and the equatorial upwelling region, east of it. TheWPWP registers sea-surface temperatures (SST)exceeding 281C (Yan et al., 1992) and both itstemperature and geographic extension fluctuatefrom year to year in relation to El Ni *no-SouthernOscillation (ENSO) events (e.g., Picaut et al., 1996;Inoue et al., 1996; Le Borgne et al., 2002). DuringEl Ni *no years, the WPWP expands and/or movestoward the eastern Pacific, and retracts to thewestern Pacific during La Ni *na years. Zonalextension of the equatorial divergence follows thereverse pattern. The velocity of the EquatorialUndercurrent (EUC), which has an eastwardcomponent, decreases during an El Ni *no event(Kessler and McPhaden, 1995).Previous studies have shown that radiolarian
assemblages are correlated to the eastward exten-sion of WPWP from the western Pacific (Wellinget al., 1996). Similarly, the conspicuous changes inorganic carbon flux due to the El Ni *no of 1983were reflected by changes in radiolarian speciesabundance in the region slightly north of theEquatorial Pacific (111N, 1401W) (Pisias et al.,1986). As a baseline study, the present work willshed light on the fundamental understanding ofENSO events.
2. Samples and methods
Plankton samples for this study were collectedat several depth-intervals from 0 to 1000m at fivestations (from west to east: Stations 2, 3, 6, 9, and12) in the western Equatorial Pacific duringMR98-K02 Cruise (2–15 January 1999) on boardR/V Mirai, operated by the Japan Marine Science
and Technology Center (JAMSTEC) (Table 1,Fig. 1). Station 2 was located under the influenceof the North Equatorial Counter Current(NECC), and Stations 3, 6, 9, and 12 were locatedunder the influence of the South Equatorial
Table 1
Logistic and sample information for the vertical plankton tows
conducted at five stations during R/V Mirai Cruise MR98-K02
Station No. Sample
No.
Depth
interval
(m)
Total
specimens
counted
St. 2 1 0–10 401
2 10–30 403
Latitude 05102.490N 3 30–50 403
Longitude 140107.900E 4 50–75 399
Date 2 Jan. 1999 5 75–100 399
Sampling time 0:36–3:17 6 100–125 401
Bottom depth 4175m 7 125–150 407
8 150–200 406
9 200–500 400
10 500–1000 402
St. 3 11 0–40 405
12 40–80 406
Latitude 0100.370N 13 80–120 405
Longitude 144159.940E 14 120–160 403
Date 4 Jan. 1999 15 160–200 405
Sampling time 0:23–3:21 16 200–500 404
Bottom depth 3638m 17 500–1000 418
St. 6 18 0–40 406
19 40–80 399
Latitude 0101.380N 20 80–120 401
Longitude 159159.880E 21 120–160 405
Date 7–8 Jan. 1999 22 160–200 396
Sampling time 23:58–2:45 23 200–500 421
Bottom depth 2800m 24 500–1000 431
St. 9 25 0–40 405
26 40–80 405
Latitude 0101.960N 27 80–120 405
Longitude 174158.740E 28 120–160 405
Date 12 Jan. 1999 29 160–200 405
Sampling time 0:32–2:52 30 200–500 405
Bottom depth 4831m 31 500–750 405
St. 12 32 0–40 406
Latitude 0100.550N 33 40–80 404
Longitude 170107.960W 34 80–120 405
Date 14–15 Jan. 1999 35 120–160 405
Sampling time 0:07–2:09 36 160–200 404
Bottom depth 5397m 37 200–500 406
H. Yamashita et al. / Deep-Sea Research II 49 (2002) 2823–28622824

Current (SEC). Samples from below 750m atStation 9 and below 500m at Station 12 were notcollected due to strong currents. The samples wereobtained using a flow-metered closing planktonnet with 63-mm mesh size, between 1:00 and 3:00a.m. at each station, and preserved in 4% bufferedformaldehyde. The amount of water filtered in thenet was computed by the UNESCO formula(Cassie, 1968).Using a Folsom plankton splitter, an optimal
concentration was attained by repeated splitting(e.g., mainly 1/8–1/32 with a range of 1/8–1/128aliquot size) in order to perform the most efficientand accurate radiolarian counts. The split samplewas heated to 801C after adding 5ml of 1 N-hydrochloric acid and 30ml of 10%-hydrogenperoxide in order to digest calcium carbonate andorganic matter. The digested sample was filteredthrough 47mm Gelmans grid membrane filterswith 0.45 mm sized pores. At the end of thefiltration, the filter was rinsed with distilled waterand dried overnight in an oven at 401C. Thefiltered sample was mounted on a standard glassslide with Canada Balsam.In the eastern Equatorial Atlantic the shells
smaller than 40–60 mm (mostly juvenile) represent50% of total polycystine shells (Boltovskoy et al.,
1993). Thus, theo63-mm radiolarian shells cannotbe disregarded in the total abundance. Because oursamples were obtained with a 63-mm mesh net, ourresults in the western Equatorial Pacific are likelyto represent partial abundances.For diversity indices, the Shannon–Wiener log-
base 2 formula was used (Shannon and Weaver,1949). A cluster analysis, using the Pearson’scorrelation coefficient average linkage methodwith Systat v. 5.2, was applied to the radiolarianstanding stocks to test the relationships betweenthe taxa. One hundred and one radiolarian taxa,which were determined to have sufficient countnumbers, were applied to the cluster analysis. Therelatively rare radiolarian taxa were eliminated inthe cluster analysis computations.Species identification of radiolarians was per-
formed using the following works: Bj�rklund(1976), Renz (1976), Nigrini and Moore (1979),Takahashi and Honjo (1981), and Takahashi(1991). Taxonomic identification down to specieslevel was made on complete specimens (in the caseof broken specimens, >50% of the skeleton) formost of the abundant polycystine (Nassellaria andSpumellaria) and phaeodarian radiolarians. Whenradiolarians could not be identified to species level,they were recorded in the following categories:
140°E 160° 180° 160°W15°S 15°S
10° 10°
5° 5°
0° 0°
5° 5°
10° 10°
15°N 15°N
0 400
kmLongitude
Latit
ude
Station 2
Station 3 Station 6 Station 9 Station 12
WPWP Region Upwelling RegionSalinity Front
NEC
NECC
SEC
SEC
Fig. 1. Locations of the vertical plankton tow stations and oceanographic features during Cruise MR98-K02 in the western and
central Equatorial Pacific. Major surface currents (NEC: North Equatorial Current, NECC: North Equatorial Countercurrent, SEC:
South Equatorial Current) and characterized water masses (WPWP Region, salinity front, Upwelling Region).
H. Yamashita et al. / Deep-Sea Research II 49 (2002) 2823–2862 2825
‘‘other nassellarians’’, ‘‘other spumellarians’’, and‘‘other phaeodarians’’. If these specimens were notidentified to the suborder level, they were recordedas ‘‘other radiolarians’’.
3. Results
3.1. Oceanographic setting
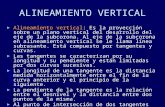
Since this study was part of a multidisciplinaryprogram conducted on board R/V Mirai, pertinenthydrographic and other physico-chemical databecame available (Suzuki et al., 1999). Thecondition during the time of sampling was LaNi *na, marked by a zonal extension of theequatorial upwelling west of the dateline (1801).La Ni *na followed the termination of the1997–1998 significant El Ni *no event. From westto east the following systems were observed: the
WPWP with low SSS (34.0=34.7), and higherthan normal SST (>29.41C). At approximately1561E, the SSS increases abruptly from 34.4 to35.0 PSU, indicating the western limit of theequatorial upwelling. The WPWP extends down toapproximately 100m where a strong thermoclinedevelops, isolating the subsurface water below thisdepth and making it oligotrophic. In this regionrainfall strongly influences the water mass condi-tion, including a ‘‘barrier layer’’ (Lukas andLindstrom, 1991). Conversely, occurrence of sur-face nutrients in the Upwelling Region make iteutrophic. Such characteristic distribution basedon the nutrient concentrations in this region can beclearly illustrated by 2-D (W–E and vertical)distributions of nutrients such as nitrate andsilicate (Figs. 2 and 3). Although the western frontof the equatorial upwelling is not well recognizedfrom SST, the edge of this region is clearly markedby a surface-salinity front (Rodier et al., 2000).
(b)
36
Temperature (°C)
Dep
th (
m)
0 10 20 300
500
100034 35 36
St. 2
34 35
St. 3
0 10 20 30
Salinity (PSU)
St. 6
34 35 36
0 10 20 30
St. 9
34 35 36
0 10 20 30
St. 12
TemperatureSalinity
34 35 36
0 10 20 30
(a)
Longitude140°E 150° 160° 170° 180° 170°W
24
Tem
pera
ture
(°C
)
25
26
27
28
29
30
31
34
Sal
inity
(P
SU
)34.5
35
35.5
Sea Surface Temperature Sea Surface Salinity
St. 2 St. 3 St. 6 St. 9 St. 12
Fig. 2. (a) Hydrographic data (sea surface temperature and salinity) obtained in the Equatorial area, taken every ten minutes during
Cruise MR98-K02. (b) Vertical profiles of temperature and salinity from 0 to 1000m at each station during Cruise MR98-K02 in the
Equatorial Pacific.
H. Yamashita et al. / Deep-Sea Research II 49 (2002) 2823–28622826
The displacement of this front is closely related tothe Southern Oscillation Index (SOI: Inoue et al.,1996) and, for example, was located at 167–1691Eduring non-El Ni *no and at 1721W during El Ni *no(Rodier et al., 2000).
3.2. Total radiolarian standing stock
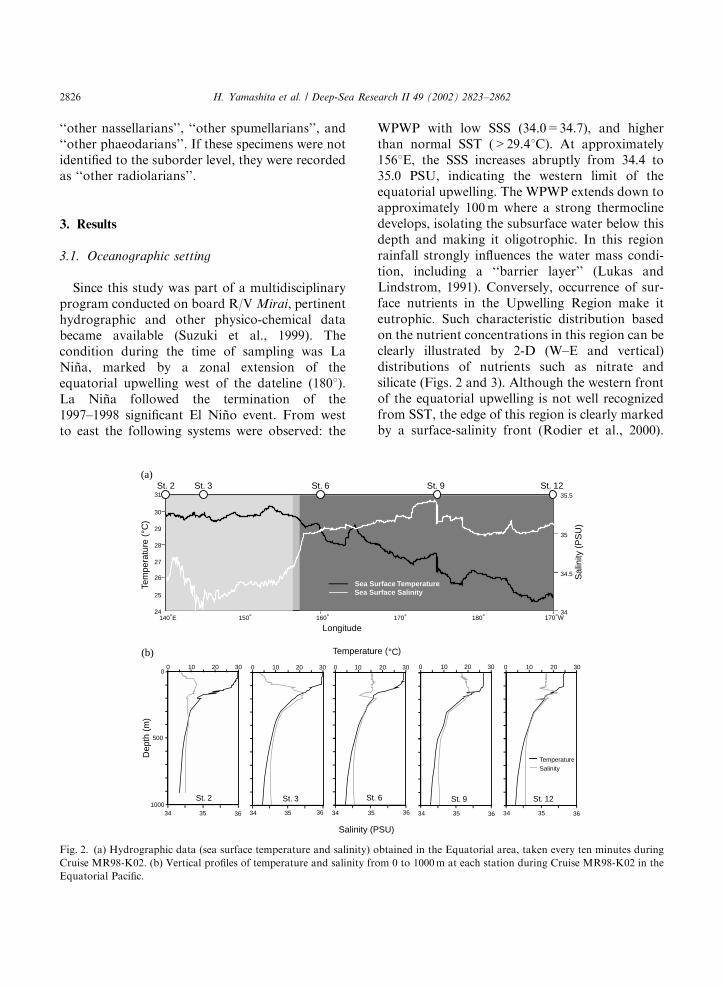
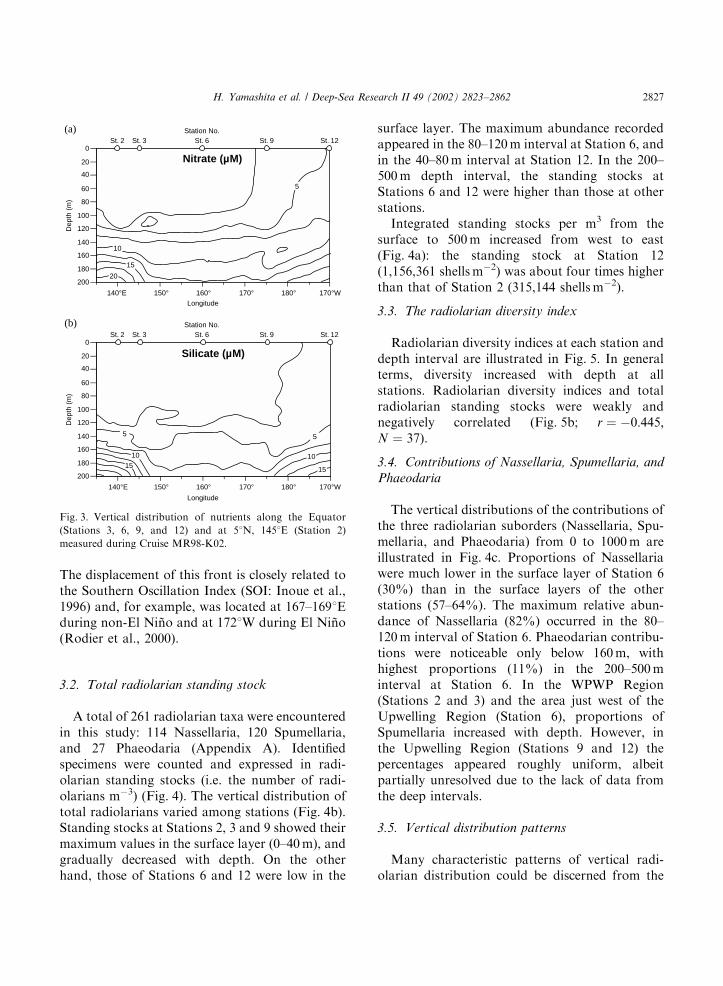
A total of 261 radiolarian taxa were encounteredin this study: 114 Nassellaria, 120 Spumellaria,and 27 Phaeodaria (Appendix A). Identifiedspecimens were counted and expressed in radi-olarian standing stocks (i.e. the number of radi-olarians m�3) (Fig. 4). The vertical distribution oftotal radiolarians varied among stations (Fig. 4b).Standing stocks at Stations 2, 3 and 9 showed theirmaximum values in the surface layer (0–40m), andgradually decreased with depth. On the otherhand, those of Stations 6 and 12 were low in the
surface layer. The maximum abundance recordedappeared in the 80–120m interval at Station 6, andin the 40–80m interval at Station 12. In the 200–500m depth interval, the standing stocks atStations 6 and 12 were higher than those at otherstations.Integrated standing stocks per m3 from the
surface to 500m increased from west to east(Fig. 4a): the standing stock at Station 12(1,156,361 shellsm�2) was about four times higherthan that of Station 2 (315,144 shellsm�2).
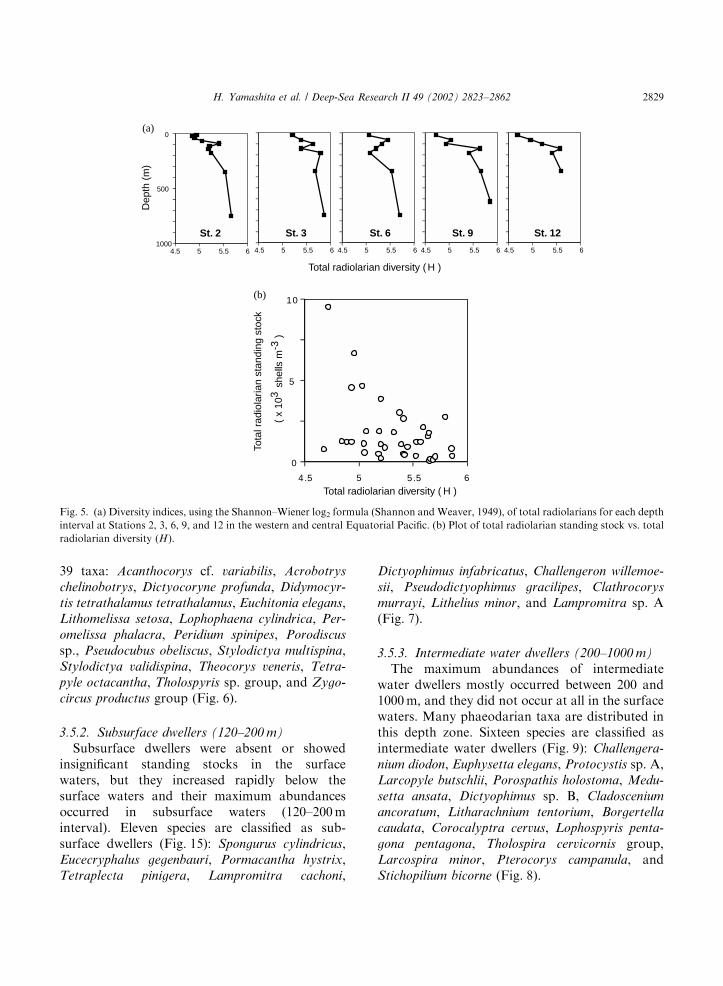
3.3. The radiolarian diversity index
Radiolarian diversity indices at each station anddepth interval are illustrated in Fig. 5. In generalterms, diversity increased with depth at allstations. Radiolarian diversity indices and totalradiolarian standing stocks were weakly andnegatively correlated (Fig. 5b; r ¼ �0:445;N ¼ 37).
3.4. Contributions of Nassellaria, Spumellaria, and
Phaeodaria
The vertical distributions of the contributions ofthe three radiolarian suborders (Nassellaria, Spu-mellaria, and Phaeodaria) from 0 to 1000m areillustrated in Fig. 4c. Proportions of Nassellariawere much lower in the surface layer of Station 6(30%) than in the surface layers of the otherstations (57–64%). The maximum relative abun-dance of Nassellaria (82%) occurred in the 80–120m interval of Station 6. Phaeodarian contribu-tions were noticeable only below 160m, withhighest proportions (11%) in the 200–500minterval at Station 6. In the WPWP Region(Stations 2 and 3) and the area just west of theUpwelling Region (Station 6), proportions ofSpumellaria increased with depth. However, inthe Upwelling Region (Stations 9 and 12) thepercentages appeared roughly uniform, albeitpartially unresolved due to the lack of data fromthe deep intervals.
3.5. Vertical distribution patterns
Many characteristic patterns of vertical radi-olarian distribution could be discerned from the
0
20
40
60
80
100
120
140
160
180
200
140°E 150° 160° 170° 180° 170°W
St. 2 St. 3 St. 6 St. 9 St. 12
Dep
th (
m)
Longitude
Station No.
5
20
10
15
Nitrate (µM)
(a)
0
20
40
60
80
100
120
140
160
180
200
140°E 150° 160° 170° 180° 170°W
St. 2 St. 3 St. 6 St. 9 St. 12
Dep
th (
m)
Longitude
Station No.
5 5
1015
10
15
(b)
Silicate (µM)
Fig. 3. Vertical distribution of nutrients along the Equator
(Stations 3, 6, 9, and 12) and at 51N, 1451E (Station 2)
measured during Cruise MR98-K02.
H. Yamashita et al. / Deep-Sea Research II 49 (2002) 2823–2862 2827
depth plots of species abundances. Therefore, acluster analysis was used in order to summarize therelationships between the 101 radiolarian taxa(Fig. 9), yielding three distinctive patterns: surfacedwellers (0–120m), subsurface dwellers (120–200m), and intermediate water dwellers (200–1000m). A specific maximum abundance depthrange characterized each pattern. Outside of thisrange, the abundance level for each species wasmuch lower. Most peak abundance intervals weregenerally narrow and sharply bounded by lowerabundances. Furthermore, these vertical distribu-tion patterns were slightly different at each of the
stations. For example, the maximum abundancesof Didymocyrtis tetrathalamus tetrathalamus (sur-face dweller) occurred in the 40–80m interval atStation 9, while at Stations 3 and 6 this taxonappeared in the upper most surface layer (0–40m).
3.5.1. Surface dwellers (0–120 m)
Surface dwellers were characterized by pro-nounced maximum abundances in the surfacewater (0–120m interval). In this study, 39 speciesof relatively high standing stocks were classified assurface dwellers (Fig. 9). The following speciesrepresented the most numerous 17 taxa among the
Total radiolarian standing stock ( x103 shells m-3 )
Dep
th (
m)
5 100
St. 3
5 100
St. 6
5 100
St. 9
NO DATA
5 100
St. 12
NO DATA
0
1000
500
5 100
St. 2
(b)
Dep
th (
m)
Percent 0 50 100
St. 2 St. 3
0 50 100
St. 6
0 50 100
St. 9
0 50 100
Nassellaria
Spumellaria
Phaeodaria
Others
St. 12
0 50 100
(c)
Tota
l rad
iola
rian
stan
ding
stoc
k (x10
6 she
lls m
-2)
0
0.5
1.0
1.5
2 3 6 9 12Station No.
(a)
0
500
1000
Fig. 4. (a) Radiolarians (# shellsm�2: 0–500m) at Stations 2, 3, 6, 9, and 12. (b) Depth profiles of total radiolarians (# shellsm�3: 0–
1000m, where available) at Stations 2, 3, 6, 9, and 12. (c) Vertical distribution of relative abundances of suborders (Nassellaria,
Spumellaria, and Phaeodaria: 0–1000m, where available) at Stations 2, 3, 6, 9, and 12.
H. Yamashita et al. / Deep-Sea Research II 49 (2002) 2823–28622828
39 taxa: Acanthocorys cf. variabilis, Acrobotrys
chelinobotrys, Dictyocoryne profunda, Didymocyr-
tis tetrathalamus tetrathalamus, Euchitonia elegans,Lithomelissa setosa, Lophophaena cylindrica, Per-
omelissa phalacra, Peridium spinipes, Porodiscus
sp., Pseudocubus obeliscus, Stylodictya multispina,Stylodictya validispina, Theocorys veneris, Tetra-
pyle octacantha, Tholospyris sp. group, and Zygo-
circus productus group (Fig. 6).
3.5.2. Subsurface dwellers (120–200 m)
Subsurface dwellers were absent or showedinsignificant standing stocks in the surfacewaters, but they increased rapidly below thesurface waters and their maximum abundancesoccurred in subsurface waters (120–200minterval). Eleven species are classified as sub-surface dwellers (Fig. 15): Spongurus cylindricus,Eucecryphalus gegenbauri, Pormacantha hystrix,Tetraplecta pinigera, Lampromitra cachoni,
Dictyophimus infabricatus, Challengeron willemoe-
sii, Pseudodictyophimus gracilipes, Clathrocorys
murrayi, Lithelius minor, and Lampromitra sp. A(Fig. 7).
3.5.3. Intermediate water dwellers (200–1000 m)
The maximum abundances of intermediatewater dwellers mostly occurred between 200 and1000m, and they did not occur at all in the surfacewaters. Many phaeodarian taxa are distributed inthis depth zone. Sixteen species are classified asintermediate water dwellers (Fig. 9): Challengera-
nium diodon, Euphysetta elegans, Protocystis sp. A,Larcopyle butschlii, Porospathis holostoma, Medu-
setta ansata, Dictyophimus sp. B, Cladoscenium
ancoratum, Litharachnium tentorium, Borgertella
caudata, Corocalyptra cervus, Lophospyris penta-
gona pentagona, Tholospira cervicornis group,Larcospira minor, Pterocorys campanula, andStichopilium bicorne (Fig. 8).
0
500
1000
Total radiolarian diversity ( H )
Dep
th (
m)
4.5 5 5.5 6
St. 2 St. 3
4.5 5 5.5 6
St. 6
4.5 5 5.5 6
St. 9
4.5 5 5.5 6
St. 12
4.5 5 5.5 6
(a)
(b)
0
5
10
4.5 5 5.5 6
Tota
l rad
iola
rian
stan
ding
sto
ck
Total radiolarian diversity ( H )
( x
103
)
she
lls m
-3
Fig. 5. (a) Diversity indices, using the Shannon–Wiener log2 formula (Shannon and Weaver, 1949), of total radiolarians for each depth
interval at Stations 2, 3, 6, 9, and 12 in the western and central Equatorial Pacific. (b) Plot of total radiolarian standing stock vs. total
radiolarian diversity (H).
H. Yamashita et al. / Deep-Sea Research II 49 (2002) 2823–2862 2829

