
Water stress reveals differential antioxidant responses of tolerant and non-tolerant sugarcane...
12
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/authorsrights
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Water stress reveals differential antioxidant responses of tolerant and non-tolerant sugarcane...

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/authorsrights

Author's personal copy
Research article
Water stress reveals differential antioxidant responses of tolerantand non-tolerant sugarcane genotypes
Luis F. Boaretto a, Giselle Carvalho a, Lucélia Borgo a, Silvana Creste b, Marcos G.A. Landell b,Paulo Mazzafera c, Ricardo A. Azevedo a,*
aDepartamento de Genética, Universidade de São Paulo, Escola Superior de Agricultura “Luiz de Queiroz”, Avenida Pádua Dias 11, CP 9,13418-900 Piracicaba, SP, BrazilbCentro Cana de Açúcar, Instituto Agronômico de Campinas, CP 206, 14001-970 Ribeirão Preto, SP, BrazilcDepartamento de Biologia Vegetal, Instituto de Biologia, Universidade Estadual de Campinas, CP 6109, 13083-970 Campinas, SP, Brazil
a r t i c l e i n f o
Article history:Received 5 September 2013Accepted 13 November 2013Available online 21 November 2013
Keywords:Abiotic stressAntioxidant enzymesReactive oxygen speciesDrought stress
a b s t r a c t
The biochemical responses of the enzymatic antioxidant system of a drought-tolerant cultivar (IACSP 94-2094) and a commercial cultivar in Brazil (IACSP 95-5000) grown under two levels of soil waterrestriction (70% and 30% Soil Available Water Content) were investigated. IACSP 94-2094 exhibited oneadditional active superoxide dismutase (Cu/Zn-SOD VI) isoenzyme in comparison to IACSP 95-5000,possibly contributing to the heightened response of IACSP 94-2094 to the induced stress. The totalglutathione reductase (GR) activity increased substantially in IACSP 94-2094 under conditions of severewater stress; however, the appearance of a new GR isoenzyme and the disappearance of anotherisoenzyme were found not to be related to the stress response because the cultivars from both treatmentgroups (control and water restrictions) exhibited identical changes. Catalase (CAT) activity seems to havea more direct role in H2O2 detoxification under water stress condition and the shift in isoenzymes in thetolerant cultivar might have contributed to this response, which may be dependent upon the locationwhere the excessive H2O2 is being produced under stress. The improved performance of IACSP 94-2094under drought stress was associated with a more efficient antioxidant system response, particularlyunder conditions of mild stress.
� 2013 Elsevier Masson SAS. All rights reserved.
1. Introduction
Brazil leads the world in the production of ethanol and sugarderived from sugarcane and according to Companhia Nacional deAbastecimento (Companhia Nacional de Abastecimento, 2012), the2012/2013 harvest season in Brazil produced approximately 602million tons on 8.56 million hectares. A challenge to increaseethanol and sugar from sugarcane in Brazil has been the limitationof the crop expansion to marginal lands, where rainfall distributionis irregular or insufficient to support the entire crop cycle (Pireset al., 2008; Cheavegatti-Gianotto et al., 2011). Additionally,
sugarcane has been considered as the main feedstock for the pro-duction of second generation ethanol from biomass (Boaretto andMazzafera, 2013).
On a global scale, thewater available for crops is limited, coveringonly 20%of the total areaused for agriculture (Rosegrant et al., 2009).Accordingly, although the physiological effects produced by droughtin plants are well studied and characterised, this remains as aresearch topic of high priority (Medici et al., 2014). One of the mostwell-known effects of water restriction in plants is the reduction ingrowth and development due to the cessation of cell expansion.Decrease in the water loss is a physiological mechanism that iscontrolled partly or completely by stomatal closure, culminating in achange in the leaf water status that is directly related to carbonassimilation and photosynthetic rates (Benesová et al., 2012).
The correlation between drought stress and the responses of theantioxidant systems in sugarcane is poorly understood, largelybecause the complexity of the sugarcane genome precludes athorough understanding of the genetic, physiological, andbiochemical events underlying this phenomenon (Azevedo et al.,2011). A recent report revealed how complex this can be in
Abbreviations: ROS, reactive oxygen species; O2�, superoxide radical; 1O2,singlet oxygen; H2O2, hydrogen peroxide; OH�, hydroxyl radical; SOD, superoxidedismutase; CAT, catalase; GPX, glutathione peroxidase; GPOX, guaiacol peroxidase;GSH, glutathione; AsA, ascorbic acid; GSSG, oxidized glutathione; GR, glutathionereductase; SAWC, Soil Available Water Content; RWC, Relative Water Content;TBARS, thiobarbituric acid reactive substances; MDA, malondialdehyde; PAGE,polyacrylamide gel electrophoresis.* Corresponding author. Tel.: þ55 19 3429 4475; fax: þ55 19 3447 8620.
E-mail address: [email protected] (R.A. Azevedo).
Contents lists available at ScienceDirect
Plant Physiology and Biochemistry
journal homepage: www.elsevier .com/locate/plaphy
0981-9428/$ e see front matter � 2013 Elsevier Masson SAS. All rights reserved.http://dx.doi.org/10.1016/j.plaphy.2013.11.016
Plant Physiology and Biochemistry 74 (2014) 165e175

Author's personal copy
sugarcane in which although drought has caused reductions in leafgas exchange in the three genotypes studied, photosynthesis in onegenotype was related to both non-stomatal and stomatal limita-tions, while photosynthesis in the other two genotypes was limitedonly stomatally (Ribeiro et al., 2013).
In fact, for themajority of plant specieswhen submitted towaterdeficit the reactive oxygen species (ROS) scavenging system is acti-vated (Kar, 2011). The plant stress defense system response variesdepending on the time length of exposure and severity of the stress,tissue/organ and stage of development (Dat et al., 2000; Douradoet al., 2013). Up to a certain level ROS production under stress maywork as a signal for triggering acclimatory/defense responses viatransduction pathways, which have H2O2 as secondary messenger(Cruz de Carvalho, 2008; Miller et al., 2010). Over a certain level,however, ROS may have deleterious effects to plant cells. As aconsequence, defences against ROS are activated, involving non-enzymatic and enzymatic antioxidant mechanisms that operatetogether in an efficient system of detoxification (Gratão et al., 2005;Impa et al., 2012). The non-enzymatic system involves flavonoids,alkaloids, phenolic compounds, tocopherol, and carotenoids, all ofwhich are able to donate electrons to the glutathione-ascorbatecycle in which the oxidised glutathione is regenerated to reducedglutathione (Gratão et al., 2005). The enzymatic antioxidantresponse includes a wide range of enzymes, such as superoxidedismutase (SOD), ascorbate peroxidase (APX), and catalase (CAT),amongothers (Gratão et al., 2005;Mittler, 2002;Gallegoet al., 2012).Other aspects and mechanisms involved in plant responses andstress tolerance may also be involved (Dourado et al., 2013).
Only in recent years more attention has been given to the un-derstanding of the antioxidative defense system in plants subjectedto drought and water deficit in plant tissues (Ghane et al., 2012;Doupis et al., 2013; Kaur et al., 2013). These studies were carriedout with different plant species, but most of them exhibitedincreased antioxidant enzymes activities. For instance, activityincreases in CAT, GR and other peroxidases have been observed ingrass species, particularly for tolerant lines under water deprivationcondition (Selote and Khanna-Chopra, 2006; Duan et al., 2009;Simova-Stoilova et al., 2009). Recent data indicated that the upregulation of SOD and CAT activities under stress conditionsappears to be one of the general mechanisms used for droughttolerant crops (Devi et al., 2012). Furthermore, a remarkable cor-relation between SOD and CAT activities and H2O2, MDA and pro-line contents with drought tolerance observed for some genotypessuggest that by combining physiological, morphological, geneticaland biochemical approaches specific traits may be further identi-fied that are related to drought tolerance (Kaur et al., 2013).
