
Valbro: A new site of vertebrates from the early Oligocene (MP22) of France (Quercy). III -...
13
Annales de Paléontologie 101 (2015) 29–41 Disponible en ligne sur ScienceDirect www.sciencedirect.com Original article Valbro: A new site of vertebrates from the early Oligocene (MP 22) of France (Quercy). III - Amphibians and squamates Valbro : un nouveau site à vertébrés de l’Oligocène inférieur (MP 22) de France (Quercy). III - Amphibiens et squamates Jean-Claude Rage ∗ , Marc Augé Sorbonne Universités, CR2P, CNRS, UPMC-Paris6, Muséum national d’Histoire naturelle (MNHN), CP 38, 8, rue Buffon, 75231 Paris Cedex 05, France a r t i c l e i n f o Article history: Received 26 August 2014 Accepted 17 October 2014 Available online 15 November 2014 Keywords: Amphibia Squamata Oligocene France Europe a b s t r a c t The Valbro locality (Phosphorites du Quercy, France) has yielded a relatively diverse fauna of amphibians and squamate reptiles from the early Oligocene. Amphibians include two caudatans (the salamandrids cf. Megalotriton sp. and Salamandra sp.) and two anurans (an alytid of the ‘Discoglossus group’ and an inde- terminate pelobatid). Squamates are more diverse, they comprise an agamid (Uromastyx europaeus), an indeterminate lacertid, an anguid (Dopasia sp.), a platynotan (Necrosaurus sp.), two amphisbaenians (an amphisbaenid and a blanid), an anilioid-grade snake (Eoanilius aff. E. oligocenicus), three boids and a colu- brid (Coluber cadurci). The fauna includes taxa known also in the Eocene, i.e. which were already present in the Phosphorites before the Grande Coupure, and immigrants that entered western Europe after this event. The stratigraphic overlap of the holdovers Necrosaurus, ‘boid C’ and cf. Megalotriton sp., and of the newcomers U. europaeus, E. aff. E. oligocenicus and C. cadurci suggests a MP 22 (early Oligocene) age. This precise age was already inferred from mammals of the locality. Taxa indicative of arid environment and aquatic (in fresh water) forms are associated in this fauna, as is possible in karstic areas. © 2014 Elsevier Masson SAS. All rights reserved. Mots clés : Amphibia Squamata Oligocène France Europe r é s u m é Le gisement de Valbro (Phosphorites du Quercy, France) a fourni une faune d’amphibiens et squamates de l’Oligocène inférieur relativement diversifiée. Les amphibiens comprennent deux urodèles (les Sala- mandridae cf. Megalotriton et Salamandra sp.) et deux anoures (un Alytidae du « groupe Discoglossus » et un Pelobatidae indéterminé). Les squamates sont plus diversifiés ; ils incluent un Agamidae (Uro- mastyx europaeus), un Lacertidae indéterminé, un Anguidae (Dopasia sp.), un platynote (Necrosaurus sp.), deux amphisbaeniens (un Amphisbaenidae et un Blanidae), un serpent de grade anilioïde (Eoanil- ius aff. E. oligocenicus), trois Boidae et un Colubridae (Coluber cadurci). Cette faune comprend des taxons présents dans l’Eocène, donc connus dans les Phosphorites avant la Grande Coupure, et des immigrants qui ont atteint l’Europe de l’ouest après cet événement. Stratigraphiquement, le chevauchement de Necrosaurus, du « Boidae C » et de cf. Megalotriton, d’une part, et des nouveaux arrivants U. europaeus, E. aff. E. oligocenicus et C. cadurci, d’autre part, correspond au niveau-repère MP 22 de l’Oligocène inférieur. Cet âge précis avait déjà été proposé sur la base des mammifères. Dans cette faune sont associés des tax- ons de milieux arides et des taxons aquatiques d’eau douce ; une telle association est possible en milieu karstique. © 2014 Elsevier Masson SAS. Tous droits réservés. ∗ Corresponding author. E-mail address: [email protected] (J.-C. Rage). 1. Introduction The Phosphorites du Quercy (southwestern France) comprise numerous fissures and cavities in-filled by phosphatic sediments. Many of them were entirely or partly emptied for exploitation of http://dx.doi.org/10.1016/j.annpal.2014.10.002 0753-3969/© 2014 Elsevier Masson SAS. All rights reserved.
Transcript of Valbro: A new site of vertebrates from the early Oligocene (MP22) of France (Quercy). III -...

O
VF
V(
JS
ARAA
KASOFE
MASOFE
0
Annales de Paléontologie 101 (2015) 29–41
Disponible en ligne sur
ScienceDirectwww.sciencedirect.com
riginal article
albro: A new site of vertebrates from the early Oligocene (MP 22) ofrance (Quercy). III - Amphibians and squamates
albro : un nouveau site à vertébrés de l’Oligocène inférieur (MP 22) de FranceQuercy). III - Amphibiens et squamates
ean-Claude Rage ∗, Marc Augéorbonne Universités, CR2P, CNRS, UPMC-Paris6, Muséum national d’Histoire naturelle (MNHN), CP 38, 8, rue Buffon, 75231 Paris Cedex 05, France
a r t i c l e i n f o
rticle history:eceived 26 August 2014ccepted 17 October 2014vailable online 15 November 2014
eywords:mphibiaquamataligoceneranceurope
a b s t r a c t
The Valbro locality (Phosphorites du Quercy, France) has yielded a relatively diverse fauna of amphibiansand squamate reptiles from the early Oligocene. Amphibians include two caudatans (the salamandrids cf.Megalotriton sp. and Salamandra sp.) and two anurans (an alytid of the ‘Discoglossus group’ and an inde-terminate pelobatid). Squamates are more diverse, they comprise an agamid (Uromastyx europaeus), anindeterminate lacertid, an anguid (Dopasia sp.), a platynotan (Necrosaurus sp.), two amphisbaenians (anamphisbaenid and a blanid), an anilioid-grade snake (Eoanilius aff. E. oligocenicus), three boids and a colu-brid (Coluber cadurci). The fauna includes taxa known also in the Eocene, i.e. which were already presentin the Phosphorites before the Grande Coupure, and immigrants that entered western Europe after thisevent. The stratigraphic overlap of the holdovers Necrosaurus, ‘boid C’ and cf. Megalotriton sp., and of thenewcomers U. europaeus, E. aff. E. oligocenicus and C. cadurci suggests a MP 22 (early Oligocene) age. Thisprecise age was already inferred from mammals of the locality. Taxa indicative of arid environment andaquatic (in fresh water) forms are associated in this fauna, as is possible in karstic areas.
© 2014 Elsevier Masson SAS. All rights reserved.
ots clés :mphibiaquamataligocèneranceurope
r é s u m é
Le gisement de Valbro (Phosphorites du Quercy, France) a fourni une faune d’amphibiens et squamatesde l’Oligocène inférieur relativement diversifiée. Les amphibiens comprennent deux urodèles (les Sala-mandridae cf. Megalotriton et Salamandra sp.) et deux anoures (un Alytidae du « groupe Discoglossus »et un Pelobatidae indéterminé). Les squamates sont plus diversifiés ; ils incluent un Agamidae (Uro-mastyx europaeus), un Lacertidae indéterminé, un Anguidae (Dopasia sp.), un platynote (Necrosaurussp.), deux amphisbaeniens (un Amphisbaenidae et un Blanidae), un serpent de grade anilioïde (Eoanil-ius aff. E. oligocenicus), trois Boidae et un Colubridae (Coluber cadurci). Cette faune comprend des taxonsprésents dans l’Eocène, donc connus dans les Phosphorites avant la Grande Coupure, et des immigrantsqui ont atteint l’Europe de l’ouest après cet événement. Stratigraphiquement, le chevauchement de
Necrosaurus, du « Boidae C » et de cf. Megalotriton, d’une part, et des nouveaux arrivants U. europaeus, E.aff. E. oligocenicus et C. cadurci, d’autre part, correspond au niveau-repère MP 22 de l’Oligocène inférieur.Cet âge précis avait déjà été proposé sur la base des mammifères. Dans cette faune sont associés des tax-ons de milieux arides et des taxons aquatiques d’eau douce ; une telle association est possible en milieukarstique.∗ Corresponding author.E-mail address: [email protected] (J.-C. Rage).
http://dx.doi.org/10.1016/j.annpal.2014.10.002753-3969/© 2014 Elsevier Masson SAS. All rights reserved.
© 2014 Elsevier Masson SAS. Tous droits réservés.
1. Introduction
The Phosphorites du Quercy (southwestern France) comprisenumerous fissures and cavities in-filled by phosphatic sediments.Many of them were entirely or partly emptied for exploitation of

3 de Palé
pfbrta1C
epfifDFw
opaC
Vtel
2
t4rbznticlaTdcataTttl
Evftt
0 J.-C. Rage, M. Augé / Annales
hosphate rock during the 19th and early 20th centuries. Sedimentsrom these fissures have generally produced rich faunas of verte-rates. From a stratigraphic point of view, the ages of the faunasange from the early Eocene (standard level MP 8 + 9; Ypresian)o the early Miocene (MN 3; early Burdigalian). However, almostll fossiliferous localities range from the late middle Eocene (MP6; late Bartonian) to the late, but not latest, Oligocene (MP 28;hattian) (Legendre et al., 1997; Rage, 2006; Sigé et al., 1991).
The recently recovered locality of Valbro is one of the partlymptied cavities. Cuttings resulting from the exploitation of phos-hate rock remain close to the sinkhole; they contain fossils. Onlyllings of four gallery openings remain in situ in the sinkhole. These
our fillings are fossiliferous and they are named Valbro A, B, C and, whereas cuttings are termed Valbro déblais (Peigné et al., 2014).ossils were collected directly from the sediment and after screenashing.
Peigné et al. (2014) studied various mammals (rodents, hyaen-dontids, carnivores) from the locality and they provided areliminary list of the vertebrates. They assigned the Valbro fauna
MP 22 (early Oligocene) age. Birds were examined by Mourer-hauviré (2014).
The present article deals with amphibians and squamates fromalbro. It should be noted that three amphibian and one-squamate
axa are added to the provisional faunal list published in Peignét al. (2014). The material is curated in the Institut des Sciences de’Évolution (ISEM), Université de Montpellier 2, France.
. Systematic Palaeontology
Class LISSAMPHIBIA Haeckel, 1866Order CAUDATA Scopoli, 1777Family: SALAMANDRIDAE Goldfuss, 1820
Genus Megalotriton von Zittel, 1890
cf. Megalotriton sp.Fig. 1A–C.Material. Valbro D: one anterior-trunk vertebra (VBO 514).Description. A comparatively large vertebra is referred to this
axon (width across prezygapophyses: 6.5 mm; length of centrum:.7 mm). It comes from the anteriormost portion of the dorsalegion as shown by its shortness and anterior location of its rib-earers. The shape of the specimen was altered pathologically; theygapophyses are not at the same level and the posterior part of theeural arch markedly rises on the right side but apparently not onhe other side. However, the overall morphology of the vertebras not modified by these deformations. The centrum is opistho-oelous, the condyle being well ossified. On the anterior face, aarge foramen opens on either side of the condyle. Ventral crestsre lacking; consequently, the centrum is well limited laterally.he rib-bearers are robust, with a large diameter and markedlyivergent; the ventral one is oriented latero-ventrally. They are notlearly oriented posterolaterally. On each side, the two rib-bearersre connected by a well-developed bony lamina that extends up toheir tips. As a result of the anterior position of the rib-bearers, thenterior zygapophyseal ridge is strongly reduced, perhaps absent.he posterior zygapophyseal ridge is present but it does not con-act the dorsal rib-bearer, thus leaving a broad notch posterior tohe latter. On each side, three vertically aligned foramina pierce theateral wall, just behind the rib-bearer.
