
Volatile organic compounds from Italian vegetation and their interaction with ozone
9
Volatile organic compounds from Italian vegetation and their interaction with ozone Carlo Calfapietra * , Silvano Fares, Francesco Loreto CNR-Istituto di Biologia Agroambientale e Forestale, Via Salaria Km. 29.300, 00015 Monterotondo Scalo (Roma), Italy The capacity to emit isoprenoids confers to plants tolerance to O 3 and alters the flux of O 3 between atmosphere and biosphere. article info Article history: Received 22 September 2008 Accepted 26 September 2008 Keywords: VOC Ozone flux Isoprene Monoterpene Antioxidant abstract Volatile Organic Compounds (VOCs) emitted from vegetation (particularly isoprenoids) represent an important source of atmospheric hydrocarbons almost double the anthropogenic source. When biogenic VOC mix with NO x in the presence of UV radiation, ozone (O 3 ) is formed. In Italy, optimal conditions for O 3 formation in terms of VOC/NO x ratios and abundance of UV radiation occur for long periods of the year. Moreover, Italian vegetation includes several species that are strong and evergreen isoprenoid emitters, and high temperatures for part of the year further stimulate these temperature-dependent emissions. We review emission of isoprenoids from Italian vegetation, current knowledge on the impact of rising O 3 levels on isoprenoid emission, and evidence showing that isoprenoids can increase both the O 3 flux to the plant and protection against oxidative stress because of their antioxidant functions. This trait not only influences plant tolerance to O 3 but also may substantially alter the flux of O 3 between atmosphere and biosphere. Ó 2008 Elsevier Ltd. All rights reserved. 1. Emission of isoprenoids from Italian vegetation Isoprenoid emission from vegetation has been discovered some decades ago when scientists started to investigate the origin of the ‘‘blue haze’’ often observed above forests (Went, 1960). Isoprenoids represent an important class of Volatile Organic Compounds (VOCs), including compounds that are emitted in large amounts by plants and have important implications for the chemistry of the atmosphere. It is estimated that VOC emission from plants accounts for 1150 Tg Cyear 1 , almost double the anthropogenic emission of VOCs (Guenther et al., 1995). Isoprene (C 5 H 8 , 2-methyl- 1,3,-butadiene), the simplest and most volatile isoprenoid, makes up as much as 80 % of the carbon emitted as VOC emissions from plants. Monoterpenes (C 10 H 16 ), isoprenoids equivalent to two isoprene units, are also abundantly emitted by vegetation, although the physical properties of these molecules make their volatility and, their emission rates lower than for isoprene (Nii- nemets et al., 2004). Many monoterpenes can be formed by different enzymes that synthesize the C-10 molecule (synthases) and then rearrange the molecule (isomerases). Acyclic (e.g. myr- cene), or cyclic monoterpenes with one ring (d-limonene) or two ring structures (a- and b-pinene, 3-carene) are all emitted by vegetation. Monoterpene emission can be distinguished in light- dependent and in a light-independent manner. The light-inde- pendent ones are expected to follow different responses to envi- ronmental changes especially to changes in temperatures because of the storage in the leaf (Copolovici and Niinemets, 2005). Sesquiterpenes are a third class of volatile isoprenoids that are formed by three isoprene units. Like monoterpenes, sesquiterpenes may be acyclic or contain rings, including many unique combina- tions. Among sesquiterpenes, b-caryophyllene is the most abun- dant compound (Bonn and Moortgat, 2003). Contrary to more volatile isoprenoids, sesquiterpenes have not been extensively investigated up to now. Because of their low abundance in the gas phase, and also because of their very high reactivity, resulting in a lifetime of few seconds once sesquiterpenes are emitted in the atmosphere from vegetation, sesquiterpenes in the atmosphere are difficult to measure. However, recent studies suggest that their role is crucial and deserves a more detailed investigation, both in plant– herbivore interactions (Gershenzon and Dudareva, 2007), and for understanding the chemical reactions driven by biogenic VOCs in the atmosphere (Bonn and Moortgat, 2003). The homoterpenes represent another category of VOC (either C 11 or C 16 ) emitted by plants during flowering or when they are damaged (Boland and Ga ¨bler, 1989; Degenhardt and Gershenzon, 2000) but at the moment it is still difficult to assess what is the importance of these compounds in the context of Italian vegetation. Many plant species emit isoprenoids although both qualitatively and quantitatively the emission rates are very different in the * Corresponding author. Tel.: þ39 06 90672530; fax: þ39 06 9064492. E-mail address: [email protected] (C. Calfapietra). Contents lists available at ScienceDirect Environmental Pollution journal homepage: www.elsevier.com/locate/envpol 0269-7491/$ – see front matter Ó 2008 Elsevier Ltd. All rights reserved. doi:10.1016/j.envpol.2008.09.048 Environmental Pollution 157 (2009) 1478–1486
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Volatile organic compounds from Italian vegetation and their interaction with ozone

lable at ScienceDirect
Environmental Pollution 157 (2009) 1478–1486
Contents lists avai
Environmental Pollution
journal homepage: www.elsevier .com/locate/envpol
Volatile organic compounds from Italian vegetation and their interactionwith ozone
Carlo Calfapietra*, Silvano Fares, Francesco LoretoCNR-Istituto di Biologia Agroambientale e Forestale, Via Salaria Km. 29.300, 00015 Monterotondo Scalo (Roma), Italy
The capacity to emit isoprenoids confers to plants tolerance to O3 and
alters the flux of O3 between atmosphere and biosphere.a r t i c l e i n f o
Article history:Received 22 September 2008Accepted 26 September 2008
Keywords:VOCOzone fluxIsopreneMonoterpeneAntioxidant
* Corresponding author. Tel.: þ39 06 90672530; faE-mail address: [email protected] (C. Cal
0269-7491/$ – see front matter � 2008 Elsevier Ltd.doi:10.1016/j.envpol.2008.09.048
a b s t r a c t
Volatile Organic Compounds (VOCs) emitted from vegetation (particularly isoprenoids) represent animportant source of atmospheric hydrocarbons almost double the anthropogenic source. When biogenicVOC mix with NOx in the presence of UV radiation, ozone (O3) is formed. In Italy, optimal conditions forO3 formation in terms of VOC/NOx ratios and abundance of UV radiation occur for long periods of theyear. Moreover, Italian vegetation includes several species that are strong and evergreen isoprenoidemitters, and high temperatures for part of the year further stimulate these temperature-dependentemissions. We review emission of isoprenoids from Italian vegetation, current knowledge on the impactof rising O3 levels on isoprenoid emission, and evidence showing that isoprenoids can increase both theO3 flux to the plant and protection against oxidative stress because of their antioxidant functions. Thistrait not only influences plant tolerance to O3 but also may substantially alter the flux of O3 betweenatmosphere and biosphere.
� 2008 Elsevier Ltd. All rights reserved.
1. Emission of isoprenoids from Italian vegetation
Isoprenoid emission from vegetation has been discovered somedecades ago when scientists started to investigate the origin of the‘‘blue haze’’ often observed above forests (Went, 1960). Isoprenoidsrepresent an important class of Volatile Organic Compounds(VOCs), including compounds that are emitted in large amounts byplants and have important implications for the chemistry of theatmosphere. It is estimated that VOC emission from plantsaccounts for 1150 Tg C year�1, almost double the anthropogenicemission of VOCs (Guenther et al., 1995). Isoprene (C5H8, 2-methyl-1,3,-butadiene), the simplest and most volatile isoprenoid, makesup as much as 80 % of the carbon emitted as VOC emissions fromplants. Monoterpenes (C10H16), isoprenoids equivalent to twoisoprene units, are also abundantly emitted by vegetation,although the physical properties of these molecules make theirvolatility and, their emission rates lower than for isoprene (Nii-nemets et al., 2004). Many monoterpenes can be formed bydifferent enzymes that synthesize the C-10 molecule (synthases)and then rearrange the molecule (isomerases). Acyclic (e.g. myr-cene), or cyclic monoterpenes with one ring (d-limonene) or tworing structures (a- and b-pinene, 3-carene) are all emitted byvegetation. Monoterpene emission can be distinguished in light-
x: þ39 06 9064492.fapietra).
All rights reserved.
dependent and in a light-independent manner. The light-inde-pendent ones are expected to follow different responses to envi-ronmental changes especially to changes in temperatures becauseof the storage in the leaf (Copolovici and Niinemets, 2005).Sesquiterpenes are a third class of volatile isoprenoids that areformed by three isoprene units. Like monoterpenes, sesquiterpenesmay be acyclic or contain rings, including many unique combina-tions. Among sesquiterpenes, b-caryophyllene is the most abun-dant compound (Bonn and Moortgat, 2003). Contrary to morevolatile isoprenoids, sesquiterpenes have not been extensivelyinvestigated up to now. Because of their low abundance in the gasphase, and also because of their very high reactivity, resulting ina lifetime of few seconds once sesquiterpenes are emitted in theatmosphere from vegetation, sesquiterpenes in the atmosphere aredifficult to measure. However, recent studies suggest that their roleis crucial and deserves a more detailed investigation, both in plant–herbivore interactions (Gershenzon and Dudareva, 2007), and forunderstanding the chemical reactions driven by biogenic VOCs inthe atmosphere (Bonn and Moortgat, 2003).