In the present study, the general objective was to evaluate thebiochemical response of the enzymatic antioxidant system of twosugarcane cultivars subjected to drought stress. IACSP 94-2094 isconsidered a rustic cultivar and exhibits tolerance to drought(Ribeiro et al., 2013; Landell et al., 2005); cultivar IACSP 95-5000 isa commercial cultivar that is widely grown in several sugarcaneareas in Brazil (Marcos, 2012). Different levels of water stress wereimposed by establishing 70% and 30% of the Soil Available WaterContent (SAWC), thus simulating conditions of mild and severewater stress (Inman-Bamber and De Jager, 1986), respectively. Therelative water content (RWC) and chlorophyll content (using theSPAD index) were also used to monitor the sugarcane plants.
2. Results
2.1. Relative water content (RWC) and SPAD index
The cultivars IACSP 94-2094 and IACSP 95-5000 were grownunder glasshouse conditions and subjected to one of two different
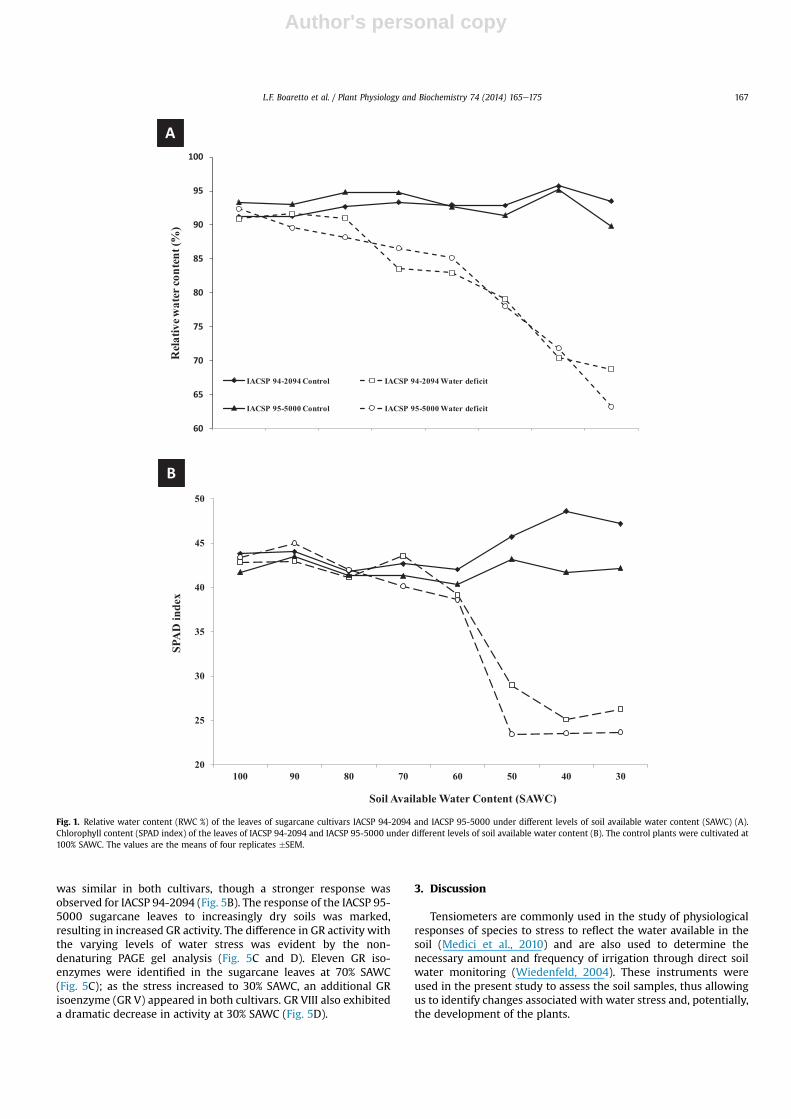
levels of soil water stress (70% SAWC and 30% SAWC). The differ-ential responses of RWC and the SPAD index to water deficiencywere compared between the leaves of well-watered plants of bothcultivars. The RWC value of the stressed IACSP 94-2094 plants washigh to levels of approximately 80% SAWC; the reduction in RWCwas significant only at 70% SAWC, decreasing from 90.9% to 83.5%and substantially decreasing thereafter until reaching approxi-mately 68.7% at 30% SAWC. For IACSP 95-5000, a reduction of thesame magnitude (on the order of 10% of RWC) was observed at80% SAWC, and the RWC value reached 63.1% at 30% SAWC(Fig. 1A).
The estimated leaf chlorophyll content (or SPAD index)remained high to 60% SAWC in both the control and stressed plantsof the two cultivars, whereas a sharp drop in the SPAD index wasobserved in both cultivars subjected to water stress at 50% SAWC.However, the SPAD index of IACSP 94-2094 remained higher thanthat of IACSP 95-5000 at 50%, 40%, and 30% SAWC (Fig. 1B).
2.2. Determination of hydrogen peroxide levels and lipidperoxidation
No differences in the H2O2 content were observed between thecontrol plants and plants subjected to the 70% SAWC stress treat-ment of both cultivars (Fig. 2A). In contrast, an increase in the H2O2content was observed only in IACSP 95-5000 under the moresevere stress treatment (30% SAWC) (Fig. 2B), whereas IACSP 94-2094 exhibited a reduction in the H2O2 content at 30% SAWC. TheH2O2 content was similar in the controls of both cultivars for the30% SAWC treatment (Fig. 2B).
The MDA levels followed a similar trend as that observed for theH2O2 content, decreasing in both cultivars at 70% SAWC (Fig. 2C).However, with the intensification of the water restriction to 30%SAWC, the MDA content increased more significantly in the treatedplants than in the control plants (Fig. 2D).
2.3. Antioxidant enzymes
The SOD activity at 70% SAWC decreased by 30.7% in the IACSP94-2094 cultivar (Fig. 3A); the opposite effect was observed at 30%SAWC when the lower water content in the soil promoted anincrease in SOD activity of up to 29.6% in the tolerant cultivar(Fig. 3B). The SOD isoenzyme profile and activity were also deter-mined using non-denaturing PAGE (Fig. 3C and D). Eight SOD iso-enzymes were detected in the IACSP 94-2094 extracts: SOD I and IIwere characterised as Mn-SOD, whereas SOD III to VIII were char-acterised as Cu/Zn-SOD. The sugarcane cultivar IACSP 95-5000 hadthe same SOD isoenzyme profile as the IACSP 94-2094 cultivar, withthe exception of a complete lack of isoenzyme VI. SODs II, VII, andVIII accounted for the majority of SOD activity in both cultivars.
The basal activity of CAT at 70% SAWC was greater in IACSP 95-5000 than in IACSP 94-2094 (Fig. 4A). However, substantial in-creases in the total CAT activity were observed for both cultivarsonly at 30% SAWC (Fig. 4B). The assessment of the CAT activity bynon-denaturing PAGE revealed the presence of two distinct CATisoenzymes (I and II) in IACSP 94-2094 at 70% SAWC (Fig. 4C and D).However, a distinct modulation in CAT activity was observed inIACSP 94-2094 subjected to mild water stress: although CAT I wasactive only in the control, CAT II was active in the mildly stressedplants (Fig. 4C). This pattern was not observed for IACSP 95-5000,with both isoenzymes (CAT I and II) equally active in all treatments(Fig. 4D).
