
Immunodeficiency Virus Type 1 Infection Pathogenesis and ...

Downloaded from www.microbiologyresearch.org by
IP: 23.22.24.125
On: Sun, 14 Feb 2016 08:37:18
Journal of General Virology (1993), 74, 2595 260l. Printed in Great Britain 2595
Virus-free survival and down-regulation of CD4 in C8166 cells infected with human immunodeficiency virus type 1 at low density
A. Benedetto , t* A. R. Garbugl ia , ~ A. D i Caro , ~ E. L o Prest i , ~ E. Alfani z and C. Delfini z
1 Centre o f Virology, S. Camitto Hospital, U S L R M 10, Circonvallazione Gianicotense 85, 00152 Rome and 2 Laboralory o f Cell Biology, Istituto Superiore di Sanith, Viale Regina Elena 299, 00161 Rome, l ta ly
Compared with other T cell lines, C8166 lymphocytes are particularly susceptible to human immunodeficiency virus (HIV) infection and the outcome is invariably cell death. The results reported in this study demonstrate that the virus-induced cytolysis is strongly dependent on the initial cell density of C8166 cultures. Cultures diluted to 50 to 500 cells/ml almost completely main- tained their cell duplication rate and released infectious virus into the medium. HIV infection of diluted C8166 cells is a simple and easily reproducible procedure for
obtaining persistently infected cultures. These cultures contained genomic and extragenomic HIV DNA, the latter being assayed by PCR for two-long terminal repeat circular forms. The status of persistent infection dis- appeared within 2 months. The recovery is due to the replacement of CD4 down-regulated infected cells by overgrowing uninfected cell variants, which are tran- scriptionally inactive for CD4. The mechanisms under- lying the emergence of these variants in persistently infected cultures are considered.
Introduct ion
Human immunodeficiency virus (HIV) is cytopathic for cells of the T lymphocyte lineage both in vitro and in the blood of infected patients. HIV-induced cytopathicity is characterized by syncytium formation and single cell lysis, the latter being prevalent in vivo and contributing to T4 lymphocyte depletion in AIDS patients (Rosenberg & Fauci, 1991).
Many reports have suggested that cytopathicity and host cell range are attributes of the viral genotype (Sakai et al., 1988, 1991; York-Higgins et al., 1990; Kim et al., 1990; Cheng-Mayer et al., 1989), but the host cell also plays a role in HIV-induced cytopathicity. The surface concentration of CD4 receptor can be considered as the critical factor involved in HIV infection and cyto- pathicity for T lymphocytes and derived cell lines. In particular, it has been suggested that the CD4 down- regulation protects the infected cells from death, by inhibiting the cellular accumulation of extragenomic provirus following multiple cycles of reinfection (Steven- son et al., 1988; Marshall et al., 1992).
C8166 T lymphocytes are characterized by extreme permissiveness to HIV infection, as demonstrated by accelerated virus replication, followed in turn by massive cytolysis (Somasundaran & Robinson, 1988). Estab- lishment of persistently infected C8166 cultures has not been reported to date.
In this study, we demonstrate that the survival of C8166 cells following HIV infection correlates with the
initial density of the culture: cells diluted to 50 to 500 cells/ml and infected with HIV maintained their dupli- cation rate and released fully infectious virus into the medium. Persistently infected cultures, therefore, were obtained and these carried integrated provirus as well as extragenomic HIV DNA. These cultures were grown for at least 2 months. During this period, the infected cells progressively decreased and completely disappeared within 50 days. The cultures that had recovered from the infection were virus-free and negative for CD4 mRNA.
M e t h o d s
Cell cultures and cloning procedures. C8166, a human T cell line immortalized by human T cell leukaemia virus (Salahuddin et al., 1983), was routinely maintained in RPMI 1640 medium supplemented with 10 % fetal calf serum (FCS) (RPMI), in Falcon bottles, by splitting the cultures at a 1:4 ratio every 4 days. Clonal subpopulations were obtained by taking cell suspensions at densities of 100, 1000 and 10000 cells/ml in RPMI 1640 medium supplemented with 20 % FCS. To 9 vol of these cell suspensions was added 1 vol of 3 % Bacto Agar, previously dissolved and kept at 42 °C. The agar~ell suspension was distributed in Falcon dishes (3 cm diam., 2 ml dish) and then incubated at 37_+0.5 °C in a 5% CO 2 95% air humidified atmosphere, until macroscopic expansion of clones was achieved.
Production and titration of virus stocks. The H9/HTLV-IIIB cell line was kindly provided by Dr G. B. Rossi (Istituto Superiore di SanitA, Rome, Italy). This line was maintained in RPMI at 37 °C in a CO 2 atmosphere and passaged every 5 days at a 1:5 split ratio. Medium from exponentially growing cells was collected, filtered through 0-45 btm Millipore filters, and employed as virus stock (HIV/H9). The titration of HIV/H9 stocks was performed in 96-well fiat-bottomed Falcon plates. A sample (0-1 ml) of 10-fold dilutions of virus stocks (10 wells
0001-1774 © 1993 SGM

Downloaded from www.microbiologyresearch.org by
IP: 23.22.24.125
On: Sun, 14 Feb 2016 08:37:18
2596 A. Benede t to and others
for each dilution) was added to wells containing 5 x 104 C8166 cells in 0.1 ml. The cultures were incubated and monitored daily for up to 120 h. The wells were scored as positive when at least one syncytium could be observed. Virus titres, expressed as TCIDs0, were calculated by the formula of Reed & Muench (1938).
