
Untitled - Eco Astronomy Sri Lanka
129
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Untitled - Eco Astronomy Sri Lanka


WILDLANKA Vol.9, No.2, pp. 173 - 300, 2021.
Copyright 2021 Department of Wildlife Conservation, Sri Lanka.
FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN
ARAVINDA RAVIBHANU SUMANARATHNA1*, KAMAL ABEWARDHANA2,
JINADASA KATUPOTHA3 and MAJDAAOUITITEN4
1,2,3,4 Department of Research & Innovation -
South Asian Astrobiology & Earth Sciences Research Unit of Eco Astronomy Sri Lanka. 1,2 United Nations Association of Sri Lanka: 39/1, Cyril Jansz, Mawatha, Panadura.
3 Department of Geography University of Sri Jayewardenepura,
Gangodawila, Nugegoda, 10250, Sri Lanka. 4 Beijing Forestry University School of Ecology and Nature Conservation,
Beijing, China.
ABSTRACT: The fossils are preserved remains of body parts or traces of ancient organisms. Sri Lanka
is a continental island that evolved via unique geological formations, including fossil remains. This island
represents many fossils belonging to three different geological periods, for instance: the Jurassic period,
Miocene period, and Pleistocene epoch. Most of the Pleistocene fossils were found in terrestrial deposits
(alluvial) from the Sabaragamuwa basin called Ratnapura fauna. Thus, our investigations focused on
documenting samples of fossils gathered, under the project called “The Paleo World of Sabaragamuwa
Basin - Sri Lanka” conducted by Eco Astronomy Inc (Sri Lanka). Considering the geological time scale,
we are looking for reporting samples that approximately belong to the Quaternary period. As we know,
the Quaternary period of the Earth’s geographic history includes two geologic epochs viz., which are:
the Pleistocene (2.58 Myr to 0.0012 Myr), and the Holocene (0.0012 Myr to Present). Both epochs have
changed and divided the fauna’s equilibrium and human’s cultural phases based on climate and sea-
level fluctuations that took place during these periods. Some of the sections in those epochs has occurred
during the last glacial maximum (LGM) and represent the mean sea level was much lower compared
with the present records. Therefore, the quaternary period shows the open accessibility to migration of
mammalian mega faunal species, that lived during the transition from the Pleistocene to the Holocene
epoch. Most probably, the terrestrial climate change has impacted them and caused the extinction of
those megafaunas. The gathered data details were synchronized via the technical aspect of sampling
photography, toy photography, and virtual reality for analyses and reconstruction purposes.
KEY WORDS: fossils, Ratnapura fauna, paleontology, Sri Lanka, reconstruction.
INTRODUCTION Fossils and Fossilization
Fossils (from Latin word fossils, literally
“digging”) are the preserved pieces of evidence
or traces of fauna, flora, and other organisms
that once lived on Earth. Fossilization is an
exceptionally rare phenomenon that occurred in
specific conditions. The process of fossilization
changes due to the tissue type and external
and environmental conditions. In other words,
fossilization is the process by which a plant or an
animal becomes a fossil, and for that to happen,
the body part needs to pass through crucial
steps of the fossilization process. Prothero
(2013) explained the permineralization,
casts and molds, authigenic mineralization,
replacement and recrystallization and adpression
(compression -impression) by Shute (1987).
As well, others discussed the soft tissue, cell
and molecular preservation (Embery, et al.,
2003; Schweitzer et al., 2014; Zylberberg &
Laurin, 2011), carbonization and coalification
(Prothero, 2013), Bioimmuration (Wilson et al.,
1994; Taylor, 1990).

174 WILDLANKA [Vol. 9 No. 2
Most organic components of the formerly-
living being tend to decompose relatively quickly
following the death of this organize. In order
for an organic organism to be fossilized (Fig.
01), the remains normally need to be covered
by sediment as soon as possible (Sumanarathna,
2020). However, there are exceptions to this,
such as if an organism becomes preserved due to
the amberization (Grimaldi, 2009), or because
the organism become completely frozen, or due
to desiccation, or if the organism was found in
an anoxic environment. Much farther, according
to some interesting scientific papers records,
we have noticed thought some analysis of the
fossils historical data, when it’s a concern of the
process of fossilization, a very small number
or amount of prehistoric organic being got
fossilized. In order for this phenomenon to take
place, conditions had to be exactly favorable.
Generally speaking, studies proved that only the
hard parts of an organism can become fossilized,
such as the teeth, the claws, the shells, and the
bones. In fact, the soft body parts are usually
lost either due to the decomposition process,
except for some specific situation under some
special conditions.
There are many ways for an organism
to get preserved, this document will aim to
explain the general samples preserved, known
as well as fossilized organisms’, we are going
to mention the way in which most fossils used
to be formed and created. Usually, fossils
occur in sedimentary rock, and it is very rare
to encounter such phenomenon in metamorphic
rock (Bucher, 1953; Franz et al., 1991; Hill,
1985; Laborda-López et al., 2015; Wright and
Ghent, 1973). The best scenario for an excellent
fossilization would be under some specific
condition in which an organism get buried at the
bottom of a lake, where it gets then covered by
a lot of sediment.
In this type of environment, the organism
is protected from other animals and natural
elements that would cause the body’s
fragmentation as it gets breakdown. It is crucial
that the body must be in an environmental
condition, that allow the rapid burial processing
mechanisms. The areas in which there is a high
rate of sediment deposition are ideal because
of the presence of minerals that will lead to
the increase of the pressure. Additionally with
some extreme conditions the records have
proved that the ammonites and meta fossils
are recorded which assemblages developed by
metasomatic exchange of silica, sourced from
the matrix with CaO sourced from the calcite
shells. As well, we need to highlight that the
metamorphic transformations, in the particular
reaction of calcite with silica-bearing fluids
attend to form wollastonite and we can notice
the transformation of pyrite to pyrrhotite (Shaw,
2019).
There are other ways of preservation, such
as the petrification phenomenon. In fact, the
petrified wood happens under extreme incident
conditions and parameters. For instance: if for
long time ago, dead logs were washed into a
river and buried in the sand. Water with alkaline
and dissolved silica went down through the
sediments, and get in contact with the logs.
The logs decayed, will release carbon dioxide,
which dissolved in the water and formed
carbonic acid.
The alkaline water will then be neutralized,
and the silica evaporate out of the solution.
Very slowly, the cellulose of the wood will be
replaced, molecule by molecule, by the silica
instead of the cellulose. Eventually, the wood
gets replaced in perfect detailed process by
minerals. If other minerals existed during this
situation, also, the wood could be stained with
pretty colors (Dávid, 2011).
Organisms can also be preserved by
carbonization. If a leaf falls into a stagnant,
oxygen-poor swamp, it may not decay. If it
gets covered in silt and subjected to heat and
pressure, most of the leaf’s organic material is
released as methane, water, and carbon dioxide.
The remainder is a thin film of carbon, showing
the imprint of the leaf. Insects and fish can be
preserved in this way too. (Sumanarathna,
2021).
Paleontology and Paleontologist
After having a brief introduction of fossils
fossilization phenomenon and their different
preservations conditions, let’s see in depth
what is the board disciplines that we going to

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 175
FIGURE 01: Project Dynamics of Himachal Pradesh: Upgrading Prometheus of Paleo Himachal
Ammonites fossils from Ladakh high altitude mountains © Aravinda Ravibhanu 2019 (Sumanarathna,
2020).
apply for study fossils. Conducting this type of
research, it is called Paleontology, it is indeed
concerning of conducting scientific studies of
the ancient life form encountered on Earth as
based on fossils formation. Within evolutionary
biology field of researches and studies, the
time parameter has long been considered the
province of the paleontologist. Strictly speaking,
this is not completely true. Time itself could be
considered as it is just as elusive parameter for
the paleontologist, as it is as well for anybody
else.
Paleontologists do not trafic in time
themselves, but rather in events arrayed
in time, in processes occurring over time,
and in techniques for the measurement and
representation of time. That said, paleontology
does have something distinctive to contribute
to our understanding of the deep past, namely
its ability to draw justifiable inferences about
the temporal sequence of events in the history
of life. Although, absolute dating based on
radioactive decay of atomic nuclei and other
physical methods have made it possible to get
better and great estimates of the ages of rocks
and fossils, paleontology throughout its history
has reconstructed the history of life without
explicit consideration of duration (Fig.02),
but rather by using the relative positions and
presumed temporal sequence of fossil bearing
strata, and the biological affinities among
the fossil remains those strata contain.
Despite refinements in absolute dating,
and the introduction of molecular methods, the
fossil and event-based relative timescale have
remained the backbone of the paleontological
conception of time. Time in paleontology
presents a menu of philosophical issues: the
phenomenology of time, the nature of relative
and absolute time, and the epistemic issues
arising out of attempts to reconstruct the past
from its present-day traces. We will begin by
contemplating the phenomenology of time in
the work of the practicing paleontologist. From

176 WILDLANKA [Vol. 9 No. 2
FIGURE 02: 145 million years old, world famous - berlin Archaeopteryx second specimen (replica)
at LKCNHM- Singapore. Image © Aravinda Ravibhanu 2017.
research in the psychology of time perception,
we know that our sense of time is event-based
(Resnick et al., 2012).
Thus, our general sense of time arises out
of the first-person perspective we have on our
lived experience, augmented by reflection on
the events of history. Yet these senses of time
are inadequate to paleontology. Consider that
the oldest known fossils on Earth, of anaerobic
bacteria, were found in Australian rocks dated
3.4 billion years before present (Wacey et al.,
2011). The timespan over which the events