The GR activity was altered in both cultivars by the mild stressimposed at the 70% SAWC level (Fig. 5A); however, the GR activitydecreased in IACSP 94-2094, whereas it increased in IACSP 95-5000. Under the severe water stress at 30% SAWC, the GR activity
L.F. Boaretto et al. / Plant Physiology and Biochemistry 74 (2014) 165e175166
Author's personal copy
was similar in both cultivars, though a stronger response wasobserved for IACSP 94-2094 (Fig. 5B). The response of the IACSP 95-5000 sugarcane leaves to increasingly dry soils was marked,resulting in increased GR activity. The difference in GR activity withthe varying levels of water stress was evident by the non-denaturing PAGE gel analysis (Fig. 5C and D). Eleven GR iso-enzymes were identified in the sugarcane leaves at 70% SAWC(Fig. 5C); as the stress increased to 30% SAWC, an additional GRisoenzyme (GR V) appeared in both cultivars. GR VIII also exhibiteda dramatic decrease in activity at 30% SAWC (Fig. 5D).
3. Discussion
Tensiometers are commonly used in the study of physiologicalresponses of species to stress to reflect the water available in thesoil (Medici et al., 2010) and are also used to determine thenecessary amount and frequency of irrigation through direct soilwater monitoring (Wiedenfeld, 2004). These instruments wereused in the present study to assess the soil samples, thus allowingus to identify changes associated with water stress and, potentially,the development of the plants.
Fig. 1. Relative water content (RWC %) of the leaves of sugarcane cultivars IACSP 94-2094 and IACSP 95-5000 under different levels of soil available water content (SAWC) (A).Chlorophyll content (SPAD index) of the leaves of IACSP 94-2094 and IACSP 95-5000 under different levels of soil available water content (B). The control plants were cultivated at100% SAWC. The values are the means of four replicates �SEM.
L.F. Boaretto et al. / Plant Physiology and Biochemistry 74 (2014) 165e175 167

Author's personal copy
Two sugarcane cultivars were used to study the enzyme anti-oxidant system and to evaluate the responses of the cultivars to soilwater restriction. In previous studies, cultivar IACSP 94-2094 wascharacterised as tolerant when compared to susceptible cultivars(Landell et al., 2005). Although no physiological alterations wereobserved in the IACSP 94-2094 plants (leaf water potential, carbondioxide assimilation, and intercellular CO2 concentration) at theinitial developmental stage, this cultivar exhibited decreases inthese parameters at the vegetative stage (approximately 184 daysafter planting). In contrast, no alterations in height and biomassproduction were detected when compared to the susceptiblecultivar (Machado et al., 2009).
The IACSP 95-5000 cultivar is widely grown throughout SãoPaulo State and other regions of Brazil (Dinardomiranda et al.,2008). Although the drought tolerance of this cultivar has notbeen characterised in detail, field observations suggest that it issomewhat more susceptible to drought than IACSP 94-2094(Machado and Ribeiro, 2012). Therefore, IACSP 95-5000was chosenas a reference in our study.
In our approach, we observed that IACSP 94-2094 retainedwater during dehydration, maintaining higher leaf RWC values thanIACSP 95-5000 (Fig. 1A). Water stress tolerance is frequently asso-ciated with the maintenance of RWC, as previously reported forwheat genotypes (Rampino et al., 2006), sugarcane cultivars(Landell et al., 2005; Cia et al., 2012; Silva et al., 2007), and commonand tepary beans (Türkan et al., 2005). Although we have notphysiologically characterised IACSP 95-5000 for water stresstolerance, its lower RWC appears to indicate that it has less abilityto retain the water content of its leaves. In a previous report pub-lished by our group (Cia et al., 2012), RWC was used to measure thewater status of tissues to follow the antioxidant response oftolerant (SP83-2847 and SP83-5073) and sensitive (SP90-3414 and
SP90-1638) sugarcane cultivars in a time-course experiment. Thesensitive genotypes exhibited lower RWC values than the tolerantgenotypes at 3 days after complete water suppression, and thisdifference persisted throughout the 20 days of the imposed stress.
An increased rate of lipid peroxidation accompanied by an in-crease in H2O2 content was observed in the IACSP 95-5000 cultivarunder conditions of severe stress (Fig. 2BeD). The increase in theH2O2 content detected in a number of species has been correlatedwith membrane damage, and the extent of this damage iscommonly used as a measure of stress tolerance (Gratão et al.,2005; Türkan et al., 2005). In addition to being an indicator ofstress, H2O2 is a secondary messenger that acts locally with othermolecules; therefore, the H2O2 stress response signalling pathwaymay promote the accumulation of other molecules that directly orindirectly contribute to the maintenance of the cellular redox-status (Maxwell et al., 2002; Mori and Schroeder, 2004). A highrate of H2O2 is produced under conditions of drought stress; in thissituation, a rapid response of the antioxidant system is required forH2O2 to function as an effective secondary messenger (Cruz deCarvalho, 2008).
The role of the antioxidant system is closely related to thescavenging of the excessive amounts of ROS produced in the cellwhen the redox state is disturbed by a stressful condition. In thisstudy, stress was a result of drought, and SOD played a key role inthe defence against superoxide (O2�) in different cell compart-ments (Alscher et al., 2002). According to our results, an additionalSOD isoenzyme (SOD VI) characterised as Cu/Zn-SOD was observedin cultivar IACSP 94-2094. Drought-tolerant and susceptible maizeinbred lines was evaluated and observed that the tolerance of thedrought-resistant line appeared to be linked to the presence of Cu/Zn-SOD isoenzymes (Malan et al., 1990). In agreement with ourobservations, SOD VI appears to play a key role in the drought
Fig. 2. Hydrogen peroxide (H2O2) content (mmol g�1 fresh weight) in the leaves of two cultivars, IACSP 2094 and IACSP 95-5000, under 70% and 30% of soil available water content(SAWC) (A and B). Lipid peroxidation was measured as the malondialdehyde (MDA) content (nmol g�1 fresh weight) in the leaves of IACSP 94-2094 and IACSP 95-5000 under 70%and 30% SAWC (C and D). The control plants were cultivated at 100% SAWC. The values are the means of four replicates �SEM. C e Control. WD e Water deficit.
L.F. Boaretto et al. / Plant Physiology and Biochemistry 74 (2014) 165e175168

Author's personal copy
Fig. 4. Specific activity of catalase (CAT) (mmol min�1 mg�1 protein) in the leaves of two cultivars, IACSP 94-2094 and IACSP 95-5000, under 70% and 30% of soil available watercontent (SAWC) (A and B). Activity staining for catalase (CAT) following non-denaturing polyacrylamide gel electrophoresis of leaf extracts of IACSP 94-2094 and IACSP 95-5000under 70% and 30% SAWC (C and D). The control plants were cultivated at 100% SAWC. The values are the means of four replicates�SEM. C e Control. WD eWater deficit. S e bovineliver CAT standard.
Fig. 3. Specific activity of superoxide dismutase (SOD) (mmol min�1 mg�1 protein) in the leaves of two cultivars, IACSP 94-2094 and IACSP 95-5000, under 70% and 30% of soilavailable water content (SAWC) (A and B). Activity staining for superoxide dismutase (SOD) following non-denaturing polyacrylamide gel electrophoresis of leaf extracts of IACSP94-2094 and IACSP 95-5000 under 70% and 30% SAWC (C and D). The control plants were cultivated at 100% SAWC. The values are the means of four replicates �SEM. C e Control.WD e Water deficit. S e SOD standard from bovine liver.