Discussion. The largest salamander from the Palaeogene ofurope is a rare salamandrid described as Megalotriton filholi by
on Zittel (1890). The original description was based on materialrom the Phosphorites du Quercy. According to von Zittel (1890)he precise provenance of the first described specimen is Escamps;his is reliable because, during recent excavations, the localityontologie 101 (2015) 29–41
produced several specimens belonging to M. filholi. Escamps is oflate Eocene (MP 19) age (Legendre, 1987). This species has neverbeen revised, only short lists of characters were provided by Estes(1981) and Duffaud (2000). M. filholi was identified in the newcollections of the Phosphorites in which it was recovered in the lateEocene, from MP 18 and MP 19 (Rage, 2006). However, Duffaud(2000) reported a large salamandrid from the Oligocene (MP21–MP 23) of the Phosphorites and he referred it to as Megalotritonaff. filholi, an identification taken up by Rage (2006).
The vertebra from Valbro (VBO 514) displays several featuresthat are consistent with M. filholi from the Eocene: vertebra largeand massive; rib-bearers strong, with a large diameter, almostperpendicular to vertebral axis; zygapophyseal facets wider thananteroposteriorly long. However, VBO 514 clearly differs fromanterior-trunk vertebrae of M. filholi from the Eocene in hav-ing marked notches posterior to the dorsal rib-bearers, longerrib-bearers, more horizontal dorsal rib-bearers, more ventrallydirected ventral rib-bearers, and in lacking ventral crests. In addi-tion, VBO 514, as all related specimens from the Oligocene, is clearlysmaller than those from the Eocene; in anterior dorsal vertebrae ofM. filholi from the Eocene, the width across prezygapophyses can beover 13 mm and the centrum length over 10 mm. These differencesshow that the Oligocene form is not M. filholi and even the referralto the genus Megalotriton might be questioned.
In addition, it is worth noting that two of the three foraminalocated behind the rib-bearer of VBO 514 may correspond to theexits of roots of the spinal nerve. This condition was not observedin Salamandridae; in this family, there is a single foramen for thenerve (Edwards, 1976). This may call into question the referralof VBO 514 to the Salamandridae. However, this feature remainsto be examined in all members of the family. The number andposition of foramina appear to be somewhat variable in M. filholifrom the Eocene.
Genus Salamandra Laurenti, 1768
Salamandra sp.Fig. 1D–F.Material. Valbro déblais: one trunk vertebra (VBO 519).Description. An incomplete mid-trunk vertebra represents a
second salamander. It is smaller than that referred to as cf.Megalotriton (width across prezygapophyses: 5.7 mm; length ofcentrum: 4.2 mm). The vertebra resembles those of Salamandra.It is more elongate and less robust than that of cf. Megalotriton,it being understood that the elongation largely reflects the posi-tion in the vertebral column. Its neural arch is wide and stronglydepressed. Posteriorly, the neural arch forms two projections thatborder a median notch. The neural spine is long and low. The artic-ular facets of the prezygapophyses are large and oblique in dorsalaspect. The rib-bearers, located at mid-length, are broken off buttheir bases show that they were not strong. A large spinal foramenis located on each side, posterior to the rib-bearer; it faces postero-laterally. The zygapophyseal ridges are well developed; the anteriorone is horizontal and it contacts the dorsal rib-bearer. The centrumis opisthocoelous and the anterior condyle is well ossified. A poorlymarked neck separates the condyle from the centrum. Posteriorto the condyle, a transverse ridge whose anterior limit is sharplymarked extends to both prezygapophyses. In lateral view, the ven-tral surface of the centrum is arched dorsally. This single specimensuggests assignment to Salamandra.
Discussion. Vertebrae displaying the morphology of Salaman-
dra, or a very close morphology, are known since the Palaeocene(Estes, 1981). Salamandra was reported, as S. sansaniensis Lartet,1851, from the latest middle Eocene (MP 16) of the Phosphorites duQuercy (Rage, 1988a). However, Duffaud (2000) referred the latter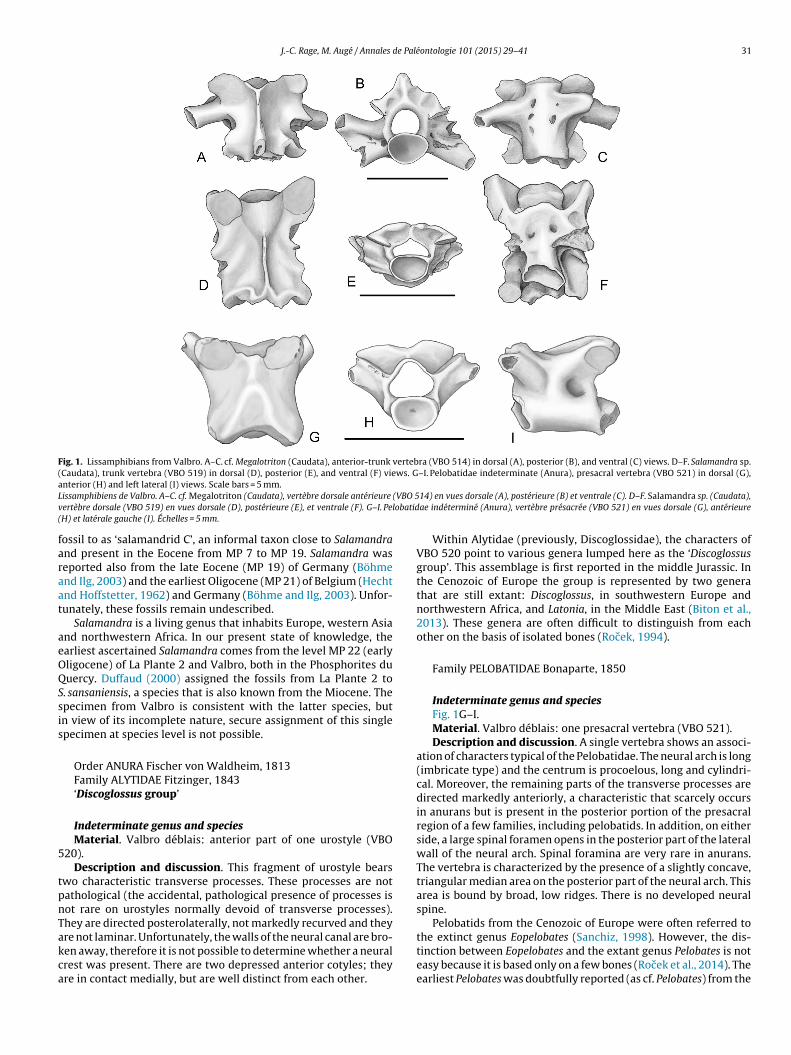
J.-C. Rage, M. Augé / Annales de Paléontologie 101 (2015) 29–41 31
Fig. 1. Lissamphibians from Valbro. A–C. cf. Megalotriton (Caudata), anterior-trunk vertebra (VBO 514) in dorsal (A), posterior (B), and ventral (C) views. D–F. Salamandra sp.(Caudata), trunk vertebra (VBO 519) in dorsal (D), posterior (E), and ventral (F) views. G–I. Pelobatidae indeterminate (Anura), presacral vertebra (VBO 521) in dorsal (G),aL (VBO 5v lobatid(
faraat
aeOQSsis
5
tpnTakca
nterior (H) and left lateral (I) views. Scale bars = 5 mm.issamphibiens de Valbro. A–C. cf. Megalotriton (Caudata), vertèbre dorsale antérieure
ertèbre dorsale (VBO 519) en vues dorsale (D), postérieure (E), et ventrale (F). G–I. PeH) et latérale gauche (I). Échelles = 5 mm.
ossil to as ‘salamandrid C’, an informal taxon close to Salamandrand present in the Eocene from MP 7 to MP 19. Salamandra waseported also from the late Eocene (MP 19) of Germany (Böhmend Ilg, 2003) and the earliest Oligocene (MP 21) of Belgium (Hechtnd Hoffstetter, 1962) and Germany (Böhme and Ilg, 2003). Unfor-unately, these fossils remain undescribed.
Salamandra is a living genus that inhabits Europe, western Asiand northwestern Africa. In our present state of knowledge, thearliest ascertained Salamandra comes from the level MP 22 (earlyligocene) of La Plante 2 and Valbro, both in the Phosphorites duuercy. Duffaud (2000) assigned the fossils from La Plante 2 to. sansaniensis, a species that is also known from the Miocene. Thepecimen from Valbro is consistent with the latter species, butn view of its incomplete nature, secure assignment of this singlepecimen at species level is not possible.
Order ANURA Fischer von Waldheim, 1813Family ALYTIDAE Fitzinger, 1843‘Discoglossus group’
Indeterminate genus and speciesMaterial. Valbro déblais: anterior part of one urostyle (VBO
20).Description and discussion. This fragment of urostyle bears
wo characteristic transverse processes. These processes are notathological (the accidental, pathological presence of processes isot rare on urostyles normally devoid of transverse processes).hey are directed posterolaterally, not markedly recurved and they
re not laminar. Unfortunately, the walls of the neural canal are bro-en away, therefore it is not possible to determine whether a neuralrest was present. There are two depressed anterior cotyles; theyre in contact medially, but are well distinct from each other.14) en vues dorsale (A), postérieure (B) et ventrale (C). D–F. Salamandra sp. (Caudata),ae indéterminé (Anura), vertèbre présacrée (VBO 521) en vues dorsale (G), antérieure
Within Alytidae (previously, Discoglossidae), the characters ofVBO 520 point to various genera lumped here as the ‘Discoglossusgroup’. This assemblage is first reported in the middle Jurassic. Inthe Cenozoic of Europe the group is represented by two generathat are still extant: Discoglossus, in southwestern Europe andnorthwestern Africa, and Latonia, in the Middle East (Biton et al.,2013). These genera are often difficult to distinguish from eachother on the basis of isolated bones (Rocek, 1994).
Family PELOBATIDAE Bonaparte, 1850
Indeterminate genus and speciesFig. 1G–I.Material. Valbro déblais: one presacral vertebra (VBO 521).Description and discussion. A single vertebra shows an associ-
ation of characters typical of the Pelobatidae. The neural arch is long(imbricate type) and the centrum is procoelous, long and cylindri-cal. Moreover, the remaining parts of the transverse processes aredirected markedly anteriorly, a characteristic that scarcely occursin anurans but is present in the posterior portion of the presacralregion of a few families, including pelobatids. In addition, on eitherside, a large spinal foramen opens in the posterior part of the lateralwall of the neural arch. Spinal foramina are very rare in anurans.The vertebra is characterized by the presence of a slightly concave,triangular median area on the posterior part of the neural arch. Thisarea is bound by broad, low ridges. There is no developed neuralspine.