CD4 expression on the cell surface. The levels of CD4 on infected or uninfected cell surfaces were determined by flow cytometry, by fluorescence-activated cell sorting (FACS) using a FACScan cytometer, using Consort 30 software (Becton-Dickinson), by staining the cells with ftuorescein isothiocyanate (FITC)-labelled OKT4 monoclonal antibody (MAb) (Ortho Diagnostic). In some experiments the presence of CD4 on cells (fixed with 2 % parafornlaldehyde) was assayed by indirect immunofluorescence with OKT4 MAb and FITC-conjugated goat anti-mouse IgG (Sigma).
Calculation of the number of infectious cells in C8166 cultures. Cells from infected cultures were diluted 10-fold in RPMI and distributed in wells (10 wells for each cell dilution) that had been seeded with uninfected C8166 cells (104 cells/well). The plates were incubated at 37 °C for 6 to 8 days and observed for the emergence of syncytia. The minimum number of cells needed for inducing syncytia in 50 % of the previously uninfected C8166 cell cultures was calculated according to the Reed & Muench (1938) formula and expressed as ISCs0.
Purification and J~'actionation of cellular DNA. Genomic and extrachromosomal fractions were separated by the Hirt procedure (Hirt, 1967). The phenol~zhloroform method (Sambrook et al., 1989) was used for extracting DNA from each fraction previously treated with 60 lag/ml proteinase K. Genomic DNA was further purified in 1% agarose Nusieve gel. High M,. DNA was excised and extracted (twice) with phenol-chloroform. Genomic (1 p.g) and extragenomic (from 105 cells) DNAs were analysed by PCR.
Reverse transcription of CD4 mRNA. Total cellular RNA was extracted from 10~cells by the guanidinium thiocyanate method (Chomczynski & Sacchi, 1987) and then dissolved in 30lal of diethylpyrocarbonate-treated water and 10 lag of RNasin was added. A sample (5 ~tl) of RNA solution, denatured at 70 °C, was reverse- transcribed in 20 lal of a mixture containing 50 units of Moloney leukaemia virus reverse transcriptase (Bethesda Research Labor- atories), 10 mM-Tri~HC1 pH 8'3, 7.5 mM-KC1, 2.5 mM-MgC12, 200 laM- dNTP and 0"5 laM of a residue 501 to 472 oligonucleotide (see legend to Fig. 1) as CD4 antisense primer. After 1 h of incubation at 37 °C, 10 lal of the synthesized cDNAs were subjected to PCR analysis.
PCR of DNA and RNA. Genomic and extragenomic DNAs from infected and uninfected C8166 cells and cDNA/CD4 from C8166, HeLa and Jurkat cells were added to 100 lal of amplification mixture containing 0.5 laN of specific primers, 200 laM-dNTP, 50 laM-KC1, 10 mM-Tris-HC1 pH 8-3, 1.5 mM-MgC12, 2.5 units of Amplitaq DNA polymerase (Perkin-Elmer Cetus) and 2 mg/ml gelatin. The same reaction mixture with minor modifications (100laM-dNTP, 5 mM- MgC12, 0'1 laM specific primers) was used for the amplification of 2-LTR (long terminal repeat) circular DNA (Pang et al., 1990).
The samples were subjected to 30 cycles of amplification in the Perkin-Elmer DNA Thermal Cycler. The time and temperature conditions of each cycle were: 2 min at 94 °C, 1.5 min at 55 °C and 2 rain at 72 °C, for HIV and CD4; 1 min at 95 °C, 2 min at 61 °C and 3 min at 72 °C, for mitochondrial DNA; 1 min at 91 °C and 2 min at 65 °C, for 2-LTR circular DNA.
Aliquots (15 lal) of amplified samples were electrophoresed on 2% agarose gel in TBE buffer (90 mM-Tris-HC1 pH 8.0, 90 mM-boric acid, 2 mM-EDTA) containing ethidium bromide. The bands were trans- ferred to Hybond N ÷ membrane (Amersham) in 0.4 M-NaOH under slight vacuum and prehybridized at 55°C for 2 h in 4xSSC, 5 x Denhardt's solution, 0.5 % SDS and 10 lag/ml of denatured salmon
sperm DNA. The hybridization was carried out at 55 °C overnight in the above mixture containing 10 pmol of the T4 polynucleotide kinase a2P-labelled specific probes. After washing with 2 x SSC, 0-1% SDS at room temperature for 15 min, the membranes were autoradiographed.
