
X chromosome map at 75-kb STS resolution, revealing extremes of recombination and GC content
14
10.1101/gr.7.3.210 Access the most recent version at doi: 1997 7: 210-222 Genome Res. R Nagaraja, S MacMillan, J Kere, et al. recombination and GC content. X chromosome map at 75-kb STS resolution, revealing extremes of References http://genome.cshlp.org/content/7/3/210#related-urls Article cited in: http://genome.cshlp.org/content/7/3/210.refs.html This article cites 49 articles, 17 of which can be accessed free at: service Email alerting click here top right corner of the article or Receive free email alerts when new articles cite this article - sign up in the box at the http://genome.cshlp.org/subscriptions go to: Genome Research To subscribe to Copyright © Cold Spring Harbor Laboratory Press Cold Spring Harbor Laboratory Press on July 15, 2011 - Published by genome.cshlp.org Downloaded from
-
Upload
thermofisher -
Category
Documents
-
view
0 -
download
0
Transcript of X chromosome map at 75-kb STS resolution, revealing extremes of recombination and GC content

10.1101/gr.7.3.210Access the most recent version at doi: 1997 7: 210-222Genome Res.
R Nagaraja, S MacMillan, J Kere, et al. recombination and GC content.X chromosome map at 75-kb STS resolution, revealing extremes of
References
http://genome.cshlp.org/content/7/3/210#related-urlsArticle cited in:
http://genome.cshlp.org/content/7/3/210.refs.htmlThis article cites 49 articles, 17 of which can be accessed free at:
serviceEmail alerting
click heretop right corner of the article orReceive free email alerts when new articles cite this article - sign up in the box at the
http://genome.cshlp.org/subscriptions go to: Genome ResearchTo subscribe to
Copyright © Cold Spring Harbor Laboratory Press
Cold Spring Harbor Laboratory Press on July 15, 2011 - Published by genome.cshlp.orgDownloaded from

RESEARCH
X Chromosome Map at 75-kb STS Resolution, Revealing Extremes of Recombination and
GC Content Ramaiah Nagaraja, Sandra MacMillan, Juha Kere, Carmela Jones,
Stephanie Griffin, Matthew Schmatz, Jennifer Terrell, Michael Shomaker, Christopher Jermak, Christian Hott,
Mochubeloa Masisi, Steven Mumm, Anand Srivastava, Giuseppe Pilia, Terence Featherstone, Richard Mazzarella, Sheila Kesterson,
Brigid McCauley, Brian Railey, Frank Burough, Volker Nowotny, Michele D'Urso, David States, Bernard Brownstein, and
David Schlessinger ~
Center for Genetics in Medicine, Washington University School of Medicine, St. Louis, Missouri 63110
A YAC/STS map of the X chromosome has reached an inter-STS resolution of 75 kb. The map density is sufficient to provide YACs or other large-insert clones that are cross-validated as sequencing substrates across the chromosome. Marker density also permits estimates of regional gene content and a detailed comparison of genetic and physical map distances. Five regions are detected with relatively high G + C, correlated with gene richness; and a 17-Mb region with very low recombination is revealed between the Xq13.3 {XISTJ and Xq21.3 XY homology loci.
[The STSs and YACs can be accessed at http'//genome.wustl.edu/cgm/cgm.html/.]
In addition to sustaining an exponentially increas- ing rate of gene finding (Collins 1995), yeast artifi- cial chromosome/sequence-tagged site (STS/YAC)- based maps (Burke et al. 1987; Olson et al. 1989) have begun to reveal additional features of chromo- some structure and dynamics. For example, during the development of maps for subportions of the X chromosome, the existence of a second "pseudoau- tosomal" region at the Xq terminus of the chromo- some was demonstra ted (Freije and Schlessinger 1992; Freije et al. 1992; Li and Hamer 1995), fol- lowed by the discovery that the region shows a unique phenomenon of gene inactivation on both the X and Y homologs (D'Esposito et al. 1996). In another instance, it was shown that a cluster of genes in a delimited segment of XpI 1 escape X in- activation (Miller et al. 1995). As the density of markers across the chromosome has increased be- yond the 100-kb resolution goal suggested for the
1Corresponding author. E-MAIL [email protected]; FAX (314) 362-3203.
"genome init iative," addit ional features are re- vealed, as described here.
The average inter-STS distance o f - 7 5 kb has been achieved by the placement of 2091 STSs on cognate YACs across the 160 Mb of the chromo- some. Collectively, the STSs sample -1% of X- specific sequences. About half of the STSs (962) are made from YAC insert ends (Kere et al. 1992), and another 592 are from randomly derived unique X- chromosomal sequences. However, the STSs also in- clude 97 expressed sequence tags (ESTs) and 190 gene-specific STSs from known genes, as well as 192 dinucleotide and 38 tri- and tetranucleotide repeat markers that detect polymorphism. As a result, the YAC/STS map can be integrated with transcriptional and genetic maps.
RESULTS
Mapping Strategy and Performance
We used a modified "all-walking" form of STS con- tent mapping (Kere et al. 1992) in which STSs were
210 ~GENOME RESEARCH 7:210-222 01997 by Cold Spring Harbor Laboratory Press ISSN 1054-9803/97 $5.00
Cold Spring Harbor Laboratory Press on July 15, 2011 - Published by genome.cshlp.orgDownloaded from

derived systematically from YAC insert ends at the edges of contigs to ensure that subsequent screening of YAC libraries would move progressively into new territory. From the screening results, SEGMAP (see Methods) efficiently generates the most probable orders of clones and STSs, and the likely size of cur- rent contigs.
Based on simulations, we expected that an essen- tially complete map should have been assembled after -600 screenings; instead, >1500 were required. One of the unexpected factors that increased the effort was the smaller size and fourfold higher chimera fre- quency than had been estimated previously for some YAC collections (for review, see Nagaraja et al. 1994). Also, the amount of insert ends that yielded STSs was lower than anticipated, because >60% were either co- cloned sequences from other chromosomes, con- tained repetitive sequence elements, or had a nucleo- tide composition too high in A and T for STS devel- opment. Nevertheless, the intuitive strength of STS content mapping to achieve map closure was realized in the final year of mapping, with a drop in the num- ber of contigs from 120 to 13.
Map Features
The 2091 STSs are aligned and ordered in 5423 YACs (see Methods) across the X chromosome (excluding the 3-Mb centromeric region, which remains un- mapped). The DNA contents of the YACs have all been estimated to be + 10% by pulsed-field gel elec- trophoresis (PFGE) compared to the size of yeast chromosomes (Goffeau et al. 1996), and provide an average of 10-fold clone coverage of each STS on the chromosome.
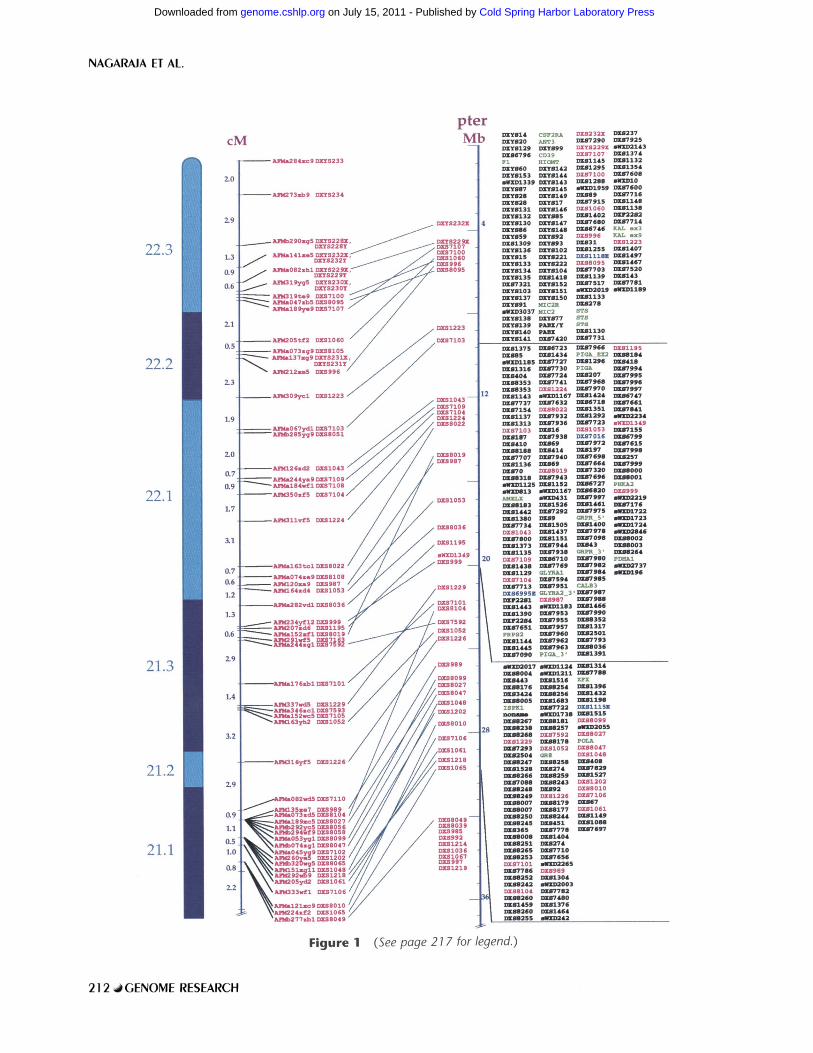
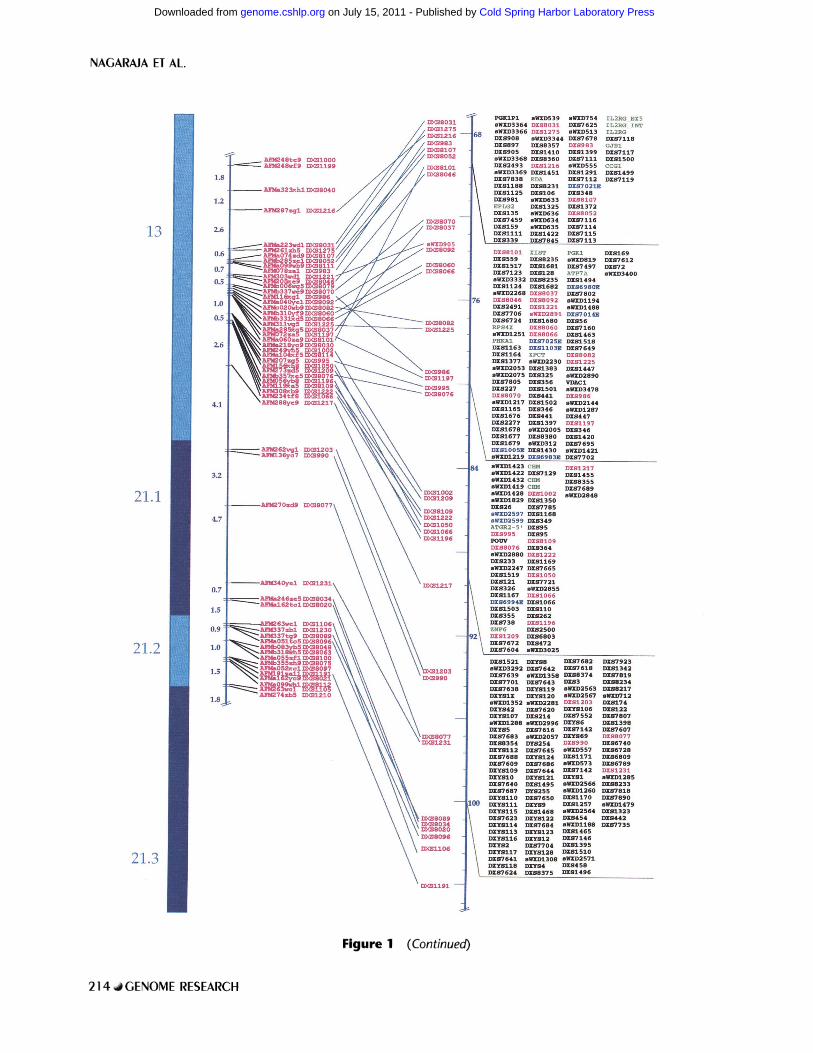
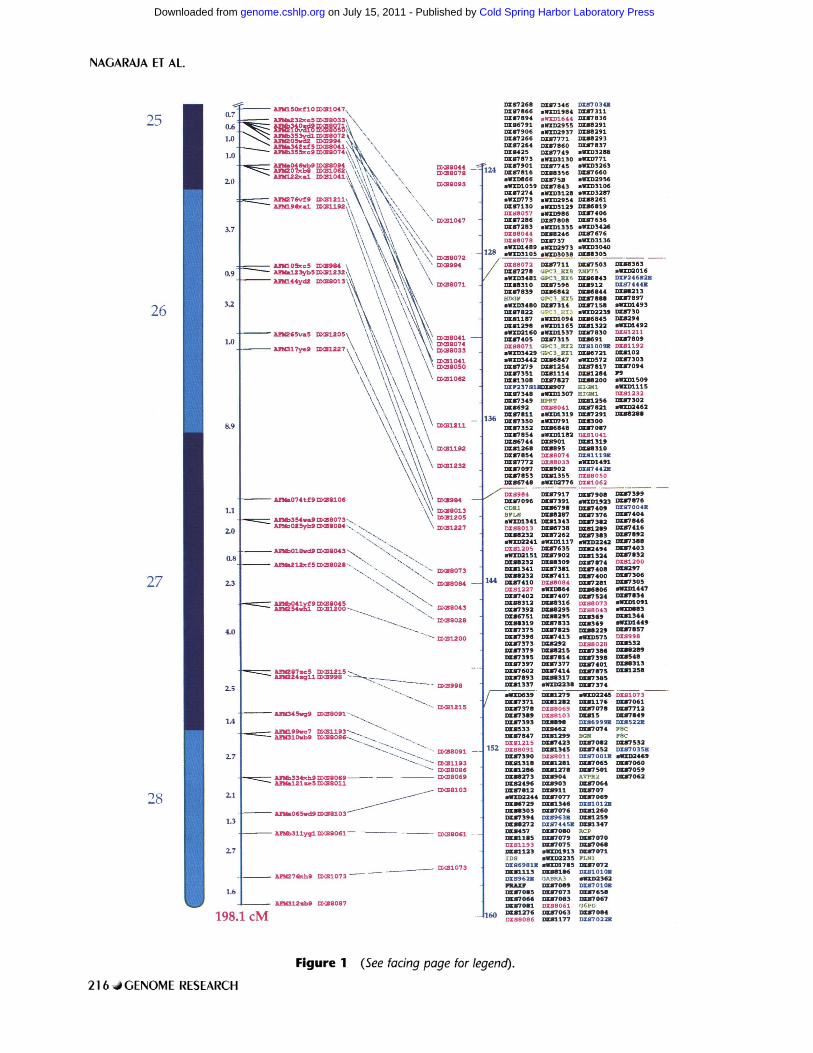
In Figure 1, which is a photograph of a WEB representation, parallel vertical components show the following. (1) An ideogram of the chromosome with approximate cytogenetic band positions. (2) The Centre d'Etude du Polymorphisme Humain (CEPH)/Genethon 198.1-cM genetic linkage map (Dib et al. 1996) normalized to the same overall length as the 160-Mb physical map (with its own index scale farther right). (3) Lines joining the cor- responding positions of dinucleotide repeat markers that are placed on both the physical and genetic linkage maps; crossed lines indicate discordances, which are usually minor. (4) Boxed cohorts of or- dered STSs in 10-Mb intervals from Xpter to Xqter; polymorphic (CA)n markers, genes, and ESTs are color-coded (see Methods and Fig. 1 legend).
