
Vascular plant diversity and climate change in the alpine zone of the Snowy Mountains, Australia
11
This article was downloaded by: [UGR-BTCA Gral Universitaria] On: 21 November 2012, At: 00:53 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Plant Biosystems - An International Journal Dealing with all Aspects of Plant Biology: Official Journal of the Societa Botanica Italiana Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/tplb20 Vascular plant diversity and climate change in the upper zone of Sierra Nevada, Spain M. R Fernández Calzado a , J. Molero Mesa a , A. Merzouki a & M. Casares Porcel a a Faculty of Pharmacy, Department of Botany, University of Granada, Campus de Cartuja, 18071, Granada, Spain Accepted author version posted online: 13 Jul 2012.Version of record first published: 17 Aug 2012. To cite this article: M. R Fernández Calzado, J. Molero Mesa, A. Merzouki & M. Casares Porcel (2012): Vascular plant diversity and climate change in the upper zone of Sierra Nevada, Spain, Plant Biosystems - An International Journal Dealing with all Aspects of Plant Biology: Official Journal of the Societa Botanica Italiana, 146:4, 1044-1053 To link to this article: http://dx.doi.org/10.1080/11263504.2012.710273 PLEASE SCROLL DOWN FOR ARTICLE Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. The publisher does not give any warranty express or implied or make any representation that the contents will be complete or accurate or up to date. The accuracy of any instructions, formulae, and drug doses should be independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings, demand, or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with or arising out of the use of this material.
Transcript of Vascular plant diversity and climate change in the alpine zone of the Snowy Mountains, Australia

This article was downloaded by: [UGR-BTCA Gral Universitaria]On: 21 November 2012, At: 00:53Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Plant Biosystems - An International Journal Dealingwith all Aspects of Plant Biology: Official Journal of theSocieta Botanica ItalianaPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/tplb20
Vascular plant diversity and climate change in theupper zone of Sierra Nevada, SpainM. R Fernández Calzado a , J. Molero Mesa a , A. Merzouki a & M. Casares Porcel aa Faculty of Pharmacy, Department of Botany, University of Granada, Campus de Cartuja,18071, Granada, SpainAccepted author version posted online: 13 Jul 2012.Version of record first published: 17 Aug2012.
To cite this article: M. R Fernández Calzado, J. Molero Mesa, A. Merzouki & M. Casares Porcel (2012): Vascular plant diversityand climate change in the upper zone of Sierra Nevada, Spain, Plant Biosystems - An International Journal Dealing with allAspects of Plant Biology: Official Journal of the Societa Botanica Italiana, 146:4, 1044-1053
To link to this article: http://dx.doi.org/10.1080/11263504.2012.710273
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form toanyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae, and drug doses shouldbe independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims,proceedings, demand, or costs or damages whatsoever or howsoever caused arising directly or indirectly inconnection with or arising out of the use of this material.

Vascular plant diversity and climate change in the upper zone ofSierra Nevada, Spain
M. R. FERNANDEZ CALZADO, J. MOLERO MESA, A. MERZOUKI, &
M. CASARES PORCEL
Faculty of Pharmacy, Department of Botany, University of Granada, Campus de Cartuja, 18071, Granada, Spain
AbstractThis study examines the effects of altitudinal, temperature and aspect gradients on vascular plant species richness onmountain tops in Sierra Nevada (Spain) at different spatial scales (1 m2 quadrats, plot clusters of 4 m2, upper summit areadown to the 5-m contour line, entire summit down to the 10-m contour line). The methodology follows the GlobalObservation Research Initiative in Alpine Environments (GLORIA) programme. Floristic and soil temperature data of eightsummits sites in two neighbouring regions of the high part of Sierra Nevada (from 2668 m to 3327 m a.s.l.) were used in thisstudy. In total, 102 taxa were recorded (84 genera; 29 families). The species richness decreased, whereas the proportion ofendemic taxa increased with elevation. There were significant linear relationships between species richness and altitude andaverage soil temperature at each spatial scale. However, there was no significant relationship between species richness andaspect variables. Facing continued climate change, the high-altitude flora of Sierra Nevada is expected to be particularlyvulnerable and prone to warming-induced biodiversity losses due to the high proportion of endemic taxa, ranging from 23%at lower elevations up to 67% at higher ones.
Keywords: Altitude gradient, biodiversity, climate change, endemics plants, mountain, temperature
Introduction
Since the last century, the influence of climate change
on local-large scale biodiversity pattern has been
emphasized by several studies. Warming-induced
species migrations lead to latitudinal and altitudinal
shifts in species’ distribution ranges and may result in
species extinctions, especially in highly fragmented
ecosystems (Ozenda & Borel 1991; Grabherr et al.
1994; Hansen et al. 2001; Houghton et al. 2001;
Pearson & Dawson 2003; Araujo et al. 2005a,b;
Broennimann et al. 2006; Thuiller et al. 2006).
In this context, high mountain regions are of
particular interest as they usually harbour small-
scaled and fragmented ecosystems and, in some
regions, species with strongly restricted distribution
ranges (Nagy & Grabherr 2009). High mountain
areas appear to be very suitable places to study
the impacts of current climate change on natural
ecosystems. This is because they exhibit steep
ecological gradients and narrow ecotones dominated
by abiotic factors in relation to climate conditions,
and scarce anthropogenic disturbance compared to
lower altitudes (Theurillat 1995; Grabherr et al.
1994, 2001; Korner 1994, 2002; Pauli et al. 1996,
2004; Dullinger et al. 2007). Therefore, the long-
term monitoring programme GLORIA (Global
Observation Research Initiative in Alpine Environ-
ments: www.gloria.ac.at) was initiated at the turn of
the century.
The field work of GLORIA started in 2001 by
applying a common monitoring protocol in summit
areas of 18 European mountain regions with Sierra
Nevada among them. This protocol involved the
recording of the composition and cover of vascular
plant species and soil temperatures (Grabherr et al.
2000; Pauli et al. 2003, 2004). The peculiarities of
the highest zone of Sierra Nevada (geographical
location, Mediterranean climate, high floristic biodi-
versity and endemism rate) and its being considered
as one of the most sensible and vulnerable zones to
climate change effects in Spain (Fernandez-Gonzalez
et al. 2005) motivated us to extend the number of
summit sites (Molero Mesa et al. 2009). Currently,
Correspondence: M. R. Fernandez Calzado, Department of Botany, Faculty of Pharmacy, University of Granada, Campus de Cartuja, 18071, Granada, Spain.
