
Weerts, H.J.T., J. Opdebeeck, A. Otte & J. van Doesburg, 2015: Vondsten nieuwe tijd, in: M. Kuitems...
53
Rapport 4 van het geo-archeologisch en paleontologisch onderzoek zandwingebied en buitencontour Maasvlakte 2 Succesvol botvangen M. Kuitems D. De Loecker T. van Kolfschoten W. Borst J. van Doesburg H.P. van der Es J. Opdebeeck A. Otte J.W.F. Reumer O. van Tongeren H.J.T. Weerts F. Wesselingh Rapport 4 van het geo-archeologisch en paleontologisch onderzoek zandwingebied en buitencontour Maasvlakte 2
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Weerts, H.J.T., J. Opdebeeck, A. Otte & J. van Doesburg, 2015: Vondsten nieuwe tijd, in: M. Kuitems...

een auteur
Rapport 4 van het geo-archeologisch en paleontologisch onderzoek zandwingebied en buitencontour Maasvlakte 2
Succesvolbotvangen
M. KuitemsD. De LoeckerT. van KolfschotenW. BorstJ. van DoesburgH.P. van der EsJ. OpdebeeckA. OtteJ.W.F. ReumerO. van TongerenH.J.T. WeertsF. Wesselingh
Rap
po
rt 4 van h
et geo
-archeo
log
isch en
paleo
nto
log
isch o
nd
erzoek zan
dw
ing
ebied
en b
uiten
con
tou
r Maasvlakte 2

Succesvol botvangen
Rapport 4 van het geo-archeologisch en paleontologisch onder-
zoek zandwingebied en buitencontour Maasvlakte 2
M. KuitemsD. De Loecker
T. van KolfschotenW. Borst
J. van DoesburgH.P. van der Es
J. OpdebeeckA. Otte
J.W.F. ReumerO. van Tongeren
H.J.T. WeertsF. Wesselingh

Colofon
Succesvol botvangen
Rapport 4 van het geo-archeologisch en paleontologisch onderzoek zandwingebied en buitencontour
Maasvlakte 2
Onderzoek 2009-2013
In opdracht van:
Havenbedrijf Rotterdam N.V. Projectorganisatie Maasvlakte 2
Auteurs:
Faculteit der Archeologie, Universiteit Leiden: M. Kuitems, D. De Loecker, T. van Kolfschoten
Havenbedrijf Rotterdam: W. Borst, O. van Tongeren
Rijksdienst voor het Cultureel Erfgoed: J. van Doesburg, J. Opdebeeck, A. Otte, H.J.T. Weerts
Natuurhistorisch Museum Rotterdam: H.P. van der Es, J.W.F. Reumer
Naturalis Biodiversity Center: F. Wesselingh
Illustraties:
Conform vermelding onderschriften
Opmaak, ontwerp en omslag:
Archol BV, Leiden
©Faculteit der Archeologie, Universiteit Leiden
ISBN/EAN:978-90-8961-000-3
Faculteit der Archeologie, Universiteit Leiden
Postbus 9515
2300 RA Leiden
http://www.archeologie.leidenuniv.nl

Inhoud
1 Inleiding 5
2 Methoden 7
3 Fossiele zoogdieren 13
4 Tafonomie van de fossiele zoogdieren 17
5 Mollusken 31
6 Vondsten nieuwe tijd 35
7 Relatie tracks en vondsten 41
8 Discussie en conclusie 47
9 Literatuur 49
De bijgeleverde CD bevat de bijlagen waarnaar wordt verwezen in het rapport


InleIdIng 5
1 Inleiding
D. De Loecker
Een rijk bodemarchiefSinds de tweede helft van de 19de eeuw worden er regelmatig pliocene, pleistocene
en holocene faunaresten opgevist uit de Noordzee (Mol et al. 2008; Mol 2012).
Dit resulteerde aanvankelijk in een verhoogde interesse voor het verzamelen van
dierlijke fossielen, afkomstig uit verdronken paleo-landschappen. Door een intense
en gestructureerde samenwerking tussen (semi-)professionele paleontologen/
archeologen en de commerciële boomkorvisserij zijn er de laatste zes decennia
duizenden botten geborgen van de Noordzeebodem (Heuff et al. 2010). Daardoor kan
de Noordzee intussen gezien worden als één van de rijkste gebieden in de wereld voor
het vinden van fossiel zoogdiermateriaal.
De fossielen worden normaal gesproken opgevist door kotters die het materiaal als
bijvangst in hun netten tegenkomen (o.a. Mol 1991; van Kolfschoten en Laban 1995; Mol
en de Vos 1995; Glimmerveen et al. 2004; Mol et al. 2006, 2007, 2008; Mol en Post 2010;
Mol 2012). Ook commerciële baggerwerkzaamheden, zoals grind- en zandwinning, voor
de kust van Engeland en Nederland leverden een overvloed aan spectaculaire vondsten
op (o.a. Peeters et al. 2009; Hublin et al. 2009; Wessex Archaeology 2010, 2011a en
b). Naast de enorme hoeveelheid botmateriaal worden sporadisch vroeg/midden-
paleolithische en vroeg-mesolithische artefacten aangetroffen (o.a. Louwe Kooijmans
1971; Verhart 1988, 1995, 2001, 2004; Glimmerveen et al. 2004, 2006; De Loecker 2010;
Pieters et al. 2010; Hijma et al. 2011; Wessex Archaeology 2011a en b; Momber et al.
2011) en zeer uitzonderlijk komen er ook fossiele menselijke resten van de bodem van
de zee. Recentelijk werd een schedelfragment uit de Noordzee afkomstig van een jong
volwassen mannelijke Neanderthaler (Homo neanderthalensis) ( Hublin et al. 2009;
Amkreutz et al. 2010).
Aanleiding tot onderzoekHet opgeviste dierlijk botmateriaal kan naast zijn waarde voor paleontologische
studies informatie verschaffen over menselijke activiteiten; bijvoorbeeld aan de
hand van mogelijke snij- en slachtsporen op botten. Verder kunnen de faunaresten
ons informeren over o.a. dateringen, klimaat en landschap ten tijde van depositie.
Het probleem is dat van vrijwel al deze zorgvuldig gedocumenteerde vondsten een
(gedetailleerde) landschappelijke, ecologische en stratigrafische context ontbreekt.
Zonder verder onderzoek kunnen deze bot- en artefactverzamelingen omschreven
worden als:
“… a result of post-depositional processes both natural as well as industrial. A cautious,
scientific approach would be to suggest that from this perspective, the dredged finds
should be treated as a complex palimpsest ‘surface scatter’ with a restricted value for
early human behavioural inferences.” (Tizzard et al. in press).
Met andere woorden, deze unieke vondsten illustreren de hoge wetenschappelijke
potentie van het Noordzeebekken, maar de losse vondsten kunnen moeilijk in een
archeologische context geplaatst worden (Nieuwenkamp 2012). Onderzoek moet zich
daarom richten op vragen zoals:

6 Botvangen
· Waar komen de vondsten precies vandaan? (stratigrafische context)?
· Wanneer leefden deze dieren en mensen? (relatieve en absolute datering)?
· Wat is het chronologisch kader?
· Wat waren de klimatologische omstandigheden waaronder de dieren en mensen
leefden? (ecologische context)?
· Hoe zag het pleistocene en holocene landschap eruit? (landschappelijke context)?
· Wat was de positie van de rivierlopen in verschillende periodes? (landschappelijke
context)?
· Etc…
Antwoorden op dergelijke vragen zouden dan weer een grote bijdrage kunnen leveren
aan thema’s zoals (cf. Deeben et al. 2006):
· De kolonisatie en vroegste bewoningsgeschiedenis van Europa;
· Het gebruik van het landschap en nederzettingssystemen tijdens bepaalde
perioden;
· Voedseleconomieën, relatie mens en milieu tijdens bepaalde perioden;
· Culturele tradities, sociale relaties en interacties.
Helaas is dit aan de hand van ‘losse’ vondsten niet mogelijk. Archeologen beschikken
echter over een breed scala aan analytische disciplines en technieken om paleoland-
schappen te bestuderen. Deze, voornamelijk uit de aardwetenschappen afkomstige
vakken, bevatten o.a. sedimentologie, lithologie, micromorfologie, paleontologie,
botanie, malacologie. Door inpassing van dergelijke vakgebieden in het archeologisch
onderzoek van de Noordzeebodem, zouden kennisleemten opgevuld kunnen worden.
In het bijzonder een goede lithostratigrafische studie is van cruciaal belang voor
het verschaffen van een context aan, en een datering van, de gevonden objecten.
Concluderend kunnen we stellen dat de focus op het verzamelen van objecten moet
verplaatsen naar een focus op de context van de objecten (Nieuwenkamp 2012: 26).
Een mogelijkheid hiervoor werd geboden door de aanleg van Maasvlakte 2. Sinds 2008 is
er in het kader van het Maasvlakte 2 Project onderzoek gedaan naar de archeologische
en paleontologische relicten die aangetroffen werden tijdens de uitbreiding van de
Rotterdamse haven. Grondig geplande vistochten in het zandwingebied, ca. 11 km
voor de Nederlandse kust, hebben in dit onderzoek een belangrijke rol gehad. Met
behulp van kotters, voorzien van ‘bottom trawls’, werd de zeebodem doelgericht
afgezocht naar botfragmenten en archeologische objecten (lokaliseren en karteren
van vondstrijke gebieden). Verder hadden de verschillende surveys als doel, het
plaatsen van het geborgen materiaal in een stratigrafisch/geologisch, ecologisch en
landschappelijk kader. Naast het verzamelen van vondstmateriaal werd er namelijk ook
gestart met een litho- en chronostratigrafische reconstructie van het zandwingebied,
waaraan de vondsten later gekoppeld zouden worden.
Het complete archeologische, paleontologische en geologische Maasvlakte 2
onderzoek zou uiteindelijk een landschappelijke, ecologische en stratigrafische context
moeten geven aan de paleontologische en archeologische vondsten.
Dit ‘vistochten’ rapport presenteert eerst kort de gevolgde strategie en methodologie
tijdens de verschillende surveys en analyses. Vervolgens wordt er een overzicht
gegeven van de geborgen vondsten.

Methoden 7
2 Methoden
D. De Loecker, O. van Tongeren
Van de OD-7, via de BRA-7, tot de BRA 5In oktober 2009 is de boomkotter OD-7, met aan boord een bemanning van amateur-
paleontologen, vertrokken om de eerste survey in het kader van het Maasvlakte 2
Project uit te voeren. Tijdens deze expeditie werd per track aan weerszijden van het
schip een net uitgezet. Na iedere track zijn de netten opgehaald. Deze tocht heeft
o.a. geresulteerd in de vangst van het grootste wolharige mammoetdijbeen dat tot
op heden gevonden is op Nederlandse bodem (Moeliker 2010). Het 133 cm lange bot
werd aangeboden aan het Natuurhistorisch Museum in Rotterdam (nmr) tijdens het
ondertekenen van een convenant met het Havenbedrijf (medio 2010). In het convenant
werd vastgelegd dat alle vondsten die op de in aanbouw zijnde Maasvlakte 2 worden
gevonden aan het nmr worden aangeboden.
Naar aanleiding van deze geslaagde tocht en met de achterliggende gedachte om de
context van de vondsten te achterhalen, is binnen de projectorganisatie nagegaan of
het mogelijk was om in 2010 vervolg te geven aan dit soort surveys. Bij de uiteindelijke
positieve beslissing heeft een aantal factoren meegespeeld:
· Organisatorische en financiële haalbaarheid: aangezien het Havenbedrijf zes maal
per jaar, gedurende een week (maandag tot vrijdag), in eigen beheer slibmetingen
uitvoert langs de Nederlandse kust, was er de mogelijkheid om interdisciplinair te
werk te gaan. Voor dit slibonderzoek is de BRA-7 gecontracteerd, een Eurokotter
die vergelijkbaar is met de OD-7.
· Wetenschappelijke meerwaarde: naast het traceren en verzamelen van vondsten,
zouden vervolg-surveys gebruikt kunnen worden om een methodologie te
ontwikkelen waarbij zoveel mogelijke contextuele informatie verzameld kon
worden (strategie van vissen aanpassen).
In februari 2011 is tijdens een ‘slibweek’ als proef door de bemanning van de BRA-7
en de aanwezige surveyors (Havenbedrijf) en toezichthouders (projectorganisatie), in
de zandwinput naar botten gevist. Het resultaat was dat dit ook met de boomkotter
BRA-7, voorzien van haar ‘bottom trawls’, uitstekend te doen was. Vervolgens is
besloten om op de resterende vijf zaterdagen volgend op een slibweek vistochten in de
zandwinput uit te voeren. Deze tochten hebben kwantitatief en kwalitatief waardevol
materiaal opgeleverd. Bovendien begon er, door de expertise van de bemanning
(meestal dezelfde kapitein, bemanning en [semi-]professionele wetenschappers), een
inzicht te ontstaan over de samenstelling en de kwantiteit van de vondsten per beviste
locatie en per track.
Dit alles resulteerde in een overleg met de Rijksdienst voor het Cultureel Erfgoed.
Mede omdat er artefacten aangetroffen konden worden, werd er besloten om in 2011
een stevig wetenschappelijk onderbouwde en multidisciplinaire survey-week op te
zetten (2011, week 23). Vervolgens heeft er op 9 september 2011 nog een vistocht
plaatsgevonden met de BRA 5.
Toenemende nauwkeurigheidTijdens de verschillende vistochten (zie Tabel 2.1) is in toenemende mate aandacht
besteed aan de nauwkeurigheid van het vastleggen van de beviste tracks. Tijdens
de eerste vistocht met de OD-7 werd uitsluitend het beginpunt en eindpunt van de

8 Botvangen
vistrack vastgelegd. Met de BRA-7 werd daarna t/m juni 2011 de gehele vistrack
vastgelegd en tenslotte zijn in september 2011 met de BRA 5 tijdens de track
bovendien de tijdstippen vermeld van start vieren, net aan de grond, stop vieren, start
halen, net los van de grond en stop halen.
Bij eerdere vistochten werd het materiaal alleen per track geanalyseerd. Omdat
was opgevallen dat de inhoud van het stuurboordnet vaak in grote mate verschilde
van het bakboordnet zijn vanaf de visweek in juni 2011 de vondsten per net apart
genoteerd. Vanaf deze visweek werd de opgeviste inhoud van de twee netten
gescheiden verzameld in een stortbak en de stuurboord- en bakboordvondsten kregen
een eigen vondstformulier toegekend. Na het nemen van een overzichtsfoto (per
verzameleenheid) is het materiaal vervolgens vanuit de stortbak omhoog getakeld
door middel van een lopende band alwaar ter plaatse de inhoud van de individuele
netten nauwkeurig werd gesorteerd op vondstcategorie. Diverse (semi-)professionele
wetenschappers concentreerden zich bij het uitzoeken op botmateriaal, vuursteen,
steen, hout, schelpen, veen, klei, ander materiaal en de administratie.
Ook kreeg Deltares de opdracht om tijdens deze visweek diverse onderzoeken uit te
voeren met behulp van een Side Scan sonar, een Subbottom Profiling met boomer en
Figuur 2.1 Bathymetrische opname (augustus 2011) van de noordelijke zandwinput. De diepte ver-loopt van ca. 20m -NAP aan de noordrand tot ca 40m –NAP in het midden van de put.
Datum van uitvoering Schip en maaswijdte Locatie van uitvoering
1-10-2009 Kotter OD-7, # 10cm Zandwingebied en stukje van Eurogeul
2-10-2009 Kotter OD-7, # 10cm Zandwingebied en stukje van Eurogeul
17-2-2010 Kotter BRA-7, # 10cm Zandwingebied
24-4-2010 Kotter BRA-7, # 10cm Zandwingebied
26-6-2010 Kotter BRA-7, # 10cm Zandwingebied
18-8-2010 Kotter BRA-7, # 10cm Zandwingebied
16-10-2010 Kotter BRA-7, # 10cm Zandwingebied
9-12-2010 Kotter BRA-7, # 10cm Zandwingebied
6-6-2011 Kotter BRA-7, # 8 & 2cm Zandwingebied
7-6-2011 Kotter BRA-7, # 8 & 2cm Zandwingebied
8-6-2011 Kotter BRA-7, # 8 & 2cm Zandwingebied
9-6-2011 Kotter BRA-7, # 8 & 2cm Zandwingebied
10-6-2011 Kotter BRA-7, # 8 & 2cm Zandwingebied
9-9-2011 Kotter BRA 5, # 8 & 2cm Zandwingebied
Tabel 2.1 Overzicht van de paleontologische vistochten.