Determination of CD4 mRNA by Northern blotting. Total RNA was extracted from 107 cells with 4 N-guanidinium thiocyanate followed by centrifugation in CsC1 solution (density 1.8g/ml) according to Chirgwin et al. (1979). Twenty lag of RNA was denatured at 65 °C for 15 rain, electrophoresed on 1% agarose-2.2 N-formaldehyde gel, and transferred to nylon N ÷ membranes with 20 x SSC under slight vacuum. RNA blots were prehybridized for 4 to 6 h at 42 °C in a solution containing 5 x SSC, 50 % formamide, 5 x Denhardt's solution, 0-5 % SDS, and 100 lag/ml of denatured sonicated salmon sperm DNA. Hybridization was performed at 42 °C for 24 h in the same solution supplemented with 5x 108d.p.m./ml of 32P-labelled CD4 probe prepared from the T4-pMV7 plasmid (Maddon et al., 1986). After two washings in 2 x SSC and 0-5 % SDS at room temperature, followed by two washings in 0.2 x SSC and 0-2 % SDS at 65 °C, the membranes were autoradiographed at - 7 0 °C on Kodak X-Omat G film with an intensifying screen (Du Pont). Estimation of the quantity of each RNA was performed by parallel Northern blotting using a 32P-labelled fl-actin probe.
Search for CD4-negative cells by negative selection using immuno- magnetic beads. Uninfected C8166 cells (4x l0 s ceils), suspended in 1 ml of PBS with 3 % FCS, were mixed with M-450 CD4 beads (Dynal) at the ratio of 40 beads/cell. After 30 min incubation in an end-over- end rotor at 4 °C, the CD4-positive cells that were not bound to the beads were recovered by use of an MPC-1 magnet (Dynal) and discarded. The unbound cells were collected, regrown and assayed for surface CD4 expression by FACS analysis.
Complement-induced immunolysis. C8166 cells (2 x 106 cells in 0.2 ml of serum-free RPMI) were treated with 15 lag of T4 MAb (Coulter) and 2 units of guinea-pig complement (Institute Behring, Italy). After 2 h of incubation at 37 °C, the cells were subjected to cloning procedures, as described above.
Results
Cell infection and C D 4 receptor down-regulat ion
I n o u r e x p e r i m e n t a l p r o t o c o l , C 8 1 6 6 cel ls w e r e s e e d e d a t
a d e n s i t y o f 5 x 104 c e l l s / m l i n R P M I c o n t a i n i n g c u l t u r e
m e d i u m o f H 9 / H T L V - I I I B cel ls ; t h e f ina l c o n c e n t r a t i o n
o f v i r u s w a s 10 G7 T C I D ~ 0 / m l , e q u i v a l e n t to a n m.o . i , o f
a b o u t 100 T C I D s 0 p e r cell. T h e c u l t u r e s w e r e i n c u b a t e d
a t 37 °C ( t a k e n as t he s t a r t i n g t i m e o f i n f e c t i o n ) . R a r e
s y n c y t i a w e r e a l r e a d y v i s ib le 24 h p o s t - i n f e c t i o n (p. i . )
a n d i n c r e a s e d in n u m b e r a n d size r e a c h i n g m a x i m u m
v a l u e s a t 72 h p.i . S y n c y t i a p r o g r e s s i v e l y d i s a p p e a r e d
t h e r e a f t e r w h e r e a s g e n e r a l d a m a g e t o s ing le cells in -
c r e a s e d . A t 6 to 8 d a y s p.i . a l l c u l t u r e s a p p e a r e d to h a v e
d e g e n e r a t e d . T h e s a m e r e s u l t w a s f o u n d w h e n c u l t u r e s
we re e x p o s e d to t h e v i r u s f o r 0 to 24 h, f o l l o w e d b y
r e p l a c e m e n t o f t he v i r u s i n o c u l u m w i t h f r e s h R P M I . T h e
eff icacy o f t h e s e p r o t o c o l s in i n f e c t i n g t h e cells o f t he
c u l t u r e was a s se s sed b y c a l c u l a t i n g t he m i n i m a l n u m b e r
o f cells a b l e to i n d u c e s y n c y t i a w h e n c o c u l t i v a t e d w i t h
u n i n f e c t e d C 8 1 6 6 cell c u l t u r e s ( e x p r e s s e d as ISCs0 ). T h e
resu l t s , s h o w n in T a b l e 1, d e m o n s t r a t e d t h a t m o s t cells

Downloaded from www.microbiologyresearch.org by
IP: 23.22.24.125
On: Sun, 14 Feb 2016 08:37:18
H I V infection: role of cell density 2597
Table 1. Cellular infectivity o f cultures infected with HIV-1
Cellular infectivity Time (h) p.i. (ISCs0)* Dead cells (%)t (m.o.i. 100
TCID~o/cell ) 1 ~: 2 1 2
24 0.6 1-2 N D § N D
96 0"3 0"7 98 94
* The numbers indicate exponents to the base 10. t Determined 144 h p.i. by the trypan blue exclusion method. :~ 1, First experiment; 2, second experiment. §NO, Not determined.
(a) 150
24 h i
0 . . . . ;i;,, I ........