Tel: 0034 607 678 047. Fax: 0034 958 243 916. Email: [email protected]
Plant Biosystems, Vol. 146, No. 4, December 2012, pp. 1044–1053
ISSN 1126-3504 print/ISSN 1724-5575 online ª 2012 Societa Botanica Italiana
http://dx.doi.org/10.1080/11263504.2012.710273
Dow
nloa
ded
by [
UG
R-B
TC
A G
ral U
nive
rsita
ria]
at 0
0:53
21
Nov
embe
r 20
12
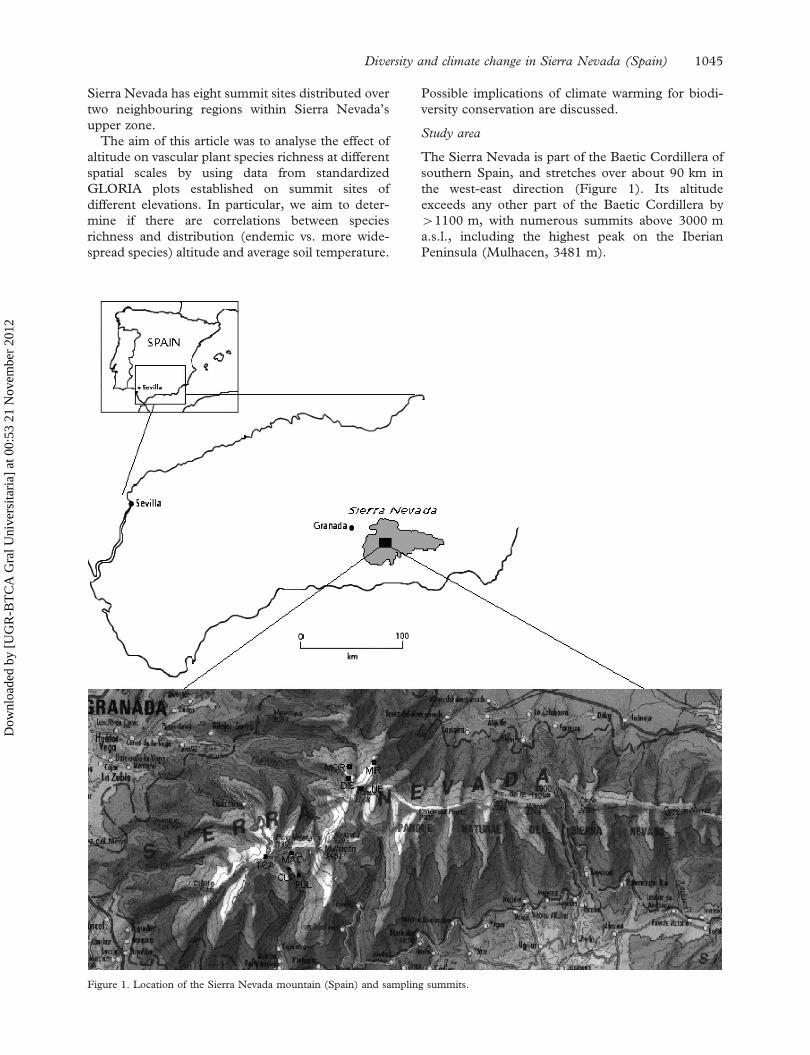
Sierra Nevada has eight summit sites distributed over
two neighbouring regions within Sierra Nevada’s
upper zone.
The aim of this article was to analyse the effect of
altitude on vascular plant species richness at different
spatial scales by using data from standardized
GLORIA plots established on summit sites of
different elevations. In particular, we aim to deter-
mine if there are correlations between species
richness and distribution (endemic vs. more wide-
spread species) altitude and average soil temperature.
Possible implications of climate warming for biodi-
versity conservation are discussed.
Study area
The Sierra Nevada is part of the Baetic Cordillera of
southern Spain, and stretches over about 90 km in
the west-east direction (Figure 1). Its altitude
exceeds any other part of the Baetic Cordillera by
41100 m, with numerous summits above 3000 m
a.s.l., including the highest peak on the Iberian
Peninsula (Mulhacen, 3481 m).
Figure 1. Location of the Sierra Nevada mountain (Spain) and sampling summits.
Diversity and climate change in Sierra Nevada (Spain) 1045
Dow
nloa
ded
by [
UG
R-B
TC
A G
ral U
nive
rsita
ria]
at 0
0:53
21
Nov
embe
r 20
12

Eight summits, assembled in two neighbouring
regions (target regions) of the high part of Sierra
Nevada, were selected and permanently marked
(Figure 1), using criteria specified in the GLORIA
field manual (Pauli et al. 2004). The Sierra Nevada
west target region (coded ES-SNE) is situated in the
central-occidental zone of the range, with four
summit sites: Machos (MAC) 3327 m; Tosal Car-
tujo (TCA) 3150 m; Cupula (CUP) 2968 m and
Pulpitito (PUL) 2778 m. The second target region,
Sierra Nevada northeast (coded ES-SNN), is located
in the oriental part of the high Sierra Nevada, also
with four summit sites: Cuervo (CUE) 3144 m;
Diegisa (DIE) 2800 m; Miron (MIR) 2717 m and
Monte Rosa (MOR) 2668 m. Species data from the
ES-SNE sites were from 2001, and those from the
ES-SNN sites from 2004 to 2006.
All of the summits are exposed to the same local
climate, namely a Mediterranean bioclimate in its
pluviseasonal oceanic variant (Rivas-Martınez et al.
2007), with a pronounced summer drought at all
altitudes and with precipitation occurring almost
exclusively as snow during winter at altitudes above
2500 m. They also have similar siliceous bedrock
except for PUL, having a less acidic substrate. The
vegetation changes between the lower summits (i.e.
PUL or MOR) where dwarf shrub communities are
predominant in the landscape, and in the highest
summits such us MAC, TCA, CUE, where the
vegetation is composed of scattered psycroxerophytic
grasses and scree plants.
Materials and methods
Sampling design
The sampling design and the recording methods
follow the GLORIA field manual (Pauli et al. 2004).
Firstly, the survey area on each summit was defined
as a polygon with four corners, at each point of the
compass (N, S, E and W), and at an altitude
difference of 5 m form the highest summit point
(HSP) and a lower area extending from five vertical
metres below the summit to 10 vertical metres below
the summit. Intersection lines from the HSP to the
NE, SE, SW and NW directions resulted in eight
summit area sections (SASs; four upper and four
lower ones). Recording in each SAS included a
complete list of all vascular plants species, and
visually estimated percentage top cover of surface
types such as total vegetation cover (vascular plants),
solid rock, scree, etc.
Secondly, to evaluate the quantitative floristic
compositions at each aspect, four quadrat clusters
(3 m63 m resulting in nine 1-m2 quadrats) were
established at each cardinal direction (N, E, S, W).
The vegetation sampling (vascular plant species
and their percentage cover) was recorded in the
four corners-quadrats of each 3 m63 m quadrat
cluster.
Thirdly, data-loggers (StowAway TidbiT, Onset
Corporation, MA, USA) were installed at 10 cm
below the surface in each central quadrat in the
3 m63 m quadrat clusters, to measure the soil
temperature every hour. Temperature was recorded
from August 2001 to July 2008 in the ES-SNE target
region, and from October 2005 to March 2008 in the
ES-SNN target region.
Data analysis
The effect of aspect and altitude on species richness
at each spatial scale, i.e. (1 m2 and 4 m2 of one plot
cluster in the upper summit area (from the top down
to the 5-m level), and the entire summit area (from
the top to the 10-m level) was tested by Analysis of
Covariance (ANCOVA) with altitude as the covari-
ate. Linear regression was used to analyse the
relation between species richness, altitude and
average soil temperature. Both analyses were per-
formed using the statistical package SPSS.