The homoterpenes represent another category of VOC (eitherC11 or C16) emitted by plants during flowering or when they aredamaged (Boland and Gabler, 1989; Degenhardt and Gershenzon,2000) but at the moment it is still difficult to assess what is theimportance of these compounds in the context of Italian vegetation.
Many plant species emit isoprenoids although both qualitativelyand quantitatively the emission rates are very different in the

Table 1Main tree species of Italian vegetation listed from the most to the less abundant interms of coverage in natural ecosystems (excluding plantations; from Lenz et al.,2001) with their emission trait (adapted from Steinbrecher et al., in press). Emissionrates for the different species are listed as:
Species Isopreneemission
Monoterpeneemission
Approximate abundancein Italy (%)
Fagus sylvatica High 18Quercus pubescens High Low 13Quercus cerris Low 11Castanea sativa Medium 8Picea abies Low Low 6Larix decidua Medium 5Ostrya carpinifolia 4Pinus nigra Medium 3Quercus ilex High 3Pinus sylvestris Medium 3Quercus petraea High Low 2Abies alba Low Low 1Alnus cordata Low 1Acer platanoides Low 1Robinia pseudoacaciaa Medium <1Fraxinus sp. <1Pinus pineaa Medium <1Quercus robur High Low <1Pinus halepensis Low <1Pinus pinaster Low <1Cupressus sempervirens Low <1Quercus suber Low Medium <1Betula pendula Low <1Salix sp.a High Low <1Populus sp.a High <1Carpinus betulus Low <1Tilia cordata <1Eucalyptus sp.a High Medium <1Mediterranean macchiab Low N/A
Empty cells: emission rates close to 0, Low: emission rates ranging from 0.1 to3 mg g�1 h�1; Medium: emission rates ranging from 3 to 20 mg g�1 h�1; High:emission rates above 20 mg g�1 h�1; Monoterpene emission includes both themonoterpene synthesis emission (light and temperature dependent) and themonoterpene pool emission (temperature dependent).
a These species are commonly used in plantations, thus increasing considerablythe potential emission at national scale.
b This ecosystem includes a large number of shrub species which are generallymonoterpenes emitter (BEMA, 1997).
C. Calfapietra et al. / Environmental Pollution 157 (2009) 1478–1486 1479
different species (Wiedinmyer et al., 2004). In terms of globalinventory, forests and especially fast growing tree plantations arethe biggest source of isoprenoids; trees such as oaks, poplars, andeucalyptus are strong emitters. Most of the deciduous trees emitisoprene, whereas coniferous woodland such as pine trees, cedarsand firs emit predominantly monoterpenes (Seufert et al., 1997;Guenther et al., 1994). Some species emit both isoprene andmonoterpenes such as spruce, eucalyptus and myrtle, whereas theemission of isoprenoids is negligible or absent altogether in severalother species (Steinbrecher, 1997; Kesselmeier and Staudt, 1999.).
Italian vegetation includes a large number of forest species,reflecting the environmental and climatological areas of thecountry that span from alpine to semiarid environments. Becauseof this wide range of ecosystems, the variability in terms of iso-prenoid emission is particularly large over the Italian territory(BEMA, 1997; Lenz et al., 2001).
An inventory of isoprenoid emission by forest tree species isunder construction for Italy under the project FISR-CarboItaly(Papale, 2006), based on data of basal emission of each singlespecies, i.e. the emission measured under standard environmentalconditions (Guenther et al., 1991), which is then spatially integratedby knowing the species-specific land cover with satellite images. Asalpine forests are dominated by spruce, firs, larch, pines and, toa lesser extent, beech, monoterpenes predominate over isopreneand, given the low temperatures in alpine areas, usually the totalemissions per unit area are quite low. In the Po Valley, a big sourceof isoprene is represented by poplar plantations given their highpotential both in terms of yield and isoprene emission. Althoughtree plantations in Italy currently occupy 122,000 ha, representingonly 1.5 % of the total forested area (INFC, 2007), the potential forincrease of isoprene emission is represented by the spreading ofshort rotation plantations characterized by high planting densities.On the Apennines, forests are dominated at higher elevations bybeech, which is a quite strong emitter of light and temperature-dependent monoterpenes (Dindorf et al., 2006). At lower elevationsApennine forests are dominated by oaks and, in some particularlysuitable areas, by chestnuts. While chestnut is supposed to bea medium emitter (Table 1), oaks include both species among thestrongest isoprenoid emitters and species characterized by veryweak emissions (Loreto, 2002; Table 1). This variability makes thecharacterization of spatial emissions very difficult. For instanceforests of central and southern Italy are dominated by a co-occur-rence of Quercus pubescens and Quercus cerris, the former beinga strong isoprene emitter, the latter known to be a modest (almostnegligible) emitter. Moving further south or closer to the sea cost,evergreen oaks begin to be predominant. However, while Quercusilex is a strong monoterpene emitter, the emission by Quercus suberis more controversial since reports have described this species asa very low monoterpene emitter (Steinbrecher et al., 1997) whilerecently it has been reported as a medium monoterpene emitter(Steinbrecher et al., in press) (Table 1). Finally, most of the coastsand large areas of southern Italy are dominated by the typicalMediterranean macchia, characterized by evergreen shrubs orsmall trees of different species. Most of these species are known tobe monoterpene emitters although for most of them the amountsemitted are low (Owen et al., 1997).
As briefly mentioned above, leaf temperature is the environ-mental factor that exerts significant control of isoprenoid emission.This reflects the strong temperature-dependence of synthases thatcatalyze isoprenoid formation (Monson et al., 1992) and thetemperature-dependence of volatility. Consequently, high emis-sions of isoprenoids are expected during summer, which makesItaly and the Mediterranean likely significant sources of isoprenoidemissions. Hot periods with temperature ranging around 30 �C aregenerally associated by metabolic limitations that limit primarymetabolism of plants, e.g. heavy drought stress. However, it is now
firmly established that isoprenoid biosynthesis is resistant todrought stress and may even be stimulated when plants areexposed to recurrent stress periods before the final inhibition whenthe stress becomes too limiting (Sharkey and Loreto, 1993; Funket al., 2004; Brilli et al., 2007).
2. Role of isoprenoids in the chemical reactivity of thetroposphere
A very important implication of biogenic emission of iso-prenoids is that these compounds play a fundamental role in theformation of the photochemical smog in the troposphere, leadingto the formation of ozone (O3) and other secondary pollutants suchas peroxyacyl nitrates (PAN), aldehydes and ketones, hydrogenperoxide (H2O2), secondary organic aerosol (SOA) and particlematters (PM) (Fehsenfeld et al., 1992; Fuentes et al., 2000).
In particular the reaction VOC-OX (defined as VOC compoundswith high potential of photochemical O3 formation) with OHradicals produces other radicals:
VOC L OX D OH / R�D ROH�
which can react with oxygen generating peroxide radicals:
R�D O2 / RO2�
ROH D O2 / RðOHÞO2�

C. Calfapietra et al. / Environmental Pollution 157 (2009) 1478–14861480
peroxide radicals, reacting with nitric oxide (NO), produce nitrogendioxide (NO2):
RO2�D NO / RO�D NO2
RðOHÞO2�D NO / RðOHÞO�D NO2
These reactions lead to an increase of the concentrations of NO2
to the detriment of NO, with consequent increase of O3 concen-tration, produced by the reactions:
NO2 D hv / NO D O�
OL D O2 / O3
In summary, the presence of biogenic VOC, and in particular ofisoprenoids, alters the cycle NO–NO2–O3 responsible for O3
formation and degradation (Ciccioli et al., 1999; Atkinson, 2000; DiCarlo et al., 2004). However, isoprene and monoterpenes havea different potential of O3 formation (POF), which can be expressedin terms of grams of O3 produced per gram of molecule. For instance,the POF is about 9.1 for isoprene but only 3.3 for a-pinene, the mostabundant monoterpene usually emitted by vegetation (Carter,1994). The species-specific emission rates of isoprenoids, togetherwith the POF and the meteorological and climatic conditions of theatmosphere, ultimately determine the potential impact of biogenicisoprenoids on the atmospheric chemistry. In particular, the ratio ofnon-methane hydrocarbons (i.e. VOCs) to NOx is probably the mostimportant parameter to characterize the efficiency of O3 formationin VOC–NOx–air mixtures. At low VOC-limited ratios (< 5), O3
formation is slow and inefficient. Decreasing NOx levels may resultin unexpected increase of O3 formation under VOC-limited condi-tions that are often common in urban areas. At high ratios (>15–20),O3 formation is limited by availability of NOx rather than VOC (NOx-limited). Ratios between 5 and 15 are considered transitional, withan optimal production of O3 estimated in correspondence of VOC/NOx ratio around 8 (Derwent et al.,1996). The principle that the ratioof VOC to NOx is the driver of ozone production explains why, inmany occasions, O3 levels are higher in the countryside rather thanin urban areas, where VOCs are low. This is particularly true forCentral and Southern Italy where the air masses rich of NOx
produced in the large urban areas along the sea cost are transportedby air circulation, characterized by diurnal sea breezes in a regime ofsteady atmospheric conditions, towards the hills. Over the hills, theair transported from urban areas mixes with large amounts ofbiogenic VOCs, produced especially from oak forests. As a result,high peaks of O3 are registered daily in many rural areas, eventhough these are far away from sources of NOx (Millan et al., 1997).