48 h 72 h
2;- ;~
I I I I I I i i i i 1 1 i I I I I i i i I i i i i i i i i
10 o 101 10 o
96 h
i i l l l n l I i i r l
10 o 101 lO 1 10 o 10 l
(b) C8166 24 48 72 96 H J
Fig. 1. Down-regulation of CD4 receptor in the course of HIV acute infection. M.o.i. was about 100 TCID~0/cell. (a) FACS profiles of HIV-infected C8166 cells at different times p.i. The solid line indicates staining with FITC-conjugated OKT4 MAb; the dotted line indicates unstained cells. (b) PCR analysis of CD4 mRNA in C8166 cells at the times (h) indicated, using oligonucleotides of residues 145 to 174 and 501 to 472 as primers, and residues 336 to 357 as probe (Maddon et al., 1985)• HeLa (H) and Jurkat (J) cells were used as negative and positive controls, respectively.
were already infected 24 h after the initial exposure to the virus. The subsequent rate of cell reinfection will depend on the a m o u n t of virus released into the med ium and on the n u m b e r of C D 4 molecules present on the surface of infected cells. C D 4 expression in C8166 lymphocytes was invest igated by F A C S analysis at var ious t ime intervals dur ing the course o f H I V infection. As shown in Fig. 1, C D 4 down-regula t ion begins 72 h p.i. and reaches its m a x i m u m 96 h p.i., in the absence of any appreciable change in cellular levels o f C D 4 m R N A .
Influence of cell density o f infected cultures on survival rate and virus expression
An easy way to reduce or prevent cell reinfection wi thout using specific ant ibodies or drugs is to infect cultures seeded at very low density; in this way, the concent ra t ion
Fig. 2. Microscopic observations (a, b) and immunofluorescence analysis of HIV antigens (c,d) of cells infected with HIV/H9 (10nTTCIDs0/ml). Panels (a) and (c) show cultures seeded at 5 x 104 cells/rot and observed 72 h p.i.; (b) and (d) show cultures seeded at 50 cells/ml and observed 12 days p.i. Indirect immunofluorescence was performed using a pool of sera from HIV-l-seropositive patients, as previously described (Szab6 et al., 1993).
o f the new virus p roduced by the cells is lowered. Serial 10-fold dilutions of C8166 cells (10 ~, 10 a, 102, 101 cells/ 0.2 ml/wel l , 20 wells for each dilution) were seeded in multi-well plates in the presence o f H I V - 1 / H 9 (final concent ra t ion was 106•7 T C I D S m l ). The cultures were incubated at 37 °C. The survival o f cells in each well was determined using microscopy. Three or 4 days p.i. all cultures seeded at 104 cells/well showed large syncytia (Fig. 2 a) and intense HIV-specif ic fluorescence (Fig. 2 c); later (6 to 8 days p.i.) these cultures appeared to have complete ly degenerated.
In more dilute cultures the ou tcome of infection, however , differed significantly: the percentage o f sur- viving and overgrowing cultures increased as the initial cell density decreased (Fig. 3). Ten to 15 days p.i., in the major i ty of the cultures initially seeded at 101 cells per well, the cells appeared clustered (Fig. 2b) and showed weak HIV-specif ic fluorescence (Fig. 2d). Fif teen days p.i., cell counts pe r fo rmed on 10 r andomly chosen cultures (initial cell density 101 cells per well) had mean values of 32 (_+ 8) x 103 cells per well, indicating a growth rate o f abou t 0"8 cell duplicat ions per day. Serially diluted cells seeded in two addi t ional plates and infected with H I V as above were again given fresh H I V / H 9 , 24 h

Downloaded from www.microbiologyresearch.org by
IP: 23.22.24.125
On: Sun, 14 Feb 2016 08:37:18
2598 A. Benedetto and others
100
75
r~
" 50 -~. -~.
25
I I I I
I I I V
101 102 103 104
Density (cells/ml)
Fig. 3. Survival of HIV-infected C8166 cell cultures seeded at different ceil densities and infected with HIV/H9 (106-~ TCIDs0/ml ). ©, Cells infected only once; ~ , cells reinfected 24 h p.i_; 0 , cells reinfected 72 h p.i.
(a)
Genomic fraction
1 2 3 4 5 6 7 8
Hirt fraction
(b)
1 2 3 4 5 6 7 8
Mitochondrial D N A
Fig. 4. (a) PCR analysis of D N A from genomic ( U ~ g a g region of HIV) and Hirt (2-LTR circle) fractions of cultures seeded at different cell densities and infected with HIV-1 (10 *'7 TCIDs0/ml ). The cultures were seeded at 5 x 104 cells/ml in acute infection (lanes 1 to 5) and at 50 cells/ml in chronic infection (lanes 6 to 8). Lane 1, 24 h; lane 2, 48 h; lane 3, 72 h; lane 4, 96 h; lane 5, 96 h p.i. of a culture treated with AZT (20 nM) in the 24 to 96 h time interval; lane 6, 12 days; lane 7, 20 days; lane 8, 30 days p.i. The U5 gag region was amplified using the primers M667 and M661 reported by Pang et al. (1990). Oligonucleotides M667 and M884 were used to detect 2-LTR circles of HIV-I DNA (Pang et al., 1990). Southern blot hybridization was performed with the oligonucleotide residues 619 to 638 (Sanchez-Pescador et al., 1985) as a probe. (b) Ethidium bromide staining of mitochondrial DNA amplified from the Hirt fraction of (a). PCR was performed according to Stevenson et al. (1988) using oligonucleotides of residues 247 to 267 and 673 to 653 as primers (Anderson et al., 1981).
and 72 h after cell seeding, respectively. The cultures reinfected at 72 h showed rates of cell survival similar to those of cultures infected only once. In contrast, cultures reinfected at 24 h showed a lower survival rate (Fig. 3). These results indicate that 24 h after the first exposure to the virus, most of the cells were still susceptible to the killing effects of a second inoculum whereas at 72 h p.i. the cells became resistant to superinfection. This re- sistance to superinfection, exhibited by the low density cultures, coincides temporarily with strong down-regu- lation of the CD4 receptor, as determined by indirect immunofluorescence assay (IFA) using the OKT4 MAb (not shown).
