
Vertical distribution and temporal variation of marine planktonic archaea in the Gerlache Strait,...
11
Vertical distribution and temporal variation of marine planktonic archaea in the Gerlache Strait, Antarctica, during early spring Ramon Massana, Lance T. Taylor, Alison E. Murray, Ke Y. Wu, Wade H. Jefsyey,’ and Edward F. DeLong2 Marine Science Institute, University of California, Santa Barbara, Santa Barbara, California, California 93106 Abstract A station located in the Gerlache Strait (Antarctic Peninsula) was sampled during early spring to determine the vertical distribution of marine planktonic archaea and to further describe the dynamic environment where they live. Chlorophyll concentration indicated that sampling occurred during the early stages of austral spring algal devel- opment. As expected, prokaryote abundance was higher at surface than at depth, and prokaryotic activity estimated by leucine incorporation was low at the surface and extremely low at 500 m. The relative abundance of planktonic archaeal, eucaryal, and bacterial ribosomal RNA was determined by quantitative rRNA hybridization, and the performance of two different universal probes used to normalize group-specific probe hybridization response was compared. Archaeal rRNA was detected in all samples analyzed, and was more abundant at depth (up to 25% of total rRNA) than at the surface. In both years, the archaeal signal decreased during the sampling period, particularly at surface. Most of the archaeal signal was attributable to group I archaea, affiliated with the kingdom Crenarchaeota. Planktonic euryarchaeotes (group II archaea) showed a larger contribution to the archaeal assemblage in surface waters than at depth. In total, our results verify that planktonic archaea are dynamic and abundant components in marine picoplankton assemblages of the Antarctic Peninsula. The Southern Ocean includes all the water surrounding the Antarctic continent south of the Antarctic Convergence (Vincent 1988; Karl 1993). It is one of the largest contiguous ecosystems on Earth and is one of the least understood, be- ing undersampled with respect to other more accessible lo- cations. In general, the Southern. Ocean is characterized by oligotrophic conditions in terms of biomass and production. However, certain regions have a seasonal cycle that includes a very high productivity and an active and dynamic food web. The Gerlache Strait, a region west of the Antarctic Peninsula, is one of these productive areas (Holm-Hansen and Vemet 1990; Holm-Hansen and Mitchell 1991). The biological cycle in Antarctic waters is strongly con- ditioned by the seasonal cycle. After a cold and dark winter with minimal primary productivity, the increase of daylength as well as the increase of the stability of the water column coincide with the development of algal blooms, particularly in coastal areas and in the marginal ice zone. The phyto- plankton nurtures extensive krill populations that serve as I University of West Florida, Center for Environmental Diagnos- tics and Bioremediation, 11000 University Parkway, Pensacola, Florida 32514. 2 Corresponding author, now at: Monterey Bay Aquarium Re- search Institute, PO. Box 628, 7700 Sandholdt Rd., Moss Landing, California 95039. Acknowledgments We are indebted to the captain and crew of the RV Polar Duke, as well as Antarctic Support Associate employees, for continuous help and support during the cruise. Special thanks to Odd-Helge Tiegen and Pare for solving what we thought to be insoluble prob- lems. R.M. was recipient of a postdoctoral fellowship from the Spanish Ministry of Education and Science. This work was sup- ported by National Science Foundation grants OCE 95-29804 and OPP 94-18442 to EED., and OPP-9419037 to W.H.J. food for fish, birds, and mammals. This food chain, once thought to be extremely simple, has been modified to include nutrient cycling through heterotrophic and phototrophic mi- croorganisms (Vincent 1988; Karl 1993). Prokaryotes are recognized as important components of marine food webs, both as remineralizers and as conduits for the conversion of dissolved organic matter to particulate or- ganic matter, which is incorporated into higher trophic levels (Azam et al. 1983; Fenchel 1988). With the development of new methods for studying microbial diversity, recent efforts have focused on the phylogenetic identification of naturally occurring planktonic prokaryotes. Many new phylogenetic types within the domain Bacteria have been recently dis- covered in the marine plankton (Giovannoni et al. 1990; Britschgi and Giovannoni 1991; Schmidt et al. 1991; Fuhr- man et al. 1993; Mullins et al. 1995), as have a few groups within the domain Archaea (DeLong 1992; Fuhrman et al. 1992). The presence and high relative abundance of archaea in cold and oxic marine waters were initially surprising, be- cause no cultured archaea (methanogens, halophiles, and thermophiles) are known to live in such habitats. Neverthe- less, marine archaea have been detected in many different regions of the world oceans (DeLong 1992; Fuhrman et al. 1992; Massana et al. 1997a), including Antarctic coastal wa- ters (DeLong et al. 1994). Only two major phylogenetic ar- chaeal groups have so far been consistently detected in di- verse marine waters using molecular phylogenetic approaches. Group I is affiliated with the archaeal kingdom Crenarchaeota, and group II belongs to a lineage associated with the archaeal kingdom Euryarchaeota. The vertical dis- tribution of these two groups in temperate coastal waters has been reported to be different, with group II being prevalent at surface and group I at depth (Massana et al. 1997a). In a previous study, we reported a high archaeal signal in 607
Transcript of Vertical distribution and temporal variation of marine planktonic archaea in the Gerlache Strait,...

Vertical distribution and temporal variation of marine planktonic archaea in the Gerlache Strait, Antarctica, during early spring
Ramon Massana, Lance T. Taylor, Alison E. Murray, Ke Y. Wu, Wade H. Jefsyey,’ and Edward F. DeLong2 Marine Science Institute, University of California, Santa Barbara, Santa Barbara, California, California 93106
Abstract
A station located in the Gerlache Strait (Antarctic Peninsula) was sampled during early spring to determine the vertical distribution of marine planktonic archaea and to further describe the dynamic environment where they live. Chlorophyll concentration indicated that sampling occurred during the early stages of austral spring algal devel- opment. As expected, prokaryote abundance was higher at surface than at depth, and prokaryotic activity estimated by leucine incorporation was low at the surface and extremely low at 500 m. The relative abundance of planktonic archaeal, eucaryal, and bacterial ribosomal RNA was determined by quantitative rRNA hybridization, and the performance of two different universal probes used to normalize group-specific probe hybridization response was compared. Archaeal rRNA was detected in all samples analyzed, and was more abundant at depth (up to 25% of total rRNA) than at the surface. In both years, the archaeal signal decreased during the sampling period, particularly at surface. Most of the archaeal signal was attributable to group I archaea, affiliated with the kingdom Crenarchaeota. Planktonic euryarchaeotes (group II archaea) showed a larger contribution to the archaeal assemblage in surface waters than at depth. In total, our results verify that planktonic archaea are dynamic and abundant components in marine picoplankton assemblages of the Antarctic Peninsula.
The Southern Ocean includes all the water surrounding the Antarctic continent south of the Antarctic Convergence (Vincent 1988; Karl 1993). It is one of the largest contiguous ecosystems on Earth and is one of the least understood, be- ing undersampled with respect to other more accessible lo- cations. In general, the Southern. Ocean is characterized by oligotrophic conditions in terms of biomass and production. However, certain regions have a seasonal cycle that includes a very high productivity and an active and dynamic food web. The Gerlache Strait, a region west of the Antarctic Peninsula, is one of these productive areas (Holm-Hansen and Vemet 1990; Holm-Hansen and Mitchell 1991).
The biological cycle in Antarctic waters is strongly con- ditioned by the seasonal cycle. After a cold and dark winter with minimal primary productivity, the increase of daylength as well as the increase of the stability of the water column coincide with the development of algal blooms, particularly in coastal areas and in the marginal ice zone. The phyto- plankton nurtures extensive krill populations that serve as
I University of West Florida, Center for Environmental Diagnos- tics and Bioremediation, 11000 University Parkway, Pensacola, Florida 32514.
2 Corresponding author, now at: Monterey Bay Aquarium Re- search Institute, PO. Box 628, 7700 Sandholdt Rd., Moss Landing, California 95039.
