
Xbap,a Vertebrate Gene Related to bagpipe, Is Expressed in Developing Craniofacial Structures and in...
11
DEVELOPMENTAL BIOLOGY 181, 223 –233 (1997) ARTICLE NO. DB968416 Xbap, a Vertebrate Gene Related to bagpipe, Is Expressed in Developing Craniofacial Structures and in Anterior Gut Muscle Craig S. Newman, Matthew W. Grow, Ondine Cleaver, Frank Chia, and Paul Krieg 1 Center for Developmental Biology, Department of Zoology, University of Texas at Austin, Austin, Texas 78712 The Drosophila bagpipe (bap) gene is involved in the specification of the musculature of the embryonic midgut. We report the isolation and characterization of a Xenopus sequence, Xbap, which is closely related to bap. Xbap is also expressed in the developing musculature of the midgut, suggesting that this developmental role of bagpipe is evolutionarily conserved. However, a second, novel role in development is suggested by the observation that Xbap is also expressed in a region of the developing facial cartilage. Using a combination of cartilage staining and comparison to the goosecoid head expression pattern, we show that Xbap expression marks the precursors to the basihyobranchial, palatoquadrate, and possibly Meckel’s cartilages. This vertebrate bagpipe sequence therefore is expressed in both mesodermally and neural crest-derived tissues. q 1997 Academic Press INTRODUCTION appears to be homologous to the murine Pax-6 and human Aniridia genes (Quiring et al., 1995). The embryonic expres- sion patterns of these three genes are very similar, being Homeodomain-containing proteins have been implicated expressed in the developing central nervous system and eye. in the control of a variety of developmentally important Ectopic expression of ey is capable of inducing the forma- events including the patterning of the anterior/posterior tion of essentially normal eyes in various parts of the fly axis and organogenesis. In virtually all cases, expression of body, and the mouse Pax-6 sequence can effectively mimic the homeobox gene precedes any overt, cellular differentia- this activity (Halder et al., 1995). It thus appears that both tion, suggesting that homeodomain proteins act as molecu- the sequence and the function of these genes has been con- lar switches, instructing cells to adopt particular states of served during evolution. Another such example of conserva- determination. In this way, cells that are initially identical tion in the role of homeodomain proteins between verte- can go on to different fates by expressing different comple- brates and invertebrates comes from the study of the devel- ments of homeobox genes. Specific homeobox genes are opment of the heart. In Drosophila, the tinman gene (tin) implicated in the specification and subsequent maturation is required for development of the dorsal vessel, the fly of a number of vertebrate organs and organ systems. Exam- equivalent of the heart, and for the development of the mus- ples include Hox11 in the spleen (Roberts et al., 1994; Dear culature of the midgut (Bodmer, 1993). Knockout of Nkx2-5, et al., 1995), Hlx in the gut and liver (Hentsch et al., 1996), the putative mouse homolog of tinman results in abnormal Nkx2-1/TTF-1 in the thyroid (Lazarro et al., 1991; Guazzi development of the cardiac musculature, suggesting that et al., 1990, 1994), and Pdx1 in the pancreas (Jonsson et al., the two genes share similarity at both the sequence and 1994). functional level (Komuro and Izumo, 1993; Lints et al., In several cases, the insect and vertebrate homologs have 1993; Lyons et al., 1995). been found to play a role in the development of correspond- The tinman gene is related in sequence and is geno- ing organs. For example, the Drosophila eyeless (ey) gene mically linked to the bagpipe (bap) gene. Both are members of the NK2 class of homeobox genes, which now contains many members from a wide variety of animal phyla (Har- 1 To whom correspondence should be addressed. Fax: (512) 471- 9651. E-mail: [email protected]. vey, 1996). bap transcripts are found in a subset of tin- 223 0012-1606/97 $25.00 Copyright q 1997 by Academic Press All rights of reproduction in any form reserved.
Transcript of Xbap,a Vertebrate Gene Related to bagpipe, Is Expressed in Developing Craniofacial Structures and in...

DEVELOPMENTAL BIOLOGY 181, 223–233 (1997)ARTICLE NO. DB968416
Xbap, a Vertebrate Gene Related to bagpipe,Is Expressed in Developing CraniofacialStructures and in Anterior Gut Muscle
Craig S. Newman, Matthew W. Grow, Ondine Cleaver,Frank Chia, and Paul Krieg1
Center for Developmental Biology, Department of Zoology, University of Texas at Austin,Austin, Texas 78712
The Drosophila bagpipe (bap) gene is involved in the specification of the musculature of the embryonic midgut. We reportthe isolation and characterization of a Xenopus sequence, Xbap, which is closely related to bap. Xbap is also expressedin the developing musculature of the midgut, suggesting that this developmental role of bagpipe is evolutionarily conserved.However, a second, novel role in development is suggested by the observation that Xbap is also expressed in a region ofthe developing facial cartilage. Using a combination of cartilage staining and comparison to the goosecoid head expressionpattern, we show that Xbap expression marks the precursors to the basihyobranchial, palatoquadrate, and possibly Meckel’scartilages. This vertebrate bagpipe sequence therefore is expressed in both mesodermally and neural crest-derived tissues.q 1997 Academic Press