Integrated and extragenomic HIV DNA in acutely and persistently infected cultures
The presence of integrated and extragenomic HIV DNA was determined by PCR analysis of the U5-gag region (genomic fraction) and 2-LTR circle molecules (Hirt fraction) in cultures seeded at a cell density of 5 x 104 cells/ml (acute infection) and 50 cells/ml (chronic infection). As shown in Fig. 4, the amplification of DNA extracted from the cells collected 24 h p.i. gave strong signals for the gag region in the genomic DNA fraction
and very weak ones for the 2-LTR circular molecules in the Hirt fraction. At later times the 2-LTR circle positivity increased considerably. This increase was almost completely inhibited when azidothymidine (AZT) (20 riM) was added to the cells 24 h p.i., suggesting that the accumulation of extragenomic DNA was due to multiple rounds of cell reinfection.
We also investigated HIV DNA forms in the genomic and extragenomic fractions of persistently infected cultures obtained from highly diluted cells. To this end a diluted cell suspension (50 cells/ml) was distributed in multi-well plates (0.2 ml/well) and infected with HIV/H9 at the final concentration of 106.7 TCIDs0/ml. Ten days after seeding, most of the wells contained fully growing cultures. The cells of each well were mixed together, resuspended in fresh medium at a density of 105 celts/ml and reincubated. This culture was maintained by diluting the cells 1 : 5 at every passage. The cells were harvested 12, 20 and 30 days p.i. Genomic and extragenomic DNAs were extracted from each cell sample and analysed by PCR as above. Strong signals for the gag region were obtained in the genomic fractions. All cell samples were PCR-positive for 2-LTR DNA circles, indicating the presence of extrachromosomal HIV DNA during the growth of the persistently infected cultures (Fig. 4).
Downloaded from www.microbiologyresearch.org by
IP: 23.22.24.125
On: Sun, 14 Feb 2016 08:37:18
HIV infection : role of cell density 2599
Table 2. Follow-up of persistently infected S1 culture
Number of infected cells HIV D N A t
Cultivation IFA-positive time* cells (%) ISCs0 § Genomic Extragenomic
Surface CDa:~
CD4 m R N A (PCR)
12 71 0.5 + + 20 23 1.2 + ND 30 28 NDII + + 40 3 > 4 ND ND 60 0 ND -- --
+ +
Weak
* Days after cell seeding (10 cells per well). t PCR analysis of U5-gag in genomic fraction and of 2-LTR circle in Hirt fraction. :~ FACS analysis with OKT4 MAb. §The numbers indicate exponents to the base 10. I] ND, Not determined.
1 2 3 4 5
CD4 m R N A ~ ~ 3 kb
fl-Actin
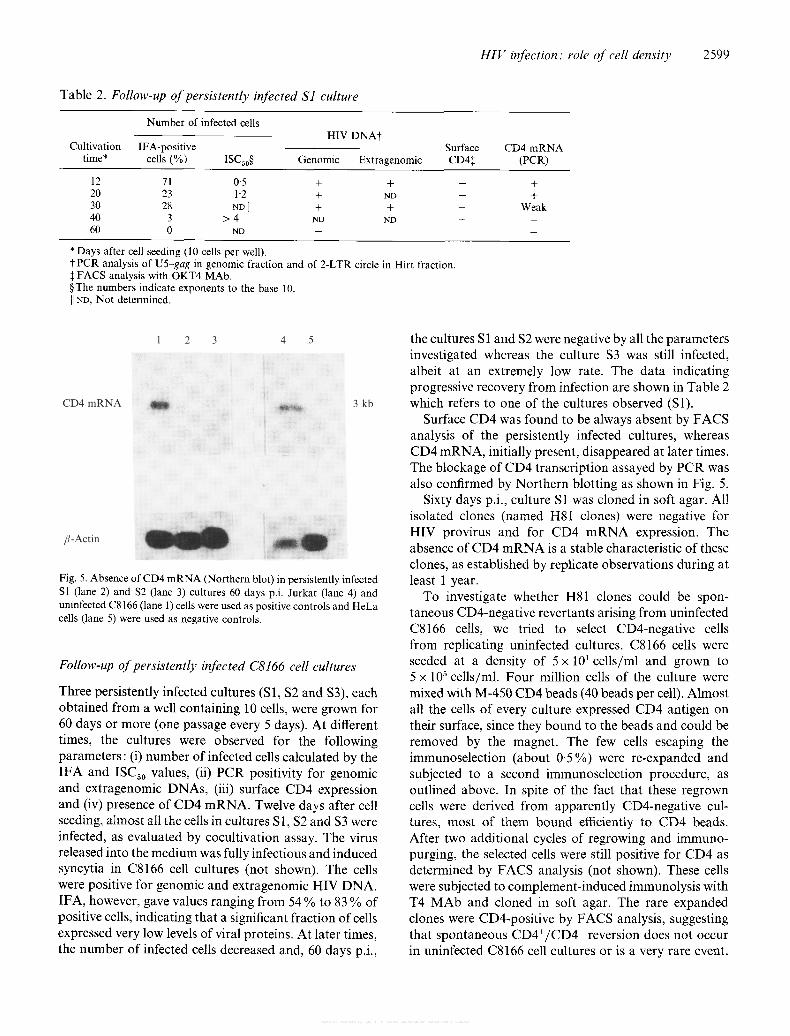
Fig. 5. Absence of CD4 m R N A (Northern blot) in persistently infected Sl (lane 2) and S2 (lane 3) cultures 60 days p.i. Jurkat (lane 4) and uninfected C8166 (lane 1) cells were used as positive controls and HeLa cells (lane 5) were used as negative controls.