Pelobatids from the Cenozoic of Europe were often referred to
the extinct genus Eopelobates (Sanchiz, 1998). However, the dis-tinction between Eopelobates and the extant genus Pelobates is noteasy because it is based only on a few bones (Rocek et al., 2014). Theearliest Pelobates was doubtfully reported (as cf. Pelobates) from the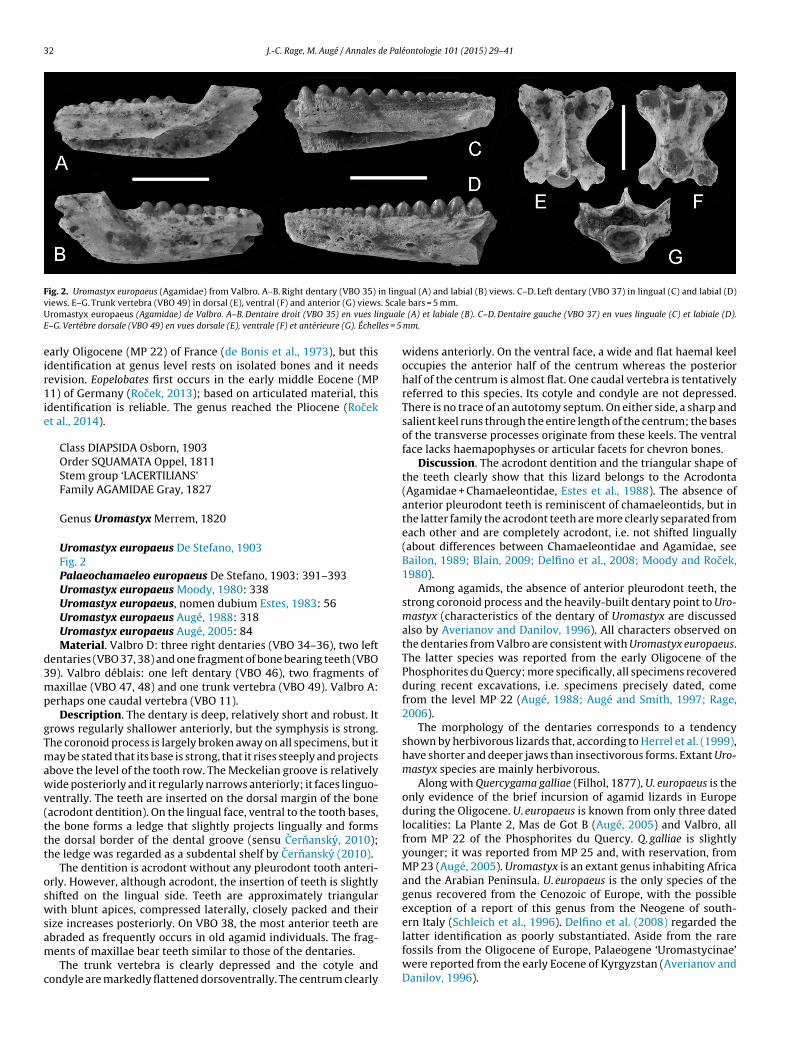
32 J.-C. Rage, M. Augé / Annales de Paléontologie 101 (2015) 29–41
Fig. 2. Uromastyx europaeus (Agamidae) from Valbro. A–B. Right dentary (VBO 35) in lingual (A) and labial (B) views. C–D. Left dentary (VBO 37) in lingual (C) and labial (D)v . ScaleU ngualeE les = 5
eir1ie
d3mp
gTmawv(ttt
oswsam
c
iews. E–G. Trunk vertebra (VBO 49) in dorsal (E), ventral (F) and anterior (G) viewsromastyx europaeus (Agamidae) de Valbro. A–B. Dentaire droit (VBO 35) en vues li–G. Vertèbre dorsale (VBO 49) en vues dorsale (E), ventrale (F) et antérieure (G). Échel
arly Oligocene (MP 22) of France (de Bonis et al., 1973), but thisdentification at genus level rests on isolated bones and it needsevision. Eopelobates first occurs in the early middle Eocene (MP1) of Germany (Rocek, 2013); based on articulated material, this
dentification is reliable. The genus reached the Pliocene (Rocekt al., 2014).
Class DIAPSIDA Osborn, 1903Order SQUAMATA Oppel, 1811Stem group ‘LACERTILIANS’Family AGAMIDAE Gray, 1827
Genus Uromastyx Merrem, 1820
Uromastyx europaeus De Stefano, 1903Fig. 2Palaeochamaeleo europaeus De Stefano, 1903: 391–393Uromastyx europaeus Moody, 1980: 338Uromastyx europaeus, nomen dubium Estes, 1983: 56Uromastyx europaeus Augé, 1988: 318Uromastyx europaeus Augé, 2005: 84Material. Valbro D: three right dentaries (VBO 34–36), two left
entaries (VBO 37, 38) and one fragment of bone bearing teeth (VBO9). Valbro déblais: one left dentary (VBO 46), two fragments ofaxillae (VBO 47, 48) and one trunk vertebra (VBO 49). Valbro A:
erhaps one caudal vertebra (VBO 11).Description. The dentary is deep, relatively short and robust. It
rows regularly shallower anteriorly, but the symphysis is strong.he coronoid process is largely broken away on all specimens, but itay be stated that its base is strong, that it rises steeply and projects
bove the level of the tooth row. The Meckelian groove is relativelyide posteriorly and it regularly narrows anteriorly; it faces linguo-
entrally. The teeth are inserted on the dorsal margin of the boneacrodont dentition). On the lingual face, ventral to the tooth bases,he bone forms a ledge that slightly projects lingually and formshe dorsal border of the dental groove (sensu Cernansky, 2010);he ledge was regarded as a subdental shelf by Cernansky (2010).
The dentition is acrodont without any pleurodont tooth anteri-rly. However, although acrodont, the insertion of teeth is slightlyhifted on the lingual side. Teeth are approximately triangularith blunt apices, compressed laterally, closely packed and their
ize increases posteriorly. On VBO 38, the most anterior teeth are
braded as frequently occurs in old agamid individuals. The frag-ents of maxillae bear teeth similar to those of the dentaries.The trunk vertebra is clearly depressed and the cotyle andondyle are markedly flattened dorsoventrally. The centrum clearly
bars = 5 mm. (A) et labiale (B). C–D. Dentaire gauche (VBO 37) en vues linguale (C) et labiale (D).mm.
widens anteriorly. On the ventral face, a wide and flat haemal keeloccupies the anterior half of the centrum whereas the posteriorhalf of the centrum is almost flat. One caudal vertebra is tentativelyreferred to this species. Its cotyle and condyle are not depressed.There is no trace of an autotomy septum. On either side, a sharp andsalient keel runs through the entire length of the centrum; the basesof the transverse processes originate from these keels. The ventralface lacks haemapophyses or articular facets for chevron bones.
Discussion. The acrodont dentition and the triangular shape ofthe teeth clearly show that this lizard belongs to the Acrodonta(Agamidae + Chamaeleontidae, Estes et al., 1988). The absence ofanterior pleurodont teeth is reminiscent of chamaeleontids, but inthe latter family the acrodont teeth are more clearly separated fromeach other and are completely acrodont, i.e. not shifted lingually(about differences between Chamaeleontidae and Agamidae, seeBailon, 1989; Blain, 2009; Delfino et al., 2008; Moody and Rocek,1980).
Among agamids, the absence of anterior pleurodont teeth, thestrong coronoid process and the heavily-built dentary point to Uro-mastyx (characteristics of the dentary of Uromastyx are discussedalso by Averianov and Danilov, 1996). All characters observed onthe dentaries from Valbro are consistent with Uromastyx europaeus.The latter species was reported from the early Oligocene of thePhosphorites du Quercy; more specifically, all specimens recoveredduring recent excavations, i.e. specimens precisely dated, comefrom the level MP 22 (Augé, 1988; Augé and Smith, 1997; Rage,2006).
The morphology of the dentaries corresponds to a tendencyshown by herbivorous lizards that, according to Herrel et al. (1999),have shorter and deeper jaws than insectivorous forms. Extant Uro-mastyx species are mainly herbivorous.
Along with Quercygama galliae (Filhol, 1877), U. europaeus is theonly evidence of the brief incursion of agamid lizards in Europeduring the Oligocene. U. europaeus is known from only three datedlocalities: La Plante 2, Mas de Got B (Augé, 2005) and Valbro, allfrom MP 22 of the Phosphorites du Quercy. Q. galliae is slightlyyounger; it was reported from MP 25 and, with reservation, fromMP 23 (Augé, 2005). Uromastyx is an extant genus inhabiting Africaand the Arabian Peninsula. U. europaeus is the only species of thegenus recovered from the Cenozoic of Europe, with the possibleexception of a report of this genus from the Neogene of south-ern Italy (Schleich et al., 1996). Delfino et al. (2008) regarded the
latter identification as poorly substantiated. Aside from the rarefossils from the Oligocene of Europe, Palaeogene ‘Uromastycinae’were reported from the early Eocene of Kyrgyzstan (Averianov andDanilov, 1996).
de Palé
fi4v
titpaesc
ooaaassrits
bftdsligacv
lsntarw(otl(fftt
J.-C. Rage, M. Augé / Annales
Family LACERTIDAE Batsch, 1788
Indeterminate genus and species
Material. Valbro A: one incomplete left maxilla (VBO 12), oneragment of right dentary (VBO 13) and one fragment of bone bear-ng teeth (VBO 14). Valbro D: two anterior caudal vertebrae (VBO0). Valbro déblais: one trunk vertebra (VBO 50) and one caudalertebra (VBO 51).
Description. The maxilla is the most significant bone. It bears 15ooth loci, eight teeth being preserved. A large maxillary foramens located at the middle-length of the dorsal face of the supraden-al table. An elongate, shallow depression occurs in the posteriorart of this dorsal face. The insertion of teeth is pleurodont. Teethre vertical and their bases are not expanded. The tooth apices areither simple or bicuspid. The accessory cuspid is weak, scarcelyeparated from the main cuspid, and located anteriorly. The mainuspid does not clearly projects and it is not pointed.
The trunk vertebra is comparatively high and short. Anteri-rly, the neural arch forms a small projection that is reminiscentf a pseudozygosphene (Hoffstetter and Gasc, 1969). The paradi-pophyses are rounded and located beneath the prezygapophyses,t mid-height of the vertebra. The ventral face of the centrum bears
broad haemal keel whose ventral surface is slightly hollowed by aagittal depression in its anterior half. The caudal vertebrae do nothow autotomy septa, they likely come from the anterior caudalegion. The bases of the broken off transverse processes are locatedn the middle of the vertebrae; they originated from a long horizon-al keel that runs through the entire lateral face of the vertebrae. Atrong median keel occurs on the ventral face of the centrum.
Discussion. The maxilla, although incomplete, displays a com-ination of features that is known only in Lacertidae: maxillaryoramen large and located at the middle-length of the supradentalable; presence of a shallow depression on the posterior part of theorsal face of the supradental table; teeth bicuspid with the acces-ory cuspid located anteriorly. The assignment of the vertebrae isess secure. As far as the trunk vertebra is concerned, the referrals based on the presence of a projection looking like a pseudozy-osphene and on the presence of rounded paradiapophyses locatedt half the height of the vertebra. The location of the transverse pro-esses on caudal vertebrae is consistent also with anterior caudalertebrae of lacertids.
The Oligocene of the Phosphorites du Quercy produced severalacertid species (Augé, 2005). The teeth of only one of thesepecies, Lacerta s.l. filholi Augé, 1988, are not amblyodont (i.e.,ot enlarged); however, they are clearly different from those ofhe lacertid from Valbro, most of them being tricuspid with ancute main cusp. Oligocene lacertids from outside the Phospho-ites du Quercy remain poorly studied in Europe. Lacerta filholias recovered from the earliest Oligocene (MP 21) of Belgium
Augé and Smith, 2009) but in the Phosphorites du Quercy it firstccurs only in MP 22; outside of the Quercy, the species reacheshe early Miocene (Augé, 2005). In Germany, non-amblyodontacertids include Plesiolacerta eratosthenesi, described from MP 28Cernansky and Augé, 2013), and several indeterminate speciesrom MP 30 reported as Lacerta sp. (Böhme, 2008). The specimensrom Valbro differ from all named and/or reported lacertids fromhe European Oligocene. However, in view of the limited material,hey are only referred to as Lacertidae indeterminate.
Family ANGUIDAE Gray, 1825Subfamily ANGUINAE Gray, 1825
Genus Dopasia Gray, 1853 (sensu Augé, 2005)
Dopasia s.l. sp.
ontologie 101 (2015) 29–41 33
Material. Valbro A: one incomplete left dentary (VBO 15), tentrunk vertebrae (VBO 16–19) and one caudal vertebra (VBO 20).Valbro D: one trunk vertebra (VBO 41). Valbro déblais: four trunkvertebrae (VBO 52) and six caudal vertebrae (VBO 53).