Acknowledgments We are indebted to the captain and crew of the RV Polar Duke,
as well as Antarctic Support Associate employees, for continuous help and support during the cruise. Special thanks to Odd-Helge Tiegen and Pare for solving what we thought to be insoluble prob- lems. R.M. was recipient of a postdoctoral fellowship from the Spanish Ministry of Education and Science. This work was sup- ported by National Science Foundation grants OCE 95-29804 and OPP 94-18442 to EED., and OPP-9419037 to W.H.J.
food for fish, birds, and mammals. This food chain, once thought to be extremely simple, has been modified to include nutrient cycling through heterotrophic and phototrophic mi- croorganisms (Vincent 1988; Karl 1993).
Prokaryotes are recognized as important components of marine food webs, both as remineralizers and as conduits for the conversion of dissolved organic matter to particulate or- ganic matter, which is incorporated into higher trophic levels (Azam et al. 1983; Fenchel 1988). With the development of new methods for studying microbial diversity, recent efforts have focused on the phylogenetic identification of naturally occurring planktonic prokaryotes. Many new phylogenetic types within the domain Bacteria have been recently dis- covered in the marine plankton (Giovannoni et al. 1990; Britschgi and Giovannoni 1991; Schmidt et al. 1991; Fuhr- man et al. 1993; Mullins et al. 1995), as have a few groups within the domain Archaea (DeLong 1992; Fuhrman et al. 1992). The presence and high relative abundance of archaea in cold and oxic marine waters were initially surprising, be- cause no cultured archaea (methanogens, halophiles, and thermophiles) are known to live in such habitats. Neverthe- less, marine archaea have been detected in many different regions of the world oceans (DeLong 1992; Fuhrman et al. 1992; Massana et al. 1997a), including Antarctic coastal wa- ters (DeLong et al. 1994). Only two major phylogenetic ar- chaeal groups have so far been consistently detected in di- verse marine waters using molecular phylogenetic approaches. Group I is affiliated with the archaeal kingdom Crenarchaeota, and group II belongs to a lineage associated with the archaeal kingdom Euryarchaeota. The vertical dis- tribution of these two groups in temperate coastal waters has been reported to be different, with group II being prevalent at surface and group I at depth (Massana et al. 1997a).
In a previous study, we reported a high archaeal signal in
607

608 Massana et al.
Arthur Harbor (adjacent to Palmer Station, Anvers Island, Antarctic Peninsula) in surface samples collected over a 2- week period in late winter 1993 (DeLong et al. 1994). A more detailed study of this nearshore, shallow system has been conducted during early spring of 1995 and 1996 (Mur- ray et al. in press). In order to determine whether archaeal abundance is a specific characteristic of Arthur Harbor or is a more general feature of coastal Antarctic waters, we sam- pled a station in the center of the Gerlache Strait, a dynamic aquatic system with depths averaging 500-1,000 m (Niiler et al. 1991). The station was occupied for 1 month during 2 consecutive years in early spring. In 1995 we collected tem- perature and salinity data and determined the relative abun- dance of archaea, eucarya, and bacteria in plankton samples from different depths. In 1996, additional measurements, in- cluding counts of prokaryotes and flagellates, chlorophyll determinations, and leucine incorporation estimates, as well as size-fractionation experiments and attempts to measure archaeal-specific metabolic activities, were performed. In this paper we present the vertical distribution of archaea in the water column of the Gerlache Strait and their changes during 1 month in early spring, and we relate these data with other biological and physical data.
Materials and methods
Sampling and standard measurements-A station in the Gerlache Strait between Two Hummock Island and Brabant Island (64”13.11’$ 61”50.71’W) was visited with the RV Polar Duke. Sampling was conducted five times during the first cruise (13 October-l November 1995) and eight times during the second cruise (28 September-26 October 1996). Profiles of temperature, salinity, and fluorescence (Sea-Tech fluorometer) were obtained with a SeaBird SBE-9 CTD pro- filer. Seawater samples from discrete depths were collected with twelve 12-liter Niskin bottles installed on the CTD. Typically, 65-85 liters were collected at four depths (1995) or 20-40 liters at five depths (1996).
Seawater samples were processed immediately after sam- pling. During the 1995 cruise, samples were filtered by pres- sure filtration sequentially through 0.8- and 0.2-pm Supor filters (Gelman), and the 0.2-pm-filtered samples were kept frozen at -80°C until nucleic acid extraction. During the 1996 cruise, subsamples were fixed with ice-cold glutaral- dehyde (1% final concn) for microscopic counts. Seawater aliquots of 0.3-1.0 liters were filtered through 0.45-pm ny- lon filters (Cole-Parmer) for chlorophyll a (Chl a) determi- nations. Filters were dissolved overnight in 90% acetone at -20°C and the extract was read in a Turner AU10 digital fluorometer (Parsons et al. 1984). The remaining seawater (-18-38 liters) was sequentially filtered through 1.6-pm GF/A filters (Whatman) and a 0.2-pm Sterivex-GV filter (Millipore) using a peristaltic pump (Somerville et al. 1989). Sterivex filters were filled with 1.8 ml of lysis buffer (40 mM EDTA, 50 mM Tris-HCl and 0.75 M sucrose) and kept frozen at -80°C until nucleic acid extraction.
Microbial counts were performed on board with a Zeiss epifluorescence microscope installed on hydraulic table. Ali- quots of the glutaraldehyde fixed sample (10 ml) were
stained with DAPI (5 pg ml I final concn) and filtered through 0.2-pm black-stained polycarbonate filters (Poret- its). Prokaryotes and flagellates were counted by standard epifluorescence microscopy (Porter and Feig 1980). The presence or absence of chlorophyll in flagellates was checked using blue light irradiation. Heterotrophic flagellates were sized using a grid in the ocular, and biovolumes were converted to biomass with the correction factor 0.22 pg C pm-l (Barsheim and Bratbak 1987). Some fixed samples were transported to shore to determine the size of prokary- otes by image analysis (Massana et al. 1997b). The mean biovolume was determined in seven samples and averaged 0.044 pm7 cell -I, corresponding to a carbon content of 15 fg C cell-’ (Simon and Azam 1989).
Leucine incorporation experiments were initiated by add- ing PHIleucine (180 Ci mmol ‘) to 45 ml of sample ( 10 nM final concn) and incubating the bottles in the dark at in situ temperatures (between - 1.7 and -0.3”C). Subsamples of 15 ml were processed at times 0 and 4 h (duplicates) by filtering through a 0.45-pm Millipore filter, followed by three washes with 5% cold TCA and two washes with 80% cold ethanol. The filters were placed in scintillation vials, dissolved with 0.5 ml ethyl acetate, and radioassayed onboard. Leucine in- corporation was converted to prokaryote carbon production (Kirchman 1993), and doubling times were calculated from prokaryote production and biomass.
In routine practice, DAPI counts were not replicated. However, we estimated from selected samples that the co- efficient of variation for bacterial counts averaged 4.8% (range of 0.5-l 1.6; n = 15). Chlorophyll determinations were always done in duplicates, and the coefficient of vari- ation for all samples averaged 3.3% (range of O-l 1.7; n = 16). Leucine determinations were also done in duplicates, and the coefficient of variation for all samples averaged 4.1% (range of O-19.9; n = 24).
Nucleic acid analysis-Nucleic acid extracts were ob- tained from the microbial biomass collected on Supor or Sterivex filters by the lysozyme-sodium dodecyl sulfate- proteinase K treatment and phenol-chloroform extraction, as previously described (Massana et al. 1997a). The extracts were checked by agarose gel electrophoresis and quantified by a Hoechst dye fluorescence assay (Paul and Myers 1982). Extraction yields for the 1996 samples were between 0.04 and 0.25 pg DNA liter ~’ (mean of 0.14).