Follow-up of persistently infected C8166 cell cultures
Three persistently infected cultures (S1, $2 and $3), each obtained from a well containing 10 cells, were grown for 60 days or more (one passage every 5 days). At different times, the cultures were observed for the following parameters: (i) number of infected cells calculated by the IFA and ISCs0 values, (ii) PCR positivity for genomic and extragenomic DNAs, (iii) surface CD4 expression and (iv) presence of CD4 mRNA. Twelve days after cell seeding, almost all the cells in cultures SI, $2 and $3 were infected, as evaluated by cocultivation assay. The virus released into the medium was fully infectious and induced syncytia in C8166 cell cultures (not shown). The cells were positive for genomic and extragenomic HIV DNA. IFA, however, gave values ranging from 54 % to 83 % of positive cells, indicating that a significant fraction of cells expressed very low levels of viral proteins. At later times, the number of infected cells decreased and, 60 days p.i.,
the cultures S 1 and $2 were negative by all the parameters investigated whereas the culture $3 was still infected, albeit at an extremely low rate. The data indicating progressive recovery from infection are shown in Table 2 which refers to one of the cultures observed (Sl).
Surface CD4 was found to be always absent by FACS analysis of the persistently infected cultures, whereas CD4 mRNA, initially present, disappeared at later times. The blockage of CD4 transcription assayed by PCR was also confirmed by Northern blotting as shown in Fig. 5.
Sixty days p.i., culture S1 was cloned in soft agar. All isolated clones (named HS1 clones) were negative for HIV provirus and for CD4 mRNA expression. The absence of CD4 mRNA is a stable characteristic of these clones, as established by replicate observations during at least 1 year.
To investigate whether H81 clones could be spon- taneous CD4-negative revertants arising from uninfected C8166 cells, we tried to select CD4-negative cells from replicating uninfected cultures. C8166 cells were seeded at a density of 5 x 101 cells/ml and grown to 5 x 105 cells/ml. Four million cells of the culture were mixed with M-450 CD4 beads (40 beads per cell). Almost all the cells of every culture expressed CD4 antigen on their surface, since they bound to the beads and could be removed by the magnet. The few cells escaping the immunoselection (about 0'5 %) were re-expanded and subjected to a second immunoselection procedure, as outlined above. In spite of the fact that these regrown cells were derived from apparently CD4-negative cup tures, most of them bound efficiently to CD4 beads. After two additional cycles of regrowing and immuno- purging, the selected cells were still positive for CD4 as determined by FACS analysis (not shown). These cells were subjected to complement-induced immunolysis with T4 MAb and cloned in soft agar. The rare expanded clones were CD4-positive by FACS analysis, suggesting that spontaneous CD4+/CD4 - reversion does not occur in uninfected C8166 cell cultures or is a very rare event.

Downloaded from www.microbiologyresearch.org by
IP: 23.22.24.125
On: Sun, 14 Feb 2016 08:37:18
2600 A. Benedetto and others
Discussion
C8166 cells diluted to 50 to 500 cells/ml almost com- pletely maintain their cell duplication rate while they continue to release infectious virus into the medium. This is consistent with the hypothesis that the reduced rate of cell reinfection occuring in HIV-infected low density cultures leads to a slowing of virus replication. In this way, CD4 down-regulation occurs before irreversible cellular damage takes place. As expected, such a steady state can be interrupted by adding fresh virus to the cultures in the course of 48 to 72 h p.i. Subsequently, the cells become stably resistant to superinfection, permitting the establishment of persistently infected cultures. In accordance with the above hypothesis, we found that the cells of low density cultures down-regulated their CD4 receptor, starting from 72 h p.i. This was observed by IFA using OKT4 MAb, which binds to the receptor in a different position from the one involved in gp 120 binding (Sattentau et al., 1986).
The effect of superinfection on cell death has been studied by Bergeron & Sodroski (1992). They provide evidence that cell treatments inhibiting cell superinfection do not prevent the single cell lysis of C8166 cells infected with the viral clone HXBc2. Although it is not easy to compare results obtained with two different virus strains, we are inclined to think that the increased cell survival of low density cultures is due only in part to a reduced rate of superinfection. The involvement of mechanisms related to the cell crowding conditions cannot be excluded. In this context, it may be relevant that the saturation density (at the end of the culture growth) of C8166 cells is lower than that found in other T cell lines, such as CEM cells (about 7 x 105, and 1.2 x 106 cells/ml, respectively, under our experimental conditions).