The current map is based on a composite inter- active assembly of contigs made completely in the Center for Genetics in Medicine or developed in col-
X CHROMOSOME YAC/STS MAP
laboration with other groups (displayed on a World Wide Web page and published in some cases; see Methods and Acknowledgments). The 13 remaining YAC contigs range from 1.5 to 52 Mb in size, with an average of 12 Mb (the contigs are listed with their marker limits in Methods).
During the construction of the map, YAC sizes and STS content were used in SEGMAP to estimate the minimum, maximum, and average size of na- scent contigs. As STS content increased, the three size estimates converged; and by the time 100-kb inter-STS distances were approached, the sizes of contigs were in close agreement with sizes from other sources. For example, the extent of contigs was the same when assessed by rare-cutter restric- tion enzyme mapping in PFGE for regions of up to 8 Mb in Xp22 (Trump et al. 1996), Xq26 (Pilia et al. 1996), Xq27 (Zucchi et al. 1996), and Xq28 (Palm- ieri et al. 1994). Also, the total sum of successive contigs is within 5% of the 160-Mb content of the X chromosome estimated from cell-sorting experi- ments (Trask et al. 1990); and the contigs in Xq24- qter add up to about one-third the total, or about the cytogenetic fraction of the X included in the X3000.11 cell line containing that region (Nagaraja et al. 1994 and references therein). Thus, the index positions assigned to STSs along the chromosome provide a reasonable first approximation of position and can guide the allocation of regions of known size for long-range sequencing projects (Seventh X Chromosome Workshop 1996, unpubl, report).
Fewer than 20 of 2400 STSs tested thus far fail to find cognate YACs in the screening of 15 genomic DNA equivalents in YACs, suggesting that the map covers >99% of the chromosome. Some of the re- maining apparent gaps are still being filled by on- going screening. Furthermore, in two cases analyzed in Xq28 (Palmieri et al. 1994; Rogner et al. 1994; M. D'Urso, in prep.), DNA gaps in contigs were only on the order of 10-20 kb. Thus, any small amount of uncloned DNA may well be recoverable by long- range PCR amplification from chromosomal DNA preparations.
DISCUSSION
Universality of the Map
The internal consistency of the YAC/STS map in clones derived from seven different libraries of hu- man DNA is in accord with the anticipated general- ity of the structure of the X chromosome in human populations. As expected, the cohort of STSs present in a region proximal or distal to a translocation
GENOME RESEARCH J 21 1
Cold Spring Harbor Laboratory Press on July 15, 2011 - Published by genome.cshlp.orgDownloaded from
22.3
2.9
22.2
22.1
21.3
21.2
21.1
ii/i~ ~i ¸̧~I iiJ:i
i: ?: ? 7
, =,:
. ,
?
(:
. . . .
: i
. . . . .
c M
. . . . ~ 8 4 x c 9 D]~S233
2.0
AEtt273x~ 9 DXYS234
1.3
0 .9
0.6
2.1
0 . 5
1.3
L 9
NAGARAJA ET AL.
2.0
O.7
0.9
1.7
3.1
0.7" . ,,
0.6
1.2
1.3
O.6
2.9
AL~4~I76s~1 DXS7101
L4 ~ -- ,I~1r~i3 3 '~ ~lll ' l~ DXS122 9 ~ - " - A I q H M 3 4 6 z c l D)(875 g3 " ~ A ] ~ 4 a 1 5 2 w c 5 DXS7105
AFM163yh2 ~ 1 0 5 2
3.2 I /
j - AWt~16yE5 DXS1226
2.9 I . / , ~ ~ 082t~115 DXS7110
35ze7 D]~ 989 0.0 ~ A ~ s 0 7 3 s d 5 111188104
~ A ~ I M i a 1 8 9 x e 5 DY,,S8027 1 ~ ~ J U ~ M b 2 92I~5 DX~805 6
~ A J ~ 4 b 2 94Tr~ 9 D x , q 8 0 5 8
1.0 ~ ' ~ AlqHis045y~9 ~T,3 ? 102
0.8 . ~ \ \ ~ A n ~ . s l z ~ r l l m ~ 1 0 4 8 ~k \ ' ~ u m 9 2 . 5 ~ ) m=12~8
2.2 ' ~ " JUrM333~l DY~7106
. \ ~ u . , ~ 2 ~ = ~ ~..s8olo -... =- ~APM224 sf2 DXS1065
AF~277 zbl DY.$8049
pter ~ . ~ , ~ ~ ~ ~ 7
9XYS232X
~ A1+t~2 90x~5 DXYS228X DXYS22 9][ ~,s228,' ~ ~ - ~ ] ~ 7 ~ 0 ~ - / / / I~KSTI00 ~ " AFMa141xe5 D,~ZS232X, / /
DXTS232Y / / L DXS1060 / IPs~P~ ~ r
DXYS229Y ' / I ' "
+-~ ~s23o7 ~ ~ / / ~ 1 ~ . 9 ~ 1 o o ~ / / ~ n ~ o 4 7 ~ s ~x~8ogs ~ / /
~ - ' - ' - - A1q~137x~rg DXTS231~, / / / .r, s2~1, / / /
~ DXS1224 + ~ Alqda06'7)rdl D]~7103 D(XS8022
AFI~2851rg9 DXS8051
D~S8019 D)(S987
A1~125sd2 DXS1043
~ I L F I ~ I 8 4wEI - -
' A l ~ 3 5 0 z f 5 ~ 1 0 5 3
---.--------- A1~(31 l ~ f 5 D]~1224 DXS8036
D]r~l lgS
ISH33)1349 D)r~999 ~ A P M a 1 6 3 tX:l D)~8022
~' AFMa074se9 ~8108 -- A31H[12 0~J~9 Dxsg87
, ~or.q122 9 ""--"-- AFMI64zd4 DY~1053
DXSTI01 ~AFMa282vdl DXS8036 /"I/ DXS8104 //
A~M2341rE12 DX8999 / ~ I:~r~S7592 AFM207Sd6 DXS1195 AFMaI52xfl DXS8019 / ~$1052
.1~1~1989 _
/ EW~80gg //I~XS8027
/ / ~($8047
DY~804g D]LS803 g ~1~ 985 DZ8992 DX81214 D]1~1036 DXSI067 ~[89g? DXS1219
DXTB129 ~ Y B 9 9 ~TYS229Z ~rJD)2145 D~B6796 CD39 DX87107 !~J1374 F1 HIOMT !~JI1145 ! ~ J l 1 3 2 ] ~ r 8 6 0 )DXYB142 DZB1295 ]02~g1354 D~YBIS3 D~Y8144 E~ZB7100 ]~Jr/608 SlIZD1339~[Y8143 ~(XB1388 IffZD10 ]DXE887 DXYB145 ~rJ[D1959 ]0~B7600 ~ Y 8 2 8 ~X~8149 D'&B89 D~J7716 DXYB38 D~£B17 ~ r 7 9 1 5 ~0(Bl148 I)~,ZB131 i~1rB146 9Z81060 D~81138 D2'ZB132 D~BSS I~Jr1402 DD~2B2
4 DZYB130 I ~ [ ~ L 4 7 ]0~rJ;7680 D~87714 ]01YB86 ~[YS148 DXB6746 ~ ex3 D~rBS9 DXTB92 DZ8996 ~ ex9 DZg1309 l:~rB93 1 )~J ;31 DX81223 D~Z8136 D)L~8102 D~B125S D~rJ.407 I~T815 ]0~£B221 ~JSl118~ )0~B1497 i ~ T 8 1 3 3 ] ~ r8223 DZ88095 D~Jr/467 ~ Z T 8 1 3 4 DzlrB104 ~ 7 7 0 3 ]o~Jr7520 DXT8155 D~J1418 DZB1139 ]0~f143 ]~1r7521 )~T8152 ]~ur7517 ~ ! F ' / 7 8 1 ]00[Y8103 ~I1L'BIS1 IWZD2019 MIX]D1189 ~ r 8 1 3 7 D~TB150 D~J;1133 Dr£B91 NIC2R ]0~J278 MrJ[D3037MZC2 STS ]0X'ZB138 I~ 'ZB77 STS D~ZB139 PJkBX/T ST8 DXYB140 PJLE~ Dor.l11130 ~ z B 1 4 1 D]Ur7420 ~J~7731
]~B1375 DYJ~723 DXW/966 ~(Bl19S D~BSS D281434 PZG,t,_P_.~ ]~18184 IIrJDD118SDXBT?27 D ~ 1 2 9 6 !)~;418 1W~1316 ~ r 7 7 3 0 PIGA I)~87994 ])ZB404 ] :~JI7724 D~8207 D~Jr7995 DZB8353 )DX~7741 IW, B7968 ~J ;7996 i~118353 D3~1224 DXW7970 l:l~JrTg97
12 DX81143 ~rJD)1167 ]0~11424 )0~J;b'747 D=~7737 ~:Lq7632 ]~J;6718 l~Jr1661 ]021;7154 DZ88022 I]~Jr1351 )o~r/841 Dxg1137 D~Ji7932 D~J;1292 ~ D ~ 2 ~ 3 4