L.F. Boaretto et al. / Plant Physiology and Biochemistry 74 (2014) 165e175 169

Author's personal copy
tolerance exhibited by IACSP 94-2094; this isoenzyme may alsoexplain the better performance of this cultivar in the total stressresponse exhibited. However, neither the total SOD activity (Fig. 3Aand B) nor the isoenzyme band intensity profiles appeared to varysignificantly between the cultivars (Fig. 3C and D), suggesting thatany possible excess H2O2 produced by the drought conditions wasnot a result of the increased dismutation of the superoxide radicalto H2O2 by SOD.
H2O2 can be reduced to H2O by CAT and other peroxidases, andH2O2 functions as an intracellular signalling molecule that activatesor deactivates certain antioxidant enzymes (Sgherri et al., 2000).According to our results, both sugarcane cultivars exhibited in-creases in CAT activity under conditions of severe stress (30%SAWC) (Fig. 4B); however, as evidenced in Fig. 2B, this increase wassufficient to overcome the excess H2O2 production only in cultivarIACSP 94-2094. The levels of H2O2 were unaltered for the tolerantcultivar, suggesting that if H2O2 was produced in excess, CAT andpossibly other peroxidases not analysed in this study were efficientin dealing with the excessive production. This hypothesis is furtherstrengthened by the increased CAT activity exhibited by IACSP 94-2094 under severe stress (Fig. 4B). Studies of the responses of 5-day-old seedlings of indica rice (Oryza sativa indica) cultivars todrought stress conditions imposed by 20% (v/v) PEG 6000 in Mur-ashige and Skoog (MS) solution showed that the response of adrought-tolerant variety was associated with an enhanced capacityof antioxidant enzymes to address the stress and that the decline inantioxidant response induced by drought was associated with thecultivar’s sensitivity to drought (Basu et al., 2010). Similar resultswere obtained (Lascano et al., 2001) when studying drought-tolerant wheat cultivars under conditions of water deficit. Theseauthors reported increases in SOD, ascorbate peroxidase and GRactivities in tolerant cultivars.
Another important result obtained in this study was thedescription of the CAT isoenzyme profile (Fig. 4C and D). Two CATisoenzymes were detected and although they were not clearlyseparated from each other by non-denaturing PAGE, it was possibleto observed distinct responses. The IACSP 95-5000 cultivar did notexhibit anymajor changes in isoenzyme activity in either treatment(control or water restriction), with the two main CAT isoenzymesbeing observed. The same two isoenzymes were also detected inthe drought-tolerant IACSP 94-2094 cultivar; however, only CAT I,the least electronegative isoenzyme, was present in the control andonly CAT II, the least electropositive isoenzyme, was present in thetreated plants under conditions of mild stress (Fig. 4C). When theplants were subjected to severe stress (Fig. 4D), cultivar IACSP 94-2094 exhibited only CAT I isoenzyme in both treatments, whereasplants of cultivar IACSP 95-5000 subjected to water restrictionexhibited higher CAT II activity. The gel data, however, does notallow us to explain fully the overall increased in total CAT activityobserved (Fig. 4B) since a direct relationship between in vitro and ingel activity may not be always established. But, the presence of adistinct response regarding the CAT isoenzymes suggests thattolerant cultivar responded differently to the level of stress. Wehave not been able to clearly determine the organelle location ofeach isoenzymewhich would have provided more information thatmight have helped to understand the response. For instance,Soares-Netto (2001) reported the results of a search of the sugar-cane expressed sequence tag (SUCEST) database for homologoussequences involved in the oxidative stress response. This authorreported eight clusters with similarities to all three maize CATisoenzymes (CAT 1, CAT 2 and CAT 3) in the SUCEST database, whichsuggest that peroxisomal, mitochondrial and cytoplasmic iso-enzymes may be distributed among sugarcane tissues/organs.Therefore, under low mild water stress the shift observed may be
Fig. 5. Specific activity of glutathione reductase (GR) (mmol min�1 mg�1 protein) in the leaves of two cultivars, IACSP 94-2094 and IACSP 95-5000, under 70% and 30% of soilavailable water content (SAWC) (A and B). Activity staining for glutathione reductase (GR) following non-denaturing polyacrylamide gel electrophoresis of leaf extracts of IACSP 94-2094 and IACSP 95-5000 under 70% and 30% SAWC (C and D). The control plants were cultivated at 100% SAWC. The values are the means of four replicates �SEM. C e Control. WDe Water deficit. S e Saccharomyces cerevisiae GR standard.
L.F. Boaretto et al. / Plant Physiology and Biochemistry 74 (2014) 165e175170

Author's personal copy
due to different production levels of H2O2 in distinct cell organelles,resulting in different participation of distinct CAT isoenzymes.However, at this stage we cannot suggest which CAT isoenzyme ispreferentially being expressed in the early response to mild waterstress in sugarcane leaves. In another study Lambais (2001) iden-tified defense-related protein ESTs differentially expressed in sug-arcane tissues infected with Gluconacetobacter diazotrophicus orHerbaspirillum rubrisubalbicans, which resulted in 179 putativedefense-related protein EST-contigs expressed in non-infected tis-sues (leaves and roots) and/or infected tissues. One interestingresult was that G. diazotrophicus and H. rubrisubalbicans-infectedtissues exhibited induction of an EST-contig encoding CAT 3, whichsuggested that CAT 3 in G. diazotrophicus and H. rubrisubalbicansinfected tissues was likely to be directly involved in stress responseto the two endophytes by controlling the accumulation of H2O2 inthe plant tissues. Yet, this author did not comment on the organellelocation of CAT 3. Three CAT isoenzymes were also observed in astudy of nine sugarcane genotypes subjected to heat stress, but theresponses among the tolerant and sensitive genotypes varied aswell as the distribution of the isoenzymes, in some cases all threeisoenzymes were observed, whilst in others just one CAT isoen-zyme was detected (Srivastava et al., 2012). Nevertheless, theresponse of CAT in sugarcane may vary as shown in a number ofstudies and to different types of abiotic stresses. For instance, twosugarcane varieties subjected to salinity stress exhibited differentresponses by CAT (Willadino et al., 2011), but the CAT isoenzymeactivity profile was not determined. Finally, it must be kept in mindthat sugarcane has a complex genome.
Another aspect that deserves more attention in future researchis the study of stress responses to drought by C3 and C4 plantspecies and the mechanisms of tolerance involved in comparativestudies, with special attention to the role of CAT. As far as we areaware, little has been done when the antioxidant responses areconcerned. A more detailed study has been recent published byUzilday et al. (2012) who compared ROS formation and antioxidantenzymes in the C4 Cleome gynandra and C3 Cleome spinosa plantspecies under drought stress. The results indicated that both spe-cies exhibited different responses, including for CAT, but moreimportantly the antioxidant system of the C4 species was able tocope with ROS formation under drought stress, which was not truefor the C3 species. This provides further indication that CAT has animportant role and confirms the results observed in the C4 sugar-cane plant species used in this study. These results are also inagreement with the responses observed for CAT in wild-type and aCAT-deficient barley (C3) when subjected to ozone stress (Azevedoet al., 1998).