The increase of OH radicals subsequent to the oxidation of VOCsleads also to secondary reactions such as the oxidation of NO2 and SO2
to produce acids (HNO3 and H2SO4), which are responsible for wet anddry acid depositions and for the formation of fine particles (smallerthan 1 mm) and secondary organic aerosols (Andreae and Crutzen,1997). In order to assess the importance of the biogenic emissions in O3
and aerosol formation the understanding of the OH reactivity becomescrucial. In a recent study, Di Carlo et al. (2004) showed that aboveforests total OH reactivity was significantly greater than expected, andprovided measurements and calculations which are consistent withthe hypothesis that unknown reactive BVOCs, probably terpenesand/or sesquiterpenes, are responsible for the missing OH reactivity.
3. Impact of increasing O3 levels on isoprenoid biosynthesisand emission
While isoprenoid emission has a crucial impact on O3 formationin the troposphere, the concentration of O3 itself can influence the
physiology of the plants thus having important repercussion onisoprenoid biosynthesis and emission. In the long term theseinteractions can become very complex because a series of indirectand direct effects can be induced as well as negative or positivefeedbacks.
Direct effect of O3 exposure on isoprenoid emission at the leaflevel can be detected by leaf gas exchange measurements understandard conditions of light and temperature (basal emission).Unfortunately, no estimates are available on the effect of increasingO3 levels on isoprenoid emission at global scale because of thecomplex interactions at ecosystem level.
There have been several studies of the direct effects ofincreasing O3 levels on the emission of isoprenoids from strongemitters of the Italian vegetation, especially Quercus sp. and Pop-ulus sp. Loreto et al. (2004) investigated the effect of growing O3
concentrations from 100 to 200 ppb on monoterpene emission, byfumigating with the web O3 fumigation technique small trees of Q.ilex for 4 h per day, for five days. Both fumigation of the wholeplants and of single leaves resulted in a stimulation of the threemost abundant monoterpenes emitted by Q. ilex, namely a-pinene,b-pinene and sabinene. In a similar experiment performed usingthe same technique, whole trees of Q. pubescens, a species known tobe a strong isoprene emitter, were exposed to O3 levels variablesbetween 100 and 300 ppb for three days (Pinelli et al., 2004). Nodifference in isoprene emission was observed in the leaves of thebranches exposed to about 100 ppb as compared to the onesexposed to ambient O3. However, isoprene emission considerablydecreased in those leaves that were exposed to about 190 ppb, andeven more in leaves exposed to 300 ppb.
In other experiments, the O3 treatment was carried out usingcuvettes that expose to the pollutant only single leaves or part ofthe leaves, or a branch. Velikova et al. (2005) observed an increaseof isoprene emission from single leaves of Phragmites exposed to300 ppb of O3 for 3 h. The same effect was observed in leavespreviously treated with fosmidomycin to inhibit isoprene emission,in which O3 slightly stimulated the residual emission of isoprene.Fares et al. (2006) used large cuvettes to fumigate whole branchesof seedlings of Populus alba with O3 at a concentration of 150 ppbfor 11 h per day and for one month. Leaves developing inside thecuvette fumigated with O3 showed an increased isoprene emissioncompared to the leaves in cuvettes grown under ambient O3 levels.Surprisingly, the first leaf developed outside the cuvette fumigatedwith O3 was also induced to emit more isoprene suggesting that thesignal induced by O3 and stimulating isoprene biosynthesis istransported inside the plant. However, this effect was lost movingtowards the apex, i.e. in still-developing leaves, and at increasingdistance from the O3-fumigated portion of the plant. A stimulationof isoprene emission from Populus nigra seedlings exposed to highO3 levels was observed also in another experiment performed usinglarge cuvettes to fumigate the entire plants (Fares et al., 2008).
In summary, experiments that were carried out on small plants,using high concentrations of O3 (100–300 ppb) for short time (daysto weeks) to simulate an acute stress generally indicated that iso-prenoid biosynthesis is resistant to O3 stress, and that the emissionof these compounds may even be stimulated after such a period ofexposure to the pollutant. This is relevant information for mecha-nistic studies that investigate biosynthesis and functional roles ofisoprenoids. However, the above studies might not reflect real-lifeconditions that are generally characterized by chronic exposures tomuch lower levels of the pollutant, by a combined presence of highlevels of O3 and other air pollutants, and by the likely simultaneousoccurrence of environmental stresses such as high light and hightemperature when atmospheric O3 builds up. Moreover, cuvettemeasurements are usually carried out at the end of the O3 fumi-gation, using an O3-free, clean and well stirred flux of air, whichexcludes isoprenoid removal by direct reactions with the pollutant
C. Calfapietra et al. / Environmental Pollution 157 (2009) 1478–1486 1481
in the mesophyll, and in the gaseous phase, inside leaves or in theboundary layer between leaves and the atmosphere. Recently,measurements of isoprene emission have been carried out froma Populus tremuloides stand exposed to a chronic O3 exposure (1.5-fold the ambient levels) for several years in the AspenFACE exper-iment in open field conditions (Calfapietra et al., 2007). Under highO3 treatment, isoprene emission decreased significantly due toboth a reduction in isoprene synthase messenger RNA and isoprenesynthase protein and a reduction in the levels of dimethylallyldiphosphate (DMADP), the main substrate for isoprene synthesis(Calfapietra et al., 2008).
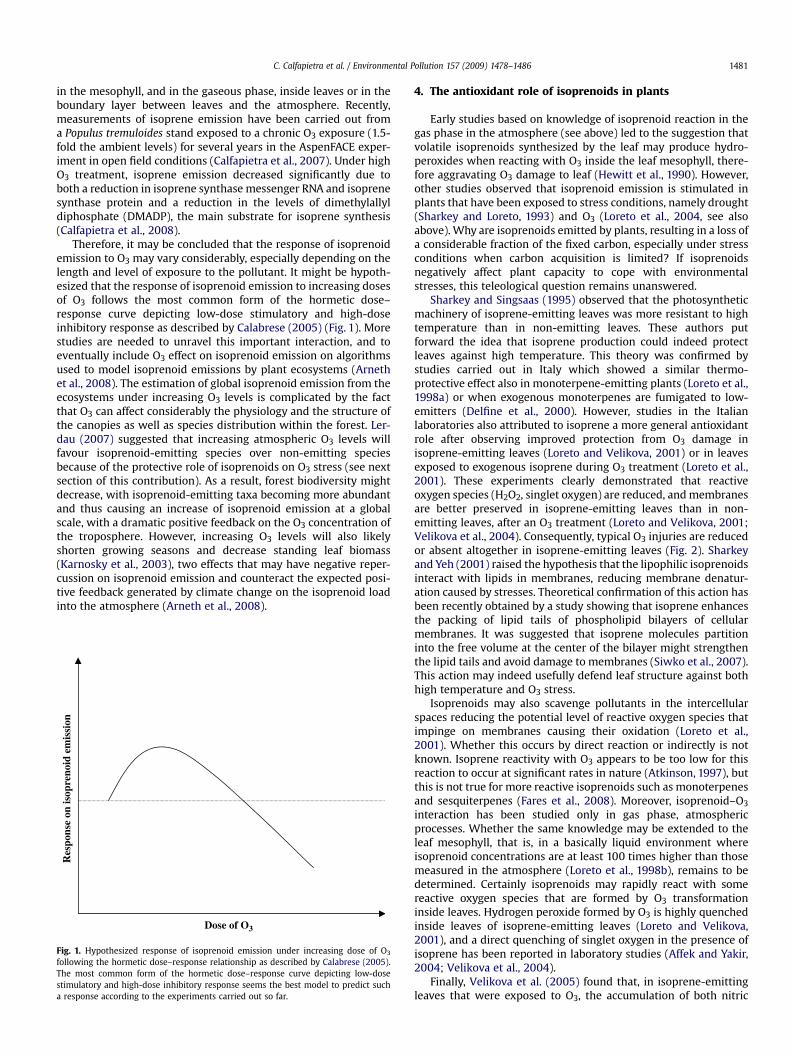
Therefore, it may be concluded that the response of isoprenoidemission to O3 may vary considerably, especially depending on thelength and level of exposure to the pollutant. It might be hypoth-esized that the response of isoprenoid emission to increasing dosesof O3 follows the most common form of the hormetic dose–response curve depicting low-dose stimulatory and high-doseinhibitory response as described by Calabrese (2005) (Fig. 1). Morestudies are needed to unravel this important interaction, and toeventually include O3 effect on isoprenoid emission on algorithmsused to model isoprenoid emissions by plant ecosystems (Arnethet al., 2008). The estimation of global isoprenoid emission from theecosystems under increasing O3 levels is complicated by the factthat O3 can affect considerably the physiology and the structure ofthe canopies as well as species distribution within the forest. Ler-dau (2007) suggested that increasing atmospheric O3 levels willfavour isoprenoid-emitting species over non-emitting speciesbecause of the protective role of isoprenoids on O3 stress (see nextsection of this contribution). As a result, forest biodiversity mightdecrease, with isoprenoid-emitting taxa becoming more abundantand thus causing an increase of isoprenoid emission at a globalscale, with a dramatic positive feedback on the O3 concentration ofthe troposphere. However, increasing O3 levels will also likelyshorten growing seasons and decrease standing leaf biomass(Karnosky et al., 2003), two effects that may have negative reper-cussion on isoprenoid emission and counteract the expected posi-tive feedback generated by climate change on the isoprenoid loadinto the atmosphere (Arneth et al., 2008).
