
Vicariance in a Cryptic Species Pair of European Pseudoscorpions (Arachnida, Pseudoscorpiones,...
13
0044-5231/04/242/04-299 $ 15.00/0 Vicariance in a Cryptic Species Pair of European Pseudoscorpions (Arachnida, Pseudoscorpiones, Chthoniidae) Christoph MUSTER 1 , Thomas SCHMARDA 2 and Theo BLICK 3 1 Staatliche Naturhistorische Sammlungen Dresden, Museum für Tierkunde, Germany 2 Institute of Zoology and Limnology, University of Innsbruck, Austria 3 Hummeltal, Germany Abstract. Following the first records of Chthonius (Ephippiochthonius) boldorii Beier, 1934 in central Europe, a species which was previously assumed to occur exclusively in Mediterranean caves, 116 series (595 specimens) of the cryptic taxa C. (E.) boldorii and C. (E.) fuscimanus Simon, 1900 (Syn. C. (E.) austriacus Beier, 1931) were re- examined. Although multivariate analyses suggest specific separation, there is only one unequivocal character for discrimination: the presence or absence of a single isolated tooth on the moveable finger of the chelicerae. The dis- tributions were found to be largely allopatric, therefore it is concluded that the species rank of the two morpho- species is justified. North of the Alps, an almost vicariant pattern emerged: east of 14° E fuscimanus occurs, west of this line boldorii occurs. The results provide a basis for discussing the relevance of minute morphological dif- ferences in pseudoscorpion taxonomy. Key words. Pseudoscorpiones, species differentiation, taxonomy, discriminant analysis, vicariance, biogeography. 1. INTRODUCTION Species delimitation in Pseudoscorpiones is often based on minute, sometimes variable characters of the external morphology. This is true for the sibling species Chthonius (E.) fuscimanus Simon, 1900 (= C. (E.) austriacus Beier, 1931) and C. (E.) boldorii Beier, 1934. The latter was originally described from a single specimen as a cavernicolous Mediterranean subspecies of the epigeous C. (E.) austriacus. BEIER (1934) con- sidered it to differ from the nominate subspecies in the length/breadth ratio of the chela, the dentation of chelal fingers, the vestigial posterior eyes and the num- ber of coxal spines. In a later determination key BEIER (1963) focused on the reduction of the posterior eyes in boldorii as discriminating feature. During re-examina- tion of the type material, MAHNERT (1980) detected the presence of a small isolated subdistal tooth on the inner margin of the cheliceral movable finger (subsequently abbreviated as di according to GABBUTT & V ACHON 1963) in C. (E.) austriacus boldorii, which is absent in the type of C. (E.) austriacus. He considered the exis- tence of such a tooth a taxonomic feature of superior value to the development of the posterior eyes (see also GABBUTT & V ACHON 1963: 79), and he therefore ele- vated boldorii to species level. However, this decision was based on only a few specimens, and neither intrapopulational nor geographic character variation was investigated. Thus, intraspecific dimorphism could not be completely excluded. Subsequent to the recognition of di as a relevant trait for species distinction, Chthonius (E.) boldorii appeared to be a widespread epigeous pseudoscorpion in central Europe. The first records in southern Switzerland (COTTI 1989, det. GARDINI) were followed by findings in Austria (SCHMARDA 1995) and by exten- sions of the known range to northern Switzerland (DEVORE-SCRIBANTE 1999) and Germany, the latter during the preparation of a threatened species list and checklist of the Bavarian Pseudoscorpiones (BLICK & MUSTER 2003, SPELDA in litt. 2002). These records encouraged us to re-examine all material of “Chtho- nius (E.) austriacus” and “C. (E.) boldorii” from the Museum of Natural History in Vienna (NHMW) and the Museum of Natural History in Stuttgart (SMNS), as well as further available series. The aims of this Zool. Anz. 242 (2004): 299–311 http://www.elsevier-deutschland.de/jcz
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Vicariance in a Cryptic Species Pair of European Pseudoscorpions (Arachnida, Pseudoscorpiones,...

0044-5231/04/242/04-299 $ 15.00/0
Vicariance in a Cryptic Species Pairof European Pseudoscorpions(Arachnida, Pseudoscorpiones, Chthoniidae)
Christoph MUSTER1, Thomas SCHMARDA2 and Theo BLICK3
1 Staatliche Naturhistorische Sammlungen Dresden, Museum für Tierkunde, Germany2 Institute of Zoology and Limnology, University of Innsbruck, Austria3 Hummeltal, Germany
Abstract. Following the first records of Chthonius (Ephippiochthonius) boldorii Beier, 1934 in central Europe, aspecies which was previously assumed to occur exclusively in Mediterranean caves, 116 series (595 specimens) ofthe cryptic taxa C. (E.) boldorii andC. (E.) fuscimanusSimon, 1900 (Syn. C. (E.) austriacusBeier, 1931) were re-examined. Although multivariate analyses suggest specific separation, there is only one unequivocal character fordiscrimination: the presence or absence of a single isolated tooth on the moveable finger of the chelicerae. The dis-tributions were found to be largely allopatric, therefore it is concluded that the species rank of the two morpho-species is justified. North of the Alps, an almost vicariant pattern emerged: east of 14° E fuscimanusoccurs, westof this line boldorii occurs. The results provide a basis for discussing the relevance of minute morphological dif-ferences in pseudoscorpion taxonomy.
Key words.Pseudoscorpiones, species differentiation, taxonomy, discriminant analysis, vicariance, biogeography.
1. INTRODUCTION
Species delimitation in Pseudoscorpiones is oftenbased on minute, sometimes variable characters of theexternal morphology. This is true for the siblingspecies Chthonius (E.) fuscimanusSimon, 1900 (= C.(E.) austriacusBeier, 1931) and C. (E.) boldoriiBeier,1934. The latter was originally described from a singlespecimen as a cavernicolous Mediterranean subspeciesof the epigeous C. (E.) austriacus. BEIER (1934) con-sidered it to differ from the nominate subspecies in thelength/breadth ratio of the chela, the dentation ofchelal fingers, the vestigial posterior eyes and the num-ber of coxal spines. In a later determination key BEIER
(1963) focused on the reduction of the posterior eyes inboldorii as discriminating feature. During re-examina-tion of the type material, MAHNERT (1980) detected thepresence of a small isolated subdistal tooth on the innermargin of the cheliceral movable finger (subsequentlyabbreviated as di according to GABBUTT & VACHON
1963) in C. (E.) austriacus boldorii, which is absent inthe type of C. (E.) austriacus. He considered the exis-tence of such a tooth a taxonomic feature of superior
value to the development of the posterior eyes (see alsoGABBUTT & VACHON 1963: 79), and he therefore ele-vatedboldorii to species level. However, this decisionwas based on only a few specimens, and neitherintrapopulational nor geographic character variationwas investigated. Thus, intraspecific dimorphismcould not be completely excluded.Subsequent to the recognition of di as a relevant traitfor species distinction, Chthonius (E.) boldoriiappeared to be a widespread epigeous pseudoscorpionin central Europe. The first records in southernSwitzerland (COTTI 1989, det. GARDINI) were followedby findings in Austria (SCHMARDA 1995) and by exten-sions of the known range to northern Switzerland(DEVORE-SCRIBANTE 1999) and Germany, the latterduring the preparation of a threatened species list andchecklist of the Bavarian Pseudoscorpiones (BLICK &MUSTER 2003, SPELDA in litt. 2002). These recordsencouraged us to re-examine all material of “Chtho-nius (E.) austriacus” and “C. (E.) boldorii” from theMuseum of Natural History in Vienna (NHMW) andthe Museum of Natural History in Stuttgart (SMNS),as well as further available series. The aims of this
Zool. Anz. 242 (2004): 299–311http://www.elsevier-deutschland.de/jcz

study are to investigate the geographic distribution ofthe two character states (di present or absent), to searchfor associated discriminating traits, and to draw con-clusions regarding the biogeographic history and thetaxonomic status of the two “forms”. By investigatingphenotypic plasticity and geographic associations, ageneral evaluation of the usefulness of single morpho-logical characters in pseudoscorpion taxonomy is pro-vided.