For quantitative rRNA hybridization experiments, nucleic acid extracts from natural samples and from controls (Sul- folobus acidocaldarius, Saccharomyces cerevisiae, and Escherichia coli) were denatured with 1.5% glutaraldehyde. A four (or six)-fold serial dilution of the extracts (50, 25, 12.5, and 6.25 ng DNA) was loaded onto nylon membranes (Hybond-N, Amersham) using a slot blotting apparatus (MilliBlot-S, Millipore), and immobilized by UV crosslink- ing (Stratalinker, Stratagene). Several identical membranes containing samples and controls were prepared to be sepa- rately hybridized with each probe. The actual amount of RNA loaded was undetermined (community DNA : RNA ra- tios are variable), but that value was not necessary to cal- culate relative abundances of the three domains, expressed as percentages of the total rRNA signal (see below). Mem-

Archaea in Antarctic waters 609
branes were preincubated at 45°C for 30 min with 10 ml of hybridization buffer (0.9 M NaCl, 50 mM Na,HPO,, 5 mM EDTA, 0.5% SDS, 10X Denhardt’s solution, 0.5 mg ml-’ polyadenosine) followed by the addition of 16S rRNA oli- gonucleotide probes end-labeled with 32P. The following probes were used (Alm et al. 1996): S-*-Univ-1390-a-A-18 (Univ-1390), S-*-Univ- 1392-a-A- 15 (Univ- 1392), S-D- Arch-0915-a-A-20 (Arch-915), S-D-Bact-0338-a-A-18 (Bact-338), and S-D-Euca-1209-a-A-16 (Euca-1209). After overnight hybridization at 45°C membranes were washed for 30 min at room temperature in 1 X SET (150 mM NaCl, 1 mM EDTA, 20 mM Tris) and 1% SDS. Membranes were then transferred to the same buffer preequilibrated to the high stringency wash temperature (37°C for Euca-1209 and Univ-1392; 45°C for Univ-1390 and Bact-338; 56°C for Arch-915), and washed for an additional 30 min, removed, and dried. Radioactivity was quantified by a radioanalytic gas-proportional counting system (Ambis Systems).
The relative eucaryal, bacterial, and archaeal rRNA abun- dance in the natural sample was calculated in three steps. First, the slope (cpm probe bound per unit rRNA) for each natural sample rRNA mixture was separately determined with each domain-specific probe. Second, this domain-spe- cific slope was multiplied by a domain-specific correction factor. (The correction factor was obtained in each experi- ment by dividing the slope of the universal probe bound, by the slope of corresponding domain-specific probe bound, to an identical control rRNA extracted from a pure culture.) Third, this corrected domain-specific slope was divided by the slope of the universal probe bound to the same natural sample, and the ratio was expressed as a percentage.
Probes specific for the marine crenarchaeotal group I (S- O-Cenar-0554-a-A-20, GI-554) and the marine euryarchaeo- tal group II (S-0-ArgII-0554-a-A-20, GII-554) were also in- cluded in hybridization experiments (Massana et al. 1997a). The dissociation temperature (T,) of these probes, deter- mined by the elution T,, method (Zheng et al. 1996) with rRNA transcripts (Polz and Cavanaugh 1997), is 48.8”C for GI-554 and 45.9”C for GII-554 (R. Massana unpubl. results). The high-stringency wash temperature was 40°C. For con- trols, we used the nucleic acid extract of a sponge that con- tains a symbiotic group I archaeon (Preston et al. 1996) and a plasmid containing a cloned rRNA gene fragment from a planktonic group II archaeon. Correction factors were ob- tained as described above, except that the group-specific probe binding was normalized to the archaeal probe instead of to the universal probe. The abundance of group I was expressed as percentage of the total archaeal signal, whereas the abundance of group II was expressed as hybridization signal (because no control rRNA is yet available for group II archaea).
Quantitative hybridization experiments were generally performed only once for each sample. We performed repli- cates from some samples (n = 12), and coefficients of vari- ation for archaeal, eucaryal, and bacterial rRNA relative abundances averaged 21.8% (range of 6.4-43.4), 18.7% (range of 3.9-41.8), and 10.3% (range of 3.0-21.6) respec- tively. We also estimated that the coefficient of variation for group I rRNA relative abundance averaged 7.3% (range of 1.5-13.3; n = 4), and the coefficient of variation for group
II hybridization signal averaged 17.2% (range of 0.6-34.6; n = 4).
Size fractionation experiments--In some samples, DAPI counts, Chl a measurements, and quantitative rRNA hybrid- ization experiments were performed on different size frac- tions of the microbial assemblage. Approximately 40 liters of seawater was collected for each sample. Half of this vol- ume was subsampled for DAPI counts, Chl a (0.2-pm nylon filters), and nucleic acids, and the other half was filtered through GF/A filters. Two GF/A filters corresponding to 1 liter of seawater were processed for Chl a. The GF/A filtrate was subsampled for DAPI counts and filtered through 0.8- pm Supor filters. Two Supor filters corresponding to 1 liter of seawater were processed for Chl a, and the filter corre- sponding to the remaining volume was stored at -80°C for nucleic acid extraction. The 0.8-pm filtrate was subsampled for DAPI counts and Chl a (0.2-pm nylon filter), and the remaining volume was filtered through a Sterivex filter. As a result of this fractionation strategy, DAPI counts were de- termined in the whole water and fractions of 0.2-1.6 and 0.2-0.8 pm; Chl a was determined in whole water and frac- tions of >1.6, 0.8-l .6, and 0.2-0.8 pm; biomass for nucleic acid extraction was collected in whole water and fractions of 0.8-1.6 and 0.2-0.8 pm.
Assay for urchaeal activity-Leucine incorporation was measured in presence of erythromycin (a bacterial, but not an archaeal protein synthesis inhibitor) and puromycin (a “universal” protein synthesis inhibitor). To obtain measur- able leucine incorporation in antibiotic treatments, the ex- periments were performed using microbial concentrates pre- pared using an Amicon ultrafiltration system (polysulfone hollow-fiber filter, 30,000 MW cutoff), which concentrated microorganisms from a large volume of seawater (prehltered through GF/A) to -1 liter. Prokaryote counts and leucine incorporation experiments (without antibiotics) were per- formed with the GF/A filtrate and the concentrate to deter- mine the initial effect of concentration on these fractions. The cell concentrate was preincubated for 1 h in separate bottles with varying amounts of erythromycin and puro- mycin, and leucine incorporation was measured as described above. The activity with each dose of antibiotic was ex- pressed as a percentage relative to the activity without an- tibiotic addition. If antibiotics performed as expected, the archaeal contribution to leucine incorporation would theo- retically be the activity remaining in the erythromycin treat- ment (only bacterial activity inhibited) minus the activity remaining in the puromycin treatment (all leucine incorpo- ration inhibited).
Results
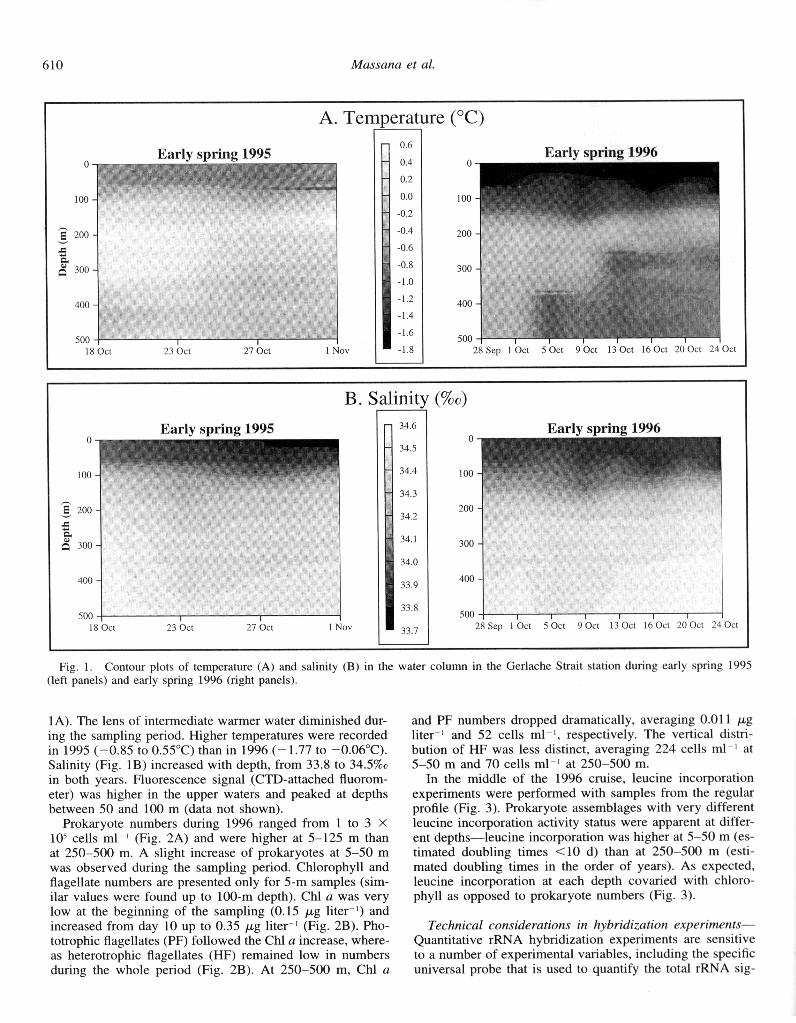
The Gerluche Strait ecosystem-A station in the Gerlache Strait was occupied during several weeks in early austral spring of 2 consecutive years. Our goals were to confirm the presence of archaea in this dynamic system and to describe the ecological setting in which they develop. In both years temperatures were lower in surface waters, increased at in- termediate depths, and decreased toward the bottom (Fig.