Description. The dentary bears five teeth plus four empty toothloci. Teeth are pleurodont and are inserted on a single surface thatis oblique in cross section. Lingually, this surface is limited by asmall, rounded ledge, but there is no projecting subdental shelf(sensu Rage and Augé, 2010). The bone lacks a sulcus dentalis. Anotch in the ledge represents the dorsal limit of the anterior inferioralveolar foramen. The notch is located below the 5th tooth positionfrom posterior. The anteroventral border of the notch forms thesplenial spine (Klembara et al., 2010). The spine is directed posteri-orly and very pointed. A large intramandibular septum divides theMeckelian groove posterior to the splenial spine. The ventral edgeof the septum remains free. Posteriorly, the septum is notched andit ends posteroventrally as a rather long spine. The ventral border ofthe bone is damaged but, apparently, the Meckelian groove largelyopened ventrally. The labial face is relatively convex transversely.The supraangular notch (sinus surangularis; Klembara et al., 2010) iselongate. The anterior part of the coronoid notch is clearly exposedon the posterior area; it extends markedly less anteriorly than thesupraangular notch. Teeth are recurved and have pointed apices.Their bases are moderately expanded and two faint carinae occuron the anterior and posterior margins. The size of teeth decreasesregularly posteriorly.
Trunk vertebrae are depressed and relatively short (centrumlength of largest vertebra: 4.9 mm). The ventral face of the centrumis flat to slightly convex, without any keel. The centrum widensanteriorly and its lateral margins are concave. Caudal vertebraeare elongate. An autotomy septum crosses the anterior part of thetransverse processes. The latter processes are located anteriorlyand are directed anterolaterally. The thin, lamellar bases of thehaemapophyses are fused to the ventral face of the centrum; theyare directed ventrally.
Discussion. The referral to the Anguidae is clearly demonstratedby the following combination of characters: teeth borne by a sin-gle, inclined surface; marked reduction or absence of the subdentalshelf; ventral opening of the Meckelian groove; lingual ledge ofdentary notched by the anterior inferior alveolar foramen. In addi-tion, the large size of the intramandibular septum and the freecondition of its ventral edge are characteristic of the Anguimor-pha. Within Anguidae, the presence of a splenial spine argues forassignment to the Anguinae. Apparently such a spine occurs only inthe latter group. Delfino et al. (2011) discussed vertebral features ofDopasia. There is no clear character on trunk vertebrae, but caudalvertebrae appear to be more significant.
Several species of Anguinae were reported from the Palaeogeneof Europe. Most have pointed, recurved teeth (i.e., ‘caniniform’teeth). The material from Valbro being limited, comparisons arerestricted to anguines from the Oligocene of Europe. In the earliestOligocene (MP 21) occurs Helvetisaurus (Augé and Smith, 2009),one of the genera that survived the Grande Coupure, that is themarked extinction event that took place at the Eocene-Oligocenetransition. Helvetisaurus is peculiar in lacking a trace of the anteriorinferior alveolar foramen on the dentary; it is thus distinct fromthe Valbro anguine. The teeth of the dentary from Valbro arerelatively similar to those of Dopasia s.l. roqueprunensis Augé, 1992(MP 21–MP 28) and Dopasia s.l. frayssensis Augé, 2005 (MP 28),both from the Phosphorites du Quercy (Augé, 1992, 2005); thesetwo species appear to be morphologically close to each otherand synonymization may be considered. Another Dopasia species
from the Phosphorites, D. coderetensis Augé, 2005 (MP 23–MP30), has less expanded tooth bases than these two species. Thespecimen from Valbro shows a tooth morphology that appearsto be more consistent with D. coderetensis than with the other
34 J.-C. Rage, M. Augé / Annales de Palé
Fig. 3. Necrosaurus sp. from Valbro: caudal vertebra (VBO 54) in dorsal (A), ventral(B), right lateral (C), anterior (D) and posterior (E) views. Scale bar = 5 mm.Necrosaurus sp. de Valbro : vertèbre caudale (VBO 54) en vues dorsale (A), ventrale (B),l
sa
t1fceeA(cut
talc(lpvao
as
versely wide mound, whose dorsal surface is flat, and a more or less
atérale droite (C), antérieure (D) et postérieure (E). Échelles = 5 mm.
pecies; however, the material in not sufficient for identificationt species level and it is only referred to as Dopasia s.l.
It should be noted that referral to the genus Dopasia rests onhe assumption that the living Anguis and Dopasia harti Boulenger,889 form a monophyletic assemblage characterized by ‘canini-orm’ teeth within Anguinae (Augé, 2005), which was partlyorroborated by Conrad (2008). However, molecular analysesither did not confirm this monophyly (Macey et al., 1999; Wienst al., 2006) or recognized a monophyletic Dopasia exclusive ofnguis (Pyron et al., 2013). Based on morphology, Klembara et al.2014) did not evidence a monophyletic Dopasia. Since there is noonsensus and no well-established phylogeny of the Anguinae, wese provisionally the name Dopasia s.l. to assign the anguine fromhe Phosphorites du Quercy.
Clade PLATYNOTA Duméril and Bibron, 1839FAMILY INDETERMINATEGenus Necrosaurus Filhol, 1876
Necrosaurus sp.Fig. 3Material. Valbro déblais: two caudal vertebrae (VBO 54, 55).Description. The vertebrae are elongate and they lack auto-
omic septa. The transverse processes are broken off but their basesre preserved; they slightly extend anteroposteriorly and they areocated more posteriorly than in anguines. The ventral face of theentrum is well limited laterally; it is flat on the smaller vertebraVBO 55) but a median groove runs through most of the centrumength of the larger vertebra (VBO 54). The latter vertebra occu-ied a more anterior position in the caudal region than the smallertebra. Two short pedicles, on which the chevron bones wererticulated, are located very close to the condyle; the articular areasf the pedicles face posteroventrally.
Discussion. The vertebrae are similar to those reported by Hechtnd Hoffstetter (1962) and Rage (1978). The absence of autotomicepta, presence of pedicles for articulation with chevron bones
ontologie 101 (2015) 29–41
located very close to the condyle, and position of the transverseprocesses argue for referral to Necrosaurus.
These vertebrae confirm the presence of Necrosaurus in theOligocene; rather common in the late Eocene, it survived theGrande Coupure but became unfrequent in the Oligocene. Itsurvived up to MP 21 in Belgium (Augé and Smith, 2009) andEngland (Klembara and Green, 2010) and up to MP 23 in Franceand Germany (Augé, 2005; Böhme and Ilg, 2003). Necrosauruswas also reported, but not described or figured, from the latestOligocene (MP 29 of Herrlingen 9) of Germany by Böhme and Ilg(2003); this isolated occurrence remains to be confirmed. Withthis in mind, it should be noted that shinisaurids were reportedfrom the latest Oligocene and early Miocene of Germany (Böhme,2008; Cernansky et al., 2014) and from the early Miocene of theCzech Republic (Klembara, 2008) and that distinction betweenNecrosaurus and shinisaurids may be problematic if the availablematerial is limited to a few isolated bones.
Clade AMPHISBAENIA Gray, 1844Family AMPHISBAENIDAE Gray, 1865
Indeterminate genus and speciesFig. 4Material. Valbro A: one left dentary (VBO 21); nine trunk verte-
brae (VBO 22); one caudal vertebra (VBO 23). Valbro déblais: oneright dentary (VBO 56); one incomplete premaxilla (VBO 57); 30trunk vertebrae (VBO 58–60); two caudal vertebrae (VBO 61). Val-bro C: perhaps one incomplete right dentary (VBO 32) and perhapsone trunk vertebra (VBO 33).
Description. The dentary is robust, short and deep. It bears ahigh coronoid process and the ventral border of the bone is straightto slightly convex. The angle formed by the ventral border, poste-rior to the symphysis, is weak and rounded. On the medial face,the Meckelian groove widely opens posteriorly whereas it is nar-row anteriorly. In the posterior part of the Meckelian groove, alarge intramandibular septum is exposed; it extends posteriorlyand its ventral edge is welded to the lateral wall of the bone atmid-depth. The articular facet for the angular forms an elongatedepression located on the ventral margin of the septum. A highcoronoid process and an angular process are present posteriorly.The dentition comprises six pleurodont, robust and unicuspid teeththat are inclined anteriorly. The anterior two teeth are robust andlarger than the more posterior ones. Tooth bases are rather broad,but the shaft clearly narrows in the upper half. The lingual face ofthe teeth is slightly concave just beneath the apex. The tooth apicesare rather blunt and they bear some striae.
The right lateral process of the premaxilla is broken off. In ante-rior aspect, the nasal process is short; some small foramina occuron its dorsalmost part whereas one larger foramen opens on eitherside of its base. Four teeth are completely preserved, including themedian one that is similar to the first two teeth of the dentary. Thepreserved lateral process bears three teeth that are smaller thanthe median one but that shows the same morphology.
Trunk vertebrae display the typical morphology of amphisbae-nians: they are depressed, the ventral face of the centrum is almostflat and its lateral margins are practically parallel, the neural spineis not well developed, and articular facets of the zygapophyses arerelatively broad. In the largest vertebra, the centrum is 3.1 mm long.These vertebrae are characterized by the presence of a low neuralspine and by the weak concavity of the posterior margin of theneural arch. The neural spine comprises a posterior low but trans-
salient anterior keel. On several vertebrae, the anterolateral bordersof the dorsal face of the mound form small anterior projectionsthat can take the shape of two short spines (Fig. 4E). The mound
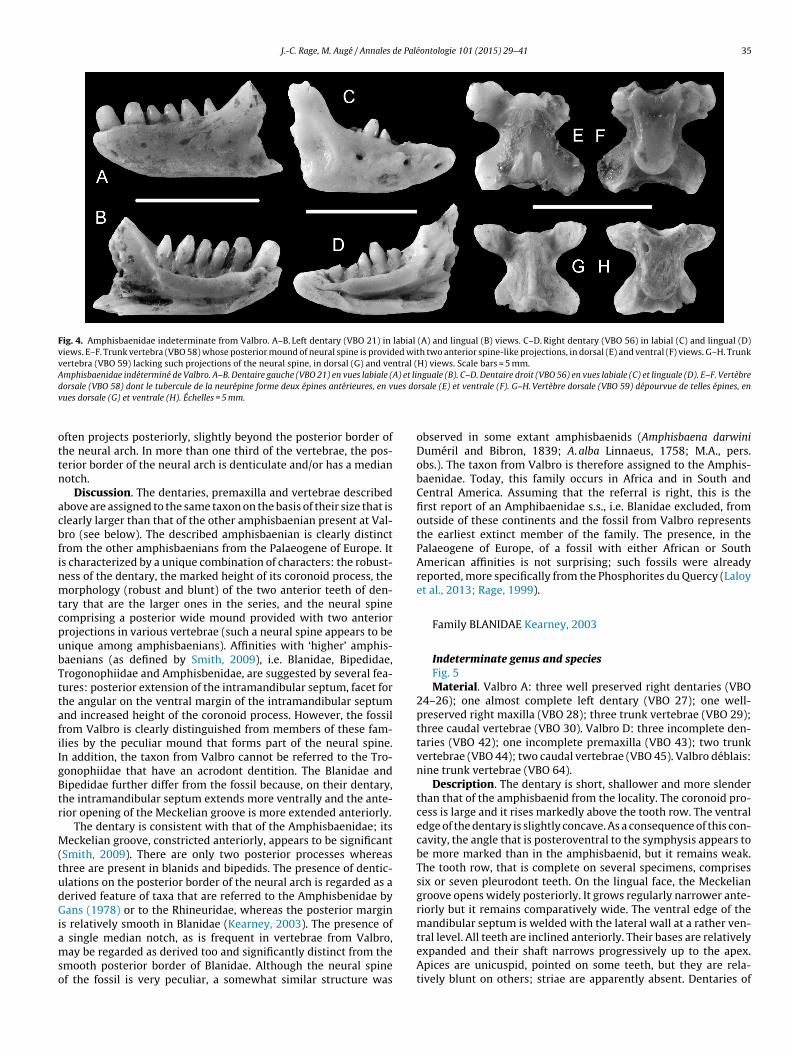
J.-C. Rage, M. Augé / Annales de Paléontologie 101 (2015) 29–41 35
Fig. 4. Amphisbaenidae indeterminate from Valbro. A–B. Left dentary (VBO 21) in labial (A) and lingual (B) views. C–D. Right dentary (VBO 56) in labial (C) and lingual (D)views. E–F. Trunk vertebra (VBO 58) whose posterior mound of neural spine is provided with two anterior spine-like projections, in dorsal (E) and ventral (F) views. G–H. Trunkvertebra (VBO 59) lacking such projections of the neural spine, in dorsal (G) and ventral (H) views. Scale bars = 5 mm.Amphisbaenidae indéterminé de Valbro. A–B. Dentaire gauche (VBO 21) en vues labiale (A) et linguale (B). C–D. Dentaire droit (VBO 56) en vues labiale (C) et linguale (D). E–F. Vertèbred ues dov
ottn
acbfinmtcpubTttafiIgBtr
M(tudGiamso
orsale (VBO 58) dont le tubercule de la neurépine forme deux épines antérieures, en vues dorsale (G) et ventrale (H). Échelles = 5 mm.
ften projects posteriorly, slightly beyond the posterior border ofhe neural arch. In more than one third of the vertebrae, the pos-erior border of the neural arch is denticulate and/or has a medianotch.