INTRODUCTION appears to be homologous to the murine Pax-6 and humanAniridia genes (Quiring et al., 1995). The embryonic expres-sion patterns of these three genes are very similar, beingHomeodomain-containing proteins have been implicatedexpressed in the developing central nervous system and eye.in the control of a variety of developmentally importantEctopic expression of ey is capable of inducing the forma-events including the patterning of the anterior/posteriortion of essentially normal eyes in various parts of the flyaxis and organogenesis. In virtually all cases, expression ofbody, and the mouse Pax-6 sequence can effectively mimicthe homeobox gene precedes any overt, cellular differentia-this activity (Halder et al., 1995). It thus appears that bothtion, suggesting that homeodomain proteins act as molecu-the sequence and the function of these genes has been con-lar switches, instructing cells to adopt particular states ofserved during evolution. Another such example of conserva-determination. In this way, cells that are initially identicaltion in the role of homeodomain proteins between verte-can go on to different fates by expressing different comple-brates and invertebrates comes from the study of the devel-ments of homeobox genes. Specific homeobox genes areopment of the heart. In Drosophila, the tinman gene (tin)implicated in the specification and subsequent maturationis required for development of the dorsal vessel, the flyof a number of vertebrate organs and organ systems. Exam-equivalent of the heart, and for the development of the mus-ples include Hox11 in the spleen (Roberts et al., 1994; Dearculature of the midgut (Bodmer, 1993). Knockout of Nkx2-5,et al., 1995), Hlx in the gut and liver (Hentsch et al., 1996),the putative mouse homolog of tinman results in abnormalNkx2-1/TTF-1 in the thyroid (Lazarro et al., 1991; Guazzidevelopment of the cardiac musculature, suggesting thatet al., 1990, 1994), and Pdx1 in the pancreas (Jonsson et al.,the two genes share similarity at both the sequence and1994).functional level (Komuro and Izumo, 1993; Lints et al.,In several cases, the insect and vertebrate homologs have1993; Lyons et al., 1995).been found to play a role in the development of correspond-
The tinman gene is related in sequence and is geno-ing organs. For example, the Drosophila eyeless (ey) genemically linked to the bagpipe (bap) gene. Both are membersof the NK2 class of homeobox genes, which now containsmany members from a wide variety of animal phyla (Har-1 To whom correspondence should be addressed. Fax: (512) 471-
9651. E-mail: [email protected]. vey, 1996). bap transcripts are found in a subset of tin-
223
0012-1606/97 $25.00Copyright q 1997 by Academic PressAll rights of reproduction in any form reserved.
AID DB 8416 / 6x16h$$581 12-28-96 00:42:15 dba

224 Newman et al.
expressing cells within the midgut and cardiac precursors Whole-Mount in Situ Hybridization and Sectioningof Drosophila (Azpiazu and Frasch, 1993) and expression is
Digoxygenin-labeled antisense RNA probes were prepared usingfirst observed in the stage 10 embryo when 11 discrete, bap-a standard protocol (Boehringer Mannheim). Xbap template wasexpressing patches of cells appear in the dorsal mesodermlinearized using NotI and transcribed using T7 RNA polymeraseof the embryo. In time, these patches fuse to form a continu-to yield a 1.8-kb product. A fragment of the goosecoid (gsc) gene
ous band of bap-expressing cells fated to form the muscula- in pGEM7z was kindly provided by A. Hemmati-Brivanlou. Forture of the midgut. The cardiac precursors of the embryo probe preparation, the plasmid was linearized with XhoI and tran-do not express bap until a somewhat later stage. bap is also scribed with SP6 RNA polymerase. Whole-mount in situ hybridiza-expressed in cells at the stomodeum and proctodeum that tion was carried out as described by Harland (1991) except thatare precursors to the visceral mesoderm of the foregut and Chaps was omitted at all steps. Double-staining in situ hybridiza-
tion was performed using BCIP and magentaphos as staining re-hindgut. While disruption of this gene results in abnormalagents (Knecht et al., 1995). For serial sectioning, whole-mount-development of midgut musculature, the foregut, hindgut,stained embryos were rinsed in TBS, dehydrated in ethanol (21 1and dorsal vessel all form normally.hr), incubated in xylene (21 10 min), incubated in paraplast (11 10In this paper, we report the isolation and characterizationmin in 1:1 xylene:paraplast; 11 30 min, and then overnight in 100%of Xbap, a bap-related gene in Xenopus. The vertebrate andparaplast), and embedded in paraplast. The 10-mm sections were
Drosophila genes are highly conserved within the homeo- cut, mounted in Permount, and observed by differential interfer-box and to a lesser extent in several other domains, and so ence optics.Xbap may represent the vertebrate homolog of the bap gene.In the Xenopus embryo, Xbap is expressed in the posteriorforegut, approximately corresponding to one of the domains
Cartilage Stainingof bap expression in the fly. Xbap is also expressed at highlevels in developing craniofacial structures, suggesting that
Staining of embryonic cartilage was carried out as described bythis homeobox sequence has acquired a novel develop-Klymkowsky and Hanken (1991) but omitting the alzarian redmental function during evolution.staining.