Dose of O3
Res
pons
e on
isop
reno
id e
mis
sion
Fig. 1. Hypothesized response of isoprenoid emission under increasing dose of O3
following the hormetic dose–response relationship as described by Calabrese (2005).The most common form of the hormetic dose–response curve depicting low-dosestimulatory and high-dose inhibitory response seems the best model to predict sucha response according to the experiments carried out so far.
4. The antioxidant role of isoprenoids in plants
Early studies based on knowledge of isoprenoid reaction in thegas phase in the atmosphere (see above) led to the suggestion thatvolatile isoprenoids synthesized by the leaf may produce hydro-peroxides when reacting with O3 inside the leaf mesophyll, there-fore aggravating O3 damage to leaf (Hewitt et al., 1990). However,other studies observed that isoprenoid emission is stimulated inplants that have been exposed to stress conditions, namely drought(Sharkey and Loreto, 1993) and O3 (Loreto et al., 2004, see alsoabove). Why are isoprenoids emitted by plants, resulting in a loss ofa considerable fraction of the fixed carbon, especially under stressconditions when carbon acquisition is limited? If isoprenoidsnegatively affect plant capacity to cope with environmentalstresses, this teleological question remains unanswered.
Sharkey and Singsaas (1995) observed that the photosyntheticmachinery of isoprene-emitting leaves was more resistant to hightemperature than in non-emitting leaves. These authors putforward the idea that isoprene production could indeed protectleaves against high temperature. This theory was confirmed bystudies carried out in Italy which showed a similar thermo-protective effect also in monoterpene-emitting plants (Loreto et al.,1998a) or when exogenous monoterpenes are fumigated to low-emitters (Delfine et al., 2000). However, studies in the Italianlaboratories also attributed to isoprene a more general antioxidantrole after observing improved protection from O3 damage inisoprene-emitting leaves (Loreto and Velikova, 2001) or in leavesexposed to exogenous isoprene during O3 treatment (Loreto et al.,2001). These experiments clearly demonstrated that reactiveoxygen species (H2O2, singlet oxygen) are reduced, and membranesare better preserved in isoprene-emitting leaves than in non-emitting leaves, after an O3 treatment (Loreto and Velikova, 2001;Velikova et al., 2004). Consequently, typical O3 injuries are reducedor absent altogether in isoprene-emitting leaves (Fig. 2). Sharkeyand Yeh (2001) raised the hypothesis that the lipophilic isoprenoidsinteract with lipids in membranes, reducing membrane denatur-ation caused by stresses. Theoretical confirmation of this action hasbeen recently obtained by a study showing that isoprene enhancesthe packing of lipid tails of phospholipid bilayers of cellularmembranes. It was suggested that isoprene molecules partitioninto the free volume at the center of the bilayer might strengthenthe lipid tails and avoid damage to membranes (Siwko et al., 2007).This action may indeed usefully defend leaf structure against bothhigh temperature and O3 stress.
Isoprenoids may also scavenge pollutants in the intercellularspaces reducing the potential level of reactive oxygen species thatimpinge on membranes causing their oxidation (Loreto et al.,2001). Whether this occurs by direct reaction or indirectly is notknown. Isoprene reactivity with O3 appears to be too low for thisreaction to occur at significant rates in nature (Atkinson, 1997), butthis is not true for more reactive isoprenoids such as monoterpenesand sesquiterpenes (Fares et al., 2008). Moreover, isoprenoid–O3
interaction has been studied only in gas phase, atmosphericprocesses. Whether the same knowledge may be extended to theleaf mesophyll, that is, in a basically liquid environment whereisoprenoid concentrations are at least 100 times higher than thosemeasured in the atmosphere (Loreto et al., 1998b), remains to bedetermined. Certainly isoprenoids may rapidly react with somereactive oxygen species that are formed by O3 transformationinside leaves. Hydrogen peroxide formed by O3 is highly quenchedinside leaves of isoprene-emitting leaves (Loreto and Velikova,2001), and a direct quenching of singlet oxygen in the presence ofisoprene has been reported in laboratory studies (Affek and Yakir,2004; Velikova et al., 2004).
Finally, Velikova et al. (2005) found that, in isoprene-emittingleaves that were exposed to O3, the accumulation of both nitric

Fig. 2. Effect of O3 exposure on Arabidopsis wild-type (Ws) and ISPS (isoprene synthase) overexpressing transgenic (IspS) plants (Sharkey et al., 2005). Plants were fumigated with300 ppb of O3 for 8 h, using the Arabidopsis gas-exchange cuvette (Tholl et al., 2006). (Picture courtesy of A. Fortunati.)
C. Calfapietra et al. / Environmental Pollution 157 (2009) 1478–14861482
oxide (NO) and H2O2 in the mesophyll was highly quenched. Thereduction of NO formation was surprising since isoprene is notbelieved to react rapidly with this relatively unreactive chemicalspecies that is formed inside stressed leaves by amino acid degra-dation. However, in very oxidative environments NO may produceperoxynitrites that are toxic and very reactive compounds (Delle-donne et al., 2002), which could react with isoprenoids (Velikovaet al., 2005). Whatever the mechanism that leads to NO removal inisoprene-emitting leaves that are exposed to O3, this effect hasimportant physiological implications. Nitric oxide and H2O2 areamong the signalling molecules that start the sequence ofbiochemical events leading to programmed cell death (Beligni et al.,2002). If both molecules are quenched by isoprene, then the die-outmessage is reduced. This might also explain the reduced cellulardamage observed in portions of leaves emitting isoprene or fumi-gated with exogenous isoprene (Loreto et al., 2001). Volatiles are bydefinition molecules that freely diffuse and can serve as messen-gers. Isoprenoids might also serve a similar role, modulatingmolecular signalling that is operated by other volatiles in leaves.
5. Modelling O3 flux when accounting for isoprenoidsdetoxification effect
As we have just summarized, several experiments support thehypothesis that isoprenoids react with reactive oxygen species inthe intercellular spaces, reducing the oxidative damage producedby O3. If this hypothesis is correct, then, first, O3 must enter theleaves before reacting with isoprenoids; second, isoprenoids are
also responsible for O3 concentration drawdown from the externalatmosphere to the mesophyll, where O3 concentration is very lowand can be treated as being zero (Laisk et al., 1989). As shown inFig. 3 the modelling of O3 flux should take into account both thestomatal and non-stomatal components. The O3 flux can be directlymeasured through cuvette systems as described in Fares et al.(2008) and Grulke et al. (2007) measuring O3 concentration at theinlet (reference) and at the outlet (sample) of a cuvette, andderiving flux by multiplying the difference in O3 concentrationbetween reference and sample for the molar flow rate andnormalizing for the leaf area enclosed in the cuvette.
Fig. 4 shows a curvilinear relationship between O3 flux, ascalculated from leaf cuvette measurements, and stomatal conduc-tance to water vapour (Gs) for isoprenoid (isoprene or mono-terpenes) emitting or non-emitting plant species. Three interestingobservations can be inferred from inspection of the relationshipsshown in this figure.
First, both relationships were best fitted by second order curves,which suggest that when stomatal conductance is very higha saturation of the scavenging capacity of the pollutant insideleaves is reached, and O3 starts to accumulate inside the mesophyll.This is of course at odds with the general assumption that O3
immediately reacts after entering stomata (Laisk et al., 1989) infavour of the idea that ozone may accumulate in the intercellularspaces (Moldau and Bichele, 2002). However, a saturation of ozonein the intercellular spaces should not occur in Mediterraneanregions when drought stress periods (always in coincidence withhigh atmospheric ozone concentrations) induce stomatal closure.
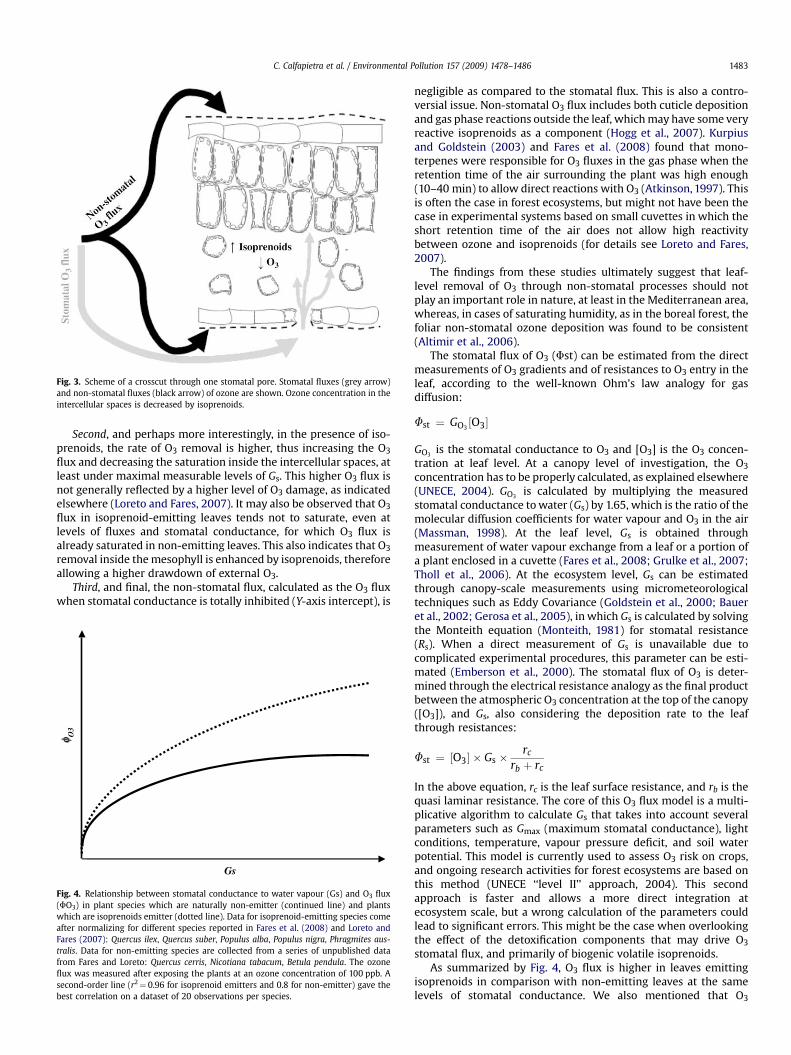
Fig. 3. Scheme of a crosscut through one stomatal pore. Stomatal fluxes (grey arrow)and non-stomatal fluxes (black arrow) of ozone are shown. Ozone concentration in theintercellular spaces is decreased by isoprenoids.