Another important pathway involved in the control of the ROSlevel is the ascorbate/glutathione cycle in which GR plays a majorrole (Chew et al., 2003; Shigeoka et al., 2002). According to ourdata, total GR activity increased substantially in the IACSP 94-2094drought-tolerant plants under severe water stress (Fig. 5B) but notmild stress. Indeed, the treated plants of this cultivar exhibited alower GR activity than the controls, clearly indicating that IACSP94-2094 was able to withstand mild stress and only requiredincreased GR activity when the stress was severe. This was not thecase for the IACSP 95-5000 commercial sugarcane cultivar, whichexhibited increased GR activity even under mild stress (Fig. 5A andB). The changes in the responses of the GR isoenzymes could notbe correlated to specific changes in any of the isoenzymes detec-ted (Fig. 5C and D); in fact, changes were observed but were notrelated to a specific cultivar or to any of the two stress conditionstested. For instance, the isoenzyme pattern was identical for bothcultivars under each stress condition, but isoenzyme GR Vappeared in all plants (control and treatments) only under con-ditions of severe stress, whereas isoenzyme GR VIII disappeared in
all plants under conditions of severe stress independent of thetreatment. These observations suggest that both GR isoenzymesare responding to the stage of plant development rather than tothe water stress imposed. It is important to bear in mind that,although the cultivars were subjected to similar conditions ofwater stress, the plants may have been at different stages ofdevelopment at the different stress levels (70% and 30%). Incontrast to the approach used in our previous report in whichwater stress was imposed by suspending the water supply andharvesting the plants on exactly the same day, independent of thewater availability in the soil (Cia et al., 2012), the approach used inthis study may have resulted in plants at different stages ofdevelopment. Therefore, the changes in the isoenzyme profileobserved cannot explain the changes in the total GR activity;instead, the changes in the total GR activity are likely due to thecombined changes in all GR isoenzymes. Increases in GR as a resultof drought stress have been previously reported for cotton(Ratnayaka et al., 2003), wheat seedlings (Keles and Oncel, 2002),beans (Torres-Franklin et al., 2007), and rice (Sharma and Dubey,2005). Moreover, the accumulation of abscisic acid (ABA) duringmild water stress was reported to trigger ROS production and aconsequent increase in antioxidant enzyme activities in youngmaize leaves (Jiang and Zhang, 2002). Similarly, cowpea leavessubjected to progressive drought stress exhibited changes in theintracellular distribution of GR enzymes, indicating that theobserved alterations in activity were related to cultivar tolerance(Contour-Ansel et al., 2006).
The integrated actions of antioxidant enzymes to preventoxidative stress suggest that an early response to drought stressoccurred in the IACSP 94-2094 cultivar under the mild stress con-dition. In contrast, cultivar IACSP 95-5000 did not show a rapid,efficient response and instead exhibited a sharp increase in H2O2levels. The main differences can be clearly observed in Fig. 6,whereby the amount of water available in the soil acted directly onthe hydric balance and, consequently, on the generation of ROS. It isclear that the IACSP 94-2094 cultivar performed better than IACSP95-5000 under both stress levels, particularly under mild stress.Although limitations may exist to determine the precise ROS levelsin plant tissues, our data suggest that any excess ROS (particularlysuperoxide and H2O2) produced were efficiently dealt by the anti-oxidant enzymes analysed. Moreover, the tolerant sugarcanecultivar had one specific SOD isoenzyme that was not observed inthe sensitive cultivar, which might suggest the involvement of thisisoenzyme in drought tolerance. GR activity also responded posi-tively to conditions of water stress, but the changes in the isoen-zyme profile were related to the developmental stage of the plantsrather than to the stress. Finally, CAT showed differential responsebetween the two genotypes as distinct isoenzyme patterns wereobserved, suggesting it may play a key role in the sugarcane stressresponse, nevertheless, more information is still needed, particu-larly if the changes observed in CAT isoenzyme activity profile andtotal activity are related to the organelle localization of the CATisoenzymes.
4. Material and methods
4.1. Plant material and growth conditions
Sugarcane cultivars IACSP 94-2094 and IACSP 95-5000, atolerant and commercial cultivar, respectively (Landell et al., 2005;Marcos, 2012), were grown under glasshouse conditions in Pira-cicaba, São Paulo (22�4203000S; 47�3803000W). One-eyed sets of bothcultivars were planted in 0.5-L plastic pots containing substrate(Plantmax, Eucatex�). After forty-five days of acclimatisation, theplants were randomly transplanted to plastic pots (two plants per
L.F. Boaretto et al. / Plant Physiology and Biochemistry 74 (2014) 165e175 171

Author's personal copy
pot) of 50 � 50 cm diameter � depth (85 L) containing latosol/organic material mixture (1:1). The plants were watered once aday, supplemented with 20 g L�1 of commercial solution (20-N;20-K2O; 20-P2O5, JR Perters Inc.) during the initial developmentstage, and maintained at 28 �C � 4 �C, until plants reached7-months old.
4.2. Determination of soil humidity and treatments
A curve representing the water content in the soil was con-structed to simulate the real conditions of irrigation and evapo-transpiration in glasshouse (Fig. 7). Tensiometers were installed inthe pots (V ¼ 100 mm; h ¼ 300 mm), and the soil moisture was
Fig. 6. General view of the antioxidant system involving enzymatic and non-enzymatic mechanisms of tolerant and commercial sugarcane cultivars based on the present work. CATecatalase; SOD e superoxide dismutase; GR e glutathione reductase; MDHA e monodehydroascorbate; DHA e dehydroascorbate; GSSG e oxidised glutathione; GSH e reducedglutathione; ABA e abscisic acid. The red squares represent a decrease in enzymatic activity, and the green squares represent an increase in enzymatic activity. The grey squaresrepresent no alteration in enzymatic activity. 70% SAWC (70% of soil available water content); 30% SAWC (30% of soil available water content). (For interpretation of the references tocolour in this figure legend, the reader is referred to the web version of this article.)
L.F. Boaretto et al. / Plant Physiology and Biochemistry 74 (2014) 165e175172

Author's personal copy
reflected as tension (kPa) (Richards, 1949). Two levels of SoilAvailable Water Capacity (SAWC) were established, when SAWCreached 70% and 30%, representing mild and severe water stress,respectively.
Theþ1 leaves with a clearly visible “dewlap”were harvested forboth genotypes at three determined periods based on SAWC: 100%SAWC (control), 70% SAWC and 30% SAWC. The leaf blades, withoutthe midrib, were immediately frozen in liquid N2, ground with amortar and pestle, and stored at �80 �C for further biochemicalanalysis.
4.3. Determination of the relative water content (RWC) and SPADindex
Leaf discs of 2.5 cm in diameter were collected from þ1 leaftillers at 100, 90, 80, 70, 60, 50, 40 and 30% SAWC, and the sampleswere immediately packed in laminated paper envelopes. The leafdisc fresh weight was determined, and the samples were thenindividually immersed in deionised-distilled water for 6 h underlow irradiance. The excess water was gently removed from the discswith absorbent paper prior to the determination of the turgidweight. The disc dry weight was determined after 72 h at 70 �C(Smart and Bingham, 1974).
The SPAD index and leaf chlorophyll content were estimatednon-destructively using a SPAD-502 chlorophyll meter (MinoltaCorp., Ramsey, USA) (Markwell et al., 1995) at the same points usedfor the RWC assay.
4.4. Protein extraction and determination
One gram of fine leaf powder was homogenised in amortar with2 mL of buffer containing 100 mM potassium phosphate buffer (pH7.5) 1 mM ethylenediaminetetraacetic acid (EDTA), 3 mM dithio-threitol, and 5% (w/v) insoluble polyvinylpolypyrrolidone. The ho-mogenate was centrifuged at 10,000 g for 30 min, and thesupernatant was sub-aliquoted and stored at �80 �C for furtheranalysis. The protein concentration was determined using bovineserum albumin as the standard (Bradford, 1976).