Discussion. The dentaries, premaxilla and vertebrae describedbove are assigned to the same taxon on the basis of their size that islearly larger than that of the other amphisbaenian present at Val-ro (see below). The described amphisbaenian is clearly distinctrom the other amphisbaenians from the Palaeogene of Europe. Its characterized by a unique combination of characters: the robust-ess of the dentary, the marked height of its coronoid process, theorphology (robust and blunt) of the two anterior teeth of den-
ary that are the larger ones in the series, and the neural spineomprising a posterior wide mound provided with two anteriorrojections in various vertebrae (such a neural spine appears to benique among amphisbaenians). Affinities with ‘higher’ amphis-aenians (as defined by Smith, 2009), i.e. Blanidae, Bipedidae,rogonophiidae and Amphisbenidae, are suggested by several fea-ures: posterior extension of the intramandibular septum, facet forhe angular on the ventral margin of the intramandibular septumnd increased height of the coronoid process. However, the fossilrom Valbro is clearly distinguished from members of these fam-lies by the peculiar mound that forms part of the neural spine.n addition, the taxon from Valbro cannot be referred to the Tro-onophiidae that have an acrodont dentition. The Blanidae andipedidae further differ from the fossil because, on their dentary,he intramandibular septum extends more ventrally and the ante-ior opening of the Meckelian groove is more extended anteriorly.
The dentary is consistent with that of the Amphisbaenidae; itseckelian groove, constricted anteriorly, appears to be significant
Smith, 2009). There are only two posterior processes whereashree are present in blanids and bipedids. The presence of dentic-lations on the posterior border of the neural arch is regarded as aerived feature of taxa that are referred to the Amphisbenidae byans (1978) or to the Rhineuridae, whereas the posterior margin
s relatively smooth in Blanidae (Kearney, 2003). The presence of
single median notch, as is frequent in vertebrae from Valbro,ay be regarded as derived too and significantly distinct from themooth posterior border of Blanidae. Although the neural spinef the fossil is very peculiar, a somewhat similar structure was
rsale (E) et ventrale (F). G–H. Vertèbre dorsale (VBO 59) dépourvue de telles épines, en
observed in some extant amphisbaenids (Amphisbaena darwiniDuméril and Bibron, 1839; A. alba Linnaeus, 1758; M.A., pers.obs.). The taxon from Valbro is therefore assigned to the Amphis-baenidae. Today, this family occurs in Africa and in South andCentral America. Assuming that the referral is right, this is thefirst report of an Amphibaenidae s.s., i.e. Blanidae excluded, fromoutside of these continents and the fossil from Valbro representsthe earliest extinct member of the family. The presence, in thePalaeogene of Europe, of a fossil with either African or SouthAmerican affinities is not surprising; such fossils were alreadyreported, more specifically from the Phosphorites du Quercy (Laloyet al., 2013; Rage, 1999).
Family BLANIDAE Kearney, 2003
Indeterminate genus and speciesFig. 5Material. Valbro A: three well preserved right dentaries (VBO
24–26); one almost complete left dentary (VBO 27); one well-preserved right maxilla (VBO 28); three trunk vertebrae (VBO 29);three caudal vertebrae (VBO 30). Valbro D: three incomplete den-taries (VBO 42); one incomplete premaxilla (VBO 43); two trunkvertebrae (VBO 44); two caudal vertebrae (VBO 45). Valbro déblais:nine trunk vertebrae (VBO 64).
Description. The dentary is short, shallower and more slenderthan that of the amphisbaenid from the locality. The coronoid pro-cess is large and it rises markedly above the tooth row. The ventraledge of the dentary is slightly concave. As a consequence of this con-cavity, the angle that is posteroventral to the symphysis appears tobe more marked than in the amphisbaenid, but it remains weak.The tooth row, that is complete on several specimens, comprisessix or seven pleurodont teeth. On the lingual face, the Meckeliangroove opens widely posteriorly. It grows regularly narrower ante-riorly but it remains comparatively wide. The ventral edge of themandibular septum is welded with the lateral wall at a rather ven-
tral level. All teeth are inclined anteriorly. Their bases are relativelyexpanded and their shaft narrows progressively up to the apex.Apices are unicuspid, pointed on some teeth, but they are rela-tively blunt on others; striae are apparently absent. Dentaries of
36 J.-C. Rage, M. Augé / Annales de Palé
Fig. 5. Blanidae from Valbro: right maxilla (VBO 28) in dorsal (A), labial (B) andlB(
bea
bttbiialdsiiisTsM(
bbimpo2
apa
ingual (C) views. Scale bar = 2 mm.lanidae de Valbro : maxillaire droit (VBO 28) en vues dorsale (A), labiale (B) et lingualeC). Échelles = 2 mm.
lanids were illustrated by, e.g. Bailon (2000), Blain (2009), Blaint al. (2007), Bolet et al. (2014), Schleich (1985, 1988) and Venczelnd S tiuca (2008).
The maxilla is well preserved. It bears 4 teeth, the second oneeing larger than the others. The labial face is smooth. Anteriorly,he nasal process rises steeply; it reaches its maximum height abovehe second tooth. From the top of the process, the posterodorsalorder goes down more or less regularly up to the posterior extrem-
ty of the maxilla. At the top of the nasal process, the frontal processs not distinctly shaped. The premaxillary process extends mediallynd is curved dorsally; it does not form a rostral process. On theingual side, a large, vertically elongate depression occurs on theorsal area of the nasal process; it likely corresponds to a contacturface with the prefrontal. The dorsal face of the supradental tables broad; it markedly rises anteriorly, forming part of the premax-llary process. Posteriorly, the lingual edge of the supradental tables notched. A depression located anteriorly on the table faces dor-olingually. Posterior to the depression, a process projects medially.eeth are pleurodont and unicuspid. The first three teeth are strong,lightly recurved and have expanded bases. The last tooth is small.axillae of blanids were illustrated in Bolet et al. (2014), Delfino
1997) and Gans and Montero (2008).The vertebrae show the generalized morphology of amphis-
aenians. They differ from those assigned to Amphisbaenidae ineing smaller (the centrum length does not exceed 2.4 mm) and
n lacking a neural spine, except a scarcely discernible ridge in theid-portion of the neural arch of some vertebrae. In addition, the
osterior margin of the neural arch lacks a median notch. Vertebraef blanids were often illustrated (e.g., Blain, 2009; Blain et al., 2007,008; Delfino and Bailon, 2000; Venczel and S tiuca, 2008).
Discussion. The referral of these skull bones and vertebrae tomphisbaenians is unquestionable. The dentaries show apomor-hies of ‘higher’ Amphisbaenia as defined by Smith (2009), anssemblage comprised of all amphisbaenians except Rhineuridae.
ontologie 101 (2015) 29–41
Within this group, the dentaries and the maxilla cannot belong tothe Trogonophiidae because teeth of the latter are acrodont. Thedentaries display characters that do not occur in amphisbaenidsbut are present in blanids and bipedids: Meckelian groove rela-tively wide anteriorly and junction between the intramandibularseptum and the lateral wall in a ventral position. Besides, the com-bination of features of the maxilla described above occurs in Blanus(Augé, 2012; Bailon, 1989) but is lacking in amphisbaenids (e.g.,Gans and Montero, 2008; Montero and Gans, 1999) and bipedids(Smith, 2009). The vertebrae are tentatively assigned to this blanid.They do not show derived characters that would permit securereferral to this taxon. However their size is consistent with theskull bones and it is smaller than that of the amphisbaenid verte-brae reported from the locality. In addition, the posterior border oftheir neural arches lacks a median notch, whereas an indentation ispresent in Togonophiidae, Rhineuridae and most Amphisbaenidae.The earliest Blanidae was reported from the earliest Eocene (MP 7)of Belgium (Folie et al., 2013).
AMPHISBAENIANS INDETERMINATEMaterial. Valbro A: five poorly preserved dentaries (VBO 31).
Valbro déblais: two poorly preserved dentaries (VBO 65); 19 trunkvertebrae (VBO 66); three caudal vertebrae (VBO 67).
Suborder SERPENTES Linnaeus, 1758ANILIOID-GRADE SNAKEGenus Eoanilius Rage, 1974
Eoanilius aff. E. oligocenicus Szyndlar, 1994Fig. 6A–CMaterial. Valbro A: one anterior (VBO 502) and one posterior
(VBO 501) trunk vertebrae. Valbro déblais: three mid-trunk verte-brae (VBO 522, 523, 524). Valbro C: perhaps one trunk vertebra(VBO 513).
Description. These small vertebrae (longest centrum = 2.4 mm)are depressed, short and wide. Their neural arches are markedlydepressed. The median notch in the posterior border of the neuralarch is well marked. The neural spine is low and short; it occupiesless than half the length of the neural arch. The anterior border ofthe neural spine is inclined posteriorly in anterior-trunk vertebraebut it rises more steeply in more posterior vertebrae. The articularfacets of the prezygapophyses are approximately horizontal andtheir major axis is oblique in dorsal aspect. Small prezygapophysealprocesses are present; they are deep but they scarcely project later-ally. The interzygapophyseal constriction is deep. The zygospheneis dorsoventrally thin, wide and provided with three lobes thatweakly project anteriorly. The narrow lateral lobes flank a broadmedian lobe whose anterior border may be slightly concave inposterior-trunk vertebrae. The massive paradiapophyses lack anytrace of division into dia- and parapophyseal areas. The cotyle andcondyle are slightly depressed. The centrum widens anteriorly butit remains comparatively narrow. The haemal keel is spatulate togladiate (sensu Auffenberg, 1963); it is clearly marked off from thecentrum in the anterior- and posterior-trunk regions.