2. MATERIALAND METHODS
This study is based on both museum collections andmaterial recently collected in Germany. A total of 595specimens were examined. According to the presenceor absence of di, 264 specimens (168�, 96�) wereidentified as C. (E.) fuscimanusand 331 (121�, 110�)as C. (E.) boldorii. The majority of the material camefrom the NHMW, including the types of C. (E.) austri-acus and C. (E.) boldorii. This collection comprises376 individuals from 87 series (stored under bothnames). 51 specimens from 11 series are stored at theSMNS. Three specimens belonging to these taxa aredeposited in the MTD (Museum für Tierkunde Dres-den). The remaining material came from private col-lections: K. LIPPOLD (111), T. SCHMARDA (5), V.DUCHAC (4), K. ENGEL (1). 44 individuals were recent-ly collected in Bavaria by T. BLICK & M.-A. FRITZE
(now in collection of C. MUSTER).For numerical morphological analysis, measurementsof 307 specimens from the whole range of distributionwere taken (C. (E.) fuscimanus66�, 41�; C. (E.)boldorii 133�, 67�). The following characters weredetermined:CL – carapace lengthCW – carapace widthFL – length of palpal femurTFF – number of teeth on fixed finger of pedipalpTMF– number of teeth on moveable finger of pedipalp
CL, CW, FL are related to body length and proportions,TFF and TMF are important taxonomic features inpseudoscorpions. Parametric (2-tailed t-test) and non-parametric (Mann-Whitney-U) tests for equality ofmeans as well as discriminant analysis were done withSPSS 11.0 for Windows.Microscopic details were investigated using the NikonEclipse 600 instrument. Measurements were takenwith ocular micrometers using BEIER’S (1963: Fig. 11)reference points. Palps were not dissected for measure-ment, to eliminate parallax errors attention was paid toprecise orientation of specimens. Photographs weretaken with the photomicrographic equipment Nikon H-III, using interference contrast with extended focusoption (Digitaloptics, Jena).
3. RESULTS
3.1. Taxonomy
With respect to the expression of the isolated subdistaltooth on the movable finger of the chelicerae, the mate-rial contained two discontinuous forms which are notlinked by intermediates in a morphocline. No singlespecimen with an asymmetric distribution of this char-acter was observed. More importantly, in the 116 seriesexamined (with a mean number of 5 individuals) thistooth is consistently either present or absent (for oneexception see 3.3.). This character constancy withinthe populations led us to reject the possibility of anintraspecific polymorphism and instead accept theexistence of two discrete taxa, for which the names C.(E.) fuscimanus(di present, Figs. 1, 10) and C. (E.)boldorii (di absent, Figs. 2, 16) are available. In con-clusion, 61 series (with 263 individuals) belong to C.(E.) fuscimanus, 54 series (with 330 individuals) to C.(E.) boldorii (for material examined, see appendix). Atone locality, one specimen of each species was found.Our study revealed that none of the diagnostic charactersgiven in BEIER (1934, 1963) allows unequivocal dis-crimination in the species pair boldorii-fuscimanus. Firstof all, the development of the posterior eyes is highlyvariable across both taxa. Digital photography using anextended focus-option illustrates the reduction of theposterior lenses in both species (Figs. 5, 6), includingspecimens from the terra typica of C. (E.) austriacus(Fig. 5). The different development of the posterior eyesmight depend on environmental circumstances in thehabitats (e. g. caves versus ground litter), but variation isobserved even within populations. In accordance withthe suggestion of MAHNERT (1980), these results provethat C. (E.) boldoriiis not a strictly cavernicolous pseu-doscorpion, as supposed by BEIER (1934, 1963). Chaeto-taxy is neither useful. For example, BEIER (1963) men-tions 11 coxal spines on leg II and 4 on leg III in C. (E.)a. austriacus, but 9 on leg II and 5 on leg III in C. (E.) a.boldorii. Our material showed the spination of the coxaeto be a highly variable and frequently asymmetric char-acter which is not of taxonomic significance (see Figs.12, 18). The same is true for the number of setae oneither side of the male genitalic aperture, which rangesfrom 7–9 in both species (Figs. 11, 17), whereas BEIER
(1963) quotes 8 of these setae in C. (E.) a. austriacus.Concerning dentation, BEIER (1934, 1963) correctlyrecognised the tendency of boldorii to bear more teethon TFF than fuscimanus. The present study confirms sig-nificant differences in the number of teeth on the fixedchelal fingers between the two taxa (Tab. 1). However,there is considerable overlapping which prevents taxondelimitation solely by this character. Similarly, the pres-ence of one to two vestigial proximal teeth on the fixed
300 C. MUSTER, T. SCHMARDA and T. BLICK
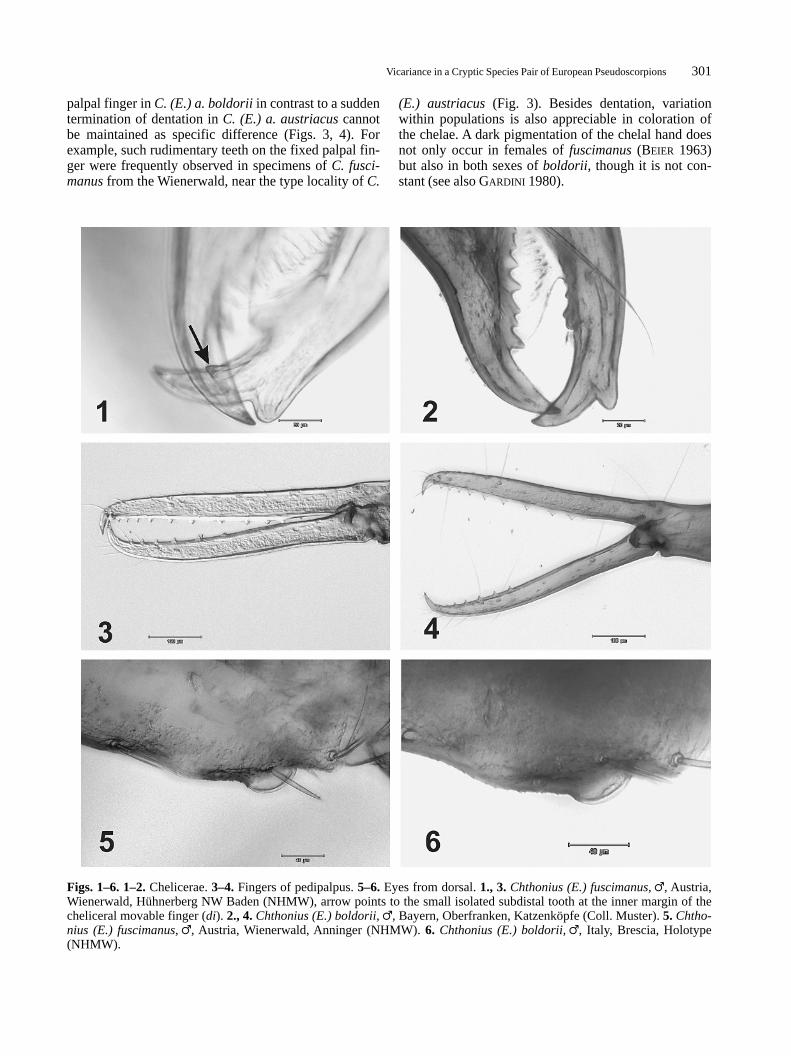
Vicariance in a Cryptic Species Pair of European Pseudoscorpions301
Figs. 1–6. 1–2.Chelicerae. 3–4. Fingers of pedipalpus. 5–6.Eyes from dorsal. 1., 3.Chthonius (E.) fuscimanus, �, Austria,Wienerwald, Hühnerberg NW Baden (NHMW), arrow points to the small isolated subdistal tooth at the inner margin of thecheliceral movable finger (di). 2., 4.Chthonius (E.) boldorii, �, Bayern, Oberfranken, Katzenköpfe (Coll. Muster). 5. Chtho-nius (E.) fuscimanus, �, Austria, Wienerwald, Anninger (NHMW). 6. Chthonius (E.) boldorii, �, Italy, Brescia, Holotype(NHMW).
palpal finger in C. (E.) a. boldoriiin contrast to a suddentermination of dentation in C. (E.) a. austriacuscannotbe maintained as specific difference (Figs. 3, 4). Forexample, such rudimentary teeth on the fixed palpal fin-ger were frequently observed in specimens of C. fusci-manusfrom the Wienerwald, near the type locality of C.
(E.) austriacus(Fig. 3). Besides dentation, variationwithin populations is also appreciable in coloration ofthe chelae. A dark pigmentation of the chelal hand doesnot only occur in females of fuscimanus(BEIER 1963)but also in both sexes of boldorii, though it is not con-stant (see also GARDINI 1980).

In general, there have been few statistical analysesdealing with the validity of measurements and propor-tions of the palps for separating closely related pseudo-scorpion species (e. g. HARVEY 1985, 1987; HEURTAULT
1980). Size differences mentioned in original descrip-tions are often challenged as larger series have beenmeasured. This is the result of our study, too: all mea-surements which were taken show wide-ranging over-lap between C. (E.) boldoriiand fuscimanus(Tab. 1).Thus, the expression of di remains the only individualdiagnostic character for a reliable separation of the twotaxa. Whether or not multivariate analysis of theremaining features can contribute to species delimita-tion is examined in the next section.