Flora Iberica (Castroviejo et al. 1986–2009) and
regional floras (Molero Mesa & Perez-Raya 1987;
Molero Mesa et al. 1996) were used as references to
define the distribution and the altitudinal ranges of
species.
Soil temperature data (hourly time series from
October 2006 to September 2007) were used in this
analysis as mean annual temperature. This was the
first time period in which vegetation sampling was
done on all eight summits.
Results
Species richness and life forms
The total number of vascular plants species of all
eight summit sites (entire summit areas down to the
10-m contour line) was 102, belonging to 84 genera
(Table I). It was impossible to distinguish the species
in five genera (Cerastium, Rumex, Viola, Veronica and
Helictotrichon) across different summits on the ES-
SNN. The taxa are included within 29 families, the
most common being the typical Mediterranean
families such as Asteraceae (17 species), Poaceae
(15), Brassicaceae (11) and Caryophyllaceae (10).
The altitudinal distribution of the species found on
the eight GLORIA summits in Sierra Nevada range
from almost sea level to a maximum altitude of
around 3500 m a.s.l., but shows a pronounced peak
between 2500 and 3000 m (Figure 2). The total
vascular plant species richness on the study summits
decreased with increasing elevation, with the excep-
tion of MIR (2717 m) and CUP (2968 m), which
1046 M. R. Fernandez Calzado et al.
Dow
nloa
ded
by [
UG
R-B
TC
A G
ral U
nive
rsita
ria]
at 0
0:53
21
Nov
embe
r 20
12
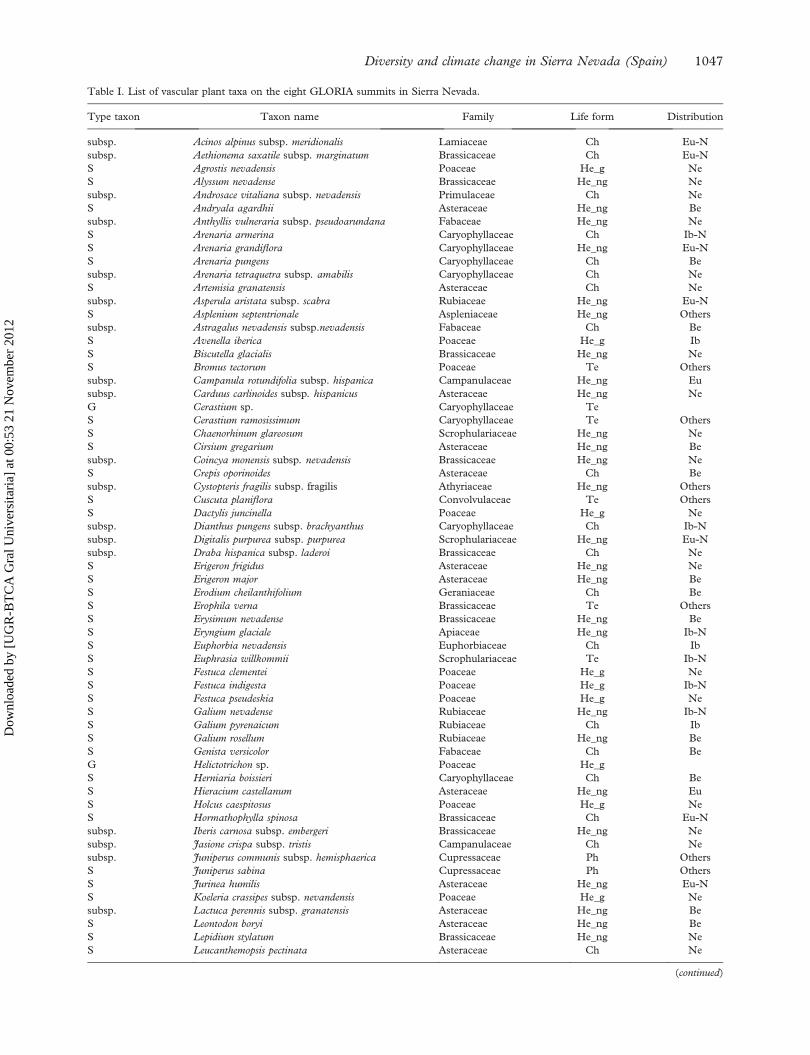
Table I. List of vascular plant taxa on the eight GLORIA summits in Sierra Nevada.
Type taxon Taxon name Family Life form Distribution
subsp. Acinos alpinus subsp. meridionalis Lamiaceae Ch Eu-N
subsp. Aethionema saxatile subsp. marginatum Brassicaceae Ch Eu-N
S Agrostis nevadensis Poaceae He_g Ne
S Alyssum nevadense Brassicaceae He_ng Ne
subsp. Androsace vitaliana subsp. nevadensis Primulaceae Ch Ne
S Andryala agardhii Asteraceae He_ng Be
subsp. Anthyllis vulneraria subsp. pseudoarundana Fabaceae He_ng Ne
S Arenaria armerina Caryophyllaceae Ch Ib-N
S Arenaria grandiflora Caryophyllaceae He_ng Eu-N
S Arenaria pungens Caryophyllaceae Ch Be
subsp. Arenaria tetraquetra subsp. amabilis Caryophyllaceae Ch Ne
S Artemisia granatensis Asteraceae Ch Ne
subsp. Asperula aristata subsp. scabra Rubiaceae He_ng Eu-N
S Asplenium septentrionale Aspleniaceae He_ng Others
subsp. Astragalus nevadensis subsp.nevadensis Fabaceae Ch Be
S Avenella iberica Poaceae He_g Ib
S Biscutella glacialis Brassicaceae He_ng Ne
S Bromus tectorum Poaceae Te Others
subsp. Campanula rotundifolia subsp. hispanica Campanulaceae He_ng Eu
subsp. Carduus carlinoides subsp. hispanicus Asteraceae He_ng Ne
G Cerastium sp. Caryophyllaceae Te
S Cerastium ramosissimum Caryophyllaceae Te Others
S Chaenorhinum glareosum Scrophulariaceae He_ng Ne
S Cirsium gregarium Asteraceae He_ng Be
subsp. Coincya monensis subsp. nevadensis Brassicaceae He_ng Ne
S Crepis oporinoides Asteraceae Ch Be
subsp. Cystopteris fragilis subsp. fragilis Athyriaceae He_ng Others
S Cuscuta planiflora Convolvulaceae Te Others
S Dactylis juncinella Poaceae He_g Ne
subsp. Dianthus pungens subsp. brachyanthus Caryophyllaceae Ch Ib-N
subsp. Digitalis purpurea subsp. purpurea Scrophulariaceae He_ng Eu-N
subsp. Draba hispanica subsp. laderoi Brassicaceae Ch Ne
S Erigeron frigidus Asteraceae He_ng Ne
S Erigeron major Asteraceae He_ng Be
S Erodium cheilanthifolium Geraniaceae Ch Be
S Erophila verna Brassicaceae Te Others
S Erysimum nevadense Brassicaceae He_ng Be
S Eryngium glaciale Apiaceae He_ng Ib-N
S Euphorbia nevadensis Euphorbiaceae Ch Ib
S Euphrasia willkommii Scrophulariaceae Te Ib-N
S Festuca clementei Poaceae He_g Ne
S Festuca indigesta Poaceae He_g Ib-N
S Festuca pseudeskia Poaceae He_g Ne
S Galium nevadense Rubiaceae He_ng Ib-N
S Galium pyrenaicum Rubiaceae Ch Ib
S Galium rosellum Rubiaceae He_ng Be
S Genista versicolor Fabaceae Ch Be
G Helictotrichon sp. Poaceae He_g
S Herniaria boissieri Caryophyllaceae Ch Be
S Hieracium castellanum Asteraceae He_ng Eu
S Holcus caespitosus Poaceae He_g Ne
S Hormathophylla spinosa Brassicaceae Ch Eu-N
subsp. Iberis carnosa subsp. embergeri Brassicaceae He_ng Ne
subsp. Jasione crispa subsp. tristis Campanulaceae Ch Ne
subsp. Juniperus communis subsp. hemisphaerica Cupressaceae Ph Others
S Juniperus sabina Cupressaceae Ph Others
S Jurinea humilis Asteraceae He_ng Eu-N
S Koeleria crassipes subsp. nevandensis Poaceae He_g Ne
subsp. Lactuca perennis subsp. granatensis Asteraceae He_ng Be
S Leontodon boryi Asteraceae He_ng Be
S Lepidium stylatum Brassicaceae He_ng Ne
S Leucanthemopsis pectinata Asteraceae Ch Ne
(continued)
Diversity and climate change in Sierra Nevada (Spain) 1047
Dow
nloa
ded
by [
UG
R-B
TC
A G
ral U
nive
rsita
ria]
at 0
0:53
21
Nov
embe
r 20
12

showed slightly higher numbers of species than the
lowest sites.