Archueu in Antarctic waters 611
J B + HF
u PF 4 d Chlu
b 5m
U 50m
-0. 100-125 ITI
--A--- 25om
+ soom
28 scp I Ott 5 Ott YOct IlOct 16Oct 20Oct 24 act
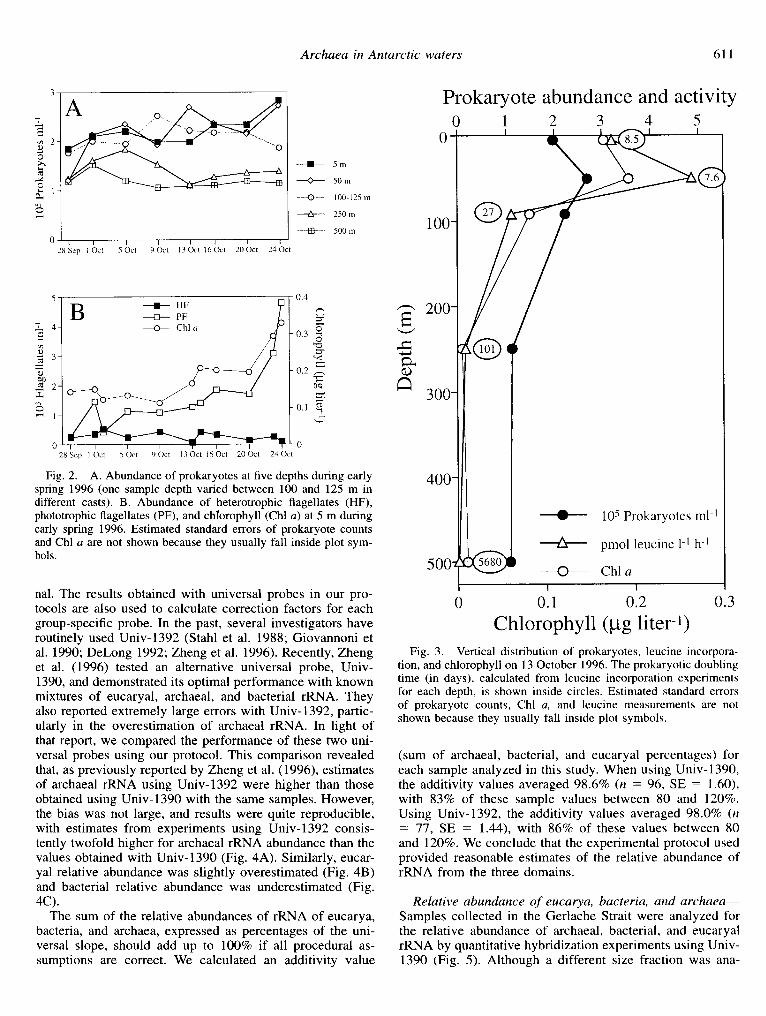
Fig. 2. A. Abundance of prokaryotes at tive depths during early spring 1996 (one sample depth varied between 100 and 125 m in different casts). B. Abundance of heterotrophic flagellates (HF), phototrophic flagellates (PF), and chlorophyll (Chl a) at 5 m during early spring 1996. Estimated standard errors of prokaryote counts and Chl a are not shown because they usually fall inside plot sym- bols.
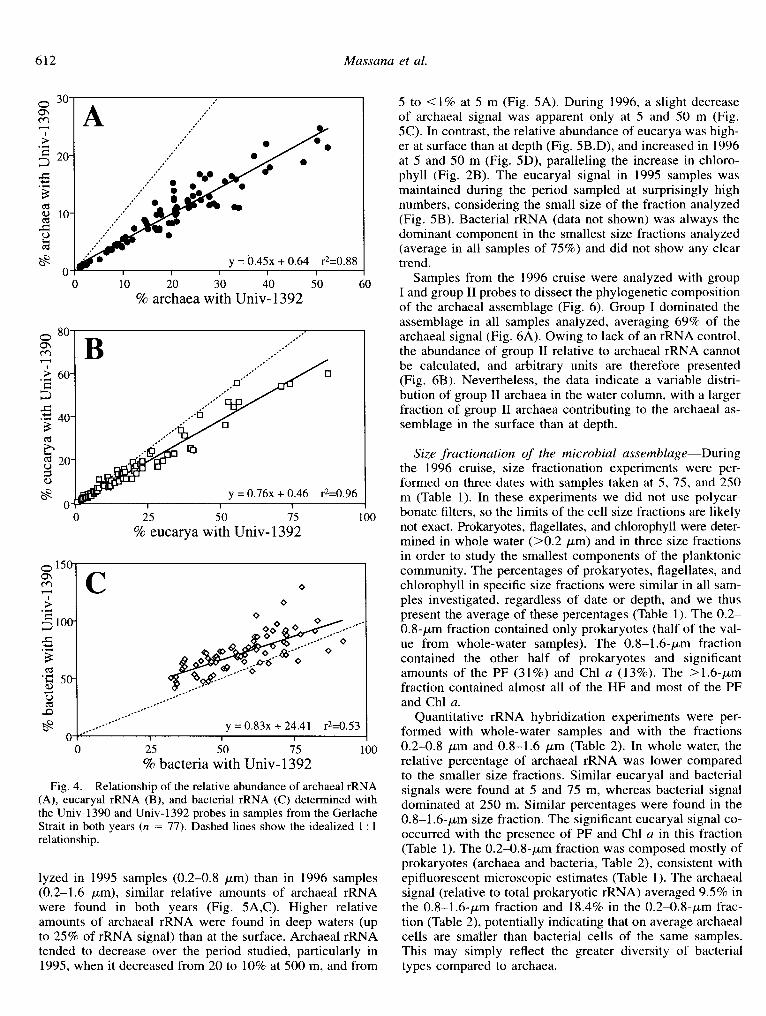
nal. The results obtained with universal probes in our pro- tocols are also used to calculate correction factors for each group-specific probe. In the past, several investigators have routinely used Univ-1392 (Stahl et al. 1988; Giovannoni et al. 1990; DeLong 1992; Zheng et al. 1996). Recently, Zheng et al. (1996) tested an alternative universal probe, Univ- 1390, and demonstrated its optimal performance with known mixtures of eucaryal, archaeal, and bacterial rRNA. They also reported extremely large errors with Univ-1392, partic- ularly in the overestimation of archaeal rRNA. In light of that report, we compared the performance of these two uni- versal probes using our protocol. This comparison revealed that, as previously reported by Zheng et al. (1996), estimates of archaeal rRNA using Univ-1392 were higher than those obtained using Univ-1390 with the same samples. However, the bias was not large, and results were quite reproducible, with estimates from experiments using Univ-1392 consis- tently twofold higher for archaeal rRNA abundance than the values obtained with Univ-1390 (Fig. 4A). Similarly, eucar- yal relative abundance was slightly overestimated (Fig. 4B) and bacterial relative abundance was underestimated (Fig. 4C).
The sum of the relative abundances of rRNA of eucarya, bacteria, and archaea, expressed as percentages of the uni- versal slope, should add up to 100% if all procedural as- sumptions are correct. We calculated an additivity value
Prokaryote abundance and activity
0
100
- 3
200
-fr 3
a 300
400
5oc
e 10” Prokaryotes ml-’
+ pmol leucine 1-t h-l
U Chl a
0 0.1 0.2 0.3
Chlorophyll (pg liter-l) Fig. 3. Vertical distribution of prokaryotes, leucine incorpora-
tion, and chlorophyll on 13 October 1996. The prokaryotic doubling time (in days), calculated from leucine incorporation experiments for each depth, is shown inside circles. Estimated standard errors of prokaryote counts, Chl a, and leucine measurements are not shown because they usually fall inside plot symbols.