3.2. Statistics of numerical characters
All numerical characters which were measured provedto overlap greatly (Tab. 1). Thus, an unequivocal deter-mination of single specimens according to these fea-tures is impossible. Nevertheless, comparison ofmeans revealed highly significant differences betweenthe two taxa in all characters except the carapacelength (2-tailed t-test, equality of variances notassumed; Tab. 1). In both sexes, C. (E.) boldorii tend tohave a smaller carapace and shorter palpal femora than
C. (E.) fuscimanus, whereas the number of teeth onboth palpal fingers is higher in the former species. Thenonparametric Mann-Whitney-U-test resulted in thesame levels of significance (data not shown).Since single numerical characters turned out to beweak discriminators, it was attempted to reach anunequivocal separation of the two taxa by means of amultivariate discriminant analysis. In the first step,this was run simultaneously with the sexes of bothtaxa as separate groups. Using this model, 85.3% ofall cases were correctly classified. The resultant scat-ter plot (Fig. 19) shows that the first discriminantfunction (Eigenvalue 3.803), which explains 82.7%of the total variance, separates males from femalesrather than C. (E.) boldorii from C. (E.) fuscimanus.Function 1 is most strongly correlated with carapacewidth and length of the palpal femur. The second dis-criminant function (Eigenvalue 0.788), which dis-criminates between the two species, is best correlatedwith the number of teeth on the fixed finger of pedi-palp, but explains only 17.1% of the total variance(the third function has no significant influence on themodel). For this reason, additional analyses were runwith males and females separately. In both sexes,carapace length has no significant influence on thediscriminant function. It was omitted in the subse-
302 C. MUSTER, T. SCHMARDA and T. BLICK
Tab. 1.Comparison of numerical characters of Chthonius (E.) fuscimanusand C. (E.) boldorii. Given are arithmetic means ±standard deviation and the extreme values. CL – prosoma length (mm), CW – prosoma width (mm), LF – length of palpalfemur (mm), TFF – number of teeth on fixed finger of pedipalpus, TMF – number of teeth on moveable finger of pedipalpus,** p < 0.01, *** p < 0.001 (2-tailed t-test, equality of variances not assumed).
fuscimanus boldorii t-test
males (n = 66) (n = 133)
CL 0.44 ± 0.017 (0.40–0.48) 0.44 ± 0.026 (0.36–0.48) n. s.CW 0.44 ± 0.021 (0.40–0.50) 0.42 ± 0.023 (0.34–0.48) ***LF 0.60 ± 0.034 (0.54–0.68) 0.57 ± 0.033 (0.46–0.66) ***TFF 10.19 ± 0.59 (9–11) 11.55 ± 0.73 (10–13) ***TMF 5.72 ± 0.51 (5–7) 6.42 ± 0.62 (5–7) ***
females (n = 41) (n = 67)
CL 0.49 ± 0.020 (0.46–0.54) 0.50 ± 0.037 (0.40–0.56) n. s.CW 0.52 ± 0.032 (0.44–0.60) 0.50 ± 0.033 (0.42–0.60) **LF 0.70 ± 0.041 (0.60–0.78) 0.66 ± 0.053 (0.56–0.78) ***TFF 10.29 ± 0.46 (10–11) 11.53 ± 0.88 (10–14) ***TMF 5.56 ± 0.50 (5–6) 6.13 ± 0.74 (5–8) ***
both sexes (n = 107) (n = 200)
CL 0.46 ± 0.031 (0.40–0.54) 0.46 ± 0.041 (0.36–0.56) n. s.CW 0.47 ± 0.046 (0.40–0.60) 0.45 ± 0.046 (0.34–0.60) ***LF 0.64 ± 0.059 (0.54–0.78) 0.60 ± 0.060 (0.46–0.78) ***TFF 10.23 ± 0.54 (9–11) 11.55 ± 0.78 (10–14) ***TMF 5.66 ± 0.51 (5–7) 6.32 ± 0.67 (5–8) ***
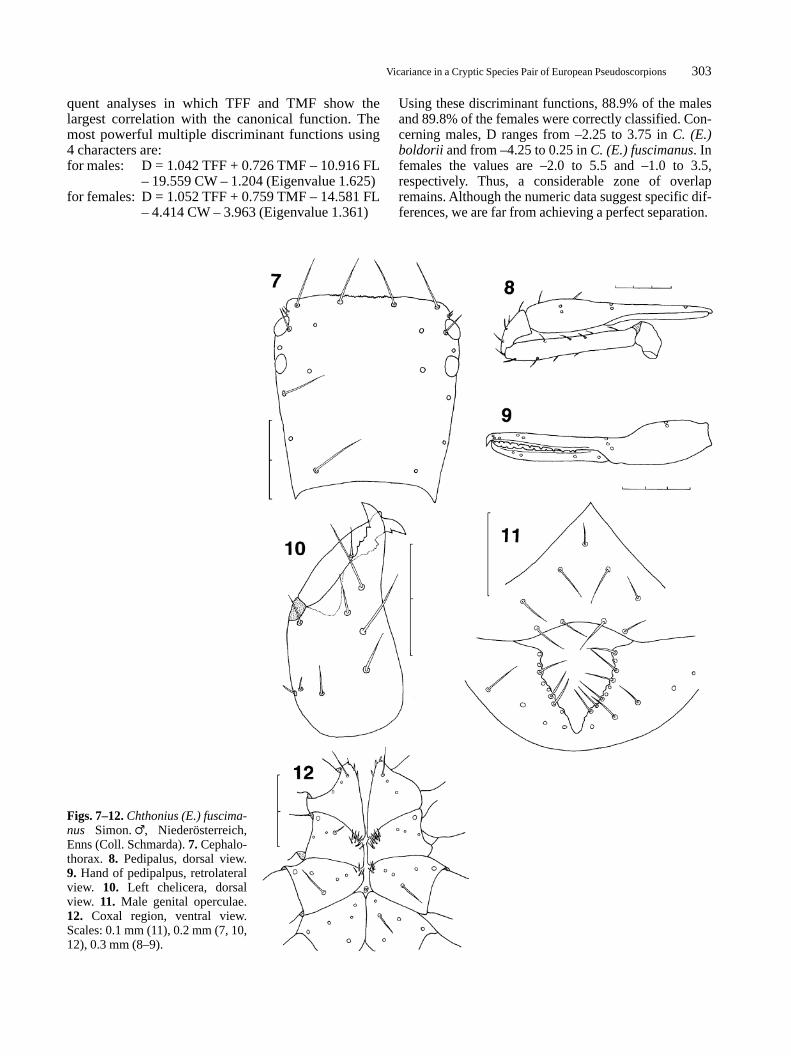
Figs. 7–12.Chthonius (E.) fuscima-nus Simon.�, Niederösterreich,Enns (Coll. Schmarda). 7. Cephalo-thorax. 8. Pedipalus, dorsal view.9. Hand of pedipalpus, retrolateralview. 10. Left chelicera, dorsalview. 11. Male genital operculae.12. Coxal region, ventral view.Scales: 0.1 mm (11), 0.2 mm (7, 10,12), 0.3 mm (8–9).
Vicariance in a Cryptic Species Pair of European Pseudoscorpions303
quent analyses in which TFF and TMF show thelargest correlation with the canonical function. Themost powerful multiple discriminant functions using4 characters are:for males: D = 1.042 TFF + 0.726 TMF – 10.916 FL
– 19.559 CW – 1.204 (Eigenvalue 1.625)for females: D = 1.052 TFF + 0.759 TMF – 14.581 FL
– 4.414 CW – 3.963 (Eigenvalue 1.361)
Using these discriminant functions, 88.9% of the malesand 89.8% of the females were correctly classified. Con-cerning males, D ranges from –2.25 to 3.75 in C. (E.)boldorii and from –4.25 to 0.25 in C. (E.) fuscimanus. Infemales the values are –2.0 to 5.5 and –1.0 to 3.5,respectively. Thus, a considerable zone of overlapremains. Although the numeric data suggest specific dif-ferences, we are far from achieving a perfect separation.

Figs. 13–18.Chthonius (E.) boldoriiBeier.�, Vorarlberg, Spiegelstein(Coll. Schmarda). 13. Cephalo-thorax. 14. Pedipalus, dorsal view.15. Hand of pedipalpus, retrolateralview. 16. Left chelicera, dorsalview. 17. Male genital operculae.18. Coxal region, ventral view.Scales: 0.1 mm (16, 17–18), 0.3 mm(13–15).