Methoden 9
een X-star (Shallow Seismic), met onder andere als doel te onderzoeken of de exacte
positie van de opgeviste objecten op de zeebodem kon worden vastgesteld in relatie
tot een eventueel zichtbare stratigrafie.
BathymetrieGaandeweg werd ook steeds minder willekeurig een track gekozen, maar werd op
basis van bathymetrische gegevens en ervaring opgedaan tijdens eerdere vistochten
gekozen voor kortere vistracks binnen een beperkte diepterange.
Tijdens de zandwinning zijn er voortdurend veranderingen in de diepte van de winput.
Voorafgaand aan elke vistocht is daarom de meest recente bathymetrische opname
van de winpunt opgevraagd bij de aannemer (PUMA). Een voorbeeld van zo’n
bathymetrische opname staat in Figuur 2.1.
Tijdens de eerste vistochten is ervan uitgegaan dat de meeste paleontologische
vondsten gedaan zouden worden op een diepte van rond de 30 meter (D. Mol pers.
comm.) en daarom was er een voorkeur om op die diepte te vissen. Later, toen
duidelijk werd dat ook op andere diepten vondsten werden gedaan, is besloten om een
deel van de zandwinput systematischer te gaan afzoeken.
De geografische positie van het schip (dus zonder loding, d.w.z. zonder bepaling van
de diepte ter plekke) wordt vijf maal per seconde vastgelegd d.m.v. het nautische
programma Quinsy. Later zijn met behulp van de bathymetrische opnamen de diepten
toegevoegd aan de vastgelegde coördinaten. Per vistrack is dus een tabel (voorbeeld
in Tabel 2.2) beschikbaar van de tijd, de X-coördinaat, de Y-coördinaat (beide in RD) en
de diepte volgens de laatste loding.
Omdat er steeds enige tijd verlopen is tussen de loding en de vistocht kunnen de
gegevens enigszins vertekend zijn, maar er is steeds gestreefd naar vissen in gebieden
waar sinds de laatste loding weinig of geen zand gewonnen is. Een andere vertekening
in de gegevens wordt veroorzaakt, doordat het net ongeveer 100 meter achter het
schip aansleept. Omdat de registratie van de track in het algemeen gestart is op het
moment dat de lier gevierd wordt en beëindigd op het moment dat het net met de lier
gehaald wordt is deze vertekening naar schatting gering. De gegevens zijn per track in
een overzicht geplaatst (Figuur 2.2).
In september 2011 is tevens genoteerd op welk tijdstip het vieren en halen van het
net startte en stopte en wanneer het net de bodem raakte of los kwam van de bodem
(dit is te zien aan de spanning op de vislijn). De tracks van 9 september zijn daarmee
gecorrigeerd voor de afstand van het net achter het schip. Deze correctie is niet voor
alle vistracks geslaagd. Figuur 2.3 laat de vistracks van 9 september 2011 zien, voor en
na correctie. In Bijlage Hoofdstuk 2 staat een overzicht van alle vistracks.
Exported layer: SURVEYDATAKP Easting Northing Depth0 54145.33 448630.633 -23.9050.3369 54145 448630.709 -23.890.674 54144.67 448630.785 -23.8731.011 54144.35 448630.862 -23.8561.347 54144.02 448630.938 -23.8371.684 54143.69 448631.014 -23.8192.021 54143.36 448631.09 -23.813….. ….. ….. …..341.39 53808.9 448606.182 -26.064341.73 53808.57 448606.139 -26.079342.06 53808.23 448606.096 -26.091342.4 53807.9 448606.053 -26.104342.73 53807.57 448606.01 -26.115343.07 53807.23 448605.968 -26.126343.41 53806.9 448605.925 -26.137343.74 53806.57 448605.882 -26.146
Tabel 2.2Voorbeeld van gegevens van een paleontologische vistrack(KP=afgelegde weg)

10 Botvangen
Max: diepte -26.15 m NAP Gevaren lengte 343.7 mMin diepte -23.79 m NAPGem. diepte -24.85 m NAP Track
1Easting Northing Depth Datum:
Start punt 54,145 448,631 -23.9 m NAP vrijdag 10 juni 2011Eind punt 53,807 448,606 -26.1 m NAP PlaatsAfstand begin - eind punt (= RECHTE LIJN) 339.7 m Zandwingebied MV2
Schip : BRA-7 Schipper : Dirk Kraak Opdrachtgever Havenbedrijf R'dam (HbR)
447500
448500
449500
51500 52500 53500 54500 55500
Nor
thin
g
Easting
Coordinaten (Eastin - Northing) van Track 001 gevaren op 10 juni 2011
-35
-33
-31
-29
-27
-25
-23
-21 0 100 200 300 400
Die
pte
in [m
NA
P]
Gevaren lengte langs de track [m]
Profiel Track - 001 gevaren op 10 Juni 2011
-45
-40
-35
-30
-25
-20
Figuur 2.2 Overzicht van de gegevens van een paleontologische vistrack. Boven de ligging ten opzichte van de zandwincontour, midden de diepteligging en onder een overzicht in getallen met rechts een box-and-whisker plot van de verdeling over de diepten.

Methoden 11
Figuur 2.3 De ligging van de vistracks van 9 september 2011 ten opzichte van de noordoosthoek van de zandwinput. Boven ongecorrigeerd, beneden gecorrigeerd voor de tijd dat het net aan de grond was. De contour van de zandwinput is aangegeven met een rode streep-stiplijn.

12 Botvangen

FossIele zoogdIeren 13
3 Fossiele zoogdieren
H.P. van der Es en J.W.F. Reumer
InleidingTijdens veertien vistochten die zijn uitgevoerd in het kader van Maasvlakte 2 zijn
botresten van grote zoogdieren gevonden. De methode van verzamelen is door middel
van kornetten, die aan weerszijden van een viskotter over de zeebodem worden
getrokken. Bij één tocht is gebruik gemaakt van mosselkorren, tijdens de overige
tochten van platviskorren.
Materiaal en methodenAl het botmateriaal (zie overzicht hieronder) is verzameld en meegenomen naar het
Natuurhistorisch Museum Rotterdam (nmr), om aldaar te worden geselecteerd. De
gehanteerde selectiecriteria voor botmateriaal zijn onder andere fragmentatiegraad,
identificeerbaarheid en staat van preservatie. In bepaalde gevallen is ook zeldzaamheid
of bijzonderheid een criterium.
De botten of botfragmenten die volgens de criteria in aanmerking kwamen voor
conservering en opname in de museumcollectie zijn grotendeels ontzilt, gereinigd
en geconserveerd met in aceton oplosbare polymeren, door de heer A. Hoekman van
North Sea Fossils/Eurocape te Urk. Een deel heeft deze behandeling niet ondergaan,
om toekomstig onderzoek (14C, DNA) mogelijk te maken. De fossielen zijn vervolgens
geregistreerd en opgenomen in de collectie, de gegevens zijn ingevoerd in de digitale
databestanden van het museum. Botten en botfragmenten die niet in de collectie
worden opgenomen zijn niet op dergelijke wijze behandeld, maar worden wel
genoemd in de bijlage (Bijlage Hoofdstuk 3). Ze zullen hetzij worden overgedragen aan
Leiden voor verder tafonomisch onderzoek, dan wel worden afgestoten.
ResultatenIn Tabel 3.1 zijn de soorten zoogdieren weergegeven die tijdens het onderzoek zijn
aangetroffen. Determinaties zijn gedaan door K. Post, soms in samenwerking met D.
Mol, indien noodzakelijk aan de hand van de vergelijkingscollectie van het nmr. De
aantallen betreffen de in de collectie opgenomen fossielen.
De aangetroffen soorten behoren vrijwel integraal tot de landzoogdieren van het
zogenaamde Mammuthus-Coelodonta Faunal Complex (Kahlke 1999), dat is het
samenstel van grote zoogdieren dat kenmerkend is voor het ecosysteem van de
mammoetsteppe (Guthrie 1990). De twee meest in het oog springende uitzonderingen
hierop zijn de otter (Lutra lutra), die tot de holocene fauna behoort, en de zadelrob
(Phoca cf. groenlandica), dat een koudeminnend zeezoogdier is dat leefde in het
estuarium van de Rijn/Maas.
Aan het materiaal is één 14C-datering verricht, en wel aan de otter (Lutra lutra), omdat
dit object in eerste instantie buiten het faunaspectrum van de mammoetsteppe valt.
Dit object (Figuur 3.1) heeft een ouderdom van 8825 +/- 45 jaar BP (laboratorium
nummer GrA-52432; zie Mol en van der Plicht 2012).
Een aantal bijzondere vondsten leidde tot publicaties in de populaire of wetenschap-
pelijke literatuur. Wij noemen hier het grootste dijbeen van mammoet (Mammuthus
primigenius) ooit in Nederland gevonden, met een lengte van 133 centimeter (Figuur
3.2, uit: Moeliker 2010), de fossiele coproliet van een hyena (Crocuta crocuta spelaea)
die de eerste vondst van een dergelijk sporefossiel uit de Noordzee betrof (Figuur 3.3,
Reumer et al. 2010; Mol et al. 2010), en de mooi geconserveerde schedel van een otter

14 Botvangen
(Lutra lutra), die de oudste tot nu toe bekende Nederlandse otter betreft (Figuur 3.1,
Mol en van der Plicht 2012). Verder zijn twee halswervels van mammoet (Mammuthus
primigenius) aangetroffen die een interessante pathologie vertonen, namelijk articula-
tiefacetten voor halsribben (Figuur 3.4, Reumer en Galis 2010).
Figuur 3.1 Schedel van otter (Lutra lutra) (inv. nr. NMR 9991-7708), het oudste exemplaar van Nederland.
Figuur 3.2 Het dijbeen van mammoet (Mammuthus primigenius) met een recordlengte van 133 cm (inv. nr. NMR 9991-6648) wordt door dhr Ronald Paul (rechts) overgedragen aan Jelle Reumer van het Natuurhistorisch Museum.

FossIele zoogdIeren 15
Proboscidea (olifantachtigen) Mammuthus primigenius (Blumenbach, 1799) (mammoet) 26Perissodactyla, Rhinocerotidae neushoorns Coelodonta antiquitatis (Blumenbach, 1799) (wolharige neushoorn) 8Perissodactyla, Equidae (paarden) Equus caballus Linnaeus, 1758 (paard) 19Artiodactyla, Cervidae (herten) Cervus elaphus Linnaeus, 1758 (edelhert) 3 Cervus cf. elaphus Linnaeus, 1758 (vermoedelijk edelhert) 3 Megaloceros giganteus (Blumenbach, 1799) (“Iers” reuzenhert) 10 Rangifer tarandus (Linnaeus, 1958) (rendier) 17 Cervidae sp. (hert, ongedetermineerd) 1Artiodactyla, Bovidae (holhoornigen) Bison priscus Bojanus, 1827 (wisent) 12 Bos sp. (rund) 1Carnivora, Mustelidae (marterachtigen) Lutra lutra (Linnaeus, 1758) (otter) 1Carnivora, Ursidae (beren) Ursus cf. arctos Linnaeus, 1758 (vermoedelijk bruine beer) 1Carnivora, Canidae (hondachtigen) Canidae sp. (hond of wolf ) 1Carnivora, Hyaenidae (hyena’s) Crocuta crocuta spelaea (Goldfuss, 1823) (grottenhyaena) 1Carnivora, Phocidae (robben) Phoca cf. groenlandica Erxleben, 1777 (vermoedelijk zadelrob) 2
Totaal 307
Tabel 3.1 De zoogdieren die tijdens het onderzoek zijn aangetroffen. De aantallen betreffen de fossielen die in de collectie van het NMR zijn opgenomen.
Figuur 3.3 De coproliet van hyena (Crocuta crocuta spelaea) (inv. nr. NMR 9991-7071), het eerste exemplaar uit de Noordzee.
Figuur 3.4 Zesde halswervel van mammoet (Mammuthus primigenius), met articulatiefacetten voor (pathologische) halsribben (inv. nr. NMR 9991-6627).