The life form spectrum over all sites revealed the
predominant role of hemicryptophytes (52 taxa: 13
graminoid and 39 non graminoid) and chamaephytes
(36 taxa) (Table I). Hemicryptophytes grow in
number with increasing altitude reaching 55.6% in
MAC, whereas chamaephytes have similar percen-
tages on the eight summits. The number of taxa
belonging to the phanerophytes declines along the
elevation gradient, becoming absent on the three
highest summits, while therophytes occurr on all the
summits, except at the highest one, with percentages
ranging between 1.9% and 20%.
Biogeographic elements and endemism
Out of the total number of vascular plant species
(except for the five unidentified ones), 35% are
endemic to Sierra Nevada (Ne), 16% to the Baetic
range of southern Spain (Be), 9% are restricted to
the Iberian Peninsula (Ib) and 7% are present in the
Iberian Peninsula and in North Africa (Ib-N), 5%
are more widely distributed in Europe (Eu) and 9%
in Europe as well as in North Africa (Eu-N). The
remaining taxa (Others, 17%) have an even wider
distribution (compare Table I).
The proportion of taxa from each of the seven
biogeographic elements on a summit varied with
Table I. (Continued).
Type taxon Taxon name Family Life form Distribution
S Linaria nevadensis Scrophulariaceae He_ng Ne
S Logfia arvensis Asteraceae Te Eu
subsp. Lotus corniculatus subsp. glacialis Fabaceae Ch Ne
S Luzula hispanica Juncaceae He_ng Ib
S Myosotis minutiflora Borraginaceae Te Others
S Nevadensia purpurea Brassicaceae He_ng Ne
S Plantago nivalis Plantaginaceae He_ng Ne
S Plantago holosteum Plantaginaceae Ch Eu
subsp. Nepeta amethystina subsp. laciniata Lamiaceae Ch Ne
S Paronychia polygonifolia Caryophyllaceae Ch Others
S Pimpinella procumbens Apiaceae He_ng Ne
S Poa ligulata Poaceae He_g Ib-N
subsp. Poa minor subsp. nevadensis Poaceae He_g Ne
S Poa nemoralis Poaceae He_g Eu
S Prunus prostrata Rosaceae Ch Others
S Polygonum aviculare Polygonaceae Te Others
S Potentilla reuteri Rosaceae He_ng Be
S Rhamnus pumila Rhamnaceae Ch Eu-N
S Ranunculus demissus Ranunculaceae He_ng Others
S Reseda complicata Resedaceae Ch Ne
G Rumex sp. Polygonaceae Te
S Saxifraga granulata Saxifragaceace He_ng Others
S Saxifraga nevadensis Saxifragaceace Ch Ne
S Scutellaria javalambrensis Lamiaceae He_ng Ib
S Sedum amplexicaule Crassulaceae Ch Others
S Sedum dasyphyllum Crassulaceae Ch Eu-N
S Sempervivum minutum Crassulaceae Ch Be
S Senecio boissieri Asteraceae Ch Ib
S Senecio nebrodensis Asteraceae He_ng Ib
subsp. Senecio pyrenaicus subsp. granatensis Asteraceae Ch Be
S Sideritis glacialis Lamiaceae Ch Ne
S Silene boryi Caryophyllaceae He_ng Ib
S Solidago virgaurea Asteraceae He_ng Others
S Teucrium aureum subsp. angustifolium Lamiaceae Ch Ib
S Thymus serpylloides Lamiaceae Ch Ne
S Trisetum glaciale Poaceae He_g Ne
S Urtica dioica Urticaceae Ch Others
G Veronica sp. Scrophulariaceae Te
G Viola sp. Violaceae Te
S Viola crassiuscula Violaceae He_ng Ne
Notes: Type of taxon: S¼ species; G¼ genus; subsp.¼ subspecies. Life form: Ph¼ phanerophytes, Ch¼ chamaephytes, He_g¼ graminoid
hemicryptophytes, He_ng¼non graminoid hemicryptophytes, Ge¼ geophytes, Te¼ therophytes. Distribution: Ne¼Nevadense (restricted
to Sierra Nevada); Be¼Baetican; Ib¼ Iberian; Ib-N¼ Iberian-North African; Eu¼European; Eu-N¼European-North African;
Others¼more widely distributed.