Tetrapyle octacantha
0 250 5000 250 500 0 250 5000 250 500
0
500
10000 250 500
NO DATA
NO DATA
West EastSTATION No.
Pseudocubus obeliscus
2500 500 0 250 5000 250 5000 250 5002500 500
0
500
1000
NO DATA
NO DATA
Lithomelissa setosa
0 1000 2000 0 1000 2000 0 1000 2000
NO DATA
0 1000 2000
NO DATA
0
500
10000 1000 2000
Didymocyrtis tetrathalamus tetrathalamus
0 5002500 500250 0 500250 0 5002500 500250
0
500
1000
NO DATA
NO DATA
0 200 400
0
500
10000 200 400 0 200 400 0 200 400 0 200 400
NO DATA
NO DATA
Peridium spinipes
2 3 6 9 12
SURFACE DWELLERS
Dep
th (
m)
Dep
th (
m)
Dep
th (
m)
Dep
th (
m)
Dep
th (
m)
Standing stock (no. shells m-3)
0 200 400
0
500
10000 200 400 0 200 400 0 200 400 0 200 400
NO DATA
NO DATA
Lophophaena cylindrica
Dep
th (
m)
Fig. 6. Vertical profiles of most abundant surface dwelling radiolarians (# shellsm�3).
H. Yamashita et al. / Deep-Sea Research II 49 (2002) 2823–28622830

Stylodictya multispina
0 100 200 3000 100 200 300 0 100 200 300 0 100 200 3000 100 200 300
0
500
1000
NO DATA
NO DATA
Acanthocorys cf. variabilis
0 100 200 3000 100 200 300
0
500
10000 100 200 300 0 100 200 300 0 100 200 300
NO DATA
NO DATA
Peromelissa phalacra
0 100 200 3000 100 200 300 0 100 200 300
NO DATA
0 100 200 300
0
500
10000 100 200 300
NO DATANO DATA
NO DATA
Dep
th (
m)
Dep
th (
m)
Dep
th (
m)
Standing stock (no. shells m-3)
Porodiscus sp.
0 100 200 0 100 200 0 100 200 0 100 2000 100 200
0
500
1000
NO DATA
NO DATA
Dictyocoryne profunda
0 50 100 150 0 50 100 150 0 50 100 1500 50 100 150
0
500
1000500 100 150
NO DATA
NO DATA
0 100 200 300
0
500
10003000 100 200 0 100 200 300
NO DATA
0 100 200 300
NO DATA
0 100 200 300
Zygocircus productus
Dep
th (
m)
Dep
th (
m)
Dep
th (
m)
West EastSTATION No.
2 3 6 9 12
SURFACE DWELLERS
Fig. 6 (continued).
H. Yamashita et al. / Deep-Sea Research II 49 (2002) 2823–2862 2831

3.5.4. Geographic changes in absolute abundance
The vertical distribution of the major speciesstanding stocks at each station is shown in Figs. 6–8. Radiolarian vertical distribution patterns varied
at each station. The following are representativeexamples.Although the surface dweller Lithomelissa setosa
was abundant at Stations 9 and 12, it showed
Euchitonia elegans
0 50 100 150 0 50 100 150 0 50 100 150 0 50 100 1500 50 100 150
0
500
1000
NO DATA
NO DATA
Stylodictya validispina
0 50 100 150 0 50 100 150 0 50 100 150 0 50 100 1500 50 100 150
0
500
1000
NO DATA
NO DATA
0 50 100 150
0
500
10000 50 100 150 0 50 100 150 0 50 100 150 0 50 100 150
NO DATA
NO DATA
Tholospyris sp. group
0 20 40 60 80
0
500
10000 20 40 60 80 0 20 40 60 80 0 20 40 60 80 0 20 40 60 80
NO DATA
NO DATA
Acrobotrys chelinobotrys
0 25 50
0
500
10000 25 50 0 25 50 0 25 50 0 25 50
NO DATA
NO DATA
Theocorys veneris
Dep
th (
m)
Dep
th (
m)
Dep
th (
m)
Dep
th (
m)
Dep
th (
m)
Standing stock (no. shells m-3)
West EastSTATION No.
2 3 6 9 12
SURFACE DWELLERS
Fig. 6 (continued).
H. Yamashita et al. / Deep-Sea Research II 49 (2002) 2823–28622832

much lower abundances at Stations 2, 3 and 6. Theboundary between these different abundancesappeared between Stations 6 and 9, and thedifference of the standing stock was considerablebetween the west and the east of this boundary.Analogous to L. setosa, the following species
showed the same pattern (Fig. 6): Pseudocubus
obeliscus, Stylodictya multispina, and Stylodictya
validispina.Collosphaera tuberosa, a colonial spumellarian,
showed an opposite pattern to L. setosa. AtStations 3 and 6 the abundances of C. tuberosa
were higher than those at Stations 9 and 12. Nospecimen of this taxon was recorded at Station 2.The abundances of C. tuberosa at Stations 9 and12 were mostly near zero. The abundance changeof C. tuberosa also occurred between Stations 6and 9 as it did in the case of L. setosa.Unlike the above species, the abundances of
Peridium spinipes, also a surface dweller, did notshow major differences among stations. Never-theless, the vertical distribution pattern variedslightly from one station to the other. Most of thespecies with such a pattern belonged to the
Phormacantha hystrix
0 50 100150 200
0
500
1000
NO DATA
NO DATA
0 50 100 150 200 0 50 100 150 200 0 50 100150 200 0 50 100150 200
Eucecryphalus gegenbauri
0 20 40 60
0
500
10000 20 40 60 0 20 40 60 0 20 40 60 0 20 40 60
NO DATA
NO DATA
0 10 20 30 40
Lithelius minor
400 10 20 30
0
500
10000 10 20 30 40 400 10 20 30 400 10 20 30
NO DATA
NO DATA
Pseudodictyophimus gracilipes
0 50 100 150
0
500
10000 50 100 150 0 50 100 150 0 50 100 150 0 50 100 150
NO DATA
NO DATA
Spongurus cylindricus
0 10 20 30
0
500
10000 10 20 30 0 10 20 30 0 10 20 30 0 10 20 30
NO DATA
NO DATA
0 50 100 150
0
500
100050 100 1500 0 50 100 150 0 50 100 150 0 50 100 150
Clathrocorys murrayi
NO DATA
NO DATA
West East
SUBSURFACE DWELLERS
STATION No.2 3 6 9 12
Dep
th (
m)
Standing stock (no. shells m-3)
Dep
th (
m)
Dep
th (
m)
Dep
th (
m)
Dep
th (
m)
Dep
th (
m)
Fig. 7. Vertical profiles of most abundant subsurface dwelling
radiolarians (# shellsm�3).
Corocalyptra cervus
0 25 50
0
500
10000 25 50 0 25 50 0 25 50 0 25 50
NO DATA
NO DATA
Litharachnium tentorium
0 20 40 60 800 20 40 60 80
0
500
10000 20 40 60 80 0 20 40 60 80 0 20 40 60 80
NO DATA
NO DATA
Borgertella caudata
0
0
500
100010050 0 50 100 0 50 100 0 50 100 0 50 100
NO DATA
NO DATA
Cladoscenium ancoratum
0 25 50 75 100
0
500
10000 25 50 75 100 0 25 50 75 100 0 25 50 75 100 0 25 50 75 100
NO DATA
NO DATA
West East
INTERMEDIATE DWELLERS
STATION No.2 3 6 9 12
Dep
th (
m)
Standing stock (no. shells m-3)
Dep
th (
m)
Dep
th (
m)
Dep
th (
m)
Fig. 8. Vertical profiles of most abundant intermediate water
dwelling radiolarians (# shellsm�3).
H. Yamashita et al. / Deep-Sea Research II 49 (2002) 2823–2862 2833

Distances0 1.0
Collosphaera tuberosaCentrobotrys thermophila
Disolenia collinaHexacontium sp.Acanthosphaera tunisEucecryphalus sestrodiscusChallengeranium diodonEuphysetta elegansProtocystis sp. ALarcopyle butschliiPorospathis holostomaMedusetta ansataDictyophimus sp. BCladoscenium ancoratumLitharachnium tentoriumBorgertella caudataCorocalyptra cervusLophospyris pentagona pentagona
Pterocorys campanulaLarcospira minorTholospira cervicornis group
Stichopilium bicorne
Eucecryphalus gegenbauriSpongurus cylindricus
Phormacantha hystrixTetraplecta pinigeraLampromitra cachoniDictyophimus infabricatusChallengeron willemoesiiPseudodictyophimus gracilipesClathrocorys murrayiLithelius minor
Conchellium capsulaLampromitra sp. A
Carpocanistrum flosculumLophospyris pentagona hyperboreaPterocanium trilobumLophophaena cf.capitoTholospira groupSpongotrochus glaciarisCarpocanistrum sp.Amphirhopalum ypsilonSpirocyrtis sp.Arachnosphaera myriacanthaAnthocrytidium ophirensePterocanium praetextum praetextum
Hexacontium axotriasHexacontium hostile
Acanthodesmia vinculataSpongodiscus bicoricarus
Eucyrtidium hexastichumStylodictya sp.Larcospira quadrangulaHexapyle sp.Porodiscus micrommaEuchitonia elegansPorodiscus sp.Stylodictya validispira
Lophophaena cylindricaLophophaena sp.
Tetrapyle octacanthaPseudocubus obeliscusLithomelissa setosaStylodictya multispinaPlegmosphaera entodictyonLophophaena decacantha groupAcrobotrys chelinobotrysPeromelissa phalacraAcanthocorys cf. variabilisPeridium spinipesActinomma arcadophorumTheocorys venerisEucyrtidium hexagonatumZygocircus productusDidymocyrtis t. tetrathulamusSemantis gracilisSpongodiscus spp. B groupSpongodiscus sp. AStaurodictyo cruciataSpongaster tetras tetrasTholospyris sp. GroupBotryocyrtis scutumZygocircus sp. piscicaudatusDictyocoryne profundaEuchitonia sp.Centrocubus octostylusLithelius solarisOctopyle groupGiraffospyris angulataLophospyris pentagona quadriforisSpongaster pentasEuchitonia cf. furcataLophospyris juvenile form groupPterocorys zancleus Hexalonche amphisiphonLitharachnium eupiliumNeosemantis distephanusAcrosphaera spinosaSpirocyrtis scalarisDictyocoryne juvenile groupBotryocyrtis elongatum
Su
rfac
e d
wel
lers
Su
bsu
rfac
e d
wel
lers
Inte
rmed
iate
w
ater
dw
elle
rsSpecies
Fig. 9. Dendrogram of average linkage cluster using correlation coefficients. Three depth zonation patterns (surface, subsurface, and
intermediate water dwellers, hatched) are distinguished.
H. Yamashita et al. / Deep-Sea Research II 49 (2002) 2823–28622834

Nassellaria and these species’ standing stocks werecomparatively high. This pattern is also seen in thefollowing species (Fig. 6): Lophophaena cylindrica,Peromelissa phalacra, and Zygocircus productus
group.
Borgertella caudata is a phaeodarian radiolarianthat dwells in the intermediate waters in the studyregion. The abundances of B. caudata were higherin the 200–500m interval at Station 6 than in theother depth-intervals and at the other stations. Thetendency of the abundance peak to deepentowards the east (Stations 3–9) is seen in B.
caudata. The following species also showed thispattern (Fig. 8): Corocalyptra cervus, Cladosce-
nium ancoratum, and Litharachnium tentorium.The maximum abundance of Didymocyrtis
tetrathalamus tetrathalamus occurred mostlyabove 200m, characterizing this taxon as a surfacedweller. Nevertheless, the standing stock of D. t.
tetrathalamus in the surface waters at Station 6was much lower than that at other stations. Thefollowing species also showed this pattern (Fig. 6):Acanthocorys cf. variabilis, Acrobotrys chelinobo-
trys, Dictyocoryne profunda, Theocorys veneris,Tholospyris sp. group, and Tetrapyle octacantha.
In addition, there are species that only occurredat Stations 2 and 3. We exclude discussions onthese radiolarian species with significantly smallstanding stocks, but they are included in AppendixB.
4. Discussion
The vertical and longitudinal distribution pat-terns defined in the studied transect showed thatthe distribution of radiolarian species is closelyrelated to water depth, distribution of watermasses, hydrographic parameters and nutrientconcentrations. This documents the factors con-trolling the distribution of individual radiolarianspecies by specific hydrographic conditions. Theintegrated standing stocks of total radiolariansperm3 from the surface to 500m increased fromwest to east (Stations 2–12). This result conformsto the observation that the zooplankton biomasseswere a few times higher in the eutrophic (upwel-ling) region than in the oligotrophic (WPWP)
region (Rodier et al., 2000; Le Borgne et al., 2002).Furthermore, Takahashi (1983) suggested thatthere is a tendency for larger radiolarian standingstock values in eutrophic/upwelling areas than inoligotrophic areas. The upper 100m of surfacewaters of the WPWP Region is oligotrophic, andoverlays eutrophic subsurface waters, while thesurface waters of the Upwelling Region areeutrophic, supporting the suggested tendency. Inthe same study area the horizontal distribution ofsurface diatoms was investigated using the samplesfrom the same cruise (Kobayashi and Takahashi,2002). The total numbers of surface water diatomsas well as centric diatom standing stocks tend toincrease from west to east, with a characteristiclow diatom standing stock near Station 6. Thus,the distribution trend of the radiolarian standingstocks is similar to that of surface diatoms. Inaddition, the vertical distribution of chlorophyll-aduring the cruise (Suzuki et al., 1999) changedsignificantly near 1601E, from low in the WPWPRegion to high in the Upwelling Region. There-fore, the radiolarian productivity in the westernEquatorial Pacific appears to have a close relation-ship with primary production (e.g., diatoms,cocolithophorids) as shown in other parts of theworld ocean. Earlier, Abelmann and Gowing(1997) suggested that increased abundances ofradiolarians in the water column indicated highernutrient conditions in the Atlantic sector of theSouthern Ocean.Radiolarian diversity indices and standing
stocks showed a weak negative correlation(Fig. 5). Diversity indices (mean H: 5.324) in thisstudy were comparatively high when comparedwith those in other studies (Kling and Boltovskoy,1995: mean H: 4.780). Note that the abovecomparison was not straightforward since Klingand Boltovskoy’s (1995) samples were obtained inthe California Current at 321N and their values didnot include phaeodarians. In the eastern subarcticPacific, the generally high indices correspond tolow fluxes of total radiolarians (Takahashi, 1997).Thus, similar relationships between the standingstock and the diversity indices are apparent for thisstudy and that of Takahashi (1997). In this regionour results support the idea that a highlyproductive region has a tendency to be less diverse
H. Yamashita et al. / Deep-Sea Research II 49 (2002) 2823–2862 2835

than an oligotrophic region (Boltovskoy andJankilevich, 1985). It appears that the radiolariandiversity indices of the near surface tend togradually increase towards the west. This tendencyis especially induced by the surface dwellers, whichhave dominant standing stocks in the eutrophicregion (Upwelling Region). The diversity indices inthe Shannon–Wiener’s expression (Shannon andWeaver, 1949) depends on two attributes: numberof species and equitability. It can be inspected bythe equitability equation as follows: H=Hmax=(P1
log2P1 þ P2 log2 P2 þyþ PSlog2 PS/log2 S)(Pielou, 1966). Our computations using the aboveequitability assessment led to the following con-clusion. It turns out that, in our samples, theequitability has a greater influence than the speciesrichness in the numerical expression of thediversity index.Radiolarian vertical distributions at Stations 6
and 12 show complex profiles from the surface to1000m. The unique vertical distributional struc-tures appear to be caused by the presence of thewater mass front and the condition of the mixedlayer. Station 6 is located within the boundarybetween the WPWP Region and the UpwellingRegion. At Station 12, the mixed layer is thickerthan at the western stations due to a morevigorous upwelling. At both Stations 6 and 12the standing stocks in the 200–500m interval werecomparatively high (Fig. 4b). Phaeodaria wereparticularly abundant (11%) in this interval atStation 6. In the Equatorial Atlantic, phaeodarianspecies appear to prefer a deeper habitat thanthose of Nassellaria and Spumellaria (Dworetzkyand Morley, 1987; Boltovskoy et al., 1993).Furthermore, at Station 6, the percent contribu-tion of Nassellaria in the surface water was muchlower than at other stations. The Nassellaria–Spumellaria ratio at Station 6 was strikinglydifferent from that of the other stations. Thisphenomenon appears to be due to the convergenceof water masses, as mentioned earlier. In a similarway, Boltovskoy and Jankilevich (1985) pointedout that in the eastern Equatorial Pacific, radi-olarian abundance and diversity dropped drama-tically in convergence regions, such as between theSEC and NECC or between the NECC and NorthEquatorial Current (NEC). Likely, the nassellar-
ian species occurred in low standing stock in thesurface water at Station 6, responding to a uniqueenvironment.In order to compare the specific depth ranges in
this study with those reported by other researchers(Renz, 1976; McMillen and Casey, 1978; Kling,1979; Dworetzky and Morley, 1987; Kling andBoltovskoy, 1995; Abelmann and Gowing, 1997),some species were concisely represented in atabular format (Fig. 10). In this study, it wasshown that most radiolarian species were char-acteristically distributed in various depth intervals.The vertical distribution profiles are not the sameat each station, because surface dwellers tend to beinfluenced by the change of variable surface waterenvironments. Abelmann and Gowing (1997) alsosuggested that radiolarian taxa of the surface-water habitats (o100m depth) were significantlyinfluenced by specific hydrographic parametersand showed restricted biogeographic distributionpatterns. It is conformable that surface dwellersdetermined in this study (Fig. 10) are mostlydistributed in the surface waters of other regions.The distribution patterns of the surface dwellersdo not change much with the geographic location.The rate of decrease of the surface dwellers withdepth closely resembles the observed temperaturedecrease. The vertical distributions of the surfacedwellers in the western Equatorial Pacific conformwell to that illustrated by Kling (1979). Inconclusion, it appears that the surface dwellersare mainly distributed in the mixed layer above thethermocline, and their distribution does notcharacteristically change in the higher/lower lati-tude locations.Two species (Didymocyrtis tetrathalamus tetra-
thalamus and Tetrapyle octacantha) are character-ized as surface dwellers in this study. They also aredistributed in the surface-zone of the central NorthPacific (Kling, 1979). Kling and Boltovskoy (1995)showed that they were practically warm-watertaxa. In the other studies these species were used asvarious indicators. For example, D. t. tetrathala-
mus is the key species for the western Pacificassemblage (Anderson et al., 1990) and T.
octacantha can be used as an indicator forupwelling regimes in mid and low latitudes(Abelmann and Gowing, 1997). In the western
H. Yamashita et al. / Deep-Sea Research II 49 (2002) 2823–28622836
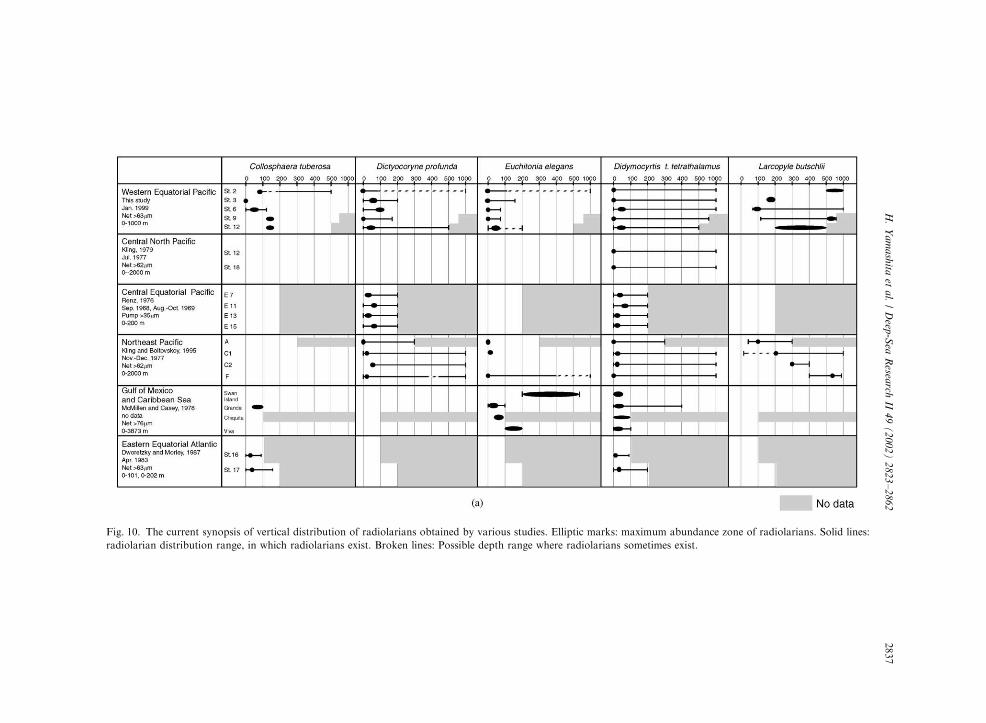
Fig. 10. The current synopsis of vertical distribution of radiolarians obtained by various studies. Elliptic marks: maximum abundance zone of radiolarians. Solid lines:
radiolarian distribution range, in which radiolarians exist. Broken lines: Possible depth range where radiolarians sometimes exist.
H.
Ya
ma
shita
eta
l./
Deep
-Sea
Resea
rchII
49
(2
00
2)
28
23
–2
86
22837
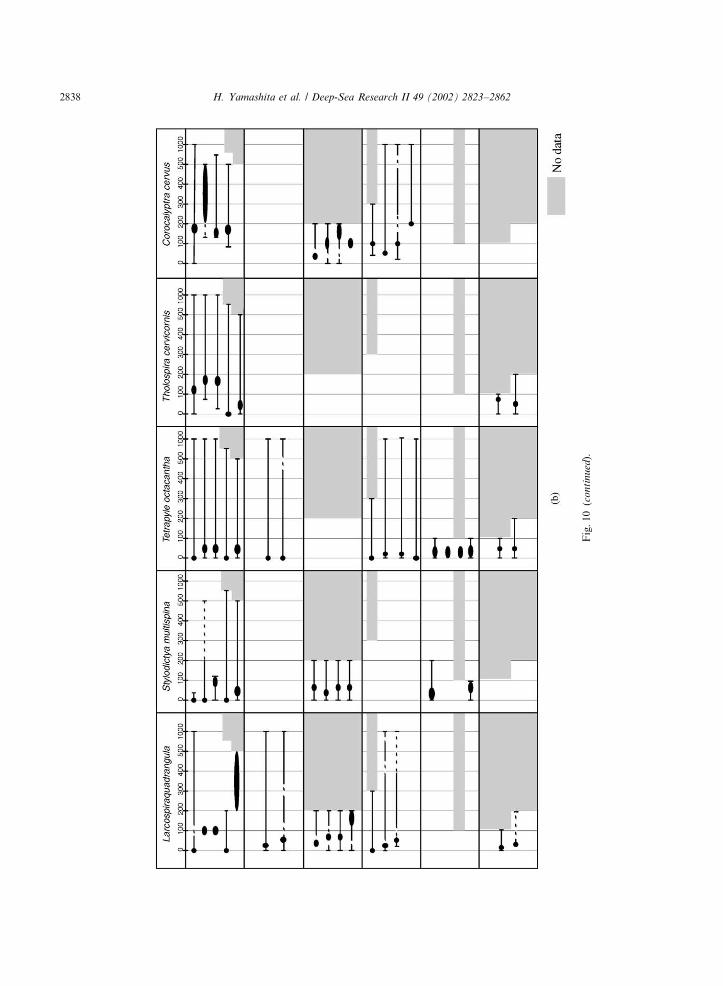
Fig.10(c
on
tin
ued).
H. Yamashita et al. / Deep-Sea Research II 49 (2002) 2823–28622838

Fig.10(c
on
tin
ued).
H. Yamashita et al. / Deep-Sea Research II 49 (2002) 2823–2862 2839