16 Botvangen

taFonoMIe 17
4 Tafonomie van de fossiele zoogdieren
M. Kuitems, D. De Loecker, T. van Kolfschoten
IntroductieIn dit hoofdstuk is het skeletmateriaal beschreven dat is verzameld tijdens acht
vistochten die hebben plaatsgevonden in 2010 op 17 februari, 24 april, 26 juni, 18
augustus, 16 oktober en 9 december en in 2011 op 6-10 juni en 9 september. Al het
skeletmateriaal van deze vistochten dat in de collectie van het nmr is opgenomen
en een gedeelte van het materiaal dat zich niet in de collectie bevindt is door Margot
Kuitems, Dimitri De Loecker en Thijs van Kolfschoten (Faculteit der Archeologie,
Universiteit Leiden) macroscopisch op tafonomie bestudeerd.
Tafonomie is de studie naar processen die hun weerslag hebben gehad op het
skeletmateriaal nadat het dier gestorven is. Deze processen hebben onder andere
invloed op verspreiding van het botmateriaal, fragmentatiegraad en conservering. Aan
de hand van tafonomische bestudering van het materiaal kan men een reconstructie
proberen te maken van de processen waaraan een bot is blootgesteld en inzicht krijgen
in de directe omgeving waarin het materiaal heeft gelegen.
In dit hoofdstuk wordt ingegaan op tafonomische aspecten van het opgeviste
skeletmateriaal (inclusief bot, gewei, ivoor, tanden en kiezen) zoals fragmentatiegraad,
verwering en modificatie van het materiaal door dierlijke en menselijke activiteit, om
antwoord te vinden op de volgende onderzoeksvragen:
- Wat is de samenstelling van het opgeviste skeletmateriaal?
- Wat is de conserveringsgraad van de verschillende materiaalcategorieën?
- Welke informatie geeft het materiaal over sortering van het materiaal en waar het
materiaal oorspronkelijk vandaan komt?
- Welke aanwijzingen voor menselijke activiteit levert het materiaal?
MateriaalDe vistochten hebben 478 fragmenten en complete elementen van skeletmateriaal
opgeleverd. Daarvan heeft Klaas Post (Urk) er 360 stuks op soort kunnen brengen.
Het merendeel van het materiaal is gedetermineerd als wolharige mammoet.
Andere soorten waarvan het materiaal afkomstig is, zijn wolharige neushoorn, paard,
hertachtige, rund, otter, hond/wolf, zadelrob en vermoedelijk bruine beer. Van twaalf
fragmenten is het type skeletelement vastgesteld, maar niet de diersoort waartoe het
behoord heeft. Van deze elementen is slechts te zeggen dat ze afkomstig zijn van een
groot zoogdier (ter grootte van neushoorn, paard, hert of rund). 106 stukken waren zo
fragmentarisch, dat ze niet meer tot diersoort of skeletelement te herleiden zijn. Een
overzicht van de hoeveelheden materiaal per diersoort is weergegeven in Tabel 4.1.
De tafonomische analyse van het materiaal heeft plaatsgevonden lang (een aantal
maanden tot twee jaar) nadat het was opgevist. Het materiaal is in de tussentijd
ontzilt en gedroogd en op basis van door Klaas Post bepaalde selectiecriteria 204
stuks in de collectie van het museum opgenomen (Tabel 4.1). Hiervan zijn 180
elementen behandeld met conserveermiddel (een oplossing van velpon en aceton) en
genummerd. Materiaal dat niet in de museumcollectie is opgenomen is wel ontzilt,
maar niet van een nummer voorzien en in de meeste gevallen niet geconserveerd.
Voor het tafonomische onderzoek zou het echter beter geweest zijn als de analyse
direct na de vistochten was uitgevoerd. In dat geval was al het materiaal nog bij elkaar
geweest en was refitten mogelijk. Een ander groot voordeel zou zijn geweest dat het

18 Botvangen
botoppervlak nog onbehandeld was, waardoor sporen van welke aard dan ook beter
zichtbaar zouden zijn geweest dan nu het geval was.
Het feit dat veel van het materiaal gedetermineerd heeft kunnen worden, heeft naast
de deskundigheid van Klaas Post te maken met factoren als de maaswijdte van het
net waarmee is gevist (die verhinderde dat grote hoeveelheden ondetermineerbare
botsplinters zijn opgevist) en de mate van conservering van het materiaal (die er
bijvoorbeeld voor zorgde dat grote delen van skeletelementen bewaard zijn gebleven).
Type skeletelement In Tabel 4.2 is per diersoort het totaal aantal (n=478) opgeviste skeletelementen
weergegeven, gesorteerd per lichaamsdeel. Van wolharige mammoet, paard, rendier,
hert en steppewisent is materiaal van de verschillende onderdelen van het lichaam
aangetroffen, namelijk van kop, romp, voorpoot, achterpoot en voet.
Zoals reeds uit Tabel 4.1 blijkt, zijn de meeste skeletelementen afkomstig van
wolharige mammoet. Deels heeft dit te maken met de verzameltechniek; in de
netten blijven grotere botten makkelijker achter dan kleinere elementen (van kleinere
diersoorten). Van de mammoet is ruim een derde van het materiaal (n=105) afkomstig
van de romp (Tabel 4.2). Dit is te verklaren doordat de romp uit het grootste aantal
skeletelementen (7 halswervels, 18 borstwervels, 1 borstbeen, 20 paar ribben, 5
lendenwervels, 22 staartwervels en 1 heiligbeen) bestaat. Daarnaast zijn bepaalde
elementen als ribben, doornuitsteeksels van de borstwervels en dwarsuitsteek-
sels van de lendenwervels gevoelig voor fragmentatie, wat kan bijdragen aan de
hoge frequentie van het aantal aangetroffen rompelementen. Ook van de overige
diersoorten is ongeveer een derde (31%) van het opgeviste materiaal toe te schrijven
aan de romp.
Van de gevonden skeletelementen van mammoet is 20% afkomstig van de voet. Ten
opzichte van de kleinere diersoorten zijn er veel voetwortelbeenderen gevonden.
Terwijl deze bij kleinere dieren moeilijk terug te vinden zijn, zijn deze bij mammoeten
natuurlijk relatief groot en daardoor goed herkenbaar. Van de andere diersoorten is
een vergelijkbaar percentage (17%) van de elementen afkomstig van de voet. Het
merendeel daarvan bestaat uit koten en sprongbeenderen. Waar fragmentatie (zoals
hierboven beschreven voor rompelementen) enerzijds kan leiden tot een toename
in aantal skeletelementen, kan een hoge mate van fragmentatie versnippering
veroorzaken, waardoor elementen niet meer gedetermineerd kunnen worden.
Bovendien zijn kleinere en poreuzere skeletelementen vatbaarder voor verwering
(Andrews 1997), dat ook nadelig kan zijn voor determinatie. Botten die minder
gevoelig zijn voor fragmentatie (zoals de hier besproken koten, wortelbeenderen
en sprongbeenderen) blijven vaak meer intact en zijn daardoor makkelijker te
determineren.
soort in collectie niet opgenomen totaalwolharige mammoet 144 128 272wolharige neushoorn 6 2 8paard 14 4 18edelhert 3 1 4reuzenhert 10 3 13rendier 13 7 20hert 1 6 7steppewisent 7 5 12rund 1 0 1groot zoogdier 0 1 1middelgroot zoogdier 0 11 11otter 1 0 1cf. bruine beer 1 0 1hond of wolf 1 0 1zadelrob 2 0 2indet. 0 106 106
Tabel 4.1 Aantal skeletelementen per diersoort dat is opgevist tijdens verschillende vistochten en al dan niet is opgenomen in de collectie van het Natuurhistorisch Museum Rotterdam.

taFonoMIe 19
Tabel 4.2 Aantal opgeviste skeletelementen per dier-soort.
soort lichaamsdeel n element n
wolharige mammoet kop 34 schedel 14
gehoorbeen 1
onderkaak 4
kies 4
stoottand 11
romp 105 halswervel 12
borstwervel 32
borstbeen 1
rib 36
lendewervel 10
wervel indet. 7
staartwervel 5
heiligbeen 2
voorpoot 47 schouderblad 13
opperarmbeen 17
spaakbeen 11
ellepijp 6
achterpoot 31 bekken 5
dijbeen 14
knieschijf 1
scheenbeen 11
poot 2 pijpbeen indet. 2
voet 53 sprongbeen 2
hielbeen 4
voetwortelbeen 15
middenvoetsbeen 26
koot 6
wolharige neushoorn kop 1 kies 1
romp 1 halswervel 1
voorpoot 2 opperarmbeen 1
spaakbeen 1
achterpoot 3 bekken 1
dijbeen 2
voet 1 voetwortelbeen 1
paard kop 2 onderkaak 1
kies 1
romp 3 halswervel 1
rib 1
heiligbeen 1
voorpoot 2 opperarmbeen 1
ellepijp 1
achterpoot 3 dijbeen 2
scheenbeen 1
voet 8 middenvoetsbeen 2
sprongbeen 2
koot 4
edelhert kop 1 gewei 1
voet 3 middenvoetsbeen 1
sprongbeen 1
hielbeen 1
reuzenhert kop 2 schedel 1
onderkaak 1
romp 8 halswervel 1
borstwervel 5
rib 1
lendewervel 1
voorpoot 2 schouderblad 1
ellepijp 1
voet 1 sprongbeen 1
rendier kop 9 gewei 8
schedel 1
romp 5 halswervel 1

20 Botvangen
De voorpoten (resp. 17% voor mammoet en 18% voor overige diersoorten) en
achterpoten (resp. 11% voor mammoet en 16% voor overige diersoorten) beslaan
respectievelijk 18% en 13% van het totale assemblage. 14% van de skeletelementen is
afkomstig van de kop. Voor mammoet worden de elementen die tot de kop behoren
gedomineerd door schedel- stoottandfragmenten. Schedels zijn gevoelig voor
fragmentatie, vooral die van grotere zoogdieren (Groot 2010). Stoottanden bestaan uit
ivoor, wat een harder materiaal is dan bot. Door de vorm zijn stoottanden desondanks
gevoelig voor fragmentatie.
Bij de overige diersoorten wordt de kop met name vertegenwoordigd door
geweifragmenten. Gewei is opgebouwd uit zeer dicht spongieus materiaal, wat het
gewei licht en stevig maakt (Groot 2010). Hertachtigen verliezen ieder jaar hun gewei,
waardoor gewei relatief vaak kan worden aangetroffen. Van de meeste opgeviste
gewei-elementen is echter niet te zeggen of het deel uitmaakte van een afgeworpen
gewei. In Tabel 4.2 is te zien dat veel geweimateriaal afkomstig is van rendieren.
Dit zou kunnen worden verklaard doordat bij rendieren zowel het mannetje als het
vrouwtje geweidragend is. Bij andere hertachtigen geldt dit alleen voor het mannetje.
Naast de zes kiezen en stoottanden die vermeld zijn in Tabel 4.2, maakt nog een
aantal gebitselementen onderdeel uit van een schedel of onderkaak. In een van
soort lichaamsdeel n element n
schouderblad 1
lendewervel 3
voorpoot 4 spaakbeen 3
ellepijp 1
achterpoot 1 scheenbeen 1
voet 1 koot 1
hert kop 2 gewei 1
schedel 1
romp 1 wervel indet. 1
voorpoot 1 schouderblad 1
achterpoot 2 bekken 1
dijbeen 1
voet 1 koot 1
steppewisent kop 1 hoornpit 1
romp 3 halswervel 1
borstwervel 2
voorpoot 5 schouderblad 2
opperarmbeen 2
spaakbeen 1
achterpoot 1 dijbeen 1
voet 2 hielbeen 1
koot 1
rund romp 1 rib 1
groot zoogdier romp 1 heiligbeen 1
middelgroot zoogdier romp 6 halswervel 1
rib 2
wervel indet. 2
heiligbeen 1
voorpoot 3 schouderblad 1
opperarmbeen 2
achterpoot 2 bekken 2
otter kop 1 schedel 1
cf. bruine beer achterpoot 1 dijbeen 1
hond of wolf romp 1 halswervel 1
zadelrob romp 1 halswervel 1
romp 1 borstwervel 1
indet. divers indet 106
Tabel 4.2 vervolgAantal opgeviste skeletelementen per dier-soort.

taFonoMIe 21
de mammoetschedels zijn nog twee kiezen aanwezig (M3), de onderkaak van het
reuzenhert bevat nog twee kiezen (m2 en m3) en in de schedel van de otter zijn
snijtanden en verder alle (pre)molaren vertegenwoordigd. Al met al kan worden
vastgesteld dat er weinig gebitselementen zijn opgevist, zeker in vergelijking met het
aantal andere skeletelementen. De reden hiervoor is niet duidelijk. Door de chemische
samenstelling van tanden en kiezen (tandbeen, glazuur en cement) en ivoor (tandbeen)
(Huisman et al. 2006) blijven gebitselementen juist relatief goed geconserveerd
(Andrews 1997).
FragmentatieOmdat ten tijde van de tafonomische analyse slechts een deel van het materiaal
dat niet is opgenomen in de collectie beschikbaar was, is om inzicht te krijgen in de
fragmentatiegraad van het materiaal een steekproef genomen van materiaal uit
verschillende vistochten, namelijk die van 18 augustus 2010 en van 6-10 juni 2011.
Om inzicht te krijgen in de fragmentatiegraad van het materiaal, was het namelijk
belangrijk om naast het materiaal dat in de museumcollectie is opgenomen, ook
materiaal te bekijken dat buiten de collectie is gehouden. Gezien het doel van een
museum, bestaat de collectie voornamelijk uit redelijk compleet materiaal. Een
materiaalstudie gebaseerd op het materiaal dat in de collectie aanwezig is, zou een
vertekend beeld opleveren van de compleetheid van het materiaal met een oververte-
genwoordiging van complete tot redelijk complete elementen.
Van de skeletelementen die op 18 augustus zijn verzameld, zijn alle vondsten bekeken
die in de collectie zijn opgenomen (50) en 79 vondsten buiten de collectie. Van deze
129 elementen, is 43% kleiner dan een kwart van de oorspronkelijke grootte van het
element. 32% van het materiaal is groter dan driekwart van de oorspronkelijke grootte,
waarvan 15% complete elementen betreft. Van het materiaal dat in juni is verzameld,
zijn 55 vondsten opgenomen in de museumcollectie. Van deze vondsten en honderd
18-Aug-10
n %
6-10 juni 2011
n %Niet opgenomen (n=79): Niet opgenomen (n=100):
0-25% 52 66 0-25% 69 69
25-50% 7 9 25-50% 10 10
50-75% 7 9 50-75% 7 7
75-100% 6 8 75-100% 4 4
compleet 1 1 compleet 1 1
onbekend 6 8 onbekend 9 9
>75% 7 9 >75% 5 5
In collectie (n=50): n % In collectie (n=55): n %
0-25% 3 6 0-25% 4 7
25-50% 0 0 25-50% 3 5
50-75% 6 12 50-75% 5 9
75-100% 18 36 75-100% 29 53
compleet 18 36 compleet 13 24
onbekend 5 10 onbekend 1 2
>75% 36 72 >75% 42 76
Totaal (n=129) n % Totaal (n=155): n %
0-25% 55 43 0-25% 73 47
25-50% 7 5 25-50% 13 8
50-75% 13 10 50-75% 12 8
75-100% 24 19 75-100% 33 22
compleet 19 15 compleet 14 9
onbekend 11 9 onbekend 10 6
>75% 43 33 >75% 47 30
Tabel 4.3 Compleetheid van een steekproef van het opgeviste skeletmateriaal.