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 177
studied by paleontologists have occurred is
so vast as to merit its own designation, Deep
Time (McPhee, 1981; Rickles’s Chap. 11 in
this volume). Deep Time dwarfs the timescales
humans are cognitively equipped to deal with. It
is immeasurably long compared to the entire span
of human history. So, the question is whether
anyone can truly grasp Deep Time. Efforts to
do so nearly always make use of appropriately
scaled spatial metaphors. It is a standard part
of the undergraduate training of geologists to
produce some spatially extended model (e.g., a
ruler, a roll of toilet paper, a piano keyboard)
such that the vastness of Deep Time can be
compared to the mere hair’s breadth (or less)
of human history that blemishes the end of it.
Spatializing time serves two roles. Existentially,
the exercise puts us in our place: as a species,
we are a relatively recent addition to Earth’s
biosphere. Cognitively and heuristically, space
often stands in for time itself in the scientific
work of the paleontologist.
GEOLOGY OF SRI LANKA
More than 90% of Sri Lanka’s surface lies
on Precambrian strata, some of it dating back
to two billion years. The granulite facies rocks
of the Highland complex (gneisses, sillimanite-
graphite gneisses, quartzite, marbles, and some
charnockites) make up most of the island, and
the amphibolite facies gneisses, magnetite,
granites, and granitic gneisses of the Vijayan
complex occur in the eastern and southeastern
lowlands. Jurassic sediments are present today
in very few areas near the western coast (Vanni
Complex), and Miocene limestones underlie the
northwestern part of the country and extend to
the south in a relatively narrow belt along the
west coast (Fig. 03A) (Cooray, 1984; Cooray
and Katupotha 1991; Katupotha and Dias 2001).
The metamorphic rock surface was created by
the transformation of ancient sediments under
intense heat and pressure during the mountain-
building processes. The theory of plate tectonics
suggests that these rocks and related rocks
forming most of south India were part of a
single southern landmass called Gondwanaland.
The beginning occurred about 200 million
years ago, the impressive forces within the
earth’s mantle began to separate the lands of
the Southern Hemisphere, and it have been
results in the locomotion of a crustal plate that
supported both India and Sri Lanka which have
caused this plate to move toward the northeast.
About 55 million years ago, the Indian plate
collided with the Asian landmass, that have
promoted the raising of the Himalayas in
northern India area, we need to mention as well
that this phenomenon is continuing to accrue
in an advanced level slowly but surely till the
present time (Fig. 03B).
However, this impressive movement and
relocation of the crustal plate didn’t result
in frequent nature catastrophes in this area,
therefore, Sri Lanka does not experience or
recorded earthquakes or major volcanic eruption
events because it rides located specifically on the
center of the plate. The island contains relatively
limited strata of sedimentation surrounding its
ancient uplands. Aside from recent deposits
along river valleys, only two small fragments
of Jurassic (140 to 190 Myr) sediment occur in
Puttalam District. In contrast, a more extensive
belt of Miocene (5 to 20 Myr) limestone is
found along the northwest coast, overlain in
many areas by Pleistocene deposit (Cooray and
Katupotha 1991; Katupotha and Dias 2001,
Katupotha 2019). The northwest coast is part
of the deep Cauvery (Kaveri) River Basin of
southeast India, which has been collecting
sediments from the highlands of India and Sri
Lanka since the breakup of Gondwanaland.
Quaternary Period in Sri Lanka
Quaternary is the era. which saw the
appearance of mankind. There is disagreement
over the duration, with some Scientists retaining
a short-time scale (600,000 years) while a
majority accepting the long time-scale of 1.8 to
2.0 million years. It comprises two epochs - the
Pleistocene and the Holocene. The Pleistocene
epoch in Sri Lanka has been subdivided based
on different types of fossils. These subdivisions
are useful in the study of paleomagnetic
changes, geological formations and Paleolithic
cultures in Sri Lanka. Radiometric dating of
fossil coral and shells collected from the western
and southern coastal zone suggested mid and
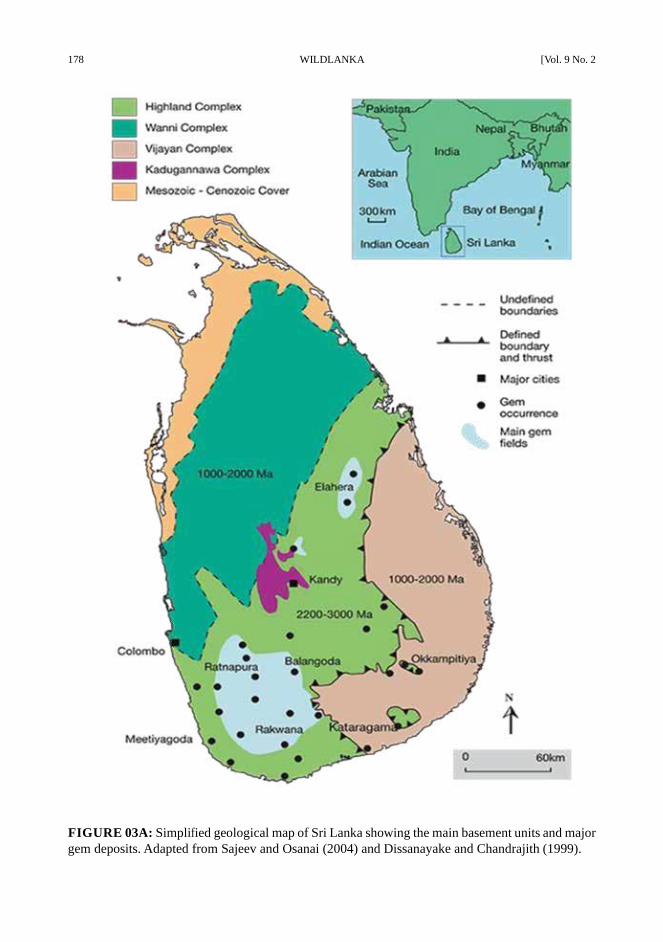
178 WILDLANKA [Vol. 9 No. 2
FIGURE 03A: Simplified geological map of Sri Lanka showing the main basement units and major
gem deposits. Adapted from Sajeev and Osanai (2004) and Dissanayake and Chandrajith (1999).

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 179
FIGURE 03B: (Left) New juxtaposition of Sri Lanka, the southern tip of India, and Madagascar
(encircled) in Gondwana. (Dissanayake and Chandrajith 1999). (Right) The position of Sri Lanka
in central Gondwana at 200 Ma (Emmel et al., 2012, Katupotha 2020).
late During the Holocene period rerecord have
documented three high sea-level episodes 6,240
yr B.P and 2,270 yr B.P (Fig. 03C).
FIGURE 03C: Eustatic 06 fluctuation in Sri
Lanka since last-Glacial Maximum
(Katupotha 1984 and 1995).
As well, during the early Holocene to LGM
and beyond the sea level found not reach the
Sabaragamuwa Basin, accordingly. However,
during the Holocene high sea level episode
the sea water did not entered to Kalu Ganga
Upper Lowlands, especially in Kuruwita and
Palmadulla basin area, but for the case of the area
covered by the fresh water marshes accordingly,
there is a need for further investigations on the
whole Quaternary period through different
disciplines in order to reveal the nature of
the paleogeography, palaeoclimatology and
paleoecology of Sri Lanka (Katupotha 1994).
The Quaternary period is a part of the
geologic time scale which is helpful to
paleontologists for the description of the timing
and relationships of events in geologic history
(Fig. 04). The Quaternary Period is popular for
the many cycles of glacial and geomorphological
growth and retreat, the extinction of many
species of mega fauna, and the spread of
humans (Table 01 & 02). This period is often
considered the “Age of Humans.” Homo erectus
appeared in Africa at the start of this period, and
as time marched on the hominid line evolved
and characterized of cranial capacity and higher
LP
LP 230,000
Third -
ABSOLUTE PROVISIONAL NORTH ALPS NORTH SRI
DATING NUMERICAL AMERICA EUROPE LANKA
ORDER
Present
H -
10,300
10,300 Main LATE
17,000 Interglacial WISCONSIN Late Wurm
17,000 Last Glaciation MAIN
30,000 WISCONSIN Main Wurm
30,000
50,000
50,000 WISCONSIN Early Wurm Weichselian Normal 6
75,000 Humid 6
67,000 - Last - Sangamanian Uznach Eemian Normal 5
128,000 Interglacial Humid 5
128,000 - Forth Glaciation ILLINOIAN RISS II WARTHE Humid 4
180,000 RISSI SAALE
180,000 Interglacial
Yarmouth Hotting Holstein Normal 3
230,000 Third Glaciation KANSAN MINDAL ELSTER Humid 3
300,000
300,000 Second Aftonian Normal 2
330,000 Interglacial
330,000 Second - NEBRASKAN GUNZ PRE-ELSTER Humid 2
470,000 Glaciation
470,000 Waalian Normal 1
538,000
538,000 First - PRE - DONAU II WEYBOURNE Humid 1
548,000 Glaciation NEBRASCAN
548,000 Tiglian
EP 585.000
585,000 DONAU I RED CRAG
600,000
e 600,000 Villa frannuchian
2,000,000
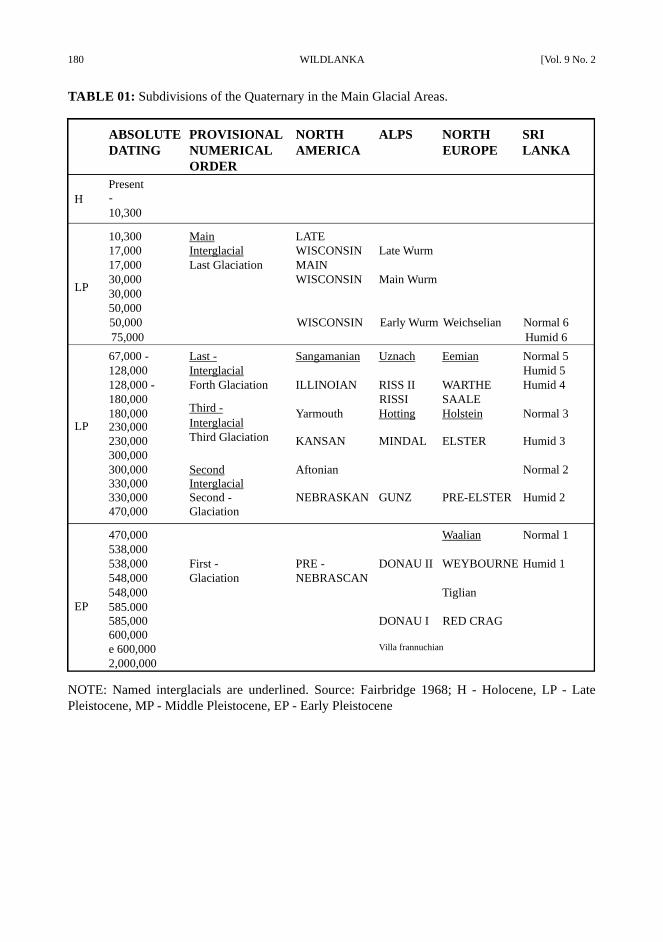
180 WILDLANKA [Vol. 9 No. 2
TABLE 01: Subdivisions of the Quaternary in the Main Glacial Areas.
NOTE: Named interglacials are underlined. Source: Fairbridge 1968; H - Holocene, LP - Late
Pleistocene, MP - Middle Pleistocene, EP - Early Pleistocene
0.01 m.y.
22.5 - 5
m.y.
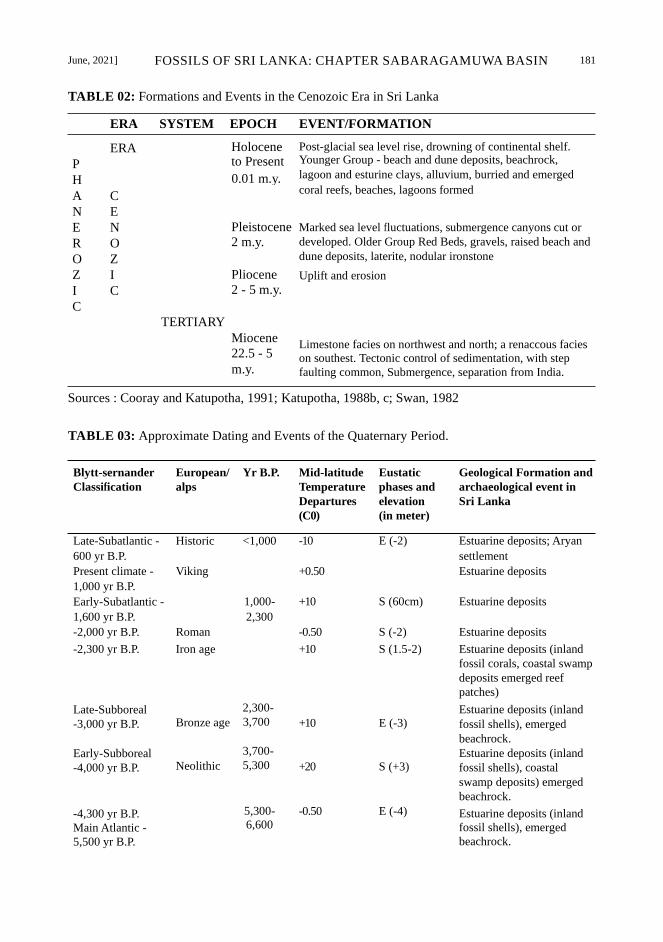
June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 181
TABLE 02: Formations and Events in the Cenozoic Era in Sri Lanka
ERA SYSTEM EPOCH EVENT/FORMATION
ERA
P
H
A C
N E
E N
R O
O Z
Z I
I C
C
Holocene Post-glacial sea level rise, drowning of continental shelf.
to Present Younger Group - beach and dune deposits, beachrock,
lagoon and esturine clays, alluvium, burried and emerged
coral reefs, beaches, lagoons formed
Pleistocene Marked sea level fluctuations, submergence canyons cut or
2 m.y. developed. Older Group Red Beds, gravels, raised beach and
dune deposits, laterite, nodular ironstone
Pliocene Uplift and erosion
2 - 5 m.y.
TERTIARY
Miocene Limestone facies on northwest and north; a renaccous facies
on southest. Tectonic control of sedimentation, with step
faulting common, Submergence, separation from India.
Sources : Cooray and Katupotha, 1991; Katupotha, 1988b, c; Swan, 1982
TABLE 03: Approximate Dating and Events of the Quaternary Period.
Blytt-sernander
Classification
Late-Subatlantic -
600 yr B.P.
Present climate -
1,000 yr B.P.
Early-Subatlantic -
1,600 yr B.P.
-2,000 yr B.P.
-2,300 yr B.P.
Late-Subboreal
-3,000 yr B.P.
Early-Subboreal
-4,000 yr B.P.
-4,300 yr B.P.
Main Atlantic -
5,500 yr B.P.
European/ Yr B.P.
alps
Historic <1,000
Viking
1,000-
2,300
Roman
Iron age
2,300-
Bronze age 3,700
3,700-
Neolithic 5,300
5,300-
6,600
Mid-latitude
Temperature
Departures
(C0)
-10
+0.50
+10
-0.50
+10
+10
+20
-0.50
Eustatic
phases and
elevation
(in meter)
E (-2)
S (60cm)
S (-2)
S (1.5-2)
E (-3)
S (+3)
E (-4)
Geological Formation and
archaeological event in
Sri Lanka
Estuarine deposits; Aryan
settlement
Estuarine deposits
Estuarine deposits
Estuarine deposits
Estuarine deposits (inland
fossil corals, coastal swamp
deposits emerged reef
patches)
Estuarine deposits (inland
fossil shells), emerged
beachrock.
Estuarine deposits (inland
fossil shells), coastal
swamp deposits) emerged
beachrock.
Estuarine deposits (inland
fossil shells), emerged
beachrock.
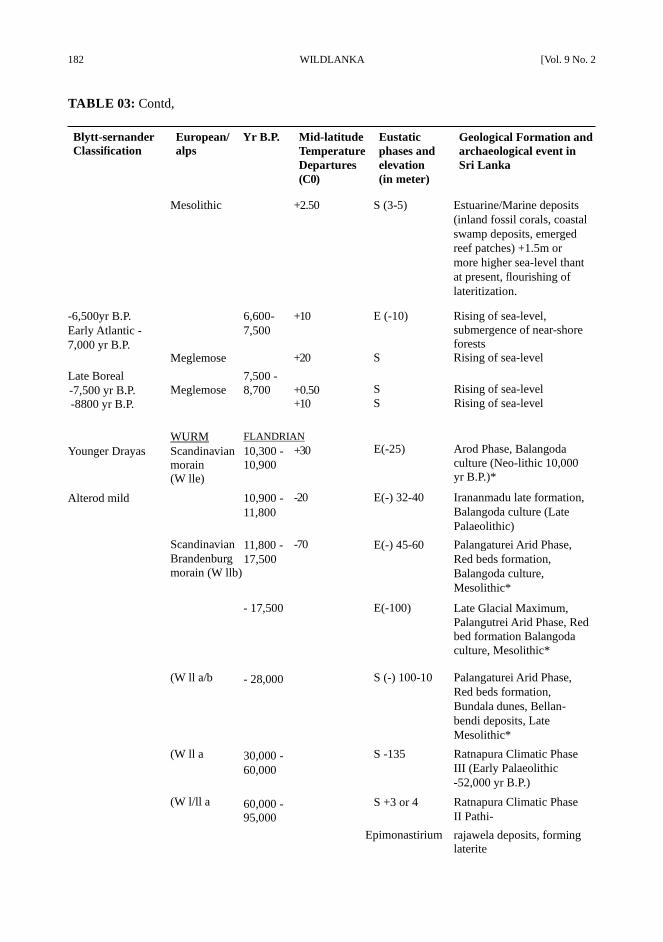
182 WILDLANKA [Vol. 9 No. 2
TABLE 03: Contd,
Blytt-sernander
Classification
European/ Yr B.P.
alps
Mid-latitude
Temperature
Departures
(C0)
Eustatic
phases and
elevation
(in meter)
Geological Formation and
archaeological event in
Sri Lanka
-6,500yr B.P.
Early Atlantic -
7,000 yr B.P.
Late Boreal
-7,500 yr B.P.
-8800 yr B.P.
Mesolithic
Meglemose
Meglemose
+2.50
6,600- +10
7,500
+20
7,500 -
8,700 +0.50 +10
S (3-5) Estuarine/Marine deposits
(inland fossil corals, coastal
swamp deposits, emerged
reef patches) +1.5m or
more higher sea-level thant
at present, flourishing of
lateritization.
E (-10) Rising of sea-level,
submergence of near-shore
forests
S Rising of sea-level
S Rising of sea-level
S Rising of sea-level
WURM FLANDRIAN
Younger Drayas Scandinavian 10,300 - +30
morain 10,900
(W lle)
E(-25) Arod Phase, Balangoda
culture (Neo-lithic 10,000
yr B.P.)*
Alterod mild 10,900 - -20
11,800
Scandinavian 11,800 - -70
Brandenburg 17,500 morain (W llb)
- 17,500
E(-) 32-40
E(-) 45-60
E(-100)
Irananmadu late formation,
Balangoda culture (Late
Palaeolithic)
Palangaturei Arid Phase,
Red beds formation,
Balangoda culture,
Mesolithic*
Late Glacial Maximum,
Palangutrei Arid Phase, Red
bed formation Balangoda
culture, Mesolithic*
(W ll a/b
(W ll a
(W l/ll a
- 28,000
30,000 -
60,000
60,000 -
95,000
S (-) 100-10
S -135
S +3 or 4
Epimonastirium
Palangaturei Arid Phase,
Red beds formation,
Bundala dunes, Bellan-
bendi deposits, Late
Mesolithic*
Ratnapura Climatic Phase
III (Early Palaeolithic
-52,000 yr B.P.)
Ratnapura Climatic Phase
II Pathi-
rajawela deposits, forming
laterite
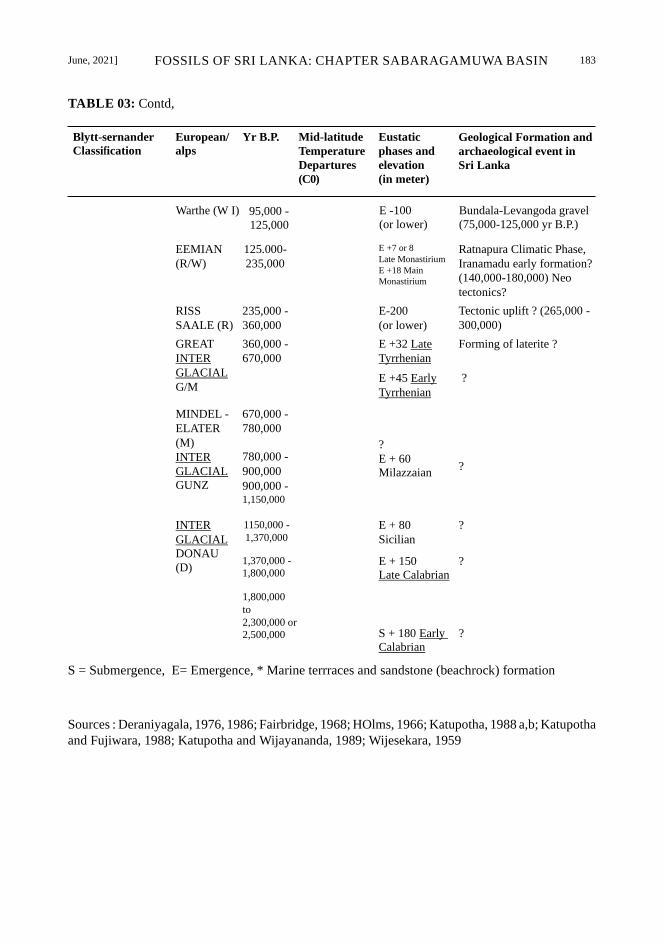
June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 183
TABLE 03: Contd,
Blytt-sernander
Classification
European/ Yr B.P.
alps
Mid-latitude
Temperature
Departures
(C0)
Eustatic
phases and
elevation
(in meter)
Geological Formation and
archaeological event in
Sri Lanka
Warthe (W I) 95,000 -
125,000
E -100
(or lower)
Bundala-Levangoda gravel
(75,000-125,000 yr B.P.)
EEMIAN
(R/W)
RISS
SAALE (R)
GREAT
INTER
GLACIAL
G/M
MINDEL -
ELATER
(M)
INTER
GLACIAL
GUNZ
125.000-
235,000
235,000 -
360,000
360,000 -
670,000
670,000 -
780,000
780,000 -
900,000
900,000 - 1,150,000
E +7 or 8
Late Monastirium
E +18 Main
Monastirium
E-200
(or lower)
E +32 Late
Tyrrhenian
E +45 Early
Tyrrhenian
?
E + 60
Milazzaian
Ratnapura Climatic Phase,
Iranamadu early formation?
(140,000-180,000) Neo
tectonics?
Tectonic uplift ? (265,000 -
300,000)
Forming of laterite ?
?
?
INTER
GLACIAL
DONAU
(D)
1150,000 -
1,370,000
1,370,000 -
1,800,000
E + 80 ?
Sicilian
E + 150 ?
Late Calabrian
1,800,000
to
2,300,000 or
2,500,000
S + 180 Early ?
Calabrian
S = Submergence, E= Emergence, * Marine terrraces and sandstone (beachrock) formation
Sources : Deraniyagala, 1976, 1986; Fairbridge, 1968; HOlms, 1966; Katupotha, 1988 a,b; Katupotha
and Fujiwara, 1988; Katupotha and Wijayananda, 1989; Wijesekara, 1959