Equatorial Pacific, T. octacantha generally in-creased eastward, similar to the geographicaldistribution shown by Lombari and Boden(1985). At all stations except Station 6, D. t.
tetrathalamus abundantly occurred in the surfacewaters. Such results obtained here are in agree-ment with the information in the literature(Anderson et al., 1990).
Lithomelissa setosa, Pseudocubus obeliscus, Sty-
lodictya validispina, and Stylodictya multispina
peaked in the surface water as well, but weremuch more abundant at Stations 9 and 12 than atStations 2, 3, and 6. The low radiolarian diversityindices in the surface of Stations 9 and 12 are dueto the high relative abundances of these species.This result suggests that these species are closelylinked with the Upwelling Region of high nutri-ents. As previously mentioned, the WPWP ischaracterized by a specific temperature and salinityrange, the boundary between the WPWP Regionand the Upwelling Region lies near 1561E(Fig. 2a). However, according to the verticaldistributions of nutrients (Fig. 3) and of chlor-ophyll-a (Suzuki et al., 1999), the water masses inthe vicinity of Station 6 (1601E) can be regarded asnitrate/chlorophyll-poor (oligotrophic) water ofthe WPWP. This nitrate/chlorophyll-poor water isin several degrees to the east of the salinity front inthis cruise. There is a gap between the salinityfront and the increase of nitrate and chlorophyll-ain the equatorial Pacific (Rodier et al., 2000). Theoccurrences of these species can be correlated withtwo different water masses with characteristicradiolarian species habitats. Therefore, radiolarianspecies are useful tracers for water masses such asthe WPWP and the Upwelling Region. Further-more, L. setosa is sensitive to changes in theabundance of background primary producers andhas a great potential to be used as an environ-mental indicator in the subarctic Pacific (Takaha-shi, 1987). In the western and central EquatorialPacific, the occurrence of L. setosa also appears tobe related to the primary production and trophiccondition.In contrast to the above four species, the
abundance of Collosphaera tuberosa was higherat Stations 3 and 6 than at Stations 9 and 12. Itappears that this taxon prefers to thrive in the
oligotrophic environments in this region. It isknown that C. tuberosa is a colonial subtropicaland tropical shallow water form (Renz, 1976;McMillen and Casey, 1978; Kling, 1979; Dwor-etzky and Morley, 1987; Gowing, 1986). Colonialradiolarians are symbiont-bearing and generallymore abundant in association with a stable watercolumn and oligotrophic open-ocean environ-ments (e.g., Swanberg and Anderson, 1985;Takahashi et al., 1995; Amaral Zettler et al.,1999). Thus, C. tuberosa may be used as anindicator for the WPWP in the western EquatorialPacific. The low standing stock of C. tuberosa atStation 2 may be interpreted as an indicator ofoligotrophic conditions as suggested by the loweststanding stock of total radiolarians. However, it isnot clear how many shells are associated with acolonial form. The presence or absence of a colonymay lead to an over-estimate or under-estimate,depending on whether the colony is caught by thenet or sediment traps or not (Gowing, 1993;Welling et al., 1996). Therefore, the species withcolonial species should be treated with caution.The maximum abundances of the subsurface
dwellers occurred in the 120–200m layer. Thethermocline and chlorophyll-a maximum occur ateach station in the upper subsurface waters(Fig. 2b; Suzuki et al., 1999). The species classifiedas subsurface dwellers seem to selectively dwell inthe relatively eutrophic water, just below thethermocline, as it was observed in the centralNorth Pacific for Pseudodictyophimus gracilipes byKling (1979). Our results on the vertical distribu-tion of this taxon generally agree with this view:the occurrence of P. gracilipes is closely associatedwith the chlorophyll-a maximum.The vertical distributions of Spongurus cylindri-
cus and Eucecryphalus gegenbauri clearly show anincreased concentration in the subsurface water.Nevertheless temperature and nutrient densityvary with depth and longitude, their standingstocks vary little among stations. Thus, depth andother physical parameters appear to be moreimportant constrains for the distribution of thesespecies than trophic conditions and sea-surfacetemperature, which tend to vary significantly.The following species, Cladoscenium ancoratum,
Corocalyptra cervus, Litharachnium tentorium, and
H. Yamashita et al. / Deep-Sea Research II 49 (2002) 2823–28622840

Borgertella caudata, are classified as intermediatewater dwellers. The characteristic distribution ofthese species was a maximum abundance in the200–500m interval at Stations 2, 6, and 12. Inparticular at Station 6, these species showedconspicuous abundance peaks in this interval. B.
caudata (Phaeodaria) significantly contributes tototal phaeodarian standing stock. Takahashi andHonjo (1981) and Gowing (1993) suggested that B.
caudata occurs in significant numbers in relativelyshallow layers. That B. caudata had high standingstocks above 500m at all stations in this regionsupports Takahashi and Honjo’s (1981) andGowing’s (1993) observations. The maximumabundance of B. caudata and the other threespecies substantially deepened towards the east(Stations 3–9). Other phaeodarian species (e.g.,Challengeranium diodon) mostly showed this ten-dency. Litharachnium tentorium was reported to bean upper subsurface (50–100m) dweller (Kling,1979). However, in this study this taxon was seenin deeper waters than the upper subsurface zonedescribed by Kling (1979). Corocalyptra cervus wasfound deeper in the water column in the tropicsthan in high latitudes (e.g., Kling and Boltovskoy,1995). The distribution patterns of deep-waterassemblages are generally related to water tem-perature (Abelmann and Gowing, 1997). In ourstudy area, there is a deeper thermocline due to thepresence of WPWP than in other study areas(Fig. 2b). Therefore, the distributions of theintermediate water dwellers in the western Equa-torial Pacific such as in this study, appear to bedeeper than those of other areas (e.g., northeastand central Pacific). It is necessary to investigatethe habitats of these intermediate water dwellers inthe other parts of the Equatorial Pacific in order toclarify their linkages with environmental para-meters.The abundances of Peridium spinipes, Perome-
lissa phalacra, Lophophaena cylindrica and Zygo-
circus productus group did not differ amongstations. The vertical distribution patterns of thesespecies are similar to those of total radiolarians.Also, note that the standing stock of these speciessignificantly contributed to that of total radiolar-ians in this region. These species appear to stand abroad temperature and salinity range as suggested
by their vertical and horizontal distributionpatterns in this region. In a similar way, P. spinipes
was found to be abundant in the upper 200m ofthe Sargasso Sea (Casey et al., 1979). Becausethese species (P. spinipes, P. phalacra, L. cylindrica
and Z. productus group) mimic the total radiolar-ian distribution pattern today, they can be wiselyused to reconstruct the past total radiolarianproductivity. One must, be cautioned however,that Nassellaria tend to undergo dissolution morerapidly than Spumellaria in the sediments (Taka-hashi, 1981).At Station 6 the standing stock of the surface
dweller D. t. tetrathalamus was clearly much lowerthan at other stations. The horizontal distributionof D. t. tetrathalamus is roughly similar to thetendency of the surface diatom standing stock(Kobayashi and Takahashi, 2002). This result maysuggest that the abundance of the potential foodsupply governs the productivity of D. t. tetra-
thalamus, which is considered to be an herbivorousradiolarian (Anderson, 1983). In this region thefollowing species showed the same pattern(Acanthocorys cf. variabilis, Acrobotrys chelinobo-
trys, Dictyocoryne profunda, Theocorys veneris,Tholospyris sp. group, and Tetrapyle octacantha),illustrating the reduction of standing stock atStation 6 (Fig. 6) compared with other stations. Itmay be that these species cannot tolerate thedensity fluctuation due to a salinity-front in thevicinity of Station 6 or are restricted by HNLC(High Nutrient-Low Chlorophyll) area near Sta-tion 6 (Rodier et al., 2000).
5. Conclusions
The total radiolarian diversity indices in thewestern Equatorial Pacific were relatively high ascompared to those of studies in other parts of theworld. Comparing station-to-station diversity var-iations in the surface waters, the total radiolariandiversity in the WPWP Region is comparativelyhigher than that in the Upwelling Region. For thediversity, the equitability of the radiolarian speciesmakes a greater contribution than the speciesrichness, according to the numerical assessmentcarried out. The eutrophic water mass in the
H. Yamashita et al. / Deep-Sea Research II 49 (2002) 2823–2862 2841

Upwelling Region tends to migrate towards thewestern Pacific due to westward flows (NEC andSEC). The total radiolarian standing stock gradu-ally increased from west to east (Stations 2–12),responding to the change in nutrient regimes. Thedistribution pattern of surface water centricdiatoms, characterized by an increase from westto east (Kobayashi and Takahashi, 2002), paral-leled that of radiolarians.The total radiolarian standing stock in this
period is higher than that of previous reports (e.g.,Renz, 1976; Kling, 1979; Kling and Boltovskoy,1995) although it is impossible to simply compareour data with other reports because the samplingperiods and methods were different.The horizontal distributions of radiolarians and
surface diatoms along the Equator were character-ized by their low abundances at Station 6, locatednear the salinity front between the WPWP andUpwelling Regions. Therefore, the component ofradiolarian species and the Nassellaria–Spumellar-ia ratio are useful to detect the front and anomalyof water masses.Four species showed much higher standing
stocks in the Upwelling Region than that in theWPWP Region: Lithomelissa setosa, Pseudocubus
obeliscus, Stylodictya multispina, and Stylodictya
validispina. In contrast to this, the standing stockof Collosphaera tuberosa was high in the WPWPRegion. These species are good eutrophic andoligotrophic indicators respectively, and importanttracers of the present and past WPWP migrationin the western Equatorial Pacific.The vertical distribution patterns of Peridium
spinipes, Lophophaena cylindrica, Peromelissa pha-
lacra, and Zygocircus productus group are nearlythe same as that of total radiolarians, and thesespecies can tolerate considerable temperature andsalinity fluctuations. These species can representthe whole radiolarian productivity in the westernEquatorial Pacific.The surface dwellers (e.g., T. octacantha; D. t.
tetrathalamus) do not geographically change muchin the depth range, compared with their distribu-tions in several locations. However, occurrences ofthe intermediate water dwellers (e.g., Corocalyptra
cervus, Litharachnium tentorium) in this region aredeeper than those in high latitude locations. Thus,
nutrient conditions and food supply, which arelinked to the thermocline depth, influence thevertical distribution of these radiolarians.
Acknowledgements
We are privileged to have had an opportunity toparticipate in the GCMAPS Program led by Dr.Hodaka Kawahata of the Geological Survey ofJapan/Tohoku University. We thank the ChiefScientist Dr. Takeshi Kawano (JAMSTEC), cap-tain, crew, and scientists on board the R/V Mirai
Cruise MR98-K02 for assistance in collectingplankton samples and their advice in variousphases. Hirofumi Asahi participated in theMR98-K02 Cruise during which the planktontows were performed. Our thanks are extendedto Professor Motoyoshi Oda of Tohoku Univer-sity and Makoto Yamazaki and Rumiko Tazoe atKumamoto University for their cooperation inobtaining and splitting the samples. We aregrateful to an anonymous referee and to Dr.Demetrio Boltovskoy of Universidad de BuenosAires for his constructive comments responsiblefor a substantial improvement of the final manu-script. We thank Dr. Richard Jordan at YamagataUniversity, and Eileen S. Takahashi at PomonaCollege, who assisted us in the English editing ofthe early versions of the manuscript. K. Masakiand F. Kobayashi, Department of Earth andPlanetary Sciences, Kyushu University providedsignificant help and useful comments for ourlaboratory work and the writing of this manu-script. This study was supported by the followingresearch programs to KT: ‘Global Carbon Cycleand Related Mapping based on Satellite ImageryProgram (GCMAPS) promoted by the Scienceand Technology Agency of Japan; and Project No.10480128 of the Ministry of Education, Scienceand Culture of Japan.
Appendix A
List of radiolarian taxa encountered in theplankton tows at Stations 2, 3, 6, 9, and 12 of
H. Yamashita et al. / Deep-Sea Research II 49 (2002) 2823–28622842

Table 2
List of radiolarian taxa with proper references, which encountered in the plankton tows at Stations 2, 3, 6, 9 and 12 during R/V Mirai
Cruise MR98-K02
Spumellaria
Acanthosphaera tunis Haeckel, Takahashi, 1991, p. 193, Pl. 8, Figs. 2 and 3.
Axoprunum stauraxonium Haeckel, Takahashi, 1991, p. 205, Pl. 14, Figs. 8–10.
Actinosphaera capillacea (Haeckel), Takahashi, 1991, p. 195, Pl. 9, Figs. 4 and 5.
Actinomma arcadophorum Haeckel, Takahashi, 1991, p. 193, Pl. 8, Figs. 8 and 9.
Arachnosphaera myriacantha Haeckel, Takahashi, 1991, p. 197, Pl. 10, Figs. 11 and 12.
Actinosphaera tenella (Haeckel), Takahashi, 1991, p. 195, Pl. 9, Fig. 1.
Actinomma sp. Takahashi, 1991, p. 203, Pl. 13, Fig. 11.
Acrosphaera spinosa (Haeckel), Takahashi, 1991, p. 179, Pl. 1, Figs. 1 and 2.
Astrosphaera hexagonalis Haeckel, Takahashi, 1991, p. 199, Pl. 11, Figs. 1–3.
Amphirhopalum ypsilon Haeckel, Nigrini and Moore, 1979, p. S75, Pl. 10, Figs. 1a-e.
Actinomma medianum Nigrini, Nigrini and Moore, 1979, p. S26, Pl. 3, Figs. 5 and 6.
Acanthosphaera teunissima (Haeckel), Renz, 1976, p. 183, Pl. 2, Fig. 11.
Amphirhopalum straussii (Haeckel), Takahashi, 1991, p. 211, Pl. 17, Fig. 4.
Cladococcus cervicornis Haeckel, Takahashi, 1991, p. 197, Pl. 10, Figs. 8–10.
Cladococcus viminalis Haeckel, Bj�rklund, 1976, p. 1131, Pl. 1, Figs. 10–12.Cladococcus abietinus Haeckel, Takahashi, 1991, p. 197, Pl. 10, Fig. 5.
Cladococcus scoparius Haeckel, Takahashi and Honjo, 1981, p. 163, Pl. 2, Fig. 11.
Cromyechinus borealis (Cleve), Takahashi and Honjo, 1981, p. 163, Pl. 2, Fig. 8.
Collosphaera huxleyi M .uller, Takahashi, 1991, p. 181, Pl. 2, Figs. 8–11.
Collosphaera tuberosa Haeckel, Nigrini and Moore, 1979, p. S1, Pl. 1, Fig. 1.
Centrocubus octostylus Haeckel, Takahashi, 1991, p. 191, Pl. 7, Fig. 1.
Centrolonche hexalonche Popofsky, Takahashi and Honjo, 1981, p. 165, Pl. 3, Fig. 18.
Collosphaera macropora Popofsky, Takahashi, 1991, p. 181, Pl. 2, Figs. 13–18.
Cromyomma villosum Haeckel, Takahashi, 1991, p. 199, Pl. 11, Figs. 10 and 11.
Collosphaera confossa Takahashi, Takahashi, 1991, p. 181, Pl. 2, Fig. 4.
Dictyocoryne truncatum (Ehrenberg), Takahashi, 1991, p. 209, Pl. 16, Fig. 14.
Disolenia collina (Haeckel), Takahashi, 1991, p. 183, Pl. 3, Figs. 5–7.
Didymocyrtis tetrathulamus tetrathulamus (Haeckel), Takahashi, 1991, p. 219, Pl. 21, Figs. 1–14.
Dorydruppa bensoni Takahashi, Takahashi, 1991, p. 207, Pl. 15, Figs. 11–14.
Druppatractus ostracion group Haeckel, Takahashi, 1991, p. 205, Pl. 14, Figs. 3 and 4.
Dictyocoryne profunda Ehrenberg, Nigrini and Moore, 1979, p. S87, Pl. 12, Fig. 1.
Disolenia quadrata (Ehrenberg), Takahashi, 1991, p. 187, Pl. 5, Figs. 1–5.
Disolenia sp. A, Takahashi, 1991, p. 187, Pl. 5, Fig. 6.
Euchitonia cf. furcata Ehrenberg, Takahashi, 1991, p. 209, Pl. 16, Fig. 8.
Euchitonia elegans (Ehrenberg), Nigrini and Moore, 1979, p. S83, Pl. 11, Figs. 1a and b.
Euchitonia sp., Takahashi, 1991, p. 209, Pl. 16, Fig. 9.
Elatomma penicillus Haeckel, Takahashi, 1991, p. 195, Pl. 9, Figs. 9 and 10.
Heliodiscus asteriscus Haeckel, Nigrini and Moore, 1979, p. S73, Pl. 9, Figs. 1 and 2.
Hexalonche anaximandri Haeckel, Renz, 1976, p. 183, Pl. 2, Fig. 8.
Hexacontium axotrias Haeckel, Takahashi, 1991, p. 203, Pl. 13, Fig. 3.
Hexacontium hostile Cleve, Takahashi, 1991, p. 203, Pl. 13, Figs. 1 and 2.
Hexastylus sp., Takahashi, 1991, p. 201, Pl. 12, Fig. 9.
Helliodiscus echiniscus Haeckel, Takahashi, 1991, p. 223, Pl. 23, Figs. 4–6.
Hexacontium sp., Takahashi, 1991, p. 201, Pl. 12, Fig. 12.
Hexalonche sp. B, Takahashi, 1991, p. 201, Pl. 12, Figs. 10 and 11.
Hexalonche amphisiphon Haeckel, Takahashi, 1991, p. 201, Pl. 12, Figs. 13 and 14.
Hexapyle sp., Takahashi, 1991, p. 223, Pl. 23, Fig. 7.
Hexalonche sp. A, Takahashi, 1991, p. 199, Pl. 11, Figs. 14 and 15.
Haliomma sp. B, Takahashi, 1991, p. 201, Pl. 12, Fig. 15.
Hexacromyum elegans Haeckel, Takahashi, 1991, p. 203, Pl. 13, Figs. 4 and 5.
Lithelius minor J�rgensen, Nigrini and Moore, 1979, p. S135, Pl. 17, Figs. 3 and 4a and b.
Larcopyle butschlii Dreyer, Takahashi, 1991, p. 221, Pl. 22, Figs. 1–4.
Larcospira quadrangular Haeckel, Takahashi, 1991, p. 223, Pl. 23, Figs. 11 and 12.
H. Yamashita et al. / Deep-Sea Research II 49 (2002) 2823–2862 2843

Larcospira minor (J�rgensen), Bj�rklund, 1976, p. 1137, Pl. 5, Figs. 2–8.Larcopyle sp. A, Takahashi, 1991, p. 221, Pl. 22, Figs. 5 and 6.
Otosphaera auriculata Haeckel, Takahashi, 1991, p. 183, Pl. 3, Fig. 13.
Porodiscus sp., Nigrini and Moore, 1979, p. S107, Pl. 14, Figs. 1 and 2a and b.
Porodiscus micromma (Harting), Takahashi, 1991, p. 217, Pl. 20, Figs. 13 and 14.
Plegmosphaera entodictyon Haeckel, Takahashi, 1991, p. 189, Pl. 6, Figs. 8,10 and 11.
Pylolena armata Haeckel, Takahashi and Honjo, 1981, p. 171, Pl. 6, Fig. 4.
Polysolenia sp., Renz, 1976, p. 181, Pl. 1, Fig. 3.
Pylospira octopyle Haeckel, Nigrini and Moore, 1979, p. S139, Pl. 17, Figs. 6a-c.
Rhizoplegma boreale (Cleve), Bj�rklund, 1976, p. 1135, Pl. 4, Figs. 1–3.Spongodiscus spp. B group, Takahashi, 1991, p. 215, Pl. 19, Figs. 2 and 3.
Stylodictya sp., Takahashi, 1991, p. 215, Pl. 19, Figs. 12 and 13.
Stylodictya validispina J�rgensen, Nigrini and Moore, 1979, p. S103, Pl. 13, Figs. 5a and b.
Stylodictya multispina Haeckel, Takahashi, 1991, p. 217, Pl. 20, Figs. 10 and 12.
Spongaster pentas Riedel and Sanfilippo, Takahashi, 1991, p. 211, Pl. 17, Figs. 12–16.
Styptosphaera sp. A, Takahashi, 1991, p. 189, Pl. 6, Figs. 12–14.
Spongodiscus sp. A, Takahashi and Honjo, 1981, p. 167, Pl. 4, Fig. 13.
Stylospongia huxleyi Haeckel, Takahashi, 1991, p. 215, Pl. 19, Fig. 8.
Spongosphaera polycantha M .uller, Takahashi, 1991, p. 191, Pl. 7, Figs. 2,3 and 5.
Spongaster tetras tetras Ehrenberg, Takahashi and Honjo, 1981, p. 167, Pl. 4, Fig. 9.
Stylochlamydium venustum (Bailey), Takahashi, 1991, p. 217, Pl. 20, Fig. 11.
Spongurus cylindricus (Haeckel), Takahashi, 1991, p. 211, Pl. 17, Figs. 5–9.
Spongosphera sp., Takahashi, 1991, p. 191, Pl. 7, Fig. 4.
Streblacantha circumtexta J�rgensen, Bj�rklund, 1976, p. 1137, Pl. 5, Figs. 9–12.Styptosphaera spongiacea Haeckel, Takahashi, 1991, p. 189, Pl. 6, Figs. 6,7 and 9.
Stylosphaera melpomene Haeckel, Takahashi, 1991, p. 205, Pl. 14, Figs. 1 and 2.
Spongotrochus sp. A, Takahashi, 1991, p. 215, Pl. 19, Fig. 7.
Spongotrochus glaciaris Popofsky, Takahashi, 1991, p. 215, Pl. 19, Fig. 10.
Siphonosphaera martensi Brandt, Takahashi, 1991, p. 185, Pl. 4, Figs. 7 and 8.
Stylodictya aculeate J�rgensen, Nigrini and Moore, 1979, p. S101, Pl. 13, Figs. 3 and 4.
Spongodiscus bicoricarus Haeckel, Takahashi, 1991, p. 215, Pl. 19, Figs. 4–6.
Siphonosphaera magnisphaera Takahashi, Takahashi,1991, p. 185, Pl. 4, Fig. 1.
Spongopyle oculosa Dreyer, Takahashi, 1991, p. 217, Pl. 20, Figs. 1–4.
Siphonosphaera socialis Haeckel, Takahashi, 1991, p. 185, Pl. 4, Figs. 9–12.
Siphonosphaera sp. A, Takahashi, 1991, p. 185, Pl. 4, Fig. 2.
Thecosphaera inermis (Haeckel), Boltovskoy and Riedel, 1987, p. 88, Pl. 2, Fig 14.
Tholospira cervicornis Haeckel group, Takahashi, 1991, p. 221, Pl. 22, Figs. 7–9.
Tholospira dendrophora Haeckel, Takahashi, 1991, p. 221, Pl. 22, Fig. 11.
Tetrapyle octacantha M .uller, Takahashi and Honjo, 1981, p. 171, Pl. 6, Figs. 5 and 6.
Xiphosphaera tesseractis Dreyer, Takahashi, 1991, p. 201, Pl. 12, Figs. 3–5.
Nassellaria
Acrobotrys chelinobotrys Takahashi, Takahashi, 1991, p. 267, Pl. 45, Figs. 22–24.
Anthocrytidium ophirense (Ehrenberg), Renz, 1976, p. 191, Pl. 6, Fig. 25.
Anthocrytidium zanguebaricum (Ehrenberg), Nigrini and Moore, 1979, p. N69, Pl. 25, Fig. 2.
Arachnocalpis ovatiretalis Takahashi, Takahashi, 1991, p. 269, Pl. 46, Figs. 12–14.
Acanthocorys cf. variabilis Popofsky, Takahashi, 1991, p. 227, Pl. 25, Fig. 1.
Acrobotrys teralans Renz, Takahashi, 1991, p. 267, Pl. 45, Figs. 18–19.
Acanthodesmia vinculata M .uller, Takahashi, 1991, p. 233, Pl. 28, Figs. 6–8.
Antarctissa strelkovi Petrushevskaya, Nigrini and Moore, 1979, p. N5, Pl. 18, Figs. 2a and b.
Antarctissa denticulate (Ehrenberg), Nigrini and Moore, 1979, p. N1, Pl. 18, Figs. 1a and b.
Acrobotrys tessarolobon Takahashi, Takahashi, 1991, p. 267, Pl. 28, Fig. 20.
Botryocyrtis elongatum Takahashi, Takahashi, 1991, p. 269, Pl. 46, Figs. 8 and 9.
Botryostrobus aquilonaris (Bailey), Nigrini and Moore, 1979, p. N99, Pl. 27, Fig. 1.
Botryocyrtis scutum (Harting), Takahashi, 1991, p. 269, Pl. 46, Figs. 6 and 7.
Callimitra annae Haeckel, Takahashi, 1991, p. 229, Pl. 26, Fig. 15.
Carpocanistrum flosculum Haeckel, Takahashi, 1991, p. 267, Pl. 45, Figs. 4,6 and 7.
Table 2 (continued )
H. Yamashita et al. / Deep-Sea Research II 49 (2002) 2823–28622844