In the course of acute infection, the 2-LTR circles accumulated. This accumulation was inhibited by treat- ing the cells with AZT, suggesting that the extragenomic DNA accumulation is due to multiple rounds of cell reinfection. This assumption is consistent with the conclusions reported by others (Besansky et al., 1991; Bergeron & Sodroski, 1992). However, as long as the mechanism(s) leading to the accumulation of extra- genomic HIV DNA in CD4 down-regulated C8166 cells is not clarified, the question remains open. We propose, therefore, that a very low concentration of surface CD4 (undetectable by FACS) could be sufficient to allow the cells to be reinfected. In keeping with this hypothesis, no correlation between unintegrated HIV DNA and surface CD4 expression was found by Besansky et al. (1991) in HIV-infected J1 cells.
The establishment of persistent infection obtained by infecting low density C8166 ceils is not permanent:
within 2 months every infected cell in the culture disappeared. This suggests that the recovery from the infection is due to the replacement of CD4 down- regulated infected cells with overgrowing uninfected cells, the latter being transcriptionally inactive for CD4. A reduced synthesis of CD4 mRNA has already been reported by Hoxie et al. (1986) in HIV-infected CEM cells. It is reasonable to assume that the uninfected/CD4 mRNA-negative cell variants were already pre-existent in uninfected C8166 cell populations. Yet, in our experiments these cell variants emerged from cultures obtained by adding virus to wells containing only 10 cells, which would imply that at least one CD4-negative cell out of ten (10%) should be present in C8166 cell populations at the start of infection. It is very unlikely, however, that so high a proportion of cell variants could have escaped detection by FACS analysis. Another possible explanation for the long-term emergence of CD4-negative uninfected cell variants could be that the high rate of spontaneous CD4+/CD4 - reversion is a peculiar property of C8166 cell cultures. A similar hypothesis has been advanced by Folks et al. (1986) for the origin of infected CD4-negative revertants in another cell line infected by HIV. We have examined this hypothesis by subjecting growing uninfected C8166 cell cultures to selective pressure against CD4-positive cells. In this context, we mimicked the cytocidal effect of HIV, by removing CD4-positive cells with immunobeads followed by complement-dependent immunolysis of immunopurged cells. In spite of this strong selection, no CD4-negative cell variants were ever detected in unin- fected C8166 cell cultures. This indicates that HIV infection is apparently indispensable for positive selec- tion of uninfected CD4 mRNA-negative variants to occur. The mechanism for this, however, remains unknown. We have isolated several uninfected clones (H81) from persistently infected C8166 cell cultures, all of which showed a similar morphology, growth behaviour and surface phenotype. A better under- standing of the genetic mechanism(s) involved in the stable CD4 inhibition of these clones would shed light on the nature of the events leading to their emergence.
The authors thank Professor R. Perez-Bercoff (University 'La Sapienza', Rome, Italy) for critical comments on the manuscript and Dr M. Equestre (lstituto Superiore di Sanitg, Rome, Italy) for his excellent assistance in the GenBank data analysis. This work was supported by grant no. 8204.14 from the Ministry of Health of Italy (V Progetto di Ricerche sull'AIDS, 1991).
References ANDERSON, S., BANKIER, A. T., BARRELL, B. G., DE BRUIJN, M. H. L.,
COULSON, A. R., DROUIN, J., EPERON, I. C., NIERLICH, D. P., ROE, B. A., SANGER, F., SCHREIER, P. H., SMITH, A. J. H., STADEN, R. & YOUNG, I.G. (1981). Sequence and organization of the human mitochondrial genome. Nature, London 290, 457~,65.

Downloaded from www.microbiologyresearch.org by
IP: 23.22.24.125
On: Sun, 14 Feb 2016 08:37:18
HIV infection" role of cell density 2601
BERGERON, L. & SODROSKI, J. (1992). Dissociation of unintegrated viral DNA accumulation from single-cell lysis induced by human immunodeficiency virus type 1. Journal of Virology 66, 5777-5787.
BESANSKY, N.J., BUTERA, S.T., SINHA, S. & FOLKS, T. M. (1991). Unintegrated human immunodeficiency virus type 1 DNA in chronically infected cell lines is not correlated with surface CD4 expression. Journal of Virology 65, 2695-2698.
CHENG-MAYER, C., WEISS, C., SETO, D. & LEVY, J. A. (1989). Isolates of human immunodeficiency virus type 1 from the brain may constitute a special group of the AIDS virus. Proceedings of the National Academy of Sciences, U.S.A. 86, 8575-8579.
CHIRGWIN, J. M., PRYZBYLA, A. E., McDONALD, R. J. & RUTTER, W, J. (1979). Isolation of biologically active ribonucleic acid from sources enriched in ribonuclease. Biochemistry 18, 5294-5299.
CHOMCZYNSKI, P. & SACCHI, N. (1987). Single-step method of RNA isolation by acid guanidinium thiocyauate-phenol-chloroform extraction. Analytical Biochemistry 162, 156-159.
FOLKS, T. M., POWELL, D., LIGHTFOOTE, M., KOENIG, S., FAUCI, A. S., BENN, S., RABSON, A., DAUGHERTY, D., GENDELMAN, H.E., HOGGAN, M. D., VENKATESAN, S. & MARTIN, M. A. (1986). Bio- logical and biochemical characterization of a cloned Leu-3 cell surviving infection with the acquired immunodeficiency syndrome retrovirus. Journal of Experimental Medicine 164, 280-290.