]0~81313 l)orJrT936 ]0~r7723 sll~D)13&9 DI~ /103 9~B16 11Z81053 )0~r~7155 ]~J187 D~Jr/938 ~,,S7016 9~J;b'~99 D1~410 ~ J 6 9 DZII7972 ]DZII7615 ]DOU;8188 I~B414 l ~ l r l r / D~J;7998 1 ~ B 7 7 0 7 ]~B7940 D~lr1698 DZlI2S7 ~ 8 1 1 3 6 ~ B 6 9 D~J7664 D~,BT999 l~rJr/0 DZB801g D~Jr/330 !~118000 ]D~118318 1)0Ur7943 DxIr1696 DTJ;8001 arJ[D1125£1~g1152 1~U~727 PBKA2 slrJD~13 MIZD1167 DXJ6820 DZ8999 • ~[ELX MILD)431 90Ur7997 MIZI~219 m~18183 ~rJ;1526 9~Jr1461 ]oorJr/176 • ~,J;1443 ]0~r7292 ]0~r797s a~r~D172~ 13~JJ1380 ~Lg9 GR~R_S' s ~ D ~ L 7 2 3 ]~J;7734 l)~irlSOS DorJr1400 ~ D 1 7 2 4 DXB1043 1 ~ B 1 4 5 7 I]0UF7978 J I S X I ~ 8 4 6 ]D2~'7800 DX~J.1S1 ~ K 7 0 9 8 D0rJ;8003 DXB1373 ]00[B7944 ]~JI45 9~18003 )~rJ1135 ~ g 7 9 3 8 (;RPR_3' ) ~ J I 2 6 4
20 ~ 8 7 1 0 9 ))0r~b'710 ~ r 7 9 8 0 P ~ I 1:0rj;1438 ~r, B7769 ~ J 7 9 8 2 IAI=D~757 DZJ1129 G L ~ I ]0~J~7984 MIXD196 DZ~104 D](BTS94 )~,g7985 ]0[][87713 Z~[B7951 CkLB3 DZ8699SE G ~ Y I ~ _ 3 ' iW, B7 9 r l ~ [ ] r 2 2 B 1 DZ8987 Z]~J7988 ][]O[81443 ~1]DDl183 z~Jr1466 ~ZB1390 DUr/SS3 9~Jr7990
t ][~]~7JI4 ]0ZB?gSS ]0~B8352 ]DwIr/6sl D~Jr7957 ]0~Jr1517 PRPS2 I~[JI7960 l)~J~S01 ! ~ B l 1 4 4 1]gji7962 ~ 1 7 7 9 5 9~11144S D~JrTSG3 ]w[BSo$6 DZBT090 PIG~_3' ]o~Jr1391
MIX]~017 i11ZD1124 ][K!11314 ]0Z88004 MIZD1211 ]DZII7788 DZB4&) ]DZJ1516 SFX ][X88176 DZB8354 DEJI1396 U~i;3434 /)~Jlr~S6 ~ d ; / 4 3 3 ]MU;800S n r J I 1 6 8 3 ][~d11198 ZSEqT1 DraB'r722 ~ r J i l l l S Z aommm stuDY1"/38 ]DZB1515 DrJI82b'? DXII8181 DZ88099
28 ]0ZB8238 DZBO257 dlZD20SS ~ J 8 2 6 8 DZ87592 DZB802"7 9Z81229 1)~Jl8178 POT~ IXB7293 ~ 1 0 5 2 D)(88047 D~JQS04 GR8 nZ81048 D~JI8247 DEB82S8 DEJI408 mj i2538 D~B274 i~B782S 1XB8266 DZB8259 ~U1152"1 I X B 7 0 8 8 DEB8243 DZ81202 D~JNI24t8 13Z892 DZS8010 1)~J8249 ~T81226 D)(87106 ]0ZB8007 i~JI8179 DKIM7 B~J;80G7 DDI8177 DX81061 DZB8250 DXB82Ji4 9~Bl149 ]~J;8245 D~;451 ]D~Jll01m lXB365 ~ $ 7 7 7 8 lXB7697 i)~J8008 Um;1404 mW83S1 D l~r74 ~1m265 D~B7710 13~J8253 DZB7656 DXS7101 IW)DDI2265 1~JI7786 DZ8989 D0(B8253 ]0~J1304 ]0~;8242 ~IX]S2003 ~1[S8104 D0rdg7783 l ~ 8 2 6 0 ~(S7480 DXI;/459 1 ~ B 1 3 7 6 • ~g8360 l~J~L464
, ~ B 8 2 5 5 ~nnD~13
F i g u r e 1 (See page 217 for legend.)
21 2 @ G E N O M E RESEARCH
Cold Spring Harbor Laboratory Press on July 15, 2011 - Published by genome.cshlp.orgDownloaded from

11.4
11.3
11.2~
11.22
11.21
11.2
12
X CHROMOSOME YAC/STS MAP
/••/D>Q•8049 E~S8039
DIS992 ]3X~1214 . IZ<~1036 EE~31067 I:D~997 D:<~1219
_
~ , ~ 1 2 ].1r~=9 D ~ 8 0 1 0 ~AR~224zf2 DXS1055 C A I ~ 2 7 7 s b l D X S 8 0 4 9 " A I q ~ 9 5 z a S D X S 8 0 3 9
~ A P M l l 2 z f 2 nKS985 ~APM072zh3 DX31036 ~AFM234~? D ~ 1 0 6 7 ~AFM283wu9 DXS1214
DIS1218 DZS1093 DZ81243 : DX81065 DZ81096 DZ~/840
RDX DZ81097 s t a i D 2 4 4 9 DZS1425 IX~D_EX79 sN][D2450 DXS7699 DIS?26 oWZD2451 DZ828 DIS1411 DZ88240 DI81086 DMD2X66 DZ81014R
~6 DZS1453 DX81214 DZB'/484 DX87181 DZ81417 D Z 8 1 3 8 2 DZ81101 DX81421 DZ81182 DZB1147 DZB1457 DIS7634 DZE7926 ]]ND_EZS9 DZS8102 DX81023 Dll]D_]rJ[56 DXS1049 DZ81017 DX81311 DZBT?20 DX81098 DMD_.EX54 DZB'/?2S I)][81020 DZB'/605 DZ88192 DIS' /2? DIS1036 DZ81371 DXB1025 DZS1067 D][87653 DX8319 DIS99? 8WZD1439 DX88049 D Z ~ 6 3 0 D IJ1370 DXB1514 D~D_46 DZ88190 ~ZB1022 DZ81513 ])ZB6817 _ DXf7783 DZ81219 DX88090 DZBT04 DZ8"/826 DX81385 DZS8039 mllZD3046 DZ88376 sWZD1248 IX4DEI44 DZt~/889 sMXD20041X4D_EX40 DZ81394 DX~r/681 IX~D_EZ33 CY~R sIIIZD2020 SIIZD3039 sWZD3438 DZ8985 D~D_27 I~3 DZ81127 ~DlntlTslIZD$8
-- #WXD2025DMDEX17 DI88113 DZB1029 D~XI9 DX8676 DZ81128 DZBT029~ DZ88014
- DZ~r7657 DMDEX8 DZ81068 DX8992 DMD_EX7 m11rJ[D3432 DZ81018 DND~X3 B11ZD3433 DZ81034 DZB8294 OTC
_ DZ81099 D~D_~X1 OTC
DZ8"/633 DZ88085 ] ~ 8 0 4 rm81267 D~B7611 o11ZD2048 ~XK7455 ZNP21 1)][81409 DXB1428 DX81408 DZ86616 DZ81116E DZB1330 DZB8308 DZ87726 DZ81069 sWZD3100 sift[D2212 ssx 8WZD881 DES6?22 81133)3519 nllZD070. D I88015 DXB6745 lW][])3518 D][B8196 DZSS042 D ~ 8 1 0 D][B1446 DZ88208 DZ82495 DZB1415 sIIZD3194 DZ8746SE DIS1056 DZB228 slrJI])2213 DIB7927B IWZD3103DX86668 slrJ[D3384 DZB722 DZB7922 DX86669 IllZ]D3386 DXB6675 DZSSS6 DZ87705 DZSl003 DXB6674 GAPDPI D~ 8WX]DgS3 D]rJ1358 DZB977 MAOA DXB1264 DZB7466M DXS1058 D][B? DX81055 DXB7467E DZS8025 DX~7700 D~B7662 DX~1011M oIFJ [D3439D][86670 8NZD3223 WASP DXB?S12 DES7152 DZ~1146 GATAI DK81379 MAOA sWZD949 HRASP sWZD3185 DZS8035 DES/290 ] ~ 6 7 6 ~XB1368 sWZD1650 ARAP1 1X#1126 ~ B 6 7 3 1 1)ZB1393 D~17880 D]Ur1240 DXB1293 NDP DKB1266 D]rJI8220 DI8993 D][B538 i]~B337 DXB'I469B DZSS012 DZS8080 TIMP D ~ 1 4 7 0 DZB1392 DZ88026 PFC D][B8227 s113])3036 ~11XD3383 ELKI D][,g8224 DXB1387 DZB819"/ DZI~I26 DEB8221 DIS120? ~ B 8 2 0 7 DZ~!629 I ] ~ J 8 2 2 8 DI81201 ] ~ 6 7 1 Z~41 TlrE3
" D][88~87 #IIZD3297 DZ81004~ D Z 8 1 0 0 ~ DX#7715 ~RE1 DZ81015]1 i~J8225. 811ZD1218 DZB1180~ ]DZB1367 DEBS76 DXB8351 DXB?015B D~Jr1338 SYP
~ 1 2 0 8 l~.31039
DZBg?g5 s l r ~ 1 8 3 ~ 1 5 9 ~ ZXDB D~B573 sl lZD2106 ~ᬬ D~B14 • X81331 ACTL1 ]D~J~77~r7 ] ~ 3 9 0 • X88226 slrJ[D2893 ~ 2 7.~) , D~B1522 D~rJ1272~ sWXD2896 ~ S l
~0 ]~[88223 DZS1199 DKB6954 D~88366 DZSl000 mlrxD2793 DZS1039 nllZD2894 DE~6951 D~B1426 #11ZD2895 DZS991 D~B1416 ] ~ 9 5 3 D ~ 6 7 5 DXK7671 s11ZD2792 D Z ~ A 4 5
r ~ 1 2 1 3 I I / ~ = 1 - -
/ / ~ ~ IIX111~16
D X S 1 0 0 8 E ] ~ 4 DX~6963 • lr£D1118 DZ88032 M~SD~DLI
AFMa323ZhlDXS8040
DXB8193 DZ8988 DXf1302 hal:B7647 :11ZD2184 DXB7828 D ~ 2 2 2 DX#423B sll~KD2794 DEB255 DZB1013E !]XB6964 - 81rXD1434 8 1 r ~ 1 8 5 DXB6961 DX88023 ~ f 4 2 3 E D]~16960 DZS8024 DES7613 ~ 4 2 9 DXB146 ~ 6 6 6 DElfT41 8 1 i r ~ 4 3 5 ] ~ r ~ 9 5 8 DKB741 DE88307 o11ZD2023 DZm6959 D ~ 8 1 3 3 4 mlIXD2108 sll:[D;r795 DZ86725 Dl~6962 D ~ 9 5 2 DX87900 DZ81204 DZB579 DX81184 DZS1044 DZIr341rl OATL2 DES6968 DZS1190 SSI FGDI IH~8580 8rXD2560DZ869S7 D]rJ429 DZB6672EDEB6965 DES8219 sIIZDX791DKB69SS DEg7622
mlrXD334?DZBl117E nl lZD3358 o l r lD3240 sIIZD3350 mlr£D3360 !~ i r7086 JHZD607 mllZD236 DKB1312 DQ~B1213 AR : I r ~ 3 4 8 1 ] E B 1 1 5 9 AP.Xxon4 : l rJ [D3349DZB1194 ARS 'E I1 D~Bg00 DTJrl AR3'0TR a~rJD)1261U~Ji1161 MIZD3361 arJ [D119SDXf1412 IArJ[D$362 DEIr7152 slrJ[D3356 :EZD336"3 DZB136 : l rxD$357 DZ86947B DXS8029 slrZD1967 DXJr1160
Figure 1 (Continued)
GENOME RESEARCH @ 21 3
Cold Spring Harbor Laboratory Press on July 15, 2011 - Published by genome.cshlp.orgDownloaded from
NAGARAJA ET AL.