184 WILDLANKA [Vol. 9 No. 2
FIGURE 04: Creative approach on displaying geologic time © Ray Troll
intelligence. The Quaternary is divided into two
epochs: the Pleistocene and the Holocene as
oldest to youngest period (Katupotha 2014).
The Pleistocene Epoch is defined as the time
period that began about 2.6 million years ago
and lasted until about 11,700 years ago. But,
Katupotha expected this period to have lasted
around 10,300 (1988a, 1994, and Table 03).
It is indicated that the most recent Ice Age
occurred then, as glaciers covered huge parts of
the planet Earth. It was followed by the current
stage, called the Holocene Epoch (11,700 to
present day).
Fossils of extinct Pleistocene fauna in Sri
Lanka are mainly found in alluvium deposits
of the Sabaragamuwa area. In additionally
Pleistocene fossils are rarely found in the
Lunugala, Adawatte gem mines near Moneragala
and Badulla, and in the Kalametiya area in
southern Sri Lanka. These Pleistocene faunal
fossils are commonly found in the vicinity of
the Rathnapura region of Sri Lanka. Hence its
name is “Ratnapura Fauna” (Deraniyagala,
1958). Ratnapura deposits have mixed up via
the redeposition mechanism and these deposits
represent specific time spots based on the most
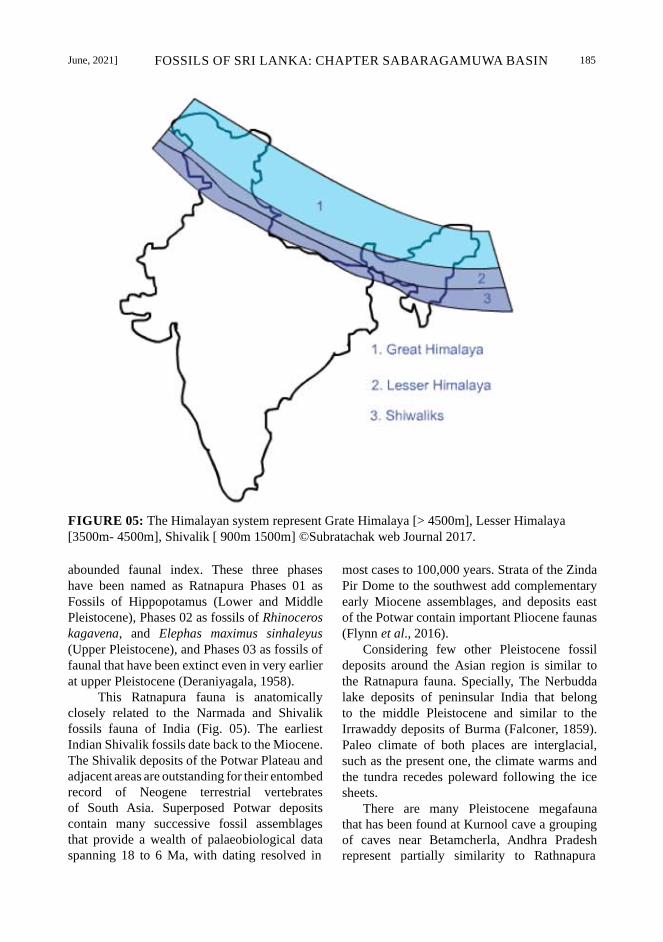
June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 185
FIGURE 05: The Himalayan system represent Grate Himalaya [> 4500m], Lesser Himalaya
[3500m- 4500m], Shivalik [ 900m 1500m] ©Subratachak web Journal 2017.
abounded faunal index. These three phases
have been named as Ratnapura Phases 01 as
Fossils of Hippopotamus (Lower and Middle
Pleistocene), Phases 02 as fossils of Rhinoceros
kagavena, and Elephas maximus sinhaleyus
(Upper Pleistocene), and Phases 03 as fossils of
faunal that have been extinct even in very earlier
at upper Pleistocene (Deraniyagala, 1958).
This Ratnapura fauna is anatomically
closely related to the Narmada and Shivalik
fossils fauna of India (Fig. 05). The earliest
Indian Shivalik fossils date back to the Miocene.
The Shivalik deposits of the Potwar Plateau and
adjacent areas are outstanding for their entombed
record of Neogene terrestrial vertebrates
of South Asia. Superposed Potwar deposits
contain many successive fossil assemblages
that provide a wealth of palaeobiological data
spanning 18 to 6 Ma, with dating resolved in
most cases to 100,000 years. Strata of the Zinda
Pir Dome to the southwest add complementary
early Miocene assemblages, and deposits east
of the Potwar contain important Pliocene faunas
(Flynn et al., 2016).
Considering few other Pleistocene fossil
deposits around the Asian region is similar to
the Ratnapura fauna. Specially, The Nerbudda
lake deposits of peninsular India that belong
to the middle Pleistocene and similar to the
Irrawaddy deposits of Burma (Falconer, 1859).
Paleo climate of both places are interglacial,
such as the present one, the climate warms and
the tundra recedes poleward following the ice
sheets.
There are many Pleistocene megafauna
that has been found at Kurnool cave a grouping
of caves near Betamcherla, Andhra Pradesh
represent partially similarity to Rathnapura