304 C. MUSTER, T. SCHMARDA and T. BLICK
3.3. Distribution
One important result of this study is the homogeneity interms of the expression of di within the investigatedpopulations, the other is the large extent of geographicseparation between the two taxa. In central Europenorth of the Alps an almost vicariant distribution patternemerged (Figs. 20, 21). Chthonius (E.) fuscimanusis theeastern species, C. (E.) boldorii the western one. Their
distribution limits roughly correspond to longitude 14°E.Saxony, Moravia and the Austrian counties Lower Aus-tria (incl. Vienna) and Styria (with the exception of thefar west) are exclusively inhabited by C. (E.) fusci-manus, whereas in Bavaria, Switzerland (DEVORE-SCRIBANTE 1999) and the Austrian counties of Vorarl-berg, Northern Tyrol, Salzburg and Carinthia only C.(E.) boldorii occurs. In Upper Austria the ranges over-lap in a small zone of sympatry. Nevertheless, only a
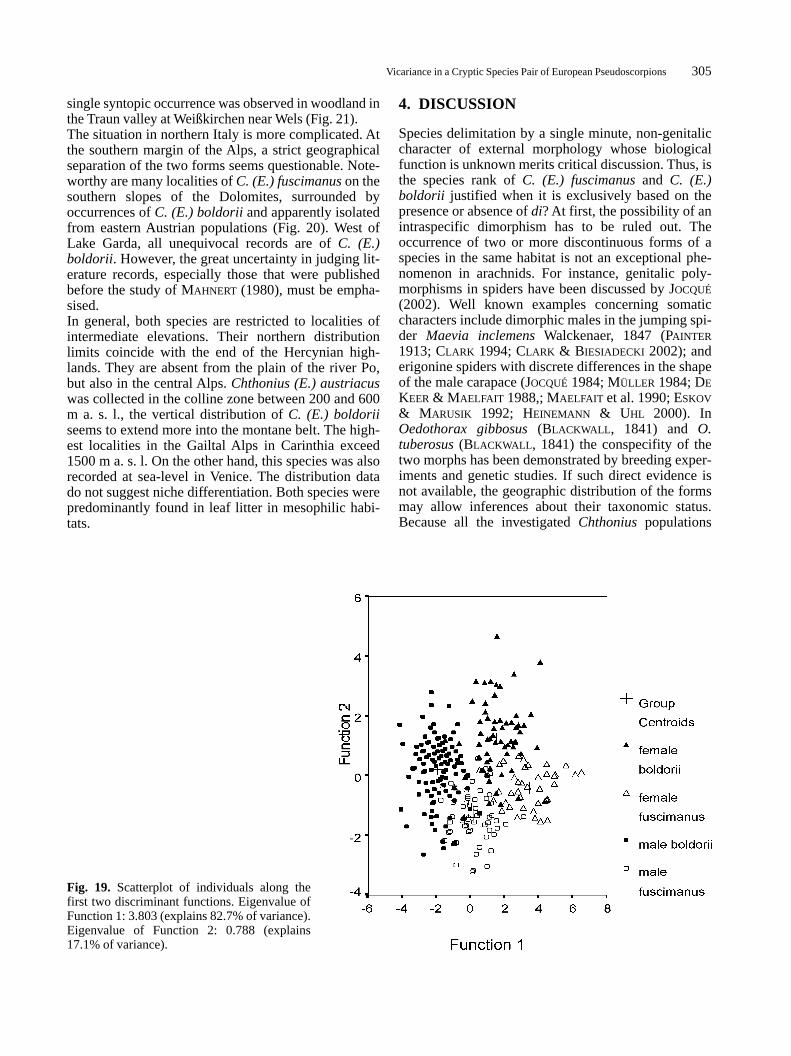
Fig. 19. Scatterplot of individuals along thefirst two discriminant functions. Eigenvalue ofFunction 1: 3.803 (explains 82.7% of variance).Eigenvalue of Function 2: 0.788 (explains17.1% of variance).
Vicariance in a Cryptic Species Pair of European Pseudoscorpions305
single syntopic occurrence was observed in woodland inthe Traun valley at Weißkirchen near Wels (Fig. 21).The situation in northern Italy is more complicated. Atthe southern margin of the Alps, a strict geographicalseparation of the two forms seems questionable. Note-worthy are many localities of C. (E.) fuscimanuson thesouthern slopes of the Dolomites, surrounded byoccurrences of C. (E.) boldoriiand apparently isolatedfrom eastern Austrian populations (Fig. 20). West ofLake Garda, all unequivocal records are of C. (E.)boldorii. However, the great uncertainty in judging lit-erature records, especially those that were publishedbefore the study of MAHNERT (1980), must be empha-sised.In general, both species are restricted to localities ofintermediate elevations. Their northern distributionlimits coincide with the end of the Hercynian high-lands. They are absent from the plain of the river Po,but also in the central Alps. Chthonius (E.) austriacuswas collected in the colline zone between 200 and 600m a. s. l., the vertical distribution of C. (E.) boldoriiseems to extend more into the montane belt. The high-est localities in the Gailtal Alps in Carinthia exceed1500 m a. s. l. On the other hand, this species was alsorecorded at sea-level in Venice. The distribution datado not suggest niche differentiation. Both species werepredominantly found in leaf litter in mesophilic habi-tats.
4. DISCUSSION
Species delimitation by a single minute, non-genitaliccharacter of external morphology whose biologicalfunction is unknown merits critical discussion. Thus, isthe species rank of C. (E.) fuscimanus and C. (E.)boldorii justified when it is exclusively based on thepresence or absence of di? At first, the possibility of anintraspecific dimorphism has to be ruled out. Theoccurrence of two or more discontinuous forms of aspecies in the same habitat is not an exceptional phe-nomenon in arachnids. For instance, genitalic poly-morphisms in spiders have been discussed by JOCQUÉ
(2002). Well known examples concerning somaticcharacters include dimorphic males in the jumping spi-der Maevia inclemensWalckenaer, 1847 (PAINTER
1913; CLARK 1994; CLARK & BIESIADECKI 2002); anderigonine spiders with discrete differences in the shapeof the male carapace (JOCQUÉ1984; MÜLLER 1984; DE
KEER& M AELFAIT 1988,; MAELFAIT et al. 1990; ESKOV
& M ARUSIK 1992; HEINEMANN & UHL 2000). InOedothorax gibbosus (BLACKWALL , 1841) and O.tuberosus(BLACKWALL , 1841) the conspecifity of thetwo morphs has been demonstrated by breeding exper-iments and genetic studies. If such direct evidence isnot available, the geographic distribution of the formsmay allow inferences about their taxonomic status.Because all the investigated Chthonius populations

306 C. MUSTER, T. SCHMARDA and T. BLICK
Fig. 20.Distribution of Chthonius (E.) boldorii(filled circles) and C. (E.) fuscimanus(open circles) in Europe, according tothe material examined and GARDINI (2000) (Italy), DEVORE-SCRIBANTE (1999) (Switzerland), MAHNERT (1980) (Croatia),STREBEL (1961) (Germany, Siebengebirge), KRUMPÁL & K IEFER (1981) (Czech Republik), RAFALSKI (1967) (Poland). Ques-tion marks signify literature records which can not unambiguously assigned to either species.
Fig. 21. Details of distribution of Chthonius (E.) boldoriiand C. (E.) fuscimanus in their contact zone in eastern Austria. Thearrow points to a single locality of syntopic occurrence at Weißkirchen near Wels.