13
21.1
21.2
21.3
~ _ . , , ~ : : : ~ ~ 4 8 t e 9 DE~1000 AET~48wE9 D:,~1199
|
J ~ AWMa323x~l D:~8040
9 ~ 8 0 3 0
2 ¸ '
ID~1217
D>~1203 D~S990
~s~5 ~ 8 0 3 4 , ! ~o 1 D~SS020,
~ 5 D~SS0(;3 5 ~ 1 ~ 8 1 0 0 5sb9 ~ 8 0 ' 7 5
D ~ 8 0 3 1 D~51275 D~S1216 ~ 9 8 3 D~$8107 D~38052
~ 8 1 0 1 D ~ 8 0 4 6
D>~$8070 D>~8037
PGKIP1 sW~D539 s r £ D 7 5 4 I L 2 R G _ ~ 5 sW][D3364 D IS8031 D] [87625 I L 2 R G , IN~ a l l ' D 3 3 6 6 DZ81275 mWZD513 IL2RG DZ8908 sW'XD3344 DrJ;7678 DZ87118 DZ8897 DZS8357 DZ8983 G~B1 DzBg05 DZ81410 DZ81399 DZ87117 8WZ] )3368DZ88360 D=[B7111 l)Z~r1500 DZ82493 DXSI216 sWZ])5S5 CCGI sW]D)3369~J~1451 D~J1291 D~81499 DZB7838 EDA DT~7112 DZB7119 D Z 8 / 1 8 8 DZ88231 ~ 0 2 1 1 t ~ Z B l 1 2 5 DZ~106 D~B348 DZ8981 sIIZD633 DZS8107 EPLG2 DZ81325 ] ~ B 1 3 7 2 ~01813S slrJD)636 DXSB052 DZB7459 SWZI~34 DZB7116 DZB159 sWZ~635 DZB?114
D~S8089 ~C~S8034 ~ 2 ~ 8 0 2 0 ~ 8 0 9 6
~ 1 1 0 6
D>~1191
- ~ DZ81111 DLg1422 DZ87115 -sWZD905 ~ Z 8 3 3 9 ,, DZB7845 D r J 7 1 1 3
" ~ 8 0 9 2 ~ DZS8101 X IST PGgl 1)][8169 DZ8559 DZ88235 sWLD819 Dzsr7612
1 ~ 8 0 6 0 Dz81517 DZB1681 DZB?497 Dr.q73 ~ 8 0 6 6 ~Z87123 ]~JT128 ATPTA #lrJ[D3400
s l r JD)3332DZ88235 DZB1494 DZB1124 D~J31682 9ZS6980H sW][D2268 DZSS03? D ~ 7 8 0 2
76 DZ88046 DZSS092 81rLD1194 DZB2491 DXS1221 s i reD1488 DZB?706 sWZD2891DZ87014R
* D ) ~ 8 0 8 2 ~Z86724 DZ81680 DZ856 " ~ 1 2 2 5 RPS4X DZ88060 Dgg7160
a i leD1251 DXS8066 DX81463 PHKA1 D]LgT025m DZB1518 DZ81163 DZBl103R 1 ) ~ 6 4 9 ~g81164 X P C T DZ88082 DZ81377 s113])2230 DZ81225 8WXD2053DZ81_383 D][B1447
~DXB986 sm~fD307SDrJ325 sw3~2890 w = x : s 7 n x ~ s o s m s 3 s e v m c x
' D~S995 ~ DZ8227 ]D~J~L50X eW]CD3478 EC~8076 Dxs8oTo m t , l Dxs986
sWZD1217 DTa1502 s i reD2144 DZS l16S DZ8346 slIZDX287 Dz~r1676 ~ [ 8 4 4 1 DZ8447 DZB227? D r J 1 3 9 7 DX81197 DxI r1678 s r J ] ) 2 0 0 5 D r J 3 4 6 DZ81677 D r J 8 3 8 0 D r J 1 4 2 0 DZB1679 8WZD312 D I J 7 6 9 5 D Z 8 1 0 0 5 E D Z B 1 4 3 0 s l r£D1421 811ZD1219DZB6983B DZIr7703
- - 84 s l iT ] )1423 CHM DZ81227 s I I Z D 1 4 2 2 D Z B 7 1 2 9 DZ81455 811][i)1432 CH~. DZ88355 s l r~D1419 C ~ Dz~r7689
D ~ 1 0 0 2 sl13]YJ.428 DX81002 s l r£D2848 D>~1209 s l l Z D / 8 2 9 D Y J i 1 3 5 0
DZ826 D r~7785 ~ 8 1 o 9 sw]D)2597 Dz~1168 ~ 1 2 2 2 s l r ~ D 2 5 9 9 D Z B 3 4 9 D>~1050 ATGR2-5' DZB95 D>~I066 DZ8995 DZBg5
PODV DXS8109 DXSS076 DX~364
- - 811ZD2880 DX81222 DZB233 ~ 1 1 6 9
1 8 W ] [ ! ) 2 2 4 7 ~ 6 6 5 1 DZ81519 DZSl050
. z m 2 1 ~ 7 2 1 ~ 1 2 1 7 I~ 1X8326 81tT])28S5
! \ ~ = m l e 7 D z m o e 6 I ~ n : s e 9 9 t z m x o e 6 ! ~ n z m s o 3 ~ z m l o
J ~ ~ S 3 5 5 i ) ] [8262 i ~ nzs'Tse Dzs1196 I_ ~ z .F6 . x s 2 s o o
- - ' ! ~ Dxs12o9 . r ~ s o 5 I ~ n x m e ~ 2 ~=,s~t2 i • Z : S T e O , ,, , m : 3 o 2 5
- x m s ~ x . n s s ~ e e ~ ~=~7~23
D>~990 DZB7639 ~11ZD1358 DX~8374 ]D][B7819 ]~J~7701 1~J~7643 1~83 1~88234 1~[B7638 l~f~B119 mlrXD2563 D r~8217 DZ l~ r lZ DXT8120 ~ [ D 2 5 6 7 =lrJ[D712 s l l z l r 1352 81r~D2281 DZ81203 1~J~174 D Z I ~ 2 ~XB?620 ! ~ 8 1 0 6 DX8122 DZ~B10? D r ~ 2 1 4 1 ]Z~/552 DZ87807 a l lZD1288 8WZD2996 DZT86 , i ~ 8 1 3 9 8 I ~ 8 5 ~ZB7616 ] ~ 7 1 4 2 IW~7607
~ 8 0 7 7 DZB7683 s i reD2057 ~X~B69 DZ~8077 ~ 1 2 3 1 D~B8354 DY8254 DIS990 ~XB6740
D]L'Y8112 DZS7645 slrJ[DSS7 DZ86728 Dg~r7688 DZT8124 DX81171 i ~ B 6 8 0 9 DZ87609 DZ87686 mIIZDS73 DZ86"789 ~ 1 0 9 D r ~ 7 6 4 4 1)Z~7142 DI81231 I ~ Z B 1 0 DZ1~121 DZT81 " sWZ~1285 I ~ 8 7 6 4 0 ] ~ B 1 4 9 5 81r~D2566 D r ~ 8 2 3 3 ~ B 7 6 8 7 D1~255 sWZD1260 ~ B 7 8 1 8 D r ~ 8 1 1 0 D ~ 7 6 5 0 DZB1170 DTJ~890
100 ] ~ ' £ B l l l ]~[T~9 D~BI257 1W3])1479 D r Z ~ 1 1 5 D~ r1468 slrJ[D2564 1X1~323 ~:H~7623 DX~B122 D r ~ 4 5 4 D r ~ 4 4 2 ~ '11 r114 ] ~ S ~ 6 8 4 oNZD1188 1XB7735 ~ r ~ B 1 1 3 ~ B 1 2 3 D~B1465 DZYr116 ~ 1 2 1~[B7146 D r Z 8 2 D r ~ 7 7 0 4 DZB1395 D ~ r 8 1 1 7 DZ~8128 ~ J 1 5 1 0 i ~ 8 7 6 4 1 sW3])1308 01r~)2571 i)ZT/r118 D g ~ 4 • ]~r~458 ! ~ r 7 6 2 4 ] ~ B 8 3 7 5 ~ J 1 4 9 6
Figure 1 (Continued)
214 @ GENOME RESEARCH
Cold Spring Harbor Laboratory Press on July 15, 2011 - Published by genome.cshlp.orgDownloaded from

X CHROMOSOME YAC/STS MAP
22.1
22.2
22.3
23
24
i i
. . . . .
=
K l ~ 2 6 3 u ~ 1 ~ 1 1 0 6 n n ~ - - - ' ~ 3 ] ~ i 3 3 7 s b l D)~S1230 ~\
"~. ~ 0 5 1 E 0 5 D:~S8096 ~ \ 1 0 i ~ A n S : ~ a 3 v ' b 5 D>mS048 ,~ \
• I ~ , ~ 3 3 t l b 3 1 8 ~ d ~ S I ~ 8 0 ~ 3 ~ ~
l 1.8i-------. s r s . o e . v ~ s ~ 8 ~ o ~
3 ~ N 3 2 8 x b l DX~8088 ,~
~ 013tlrd5 ]D:~8081 ~FM302xo9 D:~S1220
3 ~ ? . 9 1 x a 5 D ~ 8 0 5 3 \
D~91001,
~ 1 2 8 0 ~ 5 D>2L~.12
D:~8057,
DZ81441 ]DZB7617 COL4A5 5' B'I'K " 8WZD2842 D] [8137- G][,A DZ887 DX87886
- - DZS8089 D,T.B17 COL4AS.16 DZ8178 ][~B7621 COL4AS~18
- DXB8034 D][B1427 DZ87815 DX88020 DZB7799 811ZD2153 DZB1179~ DXB8112 D][111-433
~ S 1 2 0 3 DZB7610 D][B8236 DZ87851 ]~D~g990 D][B'JL01 011Z])2007 8WZD122
DZ~8096 D][B472 D][B7887 D][82490 D][B1120 C O L 4 ~ 5 3 ' DZ81106 :WZD2780 81rJ[D2542 01ir]D)1440NYCL2 D,T, S1210 DZ87693 slrJ[D'J.365 D][B8211 DX87007EDZB1504 oWZD2788 sIII~D2783 slrJ[])2840 ~ [ 8 4 5 6
D:,~8077 " - D][81153 ~ 6 6 3 ]D][B6749 D:~S1231 01rZ])2804 D][B1456 01rJ[Dl187
DZ88350 DZ81332 ]DZ87599 OWZ]D2841 ~EZD1286 ! )Zf~753 DZB7657 DZB7667 D][8287 8W][D2843 DZB1230 011Z])1198 DZB7614 :113~D467 0WJ[])1665 DZB1460 DZ87881 DZB7756 s11Z]D2783DZI~891 ][]ZI~S06 oWXD2789D][.86739 811ZD2961
100 sif t [D3844 DZB?844 8WXD3180 P~P ]DZ87882 DXSS075
I:]X88089 - DZSl191 IN[B571 DZ82506 DX~8034 DZ86734 D][B7789 DZ88048 D~S8020 DX81440 DZ81105 DXS809? D:~S8096 DZ81406 DZ87820
~Z894 ] Z S 7 8 8 3 1:D~1106 D][81348 ]DZ~/884
DZB1509 D][B7885 TBG DXB8216 ]~[81285 COL4A6
DZ87357 DZB8194 DZ87872 ~11ZD1070 I Z ~ 1 1 9 1 DZB7356 ]:0[JI7387 81rJ[D1496 ~lFJ[D768
DX~8112 D :~1230
DX~1105
DD~1210
D>~8075 DX~8048
D:~8097
~ : S 8 0 5 5 I~=S1220 D:~:S1072 ~ : S 8 0 8 8 ~¢~8081 ~ 8 0 5 3 ~ : ~ 8 0 6 4
I \ \ \ \ \ ' D~91001 . L _ _ . . _ _ _ _ ~ a 2 8 8 x d 5 ] : D ~ 8 0 3 8 ~ ~ , , ~
0 ,; ~ l ~ m o 3 = r , . ~ ~:s8o44,,\ \ \ \ \ • " - ! - . ~ ~ 6 ~ S ]~:S120~; ~ \ \ \ \ \
0.9 ~ 0 4 s y ~ s z ~ 8 o 9 3 I ~ ~ ~ ~ . o o , ~ \ \ \ \ o.~F:::~r~.:~ ~s~s \~.~, \ \ \ \
T - - - - - - - - u m s o ~ o ~ : s ~ o ~ \ \ \ ~ \ \ \ \ \ \
o . ~ T ~ : ~ ~ & ~ o ~ i O [ o ~ , \ \ \ \ \ \ \ \ \ \ \ ~ ~ XLItl~35 ~ 2
1 . 0 " 1 " ~ ~ , ~ t ~ 0 . ~ ~ , ~ , ~ ' ~ " ~ ' ~ ~ ~ Z~:~8098
1 . 0 1 ~-- ~ " ~ " ° ' ~ ' ° " \ \ \ \ \ \ %,\ Nrgg"N! ! %\ \ \ \ \ N
- - - - - - - - - - - ~ 7 6 V ~ 9 ED~1211 ][:D~8044 ~ - ~ ]~t 'N19(bral D:X~1192 DX88078
• ~ 4 1 0 5 m o 5 D~:8984
D>~80~2 D:~994
D ~ 8 0 7 1
DZB8239 DZ88088 ZXK4 DXB?032]I D][87770 D z r / ? 4 4 DZB7867 D][B7878 D][B7355 1XB7341 ]Dz]r23883JLANI~2 DZ87012B811ZD3831 DZ81310 DZB7531 DXB?300 sl lZD3125 DZB7309 DZB8314 DX87354 oWZD3117 DZf17308 D Z f / 8 7 9 D][87458 sl lZD3431 81rZD1835 DZII7272 DZ81280 ~ [B424 81rJ[D2181 D Z f / 9 0 4 ]W[B8201 ]~[B7287 811ZD1580 DZ87270 ~ B ' / ~ 4 4 DZB7339 slrJ~D773 DZB7271