1048 M. R. Fernandez Calzado et al.
Dow
nloa
ded
by [
UG
R-B
TC
A G
ral U
nive
rsita
ria]
at 0
0:53
21
Nov
embe
r 20
12
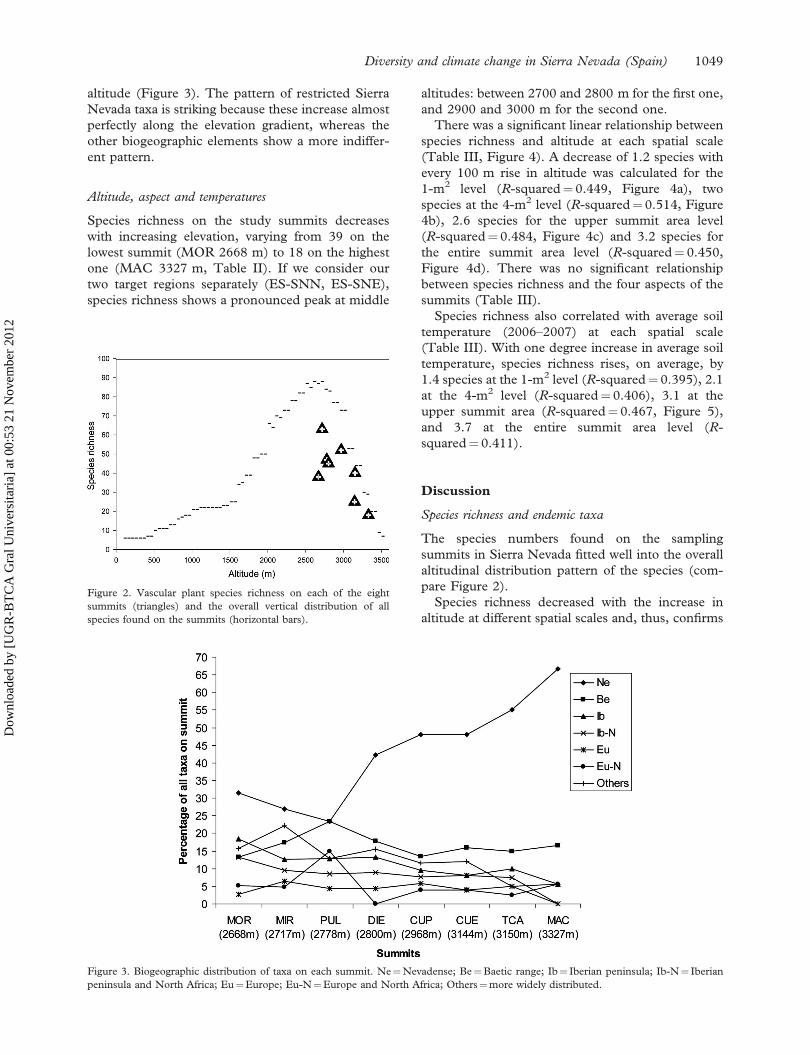
altitude (Figure 3). The pattern of restricted Sierra
Nevada taxa is striking because these increase almost
perfectly along the elevation gradient, whereas the
other biogeographic elements show a more indiffer-
ent pattern.
Altitude, aspect and temperatures
Species richness on the study summits decreases
with increasing elevation, varying from 39 on the
lowest summit (MOR 2668 m) to 18 on the highest
one (MAC 3327 m, Table II). If we consider our
two target regions separately (ES-SNN, ES-SNE),
species richness shows a pronounced peak at middle
altitudes: between 2700 and 2800 m for the first one,
and 2900 and 3000 m for the second one.
There was a significant linear relationship between
species richness and altitude at each spatial scale
(Table III, Figure 4). A decrease of 1.2 species with
every 100 m rise in altitude was calculated for the
1-m2 level (R-squared¼ 0.449, Figure 4a), two
species at the 4-m2 level (R-squared¼ 0.514, Figure
4b), 2.6 species for the upper summit area level
(R-squared¼ 0.484, Figure 4c) and 3.2 species for
the entire summit area level (R-squared¼ 0.450,
Figure 4d). There was no significant relationship
between species richness and the four aspects of the
summits (Table III).
Species richness also correlated with average soil
temperature (2006–2007) at each spatial scale
(Table III). With one degree increase in average soil
temperature, species richness rises, on average, by
1.4 species at the 1-m2 level (R-squared¼ 0.395), 2.1
at the 4-m2 level (R-squared¼ 0.406), 3.1 at the
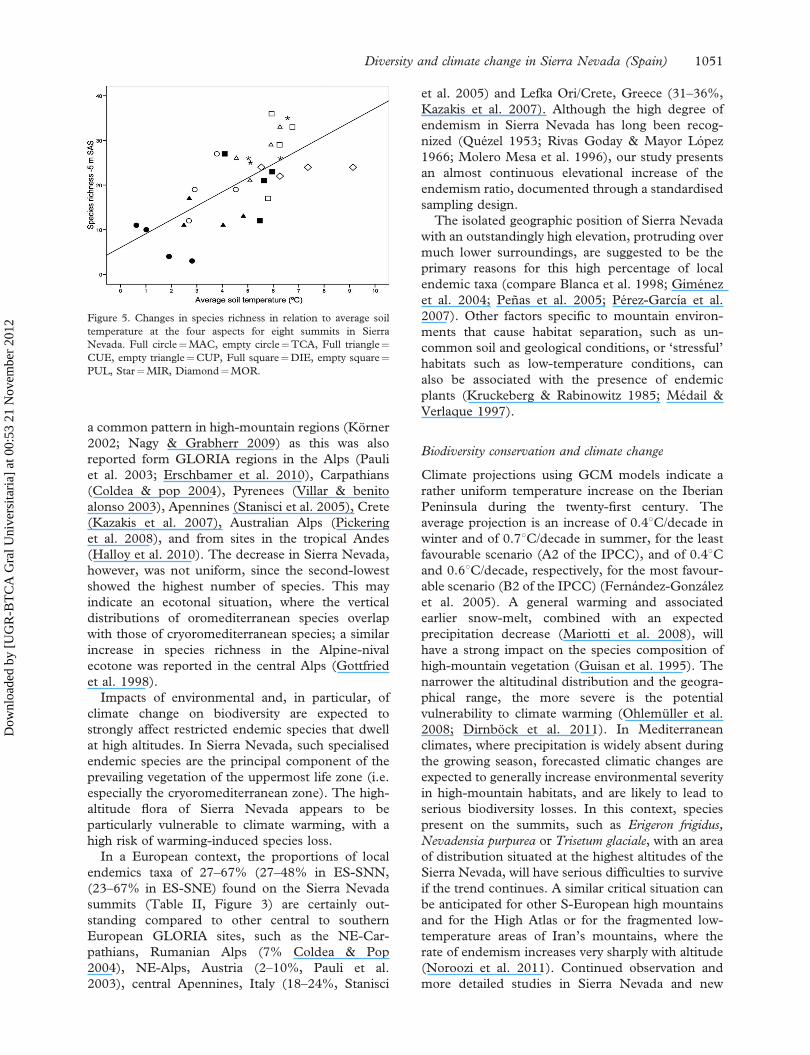
upper summit area (R-squared¼ 0.467, Figure 5),
and 3.7 at the entire summit area level (R-
squared¼ 0.411).
Discussion
Species richness and endemic taxa
The species numbers found on the sampling
summits in Sierra Nevada fitted well into the overall
altitudinal distribution pattern of the species (com-
pare Figure 2).
Species richness decreased with the increase in
altitude at different spatial scales and, thus, confirms
Figure 2. Vascular plant species richness on each of the eight
summits (triangles) and the overall vertical distribution of all
species found on the summits (horizontal bars).
Figure 3. Biogeographic distribution of taxa on each summit. Ne¼Nevadense; Be¼Baetic range; Ib¼ Iberian peninsula; Ib-N¼ Iberian
peninsula and North Africa; Eu¼Europe; Eu-N¼Europe and North Africa; Others¼more widely distributed.