(sum of archaeal, bacterial, and eucaryal percentages) for each sample analyzed in this study. When using Univ- 1390, the additivity values averaged 98.6% (n = 96, SE = 1.60), with 83% of these sample values between 80 and 120%. Using Univ-1392, the additivity values averaged 98.0% (n = 77, SE = 1.44), with 86% of these values between 80 and 120%. We conclude that the experimental protocol used provided reasonable estimates of the relative abundance of rRNA from the three domains.
Relative abundance of eucarya, bacteria, and urchaea- Samples collected in the Gerlache Strait were analyzed for the relative abundance of archaeal, bacterial, and eucaryal rRNA by quantitative hybridization experiments using Univ- 1390 (Fig. 5). Although a different size fraction was ana-
Massana et al.
E+? y = 0.45x + 0.64 r2=0.88 0 I I , I I
0 100
Fig. 4. Relationship of the relative abundance of archaeal rRNA (A), eucaryal rRNA (B), and bacterial rRNA (C) determined with the Univ-1390 and Univ-1392 probes in samples from the Gerlache Strait in both years (n = 77). Dashed lines show the idealized 1 : 1 relationship.
lyzed in 1995 samples (0.2-0.8 pm) than in 1996 samples (0.2-1.6 pm), similar relative amounts of archaeal rRNA were found in both years (Fig. 5A,C). Higher relative amounts of archaeal rRNA were found in deep waters (up to 25% of rRNA signal) than at the surface. Archaeal rRNA tended to decrease over the period studied, particularly in 1995, when it decreased from 20 to 10% at 500 m, and from
5 to <l% at 5 m (Fig. 5A). During 1996, a slight decrease of archaeal signal was apparent only at 5 and 50 m (Fig. 5C). In contrast, the relative abundance of eucarya was high- er at surface than at depth (Fig. 5B,D), and increased in 1996 at 5 and 50 m (Fig. 5D), paralleling the increase in chloro- phyll (Fig. 2B). The eucaryal signal in 1995 samples was maintained during the period sampled at surprisingly high numbers, considering the small size of the fraction analyzed (Fig. 5B). Bacterial rRNA (data not shown) was always the dominant component in the smallest size fractions analyzed (average in all samples of 75%) and did not show any clear trend.
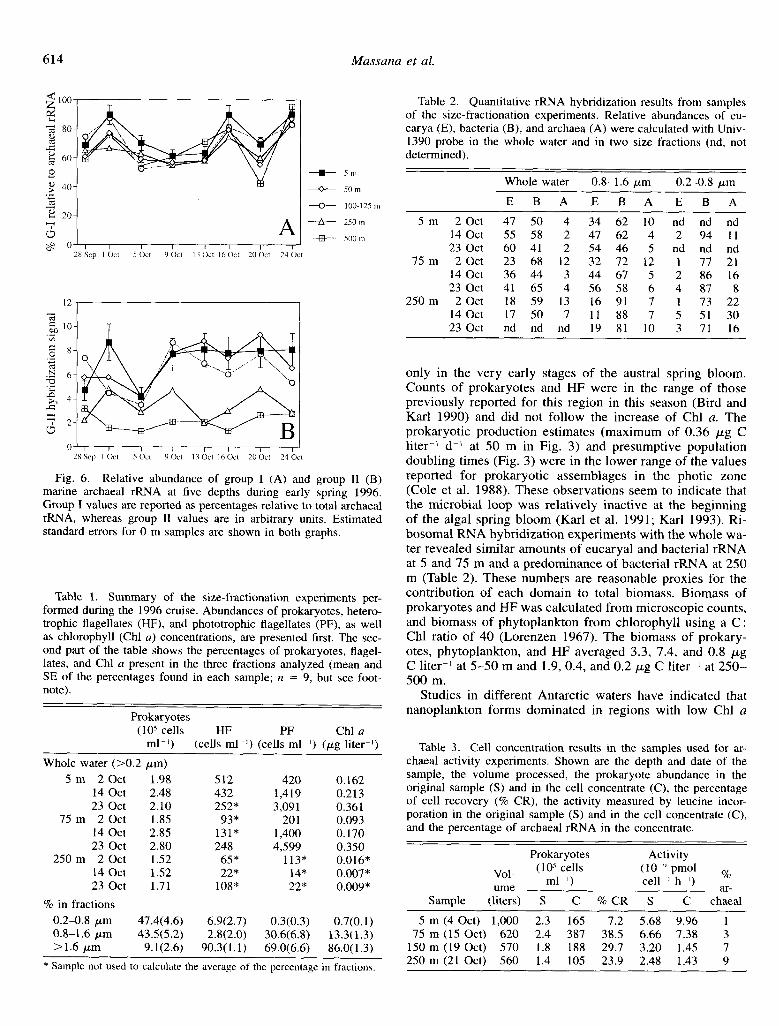
Samples from the 1996 cruise were analyzed with group I and group II probes to dissect the phylogenetic composition of the archaeal assemblage (Fig. 6). Group I dominated the assemblage in all samples analyzed, averaging 69% of the archaeal signal (Fig. 6A). Owing to lack of an rRNA control, the abundance of group II relative to archaeal rRNA cannot be calculated, and arbitrary units are therefore presented (Fig. 6B). Nevertheless, the data indicate a variable distri- bution of group II archaea in the water column, with a larger fraction of group II archaea contributing to the archaeal as- semblage in the surface than at depth.
Size fractionation of the microbial assemblage-During the 1996 cruise, size fractionation experiments were per- formed on three dates with samples taken at 5, 75, and 250 m (Table 1). In these experiments we did not use polycar- bonate filters, so the limits of the cell size fractions are likely not exact. F’rokaryotes, flagellates, and chlorophyll were deter- mined in whole water (>0.2 km) and in three size fractions in order to study the smallest components of the planktonic community. The percentages of prokaryotes, flagellates, and chlorophyll in specific size fractions were similar in all sam- ples investigated, regardless of date or depth, and we thus present the average of these percentages (Table 1). The 0.2- 0.8-pm fraction contained only prokaryotes (half of the val- ue from whole-water samples). The 0.8-1.6-pm fraction contained the other half of prokaryotes and significant amounts of the PF (31%) and Chl a (13%). The > 1.6-pm fraction contained almost all of the HF and most of the PF and Chl a.
Quantitative rRNA hybridization experiments were per- formed with whole-water samples and with the fractions 0.2-0.8 pm and 0.8-1.6 pm (Table 2). In whole water, the relative percentage of archaeal rRNA was lower compared to the smaller size fractions. Similar eucaryal and bacterial signals were found at 5 and 75 m, whereas bacterial signal dominated at 250 m. Similar percentages were found in the 0.8-1.6-pm size fraction. The significant eucaryal signal co- occurred with the presence of PF and Chl a in this fraction (Table 1). The 0.2-0.8-pm fraction was composed mostly of prokaryotes (archaea and bacteria, Table 2) consistent with epifluorescent microscopic estimates (Table 1). The archaeal signal (relative to total prokaryotic rRNA) averaged 9.5% in the 0.8-1.6-pm fraction and 18.4% in the 0.2-0.8-pm frac- tion (Table 2) potentially indicating that on average archaeal cells are smaller than bacterial cells of the same samples. This may simply reflect the greater diversity of bacterial types compared to archaea.

Archaea in Antarctic waters
“-
A -A- Om
d T -o-- 30.7om
01
---0. 140.180n
--El+- 500m
I
13 Ott 18 Ott 23 Ott 27 Ott 1 NW
4oT ,B
0 I
s*
0’ , I I I , 13 Ott IXOct 23 Ott 27 Ott 1 NW
28 Sep 1 Ott 5 act 9 Ott 13 Ott 16 Ott 20 Ott 24 Ott
I 1 I I I I I I 28 Sep I Ott 5 Ott 9 Ott 13 Ott I6 Ott 20 Ott 24 Ott
Fig. 5. Relative abundance of archaeal rRNA (A) and eucaryal rRNA (B) at four depths during early spring 1995 (two sample depths varied in different casts). Relative abundance of archaeal rRNA (C) and eucaryal rRNA (D) at five depths during early spring 1996. All data points were calculated using the Univ-1390 probe. Estimated standard errors for 0 m and 500 m samples are shown in all graphs.