08 slrZD1202 :W][D3453 DZB7746 DZIr7331 DZS1059 I]ZB7319 811Z]D770 DZB1001 ~ZB7299 DXB8081 :WZD769 ]~[B7095 DZB7360 DZ86805 ~1'~2 DZB7294 DZB7298 811ZD3116 DZB7340 DZB787? 1X87870 811ZD3118 #11ZD3430 DZB7028]I 0111~D3124D][B8195 D][B7301 =11Z])3138 slrJ[DSX25 DZ81220 mIIZD777 mWZD833 DZSl0?2 DZ88055 ][]ZB7805 D Z f / 5 9 7 DZB7307 DZB7747 ][~JI8224 81rJ[D1342 DZ86804 DZB8397 ][]ZB'/850 DZB347 ][]ZB7595 DZ87343 D][B7868 DXF17583E 1XB7345 ] [ ~ f / 7 4 8 ]DZBT848 D Z f / 3 3 6 DZB7327 :11ZD2525 DZ117277 01rZD1129 9ZB7418 01rZD3136 ][W, J 7 9 0 5 011ZD3313 DZB8399 sIIZD$127 DZB8241 :WZD400 mll fZD3479DZ87310 D Z 8 7 3 3 0 DZB7373 Jl l IZD1992DZS8296 DZS8067 D][B7328 DZE8298 DZII7347 D][B8286 mW][D3314 DZB7896 ]~[B8204 DZ88292 JWZD1949 DZB7895 sIIZ])1262 011ZD3483 mWZD3135 D][B7903 DXS8053 D][B1320 D Z ~ 7
D][87318 1ZB7282 ]DZ87668 mlir][D3086 DZB732S DZ81206 DZIrJL00 slrZD308S ]~[B'/$53 sgZD918 DZ117368 mlgZ])3325
6 DZB7316 DDIS$00 D~JJT?92 sIIZD3264 DZB7361 DZW7456 DZB8262 DZ8994 DZ i7517 ~Zl l7284 DZ118212 D]rs7384 DX88064 DZBS009 ]]][B8212 :1rZD231 DZB7861 D][B7831 DZ87780 :11ZD/781 ]DZB?920 sWZD1291 ][~B729S 8rZD1080 IXB7919 D~JJ8301 I)~JJ73t3 DZB7863 mlrJ[DS?5 DZB7777 D][B?;rT5 DZlrJ.331 DZB7313 D][B7862 1]ZB7363 DZB7297 DZB1305 DZB7296 DZB7415 ][]ZB7918 s113])1292 DZB1046 DZ87285 DZB7362 D][B6 ]DZ88218 DZIi7366 ]38" /359 D][g7276 : l l rZD/541 #IrZD2471 DZ87323 D]rJr/365 9zs8038 81rJG)1840D][B?364 ! ~ J 8 3 9 0 IXB7869 ]DZB8304 ]DZ, g7338 9] [88059 D][BT$67 DZ117659 DZB7921 DZB982 sE1[I)326"7 DZB?324 DZIM811 811ZD3145 1]ZB7335 Dr, BT511 slrJ[D3417 DZB7322 D][B8263 slrZD$134 " DZ87334 DZ87380 I I fZD3329
2A~ DX81213 ellIZD839 !]ZB7332 mWZD3427 sWZ])3369 DZB7529 #W][D$137 #11ZD3337 DZB7333 mllZD413 IW'J[D3270 DZB7628 slrJ[D1749 mlrJ[D3104 DZB7842 D r J 7 3 4 3 D r J 7 8 1 3 DZB7267 l X f / 2 6 3 ][H.8"7337 DZB?SOS DZll7413 1~[ ! )3268 1XB7654 =11ZD3366 D][111122 DZ88098 DFJ~824 D][B7910 D][87265 DZB'/824 D][JI1301 !~[B7871 DZS8093 !]ZJ7865 DZB7864 ]DZ.87288 o11ZD$338
:28 ][]Z88285 mlrJ[D1074 DZB7356 DZB7852 DZB7289 DZS1047 DZB7269 :11ZD1333 DZJ;T911 D Z f / 8 S 5 sl rJ[ ]~932 ]TF, J 7 9 o 9
Figure 1 (Continued)
GENOME RESEARCH @ 21 5
Cold Spring Harbor Laboratory Press on July 15, 2011 - Published by genome.cshlp.orgDownloaded from
NAGARAJA ET AL.
25
26
27
28
0 .7 " \ ]):[8"/866 sWZD1984 DZ87311 0.6: ~ &~ra232~¢515X~8033~ \ DZlr7894 sffZ]D1644 DZ87836
~ - ~.___ ~ ~ = ~ s ~ 8 o 4 x ~ \ . z s ~ 2 s s ~zs777z DZSe=93 DZST=6~ ~ZST8~0 nzs7837
1.0 ~ " " ~ 5 5 x © 9 ~ 8 0 7 4 \ ~ ~ DX8425 D,~lr'/749 .Ir~)3288
~ 0 4 . v ~ 8 0 ~ 4 \ \ \ , ~ \ N ~ 8 0 4 4 _ ~ 1 2 4 nzs787~ swz~3z30 swzn,,~ ~lfl~0?xb8 D~2~1062 DZff7901 DZ8774S slrJ[D3263
~0 a3'J~22xa.1. ]1::0,~.11.04.1. ,,WZD866 D][S'75B 81rJ[:D29S6
_ sWZD?73 SIIXi~954 DZ88261
DIS8057 s~D986 DZ87406
|.7 DZ~/285 I11fZ])1335 81rZD3426 DZS8044 i)~J~246 DZE7676 DIS8078 D][B737 8WXD3136 S~XD1489 sIXD~973 slrJ[D3040
\ \ \ \ ~ \ \ " ~C38072 ~ 128 8WZD3XOS 8WXD3038 IN[B8305
&]JM105xa5 I2~984 \ \ \ \ \ ~% \ \ D0~g994 / DIS8072 DZB?711 DX87503 DZ88363
k 9 . &]f lM144yd2 1 : : C ~ 8 0 1 3 ~ ~ ~ ~ \ D:~38071 . )~B8310 DZ87596 DZ8913 i)Z87444E llrJD)3481 G ~ 2 3 ~ 6 D~16843 DZ~24682~
~B7859 DZ86842 DZ86844 DZB8213 ~ HI)G¥ GPC3_EI5 !)(F~7888 1)X8~897
sWZD3480 DZ87314 DEBT158 81rJ[D1493 DZB7822 ~.'~C3,_E.~3 sWZD2239 !~J;730 ]0018118"/ s1IZD1094 !X86845 DZB294 I~81298 1N33)1165 D~B1322 I1fZD1492
JLl~M265va5 D)G~1205 \ \ \ ~ \ \ ~ ,11ZD2160 ,IfZD/537 D~B7 830 DZS1211 D:~8041 DZ87405 D0rg7315 ~rJi691 i)~r/809
33M31?ye9 ]]x~1227 D)~8033 DZ88071 GI~3_EZ2 DI81009]1 DIS1192 8r£D3429 GPC3.EX1 1)Z86721 DX8102
D:~1041 sNZD3442 DZ86847 alr£D572 !)0187303 D:~8050 DZ87279 I):[81254 DZ87817 DZgT094 ~ 1 0 6 2 " DZ87351 ~[Bl114 DZ81284 F9
DZB1308 DZ87827 DZ88200 slfZDIS09 DZF23781]~XBg07 HIGMI 81rJ[D1115 111187348 8111ZD1307 HIGNI DXS1232 DXB~ ~49 HPRT ]0~J. 256 l]~J7 302 1~8692 111188041 DZB7821 81f)G~462 DZB7811 8WZD1319 DZ87291 D~i8288
136 !~B7350 81rJ[D791 1)~8300 ])0[87352 DZ86848 l)z~rTo87 ])(][87854 01FJ[D1182 DZ~1041 ]~B6744 D~8901 !)EB1319
~ 1 1 9 2 ~[81268 DZ8895 ~B8310 D][BTB54 DZ88074 DZ811191
]D~91232 ]DZB7772 DZS8033 slrJ[D1491 ]00[8"/097 /)0[8902 DZS? 442E DZ87853 DZ81355 DXB8050 DZM748 811)G)2776 DXS1062
,!~'~a074t:f9Z:(X~8106 \ Z:Z:,~984 / DXS984 z)or, s?917 DZf/908 EXf1399 -- D~BT096 !)X87391 sBX3)1923 I)~1r/876
~c~s8013 CDRI ]1~,,B6798 DZ87409 DZS?004]~ ~ o s .~s ~=e? ~ u n ~ T s m m , o ,
~ E ~ = 0 2 5 y b g ~ 8 0 8 4 ~ . ~ slr£D1341 D~81343 DE87382 I~B7846 DZ88013 ~E86738 !)EB1289 D~r/416 I)Z88232 I)Z8"/262 ~E8~383 IX[J~892 8WZD2241 slr£D1217 01rJ[D2242 D0[J'/388
" - - - - - - - - - - - ~ g t ~ l S ~ i g l ] X H 8 0 4 3 ~ ~ ~ D/S1205 D(FJr/635 !)(X82494 l)Z]r/403 sWZD2151 ~EE7902 DZ8/324 ~E87832
----- ' ---" &FMa212xfS[D~8028~ ~ ~ ~ 1~88252 ]DE88309 ])(][117874 DZS1200 i:~38073 !~81341 DZ87381 ~ 4 0 8 DE~I97
I)E88232 l)~rJr/411 l)EIr/4oo EEr/306 D(X:S8084- 144 DZIr/410 DXS8084 !)~r/281 l)EIr/305
]31(81227 alrJ[D864 1)~B6806 aAI]D)1447 D~87402 Dx~r/407 IXB7524 I)~r/834
&-l~Mb041v~912X~8045 I)ZE6312 1X88316 DXS8073 8WJ[!)I091 E1~2 54v~. D : ~ 1 2 0 0 ~ D:~8043 ~8~392 ~ 2 9 5 DIS8043 IW'J[D883
~ 8 0 2 8 DE86751 DEB8295 D~g369 ~EB/$44 1)~.g8319 ]0Ef/l133 DEB369 81rJ[D1449
D:~91200
~ ~ F J ~ 8 7 ~ 5 ~ 1 2 1 5 ~ A3t1224z911D)~998
~ 9 9 8
i ~ 1 2 1 5 &~345t~9 D ~ S S 0 9 1 ~
~D.99vo7 ~ C 3 1 1 9 3 ~
&rd[310vb9 £ C ~ 8 0 8 ~ ]2~s8091~(:~1193 --
]:~:~8086 " ~ ~ 3 3 4 x h 9 ~ 8 0 6 9 ~ 8 0 6 9
&]rJ~s121ze51:C~8011
~ ~ J ~ l ~ 0 6 5 v d g D X : 9 8 1 0 3 ~ ~ D : ~ 8 1 0 3
I ~r~:)3111rg1D:~8061 D:~8061
! k7
198.1 cM
~ ~F~27~b~h9 1:D~1073
-, A F ~ 1 2 z b 9 D>~8087
D>~1073
DX87375 Dz]r/825 DE#8229 ~EB7857 ~B7396 I~[Irl413 811~D575 DZgg98 i~[lr7373 D~8292 DZ88028 D~B532 D:[117579 D~88215 DZ87386 DX88289 1)ZB7395 DZIr/814 Z)ZIr/398 ]XlIS48 ~[B7397 i]X1~377 DZ8740x ~ 3 1 3 ]XB?603 I)Z87414 D1[~/875 DZla.=58 ~ r / 8 9 3 ~E88317 ~ 3 8 5 1)ZI1/337 slrx]oO238 ])(8"/374
~s7~71 =s...=e- ~ I t s m m o 6 1 . z , . ~ s Dzseo6. = s ' , ~ e = , - 7 1 2 = s ~ s s Bxsoxo3 = m s E s T e , s !~87393 ]X11898 DZ86999m ~.g522E ~E8533 DZ8462 D~r/074 FSC !)ZB7847 I)0m1299 BGH FSC DZ81215 i)~Jrl423 I]E87082 ~ 5 3 2
152 DZS809~ n ~ m 3 4 s m u n 4 s 2 n~.sTo3sn • fJI7390 DZS8011 DZ~}00IE IAI~DQ469 1)E81318 i)E81281 ])EE?065 ])El7060 )0EBX286 ])E81278 i)EIr/501 i)E:B'/059 ]~88273 DZm)04 A V P R 2 ~EB7062 DZ82496 ])0[8903 I]E87 064 DZB'/SX2 ] ]Zggl l DZ8707 MIZD2244 1)E87077 1X87069 DE86729 1)E81346 DZ81012E i)E88303 DZ87076 ])E8~260 1)EZ7394 DZ8963R DE81259 l)E882r'/2 D][87445E Dg8/347 IHS457 IZET080 RCP l~JG185 D]Dr/079 Ill.S7070 DXSl193 i)E~BT075 l)E]r/068 ]0E81133 slrJ[D]L913 i)Er/071 IDS 1W/3~;B35 FLNI 9Z869811 811Z]D1785 ]0EB'/0"/2 ~81113 IZ881mS mmlOlO~ DIS962E ~ 3 8~[D2362
l]Em'089 DZS7 01GE ]0ZIr/085 I)(Z8"/073 I)~B7658 • E~W066 i]~JW083 IES7067 D~87081 DZS8061 G6PD
L60 ~J;'JL276 ~ZS'7063 l~,S70ml DIS8086 DZ,~L177 ~ 022E
216 ~J GENOME RESEARCH
Figure 1 (See facing page for legend).