22 Botvangen
elementen die niet in de collectie zijn opgenomen is 47% kleiner dan een kwart van de
oorspronkelijke grootte en 30% is groter dan driekwart van de oorspronkelijke grootte.
De precieze aantallen en percentages zijn weergegeven in Tabel 4.3.
De fragmentatie van de in Tabel 4.3 behandelde skeletelementen is deels veroorzaakt
door vraat: 9% vertoont sporen van carnivorenvraat en één rendiergeweifragment
(inv. nr. NMR 9991-7661) heeft wellicht hertenvraat aan een van de geweitakken.
Vraatsporen worden hieronder behandeld bij ‘Diersporen’. Van de in Tabel 4.3
behandelde vondsten zijn 164 elementen op breuksoort bekeken. Het grootste deel
van de breuken (73%) is oud. De overige skeletelementen zijn compleet (en bevatten
zodoende geen breuken), bevatten breuken waarvan het onduidelijk is of deze recent
dan wel oud zijn (met name het geval bij fragmenten van spongieus materiaal en
schedelfragmenten), of vertonen breuken of andere beschadigingen die van recente
aard zijn. De breuken in deze laatste categorie zijn in een aantal gevallen waarschijnlijk
veroorzaakt door het vissen zelf (bijv. beschadiging door het sleepnet).
VerweringNaast de fragmentatiegraad geeft verwering van het bot een indicatie van de
conservering. De mate van verwering geeft onder andere inzicht in hoeverre sporen op
het bot nog zichtbaar zijn en hoe lang een bot aan het oppervlak heeft gelegen. In dit
deel wordt de verwering beschreven aan de hand van alle in de collectie van het nmr
bekeken skeletelementen en een selectie van materiaal dat niet is opgenomen in de
collectie (n=179, dezelfde als in Tabel 4.3 beschreven).
Over het algemeen zijn de skeletelementen van de vistochten vrij goed bewaard
gebleven. 16% vertoont nagenoeg geen tekenen van verwering (wat overigens niet
impliceert dat botten in deze categorie per se niet gefragmenteerd zijn). Wind en
zonlicht kunnen barsten en schilferen van het bot veroorzaken. Botten die aan het
oppervlak liggen verweren daardoor meestal sneller dan botten in die zich in de
bodem bevinden (Groot 2010). Slechts 42% vertoont barsten of scheuren, waarvan
de helft barsten in de ontstaansfase betreft. In enkele gevallen komen barsten samen
voor met een afgebladderd of poreus oppervlak, maar dergelijke oppervlakken zijn
ook aangetroffen zonder barsten. Het grootste deel van de skeletelementen is dus
waarschijnlijk niet lang blootgesteld geweest aan invloeden van win en zon.
Botten die gedurende langere tijd in een bepaalde positie aan het oppervlak hebben
gelegen, vertonen aan de kant waarmee deze aan het oppervlak hebben gelegen
(sterkere) vormen van verwering, terwijl de zijde die in het sediment was ingebed geen
of minder kenmerken van verwering kan hebben. Dit is bijvoorbeeld het geval voor
het dijbeen van een jonge mammoet. In Figuur 4.1 is te zien dat hiervan de caudale
zijde sterker verweerd is (en met zeepokken begroeid is geweest) dan de craniale zijde.
Slechts een klein aantal botten is sterk verweerd, zoals te zien in Figuur 4.2.
Het merendeel van het materiaal heeft een bruine kleur (85%), variërend van
beigebruin (56%), bruin (18%), donkerbruin (5%) tot grijsbruin (6%). 12% van het
materiaal heeft een zwarte kleur. Deze elementen waren meestal sterker gefossiliseerd
dan de bruine. De overige 3% had een afwijkende kleur, bijvoorbeeld de recente
vondsten. In zekere mate kan kleur dus iets zeggen over de ouderdom, maar de kleur
van skeletelementen is voornamelijk het resultaat van inwerking van bodemmineralen
en is afhankelijk van bodemomstandigheden (bijvoorbeeld wel of niet ijzerhoudend) en
waterstand (O’Connor 2004; Groot 2010). 23% van het materiaal vertoont sporen van
ijzer, bijvoorbeeld in de vorm van roestvlekjes die vaak zijn afgezet in barstjes of holtes
in het bot. Dit laatste zou een gevolg van het schoonmaakproces kunnen zijn, waarbij
ijzeraanslag op het botoppervlak wellicht is verwijderd.
Figuur 4.1 Dijbeen fragment van een jonge mammoet (inv. nr. NMR9991-7674), waarvan de caudale zijde sterker verweerd is (en met zeepokken begroeid is geweest) dan de craniale zijde. (Foto’s: M. Knul en M. Kuitems (detail))
1 cm

taFonoMIe 23
Afzettingen van ijzeroxide hebben een conserverende werking op bot. Dit geldt
ook voor bepaalde andere concreties, die er namelijk voor zorgen dat invloeden
van buitenaf minder direct in contact staan met het botoppervlak (Huisman et al.
2006; Groot 2010). In feite is deze beschermende werking vergelijkbaar met die
van mosdiertjes en zeepokken, die verderop wordt besproken. Een redelijk vaak
aangetroffen vorm van concretie op het materiaal (11%) is een samenstelling van zand
en schelpgruis en doet een beetje aan asfalt denken (Figuur 4.3).
In aquatische omgevingen speelt mechanische verwering door watertransport en
schuren van zand vaak een belangrijke rol. Deze verwering leidt doorgaans tot
afronding en polijsting, met name van de uiteinden van het materiaal, maar ook tot
1 cm
1 cm
Figuur 4.2 Verweerde wervel van het heilig been van een mammoet (inv. nr. NMR9991-7704).(Foto: M. Kuitems)
1 cm
Figuur 4.3 Pijpbeendiafyse van groot zoogdier (inv. nr. 10-059 Leiden038) met zandconcreties en mosdiertjes. (Foto: M. Kuitems)

24 Botvangen
aantasting van het botoppervlak (Andrews 1997; Huisman et al. 2006; Groot 2010). Het
skeletmateriaal dat is opgevist uit het zandgebied van de Maasvlakte 2 vertoont echter
nauwelijks afronding of polijsting door zandschuring of watertransport. Gepolijste
randen komen echter voor, maar dan door carnivorenvraat (wordt hieronder verder
toegelicht). Uitzonderingen zijn waargenomen, zoals inv. nr. NMR 9991-7669, een
basisgeweifragment van vermoedelijk edelhert (cf. Cervus elaphus; Figuur 4.4). Dit
fragment vertoont zeer sterke afronding en wekt daardoor de indruk te zijn gerold of
anderzijds getransporteerd over een langere afstand.
Diersporen
Carnivoren Het botassemblage van de vistochten levert goede bewijzen voor carnivorenac-
tiviteit (Figuur 4.5 en 4.6). Enkele vondsten zijn twijfelgevallen (o.a. inv. nr. NMR
9991-7040, -7086 en -7669), maar een flink aantal skeletelementen vertoont duidelijke
vraatsporen van grote carnivoren, zoals hyena (o.a. inv. nr. NMR 9991-7658, -7666,
-7679, -7637 (Figuur 4.6), -7049 (Figuur 4.5) en -7455). Hyena’s en andere carnivoren
geven de voorkeur aan het knagen van de poreuzere gedeelten van het bot. De
vraatsporen zijn daarom vaak te vinden op de uiteinden van botten zoals de epifysen,
waar het sponsachtige materiaal is aan- of weggevreten. Hardere, compactere
gedeelten, zoals de diafysen van pijpbeenderen, vertonen minder vaak sporen van
carnivorenvraat (Sutcliffe 1969; O’Connor 2004; Sutcliffe 1970). Hieronder worden
sporen beschreven die kenmerkend zijn voor carnivorenvraat en die zijn aangetroffen
in het botassemblage van de vistochten:
- Pitts: ronde, ondiepe sporen op het bot (vaak op de diaphysen) veroorzaakt door
de hoektanden van carnivoren.
- Punctures: littekens in de vorm van ronde kraters in het bot (vaak op de epiphysen)
die hoektanden van carnivoren achterlaten tijdens het bijten in het bot.
- Halve punctures: soms ontstaat een breuk op de plaats van een puncture, waardoor
de puncture splijt en in profiel waarneembaar zijn.
- Scores: groeven (meestal transversaal) veroorzaakt door de tanden van de
carnivoor (Voormolen 1997).
- Scooping: doordat grote carnivoren knagen en kluiven aan het zachtere,
sponsachtige materiaal, wordt dat gedeelte van het bot uitgehold. Vaak vindt
scooping plaats aan het uiteinde van een bot. De randen van de holte zijn vaak
afgerond en soms gepolijst (Sutcliffe 1969).
0,7 cm
0,7 cm
Figuur 4.4 Sterk afgerond fragment van een afgeworpen gewei (inv. nr. NMR9991-7669), vermoedelijk van edelhert. (Foto: M. Kuitems)
Figuur 4.5 Tweede teenkootje van steppewisent (inv. nr. NMR 9991-7049) met punctures aan distale deel. (Foto: M. Knul)

taFonoMIe 25
Sporen van carnivorenvraat zijn aangetroffen zowel in het materiaal dat is opgenomen
in de museumcollectie als in het overige materiaal op verschillende skeletelementen
(zoals pijpbeenderen, borstwervels, bekken en gewei) van verscheidene diersoorten
(wolharige mammoet, wolharige neushoorn, paard, edelhert, reuzenhert en
steppenwisent).
Knaagdieren en hoefdierenSporen van carnivorenvraat zijn te verwachten in een gebied als het zandwingebied
Maasvlakte 2 waar ook andere tekens van aanwezigheid van carnivoren zijn gevonden,
zoals botresten van leeuw (Panthera leo, zie Rapport 3), grottenhyena (Crocuta crocuta
spelaea) en een coproliet van een hyena (inv. nr. NMR 9991-7071). Naast carnivoren
kunnen ook andere diersoorten hun sporen nalaten op botten, zoals verschillende
knaag- en hoefdieren (O’Connor 2004; Sutcliffe 1973).
Knaagdiervraat kenmerkt zich door relatief korte, parallelle sporen van de incisieven
Figuur 4.6 Scheenbeen van jonge mammoet (inv. nr. NMR 9991-6937) met verschil-lende typen van carnivorenvraat: pitts, (halve) punctures, scores en scooping (Foto: M. Knul)
5 cm
2 cm

26 Botvangen
van het knaagdier. Hoewel botresten van knaagdieren als bever (Castor fiber, zie
Rapport 3) en woelmuis (Mimomys savini, zie Rapport 3; Microtus agrestis en Arvicola
sp., zie Rapport 1) bekend zijn uit het zandwingebied, zijn er van deze dieren geen
vraatsporen aangetroffen op botten die gevonden zijn tijdens de vistochten.
Ook skeletmateriaal van hertachtigen is veelvuldig gevonden tijdens de vistochten.
Van herten is bekend dat ze van tijd tot tijd op botten kauwen en sabbelen om hun
calcium- en fosforgehalte in hun lichaam op peil te houden, met als gevolg dat aan
het uiteinde van een bot een vorkachtige structuur ontstaat. Tijdens de vistochten zijn
geen botten aangetroffen met overtuigende sporen van hertenvraat erop. Slechts één
rendiergeweifragment (inv. nr. NMR 9991-7661) dat zeer waarschijnlijk is afgeworpen,
vertoont mogelijk hertenvraat aan de onderste geweitak. Het gewei is daarnaast op
een aantal plekken afgebroken.
DiergangenSporen op bot en gewei van een heel andere orde vormen graafgangen van
verschillende worm- en molluskensoorten (Figuur 4.7). Organismen als boormossel,
boorworm, boorkoraal en paalworm kunnen sporen in skeletmateriaal achterlaten,
bijvoorbeeld door het graven van gangen (den Ouden en Goud 2012). Onder het
opgeviste skeletmateriaal bevinden zich meerdere fragmenten waarin gangen zijn
gevormd door organismen. In enkele gevallen is de gang niet het enige spoor dat is
achtergelaten door het organisme; In Figuur 4.8 is de schelp van een boormossel nog
in het bot aanwezig. De tunnels tasten de microstructuur van skeletelementen aan,
waardoor deze kwetsbaarder worden (Groot 2010; Huisman et al. 2006).
Diersporen op het botoppervlakVeel van de botten die zijn opgevist in het zandwingebied zijn in meer of mindere mate
bedekt met sporen van dieren die in tegenstelling tot de vraatsporen en graafgangen
onschadelijk zijn voor het bot en zelfs een positieve uitwerking kunnen hebben op de
conservering van het bot. Het gaat hier om restanten van zeepokken (Cirripedia, of
rankpotigen), mosdiertjes (Bryozoa), eikapsels van slakken (Gastropoda) als fuikhorens
(Nassariidae) en hydropoliepen (Hydrozoa) (Figuur 4.9).
Zeepokken, mosdiertjes en hydropoliepen vormen zich niet ondergronds en eikapsules
worden door slakken niet ondergronds afgezet, maar alleen boven de zeebodem.
Door de aanwezigheid ervan op opgevist materiaal, kan worden vastgesteld dat
het materiaal gedurende een bepaalde periode (gedeeltelijk) aan het oppervlak
heeft gelegen (Figuur 4.10). Door de groeisnelheid van zeepokken, mosdiertjes en
hydropoliepen is het echter lastig dan wel onmogelijk om te bepalen hoe lang en
Figuur 4.7 Geweifragment (inv. nr. NMR 9991-7701) met diergangen. (Foto: M. Knul)
Figuur 4.8 Metapode van mammoet (inv. nr. NMR 9991-7677) met boormossel. (Foto: M. Knul)

taFonoMIe 27
wanneer het materiaal blootgesteld is geweest aan het zeeoppervlak: zo is een kolonie
mosdiertjes al binnen een week gevormd. Een bot dat aan het oppervlak is komen te
liggen door een net tijdens een vistocht kan, bedekt met mosdiertjes en zeepokken, bij
een latere vistocht worden opgevist. Als het tijdens de eerdere vistocht naar boven was
gekomen, had het nog vrij kunnen zijn van deze organismen.
Hoewel dergelijke diersporen het bot niet zozeer aantasten, bemoeilijken ze wel het
ontdekken en karakteriseren van andere sporen op en kenmerken van het bot (Groot
2010; Huisman et al. 2006).
Figuur 4.9 Doornuitsteeksel van borstwervel van een groot zoogdier (inv. nr. 10-059Leiden033) met (sporen van) mosdiertjes, zeepokken en eikapsels. (Foto: M. Kuitems)
Figuur 4.10 (boven) Deze halswervel van mammoet (inv. nr. NMR 9991-7662) is aan de craniale zijde (A) begroeid met mosdiertjes en vertoont ook sporen van zeepokken. De caudale zijde (B) heeft minder begroeiing van mosdiertjes, wat erop duidt dat de caudale zijde minder aan het opper-vlak heeft gelegen dan de craniale zijde. (onder) Deze borstwervel van mammoet (inv. nr. NMR 9991-7671) is aan de craniale zijde (A) onbegroeid met mosdiertjes. De caudale zijde (B) is wel begroeid met mosdiertjes. De wervel heeft dus met de craniale kant in het sediment gelegen. (Foto: M. Kuitems)
1 cm