ons. Recent research into a cryptic neotropical pseu-doscorpion species complex, currently described as asingle species Cordylochernes scorpioides (Linnaeus,1758), revealed extensive genetic divergence in theabsence of appreciable morphological differentiation(ZEH & ZEH 1994; WILCOX et al. 1997). Sequencedivergence in the mitochondrial COI gene ranges from2.6 to 13.8% between geographic “populations”(which corresponds to divergence values foundbetween morphologically distinct, congeneric speciesin other terrestrial arthropods), whereas intrapopula-tion variability is very low. In this cited case, postzy-gotic reproductive incompatibility between the phylo-geographic lineages has been demonstrated (WILCOX
et al. 1997), thus demonstrating the presence of multi-ple biological species. However, one has to be cautiousto extrapolate this situation to primitive pseudoscorpi-ons like Chthonius, as Cordylochernesshows a com-plex biology involving pairing between males andfemales and strong competition between males.A convincing explanation for the diversification anddistribution pattern in the cryptic complex of Chtho-nius boldorii/fuscimanushardly seems feasible with-out the perspective of historical biogeography. Boththese ecologically similar species are arboreal faunalelements, inhabiting leaf litter. Physical, pollen andfossil data of the climatic changes in Europe stronglysuggest that woodland communities were restricted torefugial areas in southern Europe during glacial cycles(DE LATTIN 1967; WEBB & BARTLEIN 1992). Moleculardata from recent phylogeographic studies confirmsouthern peninsulas of Europe as major glacial refugiafor arboreal elements, and they allow the reconstruc-tion of postglacial colonization processes towards thenorth (HEWITT 1999). In the Chthoniusexample, atleast those localities north of the Alps would have beencolonized in postglacial times. A plausible scenario isthat C. (E.) boldorii survived in one or more Italianrefugia, C. (E.) fuscimanusin the Balkans. In UpperAustria we face a zone of secondary contact of thediverged genomes. Whether reproductive isolation iscomplete or gene-sharing by introgression occurs can-not be assessed without karyological or moleculardata. Hybridization often occurs where divergedgenomes met after the last ice age; the suture zones areoften narrow and remarkably stable in their positionover time (HEWITT 2001). The observed clustering ofhybrid zones in multiple taxa results from commonali-ties of refugia, rate of climate warming and barriers toexpansion (HEWITT 1999). REMINGTON (1968) hadalready speculated that such a suture zone might existin central Europe. Indeed, the parapatric pattern in theEuropean hedgehogs Erinaceus europaeus Linnaeus,1758 andE. concolorMartin, 1838 resembles the dis-tribution of Chthonius (E.) boldorii/fuscimanusin
were monomorphic with respect to the expression of di(the only exception is a locality at the limits of theirdistribution), and the two discrete forms are geographi-cally separated in large parts of their ranges, intraspe-cific polymorphism can be ruled out in this instance.Furthermore, the absence of intermediate phenotypesand the negatively associated distributions indicate thatthe two forms do not loose their divergence upon con-tact and that they will be able to continue to diverge,thus fulfilling the criteria of the genic species concept(WU 2001). In conclusion, the species rank of C. (E.)boldorii and C. (E.) fuscimanusseems well justified.But what are the driving forces behind the diversifica-tion of these species? The extraordinary diversity inmost arthropod taxa is thought to be the result of rapidand divergent genitalic evolution due to sexual selec-tion by female choice (EBERHARD 1985). While thegenitalia in the more highly developed pseudoscorpionfamilies are useful as taxonomic tools, they are of lim-ited use in the Chthoniidae (LEGG 1987). Indeed, thecomparison of the genitalia of C. (E.) boldoriiand C.(E.) fuscimanusdid not reveal any differences. This isnot really surprising, since pseudoscorpions producespermatophores. Because in Chthoniidae the sper-matophores are deposited by males without direct con-tact with females (WEYGOLDT 1966), the male genitaliacan neither function as internal courtship devices (asfemale choice hypothesis requires) nor can di be inter-preted as a secondary sexual structure. However, LEGG
(1973, 1975) proposed that the secretion which accu-mulates as a droplet on the thickened portion of thespermatophore stalk may contain pheromones that per-mit specific recognition of spermatophores by conspe-cific females. Thus, speciation genes (TING et al. 2000)in pseudoscorpions may affect biochemical rather thanmorphological traits. An explanation for the taxonomicvalue of di (which is present in many species of Ephip-piochthoniusand absent in some others) despite itsdoubtful selective function would be cohesivenesswith the unknown genes responsible for reproductiveisolation. This assumption implies, however, that a loss(or gain) of di has occurred in one species of the pairboldorii-fuscimanusindependently of losses (or gains)in other species of Ephippiochthonius. An argumentfor such a phylogenetic inference is the similar patternof presence or absence of di in the subgenus Chthonius(GABBUTT & VACHON 1963).The example of Chthonius (E.) boldorii/fuscimanusillustrates both, the fact that ‘cryptic’ species are oftenthose that have been studied superficially in terms oftheir morphology, but also the limits of distinguishingspecies through morphological characters. The view ofHEDIN (1997) that “species as morphological lineagesare currently more inclusive than basal evolutionary orphylogenetic units” may well apply to pseudoscorpi-
Vicariance in a Cryptic Species Pair of European Pseudoscorpions307

some aspects. The areas of these congeners show limit-ed overlap, meeting at approximately 15° E, with littlesuggestion of interbreeding in the wild (CORBET1988).In this case, the proposed northward expansion from,respectively, Italian and Balkan refugia has beenproved by DNA markers (SANTUCCI et al. 1998; SED-DON et al. 2001). An alternative explanation for theexistence of a narrow contact zone between the rangesof two almost vicariant congeners would be competi-tive exclusion. BROWN & L OMOLINO (1998) argued thatin the absence of direct experiments, a pattern of mutu-ally exclusive distributions of two or more species withsimilar resource requirements can serve as natural evi-dence for competition. In this respect, substitution pat-terns in other congeneric arachnid species pairs in theAlps were discussed by MUSTER (2002). An unre-solved problem remains the putative checkerboard dis-tribution (DIAMOND 1975) of C. (E.) boldorii/fusci-manusat the southern margin of the Alps. In general,haplotype diversity within species and species com-plexes of other groups is remarkably higher in southernEurope in contrast to northward expansions, probablyas a result of refugial persistence and accumulation ofvariation over several glacial cycles (HEWITT 2001).There is reason to assume that discordance betweengenetic and morphological distances, as observed insome pseudoscorpions (WILCOX et al. 1997), may haveobscured further deep phylogenetic splits.
Acknowledgements.We are indebted to the following col-legues for access to material in their care: V. Duchácˇ (HradecKrálové), J. Gruber (NHMW, Vienna), K. Lippold (Leipzig),W. Schawaller (SMNS, Stuttgart). We are grateful to V. Mah-nert (Geneva) for valuable information, and to J. Spelda(Stuttgart) for access to unpublished data. Warm thanks aredue to S. Engler (Dresden) for producing a map, to M. Nuss(Dresden) for assistance with digital photography, and to J.Dunlop (Berlin) for linguistic improvement. The helpfulcomments of two anonymous referees are gratefullyacknowledged.
REFERENCES
BEIER, M. (1934): Neue cavernicole und subterrane Pseu-doscorpione. Mitt. Höhlen- und Karstforschung 1934:53–59.
BEIER, M. (1963): Ordnung Pseudoscorpionidea (Afterskor-pione). Bestimmungsbücher zur Bodenfauna Europas.Vol. 1. VI + 313 pp., Akademie-Verlag, Berlin.
BLICK, T. & MUSTER, C. (2003): Rote Liste der Pseudoskor-pione Bayerns (1. Fassung) (Arachnida: Pseudoscor-piones). SchrR. Bayer. Landesamt f. Umweltschutz, inpress.
BLICK, T., HÄNGGI, A. & THALER, K. (2002): Checklist of thearachnids of Germany, Switzerland, Austria, Belgium andthe Netherlands (Arachnida: Araneae, Opiliones, Pseudo-
scorpiones, Scorpiones, Palpigradi). Version 2002 June 1.Internet: http://www.AraGes.de/checklist_e.html.
BROWN, J. H. & LOMOLINO, M. V. (1998): Biogeography. 2nd
edition. XII + 692 pp., Sinauer Associates, Sunderland,Massachusetts.
CLARK, D. L. (1994): Sequence ananlysis of courtshipbehaviour in the dimorphic jumping spider, Maeviainclemens. J. Arachnol. 22: 94.107.
CLARK, D. L. & BIESIADECKI, B. (2002): Mating success andalternative reproductive strategies of the dimorphic jump-ing spider, Maevia inclemens(Araneae, Salticidae). J.Arachnol. 30: 511–518.
CORBET, G. B. (1988): The family Erinaceidae: a synthesis ofits taxonomy, phylogeny, ecology and zoogeography.Mamm. Rev. 18: 117–172.
COTTI, G. (1989): Contributo alla connoscenza della faunageobia del Monte Generoso (Canton Ticino, Svizzera).Bol. Soc. Ticinese Sci Nat. 77: 13–34.
DE KEER, R. & MAELFAIT, J. P. (1988): Oedothorax gibbosus(Blackwall) and Oedothorax tuberosus (Blackwall): onespecies. Newsletter Br. arachnol. Soc. 53: 3.
DE LATTIN, G. (1967): Grundriss der Zoogeographie. 602 pp.Fischer, Jena.
DEVORE-SCRIBANTE, A. (1999): Les pseudoscorpions de laSuisse. Etude systématique, faunistique et biogéographi-que. III + 314 pp., Thesis, University of Geneva.
DIAMOND, J. M. (1975): Assembly of species communities.Pp. 342–444 in: CODY, M. L. & DIAMOND, J. M. (eds.)Ecology and Evolution of Communities. Belknap, Cam-bridge, Massachusetts.
EBERHARD, W. G. (1985): Sexual selection and animal geni-talia. 231 pp. Harvard University Press, Cambridge, Massa-chusetts.
ESKOV, K. Y. & MARUSIK, Y. M. (1992): On the Siberio-Nearctic erigonine spider genus Silometopoides(Aranei-da: Linyphiidae). Reichenbachia 29: 97–103.
GABBUTT, P. D. & VACHON, M. (1963): The external mor-phology and life history of the pseudoscorpion Chthoniusischnocheles (Hermann). Proc. Zool. Soc. London 140:75–98.
GARDINI, G. (1980): Identita’ di Chthonius tetrachelatus fus-cimanusSimon, 1900 e ridescrizione di C. (E.) nanusBeier, 1953. (Pseudoscorpionida Chthoniidae) (Pseudo-scorpioni d’ Italia XI). Ann. Mus. Civ. Stor. Nat. Genova88: 261–270.
GARDINI, G. (2000): Catalogo degli Pseudoscorpioni d’Italia.Fragmenta entomol. 32, Supplemento: 1–181.
HARVEY, M. S. (1985): The systematics of the family Sterno-phoridae (Pseudoscorpionida). J. Arachnol. 13: 141–209.
HARVEY, M. S. (1987): A revision of the genus SynsphyronusChamberlin (Garypidae: Pseudoscorpionida: Arachnida).Australian J. Zool. (suppl. ser.) 126: 1–99.