Experiments to assess the activity of archaea-We per- formed several experiments with microbial concentrates from different depths in order to assess the participation of archaea in prokaryote biomass production as estimated from leucine incorporation. The concentration protocol increased prokaryotic cell density by almost two orders of magnitude (although cell recovery was low and variable), and leucine incorporation per cell was not drastically affected (Table 3). The archaeal relative abundance in cell concentrates ranged from 1 to 9% (Table 3). Cell concentrates were used to test how different doses of erythromycin and puromycin inhib- ited leucine incorporation. However, erythromycin inhibition was always greater than puromycin inhibition (Fig. 7) and did not correlate with archaeal abundance in different ex- periments. Archaeal leucine incorporation could not be iden- tified or detected by using this particular methodological strategy.
Discussion
Microbial assemblages in the Gerlache Strait in early spring-The station studied is situated close to the center of the Gerlache Strait, over the deep continental shelf that sur- rounds the Antarctic Peninsula. This region has been studied in several RACER field campaigns during different seasons (Huntley et al. 1990, 1991). Calculated geostrophic flows (Amos et al. 1990; Niiler et al. 1991) and Lagrangian drifter
observations (Niiler et al. 1990) have demonstrated a con- sistent flow of surface waters from the southwest to the northeast along the main axis of the Gerlache Strait, with currents of lo-18 cm s ~I in the middle of the strait and residence times of a few weeks. Temperature plots (Fig. IA) confirm that this is a dynamic environment where a tongue of warmer waters at intermediate depths that disappear dur- ing the sampling period, with temperature being an indicator of different water masses (Niiler et al. 1991). The colder temperatures recorded in 1996 could be partly due to dif- ferent interannual sea ice conditions. Ice was much more widespread during 1995, potentially preventing extensive mixing and cooling of the water column. In Antarctica, vari- ations in seawater density are mainly governed by salinity, since only minor differences of temperature are found ver- tically in the water column. Therefore, density plots should be very similar to the salinity plots shown in Fig. lB, sug- gesting a marginally stratified water column. The upper mixed layer was variable, reaching depths of 100 m.
The Gerlache Strait is well known as a region with ex- tensive algal spring blooms. Chlorophyll is very low during winter (0.03-0.07 pg liter ‘; Cochlan et al. 1993) and in- creases during early spring to reach high values at the peak of the bloom (>20 pg liter ‘) between November and Jan- uary (Holm-Hansen and Vemet 1990; Holm-Hansen and Mitchell 1991). In our study, Chl a values increased from 0.15 to 0.35 pg liter I, indicating that sampling occurred
614 Massana et al.
28 Scp I Ott 5 Ott 9 act Il”Cf 16Oct 20”ct 24 ocr
o’, , I I I I 1 I 1
2x Sep I act 5 act 9 Ott 11 Ott I6 Ocr 20 Ocr 24 Ott
Fig. 6. Relative abundance of group I (A) and group 11 (B) marine archaeal rRNA at five depths during early spring 1996. Group I values are reported as percentages relative to total archaeal rRNA, whereas group II values are in arbitrary units. Estimated standard errors for 0 m samples are shown in both graphs.
Table 1. Summary of the size-fractionation experiments per- formed during the 1996 cruise. Abundances of prokaryotes, hetero- trophic flagellates (HF), and phototrophic flagellates (PF), as well as chlorophyll (Chl a) concentrations, are presented first. The sec- ond part of the table shows the percentages of prokaryotes, flagel- lates, and Chl a present in the three fractions analyzed (mean and SE of the percentages found in each sample; n = 9, but see foot- note).
Prokaryotes (lo5 cells HF PF Chl a
mll’) (cells ml-‘) (cells ml-l) (pg liter’)
Whole water (>0.2 pm)
5 m 2 Ott 1.98 14 Ott 2.48 23 Ott 2.10
75 m 2 Ott 1.85 14 Ott 2.85 23 Ott 2.80
250 m 2 Ott 1.52 14 Ott 1.52 23 Ott 1.71
512 432 252*
93* 131* 248 65* 22*
108*
420 0.162 1,419 0.213 3,091 0.361
201 0.093 1,400 0.170 4,599 0.350
113” 0.016* 14* 0.007* 22* 0.009*
% in fractions 0.2-0.8 pm 0.8-1.6 Grn
47.4(4.6) 6.9(2.7) 0.3(0.3) 0.7(0.1) 43.5(5.2) 2.8(2.0) 30.6(6.8) 13.3(1.3)
>1.6 /.&I 9.1(2.6j 90.3ii.ij 69.0(6.6j 86.0(1.3j
* Sample not used to calculate the average of the percentage in fractions.
Table 2. Quantitative rRNA hybridization results from samples of the size-fractionation experiments. Relative abundances of eu- carya (E), bacteria (B), and archaea (A) were calculated with Univ- 1390 probe in the whole water and in two size fractions (nd, not determined).
Whole water 0.8-1.6 Frn 0.2-0.8 pm
EBAEBAEBA
5m 2 Ott 47 50 4 34 62 10 nd nd nd 14 Ott 55 58 2 47 62 4 2 94 11 23 Ott 60 41 2 54 46 5 nd nd nd
75 m 2 Ott 23 68 12 32 72 12 1 77 21 14 Ott 36 44 3 44 67 5 2 86 16 23 Ott 41 65 4 56 58 6 4 87 8
250 m 2 Ott 18 59 13 16 91 7 1 73 22 14 Ott 17 50 7 11 88 7 5 51 30 23 Ott nd nd nd 19 81 10 3 71 16
only in the very early stages of the austral spring bloom. Counts of prokaryotes and HF were in the range of those previously reported for this region in this season (Bird and Karl 1990) and did not follow the increase of Chl a. The prokaryotic production estimates (maximum of 0.36 pg C liter-’ d-l at 50 m in Fig. 3) and presumptive population doubling times (Fig. 3) were in the lower range of the values reported for prokaryotic assemblages in the photic zone (Cole et al. 1988). These observations seem to indicate that the microbial loop was relatively inactive at the beginning of the algal spring bloom (Karl et al. 1991; Karl 1993). Ri- bosomal RNA hybridization experiments with the whole wa- ter revealed similar amounts of eucaryal and bacterial rRNA at 5 and 75 m and a predominance of bacterial rRNA at 250 m (Table 2). These numbers are reasonable proxies for the contribution of each domain to total biomass. Biomass of prokaryotes and HF was calculated from microscopic counts, and biomass of phytoplankton from chlorophyll using a C: Chl ratio of 40 (Lorenzen 1967). The biomass of prokary- otes, phytoplankton, and HF averaged 3.3, 7.4, and 0.8 pg C liter’ at 5-50 m and 1.9, 0.4, and 0.2 /*g C liter ’ at 250- 500 m.
Studies in different Antarctic waters have indicated that nanoplankton forms dominated in regions with low Chl a
Table 3. Cell concentration results in the samples used for ar- chaeal activity experiments. Shown are the depth and date of the sample, the volume processed, the prokaryote abundance in the original sample (S) and in the cell concentrate (C), the percentage of cell recovery (% CR), the activity measured by leucine incor- poration in the original sample (S) and in the cell concentrate (C), and the percentage of archaeal rRNA in the concentrate.
Prokaryotes Activity
Vol- (lOs cells (10 ” pm01
ume ml I) cell ’ h ‘)
%
ar- Sample (liters) S C%CRS C chaeal
5 m (4 Ott) 1,000 2.3 165 7.2 5.68 9.96 1 75 m (15 Ott) 620 2.4 387 38.5 6.66 7.38 3
150 m (19 Ott) 570 1.8 188 29.7 3.20 1.45 7 250 m (21 Ott) 560 1.4 105 23.9 2.48 1.43 9

Archaea in Antarctic waters 615
5 m (4 Ott)
100 150 m (I9 Ott) bn 250 m (21 Ott)
75 m (1.5 Ott)
0 AZibiotiZ”( pg nZ1 >
100
h *%3iotiT(pg 12-l)
100
0 Ar%thtiF(pg 12~ >
100 0 *$3ioti~(pg rZ- l>
100
Fig. 7. Percentage of leucine incorporation remaining in incubations containing varying amounts of erythromycin and puromycin (added to cell concentrates as described in Table 3. Estimated standard errors are not shown because they usually fall inside plot symbols).