Corocalyptra cervus (Ehrenberg), Takahashi, 1991, p. 243, Pl. 33, Figs. 9–12.
Cladoscenium ancoratum Haeckel, Takahashi, 1991, p. 225, Pl. 24, Figs. 9–14.
Centrobotrys thermophila Petrushevskaya, Takahashi, 1991, p. 269, Pl. 46, Figs. 1 and 2.
Cytopera aglaolampa Takahashi, Takahashi, 1991, p. 257, Pl. 40, Figs. 7 and 8.
Clathrocyclas cassiopejae Haeckel, Takahashi, 1991, p. 245, Pl. 34, Figs. 12–14.
Conarachnium facetum (Haeckel), Takahashi, 1991, p. 255, Pl. 39, Fig. 7.
Clathrocanium coarctatum Ehrenberg, Takahashi, 1991, p. 229, Pl. 26, Figs. 11–13.
Clathrocorys murrayi Haeckel, Takahashi, 1991, p. 231, Pl. 27, Figs. 4–8.
Carpocanarium papillosum (Ehrenberg), Takahashi, 1991, p. 267, Pl. 45, Figs. 16 and 17.
Cantharospyris platybursa Haeckel, Takahashi, 1991, p. 239, Pl. 31, Fig. 5.
Carpocanistrum sp., Takahashi and Honjo, 1981, p. 179, Pl. 10, Figs. 21 and 22.
Coruntella profunda Ehrenberg, Takahashi and Honjo, 1981, p. 175, Pl. 8, Fig. 9.
Corocalyptra sp., Bj�rklund, 1976, p. 1152, Pl. 15, Fig. 24.Cyrtopera longuncula Haeckel, Takahashi and Honjo, 1981, p. 177, Pl. 9, Fig. 14.
Dictyophimus butschlii Haeckel, Takahashi and Honjo, 1981, p. 175, Pl. 8, Fig. 14.
Dictyocodon palladius Haeckel, Renz, 1976, p. 187, Pl. 4, Fig. 16.
Dictyophimus infabricatus Nigrini, Takahashi, 1991, p. 251, Pl. 37, Figs. 3–5.
Dictyophimus sp. B, Takahashi, 1991, p. 255, Pl. 39, Fig. 12.
Dictyophimus macropterus (Ehrenberg), Takahashi, 1991, p. 255, Pl. 39, Figs. 8–11.
Eucyrtidium hexagonatum Haeckel, Nigrini and Moore, 1979, p. N63, Pl. 24, Figs. 4a and b.
Eucrytidium anomalum Haeckel, Renz, 1976, p. 189, Pl. 5, Fig. 8.
Eucecryphalus sestrodiscus (Haeckel), Takahashi, 1991, p. 243, Pl. 33, Figs. 5,7 and 8.
Eucyrtidium acuminatum (Ehrenberg), Nigrini and Moore, 1979, p. N61, Pl. 24, Figs. 3a and b.
Eucecryphalus gegenbauri Haeckel, Takahashi, 1991, p. 243, Pl. 33, Figs. 13–15.
Eucyrtidium hexastichum (Haeckel), Takahashi, 1991, p. 261, Pl. 42, Fig. 22.
Eucyrtidium dictyopodium (Haeckel), Takahashi, 1991, p. 261, Pl. 42, Fig. 21.
Eucecryphalus tricostatus (Haeckel), Takahashi, 1991, p. 243, Pl. 33, Fig. 4.
Giraffospyris angulata (Haeckel), Renz, 1976, p. 195, Pl. 8, Fig. 5.
Lampromitra cachoni Petrushevskaya, Takahashi, 1991, p. 243, Pl. 33, Figs. 2 and 3.
Lophospyris pentagona pentagona (Ehrenberg), Takahashi and Honjo, 1981, p. 173, Pl. 7, Figs. 20 and 21.
Lophophaena cf.capito Ehrenberg, Takahashi and Honjo, 1981, p. 171, Pl. 6, Fig. 22.
Lophophaena cylindrical (Cleve), Takahashi, 1991, p. 227, Pl. 25, Figs. 3–5.
Lophospyris juvenile form group, Takahashi, 1991, p. 233, Pl. 28, Figs. 1–4.
Lophospyris pentagona hyperborea (J�rgensen), Takahashi and Honjo, 1981, p. 173, Pl. 7, Figs. 22–26.
Liriospyris thorax (Haeckel), Takahashi, 1991, p. 239, Pl. 31, Figs. 10–13.
Lamprocyclas maritalis maritalis Haeckel, Nigrini and Moore, 1979, p. N75, Pl. 25, Fig. 4.
Lophospyris pentagona quadriforis (Haeckel), Takahashi, 1991, p. 233, Pl. 28, Fig. 5.
Lithomelissa setosa J�rgensen, Takahashi, 1991, p. 227, Pl. 25, Figs. 16–22.Lampromitra spinosiretis Takahashi, Takahashi, 1991, p. 245, Pl. 34, Figs. 1,2 and 7.
Lophophaena decacantha (Haeckel) group, Takahashi, 1991, p. 227, Pl. 25, Figs. 2 and 10.
Litharachnium tentorium Haeckel, Takahashi and Honjo, 1981, p. 175, Pl. 8, Figs. 15 and 16.
Lithostrobus hexagonalis Haeckel, Takahashi and Honjo, 1981, p. 177, Pl. 9, Fig. 10.
Liriospyris reticulata (Ehrenberg), Nigrini and Moore, 1979, p. N13, Pl. 19, Figs. 4a and b.
Lampromitra schultzei (Haeckel), Takahashi, 1991, p. 241, Pl. 32, Figs. 4 and 5.
Lipmanella virchowii (Haeckel), Takahashi, 1991, p. 257, Pl. 40, Figs. 19–21.
Lipmanella pyramidale (Popofsky), Takahashi, 1991, p. 257, Pl. 40, Fig. 18.
Lipmanella dictyoceras (Haeckel), Takahashi, 1991, p. 257, Pl. 40, Fig. 17.
Litharachnium eupilium (Haeckel), Takahashi, 1991, p. 249, Pl. 36, Figs. 1–4.
Neobotrys quadrituberasa (Popofsky), Takahashi, 1991, p. 269, Pl. 46, Fig. 3.
Neosemantis distephanus Popofsky, Takahashi, 1991, p. 231, Pl. 27, Fig. 12.
Nephrospyris renilla renilla Haeckel, Takahashi, 1991, p. 237, Pl. 30, Figs. 7–9.
Phormospyris stabilis stabilis (Goll), Takahashi, 1991, p. 237, Pl. 30, Figs. 2–5.
Pterocanium grandiporus Nigrini, Takahashi, 1991, p. 249, Pl. 36, Figs. 8 and 9.
Pterocanium praetextum praetextum (Ehrenberg), Nigrini and Moore, 1979, p. N41, Pl. 23, Fig. 2.
Phormacantha hystrix (J�rgensen), Takahashi and Honjo, 1981, p. 171, Pl. 6, Figs. 17–19.
Pseudodictyophimus gracilipes (Bailey), Takahashi and Honjo, 1981, p. 177, Pl. 9, Figs. 3 and 4.
Pterocorys campanula Haeckel, Takahashi, 1991, p. 261, Pl. 42, Figs. 5–8.
Table 2 (continued )
H. Yamashita et al. / Deep-Sea Research II 49 (2002) 2823–2862 2845

Pterocorys zancleus (M .uller), Nigrini and Moore, 1979, p. N89, Pl. 25, Figs. 11a and b.
Peridium spinipes Haeckel, Takahashi, 1991, p. 229, Pl. 26, Figs. 4–6.
Pterocanium trilobum (Haeckel), Nigrini and Moore, 1979, p. N45, Pl. 23, Figs. 4a–c.
Peromelissa phalacra Haeckel, Takahashi and Honjo, 1981, p. 173, Pl. 7, Figs. 3–5.
Phormospyris stabilis capoi Goll, Takahashi, 1991, p. 235, Pl. 29, Figs. 15–18.
Phormospyris stabilis scaphipes (Haeckel), Nigrini and Moore, 1979, N19, Pl. 20, Figs. 2a and b.
Pseudocubus obeliscus Haeckel, Takahashi, 1991, p. 229, Pl. 26, Fig. 1.
Pteroscenium pinnatum Haeckel, Takahashi, 1991, p. 249, Pl. 36, Figs. 8 and 9.
Phormostichoartus corbula (Harting), Nigrini and Moore, 1979, p. N103, Pl. 27, Fig. 3.
Plectacantha sp., Bj�rklund, 1976, p. 1138, Pl. 6, Figs. 8–11.Saccospyris preantarctica Petrushevskaya, Takahashi, 1991, p. 267, Pl. 45, Fig. 21.
Stichopilium bicorne Haeckel, Nigrini and Moore, 1979, p. N91, Pl. 26, Figs. 1a and b.
Semantis gracilis Popofsky, Takahashi, 1991, p. 225, Pl. 24, Figs. 15 and 16.
Spirocyrtis subscalaris Nigrini, Takahashi, 1991, p. 265, Pl. 44, Figs. 3–6.
Spirocyrtis scalaris Haeckel, Renz, 1976, p. 191, Pl. 6, Fig. 1.
Spirocyrtis sp., Takahashi and Honjo, 1981, p. 179, Pl. 10, Fig. 16.
Tholospyris baconiana baconiana Goll, Takahashi, 1991, p. 239, Pl. 31, Fig. 7.
Theophormis callipilium Haeckel, Takahashi, 1991, p. 241, Pl. 32, Figs. 9–12.
Theocorys veneris Haeckel, Renz, 1976, p. 189, Pl. 5, Fig. 11.
Tholospyris sp. Group, Takahashi and Honjo, 1981, p. 173, Pl. 7, Fig. 16.
Tetraphormis dodecaster (Haeckel), Takahashi, 1991, p. 241, Pl. 32, Fig. 7.
Cycladophora davisiana cornutoides (Petrushevskaya), Takahashi, 1991, p. 259, Pl. 41, Figs. 12–16.
Triacartus undulatum (Popofsky), Takahashi, 1991, p. 257, Pl. 40, Figs. 9 and 10.
Tetraphormis rotula Haeckel, Takahashi, 1991, p. 241, Pl. 32, Figs. 1–3.
Theocorythium trachelium (Ehrenberg), Nigrini and Moore, 1979, p. N93, Pl. 26, Fig. 2.
Cycladophora bicornis (Popofsky), Takahashi, 1991, p. 259, Pl. 41, Figs. 4–6 and 8–11.
Tetraplecta pinigera Haeckel, Takahashi, 1991, p. 225, Pl. 24, Figs. 1–5.
Zygocircus sp. piscicaudatus Popofsky, Takahashi, 1991, p. 231, Pl. 27, Fig. 18.
Zygocircus productus (Hertwig) group, Takahashi, 1991, p. 231, Pl. 27, Figs. 13 and 14.
Phaeodaria
Aulographonium bicorne Haecker, Takahashi, 1991, p. 303, Pl. 63, Figs. 5 and 6.
Aulospthis taumorpha Haeckel, Takahashi, 1991, p. 303, Pl. 63, Figs. 7 and 8.
Auloceras spathillaster Haeckel, Takahashi, 1991, p. 303, Pl. 63, Fig. 4.
Borgertella caudata (Wallich), Takahashi, 1991, p. 285, Pl. 54, Figs. 13–17.
Castanella macropora (Borgert), Takahashi, 1991, p. 293, Pl. 58, Fig. 12.
Challengeron lingi Takahashi, Takahashi, 1991, p. 273, Pl. 48, Figs. 1–5.
Challengeron willemoesii Haeckel, Takahashi and Honjo, 1981, p. 179, Pl. 10, Figs. 25–29.
Challengeranium diodon Haeckel, Takahashi and Honjo, 1981, p. 183, Pl. 12, Figs. 1–3.
Castanella auleata Schmidt, Takahashi, 1991, p. 293, Pl. 58, Figs. 11 and 13.
Castanidium longispinum Haecker, Takahashi, 1991, p. 291, Pl. 57, Figs. 9–13.
Conchellium capsula Borgert, Takahashi and Honjo, 1981, p. 187, Pl. 14, Figs. 1–4.
Challengerosium avicularia Haecker, Takahashi, 1991, p. 275, Pl. 49, Figs. 1–13.
Challengeron tizardi (Murray), Takahashi, 1991, p. 273, Pl. 48, Figs. 11–16.
Circoporous sexfuscinus Haeckel, Takahashi, 1991, p. 297, Pl. 60, Figs. 1,3 and 5.
Euphysetta elegans Borgert, Takahashi and Honjo, 1981, p. 183, Pl. 12, Figs. 4 and 5.
Euphysetta staurocodon Haeckel, Takahashi, 1991, p. 283, Pl. 53, Figs. 11–14.
Euphysetta lucani Borgert, Takahashi and Honjo, 1981, p. 181, Pl. 11, Fig. 7.
Lirella melo (Cleve), Takahashi, 1991, p. 287, Pl. 55, Figs. 12–18.
Medusetta ansata Borgert, Takahashi and Honjo, 1981, p. 183, Pl. 12, Figs. 6 and 7.
Protocystis sp. A, Takahashi, 1991, p. 275, Pl. 49, Fig. 14 and 15.
Protocystis xiphodon (Haeckel), Takahashi, 1991, p. 281, Pl. 52, Figs. 1–3.
Protocystis sloggetti (Haeckel), Takahashi, 1991, p. 277, Pl. 50, Figs. 12–15.
Porospathis holostoma (Cleve), Takahashi, 1991, p. 291, Pl. 57, Figs. 1–8.
Protocystis anriculata Takahashi, Takahashi, 1991, p. 277, Pl. 50, Figs. 4–7.
Table 2 (continued )
H. Yamashita et al. / Deep-Sea Research II 49 (2002) 2823–28622846
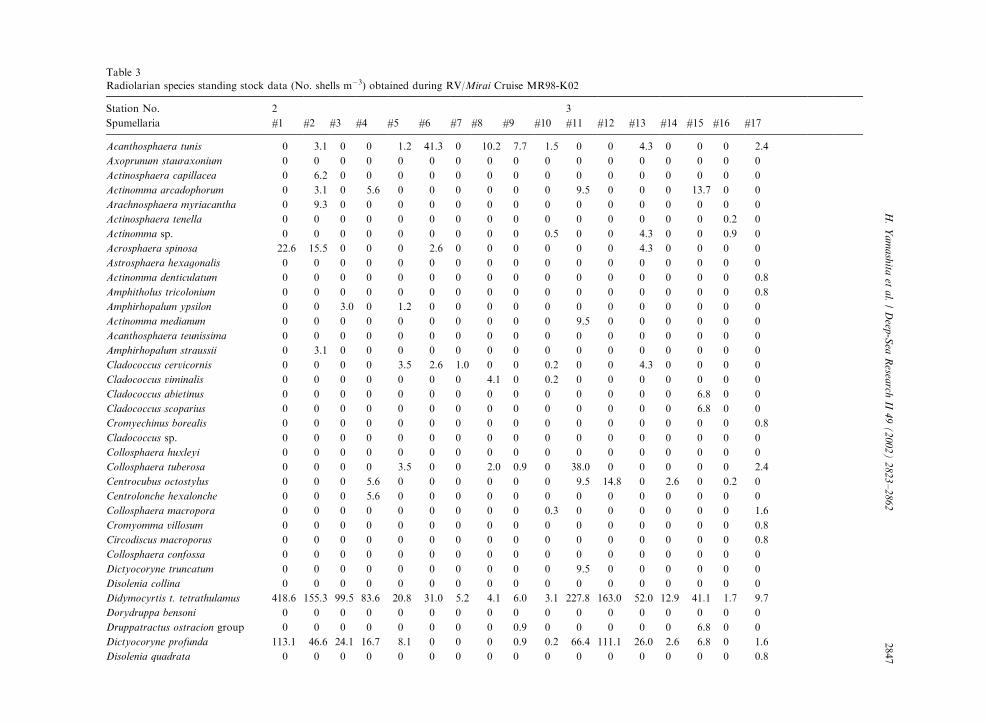
Table 3
Radiolarian species standing stock data (No. shells m�3) obtained during RV/Mirai Cruise MR98-K02
Station No. 2 3
Spumellaria #1 #2 #3 #4 #5 #6 #7 #8 #9 #10 #11 #12 #13 #14 #15 #16 #17
Acanthosphaera tunis 0 3.1 0 0 1.2 41.3 0 10.2 7.7 1.5 0 0 4.3 0 0 0 2.4
Axoprunum stauraxonium 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Actinosphaera capillacea 0 6.2 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Actinomma arcadophorum 0 3.1 0 5.6 0 0 0 0 0 0 9.5 0 0 0 13.7 0 0
Arachnosphaera myriacantha 0 9.3 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Actinosphaera tenella 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0.2 0
Actinomma sp. 0 0 0 0 0 0 0 0 0 0.5 0 0 4.3 0 0 0.9 0
Acrosphaera spinosa 22.6 15.5 0 0 0 2.6 0 0 0 0 0 0 4.3 0 0 0 0
Astrosphaera hexagonalis 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Actinomma denticulatum 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0.8
Amphitholus tricolonium 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0.8
Amphirhopalum ypsilon 0 0 3.0 0 1.2 0 0 0 0 0 0 0 0 0 0 0 0
Actinomma medianum 0 0 0 0 0 0 0 0 0 0 9.5 0 0 0 0 0 0
Acanthosphaera teunissima 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Amphirhopalum straussii 0 3.1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Cladococcus cervicornis 0 0 0 0 3.5 2.6 1.0 0 0 0.2 0 0 4.3 0 0 0 0
Cladococcus viminalis 0 0 0 0 0 0 0 4.1 0 0.2 0 0 0 0 0 0 0
Cladococcus abietinus 0 0 0 0 0 0 0 0 0 0 0 0 0 0 6.8 0 0
Cladococcus scoparius 0 0 0 0 0 0 0 0 0 0 0 0 0 0 6.8 0 0
Cromyechinus borealis 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0.8
Cladococcus sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Collosphaera huxleyi 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Collosphaera tuberosa 0 0 0 0 3.5 0 0 2.0 0.9 0 38.0 0 0 0 0 0 2.4
Centrocubus octostylus 0 0 0 5.6 0 0 0 0 0 0 9.5 14.8 0 2.6 0 0.2 0
Centrolonche hexalonche 0 0 0 5.6 0 0 0 0 0 0 0 0 0 0 0 0 0
Collosphaera macropora 0 0 0 0 0 0 0 0 0 0.3 0 0 0 0 0 0 1.6
Cromyomma villosum 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0.8
Circodiscus macroporus 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0.8
Collosphaera confossa 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Dictyocoryne truncatum 0 0 0 0 0 0 0 0 0 0 9.5 0 0 0 0 0 0
Disolenia collina 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Didymocyrtis t. tetrathulamus 418.6 155.3 99.5 83.6 20.8 31.0 5.2 4.1 6.0 3.1 227.8 163.0 52.0 12.9 41.1 1.7 9.7
Dorydruppa bensoni 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Druppatractus ostracion group 0 0 0 0 0 0 0 0 0.9 0 0 0 0 0 6.8 0 0
Dictyocoryne profunda 113.1 46.6 24.1 16.7 8.1 0 0 0 0.9 0.2 66.4 111.1 26.0 2.6 6.8 0 1.6
Disolenia quadrata 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0.8
H.
Ya
ma
shita
eta
l./
Deep
-Sea
Resea
rchII
49
(2
00
2)
28
23
–2
86
22847
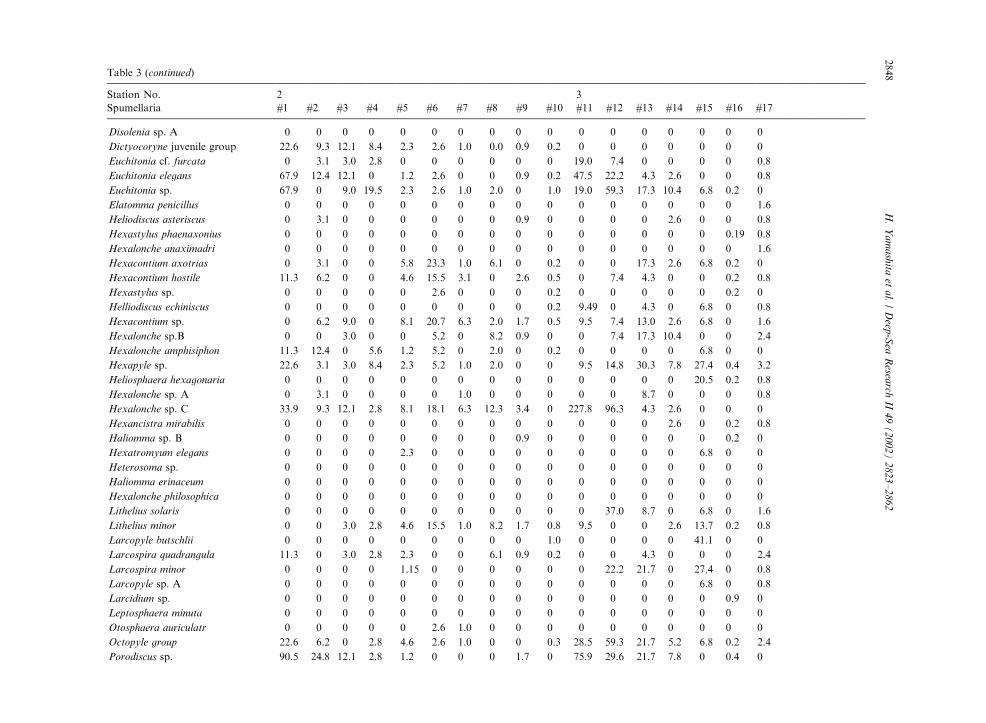
Disolenia sp. A 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Dictyocoryne juvenile group 22.6 9.3 12.1 8.4 2.3 2.6 1.0 0.0 0.9 0.2 0 0 0 0 0 0 0
Euchitonia cf. furcata 0 3.1 3.0 2.8 0 0 0 0 0 0 19.0 7.4 0 0 0 0 0.8
Euchitonia elegans 67.9 12.4 12.1 0 1.2 2.6 0 0 0.9 0.2 47.5 22.2 4.3 2.6 0 0 0.8
Euchitonia sp. 67.9 0 9.0 19.5 2.3 2.6 1.0 2.0 0 1.0 19.0 59.3 17.3 10.4 6.8 0.2 0
Elatomma penicillus 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1.6
Heliodiscus asteriscus 0 3.1 0 0 0 0 0 0 0.9 0 0 0 0 2.6 0 0 0.8
Hexastylus phaenaxonius 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0.19 0.8
Hexalonche anaximadri 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1.6
Hexacontium axotrias 0 3.1 0 0 5.8 23.3 1.0 6.1 0 0.2 0 0 17.3 2.6 6.8 0.2 0
Hexacontium hostile 11.3 6.2 0 0 4.6 15.5 3.1 0 2.6 0.5 0 7.4 4.3 0 0 0.2 0.8
Hexastylus sp. 0 0 0 0 0 2.6 0 0 0 0.2 0 0 0 0 0 0.2 0
Helliodiscus echiniscus 0 0 0 0 0 0 0 0 0 0.2 9.49 0 4.3 0 6.8 0 0.8
Hexacontium sp. 0 6.2 9.0 0 8.1 20.7 6.3 2.0 1.7 0.5 9.5 7.4 13.0 2.6 6.8 0 1.6
Hexalonche sp.B 0 0 3.0 0 0 5.2 0 8.2 0.9 0 0 7.4 17.3 10.4 0 0 2.4
Hexalonche amphisiphon 11.3 12.4 0 5.6 1.2 5.2 0 2.0 0 0.2 0 0 0 0 6.8 0 0
Hexapyle sp. 22.6 3.1 3.0 8.4 2.3 5.2 1.0 2.0 0 0 9.5 14.8 30.3 7.8 27.4 0.4 3.2
Heliosphaera hexagonaria 0 0 0 0 0 0 0 0 0 0 0 0 0 0 20.5 0.2 0.8
Hexalonche sp. A 0 3.1 0 0 0 0 1.0 0 0 0 0 0 8.7 0 0 0 0.8
Hexalonche sp. C 33.9 9.3 12.1 2.8 8.1 18.1 6.3 12.3 3.4 0 227.8 96.3 4.3 2.6 0 0 0
Hexancistra mirabilis 0 0 0 0 0 0 0 0 0 0 0 0 0 2.6 0 0.2 0.8
Haliomma sp. B 0 0 0 0 0 0 0 0 0.9 0 0 0 0 0 0 0.2 0
Hexatromyum elegans 0 0 0 0 2.3 0 0 0 0 0 0 0 0 0 6.8 0 0
Heterosoma sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Haliomma erinaceum 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Hexalonche philosophica 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Lithelius solaris 0 0 0 0 0 0 0 0 0 0 0 37.0 8.7 0 6.8 0 1.6
Lithelius minor 0 0 3.0 2.8 4.6 15.5 1.0 8.2 1.7 0.8 9.5 0 0 2.6 13.7 0.2 0.8
Larcopyle butschlii 0 0 0 0 0 0 0 0 0 1.0 0 0 0 0 41.1 0 0
Larcospira quadrangula 11.3 0 3.0 2.8 2.3 0 0 6.1 0.9 0.2 0 0 4.3 0 0 0 2.4
Larcospira minor 0 0 0 0 1.15 0 0 0 0 0 0 22.2 21.7 0 27.4 0 0.8
Larcopyle sp. A 0 0 0 0 0 0 0 0 0 0 0 0 0 0 6.8 0 0.8
Larcidium sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0.9 0
Leptosphaera minuta 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Otosphaera auriculatr 0 0 0 0 0 2.6 1.0 0 0 0 0 0 0 0 0 0 0
Octopyle group 22.6 6.2 0 2.8 4.6 2.6 1.0 0 0 0.3 28.5 59.3 21.7 5.2 6.8 0.2 2.4
Porodiscus sp. 90.5 24.8 12.1 2.8 1.2 0 0 0 1.7 0 75.9 29.6 21.7 7.8 0 0.4 0
Table 3 (continued)
Station No. 2 3
Spumellaria #1 #2 #3 #4 #5 #6 #7 #8 #9 #10 #11 #12 #13 #14 #15 #16 #17H
.Y
am
ash
itaet
al.
/D
eep-S
eaR
esearch
II4
9(
20
02
)2
82
3–
28
62
2848