HARVEY, M. S. (1991): Catalogue of the Pseudoscorpionida.726 pp., Manchester University Press, Manchester & NewYork.
HEDIN, M. C. (1997): Molecular phylogenetics at the popula-tion species interface in cave spiders of the southernAppalachians (Araneae: Nesticidae: Nesticus). Mol. Biol.Evol. 14: 309–324.
308 C. MUSTER, T. SCHMARDA and T. BLICK

HEINEMANN, S. & UHL, G. (2000): Male dimorphism inOedothorax gibbosus(Araneae, Linyphiidae): a morpho-metric analysis. J. Arachnol. 28: 23–28.
HEURTAULT, J. (1980): Quelques remarques sur les espècesfrançaises du genre Rhacochelijer Beier (Arachnides,Pseudoscorpions, Cheliferidae). Bull. Mus. Natl. Hist.Nat. Sect. A2: 161–173.
HEWITT, G. M. (1999): Post-glacial re-colonization of Euro-pean biota. Biol. J. Linn. Soc. 68: 87–112.
HEWITT, G. M. (2001): Speciation, hybrid zones and phylo-geography – or seeing genes in space and time. Mol. Ecol.10: 537–549.
JOCQUÉ, R. (1984): Linyphiidae (Araneae) from SouthAfrica. Part I: The collection of the Plant ProtectionResearch Institute. J. entomol. Soc. South Africa 47:121–146.
JOCQUÉ, R. (2002): Genitalic polymorphism – a challenge fortaxonomy. J. Arachnol. 30: 298–306.
KRUMPÁL, M. & KIEFER, M. (1981): Príspevok k poznanuiStúrikov cel’ade Chthoniidae v CˇSSR (Pseudoscorpioni-dea). Zp. Cˇs. spol. entom. 17: 127–130.
LEGG, G. (1973): Spermatophore formation in the pseu-doscorpion Chthonius ischnocheles (Chthoniidae). J.Zool., Lond. 170: 367–394.
LEGG. G. (1975): The genitalia and associated glands of fiveBritish species belonging to the family Chthoniidae (Pseu-doscorpiones: Arachnida). J. Zool., Lond. 177: 99–121.
LEGG, G. (1987): Proposed taxonomic changes to the Britishpseudoscorpion fauna (Arachnida). Bull. Br. arachnol.Soc. 7: 179–182.
MAELFAIT, J. P., DE KEER, R. & DE MEESTER, L. (1990):Genetical background of the polymorphism of Oedotho-rax gibbosus(Blackwall) (Linyphiidae, Araneae). Rev.arachnologique 9: 29–34.
MAHNERT, V. (1980): Höhlenpseudoskorpione aus Nordital-ien und der dalmatinischen Insel Krk. Atti e Memorie dellaCommissione Grotte “E. Bogan” 20: 95–100.
MÜLLER, H. G. (1984): Zur Variabilität der Kopfformen beiZwergspinnen-Männchen. Über Diplocephalus latifrons“fallaciosus” (Bertkau 1883) und Troxochrus scabriculus“cirrifrons” (Cambridge1871) (Araneae: Araneida: Liny-phiidae). Entomol. Z. 94: 337–352.
MUSTER, C. (2002): Substitution patterns in congenericarachnid species in the northern Alps. Diversity and Distri-butions 8: 107–121.
PAINTER, T. S. (1913): On the dimorphism of the males ofMaevia vittata. Zool. Jb. 37: 625–636.
RAFALSKI, J. (1967): Zaleszczotki. Pseudoscorpionidea. Kata-log Fauny Polski 32: 1–34.
REMINGTON, C. L. (1968): Suture-zones of hybrid interactionbetween recently joined biotas. Evolutionary Biol. 2:321–428.
SANTUCCI, F., EMERSON, B. C. & HEWITT, G. M. (1998):Mitochondrial DNA phylogeography of European hedge-hogs. Mol. Ecol. 7: 1163–1172.
SCHMARDA, T. (1995): Beiträge zur Kenntnis der Pseudoskor-pione von Tirol und Vorarlberg: Faunistik; taxonomischeCharakterisierung; Aktivitätsdynamik. 85 pp. + appendix,Diplomarbeit, University of Innsbruck.
SEDDON, J. M., SANTUCCI, F., REEVE, N. J. & HEWITT, G. M.(2001): DNA footprints of European hedgehogs, Eri-naceus europaeusand E. concolor: Pleistocene refugia,postglacial expansion and colonization routes. Mol. Ecol.10: 2187–2198.
STREBEL, O. (1967): Pseudoscorpiones aus dem Siebenge-birge. Decheniana-Beihefte 9: 107–108.
TING, C.-T., TSAUR, S.-C. & WU, C.-I. (2000): The phyloge-ny of closely related species as revealed by the genealogyof a speciation gene, Odysseus. Proc. Natl. Acad. Sci. 97:5313–5316.
WEBB, T. & BARTLEIN, P. J. (1992): Global changes duringthe last 3 million years: climatic controls and bioticresponses. Ann. Rev. Ecol. Syst. 23: 141–173.
WEYGOLDT, P. (1966): Vergleichende Untersuchungen zurFortpflanzungsbiologie der Pseudoscorpione. Beobach-tungen über das Verhalten, die Samenübertragungsweisenund die Spermatophoren einiger einheimischer Arten. Z.Morphol. Ökol. Tiere 56: 39–92.
WILCOX, T. P., HUGG, L., ZEH, J. A. & ZEH, D. W. (1997):Mitochondrial DNA sequencing reveals extreme geneticdifferentiation in a cryptic species complex of neotropicalpseudoscorpions. Mol. Phylogen. Evol. 7: 208–216.
WU, C.-I. (2001): The genic view of the process of specia-tion. J. Evol. Biol. 14: 851–865.
ZEH, D. W. & ZEH, J. A. (1994): When morphology misleads:interpopulation uniformity in sexual selection masksgenetic divergence in harlequin beetle-riding pseudoscor-pion populations. Evolution 48: 1168–1182.
Authors’ addresses:Dr. Christoph MUSTER (correspondingauthor), Staatliches Museum für Tierkunde, KönigsbrückerLandstraße 159, D-01109 Dresden, Germany;e-mail: [email protected];Thomas SCHMARDA, Universität Innsbruck, Institut fürZoologie und Limnologie, Technikerstraße 25, A-6020 Inns-bruck, Austria;Theo BLICK, Heidloh 8, D-95503 Hummeltal, Germany
Received: 27. 03. 2003Returned for revision: 05. 06. 2003Accepted: 15. 07. 2003Corresponding Editor: G. A. BOXSHALL
Vicariance in a Cryptic Species Pair of European Pseudoscorpions309

APPENDIX
Chthonius (Ephippiochthonius) boldorii Beier(Figs. 2, 4, 6, 13–18)Chthonius (Ephippiochthonius) austriacus boldoriiBeier, 1934Chthonius (E.) boldoriiBeier: Mahnert, 1980
Material examined:Germany: Bayern: Oberfranken, O Leutenbach (Lkr.Forchheim), Katzenköpfe, 49° 43′ N, 11° 10′ E, 400–520 m,20�, 12� (BF Jun-Okt 2000, leg. Blick & Fritze, Coll.Muster); Eggolsheim, Kautschenberg zw. Götzendorf undStackendorf, 49° 49′ N, 11° 6′ E, 450–500 m, 7�, 5� (BF1990/1991, leg. Fritze, Coll. Muster); Oberbayern, Freising,window trap in Quercus rubra, 48° 25′ N, 11° 44′ E, 1�(20.10.-20.11.2000, leg. Gossner, Coll. Muster); Berchtes-gaden, 47° 38′ N, 13° 00′ E, 600 m, 13�, 4� (17.07.1995),38�, 16� (01.06.1996), 12�, 5� (01.06.1999, all leg. &Coll. Lippold 1915/1928); NW Kiefersfelden, 47° 37′ N, 12°11′ E, 500 m, 3� (07.07.1986, leg. Schawaller, SMNS1623).Baden Württemberg:Friedrichshafen, 47° 40′ N, 09° 30′ E,4 �, 4� (01.05.1996, leg. Lippold, Coll. Lippold 1925); SWBerau, 47° 42′ N, 8° 15′ E, 1� 1 � (18.06.1993, leg. Konzel-mann, SMNS 3240); Hegau, Engen, 47° 45′ N, 8° 46′ E, 1�(10.05.1980, leg. Hahn, SMNS 339); Südschwarzwald, valleynear Wyhlen, 47° 33′ N, 7° 41′ E, 4� (26.04.1969, leg. Wey-goldt, SMNS 2975), 4� 1 � (02.08.1969, leg. Weygoldt,SMNS 2999); S Stuttgart, “Falsche Klinge”, 47° 46′ N, 9° 11′ E,11�, 15� (01.09.1987, leg. Kostenbader, SMNS 3657).Rheinland-Pfalz:Soonwald, Gräfenbachtal near Opelwiese,49° 55′ N, 7° 39′ E, 2�, 1� (22.05.1979, leg. Schawaller,SMNS 183).Hessen: Taunus, Schlangenbad, 50° 05′ N, 8° 06′ E, 1�(18.07.1978, leg. Schawaller, SMNS 22).Austria: Vorarlberg: Göfis i. Walgau, Spiegelstein, 47° 15′N, 09° 38′ E, 1� (27.07.1991, leg. Breuss, Coll. Schmarda);Übersaxen, Baschghöhle, 47° 16′ N, 09° 41′ E, 1� (13.07.1991,leg. Breuss, Coll. Schmarda); Hohenems (Bodnerhaus),47° 23′ N, 09° 41′ E, 780 m, 1� (28.11.1948, leg. ?,NHMW); Doren (Holzberg?), 47° 30′ N, 09° 53′ E, 500 m, 1�,leg. ?, NHMW).Tirol: Zahmer Kaiser, Schanz bei Ebbs, 47° 36′ N, 12° 12′ E,600 m, 1� (07.05.1994, leg. Thaler).Salzburg:W Matzing am Obertrumersee, 47° 58′ N, 13° 04′E, 2�, 1� (18.07.1960, leg. Strouhal, NHMW); Untersberg,SW Fürstenbrunn, 47° 45′ N, 12° 58′ E, 520–530 m, 2�, 3�(1948, leg. Strouhal, NHMW); Untersberg, Graben ONOVeitlbruch, 47° 45′ N, 12° 58′ E, 600 m, 1�, 3� (1948, leg.Strouhal, NHMW); Untersberg, Wildbachufer SW St. Leon-hard, 47° 44′ N, 13° 02′ E, 4� (1948, leg. Strouhal,NHMW); Hinterwinkl SW Ebenau, 47° 47′ N, 13° 10′ E,560 m, 3�, 6� (1948, leg. Strouhal, NHMW); Wiestal 6 kmNNE Hallein, 47° 44′ N, 13° 09′ E, 500 m, 6�, 2� (1948,leg. Strouhal, NHMW); NNE Hallein, 47° 42′ N, 13° 7′ E,500 m, 11�, 1� (1948, leg. Strouhal, NHMW); SSWScharfling, 47° 47′ N, 13° 22′ E, 490 m, 4�, 1�(01.01.1948, leg. Strouhal, NHMW); Tennengebirge,Wassergraben NE Werfen, 47° 29′ N, 13° 12′ E, 1 juv.(01.01.1948, leg. Strouhal, NHMW).