186 WILDLANKA [Vol. 9 No. 2
FIGURE 06: High Sea level fluctuation at Pleistocene and current environment view of
Sabaragamuwa basin from Gampaha, Ragama, Sri Lanka © Aravinda Ravibhanu.
fauna (Deraniyagala, 1958). Kurnool cave is
so significant because they contain teeth and
artifacts of early man. Systematic excavations
revealed a rich fossil assemblage that has
a bearing on past climate, environment,
ecology, and migratory patterns of some of
the mammalian groups. The existence of thick
caves sediments and ideally situated rock
shelters, which are three to four meters above
ground level, suggest that detailed excavation
is likely to yield fossil remains of early man
(Prasad, 1996).
Megafauna bonebed of the Trinil site (Java,
Indonesia) belonged to the Lower/Middle
Pleistocene (Thomas et al., 2014). Considering
the fossils in Palestine called Bethlehem
fauna partially similar to Shivalik deposits.
(Deraniyagala, 1958).
The Ratnapura fossils represent no trace
of Lateritization which occurs under humid
tropical climate conditions. This is a prolonged
process of chemical weathering of rocks leading

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 187
FIGURE 07A: Lithological column of an alluvial gem deposit in Rathnapura gravel © Aravinda
Ravibhanu 2019.

188 WILDLANKA [Vol. 9 No. 2
FIGURE 07A: [Top] Vertical mining extraction gem pit. [Bottom] Open-pit sapphire mine near
Balangoda, a large one by Sri Lankan standards, had partially filled with water from rains the week
before. Image© Andrew Lucas.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 189
to the formation of soil. It produces a variation
in the thickness, grade, chemistry and ore
mineralogy of the rock creating soil particles.
The arrangement of the same lateralized beds
in the Ratnapura series suggested few climatic
fluctuations like non laterite formation climate
present in Middle and Upper Pleistocene
(Fig. 06). However, redeposited fossils are
derived from beds of non-laterite and occur in
the upper-most or Holo -Pleistocene deposit
(Deraniyagala, 1958).
Means around the lower Pleistocene, there
are patterns to indicate red residual soil formed by
the leaching of silica and by the enrichment with
aluminum and iron oxides. Also, redeposition
has defiantly mixed up two or more horizons
of different ages which are assignable to the
Middle, Upper and Holo-Pleistocene. A marine
and a non-marine assignable that will form a
tie in have yet to be discovered (Deraniyagala,
1958). Considering the zooarchaeological
pieces of evidence around the last three decades
in Sri Lanka, revealed facts of marine fossils
belong to the lower Pleistocene.
Most of Ratnapura Fauna occurs in
association with a gem stone, dominantly rubies
and sapphires. Over 90% of Sri Lanka’s gem
mining is from secondary placer deposits that
can be classified as sedimentary gem deposits
of residual, eluvial and especially alluvial types
like Rathnapura beds (Fig. 7A & 7B). Primary
or in-situ gem occurrences are located mainly
in contact-metamorphic zones comprising of
skarn and calcium-rich rocks. Gem minerals
that are frequently found in pegmatites include
corundum, zircon, beryl, quartz varieties,
feldspar and chrysoberyl and possible to see
gem gravels at Getaheththa (Disanayake and
Rupasingha 1995).
But rubies and sapphires associated with
Ratnapura faunas are free of their pegmatite’s
matrix. This suggests that the disappearance of
FIGURE 08: Batadomba-lena pre historic cave, view from outside. Image © Aravinda Ravibhanu
2013.

190 WILDLANKA [Vol. 9 No. 2
the pegmatites from Sri Lankan Gem stones is
a comparatively recent occurrence while their
deposition apparently occurred together with
a part of Rathnapura Fauna. Uranium analysis
of Hippopotamus and Elephas maximus molar
indicated that redeposition had occurred, mixing
up beds of different ages. Considering, whole
fauna community belongs to Ratnapura fauna
suggests that it inhabited savannah country
possessing extensive bodies of water, while the
mountains probably supported forests that were
no different to present conditions (Roberts et al.,
2015; Sumanarathna et al., 2016; Wedage et al.,
2019).
When Ratnapura fauna dies around the
holocoen and Pleistocene period, most of the
body parts are typically decay completely.
But sometimes, when the conditions are just
right for the fossilization process, preserved
as fossils. Especially animal dies in a watery
environment and is buried in mud and silt like
Ratnapura beds. Soft tissues quickly decompose
leaving the hard bones or shells behind. Water
seeps into the remains, and minerals dissolved
in the water seep into the spaces within the
remains, where they form crystals. Also, the
minerals in groundwater replace the minerals
that make up the bodily remains after the water
completely dissolves the original hard parts
of the organism. A similar incident was easily
identified via fossils flora in the Sabaragamuwa
basin. Considering the Prehistoric Caves in
the Sabaragamuwa basin and the area of a
range of mountains around it, representing one
of the most important fossils belongs to the
Pleistocene and Holocene fauna in Sri Lanka.
Of these, Fahien Cave: 48,000 yrBP, Kuruwita-
Batadomba-lena Cave: 37,000 yrBP, Kitulgala
Beli-lena Cave: 31,000yrBP, Alawala Potgul-
lena Cave: 14,000 yrBP and Bellanbendi
Pelassa, as an open prehistoric human habitat in
Udawalawe: 12,000 yrBP. (Deraniyagala, 1992;
Perera, 2010; Adikari, 1998 and Bandaranayake,
1994; Premathilake & Risberg,2003).
Batadomba-lena Cave (Fig. 08) is very
unique which is located in the front head of the
Sabaragamuwa basin in the southern slop of the
highland mountains range.
The first excavation of Batabomba-lena by
Deraniyagala (P.E.P) was conducted in 1938.
The finds included fragmentary human remains
and stone artifacts. The excavation was of 4 feet
and deraniyagala assigned the assemblage of
stone artifacts, in particular, the association of
microliths and human remains, to the Balangoda
phase. A preliminary examination was made
by Deraniyagala (S.U) in 1979 revealed a rich
occupational deposit. Thereafter, an excavation
of 2.6 m was conducted.
The stratigraphic sequence of seven main
occupational layers and 3 underlying strata
directly above bed rock has been described by
Daraniyagala in 1982. Layers 1 – 3 from the
top downwards were considered to have been
described in recent times by the extraction of
guano fertilizer for village paddy fields, and
leaving of the floor by monks who used to live in
the rock shelter. The occupational deposited in
layer 4 was described as a massive homogenous
stratum with brownish sand and silt containing
stone artifacts and faunal remains. Layers 5
and 6 were considered to be the site’s major
occupational layer 7 contains many stone
artifacts including geometric microliths which
were radiocarbon dated to approx. 30,000 years
BP.
Deraniyagala named the prehistoric man
of Sri Lanka as a new subspecies called Homo
sapiens balangodensis based on the frontal
bone found during the excavation of the Ravana
Falls prehistoric cave in 1945 (Deraniyagala,
1945). He has compiled several dental and
osteological variations of this Homo sapiens
balangodensis subspecies, comparing the skulls
and other skeletal fragments of the Sinhalese
and Tamils as well as the Vedda people living in
this country today (Deraniyagala, 1945).
In additionally other human remains from
caves at Kuruvita and Telulla (Deraniyagala,
1955) and finally from a kitchen midden cum
burial mound at Bellan Biindi Piiliissa which
yielded skeletal remains of ten individuals some
of which were fragmentary. All were in flexed
postures and in association with a wealth of stone,
bone and antler artifacts (Deraniyagala, 1957a
:1957b). This Homo sapiens balangodensis
mentioned by Deraniyagala is known by the

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 191
FIGURE 09: (Left) Chopper stone artefacts of Rathnapura culture from Gem pit.
FIGURE 10: (Right): Weathered pitted artefacts of Home sapiens Location-Kuruvita © SMKA.
Image ©Aravinda Ravibhanu 2020.
FIGURE 11: Balaingoda point from Balangoda
culture in Sri Lanka | Location -Kuruvita ©
SMKA. Image ©Aravinda Ravibhanu 2020.
FIGURE 12: Bone Point with attached
neck surface from Balangoda culture Sri
Lanka | Location -Kuruvita © SMKA. Image
©Aravinda Ravibhanu 2020.

192 WILDLANKA [Vol. 9 No. 2
FIGURE 13: A view of Sabaragamuwa basin from “Balana Gala”, technically view point at the
middle of vithanakanda trail to Batadomba-lena Cave. Image ©Aravinda Ravibhanu 2013.
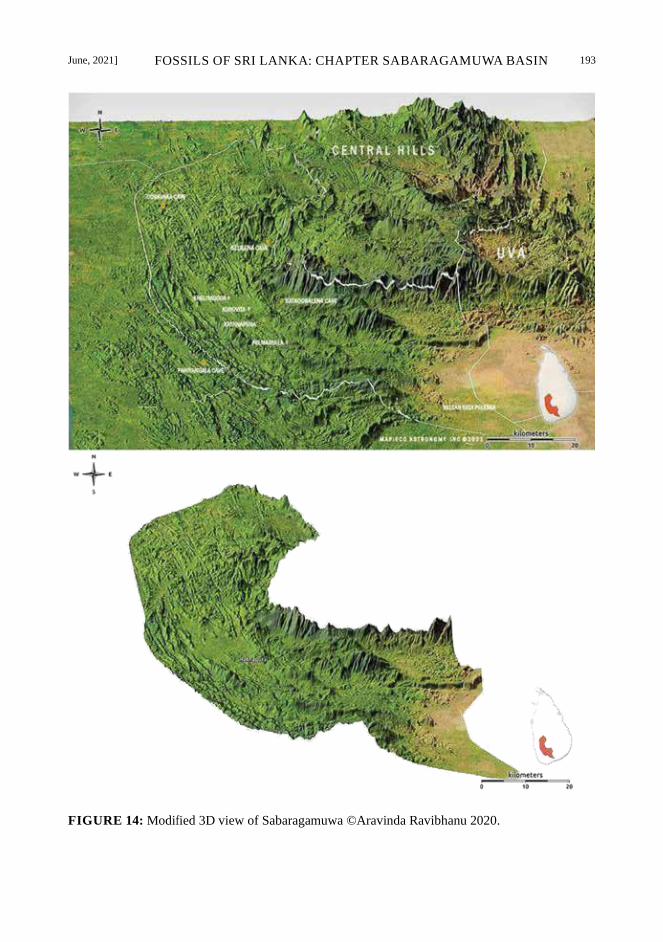
June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 193
FIGURE 14: Modified 3D view of Sabaragamuwa ©Aravinda Ravibhanu 2020.
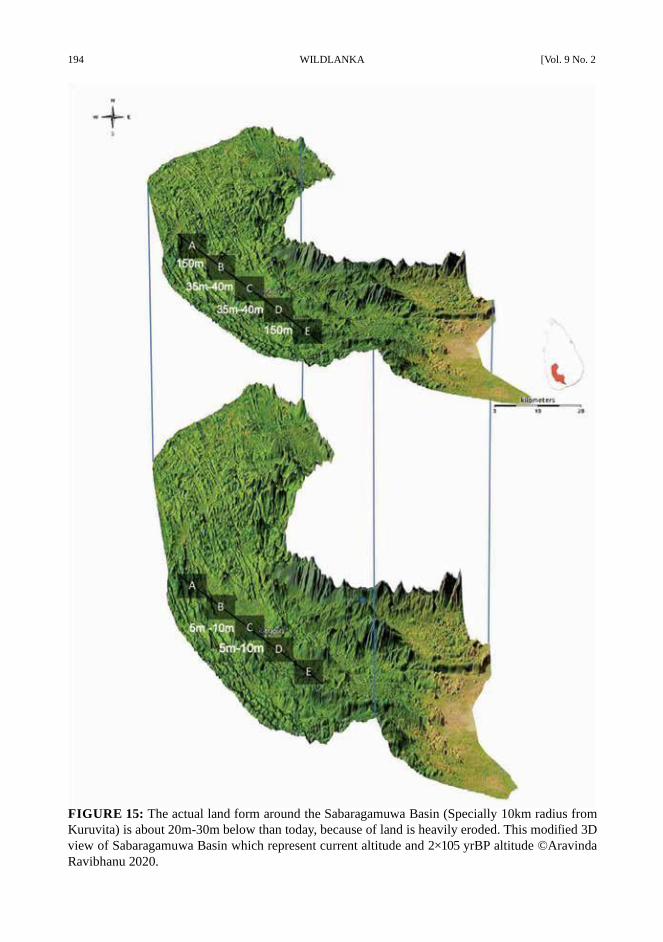
194 WILDLANKA [Vol. 9 No. 2
FIGURE 15: The actual land form around the Sabaragamuwa Basin (Specially 10km radius from
Kuruvita) is about 20m-30m below than today, because of land is heavily eroded. This modified 3D
view of Sabaragamuwa Basin which represent current altitude and 2×105 yrBP altitude ©Aravinda
Ravibhanu 2020.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 195
FIGURE 16: Paleo environment reconstruct of Sabaragamuwa basin via synchronizing geo
morphological factors of Shivalik and Ratnapura. ©Aravinda Ravibhanu 2020.