METHODS AND MATERIALSRNase Protection and RT–PCR Analysis
Isolation of Xbap SequencesEmbryonic RNA was isolated by homogenizing embryos in 50
mM Tris–Cl (pH 7.5), 50 mM NaCl, 10 mM EDTA, and 0.5% SDS.Approximately 106 plaques of a Xenopus laevis adult heart cDNAlibrary (Ji et al., 1993) were plated and screened at low stringency Proteinase K was added to 0.25 mg/ml and after 1 hr at 377C the
homogenate was phenol/chloroform extracted and isopropanol pre-(51 SSC/0.1% SDS at room temperature for 30 min) with a probecontaining the homeobox of the XNkx2-5 gene (Tonissen et al., cipitated. A further purification was achieved by resuspending the
RNA in water and precipitating with an equal volume of 8 M LiCl.1994). Of numerous recombinants detected under these low strin-gency conditions, a single clone was found to contain bap-related RNA from the equivalent of 7.5 embryos was analyzed by RNase
protection (Krieg and Melton, 1987) using probe against approxi-sequences when sequenced using the degenerate homeobox primer5*-AACCADATYTTNACYTG-3*. Additional Xbap clones were mately 400 nt of the Xbap 3* UTR. The ubiquitously expressed
Xmax sequence (Tonissen and Krieg, 1994) was used as control.isolated from a Xenopus stage 28 head cDNA library (Hemmati-Brivanlou et al., 1991), but none of these contained the N-terminal RNA was isolated from organs of an adult male using the Totally
RNA kit (Ambion). Complementary DNA was synthesized fromprotein coding region. The extreme 5* end of the coding region wastherefore isolated using inverse PCR on genomic DNA as follows. 500 ng of total RNA using standard techniques. PCR was performed
using a pair of Xbap specific primers (5*-ACCATGTTCAGAGGC-X. laevis genomic DNA isolated from red blood cells was digestedwith the restriction enzyme EcoRV, ligated into circles, and then CCTCT-3* and 5*-AGATCCTCCAGCAACAG-3*) and a pair of EF-
1a-specific primers (5*-GGTCTGCGGCGAGTTCTAAC-3* and 5*-linearized with the restriction enzyme BstXI. The genomic frag-ment was PCR amplified with the Xbap-specific primers 5*-AGA- GAGGGCATGCTCACGGGTTT-3* ) (Krieg et al., 1989). PCR con-
ditions were 35 cycles of 957C for 30 sec, 557C for 30 sec, and 727CCTGCAGTGGCCAGCCTG-3* and 5*-ATTCAGCCCTCAGTG-AGGAG-3* for 30 cycles of 1 min at 957C, 1 min at 587C, and 4 for 90 sec. Resulting Xbap products were fractionated on an agarose
gel, blotted onto Hybond-N (Amersham), and hybridized with 32P-min at 727C. The resulting product was cloned into pBluescript KS/
and the sequence of the N-terminal region of Xbap was determined. labeled Xbap probe.
FIG. 1. (A) Nucleotide and derived amino acid sequence of the Xbap coding region. The first 192 nt are derived from Xenopus genomicDNA, while the remainder of the sequence is from a cDNA clone. The two potential start methionines are marked by arrows. Thehomeodomain sequence is underlined, the TN domain is double underlined, the NK-2SD is marked by a dashed line, and the BAP domainis overlined. An intron is located at the site marked by the open arrow. (B) Comparison of the amino acid sequences of conserved regionsin bap family members. Identical amino acids are represented by a dash. The conserved tyrosine residue diagnostic of the NK-2 class ofhomeodomains is marked with an arrow.
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8416 / 6x16h$$582 12-28-96 00:42:15 dba

225Vertebrate Gene Related to bagpipe
12-28-96 00:42:15 dba

226 Newman et al.
RESULTS
Isolation of Xbap cDNA Clones
In an effort to identify novel homeobox sequences ex-pressed in the vertebrate heart, a DNA probe containing theXNkx2-5 homeobox was used to screen a X. laevis adultheart cDNA library at low stringency. Together with manystrongly positive plaques representing the XNkx2-5 gene,several plaques of lower intensity were isolated. Sequencingof homeodomain regions revealed that one of these cDNAswas 87% identical at the amino acid level to the Drosophilabagpipe gene (Fig. 1B). We have called this Xenopus se- FIG. 2. Developmental profile of Xbap expression. Approximately
7.5 embryo equivalents of total RNA from different developmentalquence Xbap.stages was subjected to RNase protection analysis using an Xbap-The nucleotide sequence of the Xbap coding region andspecific probe from the 3*UTR. A probe from the ubiquitous Maxthe conceptual translation of the Xbap protein are presentedgene was included in the same protection reaction, to ensure equalin Fig. 1A. The open reading frame is 987 nucleotides long,recovery of input RNA. The numbers at the top of the lanes indicateencoding a 329-amino acid protein with a predicted molecu-the Xenopus embryonic stage (Nieuwkoop and Faber, 1994).lar weight of 36 kDa. In addition to the homeobox, three
other regions of sequence similarity to bap can be identified(Fig. 1B). The first is the TN domain, a 10-amino acid motiflocated near the N-terminal of NK-2 class homeobox pro- transcripts at the late neurula stage of development (stageteins (Harvey, 1996). Xbap and bap are identical in 7 of 10 18). The level of Xbap mRNA increases slowly up to theresidues in the TN domain. The TN domains in Drosophila tailbud stage (stage 28) when the transcript abundancetin and bap are located 35 and 22 amino acids, respectively, climbs sharply. Xbap RNA levels then remain approxi-from the start methionine, and in all vertebrate tin homo- mately constant through the tailbud and tadpole stageslogs, the TN domain begins within 9 amino acids of the (stages 30–44). The two protected bands visible in Fig. 2 arestart methionine. In contrast, the TN domain in Xbap is probably due to the fact that Xenopus is pseudotetraploidlocated 65 amino acids from the first in-frame methionine. and usually contains two closely related copies of eachAnother in-frame