Cold Spring Harbor Laboratory Press on July 15, 2011 - Published by genome.cshlp.orgDownloaded from
X CHROMOSOME YAC/STS MAP
breakpoint or missing in particular deletions (e.g., in Xp22; Ferrero et al. 1995) are consistently found in a contiguous group in the inferred ordered list of STSs of Figure 1. As another example, YACs yWXD1 to yWXD829 in Xq24-qter, derived from an African American, show the same intrinsic content of STSs seen in the region in other YACs, derived from Cau- casians.
Overall, the results also support the fidelity of human DNA sequences cloned into YACs. For ex- ample, in a contig in Xp22.1 (Trump et al. 1996), 54 STSs were ordered across 3.5 Mb by their content in 58 YACs from five collections. The resulting map showed that an STS was negative at a site between two others in only 4 of 400 instances, consistent with 99% faithful recovery of X unique sequences.
At the average 75-kb marker density, the essen- tially universal set of STSs are sufficient not only to evaluate clone quality, but also to screen for alter- native large-insert clones, such as bacterial artificial chromosomes (BACs; Shizuya et al. 1992), as se- quencing substrates. In initial pilot work, for ex- ample, STS-based screening easily recovered mini- mal t i l ing paths of over lapping BACS for five projects of at least 1 Mb.
Recombination Indices Compared to Location
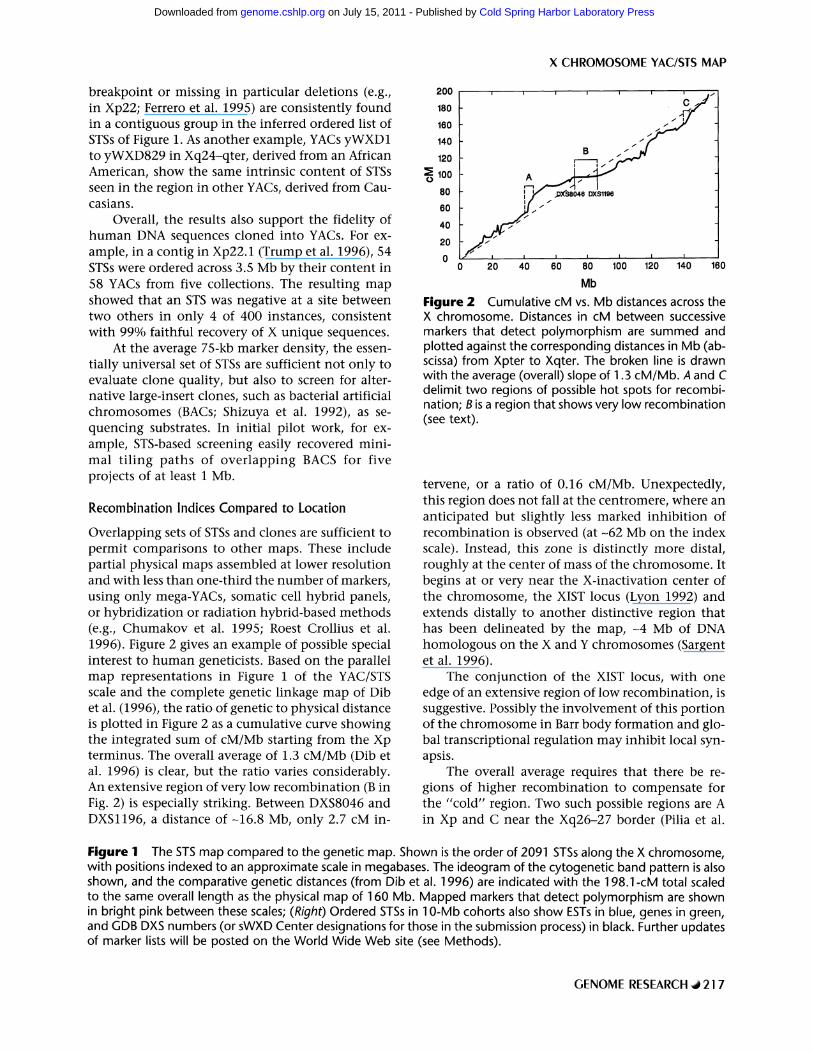
Overlapping sets of STSs and clones are sufficient to permit comparisons to other maps. These include partial physical maps assembled at lower resolution and with less than one-third the number of markers, using only mega-YACs, somatic cell hybrid panels, or hybridization or radiation hybrid-based methods (e.g., Chumakov et al. 1995; Roest Crollius et al. 1996). Figure 2 gives an example of possible special interest to human geneticists. Based on the parallel map representations in Figure 1 of the YAC/STS scale and the complete genetic linkage map of Dib et al. (1996), the ratio of genetic to physical distance is plotted in Figure 2 as a cumulative curve showing the integrated sum of cM/Mb starting from the Xp terminus. The overall average of 1.3 cM/Mb (Dib et al. 1996) is clear, but the ratio varies considerably. An extensive region of very low recombination (B in Fig. 2) is especially striking. Between DXS8046 and DXSl196, a distance o f -16 .8 Mb, only 2.7 cM in-
200
180
160
140
120
~oo 8O
60
40
20
0
i i i i i i i - ' I
/ / -
B - 2 _ ~ - ! I / -
I Y ,oX'sBO4e oxs1196 _ ~ f / I /
~,"/ _
t _
4' i i ' ' ' 0 20 0 0 0 100 120 140 160
Mb
Figure 2 Cumulative cM vs. Mb distances across the X chromosome. Distances in cM between successive markers that detect polymorphism are summed and plotted against the corresponding distances in Mb (ab- scissa) from Xpter to Xqter. The broken line is drawn with the average (overall) slope of 1.3 cM/Mb. A and C delimit two regions of possible hot spots for recombi- nation; B is a region that shows very low recombination (see text).
tervene, or a ratio of 0.16 cM/Mb. Unexpectedly, this region does not fall at the centromere, where an anticipated but slightly less marked inhibition of recombination is observed (a t -62 Mb on the index scale). Instead, this zone is distinctly more distal, roughly at the center of mass of the chromosome. It begins at or very near the X-inactivation center of the chromosome, the XIST locus (Lyon 1992) and extends distally to another distinctive region that has been delineated by the map, -4 Mb of DNA homologous on the X and Y chromosomes (Sargent et al. 1996).
The conjunction of the XIST locus, with one edge of an extensive region of low recombination, is suggestive. Possibly the involvement of this portion of the chromosome in Barr body formation and glo- bal transcriptional regulation may inhibit local syn- apsis.
The overall average requires that there be re- gions of higher recombination to compensate for the "cold" region. Two such possible regions are A in Xp and C near the Xq26-27 border (Pilia et al.
Figure 1 The STS map compared to the genetic map. Shown is the order of 2091 STSs along the X chromosome, with positions indexed to an approximate scale in megabases. The ideogram of the cytogenetic band pattern is also shown, and the comparative genetic distances (from Dib et al. 1996) are indicated with the 198.1-cM total scaled to the same overall length as the physical map of 160 Mb. Mapped markers that detect polymorphism are shown in bright pink between these scales; (Right) Ordered STSs in 10-Mb cohorts also show ESTs in blue, genes in green, and GDB DXS numbers (or sWXD Center designations for those in the submission process) in black. Further updates of marker lists will be posted on the World Wide Web site (see Methods).
GENOME RESEARCH ~ 21 7
Cold Spring Harbor Laboratory Press on July 15, 2011 - Published by genome.cshlp.orgDownloaded from
NAGARAJA ET AL.
1996; Zucchi et al. 1996), with apparent local values of recombinat ion as high as 5 cM/Mb. (A third pos- sible region of high recombinat ion is seen at 115- 120 Mb but falls in a region with relatively few ge- netic markers.)
Regional recombinat ion values can have a con- siderable impact on searches for disease genes. For example, a translocation breakpoint associated with premature ovarian failure was localized between PGK1 and DXS447 (Powell et al. 1994), a distance of 0.1 cM; however, those markers fall in the region with very low recombinat ion values and are >1 Mb apart on the chromosome. In general, the physical map and recombinat ion-based map agree in marker order where distances are appreciable on both maps; and the physical map provides further ordering in- formation for genetic markers in the regions where recombinat ion values are very low.
Estimate of Gene Concentration by GC Levels in STSs
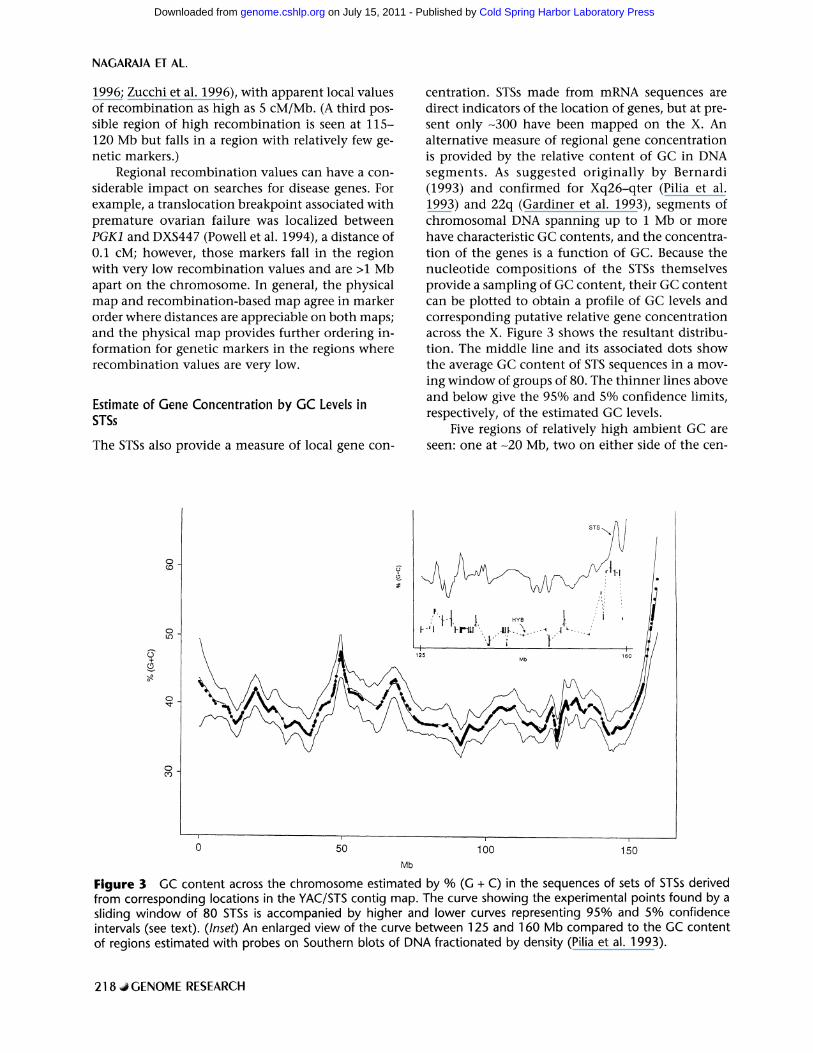
The STSs also provide a measure of local gene con-
centration. STSs made from mRNA sequences are direct indicators of the location of genes, but at pre- sent o n l y - 3 0 0 have been mapped on the X. An alternative measure of regional gene concentrat ion is provided by the relative content of GC in DNA segments . As suggested or ig ina l ly by Bernardi (1993) and confirmed for Xq26-qter (Pilia et al. 1993) and 22q (Gardiner et al. 1993), segments of chromosomal DNA spanning up to 1 Mb or more have characteristic GC contents, and the concentra- t ion of the genes is a function of GC. Because the nucleot ide composi t ions of the STSs themselves provide a sampling of GC content, their GC content can be plotted to obtain a profile of GC levels and corresponding putative relative gene concentrat ion across the X. Figure 3 shows the resultant distribu- tion. The middle line and its associated dots show the average GC content of STS sequences in a mov- ing window of groups of 80. The th inner lines above and below give the 95% and 5% confidence limits, respectively, of the estimated GC levels.