Oberösterreich:Graben zwischen Grein/Donau und Dim-bach, 48° 17′ N, 14° 53′ E, 7�, 5� (1948, leg. Strouhal,NHMW); Traun bei Weißkirchen, Auenwald, 48° 10′ N,14° 08′ E, 300 m, 1� (05.09.1948, leg. Strouhal, NHMW);N Schönau (N Bad Schallerbach), 48° 15′ N, 13° 55′ E,430 m, 10�, 4� (30.08.1948, leg. Strouhal, NHMW); NGmunden, rechtes Traunufer, 47° 56′ N, 13° 48′ E, 415 m,1 � (11.08.1955, leg. Strouhal, NHMW); Gmundnerberg, WAltmünster am Traunsee, 47° 54′ N, 13° 44′ E, 550 m, 4�,1 � (22.07.1949, leg. Strouhal, NHMW); SSO Traunkirchen,Schindlgraben, 47° 51′ N, 13° 47′ E, 5�, 1� (17.07.1949,leg. Strouhal, NHMW); E Rindbach am Traunsee, Rindbach-tal, 47° 48′ N, 13° 50′ E, 550 m, 1� (22.07.1949, leg.Strouhal, NHMW); Offenseebach-Tal S Gmundnersee,47° 46′ N, 13° 46′ E, 590 m, 3�, 1� (17.07.1949, leg.Strouhal, NHMW); ESE Bad Ischl, Rettenbachgraben,47° 42′ N, 13° 40′ E, 470–620 m, 4� (01.08.1955, leg.Strouhal, NHMW); S Bad Ischl, alte Römerstraße, 47° 42′ N,13° 40′ E, 1� (08.08.1955, leg. Strouhal, NHMW); zw.Ischl-Ahorn und Pfandl, 47° 43′ N, 13° 36′ E, 520 m, 1�(31.07.1955, leg. Strouhal, NHMW); NNE Obertraun, S-Ende des Koppentales, 47° 34′ N, 13° 43′ E, 520 m, 3�(04.08.1952, leg. Strouhal, NHMW); Leogang nearSaalfelden, Birnbachtal, 47° 29′ N, 12° 45′ E, 3�(08.06.1988, leg. Buck, SMNS 2690); Schüttelbachgrabennear Saalfelden, 47° 26′ N, 12° 47′ E, 1� (09.06.1988, leg.Buck, SMNS 2692).Steiermark: Ober-Steiermark, Bad Aussee, Obertresen,47° 38′ N, 13° 47′ E, 680 m, 4�, 1� (01.01.1948, leg.Strouhal, NHMW).Kärnten: Gößwing-Weißbriach, 46° 41′ N, 13° 16′ E, 850 m,1 � (1948, leg. Schweiger, NHMW); Gailtaler Alpen,Spitzkogel, 46° 39′ N, 13° 25′ E, 1250–1500 m, 2� (1948,leg. Schweiger, NHMW); Gailtaler Alpen, Schlucht zw. Vel-lacher Egel und Graslitzen, 46° 40′ N, 13° 27′ E, 800–1700 m,4 �, 3� (1948, leg. Schweiger, NHMW); Villacher Alpe(Prungart), 46° 37′ N, 13° 41′ E, 1� (1948, leg. Schweiger,NHMW); Warmbach Villach, 46° 36′ N, 13° 50′ E, 1�, 1�(01.01.1948, leg. Strouhal, NHMW); O Sekirn, 46° 37′ N,14° 13′ E, 7� (1948, leg. Strouhal, NHMW); Maria Wörth,46° 37′ N, 14° 10′ E, 440 m, 2� (01.09.1976, leg. Röder,SMNS 144).Italy: Prov. Brescia:Brescia, Il Baratzo di Monte Bude-flone, 45° 33′ N, 10° 13′ E, 1�, (17.03.1929, NHMW,TYPE of Chthonius (Ephippiochthonius) austriacus boldoriiBeier, 1934); Franciacorta, Camignone dint. Bura de SanFaustii, 45° 38′ N, 10° 15′ E, 1� (20.08.1981, leg. Inzaghi,NHMW).Prov. Trentino:Riva del Garda, 45° 53′ N, 10° 49′ E, 1�, 1�(11.04.1962, leg. Thaler, NHMW).Prov. Veneto:Recoaro, 45° 43′ N, 11° 13′ E, 750 m, 1�, 1�(date?, leg. ?, NHMW).Croatia: Istria: SO Cerovlje, 45° 17′ N, 13° 55′ E, 1�(01.01.1948, leg. Strouhal, NHMW).
Chthonius (Ephippiochthonius) fuscimanusSimon (Figs.1, 3, 5, 7–12)Chthonius tetrachelatus fuscimanusSimon, 1900Chthonius (Ephippiochthonius) austriacusBeier, 1931Chthonius (E.) austriacus austiacusBeier, 1963Chthonius (E.) fuscimanusE. Simon: Harvey, 1991
310 C. MUSTER, T. SCHMARDA and T. BLICK

Note on nomenclature: GARDINI (1980) detected the syn-onymy of Chthonius tetrachelatus fuscimanus Simon withChthonius (E.) austriacusBeier. Because the name fusci-manusSimon had not been mentioned in literature for morethan 80 years, he proposed regarding it a nomen oblitum.However, according to article 23.9 of the ICZN, such a deci-sion is only permitted if, among other requirements, the oldersynonym has not been used since 1899. This is obviously nottrue, therefore HARVEY (1991) is right to list C. (E.) fusci-manusas valid name. The current use of C. (E.) austriacus, e.g. in some European checklists (BLICK et al. 2002), should beavoided. Otherwise a proposal must be submitted to the ICZN.