28 Botvangen
Sporen van menselijke activiteitenEen aantal objecten dat door de mens is vervaardigd sinds de 16de eeuw na Chr. is
beschreven in het hoofdstuk Vondsten Nieuwe Tijd elders in dit rapport. Daarentegen
hebben de vistochten slechts enkele skeletelementen opgeleverd die duidelijke sporen
van menselijk handelen vertonen.
Sporen die door de mens zijn veroorzaakt op skeletmateriaal kunnen verschillende
ontstaanswijzen hebben. Het skeletmateriaal kan bijvoorbeeld gebruikt zijn voor
de voedselvoorziening van de mens en daardoor sporen bevatten van processen
als slachten, fileren, verhitten en het exploiteren van merg. Het materiaal kan ook
zijn omgevormd tot een sieraad of werktuig (Davis 1987). Het moge duidelijk zijn
dat verschillende activiteiten leiden tot verschillende sporen op het materiaal, die
in meer of mindere mate zichtbaar zijn voor het blote oog. Carnivoren en processen
als trampling kunnen resulteren in krassen die lijken op snijsporen. De conserveer-
middellaag die op het materiaal is aangebracht maakt het ontdekken van bepaalde
sporen moeilijk en soms zelfs onmogelijk en draagt eraan bij dat onderscheid tussen
snijsporen en ‘pseudo-snijsporen’ soms lastig is. Twee vondsten die duidelijke sporen
van menselijke activiteit vertonen, zullen hieronder worden besproken.
De negende track van de vistocht die plaatsvond op 26 juni 2010 heeft een ribfragment
van een rundachtige uit het Holoceen opgeleverd (inv. nr. NMR 9991-7474). Deze
vondst is niet met conserveermiddel geïmpregneerd. De rib is vrijwel compleet, maar
mist een deel van het proximale uiteinde. De rib vertoont zowel aan de binnen- als
de buitenzijde snijsporen die zijn veroorzaakt door een metalen object. De sporen
zijn V-vormig, wat karakteristiek is voor snijsporen (O’Connor 2004). De V-vormige
inkepingen zijn bedekt door een laagje roest, wat contact met een mes of een ander
metalen voorwerp hoogstwaarschijnlijk maakt.
Op 7 juni is 2011 is tijdens track 1 een deel van een edelhertgewei in het rechter net
terechtgekomen (11-064Leiden059). Het betreft een deel van het buitenste deel
van de schacht met een maximale lengte van 9,94 cm en een maximale breedte van
4,59 cm. Aan de binnenzijde is gedeeltelijk nog sponsachtige structuur aanwezig.
In het zandwingebied zijn meerdere (edelhert)geweifragmenten gevonden, maar
dit fragment is een typisch voorbeeld van ‘productieafval’. Aan weerszijden van het
fragment zijn twee duidelijke sporen waarneembaar (Figuur 4.11). Deze zijn V-vormig
en typisch voor snijsporen vervaardigd met een vuurstenen werktuig. Dergelijke
sporen maakte men om gewei in te snijden om het vervolgens te kunnen breken.
Waarschijnlijk is er tijdens het snijden of breken iets misgegaan. Op basis van typologie
is de vondst niet in een specifieke periode te plaatsen. Deze vondst is niet opgenomen
in de collectie van het nmr, maar wel geïmpregneerd met conserveringsmiddel.
Discussie en conclusieTijdens de twaalf visdagen zijn honderden skeletelementen opgevist van een
breed scala aan diersoorten. Het merendeel daarvan behoort tot de categorie
‘mammoetsteppe fauna’, maar vondsten van zeezoogdieren (zadelrob), vissen (o.a.
steur en tonijn) en holocene dieren (otter) en objecten (zie hoofdstuk Vondsten Nieuwe
Tijd) zijn ook gedaan.
Het is bekend dat de plaats van de huidige zandwinput niet altijd in zee heeft gelegen,
maar tijdens verschillende koudere perioden, waarbij veel water was opgeslagen in
de ijskappen, in een landschap lag dat werd doorkruist door een aantal grote rivieren.
De resultaten van het huidige tafonomieonderzoek, voornamelijk het feit dat de
skeletelementen niet of nauwelijks zijn afgerond, wijzen erop dat de meeste vondsten
niet ver getransporteerd zijn. De uiterlijke kenmerken van het botmateriaal wijzen erop
dat het afkomstig is van dieren die in of rondom het gebied van het zandwingebied

taFonoMIe 29
hebben geleefd en zijn gestorven. Het materiaal lijkt dus niet afkomstig te zijn van
dieren die landinwaarts hebben geleefd en gestorven zijn en waarvan skeletelementen
met de voormalige rivieren zijn meegevoerd richting de locatie van de huidige
zandwinput. In dat geval zou het materiaal sterker afgerond moeten zijn en zouden de
veelvuldige aangetroffen sporen van vraat door grote carnivoren niet of minder goed
herkend hebben kunnen worden.
Verschillende typen skeletmateriaal is teruggevonden, namelijk bot (zowel compact
bot als poreuzer, spongieus bot), gewei en gebitselementen. Hoorn is niet opgevist.
Opvallend is dat er relatief zeer weinig (stoot)tanden en kiezen zijn gevonden.
Het meeste skeletmateriaal verkeert in een goede staat van conservering. Van het
merendeel was het mogelijk om het skeletelement en de diersoort vast te stellen. Het
materiaal vertoont weinig barsten en afbladdering, wat erop wijst dat het materiaal
niet lang is blootgesteld aan wind en zonlicht. De weinige barsten en de lage mate
van afronding doen vermoeden dat het materiaal vrij snel is ingebed in de bodem, niet
ver van de plek waar het dier waarvan de skeletelementen afkomstig is, is gestorven.
In een later stadium, ten tijde van een hoge zeespiegel, heeft veel van het materiaal
onder water wel aan het bodemoppervlak gelegen. Hier vormen de (sporen van)
kolonies mosdiertjes en zeepokken, evenals eikapsels van zeeslakken aanwijzingen
voor.
Er zijn weinig sporen op het materiaal gevonden, ondanks de lage mate van afronding
en verwering. Dit impliceert echter niet dat er geen sporen op de botten aanwezig
zijn (geweest). Processen als schoonmaken en impregneren van de botten vormen
een belemmering voor het herkennen en identificeren van sporen. Ook kolonies van
mosdiertjes en zeepokken en concreties kunnen ervoor hebben gezorgd dat bepaalde
sporen aan het zicht zijn onttrokken. Desondanks is een aantal zeer duidelijke sporen
Figuur 4.11 Fragment van edelhertgewei (inv. nr. 11-064Leiden059) met snijsporen aan weerszijden. (Fotos: A. Ramcharan (boven) en M. Kuitems (onder))

30 Botvangen
van carnivorenvraat gevonden. De typische kenmerken van carnivorenvraat, zoals pitts,
(halve) punctures en scooping zijn allemaal meermaals vertegenwoordigd. Bovendien
zijn twee vondsten gedaan die bewijs vormen van menselijk handelen. Het betreft hier
een ribfragment en een geweifragment waarop evidente snijsporen waarneembaar
zijn. Daarnaast bevat een aantal skeletelementen minder duidelijke sporen die wellicht
ook toe te schrijven zijn aan sporen van menselijke activiteit en diervraat.
Om in de toekomst beter tafonomisch onderzoek te kunnen verrichten aan materiaal
dat is opgevist uit de Noordzee, is het nodig dat het onderzoek plaatsvindt als al het
materiaal van een vistocht nog bij elkaar is. Ook is het aan te raden om materiaal
van de verschillende vistochten tegelijkertijd te kunnen bestuderen, om refitten
optimaal mogelijk te maken en om stukken beter met elkaar te kunnen vergelijken.
Bovendien is het van belang dat de tafonomiestudie plaatsvindt alvorens het materiaal
wordt geïmpregneerd, zodat eventuele sporen, concreties en dierkolonies goed te
herkennen, karakteriseren en te kwantificeren zijn.

Mollusken 31
5 Mollusken
F. Wesselingh, D. De Loecker, W. Borst, O. van Tongeren
InleidingIn deze studie hebben we gekeken naar de samenstelling van de fossiele
molluskenfauna die is verzameld tijdens verschillende expedities in het zandwingebied
van Maasvlakte 2.
Met de inventarisatie willen we (1) de fauna karakteriseren, (2) verkennen of de fossiele
schelpen kunnen bijdragen in het “karteren” en karakteriseren van de stratigrafie
in het zandwingebied en (3) verkennen of er eventueel materiaal met menselijke
gebruikssporen aanwezig is.
MethodenTijdens de vistochten op 6-10 juni 2011 en 9 september 2011 is molluskenmateriaal
verzameld (Tabel 5.1). Een groot deel van de fossiele schelpen bestaat uit exemplaren
van de oester (Ostrea edulis). Deze konden gezien de snelheid van werken niet allen
worden verzameld. Per track is derhalve een kwantitatieve schatting gemaakt van de
oesters. De overige schelpen zijn allemaal verzameld, zowel de recente als de fossiele
schelpen. In dit rapport worden alleen de schelpen behandeld die we als ‘fossiel’
karakteriseren: dit zijn schelpen die ondoorzichtig/mat zijn, waarvan de primaire kleur
Soort (wetenschappelijke naam) Soort (Nederlandse naam) aantal %
Euspira catena Gewone tepelhoren 3 0
Euspira cf catenoides Tepelhoren 1 0
Buccinum undatum Wulk 43 5
Nassarius reticulatus sl Gevlochten fuikhoren 1 0
Mytilus edulis Mossel 3 0
Ostrea edulis Oester 23 3
Mimachlamys varia Bonte mantel 6 1
Anomia ephippium Paardenzadel 4 0
Astarte borealis Grote Astarte 1 0
Cerastoderma glaucum Brakwaterkokkel 7 1
Cerastoderma edule Gewone kokkel 53 6
Cerastoderma edule f major Kokkel 6 1
Acanthocardia tuberculata Geknobbelde hartschelp 504 55
Acanthocardia echinata Gedoornde hartschelp 1 0
Laevicardium oblongum crassum Noorse hartschelp 2 0
Laevicardium oblongum castanea Noorse hartschelp 34 4
Spisula subtruncata Halfgeknotte strandschelp 3 0
Spisula solida Stevige strandschelp 15 2
Spisula elliptica Ovale strandschelp 2 0
Mactra stultorum Grote strandschelp 8 1
Mactra glauca Brede strandschelp 29 3
Mactra stultorum plistoneerlandica Grote strandschelp 33 4
Lutraria lutraria sl Otterschelp 10 1
Lutraria magna Gebogen otterschelp 1 0
Macoma balthica Nonnetje 10 1
Artica islandica Noordkromp 1 0
Dosinia lupinus Dichtgestreepte artemisschelp 2 0
Dosinia exoleta Artemisschelp 1 0
Venerupis senescens Grijze tapijtschelp 6 1
Mya truncata Afgeknotte gaper 7 1
Zirfaea crispata Ruwe boormossel 93 10
Pholas dactylus Pholade 1 0
som 914 100
Tabel 5.1 Resultaten van de telmonsters.

32 Botvangen
is verdwenen en waar geen vers organisch materiaal meer in de schelp aanwezig is
waardoor de schelp enigszins bros is.
De schelpen zijn gedetermineerd door Frank Wesselingh. Bij de tellingen is
aangehouden dat elk fragment van een slak met daarin een stuk van de spil (columella)
als “1” is geteld. Elk fragment van een tweekleppige waarvan meer dan de helft van het
slot bewaard is, is als “1/2” geteld (daar twee kleppen een exemplaar vormen). Soorten
waarvan alleen een fragment is gevonden dat niet aan een van bovenstaande criteria
voldoet, zijn opgenomen als respectievelijk “1” of “1/2” exemplaar. De aantallen van
de tweekleppigen zijn vervolgens naar boven afgerond op het hele getal om tot een
minimum aantal individuen te komen.
De volgende afkortingen worden gebruikt:
f – forma
sl – determinatie in de ruime zin
Zo omvat ‘Nassarius reticulatus sl’ de soort Nassarius reticulatus en Nassarius nitidus,
omdat de juveniele stadia in afgesleten materiaal vaak niet te onderscheiden zijn. De
slotfragmenten van Lutraria lutraria en L. angustior zijn ook niet te onderscheiden en
bijeengebracht als ‘Lutraria lutraria sl’.
Tijdens de verwerking van het materiaal is voor een aantal algemene soorten een
kwalitatieve beoordeling gemaakt van de conserveringstoestand.
ResultatenIn totaal zijn 144 monsters bekeken (Tabel 5.1), waarin 32 taxa (soorten en herkenbare
vormen van soorten) werden aangetroffen.
De fossiele schelpenfauna wordt gedomineerd door de geknobbelde hartschelp
(Acanthocardia tuberculata; Figuur 5.1) die 55% van de vondsten uitmaakt. Deze
relatief grote soort is een indicator voor Eemien-afzettingen. Vandaag de dag komt
de soort voor van het Kanaalgebied en de Britse eilanden tot aan Noord-Afrika en in
de Middellandse Zee. De exemplaren tonen grote verschillen in conserveringsgraad,
van sterk afgesleten (indicatief voor omwerking) tot uitzonderlijk goed geconserveerd.
Het voorkomen van zeer goed geconserveerde exemplaren (geen slijtage aan de top,
fijne sculptuurdetails goed bewaard, in verschillende monsters resten van kleurbanden
aanwezig) in bepaalde opeenvolgende tracks (gebieden) doet vermoeden dat ter
plaatse in situ Eemien-afzettingen in het wingebied aanwezig zijn.
Andere algemene soorten zijn de ruwe boormossel (Zirfaea crispata: 10%), de kokkel
(Cerastoderma edule: 6%), wulk (Buccinum undatum: 5%), de “zuidelijke” Noorse
hartschelp (Laevicardium oblongum castaneum: 4%) en de fossiele grote strandschelp
(Mactra stultorum plistoneerlandica: 4%). De laatste twee soorten zijn Eemien-
indicatoren, de eerste drie soorten kunnen zowel een laat-pleistocene als (vroeg-)
holocene ouderdom hebben.
Aangezien vrijwel uitsluitend grote schelpen zijn opgevist, is het karakter van de
fauna anders dan die aangetroffen is op de strandvakken van de buitencontour en in
de boringen (zie respectievelijk Rapport 3 en Rapport 2 van het geo-archeologisch
en paleontologisch onderzoek zandwingebied en buitencontour Maasvlakte 2). De
opgeviste fauna heeft een duidelijke ‘bias’ in de vorm van oververtegenwoordiging van
grote soorten, en daarbinnen blijken Eemien-indicatoren (Acantohocardia tuberculata,
Mactra stultorum plistoneerlandica, Mactra glauca, Anomia ephippium, Mimachlamys
varia, Lutraria magna, Pholas dactylus en Venerupis senescens) ten opzichte van
de andere fauna’s oververtegenwoordigd te zijn. Een ‘koele’ (koud-gematigde)