196 WILDLANKA [Vol. 9 No. 2
FIGURE 17: Elevation topographic map of Sabaragamuwa province © Sachith & Katupotha 2021.
simple common name of Balangoda Man.
According to the research conducted by the
American anthropologist, Professor Kenneth
Kennedy and Dr. Siran Deraniyagala reviled
this human being has been identified as a
modern human and not as another subspecies.
According to the prehistoric excavations
carried out by the Department of Archeology
and the Postgraduate Institute of Archeology
at the University of Kelaniya over the past four
decades, it is clear that the Balangoda human
Era dates back to approximately 40,000 years
ago and 3800 years today (Deraniyagala, 1992;
Perera, 2010).
Balangoda Man is a Homo sapiens which
is dominantly represented the Mesolithic
period via stone tool technology (Fig. 09 – 12).
Information that they lived in Sri Lanka has been
obtained from the prehistoric settlements as
rock shelter and open habitat mentioned above.
These humans made geometric microliths by
applicable quartz (Perera, 2010 and 2011).
The Balangoda man of the Middle Ages
made tools out of animal bones and they were
mostly pointed called bone points. These humans
provided their food by hunting and gathering,
as well as Asian palm civet species, Buffaloes,
grizzled giant squirrel, tortoises, hamburgers,
rabbits, frogs, as well as turtles, pythons, lizards,
wild boars, hawksbills, deer and sambar deer
was added to their diet includes aquatic snails
(Palodomus sp.) and tree snails are also part of
their diet which includes Canarium zeylanicum,
Artocarpus nobilis, Elaeocarpus sp.
It is noteworthy that a large number of fossils
belonging to the Pleistocene period were found
in the vicinity of the Sabaragamuwa Basin (Fig.
13). The Sabaragamuwa Basin extends from
Getahetta to Eheliyagoda, Kuruwita, Ratnapura,
Pelmadulla, Godakawela and Rakwana. Those
fossils can be found in the forest ecosystem as
well as inland wetland and agro-ecosystems in
the Sabaragamuwa Basin (Fig. 14 – 16).
Considering the current climatic conditions
of Ratnapura’s region is classified as tropical.
The rainfall in Ratnapura is significant, with

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 197
precipitation even during the driest month which
could be so similar to the highest interglacial
time in the late Pleistocene. The present climate
here is classified as Af by the Köppen-Geiger
system. The average annual temperature in
Ratnapura is 24.3 °C (75.7 °F). The rainfall here
is around 4460 mm (175.6 inches per year).
However, the present river terraces around
the Sabaragamuwa basin, do not provide as
much information as a drier region. Traces of
such terraces are scattered along the Kalani
river as patches of ancient gravelly alluvium
between Kaduwela and Hanvella, Malvana,
Palugama, Mapitigama (Deranigalagala 1958)
excepting few tributary valleys like Puugoda.
Vestigial beds of high-level gravels occur in the
vicinity of Sithawaka.
The old Ratnapura alluvial gravel occurs
belong the gem gravel in the Kuruvita area and
the old lacustrine alluvium occur at 20m to 27m
as bedded clays (Fig. 14 -15), silts, and gravel
(Deraniyagala,1958). However, currently, there
are few records similarity factors that can be
seen unit 50m to 60m includes clear deposition
of fossils which mix-up with the gem gravels
(Fig. 15).
Fossils of Sabaragamuwa Basin – Sri Lanka Family Felidae | Panthera leo sinhaleyus
(extinct)
Panthera leo (the lion) fossils laid upon
the gem field at a depth of 6.5m below the
surface from a gem pit about four miles away at
Pahala Vela, Galadande Mandiya, Gonapitiya,
Kuruwita near the Kuru Ganga. The holotype is
a third lower left carnassial in the Deraniyagala
collection at the British Museum (Deraniyagala,
1958). This race is restricted to Sri Lanka;
originally the lion appears to have inhabited
Sri Lanka and India and was possibly replaced
by the Bengal tiger that invaded India from the
Northeast.
The similarity between the African name
is “Simba” meaning Lion, and the Indian
equivalent Simha suggests that one is derived
from the other. The lack of lion fossils in Africa
suggests that the African is derived from the
Indian Panthera leo sinhaleyus also known as
the Sri Lankan Lion, which was a prehistoric
subspecies of lion, known to be endemic to
Sri Lanka (Fig. 18). It appears to have become
extinct prior to the arrival of the modern human
culture, which was estimated to be around
39,000 years ago.
This lion is only known from two teeth, found
in alluvial deposits at Kuruwita. Deraniyagala
cited fossils of three lion teeth found from the
island; first in 1936, second in 1947 and the third
in 1961. Manamendra-Arachchi et al. (2005)
described that Deraniyagala did not explain
explicitly how he diagnosed the holotype of
this subspecies as belonging to a lion, though he
justified its allocation to a distinct subspecies of
a lion by its being “narrower and more elongate”
than those of recent lions in the British Natural
History Museum collection.
The lion has been one of the most widespread
mammals, having enjoyed a Pleistocene range
that included Africa, Eurasia, North America
and tropical South America, while the fossil
record confirms that the species range in the
Indian subcontinent did extend south to the
21º N and east to 87º E (Pilgrim 1931; Dutta
1976), approximately a line joining Gujurat to
Bengal, but there is no evidence of the existence
of the lion in Asia east of Bengal or anywhere
in peninsular India and Sri Lanka, except for
Panthera leo sinhaleyus. Panthera leo fossilis,
also known as the Early Middle Pleistocene
European cave lion, is an extinct feline of the
Pleistocene epoch.
Family Felidae | Panthera tigris (extinct)
Panthera tigris is a member of the Felidae
family and the largest of four “big cats” in
the genus Panthera. The Panthera tigris tigris
(Bengal tiger) is a tiger subspecies native to
India, Bangladesh, Nepal and Bhutan. The
pattern of genetic variation in the Bengal tiger
corresponds to the premise that tigers arrived
in India approximately 12,000 years ago.
Kitchener and Dugmore (2000) considered
that the changing biogeographical range of
the Panthera tigris through the last glacial-
interglacial cycle, based on habitat associations
of modern tiger specimen records, and
environmental reconstructions from the LGM.
198 WILDLANKA [Vol. 9 No. 2
These cycles indicate that the numerous
glacial cycles that span the evolutionary history
of the tigers since their appearance in the fossil
record about 2 Myr ago and the oldest tiger
fossils (around 2 Myr old) are from northern
China and Java. The key issue is to determine
the extent to which ancestral populations of the
tiger were geographically isolated. However,
Pleistocene glacial and interglacial fluctuations
and other geological events probably caused
repeated geographic restrictions and expansions
of tigers (Hemmer,1987; Kitchener and
Dugmore, 2000) estimated the most recent
common ancestor for tiger mt DNA haplotypes
was 72,000–108,000 years ago, with a lower
and upper bound of 39,000 years and 157,000
years, respectively.
The recent history of tigers in the Indian
subcontinent is consistent with the lack of tiger
fossils from India prior to the late Pleistocene
and the absence of tigers from Sri Lanka, which
was separated from the subcontinent by rising
sea levels in the early Holocene. However, a
recent study of two independent fossil finds
from Sri Lanka, one dated to approximately
16,500 years ago, tentatively classifies them
as being a tiger (Manamendra-Arachchi et al.,
2005).
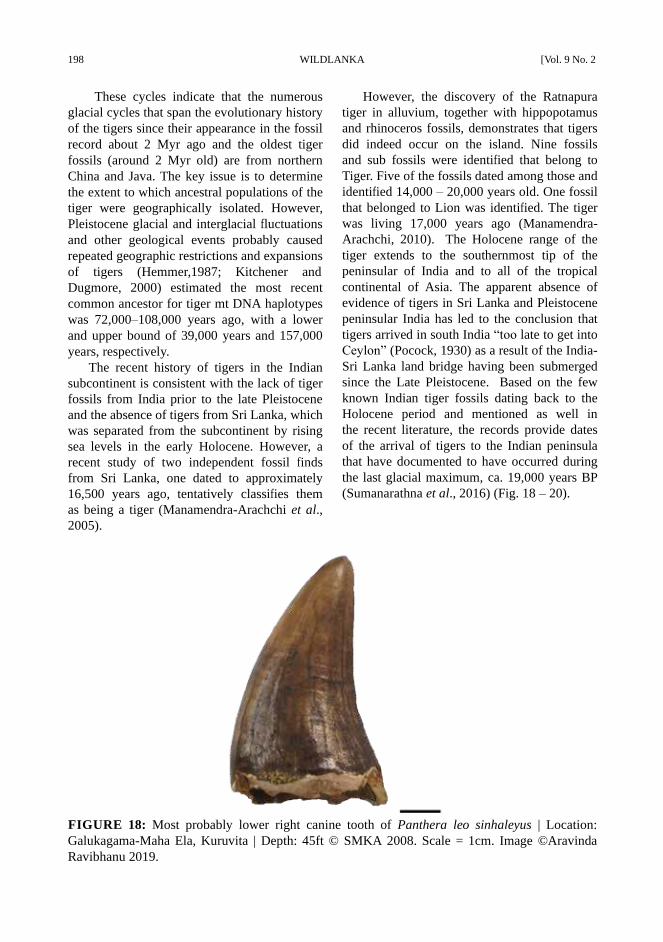
However, the discovery of the Ratnapura
tiger in alluvium, together with hippopotamus
and rhinoceros fossils, demonstrates that tigers
did indeed occur on the island. Nine fossils
and sub fossils were identified that belong to
Tiger. Five of the fossils dated among those and
identified 14,000 – 20,000 years old. One fossil
that belonged to Lion was identified. The tiger
was living 17,000 years ago (Manamendra-
Arachchi, 2010). The Holocene range of the
tiger extends to the southernmost tip of the
peninsular of India and to all of the tropical
continental of Asia. The apparent absence of
evidence of tigers in Sri Lanka and Pleistocene
peninsular India has led to the conclusion that
tigers arrived in south India “too late to get into
Ceylon” (Pocock, 1930) as a result of the India-
Sri Lanka land bridge having been submerged
since the Late Pleistocene. Based on the few
known Indian tiger fossils dating back to the
Holocene period and mentioned as well in
the recent literature, the records provide dates
of the arrival of tigers to the Indian peninsula
that have documented to have occurred during
the last glacial maximum, ca. 19,000 years BP
(Sumanarathna et al., 2016) (Fig. 18 – 20).
FIGURE 18: Most probably lower right canine tooth of Panthera leo sinhaleyus | Location:
Galukagama-Maha Ela, Kuruvita | Depth: 45ft © SMKA 2008. Scale = 1cm. Image ©Aravinda
Ravibhanu 2019.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 199
FIGURE 19: Panthera leo sinhaleyus & 3D reconstructed paleo environment of Sabaragamuwa
basin, upper image - partially wet condition and lower image - partially dry conditions ©Aravinda
Ravibhanu, Adeepa Nisal 2019.