Material examined:Germany: Sachsen:Seußlitz, 51° 15′ N, 13° 26′ E, 130 m,4 �, 4� (11.05.1990, leg. Lippold, Coll. Lippold 1853); Säch-sische Schweiz, Schmilka, 50° 54′ N, 14° 16′ E, 240 m, 1�(02.06.1982, leg. Lippold, Coll. Lippold 822), 4�, 2�(15.07.1987, leg. Lippold, Coll. Lippold 1513); Gr. WinterbergS Kuppe, 50° 54′ N, 14° 14′ E, 420–490 m, 2� (20.11.1972,leg. Krause, MTD), 1� (22.08.1973, leg. Krause, MTD).Czech Republik: Moravia: Skalni Mlyn, 49° 21′ N, 16° 45′ E,2 � (21.05.1993, leg. Ruzicka, Coll. Muster); Prasecˇi ucho,49° 21′ N, 16° 45′ E, 2� (13.05.1995, leg. Duchac, Coll.Muster).Austria: Niederösterreich/Wien:Wienerwald, Waldlgrabenzw. Leopoldsberg und Kahlenberg, 48° 17′ N, 16° 20′ E, 1�(1948, leg. Strouhal, NHMW); Kahlengebirge, Sauberg,Graben , 48° 16′ N, 16° 17′ E, 300 m, 1�, 1� (09.05.1948,leg. Strouhal, NHMW); Marbachtal bei Kierling, 48° 19′ N,16° 17′ E, 350 m, 15�, 5� (27.06.1948, leg. Strouhal,NHMW); Marbachtal nördl. Kierling, 48° 19′ N, 16° 17′ E,270 m, 6�, 3� (04.05.1952, leg. Strouhal, NHMW); Höfleina. d. Donau, S Graben, 48° 21′ N, 16° 16′ E, 300 m, 5�, 5�(27.06.1948, leg. Strouhal, NHMW); Eschengraben NEAnninger, Eschenbrünnl, 48° 04′ N, 16° 16′ E, 580 m, 6�, 1�(1948, leg. Strouhal, NHMW); NW Gugging, 48° 19′ N,16° 15′ E, 1� (13.06.1948, leg. Strouhal, NHMW); Weid-lingsbach, Domgraben, 48° 16′ N, 16° 15′ E, 1� (25.06.1967,leg. Kritscher, NHMW); N Hadersdorf, Hochbruckenberg SE-Hang, Buchenfallaub an Wassergraben, 48° 15′ N, 16° 14′ E,3 �, 3� (01.09.1946, leg. Strouhal, NHMW); PlökingN-Hang, NE Hintersdorf, 48° 18′ N, 16° 13′ E, 3�, 1�(13.06.1946, leg. Strouhal, NHMW); Mittleres Wasser-gspreng, 48° 05′ N, 16° 13′ E, 1�, 1� (1948, leg. Strouhal,NHMW); Tal SW Hühnerberg (NW Baden), 48° 02′ N, 16° 13′E, 10�, 13� (1948, leg. Strouhal, NHMW); Abflußgrabender Wienerquelle (N Kaltenleutgeben), 48° 08′ N, 16° 12′ E,330 m, 4�, 1� (06.06.1948, leg. Strouhal, NHMW); Baun-zen, 48° 11′ N, 16° 11′ E, 350 m, 1� (23.05.1948, leg.Strouhal, NHMW); Helenental, 48° 01′ N, 16° 11′ E, 2�(16.04.1912, leg. Penther, NHMW, TYPE SERIES of Chtho-nius (Ephippiochthonius) austriacusBeier, 1931); Neupurk-ersdorf, N Graben, 48° 12′ N, 16° 09′ E, 300 m, 2�, 1�(30.05.1948, leg. Strouhal, NHMW); Weichseltal S BadenerLindkogel, 48° 00′ N, 16° 09′ E, 230 m, 1� (01.01.1948, leg.Strouhal, NHMW); S Breitenfurt b. Wien, Thomasgraben,48° 08′ N, 16° 08′ E, 1�, 4� (06.05.1956, leg. Strouhal,NHMW); N Heiligenkreuz, Ufer des Dornbaches, 48° 5′ N,16° 08′ E, 370 m, 1 juv. (24.08.1952, leg. Strouhal, NHMW);Sulz, NW, zw. Gr. und Kl. Winkelberg, 48° 6′ N, 16° 08′ E,
430 m, 3�, 1� (03.05.1953, leg. Strouhal, NHMW); Sulz,SW, 48° 6′ N, 16° 08′ E, 400 m, 1� (24.08.1952, leg.Strouhal, NHMW); NE Troppberg, 48° 14′ N, 16° 08′ E,450 m, 18�, 1� (01.01.1948, leg. Strouhal, NHMW); Tropp-berg SE-Hang, 48° 13′ N, 16° 06′ E, 1� (17.05.1948, leg.Strouhal, NHMW); N Rekawinkel, Stiefelberg W-Hang, 48°11′ N, 16° 01′ E, 360 m, 3�, 7� (19.08.1948, leg. Strouhal,NHMW); S Rekawinkel, 48° 11′ N, 16° 01′ E, 400 m, 2�,5 � (04.07.1948, leg. Strouhal, NHMW); W Wintern (N Eich-graben), 48° 10′ N, 15° 56′ E, 310 m, 2� (25.06.1950, leg.Strouhal, NHMW); Schneeberg region, S Peisching, HoheWand, 47° 51′ N, 16° 05′ E, 10�, 1 juv. (04.06.1950, leg.Strouhal, NHMW); Würflach, Johannesbachklamm, SW Wil-lendorf an der Schneebergbahn, 47° 47′ N, 16° 03′ E, 450 m,2 �, 1� (27.08.1951, leg. Strouhal, NHMW); E Puchberg amSchneeberg, oberes Pfennigbachtal, 47° 48′ N, 15° 57′ E,650–700 m, 2� (21.08.1951, leg. Strouhal, NHMW); E Puch-berg am Schneeberg, Reitzenberg, 47° 48′ N, 15° 57′ E, 1�(02.07.1950 (?), leg. Strouhal, NHMW); SE Puchberg amSchneeberg, SE, Kienberg, SW-Hang, 47° 46′ N, 15° 54′ E,700 m, 1� (13.08.1951, leg. Strouhal, NHMW); N Prigglitz(N Gloggnitz), 47° 43′ N, 15° 56′ E, 700 m, 2� (24.08.1951,leg. Strouhal, NHMW); WNW Gutenstein, Graben NEGr. Gaiskopf, 47° 54′ N, 15° 42′ E, 600 m, 2� (07.07.1952,leg. Strouhal, NHMW); Waldviertel, W Hardegg, 48° 51′ N,15° 50′ E, 1� (1948, leg. Strouhal, NHMW); Wachau, WAggsbach-Markt, 48° 18′ N, 15° 22′ E, 3�, 3� (1948, leg.Strouhal, NHMW); Graben W Hohenwartberg (zw. Aggs-bach-Markt und Schönbühel), 48° 17′ N, 15° 22′ E, 220 m,3 �, 1� (01.01.1948, leg. Strouhal, NHMW); Scheibbs dis-trict, Reinsberg Robitzboden, 47° 59′ N, 15° 4′ E, 1�(01.01.1948, leg. Ressl, NHMW); Puchenstuben, GaisenbergW-Hang, 47° 56′ N, 15° 19′ E, 800 m, 2�, 2 juv. (12.08.1947,leg. Strouhal, NHMW), 2�, 1 juv. (18.08.1947, leg. Strouhal,NHMW); E Puchenstuben, Langseit, 47° 56′ N, 15° 19′ E,550 m, 1�, 3 juv. (12.08.1947, leg. Strouhal, NHMW); EPuchenstuben, Rotte Berg, 47° 56′ N, 15° 19′ E, 600 m, 1�,2 �, 1 juv. (12.08.1947, leg. Strouhal, NHMW); Nattersbach,linkes Seitental bei Laubenbachmühle, 47° 57′ N, 15° 17′ E,550 m, 1� (21.08.1947, leg. Strouhal, NHMW); Erlauftal(zw. Hint. Tormäuer u. Erlaufboden), 47° 52′ N, 15° 16′ E,600 m, 1�, 1� (20.08.1947, leg. Strouhal, NHMW); Schiess-wand (zw. Gösing u. Erlaufböden), 47° 53′ N, 15° 16′ E,650 m, 10�, 5��1 juv. (20.08.1947, leg. Strouhal, NHMW);Scheibbs, Blassenstein, 48° 1′ N, 15° 12′ E, 4�, 1�1 juv.,04.08.1950, leg. Franz, NHMW).Oberösterreich: Mauthausen, 48° 15′ N, 14° 31′ E, 2�(01.01.1948, leg. Strouhal, NHMW); Traun bei Weißkirchen,Auenwald, 48° 10′ N, 14° 8′ E, 300 m, 1� (05.09.1948, leg.Strouhal, NHMW); SW Grünau, vorderer Rinnbachgraben,47° 51′ N, 13° 57′ E, 2� (26.08.1964, leg. Strouhal,NHMW).Steiermark: Hartberg district, N Großhart, W Waltersdorf,47° 10′ N, 15° 57′ E, 400 m, 5�, 7� (01.01.1948, leg.Strouhal, NHMW); NW Buchgreit, S Waltersdorf, 47° 09′ N,16° 00′ E, 350 m, 7�, 3� (01.01.1948, leg. Strouhal, NHMW);Raab-Klamm bei Weiz, NE Matl-Schuster, 47° 11′ N, 15° 31′ E,1 �, 1� (01.01.1948, leg. Strouhal, NHMW).Italy: Prov. Parma: Bedonia, 44° 31′ N, 09° 37′ E, 1 juv.(01.01.1948, NHMW).
Vicariance in a Cryptic Species Pair of European Pseudoscorpions311




