methionine is located 10 amino acids up- mRNA (Kobel and Du Pasquier, 1986). To localize Xbapstream of the TN domain, and while there is no compelling expression in the embryo, we have carried out whole-mountreason to believe that translation starts at this site, the in situ hybridization studies. Using this technique, Xbapsecond methionine is contained in a context that more expression is first detected in the developing face and some-closely resembles the preferred translation initiation con- what later in anterior gut tissues. To simplify the descrip-sensus sequence (Kozak, 1986). A second region of similar- tion, the facial and gut expression patterns will be presentedity, the NK-2-specific domain (NK-2SD), is located immedi- separately.ately downstream of the homeodomain (Harvey, 1996). Craniofacial expression of Xbap is first detected in theOver the 17-amino acid stretch encompassing the NK-2SD, early tailbud embryo (stage 30) and becomes clearly defined7 residues are identical in Xenopus and Drosophila bap, slightly later as distinct, bilaterally symmetrical bars, onincluding a highly conserved stretch of hydrophobic resi- either side of the cement gland and flanking the futuredues making up the core of the domain (Fig. 1B). Both bap mouth opening (Figs. 3A–3C). As development proceeds,and Xbap are relatively diverged from the consensus NK- this region contracts, until Xbap expression is concentrated2SD sequence seen in the other NK-2 family members. A in two regions immediately ventroposterior to the stomo-final region of sequence identity is located 16 amino acids deum (Figs. 3D–3F). We have compared the facial expres-C-terminal of the NK-2SD. This five amino acid region is sion of Xbap to goosecoid (gsc), which is expressed in Meck-not present in any other NK-2 family member, and so we el’s cartilage and the hyoid arch in developing mouse (Gaunthave called it the BAP domain (Fig. 1B). An intron splits et al., 1993; Riveria-Perez et al., 1995; Yamada et al., 1995)the Xbap coding sequence within amino acid 180, 65 nucle- and zebrafish embryos (Schulte-Merker et al., 1994). In theotides upstream of the homeobox. The bap gene contains late-tailbud Xenopus embryo (stage 36) gsc is expressedan intron in the same approximate region, 78 nucleotides strongly in the ventral head region (Figs. 3G and 3H). Fromupstream of the homeodomain. the side, the gsc expression pattern appears very similar to
the Xbap pattern. However, viewed from either the dorsalor anterior aspect, gsc expression extends more toward theEmbryonic Expression of the Xbap Genemidline than Xbap (compare Figs. 3B and 3C and 3H and 3I).By tadpole stage (stage 42), the two patches of gsc expressionThe developmental expression profile of Xbap was deter-
mined by RNase protection (Fig. 2). Although not visible in almost meet at the midline of the embryo (Figs. 3J and 3K),while at the same stage Xbap expression does not extendthe figure, the original autoradiograph first reveals Xbap
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8416 / 6x16h$$582 12-28-96 00:42:15 dba

227Vertebrate Gene Related to bagpipe
as far toward the midline (Fig. 3F). In summary, although oping heart (Fig. 3A). Initially located high on the flank, thearea of expression shifts ventrally, broadening at the base,the regions of Xbap and gsc expression appear to partly
overlap, especially at the early stages, gsc expression is con- as development proceeds. Furthermore, expression progres-sively diminishes on the right side of the embryo, finallycentrated closer to the midline than Xbap.
The medial region of Xbap facial expression (Fig. 3A) is disappearing completely at about stage 40 (Figs. 5A–5C). Atthis stage, Xbap expression is still prominent on the leftfirst detected at approximately stage 32, posterior to the
initial sites of staining and below the floor of the developing side of the embryo on the surface of the developing anteriorgut tube (Fig. 5C).pharyngeal cavity. This domain is consistent with the re-
gion where the basihyobranchial cartilage will develop. Ex- Sections through stage 37 embryos, when bilateral gutexpression is still present, show that Xbap expression ispression of gsc occurs in the same region (Fig. 3G); however,
while Xbap is confined to a narrow band as viewed from localized to a thin layer of cells below the surface ectoderm(Fig. 5D). By stage 41 the staining extends much deeperthe anterior (Fig. 3B), gsc expression extends further later-
ally (Fig. 3H). below the surface of the embryo, and a thin layer of Xbap-expressing cells is separated from the lumen of the gut bySections through stained embryos allow a more precise
localization of Xbap and gsc expression in the head. A trans- a thick layer of nonexpressing endodermal tissue (Fig. 5E).The Xbap-expressing cells are located at the outer edges ofverse section through the head of a stage 37 embryo stained
for both Xbap and gsc transcripts shows that Xbap expres- the gut tube, which will go on to form the gut musculature,as assayed by antibody staining for expression of smoothsion is located in two patches, ventrolateral to the mouth
cavity, within the mesenchymal layer (Fig. 4A). A region of muscle actin (data not shown).Somewhat surprisingly, in situ hybridization studies dogsc-specific staining is located between the Xbap positive
cells and extending below the oral cavity (Fig. 4A). A hori- not reveal Xbap expression in the posterior gut region,where bap is expressed in the Drosophila embryo. To exam-zontal section reveals gsc staining anterior to the mouth
cavity, flanked by Xbap staining (Fig. 4B) and a higher mag- ine this matter more closely, we have carried out RNaseprotection and PCR assays to determine whether Xbap tran-nification of this region (Fig. 4C) shows that a small group