Diversity and climate change in Sierra Nevada (Spain) 1049
Dow
nloa
ded
by [
UG
R-B
TC
A G
ral U
nive
rsita
ria]
at 0
0:53
21
Nov
embe
r 20
12
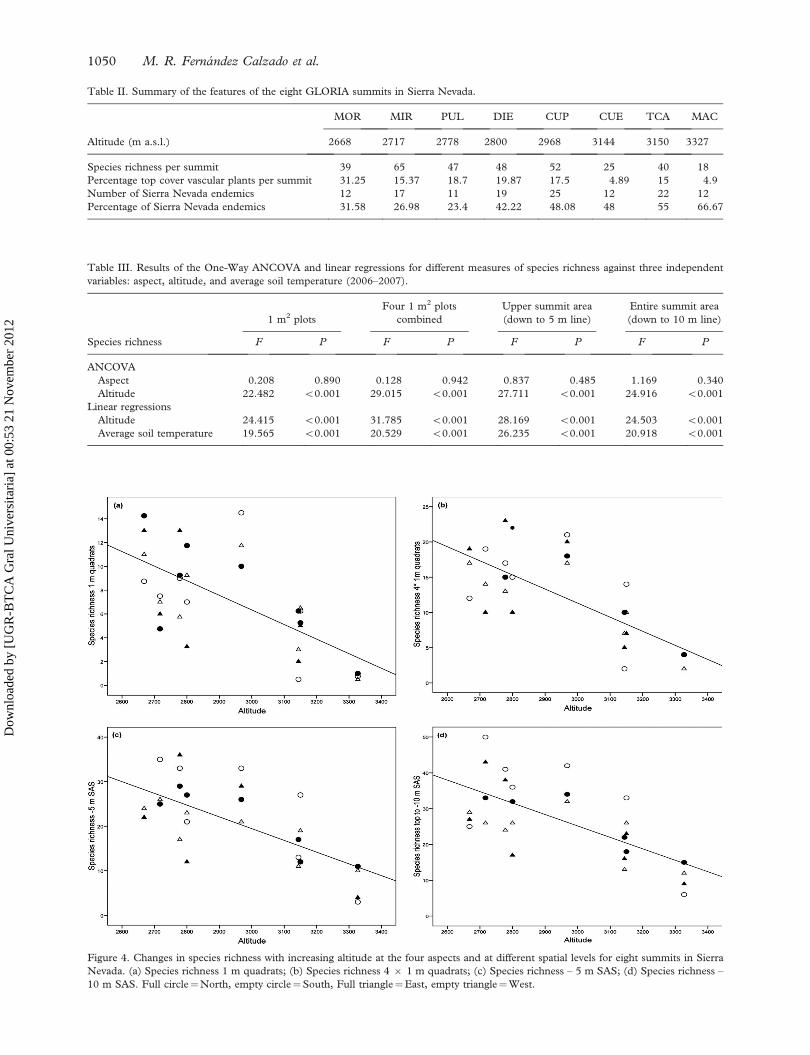
Table II. Summary of the features of the eight GLORIA summits in Sierra Nevada.
MOR MIR PUL DIE CUP CUE TCA MAC
Altitude (m a.s.l.) 2668 2717 2778 2800 2968 3144 3150 3327
Species richness per summit 39 65 47 48 52 25 40 18
Percentage top cover vascular plants per summit 31.25 15.37 18.7 19.87 17.5 4.89 15 4.9
Number of Sierra Nevada endemics 12 17 11 19 25 12 22 12
Percentage of Sierra Nevada endemics 31.58 26.98 23.4 42.22 48.08 48 55 66.67
Table III. Results of the One-Way ANCOVA and linear regressions for different measures of species richness against three independent
variables: aspect, altitude, and average soil temperature (2006–2007).
1 m2 plots
Four 1 m2 plots
combined
Upper summit area
(down to 5 m line)
Entire summit area
(down to 10 m line)
Species richness F P F P F P F P
ANCOVA
Aspect 0.208 0.890 0.128 0.942 0.837 0.485 1.169 0.340
Altitude 22.482 50.001 29.015 50.001 27.711 50.001 24.916 50.001
Linear regressions
Altitude 24.415 50.001 31.785 50.001 28.169 50.001 24.503 50.001
Average soil temperature 19.565 50.001 20.529 50.001 26.235 50.001 20.918 50.001
Figure 4. Changes in species richness with increasing altitude at the four aspects and at different spatial levels for eight summits in Sierra
Nevada. (a) Species richness 1 m quadrats; (b) Species richness 4 6 1 m quadrats; (c) Species richness – 5 m SAS; (d) Species richness –
10 m SAS. Full circle¼North, empty circle¼South, Full triangle¼East, empty triangle¼West.
1050 M. R. Fernandez Calzado et al.
Dow
nloa
ded
by [
UG
R-B
TC
A G
ral U
nive
rsita
ria]
at 0
0:53
21
Nov
embe
r 20
12
a common pattern in high-mountain regions (Korner
2002; Nagy & Grabherr 2009) as this was also
reported form GLORIA regions in the Alps (Pauli
et al. 2003; Erschbamer et al. 2010), Carpathians
(Coldea & pop 2004), Pyrenees (Villar & benito
alonso 2003), Apennines (Stanisci et al. 2005), Crete
(Kazakis et al. 2007), Australian Alps (Pickering
et al. 2008), and from sites in the tropical Andes
(Halloy et al. 2010). The decrease in Sierra Nevada,
however, was not uniform, since the second-lowest
showed the highest number of species. This may
indicate an ecotonal situation, where the vertical
distributions of oromediterranean species overlap
with those of cryoromediterranean species; a similar
increase in species richness in the Alpine-nival
ecotone was reported in the central Alps (Gottfried
et al. 1998).
Impacts of environmental and, in particular, of
climate change on biodiversity are expected to
strongly affect restricted endemic species that dwell
at high altitudes. In Sierra Nevada, such specialised
endemic species are the principal component of the
prevailing vegetation of the uppermost life zone (i.e.
especially the cryoromediterranean zone). The high-
altitude flora of Sierra Nevada appears to be
particularly vulnerable to climate warming, with a
high risk of warming-induced species loss.
In a European context, the proportions of local
endemics taxa of 27–67% (27–48% in ES-SNN,
(23–67% in ES-SNE) found on the Sierra Nevada
summits (Table II, Figure 3) are certainly out-
standing compared to other central to southern
European GLORIA sites, such as the NE-Car-
pathians, Rumanian Alps (7% Coldea & Pop
2004), NE-Alps, Austria (2–10%, Pauli et al.
2003), central Apennines, Italy (18–24%, Stanisci
et al. 2005) and Lefka Ori/Crete, Greece (31–36%,
Kazakis et al. 2007). Although the high degree of
endemism in Sierra Nevada has long been recog-
nized (Quezel 1953; Rivas Goday & Mayor Lopez
1966; Molero Mesa et al. 1996), our study presents
an almost continuous elevational increase of the
endemism ratio, documented through a standardised
sampling design.
The isolated geographic position of Sierra Nevada
with an outstandingly high elevation, protruding over
much lower surroundings, are suggested to be the
primary reasons for this high percentage of local
endemic taxa (compare Blanca et al. 1998; Gimenez
et al. 2004; Penas et al. 2005; Perez-Garcıa et al.