4.5. Hydrogen peroxide concentration and determination of lipidperoxidation
The measurement of the H2O2 content and the determination oflipid peroxidation were performed in the same extraction; 300 mgof powdered leaves was homogenised in 2 mL of 0.1% (w/v) tri-chloroacetic acid (TCA) and centrifuged at 10,000 g for 10 min at4 �C (Alexieva et al., 2001). The assay consisted of a mixture of
200 mL of supernatant from the TCA extraction, 200 mL of 100 mMpotassium phosphate buffer (pH 7.0), and 800 mL of 1 M potassiumiodide incubated for 1 h on ice in the dark. The H2O2 level wasmeasured using a spectrophotometer at 390 nm. The result wasexpressed in mmol g�1 fresh weight.
Lipid peroxidation was determined by the presence of malon-dialdehyde (MDA), a product of lipid peroxidation that reacts withthiobarbituric acid (TBA). A 250-mL of the supernatant from the TCAextraction was added to 1 mL of a solution containing 20% (w/v)TCA and 0.5% (w/v) TBA. The samples were incubated for 30 min at95 �C and centrifuged for 5 min at 10,000 g; the absorbance wasmeasured at 535 and 600 nm using a spectrophotometer. The MDAcontent was assessed by a 535 nm value e 600 nm value calcula-tion. The result was expressed in nmol g�1 fresh weight.
4.6. Analysis of antioxidant enzymes
The superoxide dismutase (SOD) activity was determinedspectrophotometrically (Giannopolitis & Ries, 1977). The assayscontained 1.790 mL of 50 mM sodium phosphate (pH 7.8), 225 mL of1 mM r-nitro blue tetrazolium chloride (NBT), 780 mL of 50 mMmethionine, 30 mL of 10 mM EDTA, 150 mL of 0.1 mM riboflavin, and25 mL of protein extract. The reaction mixture was exposed to lightfor 5 min and measured at 560 nm. The determination of the totalcatalase (CAT) activity was performed using a spectrophotometer(Monteiro et al., 2011). The reaction mixture was composed of 1 mLof 100 mM potassium phosphate buffer (pH 7.5) to which 2.5 mL ofH2O2 (30% solution) and 15 mL of biological extract were added. Theactivity was determined by the rate of H2O2 consumption at240 nm that occurred within 1 min and was expressed inmmol min�1 mg�1 protein. Lastly, the activity of glutathionereductase (GR) was determined with a spectrophotometer at412 nm for 1 min at 30 �C (Smith et al., 1988; Gratão et al., 2012).Each GR assay contained 1000 mL of 100 mM potassium phosphatebuffer (pH 7.5), 100 mL of 1 mM of oxidised L-glutathione (GSSG),100 mL of 0.1 mM of NADPH, 500 mL of 1 mM of 5,50-dithiobis(2-nitrobenzoic acid) (DTNB), and 50 mL of biological extract. The GRactivity was expressed in mmol min�1 mg�1 protein.
4.7. Determination of antioxidant enzymes by polyacrylamide gelelectrophoresis (PAGE)
The activity stainings for SOD (Beauchamp and Fridovich, 1971;Garcia et al., 2006), CAT (Woodbury et al., 1971) and GR (Martinset al., 2011) were performed using non-denaturing 12% PAGE gelsloaded with 30, 40 and 30 mg of biological extract protein, respec-tively, until migration was completed. The SOD isoenzymes wereclassified in Mn-SOD, Fe-SOD, or Cu/Zn-SOD (Azevedo et al., 1998).
4.8. Statistical analysis
The experimental design followed a complete randomised sys-tem containing four replicates. The results are expressed as themean and standard error of the mean of three independents rep-licates of the enzyme assay for each biological extract.
Acknowledgements
We thank Fundação de Amparo à Pesquisa do Estado de SãoPaulo for financial support (Fapesp Grants 2009/54676-0 andBIOEN-2008/58035-6) and Conselho Nacional de DesenvolvimentoCientífico e Tecnológico (CNPq-Brazil) for the fellowships to LFB,GC, and LB and the research fellowships to PM and RAA.
Fig. 7. Soil water retention curve.
L.F. Boaretto et al. / Plant Physiology and Biochemistry 74 (2014) 165e175 173

Author's personal copy
References
Alexieva, V., Sergiev, I., Mapelli, S., Karanov, E., 2001. The effect of drought andultraviolet radiation on growth and stress marker in pea and wheat. Plant CellEnviron. 24, 1337e1344.
Alscher, R.G., Erturk, N., Heath, L.S., 2002. Role of superoxide dismutases (SODs) incontrolling oxidative stress in plants. J. Exp. Bot. 53, 1331e1341.
Azevedo, R.A., Alas, R.M., Smith, R.J., Lea, P.J., 1998. Response of antioxidant enzymesto transfer from elevated carbon dioxide to air and ozone fumigation, in theleaves and roots of wild-type and a catalase deficient mutant of barley. Physiol.Plant 104, 280e292.
Azevedo, R.A., Carvalho, R.F., Cia, M.C., Gratão, P.L., 2011. Sugarcane under pressure:an overview of biochemical and physiological studies of abiotic stress. Trop.Plant Biol. 4, 42e51.
Basu, S., Roychoudhury, A., Saha, P.P., Sengupta, D.N., 2010. Differential antioxidativeresponse of indica rice cultivars to drought stress. Plant Growth Regul. 60, 51e59.
Beauchamp, C.H., Fridovich, I., 1971. Superoxide dismutase: improved assays and anassay applicable to acrylamide gels. Anal. Biochem. 44, 276e287.
Benesová, M., Holá, D., Fischer, L., Jedelsky, P.L., Hnilicka, F., Wilhelmová, N.,Rothová, O., Kocová, M., Procházková, D., Honnerová, J., Fridrichóva, L.,Hnilicková, H., 2012. The physiology and proteomics of drought tolerance inmaize: early stomatal closure as a cause of lower tolerance to short-termdehydration? PLoS ONE 7, e38017.
Boaretto, L.F., Mazzafera, P., 2013. The proteomes of feedstocks used for the pro-duction of second-generation ethanol: a lacuna in the biofuel era. Ann. Appl.Biol. 163, 12e22.
Bradford, M.M., 1976. A rapid and sensitive method for the quantification ofmicrogram quantities of protein utilizing the principle of protein-dye binding.Anal. Biochem. 72, 248e259.
Companhia Nacional de Abastecimento (CONAB), Web sugarcane service http://www.conab.gov.br/OlalaCMS/uploads/arquivos/12_04_10_09_19_04_boletim_de_cana.pdf. (accessed 27.08.12.).
Cheavegatti-Gianotto, A., Abreu, H.M.C., Arruda, P., Filho, J.C.B., Burnquist, W.L.,Creste, S., Ciero, L., Ferro, J.A., Figueira, A.V.O., Filgueiras, T.S., Grossi-de-Sá, M.F.,Guzzo, E.C., Hoffmann, H.P., Landell, M.G.A., Macedo, N., Matsuoka, S.,Reinach, F.C., Romano, E., Silva, W.J., Filho, M.C.S., Ulian, E.C., 2011. Sugarcane(Saccharum X officinarum): a reference study for the regulation of geneticallymodified cultivars in Brazil. Trop. Plant Biol. 4, 62e89.
Chew, O., Whelan, J., Millar, A.H., 2003. Molecular definition of the ascorbate-glutathione cycle in Arabidopsis mitochondria reveals dual targeting of anti-oxidant defense in plants. J. Biol. Chem. 278, 46869e46877.