HIRT, B. (1967). Selective extraction of polyoma DNA from infected mouse cell cultures. Journal of Molecular Biology 26, 365-369.
HOXIE, J. A., ALPERS, J.D., RACKOWSKI, J. L., HUEBNER, K., HAG- GARTY, B. S., CEDARBAUM, A. J. & REED, J. C. (1986). Alterations in T4 (CD4) protein and mRNA synthesis in cells infected with HIV. Science 234, 1123-1127.
KIM, S., IKEUCHI, K., GROOPMAN, J. & BATIMORE, D. (1990). Factors affecting cellular tropism of human immunodeficiency virus. Journal of Virology 64, 5600-5604.
MADDON, P.J., LITTMAN, D.R., GODFREY, M., MADDON, D.E., CHESS, L. & AXEL, R. (1985). The isolation and nucleotide sequence of a c-DNA encoding the T cell surface protein T4: a new member of the immunoglobulin gene family. Cell 42, 93-104.
MADDON, P. J., DALGLEISH, A. G., McDOUGAL, J. S., CLAPHAM, P. R., WEISS, R. A. & AXEL, R. (1986). The T4 gene encodes the AIDS virus receptor and is expressed in the immune system and the brain. Cell 47, 333 348.
MARSHALL, W. L., DIAMOND, D. C,, KOWALSKI, M.M. & FINBERG, R.W. (1992). High level of surface CD4 prevents stable human immunodeficiency virus infection of T-cell transfectants. Journal of Virology 66, 549~5499.
PANG, S., KONYANAGI, Y., MILES, S., WILEY, C., MINTERS, H.V. & CHEN, I. S. Y. (1990). High levels of unintegrated HIV-1 DNA in brain tissue of AIDS dementia patients. Nature, London 343, 85-89.
REED, L. J. & MUENCH, H. (1938). A simple method of estimating fifty percent endpoints. American Journal of Hygiene 27, 493-497.
ROSENBERG, Z, F. & FAUCI, A. S. (1991). Immunopathogenesis of HIV infection. FASEB Journal 5, 2382 2390.
SAKAI, K., DEWHURST, S., MA, X. & VOLSKY, n . J. (1988). Differences in cytopathogenicity and host cell range among infectious molecular clones of human immunodeficiency virus type 1 simultaneously isolated from an individual. Journal of Virology 62, 4078-4085.
SAKAI, K., MA, X., GORDIENKO, I. & VOLSKY, D. J. (1991). Recombin- ational analysis of a natural noncytopathic human immunodeficiency virus type 1 (HIV-1) isolate: role of the vif gene in HIV-1 infection kinetics and cytopathicity. Journal of Virology 65, 5765-5773.
SALAHUDDIN, S. Z., MARKHAM, P. D., WONG-STAAL, F., FRANCHINI, G., KALYANARAMAN, V.S. & GALLO, R.C. (1983). Restricted expression of human T-cell leukemia-lymphoma virus (HTLV) in transformed human umbilical cord blood lymphocytes. Virology 129, 51-64.
SAMBROOK, J., FRITSCH, E.F. & MANIATIS, T. (1989). Molecular Cloning: A Laboratory Manual, 2nd edn. New York: Cold Spring Harbor Laboratory.
SANCHEZ-PESCADOR, R., POWER, M, O., BARR, P.J., STEIMER, K. S., STEMPIEN, M- M., BROWN-SHIMMER, S. L., GEE, W. W, RENARD, A., RANDOLPH, A., LEVY, J.A., DINA, D. & LUCIW, P.A. (1985). Nucleotide sequence and expression of an AIDS-associated retro- virus (ARV-2). Science 227, 484492.
SATTENTAU, Q. J., DALGLEISH, A. G., WEISS, R. A. & BEVERLY, P. C. L. (1986). Epitopes of the CD4 antigen and HIV infection. Science 234, 1120-1123.
SOMASUNDARAN, M. & ROBINSON, H.L. (1988). Unexpectedly high levels of HIV-1 RNA and protein synthesis in a cytocidal infection. Science 242, 1554-1557.
STEVENSON, M., MEIER, C., MANN, A. M., CHAPMAN, N. & WASIAK, A. (1988). Envelope glycoprotein of HIV induces interference and cytolysis resistance in CD4 cells: mechanism for persistence in AIDS. Cell 53, 483-496.
SZAB(), B., LOCARDI, C., LO PRESTI, E., BELARDELLI, F. & BENEDETTO, A. (1993). Tumor necrosis factor-alpha increases the sensitivity of human immunodeficiency virus (HIV)-infected monocytic U937 cells to the complement-dependent cytotoxicity of sera from HIV type 1- infected individuals: role of the gpl20 protein. Journal of General Virology 74, 1271-1276.
YORK-HIGGINS, D., CHENG-MAYER, C., BAUER, D., LEVY, J. A. & DINA, D. (1990). Human immunodeficiency virus type 1 cellular host range, replication, and cytopathicity are linked to the envelope region of the viral genome. Journal of Virology 64, 4016-4020.
(Received 20 April 1993 ," Accepted 27 July1993)
Copyright © 2022 FDOKUMEN




