Mollusken 33
faunacomponent (Cerastoderma edule f major en Astarte borealis) is sterk onderverte-
genwoordigd in het materiaal, vermoedelijk omdat de betreffende soorten over het
algemeen zo klein zijn dat ze door de mazen van de netten vallen. We hebben geen
vertegenwoordigers van zoetwater en terrestrische fauna gevonden en materiaal van
brakwatersoorten (b.v. brakwaterkokkel, Cerastoderma glaucum) is zeldzaam.
Gezien het voorkomen van zeer goed geconserveerde Eemien-indicatoren is het
aannemelijk dat niet of nauwelijks omgewerkte mariene Eemien-afzettingen aanwezig
zijn in de gebieden van de volgende vistracks: 6-6-2011 tracks 11-13 (mogelijk ook in
enkele van de tracks 3-10), 7-6-2011 tracks 1-6 (mogelijk ook track 8), 8-6-2011 tracks
1-2 en 9-14, 9-6-2011 tracks 1-8 (mogelijk ook tracks 11 en 12) en 10-6-2011 tracks 6-15.
In de fauna zijn geen uitzonderlijk zeldzame soorten gevonden. Een sterk afgesleten
exemplaar van Arctica islandica (6-6-2011 track 11 links) is vermoedelijk een fluviatiel
omgewerkt exemplaar dat afkomstig is uit de vroeg-pleistocene Maassluis Formatie
of de pliocene Oosterhout Formatie die in de ondiepe ondergrond dagzomen in het
zuidelijke Zeeuwse deltagebied.
Discussie
Karakter van de faunaDoor de afmetingen van de mazen van de netten waarmee is gevist, omvat de
fossiele schelpenfauna die tijdens de vistochten is verzameld voornamelijk relatief
grote soorten, waarin Eemien-indicatoren, en met name de geknobbelde hartschelp
(Acanthocardia tuberculata) domineren. De soorten die zijn aangetroffen tijdens de
vistochten worden weliswaar ook op de strandvakken van de buitencontour gevonden,
maar de samenstelling (abundance) is wezenlijk anders. Vrijwel alle kleinere soorten (<
4 cm) die we kennen van de strandvakken ontbreken in het geviste materiaal.
Karakteriseren en karteren van de ondergrond van het zandwingebiedUit parallelstudies aan opgespoten materiaal en boringen in het zandwingebied is
gebleken dat er minimaal vier verschillende fossielhoudende geologische eenheden
voorkomen, te weten (1) holocene mariene afzettingen, (2) laat-pleistocene fluviatiel
omgewerkte en gemengde mariene, brak en zoetwater en terrestrische fauna’s, (3)
laat-pleistocene koel-gematigde mariene fauna’s en (4) midden-vroeg pleistocene
fluviatiele fauna’s. In die onderzoeken is wel veel omgewerkt Eemien-materiaal
gevonden maar zijn geen aanwijzingen voor in situ Eemien-fauna’s aangetroffen.
In het materiaal van de vistochten zijn in vergelijking met de resultaten van
bovengenoemde parallelstudies verschillende groepen sterk ondervertegenwoordigd
of helemaal afwezig, maar tegelijkertijd lijkt het aannemelijk dat in situ Eemien-
Figuur 5.1 Acanthocardia tuberculata (Linneaus, 1758). Breedte 54 mm, track 4-rechts 7-6-2011. Voorbeeld van een goed geconserveerd exemplaar waar de fijne oppervlakte details van de sculptuur goed bewaard zijn gebleven en de top van de schelp (umbo) niet is af-gesleten. (Foto: F. Wesselingh)

34 Botvangen
afzettingen wel degelijk voorkomen in het zandwingebied.
De fossiele mollusken uit de vistochten geven derhalve geen representatief beeld van
de opbouw van de ondergrond maar geven wel belangrijke aanvullende informatie ten
opzichte van boringen en strandvondsten.
Sporen van menselijke bewoningWe hebben geen schelpen en schelpresten gevonden die wijzen op menselijk gebruik
(bijv. als ornament).
ConclusiesDe fossiele schelpenfauna’s die zijn verzameld tijdens vistochten in het zandwingebied
van Maasvlakte 2 hebben een sterk ‘gebiassed’ karakter ten voordele van grote
soorten. In de fauna zijn mariene Eemien-soorten dominant. De goede conservering
in verschillende monstergebieden geeft aan dat in situ mariene Eemien-afzettingen
ter plekke in de ondergrond aanwezig moeten zijn. Sporen van menselijk gebruik van
schelpen hebben we niet aangetroffen.

vondsten nIeuwe tIjd 35
6 Vondsten nieuwe tijd
H.J.T. Weerts, J. Opdebeeck, A. Otte, J. van Doesburg
InleidingBij de vistochten in het zandwingebied van Maasvlakte 2 werden naast (fossiele)
botten, mollusken en stenen ook allerlei vondsten uit de nieuwe tijd (jonger dan
1492 na Chr.) opgevist. Het merendeel van dit vondstmateriaal is archeologisch niet
interessant. Twee vondsten zijn wel degelijk als archeologisch relevant te beschouwen:
(1) enkele houten spantjes van een klein vaartuig en (2) een baardmankruikje.
Die vondsten zijn ook in ARCHIS gemeld (Bijlage Hoofdstuk 6.1). In dit hoofdstuk
worden ze beschreven. Besloten wordt met enkele afbeeldingen van (zeer) jong
vondstmateriaal.
Spantjes van een klein vaartuigOp 1 oktober 2009 zijn door de kotter OD-7 “Adrianus” (Stellendam, schipper Jaap
Klein) enkele eikenhouten spantjes van een klein vaartuig opgevist (zie Bijlage
Hoofdstuk 6.2, track 10 voor de exacte locatie). De grootte en dikte van het hout
geven aan dat de spantjes waarschijnlijk afkomstig zijn van een klein vaartuig of een
grote sloep. De spanten zijn onderdeel van de structuur/het skelet van het schip.
Uit hun vorm is af te leiden dat het schip overnaads is gebouwd; de planken van
de scheepshuid liggen bij deze bouwwijze dakpansgewijs over elkaar. Huidplanken
van het vaartuig in kwestie zijn aan de spanten bevestigd geweest door middel van
houten pennen. Er zijn in de aangetroffen spanten geen ijzeren pennen gevonden.
De scheepshuid werd waterdicht gemaakt door de spleten tussen de planken op te
vullen met moslatjes (houten latjes of spanen) en met breeuwsel gemaakt uit teer,
mos, paardenhaar etc. Een ander constructiekenmerk van de spanten is dat aan de
onderzijde ervan waterloosgaten (loggaten) aanwezig zijn. Door deze gaten kon het
lenswater naar het laagste punt in het vaartuig stromen, waar het werd gehoosd of
met behulp van een (lens)pomp overboord werd gepompt. De aangetroffen restanten
van de spanten komen dus van de bodem van het schip. Figuren 6.1 en 6.2 geven de
Figuur 6.1 De opgeviste spanten aan boord van de OD 7 op 1 oktober 2009. (Foto: J. Opdebeeck)

36 Botvangen
opgeviste spanten weer. De spanten zijn in bezit van het Havenbedrijf Rotterdam en
bij het ter perse gaan van dit rapport te zien in Futureland, het bezoekerscentrum van
Maasvlakte 2.
De overnaadse bouw en de houten pennen zouden kunnen wijzen op restanten van
een schip van noordelijke breedte, mogelijk zelfs een Vikingschip. Om die reden zijn
uit drie spanten dwarsdoorsnedes gezaagd (Figuur 6.3) opdat met behulp van de
jaarringen de ouderdom kon worden vastgesteld. Hoewel de monsters over voldoende
jaarringen beschikken voor dendrochronologisch onderzoek, leverde dit helaas geen
ouderdom op (Bijlage Hoofdstuk 6.3a,b; RING intern Rapport nummer 2010010;
2010072, Domínguez Delmás 2010). Wel werd geconstateerd dat de zeer dichte ligging
van de jaarringen mogelijk wijst op een (zeer) noordelijke standplaats van de eiken
waaruit de spanten zijn vervaardigd (mondelinge mededeling M. Domínguez Delmás).
Om die reden is één van de kernen naar het 14C-laboratorium van de Rijksuniversiteit
Groningen gestuurd voor een ouderdomsdatering met behulp van 14C.
Voor de ouderdomsbepaling is gebruik gemaakt van een onbekend aantal ringen aan
de buitenkant van het monster; de jongste ringen. Deze 14C-datering leverde (na ijking)
een ouderdom op van 150 +- 25 jaar BP; ofwel jonger dan ca. 1670 AD (zie Bijlage
Hoofdstuk 6.4; brief van Prof.dr.ir. J. van der Plicht dd. 18 november 2011; Figuur 6.4).
Het baardmankruikjeTijdens de vistocht van 9 september 2011 is in track 14 aan de rechterkant van het
schip een vrijwel compleet baardmankruikje aangetroffen (Figuur 6.5). Het oor en de
bovenkant waren afgebroken. De breuken waren niet vers, waaruit is op te maken dat
het afbreken niet ten gevolge van de zandwinning of het bottenvissen is gebeurd. Het
kruikje was gevuld met sediment, dat is gezeefd over een 1 mm zeef. Het zeefresidu
Figuur 6.2 De spanten in de spoelbak van de RCE in Lelystad. (Foto: L. Kuhler)
Figuur 6.3 De voor dendrochronologisch onderzoek verzaagde spant. (Foto: L. Kuhler)

vondsten nIeuwe tIjd 37
bevatte geen restanten die aan een oorspronkelijke inhoud van de kruik toegewezen
konden worden, maar uitsluitend fragmenten van schelpen.
De kruik is gefabriceerd van grijs-wit steengoed, typerend voor het aardewerkproduc-
tiecentrum van Frechen bij Keulen. De kruik toont een mannenkop met baard, waaraan
het ook de naam baardmankruik te danken heeft (Friedrich 1967). Daaronder bevindt
zich een medaillon, met gestileerd bloemmotief. Op het aardewerk is, met name op de
bovenste helft, zoutglazuur aangebracht. Op basis van de stijlkenmerken van zowel de
mannenkop als het medaillon is de kruik te dateren in het 2e/3e kwart van de 17e eeuw.
Aangezien de kruik in een vistrack op de flanken van een zandwinput is gevonden, is
de oorspronkelijke archeologische context niet te achterhalen. Gezien de situering
midden op zee is het duidelijk dat het stuk aardewerk zich aan boord van een schip
heeft bevonden. Baardmankruiken werden algemeen gebruikt als keukenwaar en
schenkgerei, ook aan boord van schepen. Het is dus mogelijk dat de kruik tot de
inventaris van een schip heeft gehoord. Baardmankruiken werden echter ook op grote
schaal naar overzeese gebieden geëxporteerd. De kruik kan dus ook deel hebben
uitgemaakt van een handelslading. De kruik is in bezit van het Havenbedrijf Rotterdam
en is bij het ter perse gaan van dit rapport te zien in Futureland, het bezoekers-
centrum van Maasvlakte 2. De vondsten, zowel de spantjes als het kruikje, hebben
geen depotnummer gekregen. Het Havenbedrijf Rotterdam beslist als eigenaar over
eventuele toekomstige opslag van de vondsten.
Overig jong vondstmateriaalBij de vistochten op zoek naar botmateriaal is natuurlijk ook erg veel ander materiaal
naar boven gehaald, vooral waar met fijnmazige netten werd gevist (Figuur 6.6).
Veelal betreft het brokken veen en stukken hout, die in algemene zin vaak zijn
gedocumenteerd in de logboeken (zie bijvoorbeeld Bijlage 6.2). Enkele vermeldens-
waardige ‘vondsten’ zijn die van twee stokankers door de OD-7 op 30 september en 1
oktober 2009 (Bijlage 6.2; Figuur 6.7). Deze ankers zijn niet als archeologische vondst
aangemeld. Voor eventueel nader onderzoek zijn enkele monsters genomen van het
veen en van het hout. Onderzoek hieraan heeft (nog) niet plaatsgevonden.
Figuur 6.4 Met Intcal09 (Reimer et al. 2009) gecali-breerde ouderdom van de 14C-datering van één van de spanten. Door de aanwezigheid van een 14C-“platform” kan de ouderdom niet nauwkeuriger worden bepaald.

38 Botvangen
Naast deze grote vondsten is veel kleiner materiaal opgevist. Op Figuur 6.7 zijn o.a.
een fietsstuur en een stuk metaal te zien; een verbogen metalen platform met wieltjes.
Dat laatste is mogelijk afkomstig van de verankering van een later opgeruimde zeemijn
(Figuur 6.8).
Ook is verloren gegaan speelgoed gevonden (Figuur 6.9). Omdat dit (zeer) jonge
materiaal buiten het kader van het onderzoek valt en omdat het buiten context is
opgevist, is dit niet verder gedocumenteerd.
Figuur 6.5 Het op 9 september 2011 aangetroffen baardmankruikje. (Foto: A. Otte)
Figuur 6.6 “Volle bak”, resultaat van één vistrack. (Foto: J. Opdebeeck)

vondsten nIeuwe tIjd 39
Figuur 6.7 Foto van één van de opgeviste stokankers aan boord van de OD 7. (Foto: J. Opdebeeck)
Figuur 6.8 Zeemijn op de openlucht tentoonstelling van marine wapens, Hel (Polen). De mijn rust op een stalen onderstel met wieltjes. (Foto: Topory / Wikipedia)
Figuur 6.9 Dit smurfachtige poppetje is niet blauw, maar groen! Opgevist door de OD 7 op 9 juni 2011. (Foto: D. De Loecker)

40 Botvangen
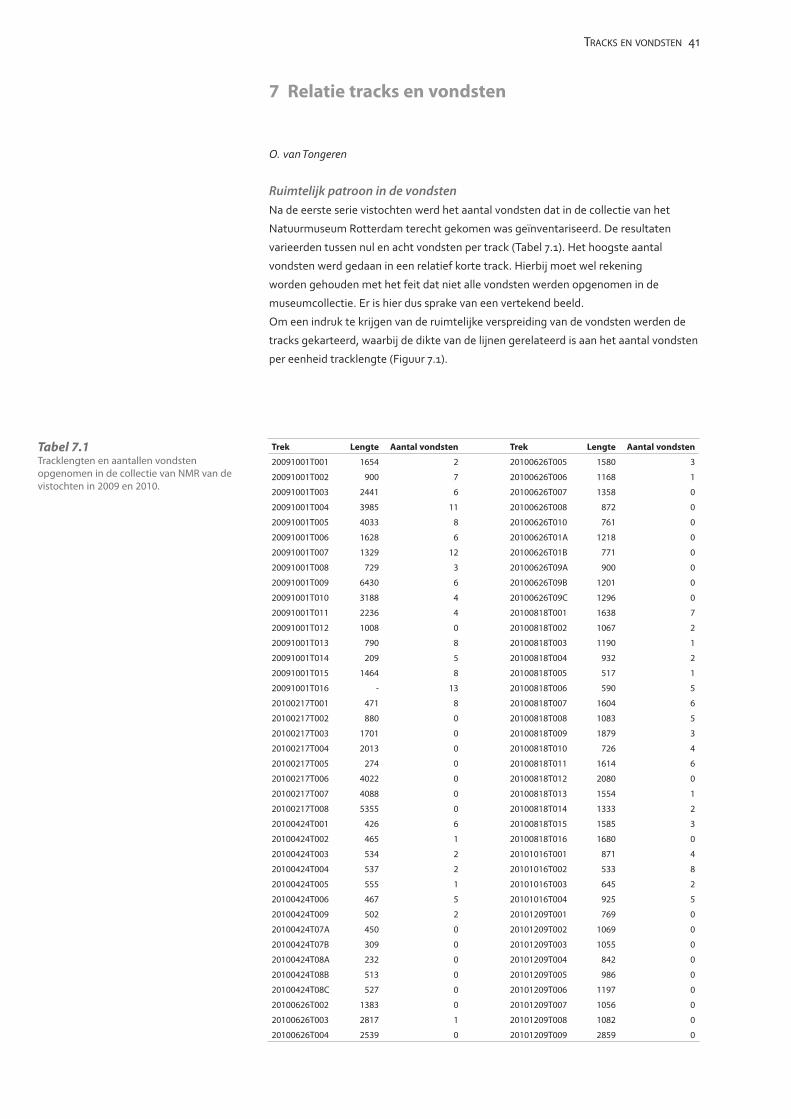
tracks en vondsten 41
7 Relatie tracks en vondsten
O. van Tongeren
Ruimtelijk patroon in de vondstenNa de eerste serie vistochten werd het aantal vondsten dat in de collectie van het
Natuurmuseum Rotterdam terecht gekomen was geïnventariseerd. De resultaten
varieerden tussen nul en acht vondsten per track (Tabel 7.1). Het hoogste aantal
vondsten werd gedaan in een relatief korte track. Hierbij moet wel rekening
worden gehouden met het feit dat niet alle vondsten werden opgenomen in de
museumcollectie. Er is hier dus sprake van een vertekend beeld.
Om een indruk te krijgen van de ruimtelijke verspreiding van de vondsten werden de
tracks gekarteerd, waarbij de dikte van de lijnen gerelateerd is aan het aantal vondsten
per eenheid tracklengte (Figuur 7.1).
Trek Lengte Aantal vondsten Trek Lengte Aantal vondsten
20091001T001 1654 2 20100626T005 1580 3
20091001T002 900 7 20100626T006 1168 1
20091001T003 2441 6 20100626T007 1358 0
20091001T004 3985 11 20100626T008 872 0
20091001T005 4033 8 20100626T010 761 0
20091001T006 1628 6 20100626T01A 1218 0
20091001T007 1329 12 20100626T01B 771 0
20091001T008 729 3 20100626T09A 900 0
20091001T009 6430 6 20100626T09B 1201 0
20091001T010 3188 4 20100626T09C 1296 0
20091001T011 2236 4 20100818T001 1638 7
20091001T012 1008 0 20100818T002 1067 2
20091001T013 790 8 20100818T003 1190 1
20091001T014 209 5 20100818T004 932 2
20091001T015 1464 8 20100818T005 517 1
20091001T016 - 13 20100818T006 590 5
20100217T001 471 8 20100818T007 1604 6
20100217T002 880 0 20100818T008 1083 5
20100217T003 1701 0 20100818T009 1879 3
20100217T004 2013 0 20100818T010 726 4
20100217T005 274 0 20100818T011 1614 6
20100217T006 4022 0 20100818T012 2080 0
20100217T007 4088 0 20100818T013 1554 1
20100217T008 5355 0 20100818T014 1333 2
20100424T001 426 6 20100818T015 1585 3
20100424T002 465 1 20100818T016 1680 0
20100424T003 534 2 20101016T001 871 4
20100424T004 537 2 20101016T002 533 8
20100424T005 555 1 20101016T003 645 2
20100424T006 467 5 20101016T004 925 5
20100424T009 502 2 20101209T001 769 0
20100424T07A 450 0 20101209T002 1069 0
20100424T07B 309 0 20101209T003 1055 0
20100424T08A 232 0 20101209T004 842 0
20100424T08B 513 0 20101209T005 986 0
20100424T08C 527 0 20101209T006 1197 0
20100626T002 1383 0 20101209T007 1056 0
20100626T003 2817 1 20101209T008 1082 0
20100626T004 2539 0 20101209T009 2859 0
Tabel 7.1 Tracklengten en aantallen vondsten opgenomen in de collectie van NMR van de vistochten in 2009 en 2010.