Cia, M.C., Guimarães, A.C.R., Medici, L.O., Chabregas, S.M., Azevedo, R.A., 2012.Antioxidant response to water deficit by drought-tolerant and sensitive sug-arcane varieties. Ann. Appl. Biol. 161, 313e324.
Contour-Ansel, D., Torres-Franklin, M.L., Cruz de Carvalho, M.H., D’Arcy-Lameta, A.,Zuily-Fodil, Y., 2006. Glutathione reductase in leaves of cowpea: cloning of twocDNA, expression and enzyme activity under progressive drought stress,desiccation and abscisic acid treatment. Ann. Bot. 98, 1279e1287.
Cruz de Carvalho, M.H., 2008. Drought stress and reactive oxygen species: pro-duction, scavenging and signaling. Plant Signal. Behav. 3, 156e165.
Cruz de Carvalho, M.H., 2008. Drought stress and reactive oxygen species. PlantSignal. Behav. 3, 156e165.
Dat, J., Vandenabeele, S., Vranová, E., Montagu, M.V., Inzé, D., Breusegem, F.V., 2000.Dual action of the active oxygen species during plant stress responses. Cell. Mol.Life Sci. 57, 779e795.
Devi, R., Kaur, N., Gupta, A.K., 2012. Potential of antioxidant enzymes in depictingdrought tolerance of wheat (Triticum aestivum L.). Indian J. Biochem. Biophys.49, 257e265.
Dinardomiranda, L.L., Vasconcelos, A.C.M., Landell, M.G.A., 2008. Cana-de-Açúcar,first ed. Instituto Agronômico de Campinas, Campinas.
Doupis, G., Bertaki, M., Psarras, G., Kasapakis, I., Chartzoulakis, K., 2013. Water re-lations, physiological behavior and antioxidant defence mechanism of oliveplants subjected to different irrigation regimes. Sci. Hortic. 153, 150e156.
Dourado, M.N., Martins, P.F., Quecine, M.C., Piotto, F.A., Souza, L.A., Franco, M.R.,Tezotto, T., Azevedo, R.A., 2013. Burkholderia sp. SCMS54 reduces cadmiumtoxicity and promotes growth in tomato. Ann. Appl. Biol. 163, 494e507.
Duan, Z.-Q., Bai, L., Zhao, Z.G., Zhang, G.P., Cheng, F.M., Jiang, L.-X., Chen, K.-M., 2009.Drought-stimulated activity of plasma membrane nicotinamide adenine dinu-cleotide phosphate oxidase and its catalytic properties in rice. J. Integr. PlantBiol. 51, 1104e1115.
Gallego, S.M., Pena, L.B., Barcia, R.A., Azpilicueta, C.E., Iannone, M.F., Rosales, E.P.,Zawoznik, M.S., Groppa, M.D., Benavides, M.P., 2012. Unraveling cadmiumtoxicity and tolerance in plants: insight into regulatory mechanism. Environ.Exp. Bot. 83, 33e46.
Garcia, J.S., Gratão, P.L., Azevedo, R.A., Arruda, M.A.Z., 2006. Metal contaminationeffects on sunflower (Helianthus annus L.), growth and protein expression inleaves during development. J. Agr. Food Chem. 54, 8623e8630.
Ghane, S.G., Lokhande, V.H., Nikam, T.D., 2012. Differential growth, physiologicaland biochemical responses of niger (Guizotia abyssinica Cass.) cultivars towater-deficit (drought) stress. Acta Physiol. Plant 34, 215e225.
Giannopolitis, C.N., Ries, S.K., 1977. Superoxide dismutase. I. Occurrence in higherplants. Plant Physiol. 59, 309e314.
Gratão, P.L., Polle, A., Lea, P.J., Azevedo, R.A., 2005. Making the life of heavy metal-stressed plants a little easier. Funct. Plant Biol. 32, 481e494.
Gratão, P.L., Monteiro, C.C., Carvalho, R.F., Tezotto, T., Piotto, F.A., Peres, L.E.P.,Azevedo, R.A., 2012. Biochemical dissection of diageotropica and Neverripe tomato mutants to Cd-stressful condition. Plant Physiol. Biochem. 56,79e96.
Impa, S.M.S., Nadaradjan, S., Jagadish, S.V.K., 2012. Drought stress induced reactiveoxygen species and anti-oxidants in plants. In: Ahmad, P., Prasad, M.N.V. (Eds.),Abiotic Stress Responses in Plants. Springer, New York, pp. 131e147.
Inman-Bamber, N.G., De Jager, J.M., 1986. The reaction of two varieties of sugarcaneto water stress. Field Crop Res. 14, 15e28.
Jiang, M., Zhang, J., 2002. Water stress-induced abscisic acid accumulation triggersthe increase generation of reactive oxygen species and up-regulates the activ-ities of antioxidant enzymes in maize leaves. J. Exp. Bot. 53, 2401e2410.
Kar, R.K., 2011. Plant responses to water stress. Role of reactive oxygen species. PlantSignal. Behav. 6, 1741e1745.
Kaur, K., Kaur, N., Gupta, A.K., Singh, I., 2013. Exploration of the antioxidative de-fense system to characterize chickpea genotypes showing differential responsetowards water deficit conditions. Plant Growth Regul. 70, 49e60.
Keles, Y., Oncel, I., 2002. Response of the antioxidant defense system to temperatureand water stress combinations in wheat seedling. Plant Sci. 163, 783e790.
Lambais, M.R., 2001. In silico differential display of defense-related expressedsequence tags from sugarcane tissues infected with diazotrophic endophytes.Genet. Mol. Biol. 24, 103e111.
Landell, M.G.A., Campana, M.P., Figueiredo, P., Vasconcelos, A.C.M., Xavier, M.A.,Bidoia, M.A.P., Prado, H., Silva, M.A., Dinardo-Miranda, L.L., Santos, A.S.,Perecin, D., Rossetto, R., Silva, D.N., Martins, A.L.M., Gallo, P.B., Kantack, R.A.D.,Cavichioll, J.C., Veiga Filho, A.A., Anjos, I.A., Azania, C.A.A.M., Pinto, L.R.,Souza, S.A.C.D., 2005. Variedades de cana-de-açúcar para o Centro-Sul do Brasil.Instituto Agronômico, Campinas, p. 33 (IAC. Boletim técnico, 197); (série tec-nologia APTA).
Lascano, H.R., Antonicelli, G.E., Luna, C.M., Melchiorre, M.N., Gómez, L.D.,Racca, R.W., Troppi, V.S., Casano, L.M., 2001. Antioxidant system response ofdifferent wheat cultivar under drought: field and in vitro studies. Aust. J. PlantPhysiol. 28, 1095e1102.
Machado, R.S., Ribeiro, R.V., 2012. Integrating Physiological, Morphological andAnatomical Traits to Understand the Differential Sucrose Yield in SugarcaneGenotype. FAPESP. Web site http://www.fapesp.br/publicacoes/pasta_bioen_jun2012.pdf (accessed 16.01.13.).
Machado, R.S., Ribeiro, R.V., Marchiori, P.E.R., Machado, D.F.S.P., Machado, E.C.,Landell, M.G.A., 2009. Biometric and physiological responses to water deficitin sugarcane at different phenological stages. Pesq. Agropecu. Bras. 12, 1575e1582.
Malan, C., Greyling, M.M., Gressel, J., 1990. Correlation between Cu/Zn superoxidedismutase and glutathione reductase, and environmental and xenobiotic stresstolerance in maize inbreds. Plant Sci. 69, 157e166.
Marcos, F.C.C., 2012. Influência de bactérias endofíticas na fisiologia de plantas decana-de-açúcar sob restrição hídrica. Master thesis in Tropical and SubtropicalAgriculture. Agronomic Institute of Campinas, Campinas.