Discussion. Eoanilius may be confused with another smallsnake, Platyspondylia, which is present in the Eocene and Oligoceneof the Phosphorites du Quercy (Rage, 2006). Both have vertebraewith a depressed neural arch, low neural spine and relativelynarrow centrum. However, the neural arch of Platyspondylia ismore strongly depressed and the neural spine of Eoanilius is
anteroposteriorly shorter. Two species of Eoanilius were described:E. europae Rage, 1974, from the late middle Eocene and late Eocene(MP 16 to MP 19, perhaps MP 20; Rage, 2006) of France andE. oligocenicus Szyndlar, 1994, from the Oligocene (MP 22 to MP
J.-C. Rage, M. Augé / Annales de Paléontologie 101 (2015) 29–41 37
Fig. 6. Snakes from Valbro. A–C. Eoanilius aff. E. oligocenicus, trunk vertebra (VBO 501) in anterior (A), right lateral (B) and dorsal (C) views. D–H. Coluber cadurci, posteriormosttrunk vertebra (VBO 533) in dorsal (D), left lateral (E), ventral (F), anterior (G) and posterior (H) views. Scale bars = 5 mm.S ues anp (G) et
2aclibsbsziEvE
21ygt(
5sv
ttQ
erpents de Valbro. A–C. Eoanilius aff. E. oligocenicus, vertèbre dorsale (VBO 501) en vostérieure (VBO 533) en vues dorsale (D), latérale gauche (E), ventrale (F), antérieure
8) of Germany. E. oligocenicus differs from E. europae in having larger neural spine (that has a well-defined anterior border), aonspicuous haemal keel, a three lobed zygosphene, comparativelyarger paradiapophyses, and distinct subcentral and lateral foram-na (Szyndlar, 1994). The presence or absence of tiny foramina maye ascribed to fossilization. The other characters show that thenake from Valbro is similar to E. oligocenicus; however, the verte-rae from Valbro slightly differ from those of Germany: their neuralpine is slightly higher and, where well preserved, the lobes of theygosphene are less projecting. The significance of these differencess unknown. However, the form from Valbro is distinct from theocene species and it may belong to E. oligocenicus; at least, it isery closely related to the latter and it is referred to as Eoanilius aff.. oligocenicus.
The earliest E. oligocenicus comes from the early Oligocene (MP2) of Germany (Szyndlar, 1994) and perhaps France (Sigé et al.,998; the present report). Venczel and Sanchiz (2006) reported theoungest occurrence of the species from Oschiri, Sardinia, whoseeological age was not constrained precisely. The geological age ofhe locality was regarded as MN 1–5; however, Rook and Kotsakis2005) assigned Oschiri to the MN 1 zone.
BOOID-GRADE SNAKESFamily BOIDAE Gray, 1825
Boidae CBoidae H Rage, 2006: 167Material. Valbro A: 15 trunk vertebrae from Valbro A (VBO
03–508). Valbro D: one trunk vertebra (VBO 515). Valbro déblais:ix trunk vertebrae (VBO 525, 526) and perhaps 11 other trunkertebrae (VBO 527).
Preliminary remarks. This unnamed boid snake was firstermed ‘Boidae C’ by de Bonis et al. (1973: tab. 2(5)). It occurs inhe late Eocene and the early Oligocene of the Phosphorites duuercy (Rage, 2006). This taxon displays comparatively important
térieure (A), latérale droite (B) et dorsale (C). D–H. Coluber cadurci, vertèbre dorsalepostérieure (H). Échelles = 5 mm.
variation and it is never very frequent in the localities, whichrendered its description difficult. However, the specimens arenow sufficiently abundant for the erection of a new taxon, but thematerial from Valbro cannot represent the base for a description.Samples from Escamps (MP 19) or Ravet (MP 21) appear to bemore appropriate. The following description provides only themain characteristics of boid C.
Description. Boid C is a small to mid-sized Boidae (centrumlength of the largest vertebra: 4.3 mm). The vertebrae show a com-bination of features that points to the Boidae: vertebrae short, wideand massive; prezygapophyseal processes small, not projecting;paradiapophyseal surface weakly subdivided; haemal keel shallow;neural arch not strongly depressed.
Within boids, boid C is characterized by its moderately high neu-ral spine whose posterior border is clearly overhanging, its narrowand projecting central lobe of zygosphene, the oblique axis of itszygapophyseal facets, the shallowness of the projecting parts of itsprezygapophyses, and its moderately vaulted neural arch whoseposterodorsal borders are weakly curved in posterior aspect butthat slightly bulge above the zygantral fossae.
Discussion. Boid C is the only snake that survived the GrandeCoupure in the Phosphorites du Quercy. It was reported from MP 16to MP 21, i.e. from the latest middle Eocene to the earliest Oligocene(Rage, 2006: tab. 3). However, another unnamed snake, namely‘Boidae H’, restricted to MP 22 (Rage, 2006: tab. 3) appears to liewithin the range of variations of boid C and we refer it to the syn-onymy of the latter. It should be noted that boid H was reportedby de Bonis et al. (1973) from the Phosphorites du Quercy; at thattime, boid H was identified from MP 22 (Mas de Got A) and MP28 (Pech du Fraysse). Subsequently, Szyndlar and Rage (2003: 14)shown that vertebrae of boid H from Pech du Fraysse were actu-ally juveniles of the large boine Bavarioboa crocheti Szyndlar and
Rage, 2003. Identification of boid C at Valbro is consistent withreferral of boid H from Mas de Got A to this taxon. Therefore,the stratigraphic range of boid C should be extended up to MP22.
3 de Palé
(nfattHd
lItltdib
o5
Vr5
ca1bmzpppvsdbncgIta(
8 J.-C. Rage, M. Augé / Annales
Boidae indeterminate AMaterial. Valbro déblais: one trunk vertebra (VBO 528).Description and discussion. This comparatively small vertebra
centrum length: 3.7 mm) differs from boid C in having a highereural spine (taking into account the fact that the vertebra is
rom the posterior region of the trunk) and a clearly overhangingnterior border of neural spine. The other features (includinghe characteristic anterior border of the zygosphene) are similaro those of boid C. This vertebra may be a variant of boid C.owever, the shape of the neural spine appears to be clearlyifferent.
Boidae indeterminate BMaterial. Valbro déblais: one trunk vertebra (VBO 529).Description and discussion. This vertebra is as large as the
argest ones of boid C; the length of its centrum reaches 4.5 mm.t differs from boid C and boid indeterminate A by its clearlyaller neural spine and more vaulted neural arch. The vertebra isess depressed, narrower than those of the two other boids fromhe locality. The zygosphene is eroded and its anterior border isamaged. This vertebra unquestionably represents a distinct boid
n the locality, but this single, slightly damaged specimen cannote compared to other extinct boids.
Booids indeterminateMaterial. Valbro A: three trunk vertebrae (VBO 509). Valbro B:
ne trunk vertebra (VBO 512). Valbro D: two trunk vertebrae (VBO16). Valbro déblais: about 30 trunk vertebrae (VBO 528–530).
Family COLUBRIDAE s.l. Oppel, 1811Genus Coluber s.l. Linnaeus, 1758
Coluber cadurci Rage, 1974Fig. 6D–HColubrinae A de Bonis et al., 1973: tab. 2(5)Coluber cadurci Rage, 1984: 44Coluber cadurci Rage, 1988b: 468Coluber cadurci Szyndlar and Schleich, 1993: 40Coluber cadurci Szyndlar, 2012: 664Material. Valbro A: six mid-trunk vertebrae (VBO 510, 511).
albro déblais: 15 mid-trunk vertebrae (VBO 531, 532), one poste-iormost trunk vertebra (VBO 533) and five caudal vertebrae (VBO34). Valbro D: perhaps one mid-trunk vertebra (VBO 517).
Description. The vertebrae show the typical morphology ofolubrines belonging to the ‘Coluber assemblage’. The specimensre mainly characterized by the following features (Rage, 1974,988b): hypapophyses absent on mid- and posterior-trunk verte-rae; haemal keel well-defined, narrow and deep; neural archoderately vaulted with posterodorsal borders barely arched;
ygosphene with a broad, rounded median lobe and two narrow,ointed lateral lobes; prezygapophyseal processes elongate andointed on mid-trunk vertebrae; neural spine low and antero-osteriorly long. However, at the time of the species description,ertebrae from the most posterior-trunk region were not available;uch one vertebra is present at Valbro, which is an opportunity toescribe it. As is usual in snakes, this posteriormost trunk verte-ra differs from those of the mid-trunk region in having a lowereural spine, a more depressed neural arch, larger and marked para-otylar notches (sensu LaDuke, 1991), and well-defined subcentralrooves (subcentral paramedian lymphatic fossae; LaDuke, 1991).
n addition to these usual variations, the posteriormost trunk ver-ebra from Valbro has blunt, shorter prezygapophyseal processesnd the central lobe of the zygosphene is wider and less projectingits anterior border is slightly concave).ontologie 101 (2015) 29–41
Discussion. This small to mid-sized ‘colubrine’ belongs to anassemblage of genera that displays a very homogenous vertebralmorphology and within which distinction of taxa is very difficult(Rage, 1974). The living genus Coluber is a member of this assem-blage and the fossil species was assigned to this genus only for thisreason, however it is not possible to demonstrate that the extinctspecies really belongs to Coluber as it is now understood.
C. cadurci is the oldest colubrid from Europe. It first occurs inthe earliest Oligocene (MP 21) of the Phosphorites du Quercy.It reaches the end of the Oligocene (MP 30), perhaps MN 2 inthe early Miocene (Rage, 2006). Szyndlar and Alférez (2005) andSzyndlar (2012) even reported cf. ‘Coluber’ cadurci from the lateearly Miocene (MN 3/4) of Spain. However, in the Phosphorites duQuercy, C. cadurci was not recovered from levels younger than MP25 (Rage, 2006).
SERPENTES INDETERMINATEMaterial. Valbro D: one caudal vertebra (VBO 518).
3. Discussion
The fauna of amphibians and squamates from Valbro is not richbut it is relatively diverse. Fifteen taxa are present (four amphibiansand eleven squamates). Only three taxa are identified at species orclose to species level. Some cannot be determined below familylevel. This results from either the low number and/or poor preser-vation of specimens or from the fact that faunas from the Oligoceneof the Phosphorites remain insufficiently studied.
3.1. Biostratigraphy
The fauna includes both taxa that were already present beforethe Grande Coupure, i.e. the extinction event that strongly affectedsquamates at the Eocene-Oligocene transition (Rage, 2012), andtaxa that reached Europe after this event. The genera Necrosaurusand perhaps Megalotriton, as well as the species boid C, wererecorded from both the Eocene and the Oligocene of the Phos-phorites du Quercy. Necrosaurus is a typical member of the lateEocene assemblages of the Phosphorites. It is rare after the GrandeCoupure; it occurs in only a few localities of early Oligocene age (MP21–MP 23), it being understood that the report of the genus fromthe late Oligocene of Germany (see above) cannot be accepted with-out reservation. The large salamander from Valbro perhaps belongsto Megalotriton that is present in the late Eocene, but this cannot bedefinitely confirmed. It cannot be demonstrated whether the formfrom the early Oligocene really belongs to this genus or is a dis-tinct taxon. These large salamanders are rare both in the Eoceneand the Oligocene, which does not permit a thorough study. Inthe Oligocene, the large salamander occurs from MP 21 to MP 23.Boidae C is present in the late Eocene and in the early Oligocene. Thesynonymization of Boidae H with Boidae C shows that this taxonreached MP 22. Boidae C is the only species from Valbro that waspresent in the Eocene; it is the only true survivor of the GrandeCoupure in the locality.
Newcomers include U. europaeus, Eoanilius aff. E. oligocenicus,Coluber cadurci and the indeterminate amphisbaenid. C. cadurci firstoccurs just after the Grande Coupure, in MP 21 of Ravet in the Phos-phorites. Although this species perhaps reaches the early Miocene(Szyndlar, 2012; Szyndlar and Alférez, 2005), in the Phosphoritesit was not recovered from the three younger levels (MP 26–MP28). C. cadurci was the most common colubrid in the Oligocene,
but specimens are never numerous in each locality. E. oligocenicusranges from MP 22 to the early Miocene. The form found at Val-bro is perhaps distinct from the species but it is, at least, very closeto it. The known stratigraphic range of U. europaeus is astonishing
de Palé
btai
OTlNEct(
3
akwrtbaianeaL1
gM
sbea
FanRfc
J.-C. Rage, M. Augé / Annales
ecause it is restricted to the level MP 22. This may be a bias, buthe species may be regarded as indicative of an early Oligocenege. The indeterminate amphisbaenid does not afford stratigraphicnformation.