C. Calfapietra et al. / Environmental Pollution 157 (2009) 1478–1486 1483
Second, and perhaps more interestingly, in the presence of iso-prenoids, the rate of O3 removal is higher, thus increasing the O3
flux and decreasing the saturation inside the intercellular spaces, atleast under maximal measurable levels of Gs. This higher O3 flux isnot generally reflected by a higher level of O3 damage, as indicatedelsewhere (Loreto and Fares, 2007). It may also be observed that O3
flux in isoprenoid-emitting leaves tends not to saturate, even atlevels of fluxes and stomatal conductance, for which O3 flux isalready saturated in non-emitting leaves. This also indicates that O3
removal inside the mesophyll is enhanced by isoprenoids, thereforeallowing a higher drawdown of external O3.
Third, and final, the non-stomatal flux, calculated as the O3 fluxwhen stomatal conductance is totally inhibited (Y-axis intercept), is
Gs
O3
Fig. 4. Relationship between stomatal conductance to water vapour (Gs) and O3 flux(FO3) in plant species which are naturally non-emitter (continued line) and plantswhich are isoprenoids emitter (dotted line). Data for isoprenoid-emitting species comeafter normalizing for different species reported in Fares et al. (2008) and Loreto andFares (2007): Quercus ilex, Quercus suber, Populus alba, Populus nigra, Phragmites aus-tralis. Data for non-emitting species are collected from a series of unpublished datafrom Fares and Loreto: Quercus cerris, Nicotiana tabacum, Betula pendula. The ozoneflux was measured after exposing the plants at an ozone concentration of 100 ppb. Asecond-order line (r2¼ 0.96 for isoprenoid emitters and 0.8 for non-emitter) gave thebest correlation on a dataset of 20 observations per species.
negligible as compared to the stomatal flux. This is also a contro-versial issue. Non-stomatal O3 flux includes both cuticle depositionand gas phase reactions outside the leaf, which may have some veryreactive isoprenoids as a component (Hogg et al., 2007). Kurpiusand Goldstein (2003) and Fares et al. (2008) found that mono-terpenes were responsible for O3 fluxes in the gas phase when theretention time of the air surrounding the plant was high enough(10–40 min) to allow direct reactions with O3 (Atkinson, 1997). Thisis often the case in forest ecosystems, but might not have been thecase in experimental systems based on small cuvettes in which theshort retention time of the air does not allow high reactivitybetween ozone and isoprenoids (for details see Loreto and Fares,2007).
The findings from these studies ultimately suggest that leaf-level removal of O3 through non-stomatal processes should notplay an important role in nature, at least in the Mediterranean area,whereas, in cases of saturating humidity, as in the boreal forest, thefoliar non-stomatal ozone deposition was found to be consistent(Altimir et al., 2006).
The stomatal flux of O3 (Fst) can be estimated from the directmeasurements of O3 gradients and of resistances to O3 entry in theleaf, according to the well-known Ohm’s law analogy for gasdiffusion:
Fst ¼ GO3½O3�
GO3is the stomatal conductance to O3 and [O3] is the O3 concen-
tration at leaf level. At a canopy level of investigation, the O3
concentration has to be properly calculated, as explained elsewhere(UNECE, 2004). GO3
is calculated by multiplying the measuredstomatal conductance to water (Gs) by 1.65, which is the ratio of themolecular diffusion coefficients for water vapour and O3 in the air(Massman, 1998). At the leaf level, Gs is obtained throughmeasurement of water vapour exchange from a leaf or a portion ofa plant enclosed in a cuvette (Fares et al., 2008; Grulke et al., 2007;Tholl et al., 2006). At the ecosystem level, Gs can be estimatedthrough canopy-scale measurements using micrometeorologicaltechniques such as Eddy Covariance (Goldstein et al., 2000; Baueret al., 2002; Gerosa et al., 2005), in which Gs is calculated by solvingthe Monteith equation (Monteith, 1981) for stomatal resistance(Rs). When a direct measurement of Gs is unavailable due tocomplicated experimental procedures, this parameter can be esti-mated (Emberson et al., 2000). The stomatal flux of O3 is deter-mined through the electrical resistance analogy as the final productbetween the atmospheric O3 concentration at the top of the canopy([O3]), and Gs, also considering the deposition rate to the leafthrough resistances:
Fst ¼ ½O3� � Gs �rc
rb þ rc
In the above equation, rc is the leaf surface resistance, and rb is thequasi laminar resistance. The core of this O3 flux model is a multi-plicative algorithm to calculate Gs that takes into account severalparameters such as Gmax (maximum stomatal conductance), lightconditions, temperature, vapour pressure deficit, and soil waterpotential. This model is currently used to assess O3 risk on crops,and ongoing research activities for forest ecosystems are based onthis method (UNECE ‘‘level II’’ approach, 2004). This secondapproach is faster and allows a more direct integration atecosystem scale, but a wrong calculation of the parameters couldlead to significant errors. This might be the case when overlookingthe effect of the detoxification components that may drive O3
stomatal flux, and primarily of biogenic volatile isoprenoids.As summarized by Fig. 4, O3 flux is higher in leaves emitting
isoprenoids in comparison with non-emitting leaves at the samelevels of stomatal conductance. We also mentioned that O3

C. Calfapietra et al. / Environmental Pollution 157 (2009) 1478–14861484
detoxification by isoprenoids led to a reduction of leaf damage. Forthis reason, when assessing the effective dose of ozone (the dose(stomatal flux� time) that causes plant injury; Massman, 2004),we should consider the role of isoprenoids. If the aim of the modelis to associate the stomatal flux of O3 to the O3 damage, a furtherresistance represented by the isoprenoids contribution to ozoneremoval should be applied in the resistance analogies shown above.As a consequence, the stomatal fluxes of ozone would be decreasedof a factor proportional to the amount of the isoprenoids emitted.This factor could also be included in flux-based metrics whichemploy the concept of effective dose (Massman, 2004), defined asthe difference between the canopy stomatal ozone flux rate anda plant defense function, which is taken to be proportional to thecanopy gross photosynthesis.
Inserting a factor in a model that takes into account the role ofisoprenoids requires a good knowledge of (a) the biochemistry andphysiology of isoprenoid formation and emission; (b) the chemistryof isoprenoid reactivity with atmospheric compounds; and (c) theecology of the ecosystem under investigation. Unfortunately, noneof these is currently refined enough to allow such a parameteriza-tion. However, some basic principles could be drafted that mayassist in rudimentary attempts of parameterization and modellingisoprenoid–O3 interactions.
(a) Isoprenoid emission is not related to stomatal conductance,because the gradient generally builds up within intercellularspaces and the atmosphere, simultaneously to stomatal closure(Loreto and Sharkey, 1990). This, contrary to what we have justelucidated about O3 modelling, makes modelling of isoprenoidemission through stomatal conductance inefficient. However,isoprenoid emission is strongly dependent on substrate avail-ability and enzymatic properties, which are reflected insignificant and well-established light and temperaturedependencies of the emission rates (Niinemets et al., 2004).The algorithm proposed by Guenther et al. (1995) exploitsthese empirical dependencies from environmental factors andcorrects a standard (or basal) emission rate (SE) measured instandard conditions (T¼ 30 �C and PPFD (PhotosyntheticPhoton Flux Density)¼ 1000 mmol m�2 s�1) by light- andtemperature-dependent correction factors. A simplified formof the equation is reported in the following equation:
Eisoprenoids¼D�SE
"aCLPPFDffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffip
#�
24 exp
�CT1ðT�TSÞ
RTST
�� �
35
1þa2PPFD2 1þexp CT2ðT�TMÞRTST
where CL, a, CT1, CT2, and TM are empirical coefficients, R is theuniversal gas constant, D is the foliar density, T is the leaftemperature and TS is the leaf temperature at standard condi-tions (303 K). A limitation to the use of this model is that thestandard emission rate is species-specific, and the variabilityaround such estimate represents a large element of uncertainty.