of cells between the domains of Xbap and gsc expression scripts are present in developing hindgut. In experimentsusing late tailbud embryos (stage 34), total RNA was iso-appears to express both sequences. In older embryos (stage
42), at the time of chondrification of the face, Xbap staining lated from the head, middle, and tail segments (Fig. 6A)and assayed for Xbap sequences by RNase protection. As amarks the anterior and lateral regions of the mouth cavity
(Fig. 4D). The medial region of Xbap expression is also a control, the same samples were probed for XNkx2-5 mRNA,which is expressed in the heart and in the developing spleenpart of the mesenchymal layer, staining a narrow patch of
cells below the posterior pharyngeal cavity (Fig. 4E). In addi- in the foregut region (K. Patterson, personal communica-tion). This probe serves to confirm that no foregut tissuetion, faint staining is visible below the surface epithelium,
within the pharyngeal arches, possibly marking the pre- was included in the tail segment. The results show that, inthe tailbud embryo, Xbap transcripts are detectable in thesumptive branchial cartilages (Fig. 4E). In comparison, the
medial region of gsc expression is visible as a broad patch anterior and middle segments of the embryos, but not inthe posterior region where the hindgut is developing (Fig.of staining across the middle third of the embryo, at approxi-
mately the level of the developing gills (Fig. 4F). This do- 6B). We have used RT–PCR to assay for Xbap expressionin later stage embryos, when gut structures are clearly elab-main of expression is distinctly broader than the equivalent
region of Xbap expression (Fig. 4E). orated. Briefly, digestive tracts from stage 46 tadpoles weredivided into foregut and hindgut sections by cutting at theThe other, major region of Xbap expression in the embryo
is the anterior gut. In tailbud embryos, Xbap transcripts are apex of the gut coil and total RNA was isolated from eachsegment. RT–PCR analysis demonstrates that Xbap tran-detected in bilateral regions dorsoposterior to the devel-
FIG. 3. Whole mount in situ hybridization pattern of Xbap (A, B, C, D, E, and F) and gsc (G, H, I, J, and K) and Alcian blue staining tovisualize cartilage (L). All embryos were photographed at 701 magnification except for (D) at 601. (A) Lateral view of a stage 36 embryoshowing both regions of facial staining and the region of gut expression (indicated by g in the figure). (B) Anterior view of the embryopictured in (A). The paired anterior regions of expression lie on either side of the future mouth opening (M). (C) Dorsal view of a stage37 embryo showing the forked appearance of the Xbap staining. (D) Lateral view of the left side of a stage 42 embryo showing stainingof the anterior gut tube and facial staining. (E) Dorsal view of a stage 40 embryo. Xbap staining now marks crescent-shaped regions oneither side of the future mouth opening (M). (F) Dorsal view of the embryo shown in (D). The region of staining corresponding to thepalatoquadrate cartilage (PQ) is indicated. (G) Lateral view of a stage 36 embryo showing gsc expression in the face. (H) Anterior view ofthe embryo pictured in (G). Compare the gsc staining pattern with that of Xbap in (B) and note the vertical nature of the bilateral patchesof staining and the broad domain of expression of the midline staining. (I) Dorsal view of the embryo shown in G and H. (J) Lateral viewof a stage 42 embryo stained for gsc transcript. (K) Dorsal view of the embryo pictured in J showing the convergence of the gsc stainingat the embryonic midline. (L) Ventral view of the head of a stage 43 embryo stained for cartilage. Compare the location of the palatoquadratecartilage (PQ) with that of Xbap staining in similar stage embryos (E and F). Meckel’s cartilage (MC), ceratohyal cartilage (CH), andbasihyobranchial cartilage (BHB) are also shown.
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8416 / 6x16h$$582 12-28-96 00:42:15 dba

228 Newman et al.
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8416 / 6x16h$8416 12-28-96 00:42:15 dba
229Vertebrate Gene Related to bagpipe
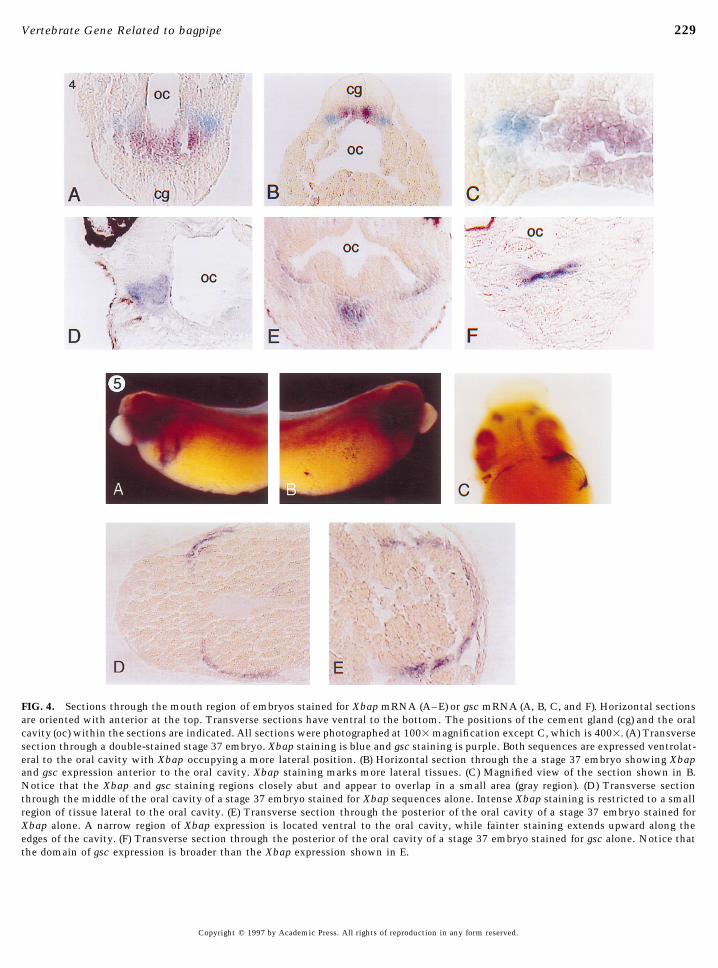
FIG. 4. Sections through the mouth region of embryos stained for Xbap mRNA (A–E) or gsc mRNA (A, B, C, and F). Horizontal sectionsare oriented with anterior at the top. Transverse sections have ventral to the bottom. The positions of the cement gland (cg) and the oralcavity (oc) within the sections are indicated. All sections were photographed at 1001magnification except C, which is 4001. (A) Transversesection through a double-stained stage 37 embryo. Xbap staining is blue and gsc staining is purple. Both sequences are expressed ventrolat-eral to the oral cavity with Xbap occupying a more lateral position. (B) Horizontal section through the a stage 37 embryo showing Xbapand gsc expression anterior to the oral cavity. Xbap staining marks more lateral tissues. (C) Magnified view of the section shown in B.Notice that the Xbap and gsc staining regions closely abut and appear to overlap in a small area (gray region). (D) Transverse sectionthrough the middle of the oral cavity of a stage 37 embryo stained for Xbap sequences alone. Intense Xbap staining is restricted to a smallregion of tissue lateral to the oral cavity. (E) Transverse section through the posterior of the oral cavity of a stage 37 embryo stained forXbap alone. A narrow region of Xbap expression is located ventral to the oral cavity, while fainter staining extends upward along theedges of the cavity. (F) Transverse section through the posterior of the oral cavity of a stage 37 embryo stained for gsc alone. Notice thatthe domain of gsc expression is broader than the Xbap expression shown in E.