Five regions of relatively high ambient GC are seen: one at -20 Mb, two on either side of the cen-
C::) cO-
~'6" 1_ I
i i 1 1
0 50 1 oo 150 Mb
Figure 3 GC content across the chromosome estimated by % (G + C) in the sequences of sets of STSs derived from corresponding locations in the YAC/STS contig map. The curve showing the experimental points found by a sliding window of 80 STSs is accompanied by higher and lower curves representing 95% and 5% confidence intervals (see text). (Inset) An enlarged view of the curve between 125 and 160 Mb compared to the GC content of regions estimated with probes on Southern blots of DNA fractionated by density (Pilia et al. 1993).
218 ~ GENOME RESEARCH
Cold Spring Harbor Laboratory Press on July 15, 2011 - Published by genome.cshlp.orgDownloaded from

X CHROMOSOME YAC/STS MAP
t romere , one in Xq26.1, and one, especial ly marked , r ising sharply in sub te lomer ic Xq28. Two lines of ev idence suppor t t h e va l id i ty of these es t imates . First, t he prof i le in Xq26 to Xq te r s t r o n g l y re- sembles t ha t f o u n d by an i n d e p e n d e n t assay of GC levels. The inset in Figure 3 compares an en la rged
s e g m e n t of the GC curve f rom Figure 3 wi th assays based on Sou the rn analyses wi th specific probes as- sayed agains t DNA f rac t iona ted accord ing to its GC c o n t e n t by cen t r i fuga t ion to equ i l ib r ium in dens i ty gradien ts (Pilia et al. 1993). The direct assays were d o n e wi th on ly a l imi ted n u m b e r of probes and wi th local izat ion less precise t h a n tha t of Fig. 2; bu t the two inferred d is t r ibut ions are genera l ly concor- dant . Especially clear is the very h igh GC level in the Xq sub te lomer ic region, w h i c h has also b e e n con- f i rmed direct ly by long- range s equenc ing and o the r m e t h o d s (see, for example , C h e n et al. 1996).
A second i n d e p e n d e n t l ine of ev idence comes f rom f luorescent in situ hybr id iza t ion (FISH) of the
h ighes t GC fract ion of h u m a n DNA (the "H3" iso- chores; Bernardi 1993) to m e t a p h a s e c h r o m o s o m e s . Such expe r imen t s have de tec ted four of the p e a k s m one in t e lomer ic Xp and th ree in Xq. High GC seg- m e n t s in the one region u n d e t e c t e d by FISH, near the c e n t r o m e r e in Xp, have also b e e n d e m o n s t r a t e d direct ly by long- range s equenc ing (G. Nyakatura , M. Platzer, B. Drescher, H. Hel lebrand, D. Schindel- hauer , A. Meindl , and A. Rosenthal , pers. comm. ) . Fur the rmore , a very similar d i s t r ibu t ion has b e g u n
to emerge f rom the p l a c e m e n t of ESTs a long the X c h r o m o s o m e (Schuler et al. 1996). Thus, groups of STSs provide a reliable sample of the GC c o n t e n t of u n i q u e sequences in di f ferent po r t ions of the X.
In the s equenc ing phase of g e n o m e analysis, strategies m a y be adapted especially for regions of h igh GC, which tend to show compress ions in se- quenc ing and of ten con ta in h igh levels of Alu se- quences (Chen et al. 1996), or for regions of low GC, which m a y conta in a n u m b e r of long, nearly identical LINE sequences (Hood et al. 1995). Similarly, the analysis of regions of h igh or low GC requires software wi th differential sensitivity to detect CpG islands or putat ive exons, and a region tha t shows low recombi- na t ion m a y cue an analysis to look for any correlated
"suppressing" sequences. It will be of interest to com- pare the locations of ho t or cold spots of recombina- t ion and h igh or low GC levels on au tosomes for which maps reach comparable STS density.
METHODS
STSs and YACs
STSs and YACs are described in detail both in the Genome
DataBase (GDB) and on the World Wide Web site at http:// genome.wustl.edu/cgm/cgm.html; STSs that are not yet as- signed DXS numbers are identified by their Washington Uni- versity Genome Center accession (sWXD) numbers. All YACs have been deposited at the American Type Culture Collection (ATCC). Contigs include 5423 YACs from seven libraries (Na- garaja et al. 1994), supplemented with 569 cosmids from the X-specific Lawrence Livermore Laboratory collection and a growing number of BACs, currently totaling 250. The YACs comprise 1360 and 829 clones from two X-specific libraries [F and X3000 (Nagaraja et al. 1994), with an average size of 240 kb]; 993 and 1153 clones from two collections made from cell lines containing multiple X chromosomes (E and I, with an average size of 330 kb); 290 clones from the CEPH mega-YAC library [(Albertsen et al. 1990), with an average size of 1014 kb], and some additional clones from the Imperial Cancer Research Fund (ICRF) (Larin et al. 1991) and original St. Louis (Nagaraja et al. 1994) YAC libraries.
Strategy and Contigs from STS Content Mapping
The original formulation of STS content mapping (Green and Olson 1990; Green and Green 1991) was based on initial PCR- based screening of YAC libraries with a series of randomly generated STSs to yield substantial contigs across most of the area of a chromosome. As coverage becomes extensive and new random STSs increasingly fall within already mapped re- gions, directed walking is initiated with STSs (pseudoends) that map nearest the ends of nascent contigs. In this project, a modified all-walking approach was adopted in which STSs were made from YAC insert ends, and a series of walking steps at many points was then instituted and reiterated (Kere et al. 1992) until contigs merged across the entire X chromo- some. This strategy was theoretically more efficient (Palazzolo et al. 1991; Kere et al. 1992) and was facilitated by the avail- ability of two collections of X-specific YACs to initiate STS development and the merger of seed contigs (Nagaraja et al. 1994).
Contigs were aligned and oriented by a combination of FISH (Srivastava et al. 1995) and published marker orders and deletion panels. The somatic cell hybrid panel used hamster cell lines containing an entire X (GM06218C) or defined frag- ments of X chromosomes [Xp21.2-qter (GMl11713); Xp11.2-qter (GM10501); Xq13-qter (GM10095); Xq21.3-qter (GMl1172); and Xq24-qter (X3000.1)].
The current 13 contigs, with their bracketing markers, index positions roughly in megabases, and relevant references to some of the significant partial published versions (in addi- tion to Schlessinger et al. 1991 and Nelson et al. 1995), are as follows, from Xpter to Xqter: Xpter-DXS6667 (0-52 Mb; Cof- fey et al. 1992; Monaco et al. 1992; Alitalo et al. 1995; Black et al. 1995; Ferrero et al. 1995; Ried et al. 1995; Trump et al. 1996; Boycott et al. 1996); OATL2-centromere (52-58.5 Mb; Miller et al. 1995); centromere; DXS62-DXS1 (62-63 Mb); DXS1-DXS1225 (63-79 Mb; Villard et al. 1995); DXS447- DXS26 (79-84 Mb; van der Maarel et al. 1995); DXS995-TBG (84-106 Mb; Vetrie et al. 1994; Forbes et al. 1996, Sargent et al. 1996); DXS7667-COL4A5 (107-109.5 Mb; Srivastava et al. 1995); DXS1210-DXS287 (109.5-113 Mb); DXS1059- DXS8088 (113-118.5 Mb); DXS7308-DXS7872 (118.5-120 Mb); ANT2-DXS7317 (120-124 Mb); DXS8081-DXS1206 (124-131 Mb); and DXS1339-Xqter (131-160 Mb; Palmieri et al. 1994; Rogner et al. 1994; Pilia et al. 1996; Zucchi et al. 1996).
GENOME RESEARCH @ 21 9
Cold Spring Harbor Laboratory Press on July 15, 2011 - Published by genome.cshlp.orgDownloaded from

NAGARAJA ET AL.
Use of SEGMAP
STS content mapping was sustained and directed by the ap- plication of the SEGMAP program (developed by C. Magness and P. Green, available from [email protected] ington.edu). The most probable marker order was inferred from YAC/STS content (Green and Green 1991) with the pro- gram. In the middle game of mapping, contig formation and software representations of maps were very sensitive to false positives, which often joined distant portions of the X chro- mosome; but these were later sorted out by additional clones and cross-testing of STS content.
SEGMAP also facilitated the integration of outside data, based on the screening of published STSs to identify their location in relation to existing contigs. The comparative lo- calization was extended to infer relative genetic and physical distances, as in Figures 2 and 3.
ACKNOWLEDGMENTS
We are indebted to many collaborators in the X community for cooperative mapping interactions, including the provision of auxiliary STSs, YACs, and cosmids. Our major collaborators include G. Rappold (XpPAR); G.B. Ferraro, A. Ballabio, D. Nel- son, T. Alitalo, and D. Trump (Xp22); K. Boycott and T. Bech- Hansen (Xp11.3-p11.4); A. Miller and H.F. Willard (the peri- centromeric region); P. Stanier and F. Cremers (Xq21.3); and especially I. Zucchi, G. Palmieri, G. Romano, and the group of M. D'urso (Xq24-qter); Jean Weissenbach and Jeff Murray (simple sequence repeat markers and genetic map informa- tion prior to publication); and GDB (additional STS informa- tion). This work was supported by GESTEC (grant HG00201) from the National Center for Genome Research.
The publication costs of this article were defrayed in part by payment of page charges. This article must therefore be hereby marked "advertisement" in accordance with 18 USC section 1734 solely to indicate this fact.
REFERENCES
Albertsen, H.M., H. Abderrahim, H.M. Cann, J. Dausset, D. LePaslier, and D. Cohen. 1990. Construction and characterization of a yeast artificial chromosome library containing seven haploid human genome equivalents. Proc. Nat. Acad. Sci. 87: 4256-4260.
Alitalo, T., F. Francis, J. Kere, H. Lehrach, D. Schlessinger, and H.F. Willard. 1995. A 6-Mb YAC contig in Xp22.1-p22.2 spanning the DXS69E, XE59, GLRA2, PIGA, GRPR, CALB3, and PHKA2 genes. Genomics 25: 691-700.
Bernardi, G. 1993. The human genome organization and its evolutionary history: A review. Gene 135: 57-66.
Black, G.C.M., M.P. Coleman, Z.Y. Chen, A.H. Nemeth, K.E. Davies, and I.W. Craig. 1995. A bidirectional YAC walk from Norrie disease (NDP) locus. Genomics 25: 644-649.
Boycott, K.M., G.R. Halley, D. Schlessinger, and T. Bech-Hansen. 1996. A 2-megabase physical contig incorporating 43 DNA markers on the human X
chromosome at p11.23-p11.22 from ZNF21 to DXS255. Genomics 33: 488-497.
Burke, D.T., G.F. Carle, and M.V. Olson. 1987. Cloning of large segments of exogenous DNA into yeast by means of artificial chromosome vectors. Science 236: 806-812.
Chen, E.Y., M. Zollo, R. Mazzarella, A. Ciccodicola, C. Chen, L. Zuo, C. Heiner, F. Burough, M. Repetto, D. Schlessinger, and M. D'Urso. 1996. Long-range sequence analysis in Xq28: Thirteen known and six candidate genes in 219.4 kb of high GC DNA between the RCP/GCP and G6PD loci. Hum. Mol. Genet. 5- 659-668.