Markwell, J., Osterman, J.C., Mitchell, J.L., 1995. Calibration of the Minolta SPAD-502leaf chlorophyll meter. Photosynth. Res. 46, 467e472.
Martins, P.F., Carvalho, G., Gratão, P.L., Dourado, M.N., Pileggi, M., Araújo, W.L.,Azevedo, R.A., 2011. Effects of the herbicides acetochlor and metalachlor onantioxidant enzyme in soil bacteria. Process. Biochem. 46, 1186e1195.
Maxwell, D., Nickels, R., McIntosh, L., 2002. Evidence of mitochondrial involvementin the transduction of signal required for the induction of gene associated withpathogen attack and senescence. Plant J. 29, 269e279.
Medici, L.O., Rocha, H.S., Carvalho, D.F., Pimentel, C., Azevedo, R.A., 2010. Automaticcontroller to water plants. Sci. Agric. 67, 727e730.
Medici, L.O., Reinert, F., Carvalho, D.F., Kozak, M., Azevedo, R.A., 2014. What aboutkeeping plants well watered? Environ. Exp. Bot. 99, 38e42.
Miller, G., Suzuki, N., Ciftci-Yilmaz, S., Mittler, R., 2010. Reactive oxygen specieshomeostasis and signalling during drought and salinity stresses. Plant CellEnviron. 33, 453e467.
Mittler, R., 2002. Oxidative stress, antioxidants and stress tolerance. Trends PlantSci. 7, 405e410.
Monteiro, C.C., Carvalho, R.F., Gratão, P.L., Carvalho, G., Tezotto, T., Medici, L.O.,Peres, L.E.P., Azevedo, R.A., 2011. Biochemical responses of the ethylene-insensitive Never ripe tomato mutant subjected to cadmium and sodiumstress. Environ. Exp. Bot. 71, 306e320.
Mori, I.C., Schroeder, J.I., 2004. Reactive oxygen species activation of plant Ca2þ
channels. A signaling mechanism in polar growth, hormone transduction,stress signaling, and hypothetically mechanotransduction. Plant Physiol. 135,702e708.
Pires, R.C.M., Arruda, F.B., Sakai, E., 2008. Irrigação e drenagem. In: Dinardo-Miranda, L.L., Vasconcelos, A.C.M., Landell, M.G.A. (Eds.), Cana-de-açúcar.Instituto Agronômico de Campinas, Campinas, p. 882.
Rampino, P., Pataleo, S., Gerardi, C., Mita, G., Perrotta, S.D., 2006. Drought stressresponse in wheat: physiological and molecular analysis of resistant and sen-sitive genotypes. Plant Cell Environ. 29, 2143e2152.
Ratnayaka, H.H., Molin, W.T., Sterling, T.M., 2003. Physiological and antioxidantresponse of cotton and spurred anoda under interference and mild drought.J. Exp. Bot. 54, 2293e2305.
Ribeiro, R.V., Machado, R.S., Machado, E.C., Machado, D.F.S.P., Magalhães Filho, J.R.,Landell, M.G.A., 2013. Revealing drought-resistance and productive patterns insugarcane genotypes by evaluating both physiological responses and stalk yield.Exp. Agric. 49, 212e224.
L.F. Boaretto et al. / Plant Physiology and Biochemistry 74 (2014) 165e175174

Author's personal copy
Richards, L.A., 1949. Methods of measuring soil moisture tension. Soil Sci. 68, 95e112.
Rosegrant, M.W., Ringler, C., Zhu, T., 2009. Water for Agriculture: maintaining foodsecurity under growing scarcity. Annu. Rev. Env. Resour. 34, 205e222.
Selote, D.S., Khanna-Chopra, R., 2006. Drought acclimation confers oxidative stresstolerance by inducing co-ordinated antioxidant defense at cellular and sub-cellular level in leaves of wheat seedlings. Physiol. Plant 127, 494e506.
Sgherri, C.L.M., Maffei, M., Navari-Izzo, F., 2000. Antioxidative enzymes in wheatsubjected to increasing intensity of water deficit and rewatering. J. PlantPhysiol. 157, 273e279.
Sharma, P., Dubey, R.S., 2005. Drought induces oxidative stress and enhances theactivity of antioxidant enzymes in growing rice seedling. Plant Growth Regul.46, 209e221.
Shigeoka, S., Ishikawa, T., Tamoi, M., Miyagawa, Y., Takeda, T., Yabuta, Y.,Yoshimura, K., 2002. Regulation and function of ascorbate peroxidase iso-enzymes. J. Exp. Bot. 53, 1305e1319.
Silva, M.A., Jifon, J.L., Da Silva, J.A.G., Sharma, V., 2007. Use of physiological pa-rameters as a fast tool to screen for drought tolerance in sugarcane. Braz. J. PlantPhysiol. 19, 193e201.
Simova-Stoilova, L., Dermirevska, K., Petrova, T., Tsenov, N., Feller, U., 2009. Anti-oxidative protection and proteolytic activity in tolerant and sensitive wheat(Triticum aestivum L.) varieties subjected to long-term field drought. PlantGrowth Regul. 58, 107e117.
Smart, R.E., Bingham, G.E., 1974. Rapid estimates of relative water content. PlantPhysiol. 53, 258e260.
Smith, I.K., Vierheller, T.L., Thorne, C.A., 1988. Assay of glutathione reductase incrude tissue homogenates using 5,5’-dithiobis(2-nitrobenzoic acid). Anal. Bio-chem. 175, 408e413.
Soares Netto, L.E., 2001. Oxidative stress response in sugarcane. Genet. Mol. Biol. 24,93e102.
Srivastava, S., Pathak, A.D., Gupta, P.S., Shrivastava, A.K., Srivastava, A.K., 2012.Hydrogen peroxide-scavenging enzymes impart tolerance to high temperatureinduced oxidative stress in sugarcane. J. Environ. Biol. 33, 657e661.
Torres-Franklin, M.L., Gigon, A., Fernandes de Melo, D., Zuily-Fodil, Y., Pham-Thi, A.T., 2007. Drought stress and rehydration affect the balance betweenMGDG and DGDG synthesis in cowpea leaves. Physiol. Plant 131, 201e210.
Türkan, I., Bor, M., Özdemir, F., Koca, H., 2005. Differential responses of lipid per-oxidation and antioxidants in the leaves of drought-tolerant P. acutifolius Grayand drought-sensitive P. vulgaris L. subjected to polyethylene glycol mediatedwater stress. Plant Sci. 168, 223e231.
Uzilday, B., Turkan, I., Sekmen, A.H., Ozgur, R., Karakaya, H.C., 2012. Comparison ofROS formation and antioxidant enzymes in Cleome gynandra (C-4) and Cleomespinosa (C-3) under drought stress. Plant Sci. 182, 59e70.
Wiedenfeld, B., 2004. Scheduling water application on drip irrigated sugarcane. Agr.Water Manage 64, 169e181.
Willadino, L., de Oliveira, R.A., da Silva, E.A., Neto, A.G., Camara, T.R., 2011. Salinitystress in two varieties of sugar cane: enzymes of the antioxidant system andchlorophyll fluorescence. Rev. Cien. Agron. 42, 417e422.
Woodbury, W., Spencer, A.K., Stahmann, M.A., 1971. An improved procedure usingferricyanide for detecting catalase isozymes. Anal. Biochem. 44, 301e305.
L.F. Boaretto et al. / Plant Physiology and Biochemistry 74 (2014) 165e175 175




