(< 1 pg liter I), whereas microplankton forms dominated in regions of high phytoplankton biomass (Hewes et al. 1990). The same trend has been observed in Gerlache waters, where the nanoplankton dominate before and after the spring algal bloom (Holm-Hansen and Mitchell 1991). We sampled in a prebloom situation and observed that a significant fraction of the Chl a (13%, Table 1) was smaller than 1.6 pm, and that this is likely due to very small PE The presence of picophytoplankton and absence of cyanobacteria have been thoroughly documented in Antarctic waters (Letelier and Karl 1989; Hewes et al. 1990). The small number of PF most likely accounts for the eucaryal rRNA signal found in 1996 samples (Fig. 5D). On the other hand, the high eucaryal signal observed in 1995 samples (Fig. 5B) cannot be ex- plained with reference to size-fractionation experiments per- formed in 1996 that indicated no eucaryal signal in the 0.2- 0.8-pm fraction (Tables 1, 2). The size structure of the 1995 microbial assemblage might have been different. Another ex- planation is that artifacts may have been introduced by pres- sure filtration in the 1995 samples, which may have broken eucaryotic cells and allowed (eucaryotic) cell material to
pass through the 0.8-pm Supor filter. Additionally, prokary- ote counts at surface during the 1995 cruise were reported to be one order of magnitude lower than in 1996 (Pakulski et al. 1996), which could also have resulted in the greater relative percentage eucaryal rRNA in our 1995 samples.
Archaeal assemblage in the Gerlache Strait-Our data generally confirm that archaea are fundamental components of prokaryotic assemblages in Antarctic coastal waters, be- ing consistently present in the dynamic waters of the Ger- lathe Strait over the period sampled. Two trends were clearly identified: a higher relative percentage of archaeal rRNA at depth, and a decrease in the relative abundance of archaeal rRNA over time, especially at the surface (Fig. 5). Archaeal and eucaryal relative abundances followed opposite trends (Fig. 5). To avoid the potential effects of varying eucaryal abundance, archaeal relative abundance was recalculated with respect to only the prokaryotic signal (archaea plus bac- teria); the same trends over depth and time were observed.
High archaeal abundance was previously reported in coastal surface waters near Anvers Island in late winter

616 Massana et al.
(DeLong et al. 1994). These reported values, when numer- ically converted to Univ-1390 values using the regression shown in Fig. 4A, correspond to 9-14% archaeal rRNA and are similar to surface-water values determined in this study at the beginning of the sampling period. In total, the data indicate a decrease in archaeal numbers from late winter to spring. We postulate that continuous sampling through a spring bloom may reveal a further decreases of archaea in the prokaryotic assemblage, with archaeal numbers rebound- ing after the onset of winter.
Cultured and characterized archaea seem to thrive in hab- itats where their specialized physiologies or adaptations to extreme physicochemical conditions allow them to develop in the absence of intense competition with other (bacterial) species. If this generality holds true for marine archaea, they may be outcompeted by bacteria when conditions favorable for higher productivity (and thus the increased competition) appear, as typically occurs during algal blooms. So far, the distribution of group I marine crenarchaeotes temporally in Antarctica and spatially in temperate waters (Massana et al. 1997a) is consistent with this hypothesis.
The use of probes specific for different groups of marine archaea allowed us to estimate the contribution of these groups to archaeal assemblages in the Gerlache Strait. Data from 1995 (not shown) and from 1996 (Fig. 6) revealed that most of the archaeal rRNA was attributable to the crenar- chaeotal group I, averaging nearly 70% of the total archaeal signal in 1996. Reliable estimates of the percentage of the euryarchaeotal group II rRNA relative to other rRNA species are not presently available. However, our data indicated a higher proportion of group II rRNA at the surface (Fig. 6B). A similar trend was observed in waters from the Santa Bar- bara Channel, where group II archaea dominated in surface waters, and group I archaea were more abundant at depth (Massana et al. 1997a). The trend in both systems suggests that group II archaea develop mainly in upper waters. In our analyses, only these two archaeal groups have been detected in surveys of planktonic archaeal rDNA clone libraries ob- tained from samples collected in the Santa Barbara Channel (575 clones; Massana et al. 1997a) and in Arthur Harbor, Antarctica (559 clones; DeLong et al. 1998).
Archaeal activity-Significant leucine incorporation was detected in presence of the universal antibiotic puromycin (Fig. 7), confirming that the use of specific antibiotics has limitations and needs to be supported by additional data (Or- emland and Capone 1988). We performed different experi- ments in attempts to relate archaeal-specific activity with archaeal abundance. The remaining activity in the presence of erythromycin was similar in the four experiments (Fig. 7) despite archaeal abundances ranging from 1 to 9% (Table 3). Therefore, we were unable to conclusively demonstrate archaeal-specific leucine incorporation. One potential expla- nation could be that the bacterial antibiotic erythromycin also inhibits marine archaea. However, resistance to this an- tibiotic has been attributed to the presence of a specific nu- cleic acid residue in the 23s rRNA (Amils et al. 1993), and group I marine archaea have the resistant genotype in their large subunit rRNA (DeLong unpubl. results). Another pos- sible explanation is that marine archaea do not generally
incorporate leucine significantly. Finally, it may also be that archaeal picoplankton were not active in the field (or were less active than their biomass suggests) or were affected by our sampling and filtration procedure.
We are currently investigating several other approaches to assess archaeal activity in natural assemblages. One strategy involves incorporation of acetate into microbial biomass and subsequent lipid extraction and fractionation in order to quantify acetate incorporation into archaeal lipids. This ap- proach assumes that marine archaea possess membrane lip- ids similar to the lipids of known, cultivated archaea, based in ether-linked phytanyl chains (Kates 1993).
This report, together with a parallel study in coastal waters near Anvers Island (Murray et al. in press) and previous results obtained from a totally different ecosystem (Massana et al. 1997a), verifies that archaea are widespread in marine waters and compose a significant fraction of marine pico- plankton biomass. The general properties of characterized and cultivated archaea suggest that the biochemical and physiological attributes of planktonic marine archaea may be unique. Their distribution and abundance are of potential biogeochemical significance. The next step in understanding the ecology and biology of these ubiquitous microbes will likely involve correlating their distributions in time and space with specific biogeochemical activities and ecological processes.
References
ALM, E. W., D. B. OERTHER, N. LARSEN, D. A. STAHL, AND L. RASKIN. 1996. The oligonucleotide probe database. Appl. En- viron. Microbial. 62: 3557-3559.
AMILS, R., P CAMMARANO, AND P LONDEI. 1993. Translation in Archaea, p. 393-438. In M. Kates et al. [eds.], The biochem- istry of the Archaea (Archaebacteria). Elsevier.
AMOS, A. E, S. S. JACOBS, AND J.-H. Hu. 1990. RACER: Hydrog- raphy of the surface waters during the spring bloom in the Gerlache Strait. Antarct. J. U.S. 25: 131-134.
AZAM, E, T. FENCHEL, J. G. FIELD, J. S. GRAY, L. A. MEYER-RI% AND E THINGSTAD. 1983. The ecological role of water-column microbes in the sea. Mar. Ecol. Prog. Ser. 10: 257-263.
BIRD, D. E, AND D. M. KARL. 1990. RACER: Bacterial growth, abundance, and loss due to protozoan grazing during the 1989 spring bloom. Antarct. J. U.S. 25: 156-157.
B~RSHEIM, K. Y., AND G. BRATBAK. 1987. Cell volume to cell carbon conversion factors for a bacterivorous Monas sp. en- riched from seawater. Mar. Ecol. Prog. Ser. 36: 171-175.
BRITSCHGI, T. B., AND S. J. GIOVANNONI. 1991. Phylogenetic anal- ysis of a natural marine bacterioplankton population by rRNA gene cloning and sequencing. Appl. Environ. Microbial. 57: 1707-1713.
COCHLAN, W. I?, J. MARTINEZ, AND 0. HOLM-HANSEN. 1993. RACER: Primary production in Gerlache Strait, Antarctica, during austral winter. Antarct. J. U.S. 28: 172-174.