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8416 / 6x16h$8416 12-28-96 00:42:15 dba

230 Newman et al.
FIG. 7. RT–PCR analysis of Xbap expression in adult tissues.Total RNA was collected from the indicated adult organs and as-sayed for Xbap transcripts using gene specific primers as describedunder Materials and Methods. RT–PCR amplification of the ubiq-uitous EF-1a sequence (visualized by ethidium bromide staining)was used as a control for RNA levels.
and stomach show significant levels of Xbap transcript andslightly lower levels are detected in the intestine, skeletalmuscle, and tongue. Heart, liver, and lung contain little orno Xbap RNA. These results indicate that the late-stageembryonic expression of Xbap in gut tissues is maintainedin the corresponding adult organs.
DISCUSSIONFIG. 6. Localization of Xbap transcripts in the tailbud embryo.(A) Stage 34 embryos were dissected into head, middle, and tail Isolation of a Vertebrate Gene Related to bapsections as indicated and total RNA was isolated. (B) Xbap tran-
We have isolated and characterized a vertebrate sequencescript distribution assayed by RNase protection. Total RNA fromclosely related to Drosophila bagpipe. The sequence of thethe head, middle, or tail segments of stage 34 embryos was assayed
using either Xbap or XNkx2-5 probe. Both XNkx2-5 and Xbap Xbap protein is highly conserved with respect to bap in aprobes generate two protected fragments, most likely due to dupli- number of important regions, including the TN domain,cated genes in the pseudotetraploid Xenopus genome (Kobel and homeodomain, NK-2SD, and the BAP domain. Within theDu Pasquier, 1986). Equal RNA input was determined with the homeodomain the Xbap protein sequence is 87% identicalMax probe. to Drosophila bap (Azpiazu and Frasch, 1993) and 83% iden-
tical to planarian Pnbap (Balavoine, 1996). While the TNdomain and NK-2SD are present in most members of the
scripts are present, at approximately equal levels, in both NK-2 class of homeodomain proteins, no function has yetthe foregut and the hindgut regions (data not been ascribed to these two regions, although in vitro studiesshown). This result indicates that Xbap expression ex- with Nkx2-5 suggests that the NK-2SD may play a role astends into hindgut tissues during the later stages of em- a transcriptional repression domain (Chen and Schwartz,bryogenesis. 1995).
Adult Expression of the Xbap Gene Expression of Xbap in the Developing GutIn the fly embryo, bap is necessary for development ofRT–PCR was used to assay for Xbap expression in a vari-
ety of adult tissues (Fig. 7). The kidney, pancreas, spleen, the visceral musculature of the midgut. In addition, bap is
FIG. 5. Xbap expression in the gut. A and B are at 301 magnification and C is at 701 magnification. (A) Lateral view of the left side ofa stage 38 embryo stained for Xbap. Note the domain of Xbap expression in the gut. (B) Lateral view of the right side of the embryopictured in A illustrating absence of gut staining. (C) Ventral view of a stage 40 embryo stained for Xbap mRNA showing expression inthe developing gut tube on the left side of the embryo. Anterior is up. (D) 1001 magnification of a horizontal section through a stage 37embryo showing Xbap transcripts in a layer of cells underneath the outer ectoderm on both sides of the embryo. Anterior is to the left.(E) 2001magnification view of a horizontal section through a stage 41 embryo showing Xbap expression on the left side, partially encirclingthe gut tube. Anterior is up.
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8416 / 6x16h$$582 12-28-96 00:42:15 dba

231Vertebrate Gene Related to bagpipe
expressed in the precursor cells of the dorsal vessel and inthe musculature of the fore and hindgut. Our studies showthat Xbap has two major expression domains in the verte-brate embryo—the developing cartilage of the head and theanterior viscera. The pattern of Xbap expression in the de-veloping gut muscle of the frog is approximately equivalentto bap expression in the gut muscle of the fly, suggestingthat this bap function may have been evolutionarily con-served. Unlike bap expression in the dorsal vessel of thefly, Xbap sequences are never observed in the heart anlageof the amphibian embryo. However, mutation of the Dro-sophila bap gene does not effect development of the dorsalvessel, suggesting either that bap does not function in thistissue or that a redundant system can replace bap function.Overall, the pattern of Xbap expression in the visceral re-gion is consistent with a role in patterning, perhaps by pro-viding the mesodermal cells of the gut with positional infor-mation along the A/P axis, or by specifying the developmentof the muscular tissue.