200 WILDLANKA [Vol. 9 No. 2
FIGURE 19: Panthera tigris & Elephas maximus maximus via 3D reconstructed paleo environment
of Sabaragamuwa basin, |Location: Mere kele close to Boodhimaluwa, Eheliyagoda ©Aravinda
Ravibhanu 2019.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 201
Panthera tigris probably differentiated in
the early Pleistocene (1.806–2.588 Ma ago)
in northcentral and northeastern China. The
earliest forms averaged smaller than those of
later Pleistocene times. Thus, it seems that the
species has reached its maximum size in the
living subspecies P. tigris altaica. The early
Pleistocene species Panthera palaeosinensis,
from northern China, appears to represent
an early tiger or a form ancestral to the tiger
(Mazak, 1981).
Researches on fossil remains have been
conducted by many scientists, for example,
Mazak (1981) summarized the fossils records
in Sri Lanka. Accordingly, fossil remains,
definitely identified as Panthera tigris, are of
lower to upper Pleistocene age and originated
from the Altai caves in central Asia, eastern
and northern China, including Choukoutien
localities, Japan, Jana River in northern Siberia,
the Ljachov Island situated off the northern
coast of Siberia, and from Sumatra and Java.
In addition, several sub-recent tigers remain
were found in the Caucasus region, India, and
Borneo. It is not clear whether the material from
Borneo represents a member of the native late
Pleistocene fauna or a later introduction by
humans (there is no reliable evidence of tigers
on Borneo within historic times). Family Elephantidae | Elephas maximus
sinhaleyus (extinct)
The Asian elephant (Elephas maximus) is
one of the most seriously endangered species of
large mammals in the world. Given its enormous
size and body mass, it is also one of the few
species of terrestrial mega herbivores that still
exist. Its present geographical distribution
extends from the Indian subcontinent in the west
to Indo-China in the east across 13 countries
including islands such as Sri Lanka, Sumatra
and Borneo. The entire population in the wild
is estimated to be between 35,000 and 55,000.
Even optimistic figures indicate that there are
only about one tenth as many Asian as African
elephants (Hendavitharana et al., 1994).
Deraniyagala found one Fossil and explained
the extinct Sri Lankan elephant as subspecies
of Elephas maximus sinhaleyus (Deraniyagala,
1958; Sumanarathna et al., 2016) (Fig. 23-
46). Deraniyagala explained the tusks are
usually present, molars smaller and mandibular
spout wider than in forma typical. In addition,
he explained that there were three recently
extinct subspecies of Elephas maximusasurus
(Mesopotamia), Elephas maximus eondaicus
(Java) and Elephas maximus rubridens (China).
The extinct elephant species that were
living 100,000 years ago have been reported as
Hypselephas hysudricus sinhaleyus (Fig. 22)
by Deraniyagala (1914, 1937) and as Elephas
hysudricus by Manamendra-Arachchi (2008).
Elephas maximus sinhaleyus was secured in
1947 from a gem pit about four miles away at
Pahala Vela, Galadande Mandiya, Gonapitiiya,
Kuruwita near the Kuru Ganga. The fossils
were laid upon the gem field at a depth of 6.5m
below the surface, and yielded Elephas maximus
sinhaleyus (Deraniyagala, 1958). It frequently
occurs in association with hippopotamus fossils
from Gatahatta as far as Ratnapura, and with
rhinoceros from Gatahatta to Pelmadulla.
The origin of Elephas maximus remained
unknown until 1936, when its fossils were
discovered in Sri Lanka, and even as recently
as 1942 the general opinion was that nothing
was known of its origin except that it appeared
suddenly rather late in the age of man. It is true
that a few isolated fossils proboscidean molars
were assigned to an extinct Japanese race of
this elephant named Elephas maximus buski
(Deraniyagala, 1958), however, those belong
to Palaooloxodon namadicua naumanni and no
Elephas maximus fossils were found in Japan.
In various other countries also isolated and
often fragmentary teeth have been ascribed to
Elephas maximus, but in every instance these
have proved to be either those of the extinct
Palaeoloxodon namadicus (Fig. 21) or the
remains of some subspecies of Elephas maximus
that had become extinct during prehistoric
or historic times. Since its earliest remains
occur only in Sri Lanka, Elephas maximus
apparently evolved from some Plio-Quaternary
proboscidean, which had become isolated here
upon the Island’s separation from Asia. During a
Pleistocene reconnection with India, the Ceylon
animal had invaded the mainland and wandered
202 WILDLANKA [Vol. 9 No. 2
FIGURE 21: Palaeoloxodon namadicus sinhaleyus is one of extinct elephant species in Sri Lanka.
Illustrated by: Deraniyagala (1958). Image ©Aravinda Ravibhanu 2016.
northwards until it encountered the Himalayan
mass if, whereupon it had spread along its base
eastwards as far as Wallace’s line (Wallace’s
Line is a boundary that separates the eco-
transitional zone between Asia and Australia).
West of the line is found organisms related to
Asiatic species; to the east, a mixture of species
of Asian and Australian origin is present, and
westwards until checked by the Mediterranean
Sea and the deserts of Arabia and North Africa.
Over this vast expanse in a belt stretching from
40 degrees north to 10 degrees south, land
subsidence, changing river systems, deepening
river gorges and expanding deserts, assisted
the mountain ranges as barriers, and resulted
in the evolution of twelve (12) subspecies
(Deraniyagala,1958).
June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 203
FIGURE 22: Hypselephas hysudricus sinhaleyus is one of extinct elephant species in Sri Lanka.
Illustrated by Deraniyagala (1958). Image ©Aravinda Ravibhanu 2016.

204 WILDLANKA [Vol. 9 No. 2
FIGURE 23: Palaeoloxodon namadicus sinhaleyus and Crocodylus sinhaleyus 3D reconstructed
paleo environment of Sabaragamuwa basin. | Location: Kahengama- Ovita kubura, Kuruwita.
©Aravinda Ravibhanu 2019.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 205
FIGURE 24: A piece of elephant tooth [Elephas maximus maximus] [vertical braking down via
line of enamel apatite] | Location: Kuruvita | Depth: 50ft © SMKA 2008 Scale = 1cm. Image:
©Aravinda Ravibhanu 2019.

206 WILDLANKA [Vol. 9 No. 2
FIGURE 25: A eroded piece of elephant molar tooth [Elephas maximus maximus] | Location:
Kuruvita | Depth: 48ft © SMKA 2010. Scale = 1cm.] Image: ©Aravinda Ravibhanu 2019.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 207
FIGURE 26: Eroded and piece of elephant tusk [Elephas maximus maximus] | Location: Kuruvita
| Depth: 55ft. © SMKA 2011. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.
208 WILDLANKA [Vol. 9 No. 2
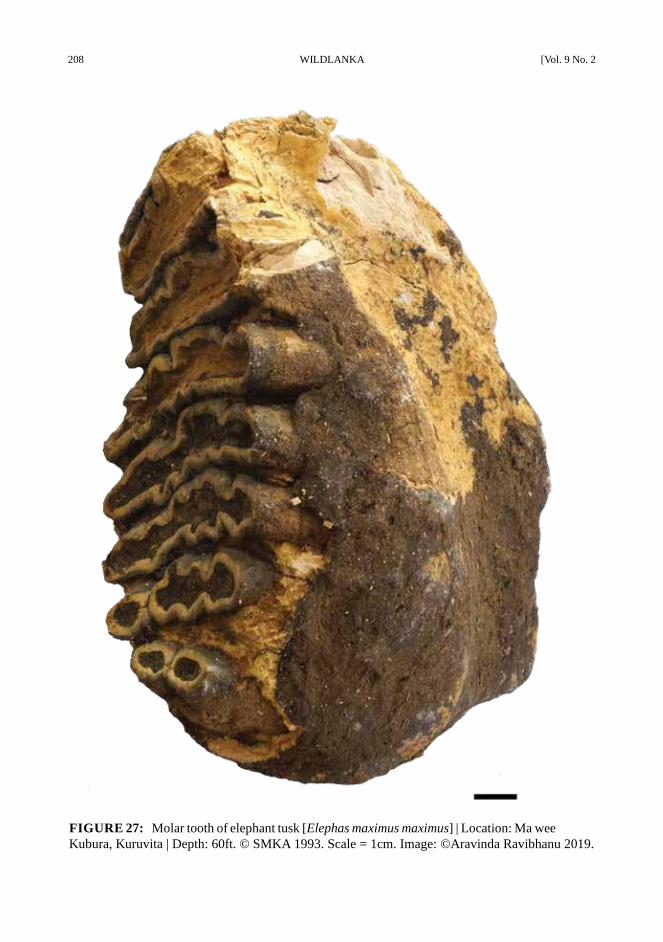
FIGURE 27: Molar tooth of elephant tusk [Elephas maximus maximus] | Location: Ma wee
Kubura, Kuruvita | Depth: 60ft. © SMKA 1993. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 209
FIGURE 28: Piece of elephant molar [Elephas maximus maximus] | Location: Kuruvita | Depth:
40ft. © SMKA 2011. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

210 WILDLANKA [Vol. 9 No. 2
FIGURE 29: Elephant upper molar [Elephas maximus maximus] | Location: Kuruvita | Depth: 45ft.
© SMKA 2007. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 211
FIGURE 30: Elephant molar [Elephas maximus maximus] | Location: Kuruvita | Depth: 65ft. ©
SMKA 2003. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

212 WILDLANKA [Vol. 9 No. 2
FIGURE 31: Piece of elephant tooth [Elephas maximus maximus] | Location: Kuruvita | Depth:
75ft. © SMKA 2003. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 213
FIGURE 32: Piece of elephant molar tooth [Elephas maximus maximus.] | Location: Kahingama,
Kuruvita | Depth: 70ft. © SMKA 1994. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

214 WILDLANKA [Vol. 9 No. 2
FIGURE 33: Piece of elephant molar tooth [Elephas sp.] | Location: Kahingama, Kuruvita | Depth:
50ft. © SMKA 1994. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 215
FIGURE 34: Piece of elephant molar tooth [Elephas maximus maximus] showing digitations on
the 2 to 3 plates | Location: Kuruvita | Depth: 35ft. © SMKA 2000. Scale = 1cm. Image: ©Aravinda
Ravibhanu 2019.

216 WILDLANKA [Vol. 9 No. 2
FIGURE 35: Piece of elephant molar tooth [Elephas sp.] | Location: Kuruvita | Depth: 55ft. ©
SMKA 2000. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 217
FIGURE 36: Piece of elephant molar tooth [Elephas sp.] | Location: Kuruvita | Depth: 50ft. ©
SMKA 2005. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

218 WILDLANKA [Vol. 9 No. 2
FIGURE 37: Piece of elephant molar tooth [Elephas sp.] | Location: Kuruvita | Depth: 45ft. ©
SMKA 2005. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 219
FIGURE 38: Piece of elephant molar tooth (plate) and eroded wear enamel figure from anterior
side [Most probably Elephas maximus sinhaleyus or Palaeoloxodon namadicus sinhaleyus] |
Location: Kuruvita | Depth: 60ft. © SMKA 2005. Scale = 1cm. Image: ©Aravinda Ravibhanu
2019.