42 Botvangen
Figuur 7.1 laat duidelijk zien dat de tracks met de OD-7 verspreid zijn over de
zandwinput en daarbuiten (rechte lijnen, trekken gedaan in 2009) en dat de tracks
met de BRA-7 vooral aan de rand van de put gedaan zijn. Dit heeft te maken met het
feit dat tussen de vistocht met de OD-7 in oktober 2009 en de eerste vistocht met
de BRA-7 de put veel dieper is geworden. Omdat vondsten verwacht werden op een
diepte van ca. 30 m is door de BRA-7 vanaf het begin meestal langs de rand van de
zandwinput gevist. De enkele langere tracks die over de bodem van de zandwinput
gedaan zijn leverden niets op en vooral langs de randen van de put werd veel materiaal
aangetroffen. In april 2010 en juni 2010 werden voor het eerst pogingen ondernomen
om vindplaatsen meer gedetailleerd te lokaliseren door kortere vistracks te maken
Figuur 7.1 Overzicht van de beviste tracks in de jaren 2009 en 2010. De aantallen vondsten per lengte-eenheid zijn aangegeven door de lijndikte (hoe dikker, hoe hoger het aantal vondsten). Stippellijnen zijn tracks waaruit geen enkele vondst in de collectie van het NMR is opgenomen. Kortere vistracks nabij de Maascenterboei en in de zuidelijke zandwin-put zijn in rood aangegeven.

tracks en vondsten 43
nabij de Maascenterboei (in de buurt van de locatie 53000,449000). Deze tracks zijn in
Figuur 7.1 in rood aangegeven, evenals de korte tracks die in december 2010 gedaan
zijn in de zuidelijke zandwinput. Helaas leverden deze tracks geen vondsten op.
Voorafgaand aan de visweek in juni 2011 werd besloten om een kansrijk deel van het
gebied nader in kaart te brengen. Gekozen werd voor de noordoostrand van de put,
omdat daar zowel in lange tracks over dieptelijnen als in dwarstracks (in de richting
van de helling) vondsten waren gedaan. Tevens werd besloten om alle vondsten te
noteren, dus niet alleen de vondsten die in de collectie werden opgenomen. Tijdens
deze week is ook uitvoerig gemeten met seismische apparatuur en side scan sonar.
Zoals blijkt uit Figuur 7.2 (links boven) is op 6 juni begonnen met een aantal tracks
tegen de helling op om een eerste indruk te krijgen van de kans op vondsten
langs de noordoostrand van de zandwinput. De track die verder van de rand van
de zandwinput is gesitueerd, ligt daar, omdat ter plekke een “taartpunt” zand is
blijven staan (zie Figuur 2.1). Aan de oostzijde (Figuur 7.2 rechts boven, 7 juni 2011),
waar de meeste vondsten waren gedaan, werden vervolgens tracks gedaan langs
dieptelijnen. Gedurende de rest van de week is ook de rest van de noordoostrand
meer gedetailleerd in kaart gebracht. Na uitwerking van de gegevens bleek inderdaad
het zwaartepunt van de vondsten in de oostelijke hoek van de zandwinput te liggen.
Daarom werd besloten om op 9 september nogmaals een vistocht te houden om in
die hoek de gegevens te vervolledigen door middel van een aantal tracks langs de
oostrand (Figuur 7.2 links onder).
In Bijlage Hoofdstuk 7 staat een tabel met alle vondsten van 2011. De vondsten zijn
voor het oostelijk deelgebied samengevat in tabellen met het totaal aantal tracks en
het totaal aantal vondsten, ingedeeld in 1 m brede diepteklassen en in ca. 200 m brede
stroken van de helling, die ongeveer samenvallen met de tracks 7, 8, 9 en 10 van 6 juni.
Dit zijn de vier meest oostelijke tracks tegen de helling van de zandwinput op (Figuur
7.2 links boven). De Figuren 7.3 t/m 7.5 geven een overzicht van het gemiddeld aantal
vondsten per net. De vondsten worden in meer detail beschreven in de hoofdstukken
over fossiele zoogdieren, mollusken en lithische artefacten elders in dit rapport.
Er is een duidelijk maximum in de botvangsten rond de 26 tot 27 m diepte en in de
strook ter hoogte van track nummer 9 (Figuur 7.3). Een overeenkomstig maximum in
de richting van west naar oost, maar dan verspreid over twee stroken (8 en 9) wordt
Figuur 7.2 Paleontologische vistracks juni en september 2011. Van links boven naar rechts onder 6-6, 7-6, 8-6, 9-6, 10-6 en 9-9. De rand van de zandwinput is aangegeven met een rode streepjeslijn.

44 Botvangen
Schelpen
Diepte 7 8 9 10 rij /trek22.5 9 2 5.5 5.523.5 10 2.3 8.7 8.66666724.5 31 14 22 20 21 21.3333325.5 20 81 12 9 24 23.8333326.5 44 60 103 21 66 66.227.5 18 56 66 14 39 38.8333328.5 23 31 28 77 40 39.7529.5 15 17 57 28 29 28.830.5 7 8 7 3 6.3 6.2531.5 20 32 123 1 44 44kolom 20 37 39 20 29 29.20833
0 20 40 60 80
22.523.524.525.526.527.528.529.530.531.5
01020304050
7 8 9 10
Figuur 7.4 Tabel met bijbehorende staafdiagrammen van het gemiddeld aantal schelpen per net per track. Van west (kolom “7”) naar oost (kolom “10”) en van ondiep (22-23m, “22.5”) naar diep (31-32m, “31.5”). De per kolom (strook) gemiddelde aantallen vondsten staan in de rij “kolom”, de per rij (diepteklasse) gemid-delde aantallen vondsten in de kolom “rij”. De staafdiagrammen geven de marginale gemid-delden (gemiddelden per diepteklasse of 200m brede strook) weer.
Stenen
Diepte 7 8 9 10 rij /trek22.5 1 0 0.5 0.523.5 0 0.7 0.3 0.33333324.5 0 0 1 0 0.3 0.33333325.5 4 1 0.5 0.5 1.2 1.16666726.5 3 8 9.5 0 6 627.5 3 1 12 3.5 5.7 5.66666728.5 8 23 30 3 16 1629.5 12 8 8 0 5.6 5.630.5 5 4 2 0 2.8 2.7531.5 10 10 22 0 11 10.5kolom 4.6 6.9 6.8 0.8 4.6 4.604167
0 5 10 15 20
22.523.524.525.526.527.528.529.530.531.5
0
2
4
6
8
7 8 9 10
Figuur 7.5 Tabel met bijbehorende staafdiagrammen van het gemiddeld aantal stenen per net per track. Van west (kolom “7”) naar oost (kolom “10”) en van ondiep (22-23m, “22.5”) naar diep (31-32m, “31.5”). De per kolom (strook) gemiddelde aan-tallen vondsten staan in de rij “kolom”, de per rij (diepteklasse) gemiddelde aantallen vondsten in de kolom “rij”. De staafdiagrammen geven de marginale gemiddelden (gemiddelden per diepteklasse of 200m brede strook) weer.
Botten
Diepte 7 8 9 10 rij /trek22.5 0 0 0 023.5 0 1.3 0.7 0.66666724.5 1 0 2 1 1.2 1.16666725.5 1 0 1 0 0.5 0.526.5 6 7 12 0 7.2 7.227.5 3 6 1 2 2.5 2.528.5 3 3 1 1 2 229.5 1 0 6 2.5 2.4 2.430.5 0 1 4 1 1.5 1.531.5 1 0 8 0 2.3 2.25kolom 1.6 2.1 3.4 0.9 2.1 2.083333
0 2 4 6 8
22.523.524.525.526.527.528.529.530.531.5
0
1
2
3
4
7 8 9 10
Figuur 7.3 Tabel met bijbehorende staafdiagrammen van het gemiddeld aantal botten/botfragmenten per net. Van west (kolom “7”) naar oost (kolom “10”) en van ondiep (22-23m, “22.5”) naar diep (31-32m, “31.5”). De per kolom (strook) gemiddelde aantallen vondsten staan in de rij “kolom”, de per rij (diepteklasse) gemiddelde aantallen vondsten in de kolom “rij”. De staafdi-agrammen geven de marginale gemiddelden (gemiddelden per diepteklasse of 200m brede strook) weer.

tracks en vondsten 45
gevonden voor de schelpen (Figuur 7.4) en stenen (Figuur 7.5). Het maximum aantal
schelpen wordt, net als het maximum aantal botten gevonden op een diepte van 26-27
m (Figuur 7.4). Het maximum aantal stenen wordt op 2 m hogere diepte gevonden
(Figuur 7.5).
Op de locatie van de maximale aantallen vondsten was in de side scan sonarbeelden
iets te zien dat leek te wijzen op de aanwezigheid van een stroomgeul. De vondsten
op deze plaats waren de aanleiding om hier langs de helling van de zandwinput een
aantal boringen te doen, waarover in het rapport Geofysisch onderzoek Maasvlakte 2
geschreven wordt.
Samenhang tussen de locaties van de vondstenOp basis van het waargenomen ruimtelijke patroon is de samenhang tussen de
vondsten nader geanalyseerd. Er (b)lijkt een verband te zijn tussen het aantal schelpen
per net per track en het aantal vondsten van gewervelden (botten/botfragmenten). Dit
verband is duidelijker dan het verband tussen het aantal stenen en het aantal botten
(Figuur 7.6). Voorafgaand aan de analyse zijn de gegevens wortelgetransformeerd om
de varianties homogeen te maken.
Er is echter wel een duidelijk verband tussen het aantal stenen dat aangetroffen
wordt 2 m onder de plaats waar de botten en schelpen vandaan komen (Figuur 7.7).
Figuur 7.6 Het geschatte lineaire verband tussen het aantal gevonden botten/botfragmenten (horizontale as) en het aantal schelpen en stenen.
Figuur 7.7 Het geschatte lineaire verband tussen het aantal gevonden botten/botfragmenten (horizontale as) en het aantal schelpen en stenen dat ongeveer twee meter lager op de helling wordt aangetroffen.

46 Botvangen
We concluderen dat in dit deelgebied botten en schelpen gezamenlijk aangetroffen
worden boven plekken waar veel stenen gevonden zijn. De verdere interpretatie van
deze samenhang laten wij graag over aan de geologen en paleontologen.
Een soortgelijk verband tussen het voorkomen van botten, stenen en schelpen
werd gevonden in het gebied bij de “taartpunt” zand die aan de noordzijde van de
zandwinput is blijven staan. Omdat daar minder waarnemingen beschikbaar zijn en het
aantal vondsten lager is, is echter een andere methode gebruikt.
De chi-kwadraattoets voor onafhankelijkheid (toepassing niet geheel valide in verband
met lage verwachte aantallen in enkele cellen) wees uit dat aantallen botten en
schelpen verschillen van de verwachte aantallen, maar dat de verdeling van de stenen
over de tracks niet duidelijk afwijkt van hetgeen verwacht wordt op basis van de rij-
(diepte-) en kolomtotalen.
Op basis van de marginale totalen van de vondsten in drie diepteklassen van 2 m en
in drie stroken zijn verwachte waarden uitgerekend voor het aantal botten stenen en
schelpen in elk van de negen cellen (drie diepteklassen maal drie stroken). Vervolgens
zijn deze verwachte waarden afgetrokken van de geobserveerde aantallen en zijn
correlaties berekend tussen deze residuen (Tabel 7.2 en 7.3).
Hier zijn dus niet de aantallen vondsten van schelpen en botten duidelijk gecorreleerd
met de aantallen vondsten van stenen twee meter lager, maar gaat wel een hoger
dan verwacht aantal botten samen met een hoger dan verwacht aantal schelpen en
met een hoger dan verwacht aantal stenen twee meter daaronder. Het residu van het
aantal schelpen is echter negatief gecorreleerd met het residu van het aantal stenen
twee meter daaronder. Mede gezien de lage aantallen waarnemingen is de conclusie
voor dit gebied niet duidelijk.
Botten Stenen Schelpen
Botten 1
Stenen 0.0906 1
Schelpen 0.4403 0.1843 1
Botten Stenen Schelpen
Botten 1
Stenen 0.2681 1
Schelpen 0.5104 -0.6628 1
Tabel 7.2 Correlatie tussen residuen (verschillen tussen geobserveerde en verwachte aantallen) van botten, schelpen en stenen (3*3 tabel, dus slechts 4 vrijheidsgraden!)
Tabel 7.3 Correlatie tussen residuen (verschillen tussen geobserveerde en verwachte aantallen) van botten, schelpen en stenen twee meter lager