2007). Other factors specific to mountain environ-
ments that cause habitat separation, such as un-
common soil and geological conditions, or ‘stressful’
habitats such as low-temperature conditions, can
also be associated with the presence of endemic
plants (Kruckeberg & Rabinowitz 1985; Medail &
Verlaque 1997).
Biodiversity conservation and climate change
Climate projections using GCM models indicate a
rather uniform temperature increase on the Iberian
Peninsula during the twenty-first century. The
average projection is an increase of 0.48C/decade in
winter and of 0.78C/decade in summer, for the least
favourable scenario (A2 of the IPCC), and of 0.48Cand 0.68C/decade, respectively, for the most favour-
able scenario (B2 of the IPCC) (Fernandez-Gonzalez
et al. 2005). A general warming and associated
earlier snow-melt, combined with an expected
precipitation decrease (Mariotti et al. 2008), will
have a strong impact on the species composition of
high-mountain vegetation (Guisan et al. 1995). The
narrower the altitudinal distribution and the geogra-
phical range, the more severe is the potential
vulnerability to climate warming (Ohlemuller et al.
2008; Dirnbock et al. 2011). In Mediterranean
climates, where precipitation is widely absent during
the growing season, forecasted climatic changes are
expected to generally increase environmental severity
in high-mountain habitats, and are likely to lead to
serious biodiversity losses. In this context, species
present on the summits, such as Erigeron frigidus,
Nevadensia purpurea or Trisetum glaciale, with an area
of distribution situated at the highest altitudes of the
Sierra Nevada, will have serious difficulties to survive
if the trend continues. A similar critical situation can
be anticipated for other S-European high mountains
and for the High Atlas or for the fragmented low-
temperature areas of Iran’s mountains, where the
rate of endemism increases very sharply with altitude
(Noroozi et al. 2011). Continued observation and
more detailed studies in Sierra Nevada and new
Figure 5. Changes in species richness in relation to average soil
temperature at the four aspects for eight summits in Sierra
Nevada. Full circle¼MAC, empty circle¼TCA, Full triangle¼CUE, empty triangle¼CUP, Full square¼DIE, empty square¼PUL, Star¼MIR, Diamond¼MOR.
Diversity and climate change in Sierra Nevada (Spain) 1051
Dow
nloa
ded
by [
UG
R-B
TC
A G
ral U
nive
rsita
ria]
at 0
0:53
21
Nov
embe
r 20
12

monitoring in other mountains in Mediterranean-
type climates are required for a thorough assessment
of the rate and magnitude of expected species
declines.
Acknowledgements
This research was supported by GLORIA-EUROPE
(2001–2003; No. EVK2-CT-2000-00056) and by
the National Parks Network (Red de Parques
Nacionales) of the Spanish Ministry of the Environ-
ment, Project 2/2003.
References
Araujo MB, Pearson RG, Thuiller W, Erhard M. 2005a.
Validation of species–climate impact models under climate
change. Glob Change Biol 11: 1504–1513.
Araujo MB, Whittaker RJ, Ladle RJ, Erhard M. 2005b. Reducing
uncertainty in projections of extinction risk from climate
change. Glob Ecol Biogeogr 14: 529–538.
Blanca G, Cueto M, Martınez-Lirola MJ, Molero Mesa J. 1998.
Threatened vascular flora of Sierra Nevada (Southern Spain).
Biol Conserv 85: 269–285.
Broennimann O, Thuiller W, Hughes G, Midgley GF, Alkemalde
JMR, Guisan A. 2006. Do geographic distribution, niche
property and life form explain plants’ vulnerability to global
change? Glob Change Biol 12: 1079–1093.
Castroviejo S, Laınz M, Lopez Gonzalez G, Montserrat T, Munoz
Garmendia F, Paiva J, et al. editors. 1986–2009. Flora Iberica.
Plantas vasculares de la Penınsula Iberica e Islas Baleares. Vols.
I-VIII, X, XIII-XV, XVIII y XXI. Madrid: Real Jardın
Botanico de Madrid, Consejo Superior de Investigaciones
Cientıficas.
Coldea G, Pop A. 2004. Floristic diversity in relation to
geomorphological and climatic factors in the subalpine-alpine
belt of the Rodna Mountains (the Romanian Carpathians).
Pirineos 158: 61–72.
Dirnbock T, Essl F, Rabitsch W. 2011. Disproportional risk for
habitat loss of high-altitude endemic species under climate
change. Glob Change Biol 17: 990–996.
Dullinger S, Kleinbauer I, Pauli H, Gottfried M, Booker R, Nagy
L, et al. 2007. Weak and variable relationships between
environmental severity and small-scale co-occurrence in alpine
plant communities. J Ecol 95: 1284–1295.
Erschbamer B, Mallaun M, Unterluggauer P, Abdaladze O,
Akhalkatsi M, Nakhutsrishvili G. 2010. Plant diversity along
altitudinal gradients in the Central Alps (South Tyrol, Italy)
and in the Central Greater Caucasus (Kazbegi region,
Georgia). Tuexenia 30: 11–29.
Fernandez-Gonzalez F, Loidi J, Moreno Sainz JC. 2005.
Impactos sobre la biodiversidad vegetal, 5. Evaluacion
preliminar de los impactos en Espana por efecto del cambio
climatico. MMA. Secretarıa General Tecnica. Madrid:
Centro de Publicaciones.
Gimenez E, Melendo M, Valle F, Gomez-Mercado F, Cano E.
2004. Endemic flora biodiversity in the south of the Iberian
Peninsula: Altitudinal distribution, life forms and dispersal
modes. Biodiver Conserv 13: 2641–2660.
Gottfried M, Pauli H, Grabherr G. 1998. Prediction of vegetation
patterns at the limits of plant life: a new view of the alpine-nival
ecotone. Arctic Alp Res 30: 207–221.
Grabherr G, Gottfried M, Pauli H. 1994. Climate effects on
mountain plants. Nature 369: 448–448.
Grabherr G, Gottfried M, Pauli H. 2000. GLORIA: A global
observation research initiative in Alpine environments. Mt Res
Dev 20: 190–191.
Grabherr G, Gottfried M, Pauli H. 2001. Long-term monitoring
of mountain peaks in the Alps. In: Burga CA, Kratochwil A,
editors. Biomonitoring: General and applied aspects on
regional and global scales. Tasks for Vegetation Science. Vol.
35. Dordrecht (the Netherlands): Kluwer Academic Publish-
ers. pp. 153–177.
Guisan A, Tessier L, Holten JI, Haeverli W, Baumgartner M.
1995. Understanding the impact of climate changing on
mountain ecosystems: an overview. In: Guisan A, et al.,
editors. Potential ecological impacts of climate change in the
Alps and Fennoscandian mountains, Ed. Conserv. Jard. Bot.
Geneve.
Halloy S, Yager K, Garcıa C, Beck S, Carilla J, Tupayachi A, et al.
2010. South America: Climate monitoring and Adaptation
Integrated across regions and disciplines. In: Settele J, et al.
editors. Atlas of biodiversity risk. Sofia: Pensoft Publishers. pp.
90–95.
Hansen A, Neilson RP, Dale VH, Flather CH, Iverson LR, Currie
DJ, et al. 2001. Global change in forests: Responses of species,
communities, and biomes. BioScience 51: 765–779.