(b) Isoprenoid concentration inside leaves is by far higher than theconcentration of the compound in the atmospheresurrounding the leaf. The volatility, the solubility, and thecoefficient of partition in liquids of the different isoprenoidsthat may be metabolically synthesized are extremely different.The concentration of isoprenoid inside leaves can be measuredor calculated (Loreto et al., 1998b; Niinemets et al., 2004).
(c) Due to the different reactivity, and oxidation rate, differentisoprenoids may react in the gas phase with different timecourses and at different rates with O3 and reactive oxygenspecies formed after O3 acquisition by leaves through stomata.Monoterpenes and sesquiterpenes are well-known to reactwith O3 faster than isoprene (Atkinson, 1997).
In theory, a correction factor should therefore be the result of atleast three components, the first describing the emission, thesecond assessing the gaseous concentration in the mesophyll andthe third assessing the capacity of reaction of isoprenoids present inthe mesophyll with O3.
6. Interactions of isoprenoid and O3 with further abiotic andbiotic factors
We know that isoprenoids emitted by plants are crucialcomponents of plant mechanisms of protection against abiotic andbiotic stresses (Gershenzon and Dudareva, 2007) but at the sametime, abiotic and biotic stresses influence considerably the iso-prenoid emission from vegetation. This may in turn indirectlychange the flux of O3 into vegetation, and the capacity of plants toresist oxidative stresses.
Warming is the climate change factor (IPCC, 2007) that willprobably have most impact on isoprenoid emission in the future, astheir emission is strongly and positively dependent on temperature(Guenther et al., 1995). As the increase in temperature is alsoexpected to further increase O3 formation in the troposphere,a dramatic positive feedback of temperature on O3 production isexpected. On the contrary the increase in CO2 concentration in theatmosphere (IPCC, 2007) is expected to decrease the basal emissionof isoprenoids although to a different extent according to thespecies and the actual level of CO2 that will be experienced (Loretoet al., 2001; Rosenstiel et al., 2003; Possell et al., 2004; Calfapietraet al., 2008). The combination of elevated concentrations of CO2 andO3 resulted in a strong decrease of isoprene emission in open fieldconditions (Calfapietra et al., 2008). However, when the stimulativeeffect on the leaf area index by elevated CO2 is taken into account,the global emission of isoprenoids at ecosystem level is expected toremain unaltered for most of ecosystems (Arneth et al., 2008).Unfortunately no models of isoprenoid emission have investigatedthe combination of the different factors of global change, includingO3, so far.
The interactions between isoprenoids and O3 also involvingbiotic stresses have been investigated in few cases (Vuorinen et al.,2004; Pinto et al., 2007). These interactions are extremely complexbecause global change can alter the already complicated tritrophiccommunication involving plants, herbivores, and carnivorouspredators or parasitoids. In many cases it has been proved thatisoprenoids can act as signalling compounds attracting parasitoidsor predators after herbivore attack to the plant (Gershenzon andDudareva, 2007). Herbivore and mechanical wounding to theleaves may stimulate emission of constitutive isoprenoids or inducede novo synthesis of isoprenoids (Loreto et al., 2006). Ozone canhave a crucial role on this interaction by damaging the leaf cuticles(Paoletti et al., 2007) and favouring the attacks by pests and insects,which might induce an increase in isoprenoid emission andenhance at the same time the non-stomatal uptake of the pollutant.Inside leaves, on the other hand, isoprenoids can drive a higherstomatal flux of O3 while reducing the overall damage suffered. Theresult of this intricate web of feedbacks and feed forwards is diffi-cult to untangle and more experiments and new detailed modelsare needed in order to provide more accurate information on theseecologically important interactions.
7. Final remarks
For a thorough comprehension of the central role for the forestecosystems in the global biosphere–atmosphere interactions theunderstanding of roles and actions of biogenic volatile organiccompounds seems more and more essential. Not only do thesecompounds play vital roles in plant defense against abiotic andbiotic stresses, they also interact with chemical processes in the

C. Calfapietra et al. / Environmental Pollution 157 (2009) 1478–1486 1485
atmosphere. The interaction with tropospheric O3 is multiple and,despite the scientific progress of the last decade, largely elusive.
New forest plantations are established every year at plantingrates of 8.5–10.5 Mha year�1, leading to an annual net gain of1.96 Mha year�1 between 1965 and 1990 (FAO, 2005). In particular,short rotation forest (SRF) plantations are gaining importance forbiomass production substituting for fossil fuel and thereforereducing GHG emissions. Isoprenoid emission is particularly rele-vant for poplar and eucalyptus species, which are used worldwidein SRF plantations. A negative feedback might derive from thisland-use change, as plantations may massively increase isoprenoidemissions. While this might have serious repercussions on theatmospheric composition of peri-urban areas characterized byheavy anthropogenic pollution, the impact on plant protectionagainst O3 and other stressors is unclear, due to the above consid-erations. Genetic engineering today allows to grow non-emittingpoplar trees (Miller et al., 2001), which might have suitable usewhenever natural sources of O3 need to be controlled. On the otherhand, such plant material might be less resistant to the environ-ment, especially when coping with high oxidative stresses.
Acknowledgments
This work was supported in part by the European ScienceFoundation Grant ‘‘VOCBAS’’, by the European Commission inMarie-Curie Research Training Network ‘‘ISONET’’ (contract MC-RTN-CT-2003-504720) and by the Marie Curie InternationalFellowship within the 6th European Community Framework Pro-gramme (‘‘GLOBALVOC’’, contract MOIF-CT-2005-007692).
The authors thank Paolo Ciccioli and Tom Sharkey for stimu-lating discussions and advice.
References
Affek, H.P., Yakir, D., 2004. Protection by isoprene against singlet oxygen in leaves.Plant Physiology 129, 269–277.
Altimir, N., Kolari, P., Tuovinen, J.-P., Vesala, T., Back, J., Suni, T., Kulmala, M., Hari, P.,2006. Foliage surface ozone deposition: a role for surface moisture? Bio-geosciences 3, 209–228.
Andreae, M.O., Crutzen, P.J., 1997. Atmospheric aerosols: biogeochemical sourcesand role in atmospheric chemistry. Science 276, 1052–1058.
Arneth, A., Schurgers, G., Hickler, T., Miller, P.A., 2008. Effects of species composi-tion, land surface cover, CO2 concentration and climate on isoprene emissionsfrom European forests. Plant Biology 10, 150–162.
Atkinson, R., 1997. Gas-phase tropospheric chemistry of volatile organiccompounds: 1, alkanes and alkenes. Journal of Physical Chemistry 26, 215–290.
Atkinson, R., 2000. Atmospheric chemistry of VOCs and NOx. Atmospheric Envi-ronment 34, 2063–2101.
Bauer, M.R., Hultman, N.E., Panek, J.A., Goldstein, A.H., 2002. Ozone deposition toa ponderosa pine plantation in the Sierra Nevada Mountains (CA): a comparisonof two different climatic years. Journal of Geophysical Research 105, 123–136.
Beligni, M.V., Fath, A., Bethke, P.C., Lamattina, L., Jones, R.L., 2002. Nitric oxide acts asan antioxidant and delays programmed cell death in barley aleurone layers.Plant Physiology 129, 1642–1650.
(Special Issue) BEMA, 1997. A European commission project on biogenic emission inthe Mediterranean area. Atmospheric Environment 31, 1–255.
Boland, W., Gabler, A., 1989. Biosynthesis of homoterpenes in higher plants. Hel-vetica Chimica Acta 72, 247–253.
Bonn, B., Moortgat, G.K., 2003. Sesquiterpene ozonolysis: origin of atmospheric newparticle formation from biogenic hydrocarbons. Geophysical Research Letters30, 1585.
Brilli, F., Barta, C., Fortunati, A., Lerdau, M., Loreto, F., Centritto, M., 2007. Response ofisoprene emission and carbon metabolism to drought in white poplar (Populusalba) saplings. New Phytologist 175, 244–254.
Calabrese, E.J., 2005. Paradigm lost, paradigm found: the re-emergence of hormesisas a fundamental dose response model in the toxicological sciences. Environ-mental Pollution 138, 378–411.
Calfapietra, C., Wiberley, A.E., Falbel, T.G., Linskey, A.R., Scarascia Mugnozza, G.,Karnosky, D.F., Loreto, F., Sharkey, T.D., 2007. Isoprene synthase expression andprotein levels are reduced under elevated O3 but not under elevated CO2 (FACE)in field-grown aspen trees. Plant Cell & Environment 30, 654–661.
Calfapietra, C., Scarascia Mugnozza, G., Karnosky, D.F., Loreto, F., Sharkey, T.D., 2008.Isoprene emission rates under elevated CO2 and O3 in two field-grown aspenclones differing for their sensitivity to O3. New Phytologist 179, 55–61.
Carter, W.P.L., 1994. Development of ozone reactivity scales for volatile organiccompounds. Journal of the Air & Waste Management Association 44, 881–899.
Ciccioli, P., Brancaleoni, E., Frattoni, M., Di Palo, V., Valentini, R., Tirone, G.P.,Seufert, G., Bertin, N., Hansen, U., Csiky, O., Lenz, R., Sharma, M., 1999. Emissionof reactive terpene compounds from orange orchards and their removal bywithin-canopy processes. Journal of Geophysical Research 104, 8077–8094.