220 WILDLANKA [Vol. 9 No. 2
FIGURE 39: Piece of elephant molar tooth (plate)with enamel loop from anterior side [Most
probably Palaeoloxodon namadicus sinhaleyus or Hypselephas hysudricus sinhaleyus] | Location:
Kuruvita | Depth: 65ft. © SMKA 2005. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.
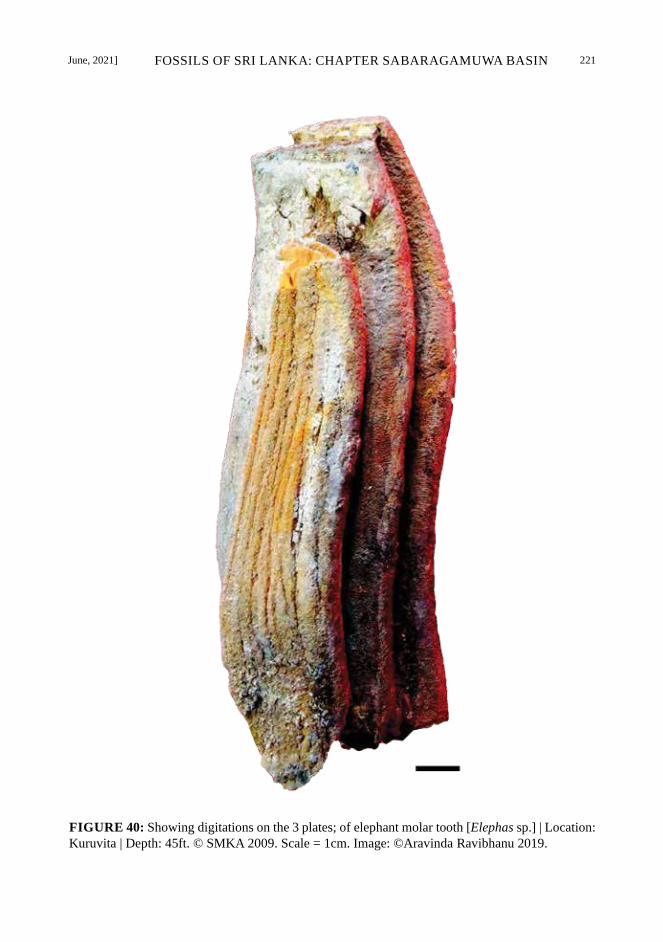
June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 221
FIGURE 40: Showing digitations on the 3 plates; of elephant molar tooth [Elephas sp.] | Location:
Kuruvita | Depth: 45ft. © SMKA 2009. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.
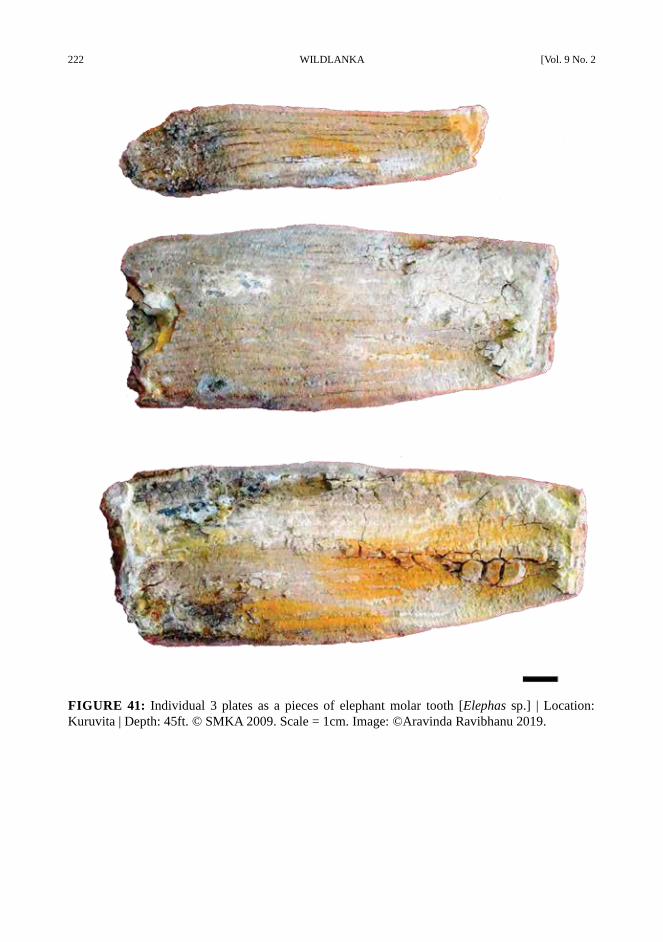
222 WILDLANKA [Vol. 9 No. 2
FIGURE 41: Individual 3 plates as a pieces of elephant molar tooth [Elephas sp.] | Location:
Kuruvita | Depth: 45ft. © SMKA 2009. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 223
FIGURE 42: Elephant [Elephas sp.] molar tooth | Location: Kuruvita | Depth: 25ft. © SMKA
2010. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

224 WILDLANKA [Vol. 9 No. 2
FIGURE 43: Most probably Elephant [Elephas sp.] plate of molar tooth | Location: Kuruvita |
Depth: 45ft. © SMKA 2007. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 225
FIGURE 44: Most probably Elephant [Elephas sp.] plate of molar tooth | Location: Kuruvita |
Depth: 40ft. © SMKA 2008. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

226 WILDLANKA [Vol. 9 No. 2
FIGURE 45: [Top] 3D reconstructed paleo environment of Sabaragamuwa basin in a wet
conditions includes Hypselephas hysudricus sinhaleyus. [Bottom] 3D reconstructed paleo
environment of Sabaragamuwa basin in a partially dry conditions includes Palaeoloxodon
namadicus sinhaleyus and Hypselephas hysudricus sinhaleyus ©Aravinda Ravibhanu 2019.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 227
FIGURE 46: 3D reconstructed paleo environment of Sabaragamuwa basin includes Elephas
maximus maximus ©Aravinda Ravibhanu & Adeepa Nisal 2019.

228 WILDLANKA [Vol. 9 No. 2
Family Rhinocerotidae | Rhinoceros
The rhinoceros’ family is characterized
by its large size (one of the largest remaining
megafaunas), with all of the species able to
reach one ton or more in weight; an herbivorous
diet; and a thick protective skin about 1.5–5 cm
thick, formed from layers of collagen positioned
in a lattice structure; relatively small brains for
mammals this size (400–600g); and a large horn.
They generally eat leafy material, although their
ability to ferment food in their hindgut allows
them to subsist on more fibrous plant matter,
if necessary. Unlike other perissodactyls, the
African species of rhinoceros lack teeth at the
front of their mouths, relying instead on their
powerful premolar and molar teeth to grind
up plant food. Both African species and the
Sumatran Rhinoceros have two horns, while the
Indian and Javan Rhinoceros have a single horn.
Rhinoceros was living 80,000 years ago.
The most known fossil remains of
Rhinoceros unicornis are estimated apparently
to be dated to probably middle Pleistocene
period (Fig 47). The direct precursor of the
living Indian rhinoceros was Rhinoceros
unicornis fossilis (synonyms R.sivalensis and
R. palaeindicus), from the upper Shiwalik beds,
within the known historic range of the species.
Rhinoceros from the Narbada or Narmada
beds is probably synonymous with Rhinoceros
unicornis Rhinoceros kendengindicus from Java
was closely related to the present species and
should probably be regarded as a subspecies of
it.
FIGURE 47: Javan and Indian rhino by CGTN Graphic and Rhinoceros unicornis at Kaziranga
National Park, India © EPA-EFE | RE Aravinda Ravibhanu 2019. Javan and Indian rhino by CGTN
Graphic.
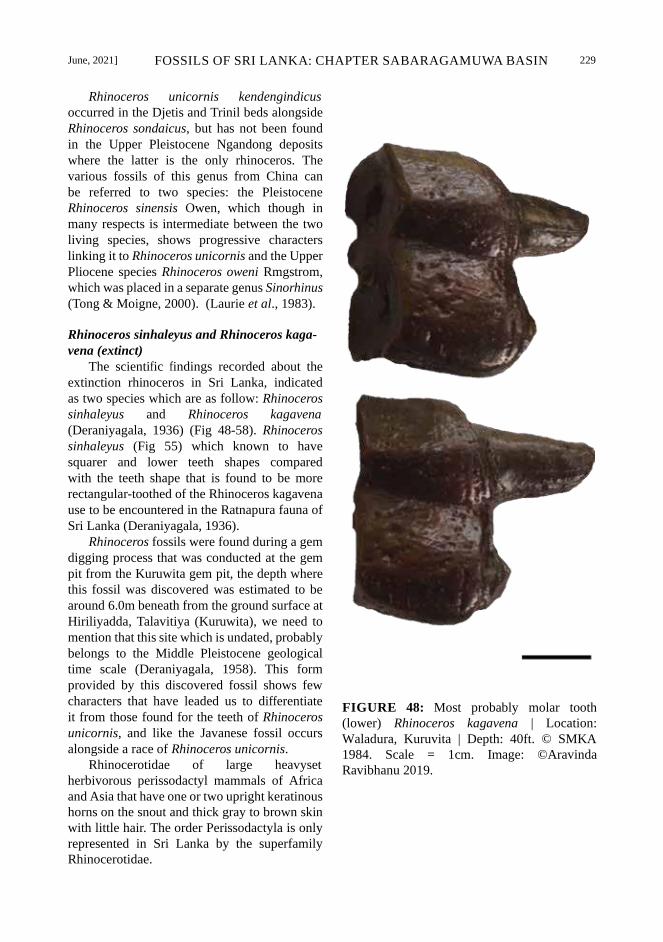
June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 229
Rhinoceros unicornis kendengindicus
occurred in the Djetis and Trinil beds alongside
Rhinoceros sondaicus, but has not been found
in the Upper Pleistocene Ngandong deposits
where the latter is the only rhinoceros. The
various fossils of this genus from China can
be referred to two species: the Pleistocene
Rhinoceros sinensis Owen, which though in
many respects is intermediate between the two
living species, shows progressive characters
linking it to Rhinoceros unicornis and the Upper
Pliocene species Rhinoceros oweni Rmgstrom,
which was placed in a separate genus Sinorhinus
(Tong & Moigne, 2000). (Laurie et al., 1983).
Rhinoceros sinhaleyus and Rhinoceros kaga-
vena (extinct)
The scientific findings recorded about the
extinction rhinoceros in Sri Lanka, indicated
as two species which are as follow: Rhinoceros
sinhaleyus and Rhinoceros kagavena
(Deraniyagala, 1936) (Fig 48-58). Rhinoceros
sinhaleyus (Fig 55) which known to have
squarer and lower teeth shapes compared
with the teeth shape that is found to be more
rectangular-toothed of the Rhinoceros kagavena
use to be encountered in the Ratnapura fauna of
Sri Lanka (Deraniyagala, 1936).
Rhinoceros fossils were found during a gem
digging process that was conducted at the gem
pit from the Kuruwita gem pit, the depth where
this fossil was discovered was estimated to be
around 6.0m beneath from the ground surface at
Hiriliyadda, Talavitiya (Kuruwita), we need to
mention that this site which is undated, probably
belongs to the Middle Pleistocene geological
time scale (Deraniyagala, 1958). This form
provided by this discovered fossil shows few
characters that have leaded us to differentiate
it from those found for the teeth of Rhinoceros
unicornis, and like the Javanese fossil occurs
alongside a race of Rhinoceros unicornis.
Rhinocerotidae of large heavyset
herbivorous perissodactyl mammals of Africa
and Asia that have one or two upright keratinous
horns on the snout and thick gray to brown skin
with little hair. The order Perissodactyla is only
represented in Sri Lanka by the superfamily
Rhinocerotidae.
FIGURE 48: Most probably molar tooth
(lower) Rhinoceros kagavena | Location:
Waladura, Kuruvita | Depth: 40ft. © SMKA
1984. Scale = 1cm. Image: ©Aravinda
Ravibhanu 2019.