Houghton JT, Ding Y, Griggs DJ, Nouger M, Van der Linden PJ,
Dai X, et al. 2001. Climate change 2001: The scientific basis.
Intergovernmental panel on climate change, Working group I.
New York: Cambridge University Press.
Kazakis G, Ghosh D, Vogiatzakis IN, Papanastasis VP. 2007.
Vascular plant diversity and climate change in the alpine zone of
the Lefka Ori, Crete. Biodiver Conserv 16: 1603–1615.
Korner C. 1994. Impact of atmospheric changes on high mountain
vegetation. In: Beniston M, editor. Mountain environments in
changing climates. London: Routledge. pp. 155–166.
Korner C. 2002. Mountain biodiversity, its causes and function:
An overview. In: Korner C, Spehn E, editors. Mountain
biodiversity – A global assessment. London, New York:
Parthenon. pp. 3–20.
Kruckeberg AR, Rabinowitz D. 1985. Biological aspects of
endemism in higher plants. Ann Rev Ecol Syst 16: 447–479.
Mariotti A, Zeng N, Yoon J-H, Artale V, Navarra A, Alpert P,
et al. 2008. Mediterranean water cycle changes: Transition to
drier 21st century conditions in observations and CMIP3
simulations. Environ Res Lett 3: 044001 (8pp).
Medail F, Verlaque R. 1997. Ecological characteristics and rarity
of endemic plants from southeast France and Corsica:
Implications for biodiversity conservation. Biologic Conserv
80: 269–281.
Molero Mesa J, Perez Raya F. 1987. La Flora de Sierra Nevada.
Avance sobre el catalogo florıstico nevadense. Ed. Universidad
de Granada, Spain.
Molero Mesa J, Perez Raya F, Gonzalez-Tejero MR. 1996.
Catalogo y analisis florıstico de la flora orofila de Sierra
Nevada. In: Chacon Montero J, Rosua Campos JL, editors.
Sierra Nevada. Vol. 2. Madrid, Spain: Conservacion y
desarrollo sostenible. pp. 271–276.
Molero Mesa J, Fernandez Calzado MR, Merzouki A, Casares
Porcel M, Gonzalez-Tejero Garcıa MR. 2009. Escenarios
fitocenologicos de observacion para el seguimiento del cambio
climatico en Sierra Nevada. In: Ramırez L, Asensio B, editors.
Proyectos de investigacion en parques nacionales: 2005–2008.
Espana: Organismo Autonomo Parques Nacionales. pp. 73–
96.
Nagy L, Grabherr G. 2009. Biology of Alpine habitats. Oxford:
Oxford University Press.
Noroozi J, Pauli H, Grabherr G, Breckle SW. 2011. The subnival–
nival vascular plant species of Iran: A unique high-mountain
flora and its threat from climate warming. Biodiver Conserv
20: 1319–1338.
1052 M. R. Fernandez Calzado et al.
Dow
nloa
ded
by [
UG
R-B
TC
A G
ral U
nive
rsita
ria]
at 0
0:53
21
Nov
embe
r 20
12

Ohlemuller R, Anderson BJ, Araujo MB, Butchart SMH, Kudrna
O, Ridgely RS, Thomas CD. 2008. The coincidence of
climatic and species rarity: High risk to small-range species
from climate change. Biol Lett 4: 568–572.
Ozenda P, Borel JL. 1991. Les consequences ecologiques
possibles des changements climatiques dans l’Arc alpin.
Rapport Futuralp. No 1. Grenoble: ICALPE.
Pauli H, Gottfried M, Grabherr G. 1996. Effects of climate change
on mountain ecosystems – Upward shifting of alpine plants.
World Resource Rev 8: 382–390.
Pauli H, Gottfried M, Dirnbock T, Dullinger S, Grabherr G.
2003. Assessing the long-term dynamics of endemic plants at
summit habitats. In: Nagy L, Grabher G, Korner C,
Thompson DBA, editors. Alpine biodiversity in Europe – A
Europe-wide assessment of biological richness and change.
Ecological Studies. Berlin, Springer: Springer. pp. 195–207.
Pauli H, Gottfried M, Hohenwallner D, Reiter K, Grabherr G.
2004. The GLORIA field manual a multi-summit approach.
Luxembourg: European Communities. Available: www.gloria.
ac.at. Accessed Jul 2012 20.
Pearson RG, Dawson TP. 2003. Predicting the impacts of
climate change on the distribution of species: Are biocli-
mate envelope models useful? Global Ecol Biogeogr 12:
361–371.
Penas J, Perez-Garcıa F, Mota JF. 2005. Patterns of endemic
plants and biogeography of the Baetic high mountains (south
Spain). Acta Bot. Gallica 152: 347–360.
Perez-Garcıa FJ, Cueto M, Penas J, Martınez-Hernandez F,
Medina-Cazorla JM, Garrido-Becerra JA, et al. 2007. Selection
of an endemic flora reserve network and its biogeographical
significance in the Baetic range (Southern Spain). Acta Bot
Gallica 154: 545–571.
Pickering CM, Hill W, Green K. 2008. Vascular plant diversity
and climate change in the alpine zone of the Snowy Mountains,
Australia. Biodiver Conserv 17: 1627–1644.
Quezel P. 1953. Contribution a l’etude phytosociologique et
geobotanique de la Sierra Nevada. Mem Soc Brot 9: 5–77.
Rivas Goday S, Mayor Lopez M. 1966. Aspectos de la vegetacion
y flora orophila del Reino de Granada. Anal Real Acad Farm
31: 345–400.
Rivas-Martınez S. 2007. Mapa de series, geoseries y geopermaseries
de vegetacion de Espana (Memoria del mapa de vegetacion
potencial de Espana. Parte I). Itinera Geobotanica 17: 1–436.
Stanisci A, Pelino G, Blasi C. 2005. Vascular plant diversity and
climate change in the alpine belt of the central Apennines
(Italy). Biodivers Conserv 14: 1301–1318.
Theurillat JP. 1995. Climate change and the alpine flora some
perspectives. In: Guisan A, Holten JI, Spichiger R, Tessier L,
editors. Potential ecological impacts of climate change in the
Alps and Fennoscandian mountains. Geneve: Conserv Jard
Bot. pp. 121–127.
Thuiller W, Broennimann O, Hughes G, Alkemalde JMR,
Midgley GF, Corsi F. 2006. Vulnerability of African mammals
to anthropogenic climate change under conservative land
transformation assumptions. Global Change Biol 12: 424–440.
Villar L, Benito Alonso J-L. 2003. La flora alpina de europa y el
cambio climatico: El caso del pirineo central (Proyecto
GLORIA-Europe). VII Congreso Nacional de la Asociacion
Espanola de Ecologıa Terrestre. Barcelona: Edita AEET. pp.
92–105.
Diversity and climate change in Sierra Nevada (Spain) 1053
Dow
nloa
ded
by [
UG
R-B
TC
A G
ral U
nive
rsita
ria]
at 0
0:53
21
Nov
embe
r 20
12




