Copolovici, L.O., Niinemets, U., 2005. Temperature dependencies of Henry’s lawconstants and octanol/water partition coefficients for key plant volatile mon-oterpenoids. Chemosphere 61, 1390–1400.
Degenhardt, J., Gershenzon, J., 2000. Demonstration and characterization of (E)-nerolidol synthase from maize: a herbivore-inducible terpene synthaseparticipating in (3E)-4,8-dimethyl-1,3,7-nonatriene biosynthesis. Planta 210,815–822.
Delfine, S., Csiky, O., Seufert, G., Loreto, F., 2000. Fumigation with exogenousmonoterpene of a non-isoprenoid- emitting oak (Quercus suber): monoterpeneacquisition, translocation, and effect on the photosynthetic properties at hightemperatures. New Phytologist 146, 27–36.
Delledonne, M., Murgia, I., Ederle, D., Sbicego, P.F., Biondani, A., Polverari, A.,Lamb, C., 2002. Reactive oxygen intermediates modulate nitric oxide signalingin the plant hypersensitive disease-resistance response. Plant Physiology andBiochemistry 40, 605–610.
Derwent, R.G., Jenkin, M.E., Saunders, S.M., 1996. Photochemical ozone creationpotentials for a large number of reactive hydrocarbons under Europeanconditions. Atmospheric Environment 30, 181–199.
Di Carlo, P., Brune, W.H., Martinez, M., Harder, H., Lesher, R., Ren, X., Thornberry, T.,Carroll, M.A., Young, V., Shepson, P.B., Riemer, D., Apel, D.E., Campbell, C., 2004.Missing OH reactivity in a forest: evidence for unknown reactive biogenic VOCs.Science 304, 722–725.
Dindorf, D., Kuhn, U., Ganzeveld, L., Schebeske, G., Ciccioli, P., Holzke, C., Koble, R.,Seufert, G., Kesselmeier, J., 2006. Significant light and temperature dependentmonoterpene emissions from European beech (Fagus sylvatica L.) and theirpotential impact on the European volatile organic compound budget. Journal ofGeophysical Research 111, D16305. 10.1029/2005JD006751.
Emberson, L.D., Ashmore, M.R., Cambridge, H.M., Simpson, D., Tuovinen, J., 2000.Modelling stomatal ozone flux across Europe. Environmental Pollution 109,403–413.
FAO, 2005. Report of the 22nd Session of the International Poplar Commission and42nd Session of its Executive Committee, Santiago, Chile, 28 November–2December 2004.
Fares, S., Barta, C., Ederli, L., Ferranti, F., Pasqualini, S., Reale, L., Brilli, F., Tricoli, D.,Loreto, F., 2006. Impact of high ozone on isoprene emission and some anatomicaland physiological parameters of developing Populus alba leaves directly orindirectly exposed to the pollutant. Physiologia Plantarum 128, 456–465.
Fares, S., Loreto, F., Kleist, E., Wildt, J., 2008. Stomatal uptake and stomatal depo-sition of ozone in isoprene and monoterpene emitting plants. Plant Biology 10,44–54.
Fehsenfeld, F., Calvert, J., Fall, R., Goldan, P., Guenther, A.B., Hewitt, N.C., Lamb, B.,Liu, S., Trainer, M., Westberg, H., Zimmerman, P., 1992. Emissions of volatileorganic compounds from vegetation and the implications for atmosphericchemistry. Global Biogeochemical Cycles 6, 390–430.
Fuentes, J.D., Lerdau, M., Atkinson, R., Baldocchi, D., Bottenheim, J.W., Ciccioli, P.,Lamb, B., Geron, C., Gu, L., Guenther, A., Sharkey, T.D., Stockwell, W., 2000.Biogenic hydrocarbons in the atmospheric boundary layer: a review. Bulletin ofthe American Meteorological Society 81, 1537–1575.
Funk, J.L., Mak, J.E., Lerdau, M.T., 2004. Stress-induced changes in carbon sources forisoprene production in Populus deltoides. Plant Cell and Environment 27,747–755.
Gerosa, G., Vitale, M., Finco, A., Manes, F., Ballarin Denti, A., Cieslik, S., 2005. Ozoneuptake by an evergreen Mediterranean Forest (Quercus ilex). Part I: microme-teorological flux measurements and flux partitioning. Atmospheric Environ-ment 39, 3255–3266.
Gershenzon, J., Dudareva, N., 2007. The function of terpene natural products in thenatural world. Nature Chemical Biology 3, 408–414.
Grulke, N.E., Paoletti, E., Heath, R.L., 2007. Comparison of calculated and measuredfoliar O3 flux in crop and forest species. Environmental Pollution 146, 640–647.
Goldstein, A.H., Hultman, N.E., Fracheboud, J.M., Bauer, M.R., Panek, J.A., Xu, M.,Qi, Y., Guenther, A.B., Baugh, W., 2000. Effects of climate variability on thecarbon dioxide, water, and sensible heat fluxes above a ponderosa pine plan-tation in the Sierra Nevada (CA). Agricultural and Forest Meteorology 101,113–129.
Guenther, A.B., Monson, R.K., Fall, R., 1991. Isoprene and monoterpene emission ratevariability: observations with eucalyptus and emission rate algorithm devel-opment. Journal of Geophysical Research 96, 799–808.
Guenther, A., Zimmerman, P., Wildermuth, M., 1994. Natural volatile organic-compound emission rate estimates for united-states woodland landscapes.Atmospheric Environment 28, 1197–1210.
Guenther, A., Hewitt, C.N., Erickson, D., Fall, R., Geron, C., Graedel, T., Harley, P.,Klinger, L., Lerdau, M., McKay, W.A., Pierce, T., Scholes, B., Steinbrecher, R.,Tallamraju, R., Taylor, J., Zimmerman, P., 1995. A global model of naturalvolatile organic compounds emissions. Journal of Geophysical Research 100,8873–8892.
Hewitt, C.N., Kok, G.L., Fall, R., 1990. Hydroperoxides in plants exposed to ozonemediate air pollution damage to alkene emitters. Nature 344, 56–58.
Hogg, A., Uddling, J., Ellsworth, D., Carroll, M.A., Pressley, S., Lamb, B., Vogel, C., 2007.Stomatal and non-stomatal fluxes of ozone to a northern mixed hardwoodforest. Tellus 59B, 514–525.

C. Calfapietra et al. / Environmental Pollution 157 (2009) 1478–14861486
INFC, 2007. I caratteri quantitativi 2005-parte I. In: Gasparini, P., De Natale, F., DiCosmo, L., Gagliano, C., Salvadori, I., Tabacchi, G., Tosi, V. (Eds.), InventarioNazionale Delle Foreste E Dei Serbatoi Forestali Di Carbonio. MIPAAF, Ispet-torato Generale Corpo Forestale dello Stato, CRA-MPF, Trento.
IPCC, 2007. Intergovernmental panel on climate change fourth assessment reportclimate change 2007: synthesis report. http://www.ipcc.ch/ipccreports/.
Karnosky, D.F., Zak, D.R., Pregitzer, K.S., Awmack, C.S., Bockheim, J.G., Dickson, R.E.,Hendrey, G.R., Host, G.E., King, J.S., Kopper, B.J., Kruger, E.L., Kubiske, M.E.,Lindroth, R.L., Mattson, W.J., McDonald, E.P., Noormets, A., Oksanen, E.,Parsons, W.F.J., Percy, K.E., Podila, G.K., Riemenschneider, D.E., Sharma, P.,Thakur, R.C., Sober, A., Sober, J., Jones, W.S., Anttonen, S., Vapaavuori, E.,Mankovska, B., Heilman, W.E., Isebrands, J.G., 2003. Tropospheric O3 moderatesresponses of temperate hardwood forests to elevated CO2: a synthesis ofmolecular to ecosystem results from the Aspen FACE project. FunctionalEcology 17, 289–304.
Kesselmeier, J., Staudt, M., 1999. Biogenic volatile organic compounds (VOC): anoverview on emission, physiology and ecology. Journal of Atmospheric Chem-istry 33, 23–88.
Kurpius, M.R., Goldstein, A., 2003. Gas-phase chemistry dominates O3 loss toa forest, implying a source of aerosols and hydroxyl radicals to the atmosphere,pp. 24–1. Geophysical Research Letters 30, 1371, doi:10.1029/2002GL016785.
Laisk, A., Kull, O., Moldau, H., 1989. Ozone concentration in leaf intercellular airspaces is close to zero. Plant Physiology 90, 1163–1167.
Lenz, R., Koble, R., Seufert, G., 2001. Species-based mapping of biogenic emissions inEurope: a case study Italy. In: Proceedings of the Eighth of the EuropeanSymposium of the Physico-chemical Behaviour of the Photochemical Pollutants,17–20 September 2001, Turin, Italy.
Lerdau, M., 2007. A positive feedback with negative consequences. Science 316,212–213.
Loreto, F., Sharkey, T.D., 1990. A gas exchange study of photosynthesis and isopreneemission in red oak (Quercus rubra L.). Planta 182, 523–531.
Loreto, F., Forster, A., Durr, M., Csiky, O., Seufert, G., 1998a. On the monoterpeneemission under heat stress and on the increased thermotolerance of leaves ofQuercus ilex fumigated with selected monoterpenes. Plant Cell & Environment21, 101–107.