dIscussIe en conclusIe 47
8 Discussie en conclusie
M. Kuitems
De vistochten, die hebben plaatsgehad op verschillende plaatsen en diepten in het
zandwingebied, hebben veel materiaal opgeleverd met een uiteenlopende datering.
Naast grote hoeveelheden levende vissen, levende zeesterren en veenbrokken zijn
skeletelementen van zoogdieren en vissen, mollusken, gebruiksvoorwerpen en grind
aangetroffen. De skeletelementen, mollusken en voorwerpen zijn in dit rapport
beschreven. Bijna alle skeletelementen van de gewervelden behoren tot zoogdieren
die kenmerkend zijn voor de mammoetsteppe: een groot ecosysteem dat zich
gedurende het Laat-Pleistoceen uitstrekte in de noordelijke contreien van Noordwest-
Europa via het voormalige Beringia tot aan Alaska. Het merendeel hiervan is afkomstig
van wolharige mammoet. Hoewel slechts een klein deel van het materiaal behoorde
tot carnivoren (vier skeletelementen en een coproliet), vormen de vraatsporen op
botten van de herbivoren bewijs dat (grote) carnivoren veelvuldig voorkwamen in de
regio van het huidige zandwingebied.
De uiterlijke kenmerken van het skeletmateriaal, zoals de zeer lage mate van
afronding, wijzen erop dat de meeste vondsten niet ver getransporteerd zijn, waaruit
geconcludeerd kan worden dat de botten behoorden tot dieren die in de omgeving
van de huidige zandwinput hebben zijn gestorven. Ten tijde van een hoge zeespiegel,
wellicht in de periode voorafgaand aan of tijdens de baggerwerkzaamheden, heeft veel
van het materiaal onder water wel aan het bodemoppervlak gelegen.
Opvallend is dat typische Eemien-componenten in de opgeviste zoogdierfauna
ontbreken, terwijl de fossiele schelpenfauna die tijdens de vistochten is verzameld
wordt gedomineerd door Eemien-indicatoren, zoals de geknobbelde hartschelp. De
goede conservering van de deze mollusken duidt erop dat in situ mariene Eemien-
afzettingen ter plekke in de ondergrond aanwezig moeten zijn. Op dit moment is
voor de discrepantie tussen de zoogdierfauna (zonder Eemien-indicatoren) en de
molluskenfauna (met Eemien-indicatoren) nog geen verklaring gevonden. Het feit
dat de Eemien-indicatoren dominant zijn in de opgeviste molluskenfauna kan echter
worden verklaard dat een aantal soorten dat veel voorkwam in het Eemien relatief
groot is; kleinere soorten (<4 cm) zijn vermoedelijk door de mazen van het net
gevallen.
Er zijn geen schelpen of schelpresten opgevist die wijzen op menselijk gebruik.
Wel is een edelhertgeweifragment gevonden met twee diepe snijsporen, die zijn
veroorzaakt met een vuurstenen werktuig. De vondst dient te worden beschouwd
als productieafval. Daarnaast is een fragment van een runderrib opgevist waarop aan
weerszijden duidelijk snijsporen waarneembaar zijn. De roestformatie in de sporen
doen vermoeden dat de snijsporen vervaardigd zijn met een metalen voorwerp.
Andere opgeviste objecten die vervaardigd zijn door de mens zijn jonger dan 1492 na
Chr. Hiertoe behoren een nagenoeg complete baardmankruik en houten spanten van
een vaartuig. Zeer jong materiaal is ook aangetroffen in de netten, maar dit is niet als
archeologisch relevant beschouwd.
Uit de samenstelling van vondsten die tijdens de verschillende visexpedities gedaan
zijn, blijkt dat in de netten materiaal uit zeer uiteenlopende perioden terecht is
gekomen. In de Synthese van het geo-archeologisch en paleontologisch onderzoek
zandwingebied en buitencontour Maasvlakte 2 zal worden getracht om de vondsten
in een geologische context te plaatsen en om een verklaring te vinden voor het wel of
niet opvissen van bepaalde (periodespecifieke) vondstassemblages tijdens specifieke
tracks.

48 Botvangen

lIteratuur 49
9 Literatuur
Amkreutz L.W.S.W., Glimmerveen, J., Hublin J.-J., Roebroeks W., Anthonis L. (2010)
Een Neanderthaler uit de Noordzee. Westerheem 2, 50-9.
Andrews P. (1997) What taphonomy can and cannot tell us. Journal of Iberian Geology
23, 53-72.
Davis S.J.M. (1987) The archaeology of animals. Yale University Press, New Haven and
London.
Deeben J.H.C., Peeters H., Raemakers D., Rensink E., Verhart L.B.M. (2006) De Vroege
Prehistorie, NOaA hoofdstuk 11 (versie 1.0). Nationale Onderzoeksagenda Archeologie.
De Loecker D. (2010) Great Yarmouth dredging licence Area 240, Norfolk, United
Kingdom. Preliminary report on the lithic artefacts, Unpublished manuscript (Faculty of
Archaeology, Leiden University, The Netherlands).
Friederich F.H.W. (1967) Baardmannen. Historie en ontwikkeling van het gebruiksaar-
dewerk versierd met gebaarde gezichten. Westerheem 16, 96-113.
Glimmerveen J., Mol D., Post K., Reumer J.W.F., van der Plicht J., van Geel B., van
Reenen, G.B.A., Pals J.P. (2004) The North Sea project: the first palaeontological,
palynological, and archaeological results. In: Flemming N.C. (ed.), Submarine prehistoric
archaeology of the North Sea (CBA Research Report 141), York, 43-52.
Glimmerveen J., Mol D., van der Plicht H. (2006) The Pleistocene reindeer of the North
Sea - initial palaeontological data and archaeological remarks. Quaternary International
142/143, 242-6.
Groot M. (2010) Handboek Zoöarcheologie. Deel 1: Materiaal en Methoden.
ACVU-HBS, Amsterdam.
Guthrie R.D. (1990) Frozen Fauna of the Mammoth Steppe. The story of Blue Babe.
Chicago, University of Chicago Press.
Heuff I., Rabelink G., Moeiliker K. (2010) 60 jaar kor en bot. Straatgras 22-3.
Hijma M.P., Westerhoff W.E., Roebroeks W., van Kolfschoten T., Cohen K. (2011)
Cultureel erfgoed in de Noordzee: hoe gaan we er mee om? ARCHEObrief 15-3, 31-6.
Hublin, J.-J., Weston D., Gunz Ph., Richards M., Roebroeks W., Glimmerveen J.,
Anthonis L. (2009) Out of the North Sea: the Zeeland Ridges Neandertal. Journal of
Human Evolution 57-6, 777-85.
Huisman D.J., Lauwerier R.C.G.M., Jans M.M.E., Cuijpers A.G.F.M., Laarman F.J. (2006)
Degradatie en bescherming van archeologisch bot. Praktijkboek Instandhouding
Monumenten Deel II – 11 Overige onderwerpen, 14. Sdu Uitgevers, Den Haag.

50 Botvangen
Kahlke R.D. (1999) The history of the origin, evolution and dispersal of the late
Pleistocene Mammuthus-Coelodonta faunal complex in Eurasia (large mammals). Hot
Springs, Mammoth Site of Hot Springs.
van Kolfschoten T., Laban C. (1995) Pleistocene terrestrial Mammal faunas from the
North Sea Area. Mededelingen Rijks geologische Dienst 52, 135-51.
Louwe Kooijmans L.P. (1971) Mesolithic bone and antler implements from the
North Sea and from the Netherlands. Berichten Rijksdienst voor het Oudheidkundig
Bodemonderzoek 20/21, 27-73.
Moeliker K. (2010) Fossielen Maasvlakte 2 naar museum. Straatgras 22 (2): 22.
Mol D. (1991) Het ijstijdlandschap van de zuidelijke Noordzee. Grondboor en hamer 1,
9-14.
Mol D. (2012) Twee etmalen korren in de Eurogeul: trok de kotter OD7 door een
mammoetkerkhof? Afzettingen WTKG 33-1, 7-10.
Mol D., de Vos J. (1995) Korren op de Oosterschelde: een zoogdierpaleontoloog als
visser en wat de fossielen van de Oosterschelde ons vertellen. Grondboor en Hamer
49-3/4, 57-61.
Mol D., Post K. (2010) Gericht korren op de Noordzee voor de zoogdierpaleontologie:
een historisch overzicht van de uitgevoerde expedities. Cranium 27 (2), 14-28.
Mol D., Post, K. Reumer J.W.F., van der Plicht H., de Vos J., van Geel B., van Reenen
G., Pals J.P., Glimmerveen J. (2006) The Eurogeul - First report of the palaeontologi-
cal, palynological and archaeological investigations of this part of the North Sea, The
Netherlands. Quaternary International 142-142, 178-85.
Mol D., van Logchem W., van Hooijdonk, K., Bakker R. (2007) De Sabeltandtijger uit de
Noordzee. Norg, DrukWare.
Mol D., de Vos J., Bakker R., van Geel B., Glimmerveen J., van der Plicht H., Post K.
(2008) Kleine encyclopedie van het leven in het Pleistoceen: mammoeten, neushoorns en
andere dieren van de Noordzeebodem. Veen Magazines, Diemen.
Mol D., Borst W., Reumer J. (2010) De eerste fossiele hyenakeutel uit de Noordzee.
Straatgras 22 (5): 91-3.
Mol D., van der Plicht H., 2012. Fossiele otters uit het Noordzeebekken dateren van ca.
9000 jaar geleden. Straatgras 24 (1), 3-5.
Mol D., Reumer J. (2010) The North Sea - Maasvlakte 2 project. In: Abstracts Vth
International Conference on Mammoths and their relatives. Le Puy en Velay, 30 August
- 5 September 2010. Quaternaire, Hors-série, Numéro 3: 196.
Momber G., Tomalin D., Scaife R., Satchell J., Gillespie J. (2011) Mesolithic occupation
at Bouldnor Cliff and the submerged prehistoric landscapes of the Solent. The Council for
British Archaeology, York.

lIteratuur 51
Nieuwenkamp R. (2012) Vondsten Zonder Context. Een studie naar de mogelijkheden
tot kwalitatief prospectieonderzoek met betrekking tot de Paleolithische archeologie van
de Noordzeebodem. Niet gepubliceerd Bachelor manuscript (Faculteit Archaeology,
Universiteit Leiden, Nederland).
O’Connor T. (2004) The archaeology of animal bones. Sutton Publishing Limited,
Gloucestershire.
den Ouden N., Goud J. (2012) Aantasting zoogdierbotten uit de Noordzee door
mariene boorders, grazers en knagers. Cranium 2, 42-7.
Peeters H., Murphy P., Flemming N. (2009) North Sea Prehistory Research and
Management Framework (NSPRMF) 2009, Amerfoort.
Pieters M., Demerre I., Lenaerts T., Zeebroek I, de Bie M., de Clercq W., Dickinson B,
Monsieur P. (2010) De Noordzee: een waardevol archief onder water Meer dan 100 jaar
onderzoek van strandvondsten en vondsten uit zee in België: een overzicht, Relicta -
Archeologie, Momumenten en landschapsonderzoek in Vlaanderen 6, 177-218.
Reimer P.J., Baillie M.G.L., Bard E., Bayliss A., Beck J.W., Blackwell P.G., Bronk Ramsey
C., Buck C.E., Burr G.S., Edwards R.L., Friedrich M., Grootes P.M., Guilderson T.P.,
Hajdas I., Heaton T.J., Hogg A.G., Hughen K.A., Kaiser K.F., Kromer B., McCormac F.G.,
Manning S.W., Reimer R.W., Richards D.A., Southon J.R., Talamo S., Turney C.S.M.,
van der Plicht J., Weyhenmeyer C.E. (2009) IntCal09 and Marine09 Radiocarbon Age
Calibration Curves, 0-50,000 Years cal BP. Radiocarbon 51 (4), 1111-50.
Reumer J., Mol D., Borst W. (2010). The first Late Pleistocene coprolite of Crocuta
crocuta spelaea from the North Sea. Deinsea 14: 15-8.
Reumer J., Galis F. (2010) An unusual high incidence of cervical ribs in mammoths
(Mammuthus primigenus). In: Abstracts Vth International Conference on Mammoths
and their relatives. Le Puy en Velay, 30 August - 5 September 2010. Quaternaire,
Hors-série, Numéro 3: 45-7.
Sutcliffe A.J. (1969) Adaptations of spotted hyenas to living in the British Isles.
Mammalian Society Bulletin 31, 10-4.
Sutcliffe A.J. (1970) Spotted hyena: crusher, gnawer, digester and collector of bones.
Nature 227, 1110-3.
Sutcliffe A.J. (1973) Similarity of bones and antlers gnawed by deer to human artefacts.
Nature 246, 428-30.
Tizzard L., A. Bicket and D. De Loecker (in press): Seabed Prehistory: Investigating the
palaeogeography and Early Middle Palaeolithic archaeology in the southern North Sea.
Wessex Archaeology Report.
Tizzard L., A.R. Bicket, J. Benjamin and D. De Loecker (in press): A Middle Palaeolithic
Site in the southern North Sea: Investigating the archaeology and palaeogeography of
Area 240. Journal of Quaternary Science.

52 Botvangen
Verhart L.B.M. (1988) Mesolithic barbed points and other implements from Europoort,
the Netherlands. Oudheidkundige Mededelingen uit het Rijksmuseum van Oudheden 68,
145-94.
Verhart L.B.M. (1995) Fishing for the Mesolithic. The North Sea: a submerged
Mesolithic landscape. In: Fischer A. (ed.) Man and sea in the Mesolithic, coastal
settlement above and below present sea level. Oxford, 291-302.
Verhart L.B.M. (2001) De zee neemt, de zee geeft. Spectaculaire vondsten uit de
Noordzee. Westerheem 50, 102-07.
Verhart L.B.M. (2004) The implications of prehistoric finds on and off the Dutch coast.
In: Flemming N.C. (ed.): Submarine Prehistoric Archaeology of the North Sea: Research
Priorities and Collaboration with Industry. CBA Research Report 141. York, Council for
Britisch Archaeology, 57-61.
Voormolen B. (1997) The vertebrate taphonomy of the Middle Pleistocene site
Schöningen 12B, Niedersachsen, Germany. Doctoraal scriptie, Universiteit Leiden.
Weerts H.J.T., Borst W., den Ouden N., Guiran T., Hijma M.P., Mol D., Otte A., Reumer
J., Roebroeks W., Smit B., van Kolfschoten T., Vellinga T., Vos P., Westerhoff W. (2011)
Maasvlakte 2 (Port of Rotterdam), a showcase of interdisciplinary research of drowned
landscapes. In: Abstracts Congres INQUA, Bern 2011.
Wessex Archaeology (2010) Seabed prehistory: Site Evaluation Techniques (Area 240),
Seabed Sampling, Ref: 70752.02.
Wessex Archaeology (2011a) Seabed prehistory: Site Evaluation Techniques (Area 240),
Synthesis, Ref: 70754.04.
Wessex Archaeology (2011b) Seabed prehistory: Site Evaluation Techniques (Area 240),
Synthesis, Appendices, Ref: 70754.04.




