Phorticum pylonium 0 0 0 0 0 0 0 0 0 0 0 7.41 0 0 0 0 0
Pylodiscus sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Porodiscus micromma 22.6 0 0 0 0 0 1.0 2.0 0.9 0.2 9.5 0 0 2.6 0 0 0
Plegmosphaera entodictyon 0 6.2 0 2.8 0 0 1.0 0 0 0 9.5 7.4 13.0 2.6 6.8 0.2 1.6
Pyolena armata 0 0 0 0 0 0 0 0 0 0 9.5 0 0 0 6.8 0 0
Polysolenia sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 6.8 0 0
Porospira odtopyle 0 0 0 0 0 0 0 0 0 0 9.5 0 0 0 0 0.2 0
Pylospira octopyle 0 0 3.0 0 0 0 0 0 0 0 0 7.4 4.3 2.6 0 0 0.8
Pityomma scoparium 0 0 0 0 0 0 0 0 0 0 0 0 0 0 6.8 0 0
Rhizoplegma boreale 0 0 0 0 0 0 0 0 0 0.2 0 0 0 0 0 0 0
Spongodiscus spp. B group 135.8 9.3 48.3 8.4 4.6 2.6 0 0 1.7 0.7 123.4 66.7 39.0 2.6 20.5 0.2 2.4
Staurancistra sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Stylodictya sp. 0 0 3.0 0 0 0 1.0 0 1.7 0 0 0 0 2.6 27.4 0.2 4.9
Stylodictya validispina 11.3 6.2 3.0 2.8 1.2 0 0 0 0 0 28.5 29.6 13.0 0 0 0 0
Stylodictya multispina 11.3 3.1 0 0 0 0 0 0 0 0 19.0 14.8 17.3 0 13.7 0.6 0
Spongaster pentas 22.6 3.1 0 0 0 0 0 0 0 0.2 104.4 29.6 56.4 7.8 27.4 1.1 6.5
Styptosphaera sp. A 0 0 0 0 0 0 0 0 0 0 0 0 4.3 0 0 0.4 2.4
Stylacontarium sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Spongodiscus sp. A 45.3 0 9.0 8.4 0 0 0 0 0 0.5 19.0 0 0 0 0 0 0
Stylospongia huxleyi 0 0 0 0 0 0 0 0 0 0 0 0 0 2.6 0 1.1 0
Spongosphaera polycantha 11.3 0 0 0 1.2 0 0 0 0 0 0 0 0 0 0 0.6 0
Spongaster tetras tetras 22.6 0 6.0 0 0 0 1.0 2.0 0 0.3 19.0 44.5 8.7 0 0 0.6 0.8
Stylochlamydium venustum 0 0 0 0 0 0 0 0 0.9 0.2 0 0 0 0 0 0 0
Spongurus cylindricus 0 0 0 0 0 0 16.8 12.3 0 0.2 0 7.4 17.3 12.9 27.4 1.1 0
Spongsphera sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 5.18 0 0.2 0
Streblacantha circumtexta 0 0 0 0 0 0 0 0 0 0 0 0 8.7 12.9 0 1.9 4.9
Styptosphaera spongiacea 11.3 0 0 0 0 0 0 0 0 0 0 7.4 0 0 0 0 0
Stylosphaera melpomene 0 0 0 0 1.2 0 0 0 0 0.2 0 0 0 0 6.8 0 0
Staurodictyo cruciata 22.6 0 6.0 2.8 1.2 0 0 0 1.7 0.3 9.5 14.8 0 0 20.5 0 0
Spongotrochus sp. A 0 0 0 0 0 0 0 0 0 0 0 7.4 0 0 0 0 0
Spongotrochus glaciaris 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Siphonosphaera martensi 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Stylodictya aculeata 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Spongodiscus bicoricarus 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Siphonosphaera magnisphaera 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Spongopyle oculosa 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Siphonosphaera socialis 0 0 0 0 0 0 0 0 0 0.2 0 0 0 0 0 0 0
Siphonosphaera sp. A 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Stylodictya sp. A 0 0 6.0 0 0 0 0 0 0 0.3 0 0 0 0 0 0 0
Thecosphaera inermis 0 0 0 0 0 2.58 0 0 0.86 0 0 0 0 0 0 0 0
Tholospira cervicornis group 11.3 3.1 3.0 8.4 3.5 28.4 18.9 18.4 13.7 4.8 19.0 51.9 52.0 51.8 130.0 2.3 7.3
H.
Ya
ma
shita
eta
l./
Deep
-Sea
Resea
rchII
49
(2
00
2)
28
23
–2
86
22849
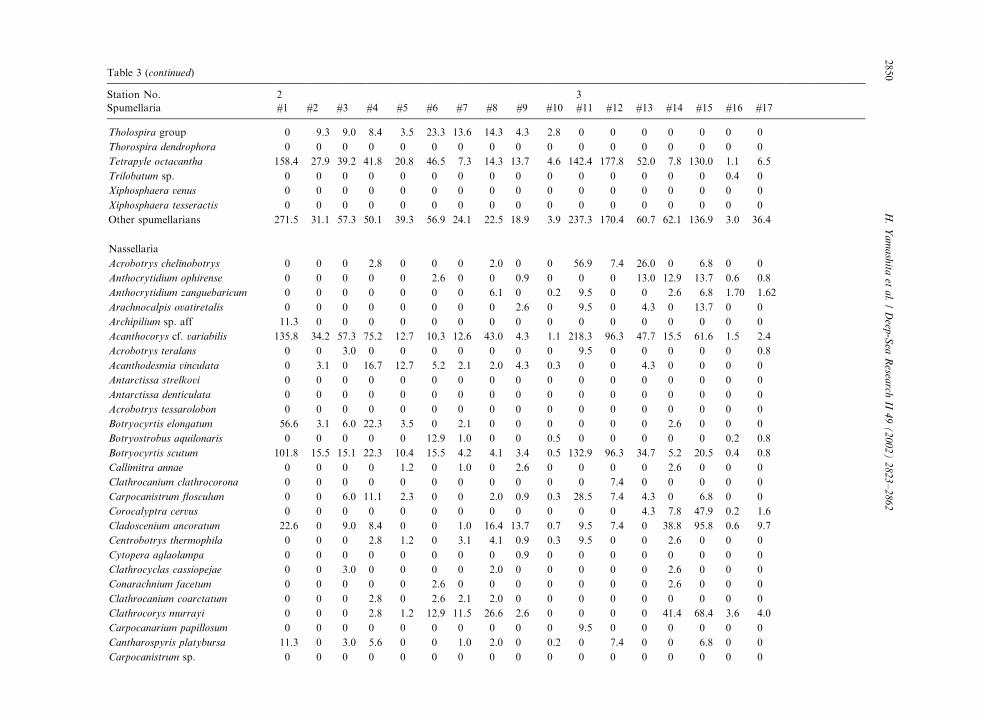
Tholospira group 0 9.3 9.0 8.4 3.5 23.3 13.6 14.3 4.3 2.8 0 0 0 0 0 0 0
Thorospira dendrophora 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Tetrapyle octacantha 158.4 27.9 39.2 41.8 20.8 46.5 7.3 14.3 13.7 4.6 142.4 177.8 52.0 7.8 130.0 1.1 6.5
Trilobatum sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0.4 0
Xiphosphaera venus 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Xiphosphaera tesseractis 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Other spumellarians 271.5 31.1 57.3 50.1 39.3 56.9 24.1 22.5 18.9 3.9 237.3 170.4 60.7 62.1 136.9 3.0 36.4
Nassellaria
Acrobotrys chelinobotrys 0 0 0 2.8 0 0 0 2.0 0 0 56.9 7.4 26.0 0 6.8 0 0
Anthocrytidium ophirense 0 0 0 0 0 2.6 0 0 0.9 0 0 0 13.0 12.9 13.7 0.6 0.8
Anthocrytidium zanguebaricum 0 0 0 0 0 0 0 6.1 0 0.2 9.5 0 0 2.6 6.8 1.70 1.62
Arachnocalpis ovatiretalis 0 0 0 0 0 0 0 0 2.6 0 9.5 0 4.3 0 13.7 0 0
Archipilium sp. aff 11.3 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Acanthocorys cf. variabilis 135.8 34.2 57.3 75.2 12.7 10.3 12.6 43.0 4.3 1.1 218.3 96.3 47.7 15.5 61.6 1.5 2.4
Acrobotrys teralans 0 0 3.0 0 0 0 0 0 0 0 9.5 0 0 0 0 0 0.8
Acanthodesmia vinculata 0 3.1 0 16.7 12.7 5.2 2.1 2.0 4.3 0.3 0 0 4.3 0 0 0 0
Antarctissa strelkovi 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Antarctissa denticulata 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Acrobotrys tessarolobon 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Botryocyrtis elongatum 56.6 3.1 6.0 22.3 3.5 0 2.1 0 0 0 0 0 0 2.6 0 0 0
Botryostrobus aquilonaris 0 0 0 0 0 12.9 1.0 0 0 0.5 0 0 0 0 0 0.2 0.8
Botryocyrtis scutum 101.8 15.5 15.1 22.3 10.4 15.5 4.2 4.1 3.4 0.5 132.9 96.3 34.7 5.2 20.5 0.4 0.8
Callimitra annae 0 0 0 0 1.2 0 1.0 0 2.6 0 0 0 0 2.6 0 0 0
Clathrocanium clathrocorona 0 0 0 0 0 0 0 0 0 0 0 7.4 0 0 0 0 0
Carpocanistrum flosculum 0 0 6.0 11.1 2.3 0 0 2.0 0.9 0.3 28.5 7.4 4.3 0 6.8 0 0
Corocalyptra cervus 0 0 0 0 0 0 0 0 0 0 0 0 4.3 7.8 47.9 0.2 1.6
Cladoscenium ancoratum 22.6 0 9.0 8.4 0 0 1.0 16.4 13.7 0.7 9.5 7.4 0 38.8 95.8 0.6 9.7
Centrobotrys thermophila 0 0 0 2.8 1.2 0 3.1 4.1 0.9 0.3 9.5 0 0 2.6 0 0 0
Cytopera aglaolampa 0 0 0 0 0 0 0 0 0.9 0 0 0 0 0 0 0 0
Clathrocyclas cassiopejae 0 0 3.0 0 0 0 0 2.0 0 0 0 0 0 2.6 0 0 0
Conarachnium facetum 0 0 0 0 0 2.6 0 0 0 0 0 0 0 2.6 0 0 0
Clathrocanium coarctatum 0 0 0 2.8 0 2.6 2.1 2.0 0 0 0 0 0 0 0 0 0
Clathrocorys murrayi 0 0 0 2.8 1.2 12.9 11.5 26.6 2.6 0 0 0 0 41.4 68.4 3.6 4.0
Carpocanarium papillosum 0 0 0 0 0 0 0 0 0 0 9.5 0 0 0 0 0 0
Cantharospyris platybursa 11.3 0 3.0 5.6 0 0 1.0 2.0 0 0.2 0 7.4 0 0 6.8 0 0
Carpocanistrum sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Table 3 (continued)
Station No. 2 3
Spumellaria #1 #2 #3 #4 #5 #6 #7 #8 #9 #10 #11 #12 #13 #14 #15 #16 #17H
.Y
am
ash
itaet
al.
/D
eep-S
eaR
esearch
II4
9(
20
02
)2
82
3–
28
62
2850
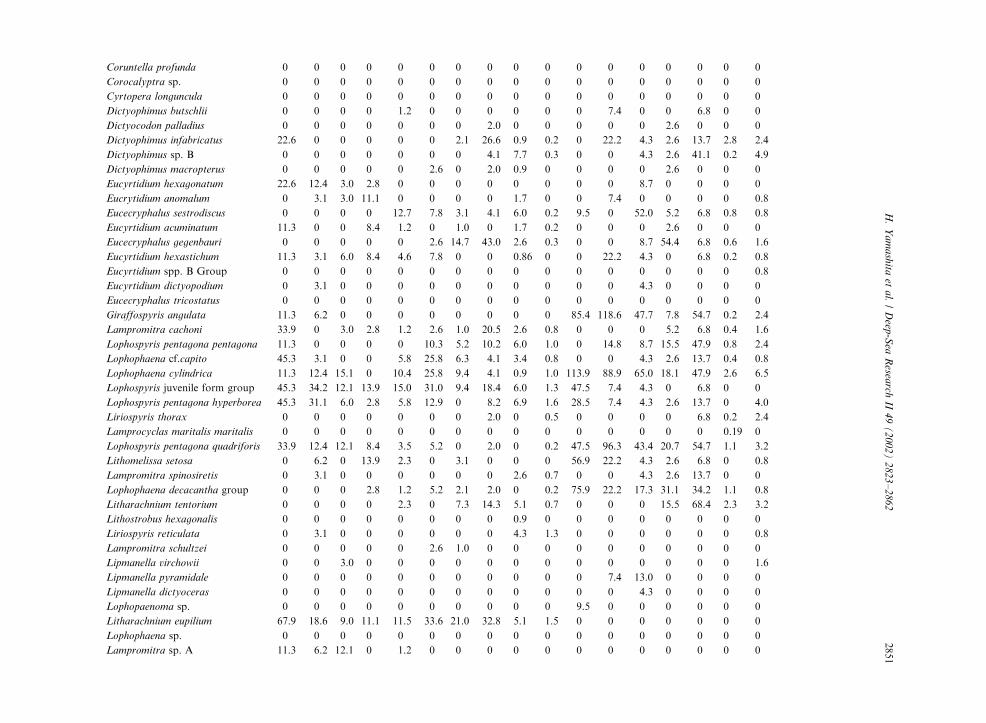
Coruntella profunda 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Corocalyptra sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Cyrtopera longuncula 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Dictyophimus butschlii 0 0 0 0 1.2 0 0 0 0 0 0 7.4 0 0 6.8 0 0
Dictyocodon palladius 0 0 0 0 0 0 0 2.0 0 0 0 0 0 2.6 0 0 0
Dictyophimus infabricatus 22.6 0 0 0 0 0 2.1 26.6 0.9 0.2 0 22.2 4.3 2.6 13.7 2.8 2.4
Dictyophimus sp. B 0 0 0 0 0 0 0 4.1 7.7 0.3 0 0 4.3 2.6 41.1 0.2 4.9
Dictyophimus macropterus 0 0 0 0 0 2.6 0 2.0 0.9 0 0 0 0 2.6 0 0 0
Eucyrtidium hexagonatum 22.6 12.4 3.0 2.8 0 0 0 0 0 0 0 0 8.7 0 0 0 0
Eucrytidium anomalum 0 3.1 3.0 11.1 0 0 0 0 1.7 0 0 7.4 0 0 0 0 0.8
Eucecryphalus sestrodiscus 0 0 0 0 12.7 7.8 3.1 4.1 6.0 0.2 9.5 0 52.0 5.2 6.8 0.8 0.8
Eucyrtidium acuminatum 11.3 0 0 8.4 1.2 0 1.0 0 1.7 0.2 0 0 0 2.6 0 0 0
Eucecryphalus gegenbauri 0 0 0 0 0 2.6 14.7 43.0 2.6 0.3 0 0 8.7 54.4 6.8 0.6 1.6
Eucyrtidium hexastichum 11.3 3.1 6.0 8.4 4.6 7.8 0 0 0.86 0 0 22.2 4.3 0 6.8 0.2 0.8
Eucyrtidium spp. B Group 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0.8
Eucyrtidium dictyopodium 0 3.1 0 0 0 0 0 0 0 0 0 0 4.3 0 0 0 0
Eucecryphalus tricostatus 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Giraffospyris angulata 11.3 6.2 0 0 0 0 0 0 0 0 85.4 118.6 47.7 7.8 54.7 0.2 2.4
Lampromitra cachoni 33.9 0 3.0 2.8 1.2 2.6 1.0 20.5 2.6 0.8 0 0 0 5.2 6.8 0.4 1.6
Lophospyris pentagona pentagona 11.3 0 0 0 0 10.3 5.2 10.2 6.0 1.0 0 14.8 8.7 15.5 47.9 0.8 2.4
Lophophaena cf.capito 45.3 3.1 0 0 5.8 25.8 6.3 4.1 3.4 0.8 0 0 4.3 2.6 13.7 0.4 0.8
Lophophaena cylindrica 11.3 12.4 15.1 0 10.4 25.8 9.4 4.1 0.9 1.0 113.9 88.9 65.0 18.1 47.9 2.6 6.5
Lophospyris juvenile form group 45.3 34.2 12.1 13.9 15.0 31.0 9.4 18.4 6.0 1.3 47.5 7.4 4.3 0 6.8 0 0
Lophospyris pentagona hyperborea 45.3 31.1 6.0 2.8 5.8 12.9 0 8.2 6.9 1.6 28.5 7.4 4.3 2.6 13.7 0 4.0
Liriospyris thorax 0 0 0 0 0 0 0 2.0 0 0.5 0 0 0 0 6.8 0.2 2.4
Lamprocyclas maritalis maritalis 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0.19 0
Lophospyris pentagona quadriforis 33.9 12.4 12.1 8.4 3.5 5.2 0 2.0 0 0.2 47.5 96.3 43.4 20.7 54.7 1.1 3.2
Lithomelissa setosa 0 6.2 0 13.9 2.3 0 3.1 0 0 0 56.9 22.2 4.3 2.6 6.8 0 0.8
Lampromitra spinosiretis 0 3.1 0 0 0 0 0 0 2.6 0.7 0 0 4.3 2.6 13.7 0 0
Lophophaena decacantha group 0 0 0 2.8 1.2 5.2 2.1 2.0 0 0.2 75.9 22.2 17.3 31.1 34.2 1.1 0.8
Litharachnium tentorium 0 0 0 0 2.3 0 7.3 14.3 5.1 0.7 0 0 0 15.5 68.4 2.3 3.2
Lithostrobus hexagonalis 0 0 0 0 0 0 0 0 0.9 0 0 0 0 0 0 0 0
Liriospyris reticulata 0 3.1 0 0 0 0 0 0 4.3 1.3 0 0 0 0 0 0 0.8
Lampromitra schultzei 0 0 0 0 0 2.6 1.0 0 0 0 0 0 0 0 0 0 0
Lipmanella virchowii 0 0 3.0 0 0 0 0 0 0 0 0 0 0 0 0 0 1.6
Lipmanella pyramidale 0 0 0 0 0 0 0 0 0 0 0 7.4 13.0 0 0 0 0
Lipmanella dictyoceras 0 0 0 0 0 0 0 0 0 0 0 0 4.3 0 0 0 0
Lophopaenoma sp. 0 0 0 0 0 0 0 0 0 0 9.5 0 0 0 0 0 0
Litharachnium eupilium 67.9 18.6 9.0 11.1 11.5 33.6 21.0 32.8 5.1 1.5 0 0 0 0 0 0 0
Lophophaena sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Lampromitra sp. A 11.3 6.2 12.1 0 1.2 0 0 0 0 0 0 0 0 0 0 0 0
H.
Ya
ma
shita
eta
l./
Deep
-Sea
Resea
rchII
49
(2
00
2)
28
23
–2
86
22851
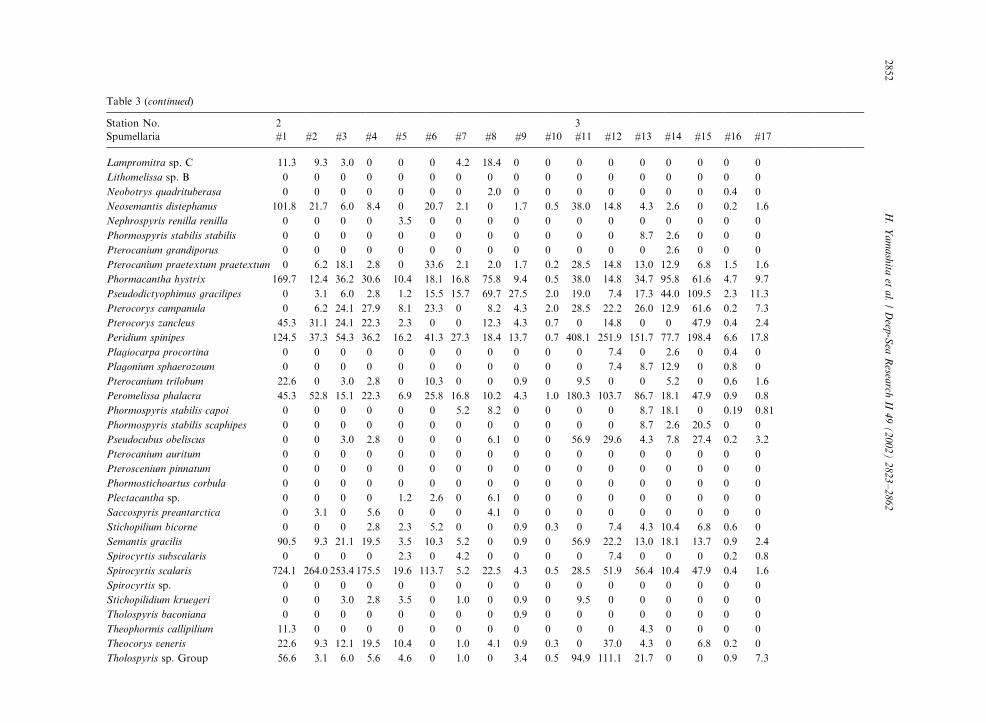
Lampromitra sp. C 11.3 9.3 3.0 0 0 0 4.2 18.4 0 0 0 0 0 0 0 0 0
Lithomelissa sp. B 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Neobotrys quadrituberasa 0 0 0 0 0 0 0 2.0 0 0 0 0 0 0 0 0.4 0
Neosemantis distephanus 101.8 21.7 6.0 8.4 0 20.7 2.1 0 1.7 0.5 38.0 14.8 4.3 2.6 0 0.2 1.6
Nephrospyris renilla renilla 0 0 0 0 3.5 0 0 0 0 0 0 0 0 0 0 0 0
Phormospyris stabilis stabilis 0 0 0 0 0 0 0 0 0 0 0 0 8.7 2.6 0 0 0
Pterocanium grandiporus 0 0 0 0 0 0 0 0 0 0 0 0 0 2.6 0 0 0
Pterocanium praetextum praetextum 0 6.2 18.1 2.8 0 33.6 2.1 2.0 1.7 0.2 28.5 14.8 13.0 12.9 6.8 1.5 1.6
Phormacantha hystrix 169.7 12.4 36.2 30.6 10.4 18.1 16.8 75.8 9.4 0.5 38.0 14.8 34.7 95.8 61.6 4.7 9.7
Pseudodictyophimus gracilipes 0 3.1 6.0 2.8 1.2 15.5 15.7 69.7 27.5 2.0 19.0 7.4 17.3 44.0 109.5 2.3 11.3
Pterocorys campanula 0 6.2 24.1 27.9 8.1 23.3 0 8.2 4.3 2.0 28.5 22.2 26.0 12.9 61.6 0.2 7.3
Pterocorys zancleus 45.3 31.1 24.1 22.3 2.3 0 0 12.3 4.3 0.7 0 14.8 0 0 47.9 0.4 2.4
Peridium spinipes 124.5 37.3 54.3 36.2 16.2 41.3 27.3 18.4 13.7 0.7 408.1 251.9 151.7 77.7 198.4 6.6 17.8
Plagiocarpa procortina 0 0 0 0 0 0 0 0 0 0 0 7.4 0 2.6 0 0.4 0
Plagonium sphaerozoum 0 0 0 0 0 0 0 0 0 0 0 7.4 8.7 12.9 0 0.8 0
Pterocanium trilobum 22.6 0 3.0 2.8 0 10.3 0 0 0.9 0 9.5 0 0 5.2 0 0.6 1.6
Peromelissa phalacra 45.3 52.8 15.1 22.3 6.9 25.8 16.8 10.2 4.3 1.0 180.3 103.7 86.7 18.1 47.9 0.9 0.8
Phormospyris stabilis capoi 0 0 0 0 0 0 5.2 8.2 0 0 0 0 8.7 18.1 0 0.19 0.81
Phormospyris stabilis scaphipes 0 0 0 0 0 0 0 0 0 0 0 0 8.7 2.6 20.5 0 0
Pseudocubus obeliscus 0 0 3.0 2.8 0 0 0 6.1 0 0 56.9 29.6 4.3 7.8 27.4 0.2 3.2
Pterocanium auritum 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Pteroscenium pinnatum 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Phormostichoartus corbula 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Plectacantha sp. 0 0 0 0 1.2 2.6 0 6.1 0 0 0 0 0 0 0 0 0
Saccospyris preantarctica 0 3.1 0 5.6 0 0 0 4.1 0 0 0 0 0 0 0 0 0
Stichopilium bicorne 0 0 0 2.8 2.3 5.2 0 0 0.9 0.3 0 7.4 4.3 10.4 6.8 0.6 0
Semantis gracilis 90.5 9.3 21.1 19.5 3.5 10.3 5.2 0 0.9 0 56.9 22.2 13.0 18.1 13.7 0.9 2.4
Spirocyrtis subscalaris 0 0 0 0 2.3 0 4.2 0 0 0 0 7.4 0 0 0 0.2 0.8
Spirocyrtis scalaris 724.1 264.0 253.4 175.5 19.6 113.7 5.2 22.5 4.3 0.5 28.5 51.9 56.4 10.4 47.9 0.4 1.6
Spirocyrtis sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Stichopilidium kruegeri 0 0 3.0 2.8 3.5 0 1.0 0 0.9 0 9.5 0 0 0 0 0 0
Tholospyris baconiana 0 0 0 0 0 0 0 0 0.9 0 0 0 0 0 0 0 0
Theophormis callipilium 11.3 0 0 0 0 0 0 0 0 0 0 0 4.3 0 0 0 0
Theocorys veneris 22.6 9.3 12.1 19.5 10.4 0 1.0 4.1 0.9 0.3 0 37.0 4.3 0 6.8 0.2 0
Tholospyris sp. Group 56.6 3.1 6.0 5.6 4.6 0 1.0 0 3.4 0.5 94.9 111.1 21.7 0 0 0.9 7.3
Table 3 (continued)
Station No. 2 3
Spumellaria #1 #2 #3 #4 #5 #6 #7 #8 #9 #10 #11 #12 #13 #14 #15 #16 #17
H.
Ya
ma
shita
eta
l./
Deep
-Sea
Resea
rchII
49
(2
00
2)
28
23
–2
86
22852
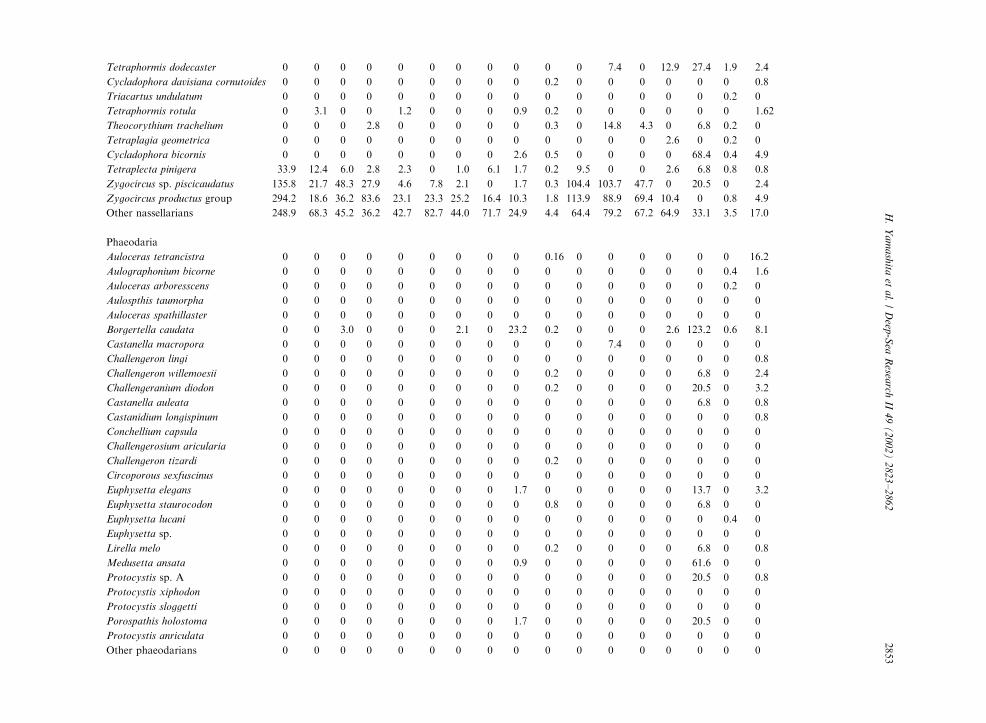
Tetraphormis dodecaster 0 0 0 0 0 0 0 0 0 0 0 7.4 0 12.9 27.4 1.9 2.4
Cycladophora davisiana cornutoides 0 0 0 0 0 0 0 0 0 0.2 0 0 0 0 0 0 0.8
Triacartus undulatum 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0.2 0
Tetraphormis rotula 0 3.1 0 0 1.2 0 0 0 0.9 0.2 0 0 0 0 0 0 1.62
Theocorythium trachelium 0 0 0 2.8 0 0 0 0 0 0.3 0 14.8 4.3 0 6.8 0.2 0
Tetraplagia geometrica 0 0 0 0 0 0 0 0 0 0 0 0 0 2.6 0 0.2 0
Cycladophora bicornis 0 0 0 0 0 0 0 0 2.6 0.5 0 0 0 0 68.4 0.4 4.9
Tetraplecta pinigera 33.9 12.4 6.0 2.8 2.3 0 1.0 6.1 1.7 0.2 9.5 0 0 2.6 6.8 0.8 0.8
Zygocircus sp. piscicaudatus 135.8 21.7 48.3 27.9 4.6 7.8 2.1 0 1.7 0.3 104.4 103.7 47.7 0 20.5 0 2.4
Zygocircus productus group 294.2 18.6 36.2 83.6 23.1 23.3 25.2 16.4 10.3 1.8 113.9 88.9 69.4 10.4 0 0.8 4.9
Other nassellarians 248.9 68.3 45.2 36.2 42.7 82.7 44.0 71.7 24.9 4.4 64.4 79.2 67.2 64.9 33.1 3.5 17.0
Phaeodaria
Auloceras tetrancistra 0 0 0 0 0 0 0 0 0 0.16 0 0 0 0 0 0 16.2
Aulographonium bicorne 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0.4 1.6
Auloceras arboresscens 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0.2 0
Aulospthis taumorpha 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Auloceras spathillaster 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Borgertella caudata 0 0 3.0 0 0 0 2.1 0 23.2 0.2 0 0 0 2.6 123.2 0.6 8.1
Castanella macropora 0 0 0 0 0 0 0 0 0 0 0 7.4 0 0 0 0 0
Challengeron lingi 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0.8
Challengeron willemoesii 0 0 0 0 0 0 0 0 0 0.2 0 0 0 0 6.8 0 2.4
Challengeranium diodon 0 0 0 0 0 0 0 0 0 0.2 0 0 0 0 20.5 0 3.2
Castanella auleata 0 0 0 0 0 0 0 0 0 0 0 0 0 0 6.8 0 0.8
Castanidium longispinum 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0.8
Conchellium capsula 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Challengerosium aricularia 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Challengeron tizardi 0 0 0 0 0 0 0 0 0 0.2 0 0 0 0 0 0 0
Circoporous sexfuscinus 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Euphysetta elegans 0 0 0 0 0 0 0 0 1.7 0 0 0 0 0 13.7 0 3.2
Euphysetta staurocodon 0 0 0 0 0 0 0 0 0 0.8 0 0 0 0 6.8 0 0
Euphysetta lucani 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0.4 0
Euphysetta sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Lirella melo 0 0 0 0 0 0 0 0 0 0.2 0 0 0 0 6.8 0 0.8
Medusetta ansata 0 0 0 0 0 0 0 0 0.9 0 0 0 0 0 61.6 0 0
Protocystis sp. A 0 0 0 0 0 0 0 0 0 0 0 0 0 0 20.5 0 0.8
Protocystis xiphodon 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Protocystis sloggetti 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Porospathis holostoma 0 0 0 0 0 0 0 0 1.7 0 0 0 0 0 20.5 0 0
Protocystis anriculata 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Other phaeodarians 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
H.
Ya
ma
shita
eta
l./
Deep
-Sea
Resea
rchII
49
(2
00
2)
28
23
–2
86
22853
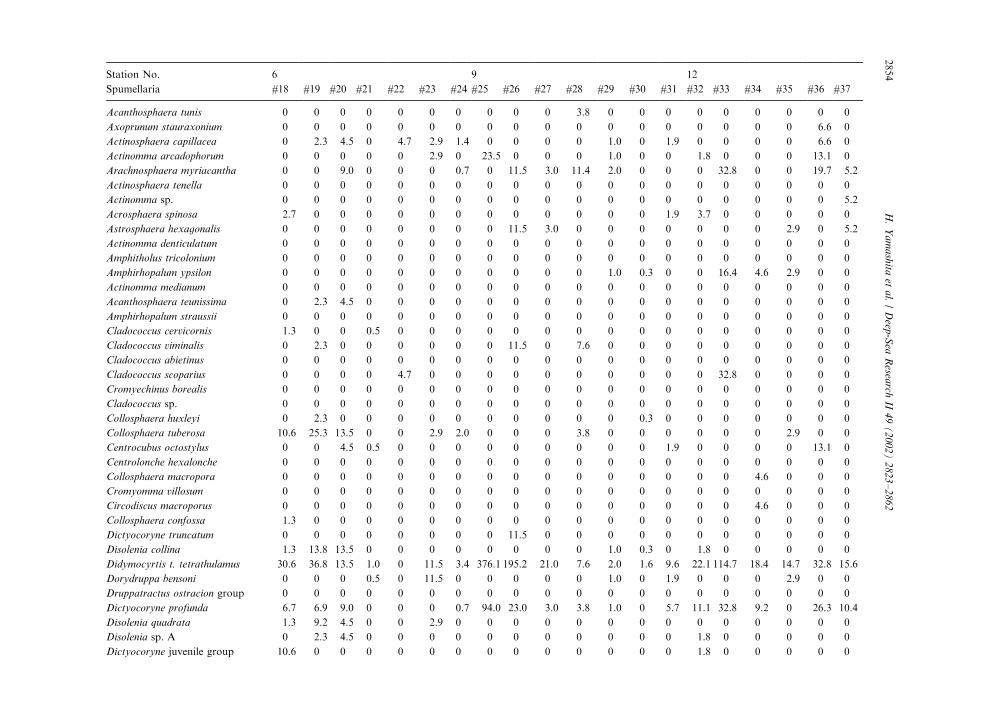
Station No. 6 9 12
Spumellaria #18 #19 #20 #21 #22 #23 #24 #25 #26 #27 #28 #29 #30 #31 #32 #33 #34 #35 #36 #37
Acanthosphaera tunis 0 0 0 0 0 0 0 0 0 0 3.8 0 0 0 0 0 0 0 0 0
Axoprunum stauraxonium 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 6.6 0
Actinosphaera capillacea 0 2.3 4.5 0 4.7 2.9 1.4 0 0 0 0 1.0 0 1.9 0 0 0 0 6.6 0
Actinomma arcadophorum 0 0 0 0 0 2.9 0 23.5 0 0 0 1.0 0 0 1.8 0 0 0 13.1 0
Arachnosphaera myriacantha 0 0 9.0 0 0 0 0.7 0 11.5 3.0 11.4 2.0 0 0 0 32.8 0 0 19.7 5.2
Actinosphaera tenella 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Actinomma sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 5.2
Acrosphaera spinosa 2.7 0 0 0 0 0 0 0 0 0 0 0 0 1.9 3.7 0 0 0 0 0
Astrosphaera hexagonalis 0 0 0 0 0 0 0 0 11.5 3.0 0 0 0 0 0 0 0 2.9 0 5.2
Actinomma denticulatum 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Amphitholus tricolonium 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Amphirhopalum ypsilon 0 0 0 0 0 0 0 0 0 0 0 1.0 0.3 0 0 16.4 4.6 2.9 0 0
Actinomma medianum 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Acanthosphaera teunissima 0 2.3 4.5 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Amphirhopalum straussii 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Cladococcus cervicornis 1.3 0 0 0.5 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Cladococcus viminalis 0 2.3 0 0 0 0 0 0 11.5 0 7.6 0 0 0 0 0 0 0 0 0
Cladococcus abietinus 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Cladococcus scoparius 0 0 0 0 4.7 0 0 0 0 0 0 0 0 0 0 32.8 0 0 0 0
Cromyechinus borealis 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Cladococcus sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Collosphaera huxleyi 0 2.3 0 0 0 0 0 0 0 0 0 0 0.3 0 0 0 0 0 0 0
Collosphaera tuberosa 10.6 25.3 13.5 0 0 2.9 2.0 0 0 0 3.8 0 0 0 0 0 0 2.9 0 0
Centrocubus octostylus 0 0 4.5 0.5 0 0 0 0 0 0 0 0 0 1.9 0 0 0 0 13.1 0
Centrolonche hexalonche 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Collosphaera macropora 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 4.6 0 0 0
Cromyomma villosum 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Circodiscus macroporus 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 4.6 0 0 0
Collosphaera confossa 1.3 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Dictyocoryne truncatum 0 0 0 0 0 0 0 0 11.5 0 0 0 0 0 0 0 0 0 0 0
Disolenia collina 1.3 13.8 13.5 0 0 0 0 0 0 0 0 1.0 0.3 0 1.8 0 0 0 0 0
Didymocyrtis t. tetrathulamus 30.6 36.8 13.5 1.0 0 11.5 3.4 376.1 195.2 21.0 7.6 2.0 1.6 9.6 22.1 114.7 18.4 14.7 32.8 15.6
Dorydruppa bensoni 0 0 0 0.5 0 11.5 0 0 0 0 0 1.0 0 1.9 0 0 0 2.9 0 0
Druppatractus ostracion group 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Dictyocoryne profunda 6.7 6.9 9.0 0 0 0 0.7 94.0 23.0 3.0 3.8 1.0 0 5.7 11.1 32.8 9.2 0 26.3 10.4
Disolenia quadrata 1.3 9.2 4.5 0 0 2.9 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Disolenia sp. A 0 2.3 4.5 0 0 0 0 0 0 0 0 0 0 0 1.8 0 0 0 0 0
Dictyocoryne juvenile group 10.6 0 0 0 0 0 0 0 0 0 0 0 0 0 1.8 0 0 0 0 0
H.
Ya
ma
shita
eta
l./
Deep
-Sea
Resea
rchII
49
(2
00
2)
28
23
–2
86
22854
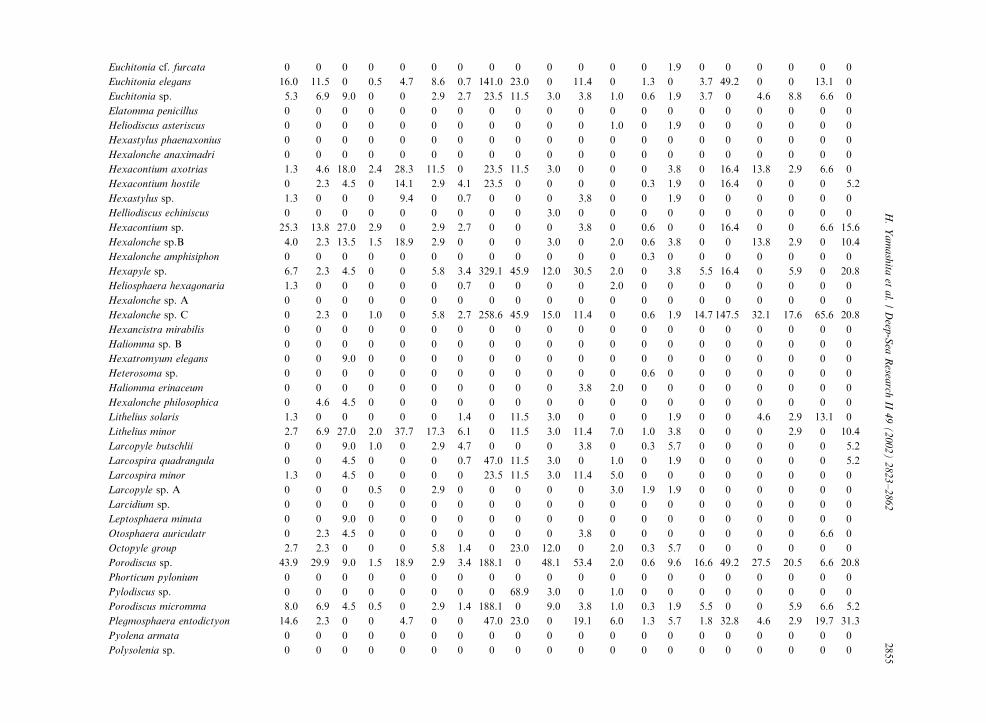
Euchitonia cf. furcata 0 0 0 0 0 0 0 0 0 0 0 0 0 1.9 0 0 0 0 0 0
Euchitonia elegans 16.0 11.5 0 0.5 4.7 8.6 0.7 141.0 23.0 0 11.4 0 1.3 0 3.7 49.2 0 0 13.1 0
Euchitonia sp. 5.3 6.9 9.0 0 0 2.9 2.7 23.5 11.5 3.0 3.8 1.0 0.6 1.9 3.7 0 4.6 8.8 6.6 0
Elatomma penicillus 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Heliodiscus asteriscus 0 0 0 0 0 0 0 0 0 0 0 1.0 0 1.9 0 0 0 0 0 0
Hexastylus phaenaxonius 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Hexalonche anaximadri 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Hexacontium axotrias 1.3 4.6 18.0 2.4 28.3 11.5 0 23.5 11.5 3.0 0 0 0 3.8 0 16.4 13.8 2.9 6.6 0
Hexacontium hostile 0 2.3 4.5 0 14.1 2.9 4.1 23.5 0 0 0 0 0.3 1.9 0 16.4 0 0 0 5.2