Loreto, F., Ciccioli, P., Brancaleoni, E., Cecinato, A., Frattoni, M., 1998b. Measurementof isoprenoid content in leaves of Mediterranean Quercus spp. by a novel andsensitive method and estimation of the isoprenoid partition between liquid andgas phase inside the leaves. Plant Science 136, 25–30.
Loreto, F., Velikova, V., 2001. Isoprene produced by leaves protects the photosyn-thetic apparatus against ozone damage, quenches ozone products, and reduceslipid peroxidation of cellular membranes. Plant Physiology 127, 1781–1787.
Loreto, F., Mannozzi, M., Maris, C., Nascetti, P., Ferranti, F., Pasqualini, S., 2001. Ozonequenching properties of isoprene and its antioxidant role in leaves. PlantPhysiology 126, 993–1000.
Loreto, F., 2002. Distribution of isoprenoid emitters in the Quercus genus around theworld: chemo-taxonomical implications and evolutionary considerations basedon the ecological function of the trait. Perspectives in Plant Ecology Evolutionand Systematics 5, 185–192.
Loreto, F., Pinelli, P., Manes, F., Kollist, H., 2004. Impact of ozone on monoterpeneemission and evidence for an isoprene-like antioxidant action of monoterpensemitted by Quercus ilex leaves. Tree Physiology 24, 361–367.
Loreto, F., Barta, C., Brilli, F., Nogues, I., 2006. On the induction of volatile organiccompound emissions by plants as consequence of wounding or fluctuations oflight and temperature. Plant, Cell and Environment 29, 1820–1828.
Loreto, F., Fares, S., 2007. Is ozone flux inside leaves only a damage indicator? Cluesfrom volatile isoprenoid studies. Plant Physiology 143, 1096–1100.
Massman, W.J., 1998. A review of the molecular diffusivities of H2O, CO2, CH4, CO,O3, SO2, NH3, N2O, NO and NO2 in air, O2 and N2 near STP. Atmospheric Envi-ronment 32, 1111–1127.
Massman, W.J., 2004. Toward an ozone standard to protect vegetation based oneffective dose: a review of deposition resistances and a possible metric.Atmospheric Environment 38, 2323–2337.
Millan, M.M., Salvador, R., Mantilla, E., Kallos, G., 1997. Photo-oxidation dynamics inthe Western Mediterranean in summer: results from European ResearchProjects. Journal of Geophysical Research 102, 8811–8823.
Miller, B., Oschinsky, C., Zimmer, W., 2001. First isolation of an isoprene synthasegene from poplar and successful expression of the gene in Escherichia coli.Planta 213, 483–487.
Moldau, H., Bichele, I., 2002. Plasmalemma protection by the apoplast as assessedfrom above-zero ozone concentration in leaf intercellular air spaces. Planta 214,484–487.
Monson, R.K., Jaeger, C.H., Adams, W.W., Driggers, E.M., Silver, G.M., Fall, R., 1992.Relationships among isoprene emission rate, photosynthesis, and isoprenesynthase activity as influenced by temperature. Plant Physiology 98, 1175–1180.
Monteith, J.L., 1981. Coupling of plants to the atmosphere. In: Grace, J., Jarvis, P.G.,Ford, E.D. (Eds.), Plants and Their Atmospheric Environment. Blackwell, Oxford,pp. 1–29.
Niinemets, U., Loreto, F., Reichstein, M., 2004. Physiological and physicochemicalcontrols on foliar volatile organic compound emissions. Trends in Plant Science9, 180–186.
Owen, S., Boissard, C., Street, R.A., Duckham, S.C., Csiky, O., Hewitt, C.N., 1997.Screening of 18 Mediterranean plant species for volatile organic compoundemissions. Atmospheric Environment 31, 101–117.
Paoletti, E., Anselmi, N., Franceschini, A., 2007. Pre-exposure to ozone predisposesoak leaves to attacks by Diplodia corticola and Biscogniauxia mediterranea. TheScientific World Journal 7 (Suppl. 1), 222–230.
Papale, D., 2006. Il Progetto CARBOITALY: una rete nazionale per la misura dei sinkforestali e agricoli italiani e lo sviluppo di un sistema di previsione dell’assor-bimento dei gas serra. Forest@ 3 (2), 165–167.
Pinelli, P., Tricoli, D., 1989. A new approach to ozone plant fumigation: The Web-O3-Fumigation. Isoprene response to a gradient of ozone stress in leaves of Quercuspubescens. Forest 1 (2), 100–108.
Pinto, D.M., Blande, J.D., Riikka Nykanen, R., Dong, W.-X., Nerg, A.-M.,Holopainen, J.K., 2007. Ozone degrades common herbivore-induced plantvolatiles: does this affect herbivore prey location by predators and parasitoids?Journal of Chemical Ecology 33, 683–694.
Possell, M., Heath, J., Hewitt, C.N., Ayres, E., Kerstiens, G., 2004. Interactive effects ofelevated CO2 and soil fertility on isoprene emissions from Quercus robur. GlobalChange Biology 10, 1835–1843.
Rosenstiel, T.N., Potosnak, M.J., Griffin, K.L., Fall, R., Monson, R.K., 2003. IncreasedCO2 uncouples growth from isoprene emission in an agriforest ecosystem.Nature 421, 256–259.
Seufert, G., Bartzis, J., Bomboi, T., Ciccioli, P., Cieslik, S., Dlugi, R., Foster, P.,Hewitt, C.N., Kesselmeier, J., Kotzias, D., Lenz, R., Manes, F., Pastor, R.P.,Steinbrecher, R., Torres, L., Valentini, R., Versino, B., 1997. An overview of theCastelporziano experiments. Atmospheric Environment 31, 5–17.
Sharkey, T.D., Loreto, F., 1993. Water-stress, temperature, and light effects on thecapacity for isoprene emission and photosynthesis of kudzu leaves. Oecologia95, 328–333.
Sharkey, T.D., Singsaas, E.L., 1995. Why plants emit isoprene. Nature 374, 769.Sharkey, T.D., Yeh, S., 2001. Isoprene emission from plants. Annual Review of Plant
Physiology and Plant Molecular Biology 52, 407–436.Sharkey, T.D., Yeh, S., Wiberley, A.E., Falbel, T.G., Gong, D., Fernandez, D.E., 2005.
Evolution of the isoprene biosynthetic pathway in Kudzu. Plant Physiology 137,700–712.
Siwko, M.E., Marrink, S.J., de Vries, A.H., Kozubek, A., Schoot Uiterkamp, A.J.M.,Mark, A.E., 2007. Does isoprene protect plant membranes from thermal shock?A molecular dynamics study. Biochimica et Biophysica Acta – Biomembranes1768, 198–206.
Steinbrecher, R., 1997. Isoprene: production by plants and ecosystem-level esti-mates. In: .Helas, G., . Slanina, J., . Steinbrecher, R. (Eds.), Biogenic VolatileOrganic Compounds in the Atmosphere. SPB Academic Publishing BV,Amsterdam, pp. 101–114.
Steinbrecher, R., Hauff, K., Rabong, R., Steinbrecher, J., 1997. Isoprenoid emission ofoak species typical for the Mediterranean area source strength and controllingvariables. Atmospheric Environment 31, 79–88.
Steinbrecher, R., Smiatek, G., Koble, R., Seufert, G., Theloke, J., Hauff, K., Ciccioli, P.,Vautard, R. VOC emissions from Natural and Semi-Natural Vegetation for Europeand neighbouring countries in the East and South: Intra-Inter-Annual Vari-ability, Atmospheric Environment, in press doi:10.1016/j.atmosenv.2008.09.072.
Tholl, D., Boland, W., Hansel, A., Loreto, F., Rose, U.S.R., Schnitzler, J.P., 2006. Practicalapproaches to plant volatile analysis. The Plant Journal 45, 540–560.
UNECE, 2004. Revised manual on methodologies and criteria for mapping criticallevels/loads and geographical areas where they are exceeded. www.icpmapping.org (September, 2007).
Velikova, V., Edreva, A., Loreto, F., 2004. Endogenous isoprene protects Phragmitesaustralis leaves against singlet oxygen. Physiologia Plantarum 122, 219–225.
Velikova, V., Tsonev, T., Pinelli, P., Alessio, G.A., Loreto, F., 2005. Localized O3-fumigation for field studies of the impact of different ozone doses on photo-synthesis, respiration, electron transport rate and isoprene emission in Medi-terranean species. Tree Physiology 25, 1523–1532.
Vuorinen, T., Nerg, A.-M., Holopainen, J.K., 2004. Ozone exposure triggers theemission of herbivore-induced plant volatiles, but does not disturb tritrophicsignalling. Environmental Pollution 131, 305–311.
Went, F.W., 1960. Blue haze in the atmosphere. Nature (London) 187, 641–643.Wiedinmyer, C., Guenther, A., Harley, P., Hewitt, C.N., Geron, C., Artaxo, P.,
Steinbrecher, R., Rasmussen, R., 2004. Global organic emissions from vegetation.In: Granier, Claire, Artaxo, Paulo, Reeves, Claire E. (Eds.), Chapter in Emissions ofAtmospheric Trace Compounds. Kluwer Academic Publishers, Dordrecht, TheNetherlands, pp. 115–170.




