COLE, J. J., S. FINDLAY, AND M. L. PACE. 1988. Bacterial produc- tion in fresh and saltwater ecosystems: A cross-system over- view. Mar. Ecol. Prog. Ser. 43: l-10.
DELONG, E. E 1992. Archaea in coastal marine environments. Proc. Natl. Acad. Sci. USA 89: 5685-5689.
-, L. L. KING, R. MASSANA, H. CITTONE, A. MURRAY, C. SCHLEPER, AND S. G. WAKEHAM. 1998. Dibyphytanylether lipids in nonthermophilic crenarchaeotes. Appl. Environ. Mi- crobiol. 64: 1133-l 138.

Archaea in Antarctic waters 617
-, K. Y. Wu, B. B. PRÉZELIN, AND R. V. M. JOVINE. 1994. High abundance of Archaea in Antarctic marine picoplankton. Nature 371: 695-697.
FENCHEL, T 1988. Marine plankton food chains. Ann. Rev. Ecol. Syst. 19: 19-38.
FUHRMAN, J. A., K. MCCALLUM, AND A. A. DAVIS. 1992. Novel major archaebacterial group from marine plankton. Nature 356: 148-149.
,-,AND-. 1993. Phylogenetic diversity of sub- surface marine microbial communities from the Atlantic and Pacific Oceans. Appl. Environ. Microbial. 59: 1294-1302.
GIOVANNONI, S. J., T. B. BRITSCHGI, C. L. MOYER, AND K. G. FIELD. 1990. Genetic diversity in Sargasso Sea bacterioplank- ton. Nature 345: 60-63.
HEWES, C. D., E. SAKSHAUG, E M. H. REID, AND 0. HOLM-HANSEN. 1990. Microbial autotrophic and heterotrophic eucaryotes in Antarctic waters: Relationships between biomass and chloro- phyll, adenosine triphosphate and particulate organic carbon. Mar. Ecol. Prog. Ser. 63: 27-35.
HOLM-HANSEN, O., AND B. G. MITCHEI.L. 1991. Spatial and tem- poral distribution of phytoplankton and primary production in the western Bransfield Strait region. Deep-Sea Res. 38: 961- 980.
, AND M. VERNET. 1990. RACER: Phytoplankton distribu- tion and rates of primary production during the austral spring bloom. Antarct. J. U.S. 25: 141-144.
HUNTLEY, M., D. M. KARL, I? NIILER, AND 0. HOLM-HANSEN. 1991. Research on Antarctic coastal ecosystem rates (RAC- ER): An interdisciplinary field experiment. Deep-Sea Res. 38: 911-941.
-,I? NIILER,~. HOLM-HANSEN, M.VERNET,E.BRINTON,A. E AMOS, AND D. M. KARL. 1990. RACER: An interdisciplin- ary study of spring bloom dynamics. Antarct. J. U.S. 25: 126 128.
KARL, D. M. 1993. Microbial processes in the Southern Oceans, p. l-63. In E. I. Friedmann [ed.], Antarctic microbiology, Wi- ley-Liss.
-, 0. HOLM-HANSEN, G. T TAYLOR, G. TIEN, AND D. E BIRD. 1991. Microbial biomass and productivity in the western Bransfield Strait, Antarctica during the 1986-87 austral sum- mer. Deep-Sea Res. 38: 1029-1055.
KATES, M. 1993. Membrane lipids of archaea, p. 261-295. In M. Kates et al. [eds.], The biochemistry of Archaea (Archaebac- teria). Elsevier.
KIRCHMAN, D. L. 1993. Leucine incorporation as a measure of biomass production by heterotrophic bacteria, p. 509-512. Zn l? E Kemp et al. [eds.], Handbook of methods in aquatic mi- crobial ecology. Lewis Publ.
LETELIER, R. M., AND D. M. KARL. 1989. Phycoerythrin-contain- ing cyanobacteria in surface waters of Drake Passage during February 1987. Antarct. J. U.S. 24: 185-188.
LORENZEN, C. J. 1967. Carbon/chlorophyll relationships in an up- welling area. Limnol. Oceanogr. 13: 202-204.
MASSANA, R., A. E. MURRAY, C. M. PRESTON, AND E. E DELONG. 1997~. Vertical distribution and phylogenetic characterization of marine planktonic Archaea in the Santa Barbara Channel. Appl. Environ. Microbial. 63: 50-56.
-, AND OTHERS. 19976. Measurement of bacterial size via image analysis of epifluorescence preparations: Description of
an inexpensive system and solutions to some of the most com- mon problems. Sci. Mar. 61: 397-407.
MULLINS, T D., T. B. BRITSCHGI, R. L. KREST, AND S. J. Glo- VANNONI. 1995. Genetic comparisons reveal the same un- known bacterial lineages in Atlantic and Pacitic bacterioplank- ton communities. Limnol. Oceanogr. 40: 148-158.
MURRAY, A. E., AND OTHERS. In press. Seasonal and spatial vari- ability of bacterial and archaeal assemblages in the coastal wa- ters off Anvers Island, Antarctica. Appl. Environ. Microbial.
NIILER, I? P., A. AMOS, AND J.-H. Hu. 1991. Water masses and 200 m relative geostrophic circulation in the western Branstield Strait region. Deep-Sea Res. 38: 943-959.
-, J. ILLEMAN, AND J.-H. Hu. 1990. RACER: Lagrangian drifter observations of surface circulations in the Gerlache and Bransfield Straits. Antarct. J. U.S. 25: 134-137.
OREMLAND, R. S., AND D. G. CAPONE. 1988. Use of “specifc” inhibitors in biogeochemistry and microbial ecology. Adv. Mi- crab. Ecol. 10: 285-383.
PAKULSKI, J. D., AND OTHERS. 1996. Iron stimulation of Antarctic bacteria. Nature 383: 133-l 34.
PARSONS, T R., Y. MAITA, AND C. M. LAIN. 1984. A manual of chemical and biological methods for seawater analysis. Per- gamon.
PAUL, J. H., AND B. MYERS. 1982. Fluorometric determination of DNA in aquatic microorganisms by use of Hoechst 33258. Appl. Environ. Microbial. 43: 1393-1399.
POLZ, M. F., AND C. M. CAVANAUGH. 1997. A simple method for quantification of uncultured microorganisms in the environ- ment based on in vitro transcription of 16S rRNA. Appl. En- viron. Microbial. 63: 1028-1033.
PORTER, K. G., AND Y. S. FEIG. 1980. The use of DAPI for iden- tifying and counting aquatic microflora. Limnol. Oceanogr. 25: 943-948.
PRESTON, C. M., K. Y. WV, T. E MOLINSKI, ANI) E. E DELoN(;. 1996. A psychrophilic crenarchaeon inhabits a marine sponge: Cenarchaeum symbiosum gen. nov., sp. nov. Proc. Natl. Acad. Sci. USA 93: 6241-6246.
SCHMIDT, ‘I M., E. E DELONG, AND N. R. PACE. 1991. Analysis of a marine picoplankton community by 16S rRNA gene clon- ing and sequencing. J. Bacterial. 173: 4371-4378.
SIMON, M., AND E AZAM. 1989. Protein content and protein syn- thesis rates of planktonic marine bacteria. Mar. Ecol. Prog. Ser. 51: 201-213.
SOMERVILLE,~. C., I. T. KNIGHT, W. L. STRAUBE, AND R. R. COL- WELL. 1989. Simple, rapid method for direct isolation of nu- cleic acids from aquatic environments. Appl. Environ. Micro- biol. 55: 548-554.
STAHL, D. A., B. FLESHER, H. R. MANSFIELD, AND L. M~NTGOM- ERY. 1988. Use of phylogenetically based hybridization probes for studies of ruminal microbial ecology. Appl. Environ. Mi- crobiol. 54: 1079-1084.
VINCENT, W. E 1988. Microbial ecosystems of Antarctica. Cam- bridge Univ. Press.
ZHENG, D., E. W. ALM, D. A. STAHL, AND L. RASKIN. 1996. Char- acterization of universal small-subunit rRNA hybridization probes for quantitative molecular microbial ecology studies. Appl. Environ. Microbial. 62: 4504-45 13.
Received: 27 June 1997 Accepted: 28 October 1997




