Hexastylus sp. 1.3 0 0 0 9.4 0 0.7 0 0 0 3.8 0 0 1.9 0 0 0 0 0 0
Helliodiscus echiniscus 0 0 0 0 0 0 0 0 0 3.0 0 0 0 0 0 0 0 0 0 0
Hexacontium sp. 25.3 13.8 27.0 2.9 0 2.9 2.7 0 0 0 3.8 0 0.6 0 0 16.4 0 0 6.6 15.6
Hexalonche sp.B 4.0 2.3 13.5 1.5 18.9 2.9 0 0 0 3.0 0 2.0 0.6 3.8 0 0 13.8 2.9 0 10.4
Hexalonche amphisiphon 0 0 0 0 0 0 0 0 0 0 0 0 0.3 0 0 0 0 0 0 0
Hexapyle sp. 6.7 2.3 4.5 0 0 5.8 3.4 329.1 45.9 12.0 30.5 2.0 0 3.8 5.5 16.4 0 5.9 0 20.8
Heliosphaera hexagonaria 1.3 0 0 0 0 0 0.7 0 0 0 0 2.0 0 0 0 0 0 0 0 0
Hexalonche sp. A 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Hexalonche sp. C 0 2.3 0 1.0 0 5.8 2.7 258.6 45.9 15.0 11.4 0 0.6 1.9 14.7 147.5 32.1 17.6 65.6 20.8
Hexancistra mirabilis 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Haliomma sp. B 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Hexatromyum elegans 0 0 9.0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Heterosoma sp. 0 0 0 0 0 0 0 0 0 0 0 0 0.6 0 0 0 0 0 0 0
Haliomma erinaceum 0 0 0 0 0 0 0 0 0 0 3.8 2.0 0 0 0 0 0 0 0 0
Hexalonche philosophica 0 4.6 4.5 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Lithelius solaris 1.3 0 0 0 0 0 1.4 0 11.5 3.0 0 0 0 1.9 0 0 4.6 2.9 13.1 0
Lithelius minor 2.7 6.9 27.0 2.0 37.7 17.3 6.1 0 11.5 3.0 11.4 7.0 1.0 3.8 0 0 0 2.9 0 10.4
Larcopyle butschlii 0 0 9.0 1.0 0 2.9 4.7 0 0 0 3.8 0 0.3 5.7 0 0 0 0 0 5.2
Larcospira quadrangula 0 0 4.5 0 0 0 0.7 47.0 11.5 3.0 0 1.0 0 1.9 0 0 0 0 0 5.2
Larcospira minor 1.3 0 4.5 0 0 0 0 23.5 11.5 3.0 11.4 5.0 0 0 0 0 0 0 0 0
Larcopyle sp. A 0 0 0 0.5 0 2.9 0 0 0 0 0 3.0 1.9 1.9 0 0 0 0 0 0
Larcidium sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Leptosphaera minuta 0 0 9.0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Otosphaera auriculatr 0 2.3 4.5 0 0 0 0 0 0 0 3.8 0 0 0 0 0 0 0 6.6 0
Octopyle group 2.7 2.3 0 0 0 5.8 1.4 0 23.0 12.0 0 2.0 0.3 5.7 0 0 0 0 0 0
Porodiscus sp. 43.9 29.9 9.0 1.5 18.9 2.9 3.4 188.1 0 48.1 53.4 2.0 0.6 9.6 16.6 49.2 27.5 20.5 6.6 20.8
Phorticum pylonium 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Pylodiscus sp. 0 0 0 0 0 0 0 0 68.9 3.0 0 1.0 0 0 0 0 0 0 0 0
Porodiscus micromma 8.0 6.9 4.5 0.5 0 2.9 1.4 188.1 0 9.0 3.8 1.0 0.3 1.9 5.5 0 0 5.9 6.6 5.2
Plegmosphaera entodictyon 14.6 2.3 0 0 4.7 0 0 47.0 23.0 0 19.1 6.0 1.3 5.7 1.8 32.8 4.6 2.9 19.7 31.3
Pyolena armata 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Polysolenia sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
H.
Ya
ma
shita
eta
l./
Deep
-Sea
Resea
rchII
49
(2
00
2)
28
23
–2
86
22855
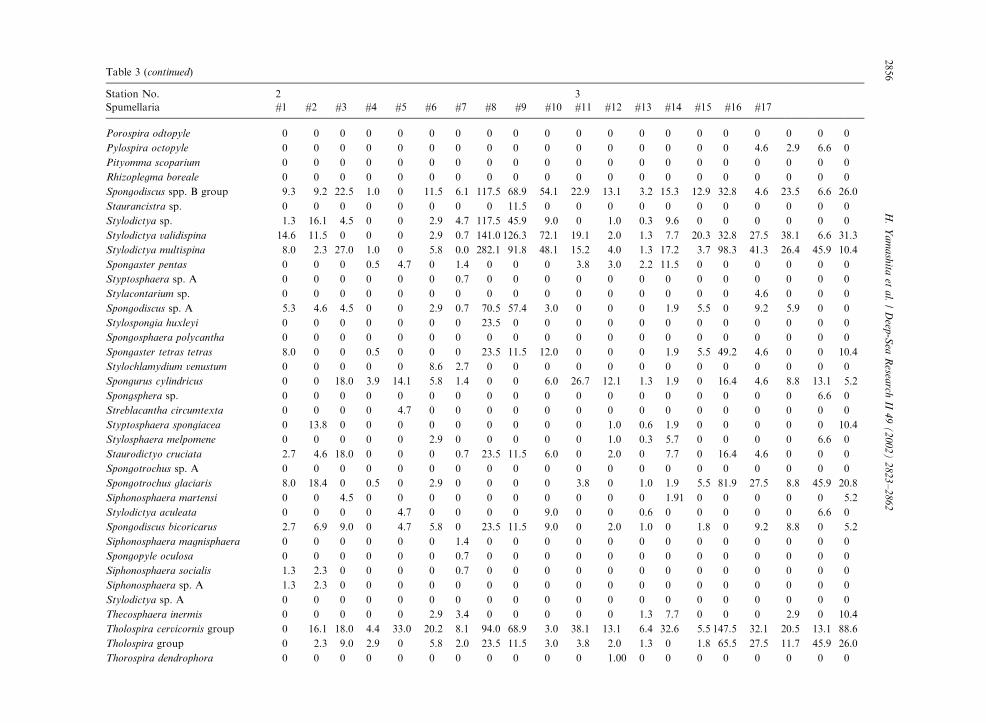
Porospira odtopyle 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Pylospira octopyle 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 4.6 2.9 6.6 0
Pityomma scoparium 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Rhizoplegma boreale 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Spongodiscus spp. B group 9.3 9.2 22.5 1.0 0 11.5 6.1 117.5 68.9 54.1 22.9 13.1 3.2 15.3 12.9 32.8 4.6 23.5 6.6 26.0
Staurancistra sp. 0 0 0 0 0 0 0 0 11.5 0 0 0 0 0 0 0 0 0 0 0
Stylodictya sp. 1.3 16.1 4.5 0 0 2.9 4.7 117.5 45.9 9.0 0 1.0 0.3 9.6 0 0 0 0 0 0
Stylodictya validispina 14.6 11.5 0 0 0 2.9 0.7 141.0 126.3 72.1 19.1 2.0 1.3 7.7 20.3 32.8 27.5 38.1 6.6 31.3
Stylodictya multispina 8.0 2.3 27.0 1.0 0 5.8 0.0 282.1 91.8 48.1 15.2 4.0 1.3 17.2 3.7 98.3 41.3 26.4 45.9 10.4
Spongaster pentas 0 0 0 0.5 4.7 0 1.4 0 0 0 3.8 3.0 2.2 11.5 0 0 0 0 0 0
Styptosphaera sp. A 0 0 0 0 0 0 0.7 0 0 0 0 0 0 0 0 0 0 0 0 0
Stylacontarium sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 4.6 0 0 0
Spongodiscus sp. A 5.3 4.6 4.5 0 0 2.9 0.7 70.5 57.4 3.0 0 0 0 1.9 5.5 0 9.2 5.9 0 0
Stylospongia huxleyi 0 0 0 0 0 0 0 23.5 0 0 0 0 0 0 0 0 0 0 0 0
Spongosphaera polycantha 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Spongaster tetras tetras 8.0 0 0 0.5 0 0 0 23.5 11.5 12.0 0 0 0 1.9 5.5 49.2 4.6 0 0 10.4
Stylochlamydium venustum 0 0 0 0 0 8.6 2.7 0 0 0 0 0 0 0 0 0 0 0 0 0
Spongurus cylindricus 0 0 18.0 3.9 14.1 5.8 1.4 0 0 6.0 26.7 12.1 1.3 1.9 0 16.4 4.6 8.8 13.1 5.2
Spongsphera sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 6.6 0
Streblacantha circumtexta 0 0 0 0 4.7 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Styptosphaera spongiacea 0 13.8 0 0 0 0 0 0 0 0 0 1.0 0.6 1.9 0 0 0 0 0 10.4
Stylosphaera melpomene 0 0 0 0 0 2.9 0 0 0 0 0 1.0 0.3 5.7 0 0 0 0 6.6 0
Staurodictyo cruciata 2.7 4.6 18.0 0 0 0 0.7 23.5 11.5 6.0 0 2.0 0 7.7 0 16.4 4.6 0 0 0
Spongotrochus sp. A 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Spongotrochus glaciaris 8.0 18.4 0 0.5 0 2.9 0 0 0 0 3.8 0 1.0 1.9 5.5 81.9 27.5 8.8 45.9 20.8
Siphonosphaera martensi 0 0 4.5 0 0 0 0 0 0 0 0 0 0 1.91 0 0 0 0 0 5.2
Stylodictya aculeata 0 0 0 0 4.7 0 0 0 0 9.0 0 0 0.6 0 0 0 0 0 6.6 0
Spongodiscus bicoricarus 2.7 6.9 9.0 0 4.7 5.8 0 23.5 11.5 9.0 0 2.0 1.0 0 1.8 0 9.2 8.8 0 5.2
Siphonosphaera magnisphaera 0 0 0 0 0 0 1.4 0 0 0 0 0 0 0 0 0 0 0 0 0
Spongopyle oculosa 0 0 0 0 0 0 0.7 0 0 0 0 0 0 0 0 0 0 0 0 0
Siphonosphaera socialis 1.3 2.3 0 0 0 0 0.7 0 0 0 0 0 0 0 0 0 0 0 0 0
Siphonosphaera sp. A 1.3 2.3 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Stylodictya sp. A 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Thecosphaera inermis 0 0 0 0 0 2.9 3.4 0 0 0 0 0 1.3 7.7 0 0 0 2.9 0 10.4
Tholospira cervicornis group 0 16.1 18.0 4.4 33.0 20.2 8.1 94.0 68.9 3.0 38.1 13.1 6.4 32.6 5.5 147.5 32.1 20.5 13.1 88.6
Tholospira group 0 2.3 9.0 2.9 0 5.8 2.0 23.5 11.5 3.0 3.8 2.0 1.3 0 1.8 65.5 27.5 11.7 45.9 26.0
Thorospira dendrophora 0 0 0 0 0 0 0 0 0 0 0 1.00 0 0 0 0 0 0 0 0
Table 3 (continued)
Station No. 2 3
Spumellaria #1 #2 #3 #4 #5 #6 #7 #8 #9 #10 #11 #12 #13 #14 #15 #16 #17H
.Y
am
ash
itaet
al.
/D
eep-S
eaR
esearch
II4
9(
20
02
)2
82
3–
28
62
2856
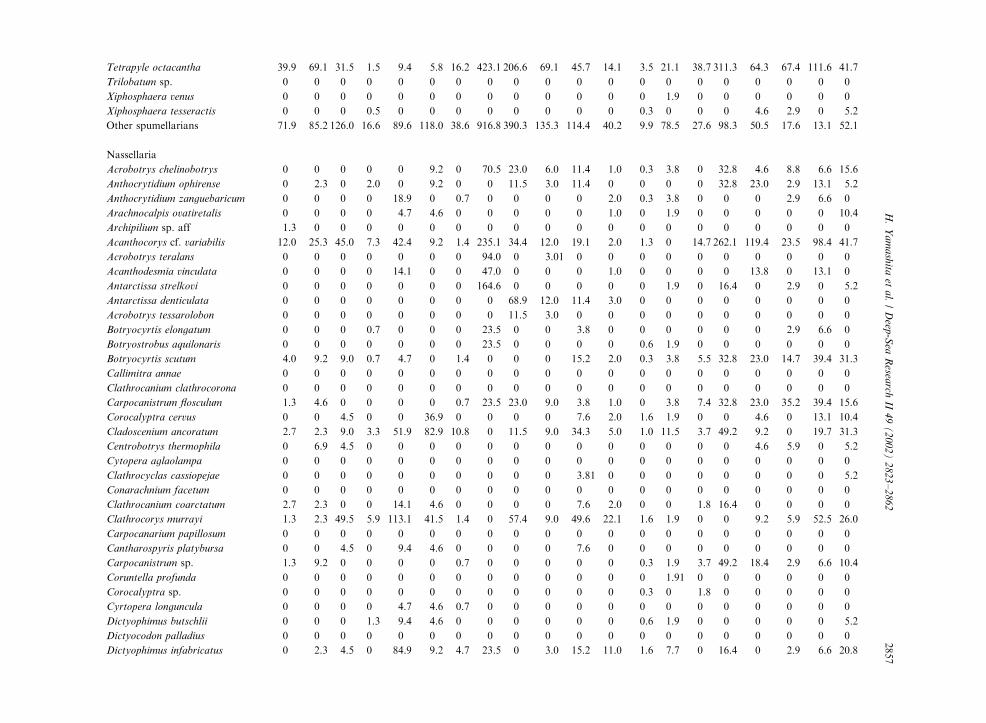
Tetrapyle octacantha 39.9 69.1 31.5 1.5 9.4 5.8 16.2 423.1 206.6 69.1 45.7 14.1 3.5 21.1 38.7 311.3 64.3 67.4 111.6 41.7
Trilobatum sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Xiphosphaera venus 0 0 0 0 0 0 0 0 0 0 0 0 0 1.9 0 0 0 0 0 0
Xiphosphaera tesseractis 0 0 0 0.5 0 0 0 0 0 0 0 0 0.3 0 0 0 4.6 2.9 0 5.2
Other spumellarians 71.9 85.2 126.0 16.6 89.6 118.0 38.6 916.8 390.3 135.3 114.4 40.2 9.9 78.5 27.6 98.3 50.5 17.6 13.1 52.1
Nassellaria
Acrobotrys chelinobotrys 0 0 0 0 0 9.2 0 70.5 23.0 6.0 11.4 1.0 0.3 3.8 0 32.8 4.6 8.8 6.6 15.6
Anthocrytidium ophirense 0 2.3 0 2.0 0 9.2 0 0 11.5 3.0 11.4 0 0 0 0 32.8 23.0 2.9 13.1 5.2
Anthocrytidium zanguebaricum 0 0 0 0 18.9 0 0.7 0 0 0 0 2.0 0.3 3.8 0 0 0 2.9 6.6 0
Arachnocalpis ovatiretalis 0 0 0 0 4.7 4.6 0 0 0 0 0 1.0 0 1.9 0 0 0 0 0 10.4
Archipilium sp. aff 1.3 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Acanthocorys cf. variabilis 12.0 25.3 45.0 7.3 42.4 9.2 1.4 235.1 34.4 12.0 19.1 2.0 1.3 0 14.7 262.1 119.4 23.5 98.4 41.7
Acrobotrys teralans 0 0 0 0 0 0 0 94.0 0 3.01 0 0 0 0 0 0 0 0 0 0
Acanthodesmia vinculata 0 0 0 0 14.1 0 0 47.0 0 0 0 1.0 0 0 0 0 13.8 0 13.1 0
Antarctissa strelkovi 0 0 0 0 0 0 0 164.6 0 0 0 0 0 1.9 0 16.4 0 2.9 0 5.2
Antarctissa denticulata 0 0 0 0 0 0 0 0 68.9 12.0 11.4 3.0 0 0 0 0 0 0 0 0
Acrobotrys tessarolobon 0 0 0 0 0 0 0 0 11.5 3.0 0 0 0 0 0 0 0 0 0 0
Botryocyrtis elongatum 0 0 0 0.7 0 0 0 23.5 0 0 3.8 0 0 0 0 0 0 2.9 6.6 0
Botryostrobus aquilonaris 0 0 0 0 0 0 0 23.5 0 0 0 0 0.6 1.9 0 0 0 0 0 0
Botryocyrtis scutum 4.0 9.2 9.0 0.7 4.7 0 1.4 0 0 0 15.2 2.0 0.3 3.8 5.5 32.8 23.0 14.7 39.4 31.3
Callimitra annae 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Clathrocanium clathrocorona 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Carpocanistrum flosculum 1.3 4.6 0 0 0 0 0.7 23.5 23.0 9.0 3.8 1.0 0 3.8 7.4 32.8 23.0 35.2 39.4 15.6
Corocalyptra cervus 0 0 4.5 0 0 36.9 0 0 0 0 7.6 2.0 1.6 1.9 0 0 4.6 0 13.1 10.4
Cladoscenium ancoratum 2.7 2.3 9.0 3.3 51.9 82.9 10.8 0 11.5 9.0 34.3 5.0 1.0 11.5 3.7 49.2 9.2 0 19.7 31.3
Centrobotrys thermophila 0 6.9 4.5 0 0 0 0 0 0 0 0 0 0 0 0 0 4.6 5.9 0 5.2
Cytopera aglaolampa 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Clathrocyclas cassiopejae 0 0 0 0 0 0 0 0 0 0 3.81 0 0 0 0 0 0 0 0 5.2
Conarachnium facetum 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Clathrocanium coarctatum 2.7 2.3 0 0 14.1 4.6 0 0 0 0 7.6 2.0 0 0 1.8 16.4 0 0 0 0
Clathrocorys murrayi 1.3 2.3 49.5 5.9 113.1 41.5 1.4 0 57.4 9.0 49.6 22.1 1.6 1.9 0 0 9.2 5.9 52.5 26.0
Carpocanarium papillosum 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Cantharospyris platybursa 0 0 4.5 0 9.4 4.6 0 0 0 0 7.6 0 0 0 0 0 0 0 0 0
Carpocanistrum sp. 1.3 9.2 0 0 0 0 0.7 0 0 0 0 0 0.3 1.9 3.7 49.2 18.4 2.9 6.6 10.4
Coruntella profunda 0 0 0 0 0 0 0 0 0 0 0 0 0 1.91 0 0 0 0 0 0
Corocalyptra sp. 0 0 0 0 0 0 0 0 0 0 0 0 0.3 0 1.8 0 0 0 0 0
Cyrtopera longuncula 0 0 0 0 4.7 4.6 0.7 0 0 0 0 0 0 0 0 0 0 0 0 0
Dictyophimus butschlii 0 0 0 1.3 9.4 4.6 0 0 0 0 0 0 0.6 1.9 0 0 0 0 0 5.2
Dictyocodon palladius 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Dictyophimus infabricatus 0 2.3 4.5 0 84.9 9.2 4.7 23.5 0 3.0 15.2 11.0 1.6 7.7 0 16.4 0 2.9 6.6 20.8
H.
Ya
ma
shita
eta
l./
Deep
-Sea
Resea
rchII
49
(2
00
2)
28
23
–2
86
22857
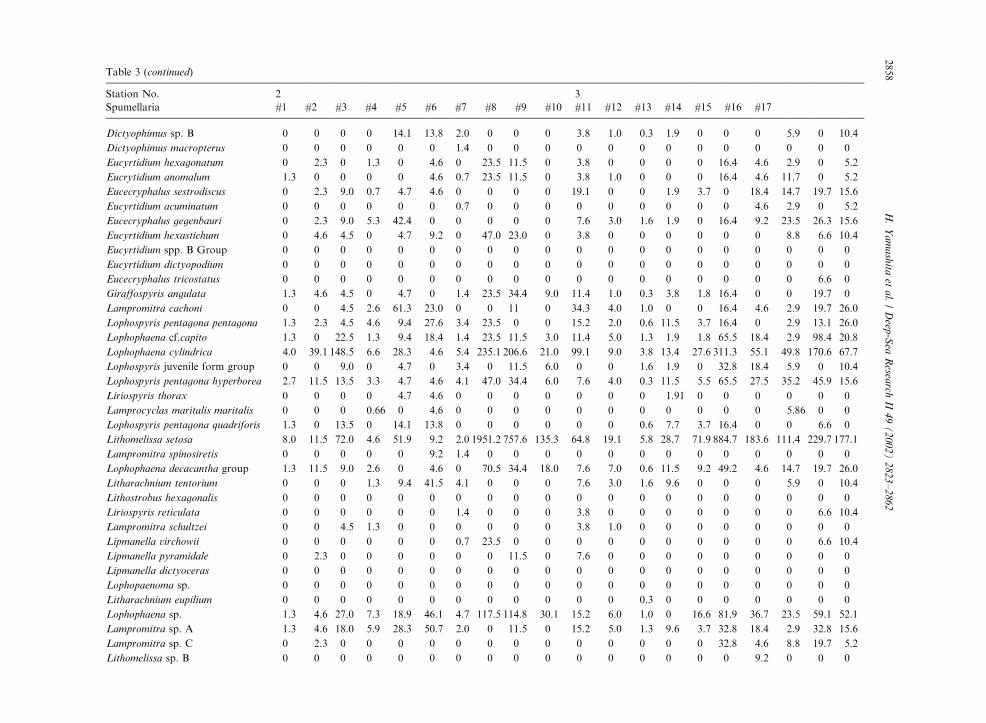
Dictyophimus sp. B 0 0 0 0 14.1 13.8 2.0 0 0 0 3.8 1.0 0.3 1.9 0 0 0 5.9 0 10.4
Dictyophimus macropterus 0 0 0 0 0 0 1.4 0 0 0 0 0 0 0 0 0 0 0 0 0
Eucyrtidium hexagonatum 0 2.3 0 1.3 0 4.6 0 23.5 11.5 0 3.8 0 0 0 0 16.4 4.6 2.9 0 5.2
Eucrytidium anomalum 1.3 0 0 0 0 4.6 0.7 23.5 11.5 0 3.8 1.0 0 0 0 16.4 4.6 11.7 0 5.2
Eucecryphalus sestrodiscus 0 2.3 9.0 0.7 4.7 4.6 0 0 0 0 19.1 0 0 1.9 3.7 0 18.4 14.7 19.7 15.6
Eucyrtidium acuminatum 0 0 0 0 0 0 0.7 0 0 0 0 0 0 0 0 0 4.6 2.9 0 5.2
Eucecryphalus gegenbauri 0 2.3 9.0 5.3 42.4 0 0 0 0 0 7.6 3.0 1.6 1.9 0 16.4 9.2 23.5 26.3 15.6
Eucyrtidium hexastichum 0 4.6 4.5 0 4.7 9.2 0 47.0 23.0 0 3.8 0 0 0 0 0 0 8.8 6.6 10.4
Eucyrtidium spp. B Group 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Eucyrtidium dictyopodium 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Eucecryphalus tricostatus 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 6.6 0
Giraffospyris angulata 1.3 4.6 4.5 0 4.7 0 1.4 23.5 34.4 9.0 11.4 1.0 0.3 3.8 1.8 16.4 0 0 19.7 0
Lampromitra cachoni 0 0 4.5 2.6 61.3 23.0 0 0 11 0 34.3 4.0 1.0 0 0 16.4 4.6 2.9 19.7 26.0
Lophospyris pentagona pentagona 1.3 2.3 4.5 4.6 9.4 27.6 3.4 23.5 0 0 15.2 2.0 0.6 11.5 3.7 16.4 0 2.9 13.1 26.0
Lophophaena cf.capito 1.3 0 22.5 1.3 9.4 18.4 1.4 23.5 11.5 3.0 11.4 5.0 1.3 1.9 1.8 65.5 18.4 2.9 98.4 20.8
Lophophaena cylindrica 4.0 39.1 148.5 6.6 28.3 4.6 5.4 235.1 206.6 21.0 99.1 9.0 3.8 13.4 27.6 311.3 55.1 49.8 170.6 67.7
Lophospyris juvenile form group 0 0 9.0 0 4.7 0 3.4 0 11.5 6.0 0 0 1.6 1.9 0 32.8 18.4 5.9 0 10.4
Lophospyris pentagona hyperborea 2.7 11.5 13.5 3.3 4.7 4.6 4.1 47.0 34.4 6.0 7.6 4.0 0.3 11.5 5.5 65.5 27.5 35.2 45.9 15.6
Liriospyris thorax 0 0 0 0 4.7 4.6 0 0 0 0 0 0 0 1.91 0 0 0 0 0 0
Lamprocyclas maritalis maritalis 0 0 0 0.66 0 4.6 0 0 0 0 0 0 0 0 0 0 0 5.86 0 0
Lophospyris pentagona quadriforis 1.3 0 13.5 0 14.1 13.8 0 0 0 0 0 0 0.6 7.7 3.7 16.4 0 0 6.6 0
Lithomelissa setosa 8.0 11.5 72.0 4.6 51.9 9.2 2.0 1951.2 757.6 135.3 64.8 19.1 5.8 28.7 71.9 884.7 183.6 111.4 229.7 177.1
Lampromitra spinosiretis 0 0 0 0 0 9.2 1.4 0 0 0 0 0 0 0 0 0 0 0 0 0
Lophophaena decacantha group 1.3 11.5 9.0 2.6 0 4.6 0 70.5 34.4 18.0 7.6 7.0 0.6 11.5 9.2 49.2 4.6 14.7 19.7 26.0
Litharachnium tentorium 0 0 0 1.3 9.4 41.5 4.1 0 0 0 7.6 3.0 1.6 9.6 0 0 0 5.9 0 10.4
Lithostrobus hexagonalis 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Liriospyris reticulata 0 0 0 0 0 0 1.4 0 0 0 3.8 0 0 0 0 0 0 0 6.6 10.4
Lampromitra schultzei 0 0 4.5 1.3 0 0 0 0 0 0 3.8 1.0 0 0 0 0 0 0 0 0
Lipmanella virchowii 0 0 0 0 0 0 0.7 23.5 0 0 0 0 0 0 0 0 0 0 6.6 10.4
Lipmanella pyramidale 0 2.3 0 0 0 0 0 0 11.5 0 7.6 0 0 0 0 0 0 0 0 0
Lipmanella dictyoceras 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Lophopaenoma sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Litharachnium eupilium 0 0 0 0 0 0 0 0 0 0 0 0 0.3 0 0 0 0 0 0 0
Lophophaena sp. 1.3 4.6 27.0 7.3 18.9 46.1 4.7 117.5 114.8 30.1 15.2 6.0 1.0 0 16.6 81.9 36.7 23.5 59.1 52.1
Lampromitra sp. A 1.3 4.6 18.0 5.9 28.3 50.7 2.0 0 11.5 0 15.2 5.0 1.3 9.6 3.7 32.8 18.4 2.9 32.8 15.6
Lampromitra sp. C 0 2.3 0 0 0 0 0 0 0 0 0 0 0 0 0 32.8 4.6 8.8 19.7 5.2
Lithomelissa sp. B 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 9.2 0 0 0
Table 3 (continued)
Station No. 2 3
Spumellaria #1 #2 #3 #4 #5 #6 #7 #8 #9 #10 #11 #12 #13 #14 #15 #16 #17H
.Y
am
ash
itaet
al.
/D
eep-S
eaR
esearch
II4
9(
20
02
)2
82
3–
28
62
2858
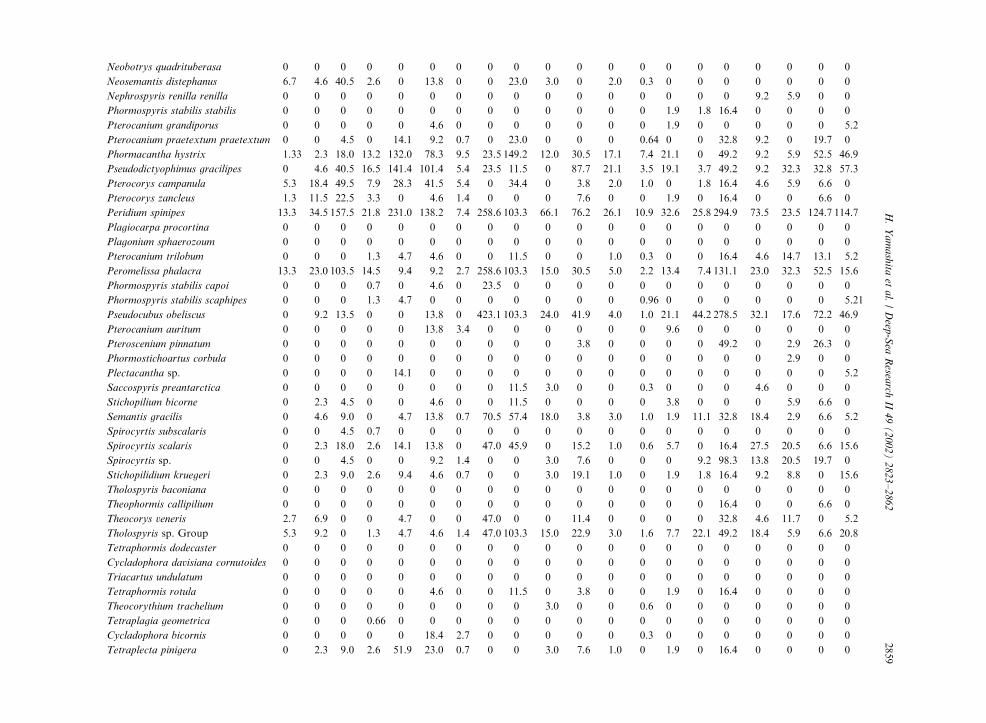
Neobotrys quadrituberasa 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Neosemantis distephanus 6.7 4.6 40.5 2.6 0 13.8 0 0 23.0 3.0 0 2.0 0.3 0 0 0 0 0 0 0
Nephrospyris renilla renilla 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 9.2 5.9 0 0
Phormospyris stabilis stabilis 0 0 0 0 0 0 0 0 0 0 0 0 0 1.9 1.8 16.4 0 0 0 0
Pterocanium grandiporus 0 0 0 0 0 4.6 0 0 0 0 0 0 0 1.9 0 0 0 0 0 5.2
Pterocanium praetextum praetextum 0 0 4.5 0 14.1 9.2 0.7 0 23.0 0 0 0 0.64 0 0 32.8 9.2 0 19.7 0
Phormacantha hystrix 1.33 2.3 18.0 13.2 132.0 78.3 9.5 23.5 149.2 12.0 30.5 17.1 7.4 21.1 0 49.2 9.2 5.9 52.5 46.9
Pseudodictyophimus gracilipes 0 4.6 40.5 16.5 141.4 101.4 5.4 23.5 11.5 0 87.7 21.1 3.5 19.1 3.7 49.2 9.2 32.3 32.8 57.3
Pterocorys campanula 5.3 18.4 49.5 7.9 28.3 41.5 5.4 0 34.4 0 3.8 2.0 1.0 0 1.8 16.4 4.6 5.9 6.6 0
Pterocorys zancleus 1.3 11.5 22.5 3.3 0 4.6 1.4 0 0 0 7.6 0 0 1.9 0 16.4 0 0 6.6 0
Peridium spinipes 13.3 34.5 157.5 21.8 231.0 138.2 7.4 258.6 103.3 66.1 76.2 26.1 10.9 32.6 25.8 294.9 73.5 23.5 124.7 114.7
Plagiocarpa procortina 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Plagonium sphaerozoum 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Pterocanium trilobum 0 0 0 1.3 4.7 4.6 0 0 11.5 0 0 1.0 0.3 0 0 16.4 4.6 14.7 13.1 5.2
Peromelissa phalacra 13.3 23.0 103.5 14.5 9.4 9.2 2.7 258.6 103.3 15.0 30.5 5.0 2.2 13.4 7.4 131.1 23.0 32.3 52.5 15.6
Phormospyris stabilis capoi 0 0 0 0.7 0 4.6 0 23.5 0 0 0 0 0 0 0 0 0 0 0 0
Phormospyris stabilis scaphipes 0 0 0 1.3 4.7 0 0 0 0 0 0 0 0.96 0 0 0 0 0 0 5.21
Pseudocubus obeliscus 0 9.2 13.5 0 0 13.8 0 423.1 103.3 24.0 41.9 4.0 1.0 21.1 44.2 278.5 32.1 17.6 72.2 46.9
Pterocanium auritum 0 0 0 0 0 13.8 3.4 0 0 0 0 0 0 9.6 0 0 0 0 0 0
Pteroscenium pinnatum 0 0 0 0 0 0 0 0 0 0 3.8 0 0 0 0 49.2 0 2.9 26.3 0
Phormostichoartus corbula 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 2.9 0 0
Plectacantha sp. 0 0 0 0 14.1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 5.2
Saccospyris preantarctica 0 0 0 0 0 0 0 0 11.5 3.0 0 0 0.3 0 0 0 4.6 0 0 0
Stichopilium bicorne 0 2.3 4.5 0 0 4.6 0 0 11.5 0 0 0 0 3.8 0 0 0 5.9 6.6 0
Semantis gracilis 0 4.6 9.0 0 4.7 13.8 0.7 70.5 57.4 18.0 3.8 3.0 1.0 1.9 11.1 32.8 18.4 2.9 6.6 5.2
Spirocyrtis subscalaris 0 0 4.5 0.7 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Spirocyrtis scalaris 0 2.3 18.0 2.6 14.1 13.8 0 47.0 45.9 0 15.2 1.0 0.6 5.7 0 16.4 27.5 20.5 6.6 15.6
Spirocyrtis sp. 0 0 4.5 0 0 9.2 1.4 0 0 3.0 7.6 0 0 0 9.2 98.3 13.8 20.5 19.7 0
Stichopilidium kruegeri 0 2.3 9.0 2.6 9.4 4.6 0.7 0 0 3.0 19.1 1.0 0 1.9 1.8 16.4 9.2 8.8 0 15.6
Tholospyris baconiana 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Theophormis callipilium 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 16.4 0 0 6.6 0
Theocorys veneris 2.7 6.9 0 0 4.7 0 0 47.0 0 0 11.4 0 0 0 0 32.8 4.6 11.7 0 5.2
Tholospyris sp. Group 5.3 9.2 0 1.3 4.7 4.6 1.4 47.0 103.3 15.0 22.9 3.0 1.6 7.7 22.1 49.2 18.4 5.9 6.6 20.8
Tetraphormis dodecaster 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Cycladophora davisiana cornutoides 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Triacartus undulatum 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Tetraphormis rotula 0 0 0 0 0 4.6 0 0 11.5 0 3.8 0 0 1.9 0 16.4 0 0 0 0
Theocorythium trachelium 0 0 0 0 0 0 0 0 0 3.0 0 0 0.6 0 0 0 0 0 0 0
Tetraplagia geometrica 0 0 0 0.66 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Cycladophora bicornis 0 0 0 0 0 18.4 2.7 0 0 0 0 0 0.3 0 0 0 0 0 0 0
Tetraplecta pinigera 0 2.3 9.0 2.6 51.9 23.0 0.7 0 0 3.0 7.6 1.0 0 1.9 0 16.4 0 0 0 0
H.
Ya
ma
shita
eta
l./
Deep
-Sea
Resea
rchII
49
(2
00
2)
28
23
–2
86
22859
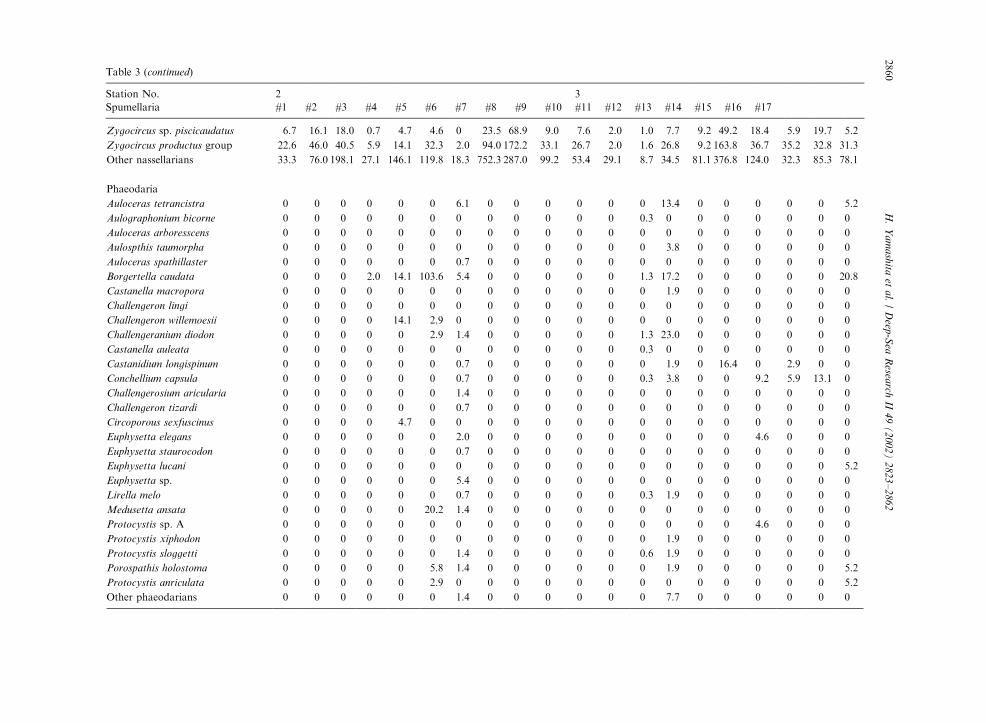
Zygocircus sp. piscicaudatus 6.7 16.1 18.0 0.7 4.7 4.6 0 23.5 68.9 9.0 7.6 2.0 1.0 7.7 9.2 49.2 18.4 5.9 19.7 5.2
Zygocircus productus group 22.6 46.0 40.5 5.9 14.1 32.3 2.0 94.0 172.2 33.1 26.7 2.0 1.6 26.8 9.2 163.8 36.7 35.2 32.8 31.3
Other nassellarians 33.3 76.0 198.1 27.1 146.1 119.8 18.3 752.3 287.0 99.2 53.4 29.1 8.7 34.5 81.1 376.8 124.0 32.3 85.3 78.1
Phaeodaria
Auloceras tetrancistra 0 0 0 0 0 0 6.1 0 0 0 0 0 0 13.4 0 0 0 0 0 5.2
Aulographonium bicorne 0 0 0 0 0 0 0 0 0 0 0 0 0.3 0 0 0 0 0 0 0
Auloceras arboresscens 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Aulospthis taumorpha 0 0 0 0 0 0 0 0 0 0 0 0 0 3.8 0 0 0 0 0 0
Auloceras spathillaster 0 0 0 0 0 0 0.7 0 0 0 0 0 0 0 0 0 0 0 0 0
Borgertella caudata 0 0 0 2.0 14.1 103.6 5.4 0 0 0 0 0 1.3 17.2 0 0 0 0 0 20.8
Castanella macropora 0 0 0 0 0 0 0 0 0 0 0 0 0 1.9 0 0 0 0 0 0
Challengeron lingi 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Challengeron willemoesii 0 0 0 0 14.1 2.9 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Challengeranium diodon 0 0 0 0 0 2.9 1.4 0 0 0 0 0 1.3 23.0 0 0 0 0 0 0
Castanella auleata 0 0 0 0 0 0 0 0 0 0 0 0 0.3 0 0 0 0 0 0 0
Castanidium longispinum 0 0 0 0 0 0 0.7 0 0 0 0 0 0 1.9 0 16.4 0 2.9 0 0
Conchellium capsula 0 0 0 0 0 0 0.7 0 0 0 0 0 0.3 3.8 0 0 9.2 5.9 13.1 0
Challengerosium aricularia 0 0 0 0 0 0 1.4 0 0 0 0 0 0 0 0 0 0 0 0 0
Challengeron tizardi 0 0 0 0 0 0 0.7 0 0 0 0 0 0 0 0 0 0 0 0 0
Circoporous sexfuscinus 0 0 0 0 4.7 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Euphysetta elegans 0 0 0 0 0 0 2.0 0 0 0 0 0 0 0 0 0 4.6 0 0 0
Euphysetta staurocodon 0 0 0 0 0 0 0.7 0 0 0 0 0 0 0 0 0 0 0 0 0
Euphysetta lucani 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 5.2
Euphysetta sp. 0 0 0 0 0 0 5.4 0 0 0 0 0 0 0 0 0 0 0 0 0
Lirella melo 0 0 0 0 0 0 0.7 0 0 0 0 0 0.3 1.9 0 0 0 0 0 0
Medusetta ansata 0 0 0 0 0 20.2 1.4 0 0 0 0 0 0 0 0 0 0 0 0 0
Protocystis sp. A 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 4.6 0 0 0
Protocystis xiphodon 0 0 0 0 0 0 0 0 0 0 0 0 0 1.9 0 0 0 0 0 0
Protocystis sloggetti 0 0 0 0 0 0 1.4 0 0 0 0 0 0.6 1.9 0 0 0 0 0 0
Porospathis holostoma 0 0 0 0 0 5.8 1.4 0 0 0 0 0 0 1.9 0 0 0 0 0 5.2
Protocystis anriculata 0 0 0 0 0 2.9 0 0 0 0 0 0 0 0 0 0 0 0 0 5.2
Other phaeodarians 0 0 0 0 0 0 1.4 0 0 0 0 0 0 7.7 0 0 0 0 0 0
Table 3 (continued)
Station No. 2 3
Spumellaria #1 #2 #3 #4 #5 #6 #7 #8 #9 #10 #11 #12 #13 #14 #15 #16 #17H
.Y
am
ash
itaet
al.
/D
eep-S
eaR
esearch
II4
9(
20
02
)2
82
3–
28
62
2860