The stratigraphic overlap of holdovers from the Eocene andligocene newcomers clearly points to an early Oligocene age.axa already known before the Grande Coupure did not remainong in existence after this event. More specifically, the overlap ofecrosaurus, Boidae C and cf. Megalotriton, on the one hand, andoanilius aff. E. oligocenicus, and U. europaeus on the other hand,learly suggests a MP 22 age for Valbro. This perfectly matcheshe age inferred from various mammalian groups by Peigné et al.2014).
.2. Palaeoenvironments
As is usual in the localities from the Phosphorites, amphibiansre by far less frequent than squamates, as frequently happens in aarstic environment. Amphibians are dependent on moisture and,ith very rare exceptions, they need water at least at the time of
eproduction. The frog of the ‘Discoglossus group’ likely requiredhe presence of permanent fresh water as all taxa of this assem-lage do. On the contrary, the lizard Uromastyx inhabits dry, aridreas. Amphisbaenians are fossorial snakelike squamates that liven dry or relatively dry areas, although some of them do not avoidquatic environments. They make tunnels into the soil, which isot consistent with proximity of water. The snake Eoanilius wasither fossorial or secretive. Moisture and likely presence of water,s pools, are not inconsistent with close dry to arid zones (Vianey-iaud et al., in press), mainly in a karstic area (Augé and Rage,995).
The presence of Uromastyx, amphisbaenians and Boidae sug-ests that temperatures were not lower than those of the presentediterranean region and were perhaps higher.It should be noted that amphisbaenians are numerous. These
quamates occur in almost all localities of the European Cenozoic,
ut at Valbro their number is unusually high. Such a ratio wasncountered only in the late Palaeocene (MP 6) of Cernay, France,nd the earliest Eocene (MP 7) of Dormaal, Belgium (Augé, 2005).ig. 7. Distribution of taxa in the loci of Valbro (note that Valbro B produced only fragment of bone assigned to an indeterminate lizard; consequently, the locus isot included in the table).épartition des taxons dans les locus de Valbro (noter que Valbro B n’a fourni qu’un
ragment d’os appartenant à un lézard indéterminé ; le locus ne figure donc pas dansette table).
ontologie 101 (2015) 29–41 39
This high number of amphisbaenians likely results from a localenvironmental cause that cannot be determined.
4. Conclusions
The assemblage of amphibians and squamates from Valbro is notrich but it is diverse (Fig. 7). The fauna includes both forms presentin the Eocene, before the Grande Coupure, and taxa that arrived inWestern Europe during the early Oligocene. This herpetofauna isindicative of a MP 22 age (early Oligocene), an age already suggestedby the mammals of the same locality (Peigné et al., 2014). Valbro isthe only fauna of amphibians and squamates from MP 22 that hasbeen entirely studied. Assemblages from MP 21 (i.e., the earlieststandard level of the Oligocene) and from MP 23 have not beendescribed yet but their compositions are known (Rage, 2006: tab.3). However, the fauna from Valbro is richer and more diverse thanthose from MP 21. It appears to be less rich and less diverse thanthe faunas from MP 23, mainly as far as amphibians are concerned.Therefore, MP 22, as illustrated by Valbro, represents a pivotal levelin the renewal of the Paleogene faunas after the Grande Coupure.
Disclosure of interest
The authors declare that they have no conflicts of interest con-cerning this article.
Acknowledgements
We first thank the team that collected the material. Weare grateful to M. Delfino and M. Venczel who provided signif-icant information and remarks that permitted to improve themanuscript. We thank S. Bailon (MNHN, Paris) for access to extantcomparative material. L. Cazes and P. Loubry took the photographs.T. Pélissié and B. Sigé initiated the series of articles devoted to theValbro fauna.
References
Auffenberg, W., 1963. The fossil snakes of Florida. Tulane Studies in Zoology 10,131–216.
Augé, M., 1988. Révision du lézard Uromastyx europaeus (Reptilia, Lacertilia) del’Oligocène franc ais. Revue de Paléobiologie 7, 317–325.
Augé, M., 1992. Une nouvelle espèce d’Ophisaurus (Lacertilia, Anguidae) del’Oligocène des Phosphorites du Quercy. Révision de la sous-famille desAnguinae. Paläontologische Zeitschrift 66, 159–175.
Augé, M., 2005. Évolution des lézards du Paléogène en Europe. Mémoires du Muséumnational d’Histoire naturelle 192, 5–369.
Augé, M., 2012. Amphisbaenians from the European Eocene: a biogeographicalreview. In: Lehmann, T., Schaal, S.F.K. (Eds.), Messel and the terrestrial Eocene.Palaeobiodiversity and Palaeoenvironments 92, 425–443.
Augé, M., Rage, J.C., 1995. Le Garouillas et les sites contemporains (Oligocène, MP25) des Phosphorites du Quercy (Lot, Tarn-et-Garonne, France) et leurs faunesde Vertébrés. 2. Amphibiens et Squamates. Palaeontographica A 236, 11–32.
Augé, M., Smith, R., 1997. Les Agamidae (Reptilia, Squamata) du Paléogène d’Europeoccidentale. Belgian Journal of Zoology 127, 123–138.
Augé, M., Smith, R., 2009. An assemblage of early Oligocene lizards (Squamata) fromthe locality of Boutersem (Belgium), with comments on the Eocene–Oligocenetransition. Zoological Journal of the Linnean Society 155, 148–170.
Averianov, A., Danilov, I., 1996. Agamid lizards (Reptilia, Sauria, Agamidae) fromthe Early Eocene of Kyrgyzstan. Neues Jahrbuch für Geologie und PaläontologieMonatshefte 12, 739–750.
Bailon, S., 1989. Les amphibiens et les reptiles du Pliocène supérieur de Balaruc II(Hérault, France). Palaeovertebrata 19, 7–28.
Bailon, S., 2000. Amphibiens et reptiles du Pliocène terminal d’Ahl al Oughlam(Casablanca, Maroc). Geodiversitas 22, 539–558.
Biton, R., Geffen, E., Vences, M., Cohen, O., Bailon, S., Rabinovich, R., Malka, Y., Oron,T., Boistel, R., Brumfeld, V., Gafny, S., 2013. The rediscovered Hula painted frog
is a living fossil. Nature Communications 4, 1–6.Blain, H.A., 2009. Contribution de la paléoherpétofaune (Amphibia & Squamata) àla connaissance de l’évolution du climat et du paysage du Pliocène supérieur auPléistocène moyen d’Espagne. Treballs del Museu de Geologia de Barcelona 16,39–170.

4 de Palé
B
B
B
B
B
C
C
C
C
d
D
D
D
D
D
D
E
E
E
E
F
G
G
H
H
H
K
K
K
K
0 J.-C. Rage, M. Augé / Annales
lain, H.A., Bailon, S., Agustí, J., 2007. Anurans and squamate reptiles from the latestearly Pleistocene of Almenara-Casablanca-3 (Castellón, East of Spain). System-atic, climatic and environmental considerations. Geodiversitas 29, 269–295.
lain, H.A., Bailon, S., Agustí, J., 2008. Amphibians and squamate reptiles fromthe latest early Pleistocene of Cueva Victoria (Murcia, southeastern Spain, SWMediterranean): paleobiogeographic and paleoclimatic implications. GeologicaActa 6, 345–361.
öhme, M., 2008. Ectothermic vertebrates (Teleostei, Allocaudata, Urodela, Anura,Testudines, Choristodera, Crocodylia, Squamata) from the Upper Oligoceneof Oberleichtersbach (Northern Bavaria, Germany). Courier ForschungsinstitutSenckenberg 260, 161–183.
öhme, M., Ilg, A., 2003. fosFARbase (access date: March 2014) http://www.wahre-staerke.com/
olet, A., Delfino, M., Fortuny, J., Almécija, S., Robles, J.M., Alba, D.M., 2014. AnAmphisbaenian skull from the European Miocene and the evolution of Mediter-ranean worm lizards. Plos One 9, e98082 (1–16).
ˇernansky, A., 2010. A revision of chamaeleonids from the Lower Miocene of theCzech Republic with description of a new species of Chamaeleo (Squamata,Chamaeleonidae). Geobios 43, 605–613.
ˇernansky, A., Augé, M., 2013. New species of the genus Plesiolacerta (Squamata: Lac-ertidae) from the upper Oligocene (MP 28) of Southern Germany and a revisionof the type species Plesiolacerta lydekkeri. Palaeontology 56, 79–94.
ˇernansky, A., Rage, J.C., Klembara, J., 2014. The Early Miocene squamates ofAmöneburg (Germany): the first stages of modern squamates in Europe.Journal of Systematic Palaeontology, http://dx.doi.org/10.1080/14772019.2014.897266.
onrad, J.L., 2008. Phylogeny and systematics of Squamata (Reptilia) basedon morphology. Bulletin of the American Museum of Natural History310, 1–182.
e Bonis, L., Crochet, J.Y., Rage, J.C., Sigé, B., Sudre, J., Vianey-Liaud, M., 1973. Nou-velles faunes de vertébrés oligocènes des Phosphorites du Quercy. Bulletin duMuséum national d’Histoire naturelle 174, 105–113.
elfino, M., 1997. Blanus from the Early Pleistocene of Southern Italy: another smalltessera from a big mosaic. In: Böhme, W., Bischoff, W., Ziegler, T. (Eds). Herpeto-logica Bonnensis, pp. 89–97.
elfino, M., Bailon, S., 2000. Early Pleistocene herpetofauna from Cava Dell’Erba andCava Pirro (Apulia, southern Italy). Herpetological Journal 10, 95–110.
elfino, M., Bailon, S., Pitruzella, G., 2011. The Late Pliocene amphibians and reptilesfrom ‘Capo Mannu D1 Local Fauna’ (Mandriola, Sardinia, Italy). Geodiversitas 33,357–382.
elfino, M., Kotsakis, T., Arca, M., Tuveri, C., Pitruzella, G., Rook, L., 2008. Agamidlizards from the Plio-Pleistocene of Sardinia (Italy) and an overview of the Euro-pean fossil record of the family. Geodiversitas 30, 641–656.
e Stefano, G., 1903. I Sauri del Quercy appartenenti alla collezione Rossignol. Attidella Società Italiana di Scienze naturali e del Museo Civico di Storia naturale inMilano 42, 382–418.
uffaud, S., 2000. Les faunes d’amphibiens du Crétacé supérieur à l’Oligocène enEurope : paléobiodiversité, évolution, mise en place. Thèse du Muséum nationald’Histoire naturelle, Paris, 221 p. (inédit).
dwards, J.L., 1976. Spinal nerves and their bearing on salamander phylogeny. Jour-nal of Morphology 148, 305–328.
stes, R., 1981. Gymnophiona, Caudata. Handbuch der Paläoherpetologie, 2. GustavFischer Verlag, Stuttgart, 115 p.
stes, R., 1983. Sauria terrestria, Amphisbaenia. Handbuch der Paläoherpetologie,10A. Gustav Fischer Verlag, Stuttgart, 249 p.
stes, R., de Queiroz, K., Gauthier, J., 1988. Phylogenetic relationships within Squa-mata. In: Estes, R., Pregill, G. (Eds.), Phylogenetic Relationships of the LizardFamilies. Stanford University Press, Stanford (California), pp. 119–281.
olie, A., Smith, R., Smith, T., 2013. New amphisbaenian lizards from the earlyPaleogene of Europe and their implications for the early evolution of modernamphisbaenians. Geologica Belgica 16, 227–235.
ans, C., 1978. The characteristics and affinities of the Amphisbaenia. Transactionsof the Zoological Society of London 34, 347–416.
ans, C., Montero, R., 2008. An atlas of amphisbaenian skull anatomy. In: Gans, C.,Gaunt, A.S., Adler, K. (Eds.), Biology of the Reptilia 21, Morphology I. Society forthe Study of Amphibians and Reptiles, Ithaca, pp. 621–738.