Expression of Xbap during CraniofacialDevelopment
The skeleton of the developing head of Xenopus has beendescribed in detail (Patterson, 1939; Trueb and Hanken,
FIG. 8. Dorsal (A), ventral (B), and lateral (C) views of the major1992) and is diagrammed in Fig. 8. The lower jaw of thecartilages in the anterior Xenopus head at stage 45. The mouth oftadpole consists of paired Meckel’s cartilages joined anteri-the tadpole is situated between the superrostral plate and the lowerorly by a single infrarostral cartilage. Posteriorly, the jawjaw, which is made up of the infrarostral and Meckel’s cartilages.articulates on the palatoquadrate cartilage. Posterior toThe approximate regions of Xbap expression are represented by
Meckel’s cartilage, and underlying the pharyngeal cavity, hatched areas. BHB, basihyobranchial cartilage; CH, ceratohyal car-are the paired ceratohyal cartilages. Positioned between the tilage; IR, infrarostral cartilage; MC, Meckel’s cartilage; PQ, palato-two ceratohyals, and extending beyond them posteriorly, is quadrate cartilage.the basihyobranchial cartilage. Mesenchymal-derived headstructures in vertebrates arise from two distinct sources(Noden, 1988). In the amphibian, muscle, blood vessels,connective tissue, and a portion of the skull arise from em- comparisons of Xbap and gsc expression, we are able to
make the following general observations. While both bapbryonic mesoderm while most of the cartilage of the embry-onic face is derived from the cranial neural crest (Stone, and Xbap are expressed in the region of the developing
mouth, they do not stain homologous structures. In the fly,1926, 1929). In particular, the cells of the mandibular crestsegment (MCS) contribute to the structures of the mandibu- bap is expressed in muscle precursors of the anterior fore-
gut. In contrast, Xbap expression marks precursors of thelar arch, including Meckel’s cartilage in the lower jaw andthe palatoquadrate (Sadaghiani and Thiebaud, 1987). In both cartilaginous elements of the face. Later in development
cartilage staining indicates that Xbap is expressed in theAmblystoma and Rana, extirpation of the MCS greatly re-duces or eliminates formation of a number of facial carti- palatoquadrate cartilage, near the point of articulation to
Meckel’s cartilage and possibly in the caudal regions oflages including Meckel’s cartilage and the palatoquadrate(Stone, 1926, 1929). The expression of Xbap in the region Meckel’s cartilage itself. The infrarostal cartilage, separat-
ing the paired Meckel’s cartilage is Xbap negative but mayof these developing cartilages suggests a mesectodermal ori-gin for some of the Xbap-positive cells. express gsc sequences (Fig. 3K). Xbap expression also marks
the forming basihyobranchial cartilage below the floor ofPrecise identification of the tissues expressing Xbap inthe frog embryo, especially at the earliest stages, is compli- the pharynx at the level of the hyoid and branchial arches
(Fig. 4E). Interestingly, unlike most other cartilage of thecated by the lack of definitive markers for facial structuresprior to differentiation of cartilage, which occurs at about facial region, the anuran basihyobranchial cartilage is not
derived from the cranial neural crest, but has its originsstage 40 (unpublished observations). To further complicatethe situation, it is not possible to distinguish between in the anterior mesoderm (Sadaghiani and Thiebaud, 1987;
Seufert and Hall, 1990). Thus, Xbap appears to be expressedchanging domains of Xbap expression and simple move-ment of the precursor structures during facial development. in both mesodermal and mesectodermal tissue within the
head.Nevertheless, by a combination of cartilage staining and
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8416 / 6x16h$$582 12-28-96 00:42:15 dba

232 Newman et al.
Harland, R. (1991). In situ hybridization: An improved wholeThe evolutionary appearance of the neural crest coincidesmount method for Xenopus embryos. Methods Cell Biol. 36,with the appearance of the vertebrate phylum, being found685–695.solely in this group of animals (Gans and Northcutt, 1983).
Harvey, R. P. (1996). NK-2 homeobox genes and heart development.It thus appears that expression of Xbap in the cartilage of theDev. Biol. 178, 203–216.jaw is not an evolutionary conservation of bap expression in
Hemmati-Brivanlou, A., de la Torre, J., Holt, C., and Harland, R.the arthropods, but represents a new role for the bap se- (1991). Cephalic expression and molecular characterization ofquence. It seems likely that, during evolution of the neural Xenopus en-2. Development 111, 715–724.crest, the bap sequence was appropriated for use in speci- Hentsch, B., Lyons, I., Li, R., Hartley, L., Lints, T. J., Adams, J. M.,fying distinct elements of the jaw. Considering this observa- and Harvey, R. P. (1996). Hlx homeo box gene is essential for antion, it may be of interest to investigate the expression pat- inductive tissue interaction that drives expansion of embryonic
liver and gut. Genes Dev. 10, 70–79.tern of the bap homolog in the jawless fish, a vertebrateJi, H., Sandberg, K., Zhang, Y., and Catt, K. J. (1993). Molecularwhich possesses a neural crest but lacks a true jaw.
cloning, sequencing and functional expression of an amphibianangiotensin II receptor. Biochem. Biophys. Res. Commun. 194,756–762.ACKNOWLEDGMENTS Jonsson, J., Carlsson, L., Edlund, T., and Edlund, H. (1994). Insulinpromoter factor 1 is required for pancreas development in mice.
We thank Dan Suefert for advice and insight and for comments Nature 371, 606–609.on the manuscript. We thank Klaus Kalthoff and Peter Vize for Klymkowsky, M. W., and Hanken, J. (1991). Whole-mount stainingcritical reading of the manuscript. We are especially grateful to of Xenopus and other vertebrates. Method Cell Biol. 36, 419–Nicola Civill and Tim Mohun for communicating the sequence of 441.an independently isolated full-length Xbap cDNA clone prior to Knecht, A. K., Good, P. J., Dawid, I. B., and Harland, R. M. (1995).publication. This work was supported by NIH Grants HD25179 Dorsal–ventral patterning and differentiation of noggin-inducedand HL52746 to P.A.K. neural tissue in the absence of mesoderm. Development 121,
1927–1936.Kobel, H. R., and Du Pasquier, L. (1986). Genetics of polyploid Xen-
opus. Trends Genet. 2, 310–315.REFERENCESKozak, M. (1986). Point mutations define a sequence flanking the
AUG initiator codon that modulates transcription by eukaryoticAzpiazu, N., and Frasch, M. (1993). tinman and bagpipe: Two ho- ribosomes. Cell 44, 283–292.
meobox genes that determine cell fates in the dorsal mesodermKomuro, I., and Izumo, S. (1993). Csx: A murine homeobox-con-
of Drosophila. Genes Dev. 7, 1325–1340.taining gene specifically expressed in the developing heart. Proc.
Balavoine, G. (1996). Identification of members of several homeo-Natl. Acad. Sci. USA 90, 8145–8149.
box genes in a planarian using a ligation-mediated polymeraseKrieg, P. A., and Melton, D. A. (1987). In vitro RNA synthesis withchain reaction technique. Nucleic Acids Res. 24, 1547–1553.