MR98-K02 Cruise is shown with proper referencesin Table 2.
Appendix B
Radiolarian species standing stock data (#shellsm�3) obtained during R/V Mirai CruiseMR98-K02 are shown in Table 3.
References
Abelmann, A., Gowing, M.M., 1997. Spatial distribution
pattern of living polycystine radiolarian taxa-baseline study
for paleoenvironmental reconstructions in the Southern
Ocean (Atlantic sector). Marine Micropaleontology 30,
3–28.
Amaral Zettler, L.A., Anderson, O.R., Caron, D. A., 1999.
Towards a molecular phylogeny of colonial spumellarian
radiolaria. Marine Micropaleontology 36, 67–79.
Anderson, O.R., 1983. Radiolaria. Springer-Verlag, New York,
pp. 355.
Anderson, O.R., Bryan, M., Bennett, P., 1990. Experimental
and observational studies of radiolarian physiological
ecology: 4. factors determining the distribution and survival
of didymocyrtis tetrathalamus tetrathalamus with implica-
tions for paleoecological interpretations. Marine Micropa-
leontology 16, 155–167.
Barber, R.T., Chavez, F.P., 1991. Regulation of primary
productivity rate in the equatorial Pacific. Limnology and
Ocenography 36, 1803–1815.
Berger, W.H., Fischer, K., Lai, C., Wu, G., 1987. Ocean
productivity and organic carbon flux, Part I: overview and
maps of primary production and export production. Scripps
Institution of Oceanography SIO Reference, San Diego,
California, pp. 87–30.
Bj�rklund, K.R., 1976. Radiolaria from the Norwegian Sea,
Leg 38 of the deep-sea drilling project. Initial Reports of the
Deep-Sea Drilling Project 38, 1101–1168.
Boltovskoy, D., Jankilevich, S.S., 1985. Radiolarian distribu-
tion in east Equatorial Pacific plankton. Oceanologica Acta
8, 101–123.
Boltovskoy, D., Riedel, W., 1987. Polycystine Radiolaria of the
California Current region: seasonal and geographic pat-
terns. Marine Micropaleontology 12, 65–104.
Boltovskoy, D., Alder, V.A., Abelmann, A., 1993. Annual flux
of Radiolaria and other shelled planktons in the eastern
Equatorial Atlantic at 853m: seasonal variations and
polycystine species-specific responses. Deep-Sea Research I
40, 1863–1895.
Casey, R., Gust, L., Leavesley, A., Williams, D., Reynolds, R.,
Duis, T., Spaw, M., 1979. Ecological niches of radiolarians,
planktonic foraminiferans and pteropods inferred from
studies on living forms in the Gulf of Mexico and adjacent
waters. Transactions-Gulf Coast Association of Geological
Societies 24, 216–223.
Cassie, R.M., 1968. Terminology. In: Tranter, D. J. (Ed.),
Zooplankton Sampling. UNESCO, Paris, pp. 123–125.
Dworetzky, B.A., Morley, J.J., 1987. Vertical distribution of
radiolaria in the eastern equatorial Atlantic: analysis of a
multiple series of closely spaced plankton tows. Marine
Micropaleontology 12, 1–19.
Gowing, M.M., 1986. Trophic biology of phaeodarian radi-
olarians and flux of living radiolarians in the upper 2000m
of the north Pacific central gyre. Deep-Sea Research 33,
655–674.
Gowing, M.M., 1993. Seasonal radiolarian flux at the
VERTEX north Pacific time-series site. Deep-Sea Research
I 40, 517–545.
Inoue, H.Y., Ishii, M., Matsueda, H., Ahoyama, M., 1996.
Change in longitudinal distribution of the partial pressure
of CO2 (pCO2) in the central and western Equatorial
Pacific, west of 160W. Geophysical Research Letters 23,
1781–1784.
Kessler, W.S., McPhaden, M.J., 1995. The 1991–1993 El Ni *no
in the central Pacific. Deep-Sea Research II 42, 233–295.
Kling, S.A., 1979. Vertical distribution of polycystine radiolar-
ians in the central north Pacific. Marine Micropaleontology
4, 295–318.
Kling, S.A., Boltovskoy, D., 1995. Radiolarian vertical
distribution patterns across the southern California Cur-
rent. Deep-Sea Research I 42, 191–231.
Kobayashi, F., Takahashi, K., 2002. Distribution of diatoms
along the equatorial transect in the western and central
pacific during the 1999 La Ni *na conditions. Deep-Sea
Research Part II 49, 2801–2821.
Koblentz-Mishke, O.J., Volkovinsky, V.V., Kobanova, J.G.,
1970. Plankton primary production of the world ocean. In:
Wooster, W. S. (Ed.), Scientific Exploration of the South
Pacific. National Academy of Sciences, Washington, DC,
pp. 183–193.
Le Borgne, R., Barber, R.T., Delcroix, T., Inoue, H.Y.,
Mackey, D.J., Rodier, M., 2002. Pacific warm pool and
divergence: temporal and zonal variations on the equator
and their effects on the biological pump. Deep-Sea Research
II 49, 2471–2512.
Lombari, G., Boden, G., 1985. Modern radiolarian global
distributions. Cushman Foundation for Foraminiferal
Research, Special Publication 16A, Washington, DC,
pp. 125.
Longhurst, A.R., 1998. Ecological Geography of the Sea.
Academic Press, London, pp. 398.
Lukas, R., Lindstrom, E., 1991. The mixed layer of the western
Equatorial Pacific Ocean. Journal of Geophysical Research
96, 3343–3357.
McMillen, K.J., Casey, R.E., 1978. Distribution of living
polycystine radiolarians in the Gulf of Mexico and
Caribbean Sea, and comparison with the sedimentary
record. Marine Micropaleontology 3, 121–145.
H. Yamashita et al. / Deep-Sea Research II 49 (2002) 2823–2862 2861