Chumakov, I.M., P. Rigault, I. Le Gall, C. Bellanne-Chantelot, A. Billault, S. Guillou, P. Soularue, G. Guasconi, E. Poullier, I. Gros, et al. 1995. A YAC contig map of the human genome. Nature 377" 175-297.
Coffey, A.J., R.G. Roberts, E.D. Green, C.G. Cole, R. Butler, R. Anand, F. Giannelli, and D.R. Bentley. 1992. Construction of a 2.6-Mb contig in yeast artificial chromosomes spanning the human dystrophin gene using an STS-based approach. Genomics 12: 474-484.
Collins, F.S. 1995. Positional cloning moves from perditional to traditional. Nature Genet. 9: 348-350.
D'Esposito, M., A. Ciccodicola, F. Gianfrancesco, T. Esposito, F. Flagiello, R. Mazzarella, D. Schlessinger, and M. D'Urso. 1996. A synaptobrevin-like gene in the Xq28 pseudoautosomal region undergoes X inactivation. Nature Genet. 13: 227-229.
Dib, C., S. Faure, C. Fizames, D. Samson, N. Drouot, A. Vignal, P. Millasseau, S. Marc, J. Hazan, E. Seboun, M. Lathrop, G. Gyapay, J. Morissette, and J. Weissenbach. 1996. A comprehensive genetic map of the human genome based on 5,264 microsatellites. Nature 380: 152-154.
Ferrero, G.B., B. Franco, E.J. Roth, B.A. Firulli, G. Borsani, J. Delmas-Mata, J. Weissenbach, G. Halley, D. Schlessinger, A.C. Chinault, H.Y. Zoghbi, D.L. Nelson, and A. Ballabio. 1995. An integrated physical and genetic map of a 35 Mb region on chromosome Xp22.3-Xp21.3. Hum. Mol. Genet. 4: 1821-1827.
Forbes, S.A., L. Brennan, M. Richardson, A. Coffey, D.R. Bentley, S. Mumm, G.E. Moore, and P. Stanier. 1996. Refined mapping and YAC contig cloning of the X-linked cleft palate and ankyloglossia locus (CPX) including the proximal X-Y homology breakpoint within Xq21.3. Genomics 31; 36-43.
Freije, D. and D. Schlessinger. 1992. A 1.6-Mb contig of yeast artificial chromosomes around the human factor VIII gene reveals three regions homologous to probes for the DXS115 locus and two for the DXYS64 locus. Am. J. Hum. Genet. 51" 66-80.
Freije, D., C. Helms, M.S. Watson, and H. Donis-Keller. 1992. Identification of a second pseudoautosomal region near the Xq and Yq telomeres. Science 258; 1784-1787.
220 @ GENOME RESEARCH
Cold Spring Harbor Laboratory Press on July 15, 2011 - Published by genome.cshlp.orgDownloaded from

Gardiner, K., B. Aissani, and G. Bernardi. 1993. A compositional map of human chromosome 21. EMBO J. 9: 1853-1858.
Goffeau, A., B.G. Barrell, H. Bussey, R.W. Davis, B. Dujon, H. Feldmann, F. Galibert, J.D. Hoheisel, C. Jacq, M. Johnston, E.J. Louis, H.W. Mewes, Y. Murakami, P. Philippsen, H. Tettelin, and S.G. Oliver. 1996. Life with 6000 genes. Science 274: 546; 563-567.
Green, E.D. and P. Green. 1991. Sequence-tagged site (STS) content mapping of human chromosomes: Theoretical considerations and early experiences. PCR Methods Applic. 1: 77-90.
Green, E.D. and M.V. Olson. 1990. Chromosomal region of the cystic fibrosis gene in yeast artificial chromosomes: A model for human genome mapping. Science 250: 94-98.
Hood, L., L. Rowen, and B.F. Koop. 1995. Human and mouse T-cell receptor loci: Genomics, evolution, diversity, and serendipity. Ann. N.Y. Acad. Sci. 758: 390-412.
Kere, J., R. Nagaraja, S. Mumm, A. Ciccodicola, M. D'Urso, and D. Schlessinger. 1992. Mapping human chromosomes by walking with sequence-tagged sites from end-fragments of yeast artificial chromosome inserts. Genomics 14: 241-248.
Larin, Z., A.P. Monaco, and H. Lehrach. 1991. Yeast artificial chromosome libraries containing large inserts from mouse and human DNA. Proc. Natl. Acad. Sci. 88: 4123-4127.
Li, L. and D.H. Hamer. 1995. Recombination and allelic association in the Xq/Yq homology region. Hum. Mol. Genet. 4 : 2 0 1 3 - 2 0 1 6 .
Lyon, M.F. 1992. Some milestones in the history of X-chromosome inactivation. Annu. Rev. Genet. 26: 17-28.
Miller, A.P., K. Gustashaw, D.J. Wolff, S.H.J. Rider, A.P. Monaco, B. Eble, D. Schlessinger, J.L. Gorski, G.-J. van Ommen, J. Weissenbach, and H.F. Willard. 1995. Three genes that escape X chromosome inactivation are clustered within a 6 Mb YAC contig and STS map in Xp11.21-p11.22. Hum. Mol. Genet. 4: 731-739.
Monaco, A.P., A.P. Walker, I. Millwood, Z. Larin, and H. Lehrach. 1992. A yeast artificial chromosome contig containing the complete Duchenne muscular dystrophy gene. Genomics 12: 465-473.
Nagaraja, R., J. Kere, S. MacMillan, M.W.J. Masisi, D. Johnson, G. Halley, B.A. Molini, K. Wein, M. Trusgnich, B. Ebel, B.H. Brownstein, B. Railey, and D. Schlessinger. 1994. Characterization of four human YAC libraries for clone size, chimerism, and X chromosome sequence representation. Nucleic Acids Res. 22:3406-3411.
Nelson, D.L., A. Ballabio, F. Cremers, A.P. Monaco, and D. Schlessinger. 1995. Report of the sixth international workshop on X chromosome mapping 1995. Cytogenet. Cell Genet. 71: 307-342.
X CHROMOSOME YAC/STS MAP
Olson, M.V., L. Hood, C. Cantor, and D. Botstein. 1989. A common language for physical mapping of the human genome. Science 245: 1434-1435.
Palazzolo, M.J., S.A. Sawyer, C.H. Martin, D.A. Smoller, and D.L. Hartl. 1991. Optimized strategies for sequence-tagged-site selection in genome mapping. Proc. Natl. Acad. Sci. 88: 8034-8038.
Palmieri, G., G. Romano, A. Ciccodicola, A. Casamassimi, C. Campanile, T. Esposito, V. Cappa, A. Lania, S. Johnson, R. Reinbold, A. Poustka, D. Schlessinger, and M. D'Urso. 1994. YAC contig organization and CpG island analysis in Xq28. Genomics 24: 149-158.
Pilia, G., R.D. Little, A. Aissani, G. Bernardi, and D. Schlessinger. 1993. Isochores and CpG islands in YAC contigs in human Xq26.1-qter. Genomics 17: 456-562.
Pilia, G., S. MacMillan, R. Nagaraja, S. Mumm, J. Weissenbach, and D. Schlessinger. 1996. YAC/STS map of 9 Mb of Xq26 at 100 kb resolution, localizing 6 ESTs, 5 genes, and 32 genetic markers. Genomics 34: 55-62.
Powell, C.M., R.T. Taggart, T.C. Drumheller, D. Wangsa, C. Qian, L.M. Nelson, and B.J. White. 1994. Molecular and cytogenetic studies of an X; autosome translocation in a patient with premature ovarian failure and review of the literature. Am. J. Med. Genet. 52: 19-26.
Ried, K., A. Mertz, R. Nagaraja, M. Trusgnich, J.H. Riley, A. Anand, H. Lehrach, D. Page, J.W. Ellison, and G. Rappold. 1995. Characterization of a YAC contig spanning the pseudoautosomal region. Genomics 29: 787-792.
Roest Crollius, H., M.T. Ross, A. Grigoriev, C.J. Knights, E. Holloway, J. Misfud, K. Li, M. Playford, S.G. Gregory, S.J. Humphray, A.J. Coffey, C.G. See, S. Marsh, et al. 1996. An integrated map of the human X chromosome. Genome Res. 6: 943-955.
Rogner, U.C., P. Kioschis, K. Wilke, W. Gong, E. Pick, A. Dietrich, U. Zechner, H. Hameister, A. Pragliola, G.E. Herman, et al. 1994. A YAC clone map spanning 7.5 megabases of human chromosome band Xq28. Hum. Mol. Genet. 3:2137-2146.
Sargent, C.A., H. Briggs, I.J. Chalmers, B. Lambson, E. Walker, and N.A. Affara. 1996. The sequence organization of Yp/proximal Xq homologous regions of the human sex chromosomes is highly conserved. Genomics 32: 200-209.
Schlessinger, D., R.D. Little, D. Freije, F. Abidi, I. Zucchi, G. Porta, G. Pilia, R. Nagaraja, S.K. Johnson, J.Y. Yoon, A. Srivastava, J. Kere, G. Palmieri, A. Ciccodicola, V. Montanaro, G. Romano, A. Casamassimi, and M. D'Urso. 1991. Yeast artificial chromosome-based genome mapping: Some lessons from Xq24-q28. Genomics 11: 783-793.
Schuler, G.D., M.S. Boguski, E.A. Stewart, L.D. Stein, G. Gyapay, K. Rice, R.E. Whie, P. Rodriguez-Tom6, A. Aggarwal, E. Bajorek, et al. 1996. A gene map of the human genome. Science 274: 540-546, 547-563.
GENOME RESEARCH @ 221
Cold Spring Harbor Laboratory Press on July 15, 2011 - Published by genome.cshlp.orgDownloaded from

NAGARAJA ET AL.
Shizuya, H., B. Birren, U.J. Kirn, V. Mancino, T. Slepak, Y. Tachiiri, and M. Simon. 1992. Cloning and stable maintenance of 300-kilobase-pair fragments of human DNA in Escherichia coli using an F-factor-based vector. Proc. Natl. Acad. Sci. 89: 8794-8797.
Srivastava, A.K., T. Featherstone, K. Wein, and D. Schlessinger. 1995. YAC contigs mapping the human COL4A5 and COL4A6 genes and DXS118 within Xq21.3-q22. Genomics 26: 502-509.
Trask, B., G. van den Engh, R. Nussbaum, C. Schwartz, and J. Gray. 1990. Quantification of the DNA content of structurally abnormal X chromosomes and X chromosome aneuploidy using high resolution bivariate flow karyotyping. Cytometry 11:184-915.
Trump, D., G. Pilia, P.H. Dixon, C. Wooding, R. Thakkar, S.E.A. Leigh, R. Nagaraja, M.P. Whyte, D. Schlessinger, and R.V. Thakker. 1996. Construction of a YAC contig and an STS map spanning 3.6 megabase pairs in Xp22.1. Human Genet. 97: 60-68.
Van der Maarel, S.M., I.HJ.M. Sholten, J.A. Maat-Kievit, I. Hubber, YJ.M. de Kok, I. de Wijs, TJ. van de Pol, H. van Bokhoven, J.T. den Dunnen, GJ. van Omman, et al. 1995. Yeast artificial chromosome cloning of the Xq 13.3-q21.31 region and fine mapping of a deletion associated with choroideremia and nonspecific mental retardation. Eur. J. Hum. Genet. 3: 207-218.
Vetrie, D., E. Kendall, A. Coffey, S. Hassock, J. Collins, C. Todd, H. Lehrach, M. Bobrow, D.R. Bentley, and A. Harris. 1994. A 6.5 Mb yeast artificial chromosome contig incorporating 33 DNA markers on the human X chromosome at Xq22. Genomics 19: 42-47.
Villard, L., J. Gecz, L. Colleaux, A.M. Lossi, J. Chelly, Y. Ishikawa-Brush, A.P. Monaco, and M. Fontes. 1995. Construction of a YAC contig spanning the Xq13.3 subband. Genomics 26: 115-122.
Zucchi, I., S. Mumm, G. Pilia, S. MacMillan, R. Reinbold, L. Susani, J. Weissenbach, and D. Schlessinger. 1996. YAC/STS map across 12 Mb of Xq27 at 25 kb resolution, merging Xq26-qter. Genomics 34: 42-54.
Received October 1, 1996; accepted in revised form January 17, 1997.
222 @ GENOME RESEARCH
Cold Spring Harbor Laboratory Press on July 15, 2011 - Published by genome.cshlp.orgDownloaded from




