SP6 RNA polymerase. Methods Enzymol. 155, 397–415.Bodmer, R. (1993). The gene tinman is required for specification ofKrieg, P. A., Varnum, S. M., Wormington, W. M., and Melton, D. A.the heart and visceral muscles in Drosophila. Development 118,
(1989). The mRNA encoding elongation factor 1-a (EF-1a) is a719–729.major transcript at the midblastula transition in Xenopus. Dev.Chen, C. Y., and Schwartz, R. J. (1995). Identification of novel DNABiol. 133, 93–100.binding targets and regulatory domains of a murine tinman ho-
Lazarro, D., Price, M., De Felice, M., and Di Lauro, R. (1991). Themeodomain factor, Nkx-2.5. J. Biol. Chem. 270, 15628–15633.transcription factor TTF-1 is expressed at the onset of thyroidDear, T. N., Colledge, W. H., Carlton, M. B., Lavenir, I., Larson, T.,and lung morphogenesis and in restricted regions of the foetalSmith, A. J., Warren, A. J., Evans, M. J., Sofroniew, M. V., andbrain. Development 113, 1093–1104.Rabbits, T. H. (1995). The Hox11 gene is essential for cell survival
Lints, T. J., Parsons, L. M., Hartley, L., Lyons, I., and Harvey, R. P.during spleen development. Development 121, 2909–2915.(1993). Nkx-2.5: A novel murine homeobox gene expressed inGans, C., and Northcutt, G. (1983). Neural crest and the origin ofearly heart progenitor cells and their myogenic descendants. De-vertebrates: A new head. Science 220, 268 –274.velopment 119, 419–431.Gaunt, S. J., Blum, M., and De Robertis, E. M. (1993). Expression
Lyons, I., Parsons, L. M., Hartley, L., Li, R., Andrews, J. E., Robb,the mouse goosecoid gene during mid-embryogenesis may markL., and Harvey, R. P. (1995). Myogenic and morphogenetic defectsmesenchymal cell lineages in the developing head, limbs andin the heart tubes of murine embryos lacking the homeobox genebody wall. Development 117, 769–778.Nkx-2.5. Genes Dev. 9, 1654–1666.Guazzi, S., Lonigro, R., Pintonello, L., Boncinelli, E., Di Lauro, R.,
Nieuwkoop, P. D., and Faber, J. (1994). ‘‘Normal Table of Xenopusand Mavilio, F. (1994). The thyroid transcription factor-1 gene islaevis (Daudin),’’ 2nd ed. Garland, New York.a candidate target for regulation by Hox proteins. EMBO J. 13,
Noden, D. M. (1988). Interactions and fates of avian craniofacial339–3347.mesenchyme. Development 103(Suppl.), 121–140.Guazzi, S., Price, M., De Felice, M., Damante, G., Mattei, M.-G.,
Patterson, N. F. (1939). The head of Xenopus laevis. Q. J. Microsc.and Di Lauro, R. (1990). Thyroid nuclear factor 1 (TTF-1) containsSci. 81, 161–234.a homeodomain and displays a novel DNA binding specificity.
Quiring, R., Walldorf, U., Kloter, U., and Gehring, W. J. (1995).EMBO J. 9, 3631–3639.Homology of the eyeless gene of Drosophila to the small eyeHalder, G., Callaerts, P., and Gehring, W. J. (1995). Induction ofgene in Mice and Aniridia in humans. Science 265, 785–789.ectopic eyes by targeted expression of the eyeless gene in Dro-
sophila. Science 267, 1788–1792. Rivera-Perez, J. A., Mallo, M., Gendron-Maguire, M., Gridley, T.,
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8416 / 6x16h$$583 12-28-96 00:42:15 dba

233Vertebrate Gene Related to bagpipe
and Behringer, R. R. (1995). goosecoid is not an essential compo- transplantation of mesectoderm in Amblystoma punctatum. J.Exp. Zool. 44, 95–131.nent of the mouse gastrula organizer, but is required for craniofa-
Stone, L. S. (1929). Experiments showing the role of migrating neu-cial and rib development. Development 121, 3005–3012.ral crest (mesectoderm) in the formation of head skeleton andRoberts, C. W., Shutter, J. R., and Korsmeyer, S. J. (1994). Hox 11loose connective tissue in Rana palustris. Wilhelm Roux Arch.controls genesis of the spleen. Nature 368, 747–749.EntwMech. Org. 118, 40–77.Sadaghiani, B., and Thiebaud, C. H. (1987). Neural crest develop-
Tonissen, K. F., Drysdale, T. A., Lints, T. J., Harvey, R. P., andment in the Xenopus laevis embryo, studied by interspecificKrieg, P. A. (1994). XNkx-2.5, a Xenopus gene related to Nkx-2.5transplantation and scanning electron microscopy. Dev. Biol.and tinman: Evidence for a conserved role in cardiac develop-124, 91–110.ment. Dev. Biol. 162, 325–328.
Schulte-Merker, S., Hammerschmidt, M., Beuchle, D., Cho, K. W., Tonissen, K. F., and Krieg, P. A. (1994). Analysis of a variant MaxDe Robertis, E. M., and Nusslein-Volhard, C. (1994). Expression sequence expressed in Xenopus laevis. Oncogene 9, 33 –38.of zebrafish goosecoid and no tail gene products in wild-type and Trueb, L., and Hanken, J. (1992). Skeletal development in Xenopusmutant no-tail embryo. Development 120, 843–852. laevis. J. Morphol. 214, 1–41.
Seufert, D. W., and Hall, B. K. (1990). Tissue interactions involving Yamada, G., Mansouri, A., Torres, M., Stuart, E. T., Blum, M.,cranial neural crest in cartilage formation in Xenopus laevis. Cell Schultz, M., De Robertis, E. M., and Gruss, P. (1995). TargetedDiffer. Dev. 32, 153–165. mutation of the murine goosecoid gene results in craniofacial
defects and neonatal death. Development 121, 2817–2922.Stone, L. S. (1926). Further experiments on the extirpation and
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8416 / 6x16h$$583 12-28-96 00:42:15 dba




