echt, M., Hoffstetter, R., 1962. Note préliminaire sur les amphibiens et les squa-mates du Landénien supérieur et du Tongrien de Belgique. Bulletin de l’InstitutRoyal des Sciences Naturelles de Belgique 38, 1–30.
errel, A., Aerts, P., Fret, J., De Vree, F., 1999. Morphology of the feeding sys-tem in agamid lizards: ecological correlates. The Anatomical Record 254,496–507.
offstetter, R., Gasc, J.P., 1969. Vertebrae and ribs of modern reptiles. In: Gans, C.(Ed.), Biology of the Reptilia 1, Morphology A. Academic Press, London and NewYork, pp. 201–310.
earney, M., 2003. Systematics of the Amphisbaenia (Lepidosauria: Squamata) basedon morphological evidence from recent and fossil forms. Herpetological Mono-graphs 17, 1–74.
lembara, J., 2008. A new anguimorph lizard from the Lower Miocene of northwestBohemia, Czech Republic. Palaeontology 51, 81–94.
lembara, J., Green, B., 2010. Anguimorph lizards (Squamata, Anguimorpha) from
the Middle and late Eocene of the Hampshire Basin of southern England. Journalof Systematic Palaeontology 8, 97–129.lembara, J., Böhme, M., Rummel, M., 2010. Revision of the anguine lizard Pseudopuslaurillardi (Squamata, Anguidae) from the Miocene of Europe, with commentson paleoecology. Journal of Paleontology 84, 159–196.
ontologie 101 (2015) 29–41
Klembara, J., Hain, M., Dobiasová, K., 2014. Comparative anatomy of the lowerjaw and dentition of Pseudopus apodus and the interrelationships of speciesof subfamily Anguinae (Anguimorpha, Anguidae). The Anatomical Record 297,516–544.
LaDuke, T.C., 1991. The fossil snakes of Pit 91, Rancho La Brea, California. Contrib-utions in Science, Natural History Museum of Los Angeles County 424, 2–28.
Laloy, F., Rage, J.C., Evans, S.E., Boistel, R., Lenoir, N., Laurin, M., 2013. A re-interpretation of the Eocene anuran Thaumastosaurus based on microCTexamination of a ‘mummified’ specimen. Plos One 8, e74874 (1–11).
Legendre, S., 1987. Mammalian reference levels MP 17–20. In: Schmidt-Kittler, N.(Ed.), International Symposium on Mammalian biostratigraphy and paleoecol-ogy of the European Paleogene. Münchner Geowissenschaftliche AbhandlungenA 10, 28.
Legendre, S., Sigé, B., Astruc, J.G., de Bonis, L., Crochet, J.Y., Denys, C., Godinot, M.,Hartenberger, J.L., Lévêque, F., Marandat, B., Mourer-Chauviré, C., Rage, J.C.,Rémy, J.A., Sudre, J., Vianey-Liaud, M., 1997. Les phosphorites du Quercy : 30 ansde recherche. Bilan et perspectives. Geobios Mémoire Spécial 20, 331–345.
Macey, J.R., Schulte, J.A., Larson, A., Tuniyev, B.S., Orlov, N., Papenfuss, T.J., 1999.Molecular phylogenetics, tRNA evolution and historical biogeography in anguidlizards and related taxonomic families. Molecular Phylogenetics and Evolution12, 250–272.
Montero, R., Gans, C., 1999. The head skeleton of Amphisbaena alba Linnaeus. Annalsof Carnegie Museum 68, 15–80.
Moody, S.M., 1980. Phylogenetic and Historical Biogeographical Relationships of theGenera in the Family Agamidae (Reptilia, Lacertilia). PhD University of Michigan,Ann Arbor, 373 p. (unpublished).
Moody, S.M., Rocek, Z., 1980. Chamaeleo caroliquarti (Chamaeleonidae: Sauria): anew species from the lower Miocene of central Europe. Vestník Ústrednihoústavu geologického 55, 85–92.
Mourer-Chauviré, C., 2014. Valbro : un nouveau site à vertébrés de l’Oligocèneinférieur (MP 22) de France (Quercy). II - Oiseaux (Aves). Annales de Paléon-tologie 100, 47–50.
Peigné, S., Vianey-Liaud, M., Pélissié, T., Sigé, B., 2014. Valbro : un nouveau siteà vertébrés de l’Oligocène inférieur (MP 22) de France (Quercy). I - Con-texte géologique ; Mammalia : Rodentia, Hyaenodontida, Carnivora. Annales dePaléontologie 100, 1–45.
Pyron, R.A., Burbrink, F.T., Wiens, J.J., 2013. A phylogeny and revised classificationof Squamata, including 4161 species of lizards and snakes. BMC EvolutionaryBiology 13, 1–53.
Rage, J.C., 1974. Les serpents des Phosphorites du Quercy. Palaeovertebrata 6,274–303.
Rage, J.C., 1978. La poche à phosphate de Ste-Néboule (Lot) et sa faune de Vertébrésdu Ludien supérieur. 5. Squamates. Palaeovertebrata 8, 201–215.
Rage, J.C., 1984. Serpentes. Handbuch der Paläoherpetologie, 11. Gustav FischerVerlag, Stuttgart, 80 p.
Rage, J.C., 1988a. Le gisement du Bretou (phosphorites du Quercy, Tarn-et-Garonne,France) et sa faune de Vertébrés de l’Eocène supérieur. I. Amphibiens et Reptiles.Palaeontographica A 205, 3–27.
Rage, J.C., 1988b. The oldest known colubrid snakes. The state of the art. In: Szyn-dlar, Z. (Ed.), Festschrift for Marian Młynarski. Acta Zoologica Cracoviensia 31,457–474.
Rage, J.C., 1999. Faunes à affinités sud-américaines dans le Paléogène d’Europe :état de la question. Allons-nous vers une nouvelle interprétation? Bulletin de laSociété géologique de France 170, 951–954.
Rage, J.C., 2006. The lower vertebrates from the Eocene and Oligocene of the Phos-phorites du Quercy (France): an overview. In: Pélissié, T., Sigé, B. (Eds.), 30millions d’années de biodiversité dynamique dans le paléokarst du Quercy.Strata 13, 161–173.
Rage, J.C., 2012. Amphibians and squamates in the Eocene of Europe: what do theytell us? In: Lehmann, T., Schaal, S.F.K. (Eds.), Messel and the terrestrial Eocene.Palaeobiodiversity and Palaeoenvironments 92, 445–457.
Rage, J.C., Augé, M., 2010. Squamate reptiles from the middle Eocene ofLissieu (France). A landmark in the middle Eocene of Europe. Geobios 43,243–268.
Rocek, Z., 1994. Taxonomy and distribution of Tertiary discoglossids (Anura) of thegenus Latonia v. Meyer, 1843. Geobios 27, 717–751.
Rocek, Z., 2013. Mesozoic and Tertiary Anura of Laurasia. In: Gardner, J.D., Nydam,R.L. (Eds.), Mesozoic and Cenozoic Lissamphibian and Squamate assemblages ofLaurasia. Palaeobiodiversity and Palaeoenvironments 93, 397–439.
Rocek, Z., Wuttke, M., Gardner, J.D., Bhullar, B.-A.S., 2014. The Euro-Americangenus Eopelobates, and a re-definition of the family Pelobatidae (Amphibia,Anura). Palaeobiodiversity and Palaeoenvironments, http://dx.doi.org/10.1007/s12549-0169-5.
Rook, L., Kotsakis, T., 2005. Il Miocene: le terre emerse del Miocene. In: BonfiglioL. (Ed.), Paleontologia dei vertebrati in Italia: evoluzione biologica, significatoambientale e paleogeografia. Memorie del Museo Civico di Storia Naturale diVerona, 2 serie, Sezione Scienze della Terra 6, 155–156.
Sanchiz, B., 1998. Salientia. Handbuch der Paläoherpetologie, 4. F. Pfeil, Munich (xii+ 275 p.).
Schleich, H.H., 1985. Neue Reptilienfunde aus dem Tertiär Deutschlands. 3. Erstnach-weis von Doppelschleichen (Blanus antiquus sp. nov.) aus dem Mittelmiozän
Süddeutschlands. Münchner Geowissenschaftliche Abhandlungen A 4,67–149.Schleich, H.H., 1988. Neue Reptilienfunde aus dem Tertiär Deutschlands. 8.Palaeoblanus tobieni n. gen., n. sp. - neue Doppelschleichen aus dem TertiärDeutschlands. Paläontologische Zeitschrift 62, 95–105.

de Palé
S
S
S
S
S
S
S
von Zittel, K.A., 1890. Handbuch der Palaeontologie. 1. Palaeozoologie. III. Vertebrata
J.-C. Rage, M. Augé / Annales
chleich, H.H., Kästle, W., Kabish, K., 1996. Amphibians and Reptiles of North Africa.Koeltz Scientific Books, Koenigstein (630 p.).
igé, B., Aguilar, J.P., Marandat, B., Astruc, J.G., 1991. Extension au Miocène inférieurdes remplissages phosphatés du Quercy. La faune de vertébrés de Crémat (Lot,France). Geobios 24, 497–502.
igé, B., Hugueney, M., Crochet, J.Y., Legendre, S., Mourer-Chauviré, C., Rage, J.C.,Simon-Coinc on, R., 1998. Baraval, nouvelle faune de l’Oligocène inférieur (MP22) des phosphorites du Quercy. Apport à la signification chronologique desremplissages karstiques. Bulletin de la Société d’Histoire naturelle de Toulouse134, 85–90.
mith, K.T., 2009. A new lizard assemblage from the earliest Eocene (zone Wa0) ofthe Bighorn Basin, Wyoming, USA: biogeography during the warmest intervalof the Cenozoic. Journal of Systematic Palaeontology 7, 299–358.
zyndlar, Z., 1994. Oligocene snakes of Southern Germany. Journal of VertebratePaleontology 14, 24–37.
zyndlar, Z., 2012. Early Oligocene to Pliocene Colubridae of Europe: a review. In:Steyer, J.S., Buffetaut, E. (Eds.), Dr. Jean-Claude Rage – an appreciation. Bulletinde la Société géologique de France 183, 661–681.
zyndlar, Z., Alférez, F., 2005. Iberian snake fauna of the early/middle Miocene tran-sition. Revista Espanola de Herpetología 19, 57–70.
ontologie 101 (2015) 29–41 41
Szyndlar, Z., Rage, J.C., 2003. Non-erycine Booidea from the Oligocene and Mioceneof Europe. Institute of Systematics and Evolution of Animals, Polska AkademiaNauk, Cracow, 109 p.
Szyndlar, Z., Schleich, H.H., 1993. Description of Miocene snakes from Peters-buch 2 with comments on the lower and middle Miocene ophidianfaunas of southern Germany. Stuttgarter Beiträge zur Naturkunde B 192,1–47.
Venczel, M., Sanchiz, B., 2006. Lower Miocene amphibians and reptiles from Oschiri(Sardinia, Italy). Hantkeniana 5, 72–75.
Venczel, M., Stiuca, E., 2008. Late middle Miocene amphibians and squamate reptilesfrom Taut, Romania. Geodiversitas 30, 731–763.
Vianey-Liaud, M., Comte, B., Marandat, B., Peigné, S., Rage, J.C., Sudre, J., in press.A new early Late Oligocene (MP 26) continental vertebrate fauna from Saint-Privat-des-Vieux (Alès Basin, Gard, Southern France). Geodiversitas.
(Pisces, Amphibia, Reptilia, Aves). R. Oldenbourg, Munich, 900 p.Wiens, J.J., Brandley, M.C., Reeder, T.W., 2006. Why does a trait evolve multiple times
within a clade? Repeated evolution of snakelike body form in squamate reptiles.Evolution 60, 123–143.




