Nigrini, C.A., Moore Jr., T.C., 1979. A guide to modern
radiolaria. Cushman Foundation for Foraminiferal Re-
search, Special Publication 16, Washington, DC, pp. 342.
Picaut, J., Ioualalen, M., Menkes, C., Delcroix, T., McPhaden,
M., 1996. Mechanism of zonal displacements of the
Pacific warm pool: implications for ENSO. Science 274,
1486–1489.
Pielou, E.C., 1966. The measurement of diversity in different
types of biological collections. Journal of Theoretical
Biology 13, 131–144.
Pisias, N.G., Murray, D.W., Roelofs, A.K., 1986.
Radiolarian and silicoflagellate response to oceanographic
changes associated with the 1983 El Ni *no. Nature 320,
259–262.
Renz, G.W., 1976. The distribution and ecology of radiolaria in
the central Pacific: plankton and surface sediments. Bulletin
of the Scripps Institute of Oceanography 22, 267.
Rodier, M., Eldin, G., Borgne, R., 2000. The western boundary
of the Equatorial Pacific upwelling: some consequences of
climatic variability on hydrological and planktonic proper-
ties. Journal of Oceanography 56, 463–471.
Shannon, C.E., Weaver, W., 1949. The Mathematical Theory
of Communication. University of Illinois Press, Urbana,
pp. 125.
Suzuki, Y., Seike, T., Kuji, T., Nishimura, T., Matumoto, K.,
1999. MR98-K02 Preliminary Cruise Report March 1999.
Japan Marine Science and Technology Center, Japan,
pp. 131–137.
Swanberg, N.R., Anderson, O.R., 1985. The nutrition of
radiolarians: trophic activity of some solitary Spumellaria.
Limnology Oceanography 30, 646–652.
Takahashi, K., 1981. Vertical flux, ecology and dissolution of
Radiolaria in tropical oceans: implications for the silica
cycle, Ph.D. Thesis, Massachusetts Institute of Technology
and the Woods Hole Oceanographic Institution, pp. 461.
Takahashi, K., 1983. Radiolaria: sinking population, standing
stock, and production rate. Marine Micropaleontology 8,
171–181.
Takahashi, K., 1987. Radiolarian flux and seasonality: climatic
and El Ni *no response in the subarctic pacific, 1982–1984.
Global Biogeochemical Cycles 1, 213–231.
Takahashi, K., 1991. Radiolaria: flux, ecology, and taxonomy
in the Pacific and Atlantic. In: Honjo, S. (Ed.), Ocean
Biocoenosis Series No. 3. Woods Hole Oceanographic
Institution Press, pp. 303.
Takahashi, K., 1997. Siliceous microplankton fluxes in the
eastern subarctic Pacific, 1982–1986. Journal of Oceano-
graphy 53, 455–466.
Takahashi, K., Honjo, S., 1981. Vertical flux of radiolaria: a
taxon-quantitative sediment trap study from the western
tropical Atlantic. Micropaleontology 27, 140–190.
Takahashi, K., Oba, T., Yamazaki, H., Ohkouchi, N., 1995.
Paleoceanographic change of the western Equatorial Pacific
from radiolarian record for 80 ka. Kaiyou-Monthly 27,
466–473 (in Japanese).
Welling, L.A., Pisias, N.G., 1998. Radiolarian fluxes, stocks,
and population residence times in surface waters of
the central Equatorial Pacific. Deep-Sea Research I 45,
639–671.
Welling, L.A., Pisias, N.G., Johnson, E.S., White, J.R., 1996.
Distribution of polycystine radiolaria and their relation to
the physical environment during the 1992 El Ni *no and
following cold event. Deep-Sea Research II 43, 1413–1434.
Yan, X.H., Ho, C.R., Zheng, Q., Klemas, V., 1992. Tempera-
ture and size variabilities of the Western Pacific Warm Pool.
Science 258, 1643–1645.
H. Yamashita et al. / Deep-Sea Research II 49 (2002) 2823–28622862