230 WILDLANKA [Vol. 9 No. 2
FIGURE 49: Most probably proximal portion of scapula bone of Rhinoceros sp.| Location:
Paradise, Kuruvita | Depth: 55ft. © SMKA 1999. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.
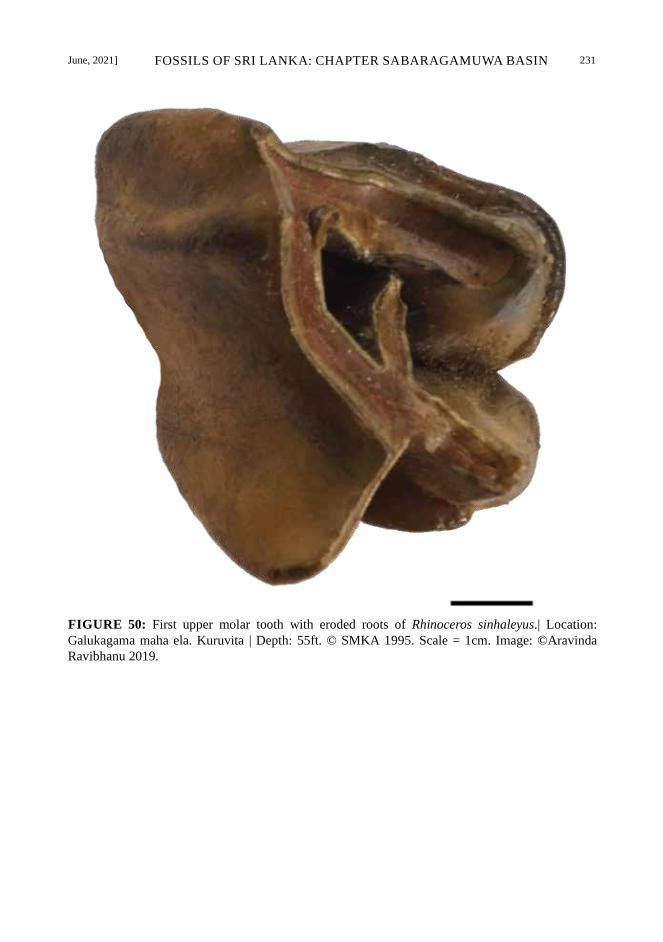
June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 231
FIGURE 50: First upper molar tooth with eroded roots of Rhinoceros sinhaleyus.| Location:
Galukagama maha ela. Kuruvita | Depth: 55ft. © SMKA 1995. Scale = 1cm. Image: ©Aravinda
Ravibhanu 2019.

232 WILDLANKA [Vol. 9 No. 2
FIGURE 51: Lower third molar tooth of Rhinoceros kagavena.| Location: Kahingama west, Kuburu
yaya Kuruvita | Depth: 57ft. © SMKA 1998. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 233
FIGURE 52: Pieces of molar tooth of Rhinoceros sinhaleyus.| Location: Kuruvita | Depth: 50ft. ©
SMKA 2000. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

234 WILDLANKA [Vol. 9 No. 2
FIGURE 53: Most probably piece of vestibular of Rhinoceros kagavena.| Location: Kuruvita |
Depth: 48ft. © SMKA 2004. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 235
FIGURE 54: Second upper molar tooth of Rhinoceros sinhaleyus | Location:.Kuruvita | Depth:
45ft. © SMKA 1995. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

236 WILDLANKA [Vol. 9 No. 2
FIGURE 55: 3D reconstructed paleo environment of Sabaragamuwa basin includes Rhinoceros
sinhaleyus | Location:.Waladura, Kuruvita ©Aravinda Ravibhanu 2019.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 237
FIGURE 56: 3D reconstructed paleo environment of Sabaragamuwa basin includes Rhinoceros
sinhaleyus (left) and Rhinoceros kagavena (right) ©Aravinda Ravibhanu 2019.

238 WILDLANKA [Vol. 9 No. 2
FIGURE 57: Most probably Rhinoceros kagavena molar tooth. | Location:.Ekneligoda, Kuruvita |
Depth: 35ft. © SMKA 1998. Scale = 1cm. Image ©Aravinda Ravibhanu 2019.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 239
FIGURE 58: 3D reconstructed paleo environment of Sabaragamuwa basin includes Rhinoceros
kagavena | Location: Paradise, Kuruvita. ©Aravinda Ravibhanu 2019.

240 WILDLANKA [Vol. 9 No. 2
Family Hippopotamidae | Hexaprotodon
sinhaleyus, (extinct)
The hippopotamus (Hippopotamus
amphibius), or hippo, from the ancient Greek
for “river horse”, is a large, mostly herbivorous
mammal in sub-Saharan Africa, and one of only
two extant species in the family Hippopotamidae,
the other is the Pygmy Hippopotamus. The
earliest known hippopotamus fossils, belonging
to the genus Kenyapotamus in Africa, roughly
16 million to 8 million years ago during
the Miocene epoch. After the elephant, the
hippopotamus is the largest land mammal and
the heaviest extant artiodactyls, despite being
considerably shorter than the giraffe.
The hippopotamus is semi-aquatic,
inhabiting rivers and lakes where territorial
bulls preside over a stretch of river and groups
of 5 to 30 females and young. During the day
they remain cool by staying in the water or mud;
reproduction and childbirth both occur in water.
They emerge at dusk to graze on grass. While
hippopotamus uses rest near each other in the
water, grazing is a solitary activity and hippos
are not territorial on land.
In the ‘Pleistocene of Ceylon’Deraniyagala
(1936, 1939, 1944 and 1958) explains his
findings of Hexaprotodon sinhaleyus and
Hexaprotodon sivalensis sinhaleyus based on
gem pits in the Ratnanapura area about seven
kilometers away at Pahala Vela, Galadande
Mandiya, Gonapitiiya, and Kuruwita near the
Kuru Ganga (Sumanarathna et al., 2016) (Fig
59-67). The fossils were laid at a depth of 6.5m
below the surface. Accordingly, Deraniyagala
revealed the fossilized remains of the lower
jaws and teeth of a Sri Lankan hippopotamus.
The lower jawbone of the hippopotamus reveals
six incisor teeth, whereas the hippopotamus
that survives in Africa has only four incisors.
The extinct Ceylon hippopotamus has been
named the Hexaprotodon sinhaleyus, (Fig 67)
means consisted of six teeth in front includes
six permanent incisors as mentioned. Genera
is classified as an extinct hippopotamus living
in the Pleistocene in Asian regions such as
Sri Lanka, India, Java, Syria and Burma.
Considering the Hexaprotodon sinhaleyus
fossils in Rathnapura beds, dominantly found
molars, incisor canine, premolar, zygomatic
arch, humerus, femur, ribs. Hexaprotodon
sinhaleyus is endemic to Sri Lanka and quite
similar to Hexaprotodon namadicus (extinct)
once lived in the Pleistocene in India. The change
in climate from heavy rainfall that fed numerous
large rivers and lakes to a more moderate
rainfall that reduced the island’s waterbodies
was probably responsible for the extinction of
the world’s second heaviest land mammal on
the island (Deraniyagala, 1958). The extinction
of this animal might have occurred sometime
shortly after the middle Pleistocene times,
since its nearest a relative, the extinct Indian
hippopotamus from former lake beds which
are now traversed by the Nerbudda (Narmada)
River, became extinct in Ionian times.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 241
FIGURE 59: Piece of molar tooth of Hexaprotodon sinhaleyus | Location: Waladura, Kuruvita
| Depth: 55ft. © SMKA 1986. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

242 WILDLANKA [Vol. 9 No. 2
FIGURE 60: Piece of molar tooth of Hexaprotodon sinhaleyus | Location: Paradise gem pit,
Kuruvita | Depth: 50ft. © SMKA 1995. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 243
FIGURE 61: Pieces of lower canine tooth - Hexaprotodon sinhaleyus | Location: Ellawala,
Pahalagama | Depth: 48ft. © SMKA 1994. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

244 WILDLANKA [Vol. 9 No. 2
FIGURE 62: Piece of metatarsal - Hexaprotodon sinhaleyus | Location: Ovita Kubura, Kuruvita |
Depth: 75ft. © SMKA 1994. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.
June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 245
FIGURE 63: Piece of molar tooth - Hexaprotodon sinhaleyus | Location: Eedandawala, Kuruvita |
Depth: 60ft. © SMKA 1987. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.
FIGURE 64: Piece of molar tooth of Hexaprotodon sinhaleyus | Location: Kuruvita | Depth: 65ft.
© SMKA 2000. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

246 WILDLANKA [Vol. 9 No. 2
FIGURE 65: Molar tooth of Hexaprotodon sinhaleyus | Location: Kuruvita | Depth: 50ft. © SMKA
2005. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 247
FIGURE 66: Pieces of lower canine tooth - Hexaprotodon sinhaleyus | Location: Ellawala,
Pahalagama | Depth: 45ft. © SMKA 1994. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

248 WILDLANKA [Vol. 9 No. 2
FIGURE 67: 3D reconstructed paleo environment of Sabaragamuwa basin includes Hexaprotodon
sinhaleyus | Location: Paradise, Kuruvita ©Aravinda Ravibhanu 2019.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 249
Family Crocodylidae | Crocodylus sinhaleyus
(extinct)
The extinct crocodile species called
Crocodylus sinhaleyus has been nomenclature
by P.E.P Deraniyagala in 1953. This extinct
crocodilian that probably attained to a length
of 12ft; known from a single tooth, Colombo
National Museums No. F. 28 (Deraniyagala
1958). Considering the dimensions of specimen,
depth of tooth 43mm, diameter of base 19mm,
circumference of base 58mm. The distal
extremely of the pulp cavity is present and when
reconstructed the tooth will probably be about
95mm long (Deraniyagala 1958). This was
discovered on 17th of July 1939 from the gem
sand in a gem pit at tunhiriya vila, Gonavitiya,
Kuruvita from a depth of 12 feet together
with teeth of rhinoceros (Deraniyagala 1958).
Also, Crocodylus porosus minikanna which
is occasionally captured in sabaragamuwa
beds (Kalu gaga- river - near rathnapura)
was the only species around 1940 decades
(Deraniyagala 1953,1958). But currently only
the Crocodylus porosus lives in the Ratnapura
area (Sumanarathna et al., 2016) (Fig 68-69).
FIGURE 68: A tooth of Crocodylus sp. | Location: Kahengama,Ovitakubura, Kuruvita | Depth:
25ft. © SMKA 2013. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

250 WILDLANKA [Vol. 9 No. 2
FIGURE 69: 3D reconstructed paleo environment of Sabaragamuwa basin includes Crocodylus
sinhaleyus | Location: Karapincha, Kuruvita ©Aravinda Ravibhanu 2019.
Family Suidae | Sus sinhaleyus (extinct)
First record of the Sus sinhaleyus by
Deraniyagala (P.E.P) who published in Journal
of Royal Asiatic Soc. (Ceylon Br.) in 1947. The
article was based on five teeth that belonged
to different individuals as each was obtained
from a separate gem pit from Potgul _kanda
near Ratnapura. As he mentioned, the heel of
the third molar is less complex and the teeth
are considerably smaller and more brachydont
than in the living Ceylon wild pig. Most
probably this species was about two-thirds the

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 251
size of the existing Sus scrofa zeylonicus. Due
to Deraniyagala (P.E.P), please following pre
rescored measurement of Sus sinhaleyus.
Dentition - The molars are sub-brachydont;
the third molar consists of three longitudinal
rows of cusps of which the median row is
reduced and interrupted by the three labial and
three lingual cusps being contiguous mesially.
This tooth possesses two ordinary cusps and
the third enlarged into the posterior talonid. The
anterior talonid is a flat, wide plate formed by
the cingulum.
Type - Colombo Museum No. F. 194, a last
lower right molar with the two anterior cusps
slightly worn. Three median cusps with their
bases hidden by the labia1 and lingual once. The
enamel on the cusps is smooth and uncrenulated.
The cusps are simple, with two median ones
anteriorly, and one median one posteriorly
on the talon, which latter is as long ss the
remainder of the tooth (PI. L). This anterior part
of the fossil tooth is not very different to that
of Sus scrofa zeylonicus but is less hypsodont,
the talon possesses only a single middle cusp
instead of two, and terminates posteriorly in
one strong cusp instead of several small once.
Compare Pl. L with Fig. 1194 in Nich Ison and
Lydekker (1889).
Locality - From the gem sand at Gönäpitiya,
Kuruvita, at a depth of 15 feet below the surface.
Paratype - Colombo Museum No. 174 b,
an unworn last, upper, left molar, from which
the talon area is missing; the width, depth and
shapo of the cusps with uncrenulated enamel
indicate that this tooth is co-specific with the
type. The unworn cusps are less prominent and
1088 differentiated from one another than in Sus
s. zeylonicus and there are fewer accessory ones
(Deraniyagala, 1957).
Make sure to consider measurement data
mention above is providing to develop 3D
reconstruction application via AR and VR. (As
a source data for VR\AR developers)
TABLE 04: A comparison of the last, lower, right molar of the two species is as follows
(Deraniyagala, 1957).
Number F 57- 143 – 16
Measurements Sus sihalayus Sus s. zeylonicus
Basal Length 32mm 45mm
Greatest width 15mm 18mm
Depth 12mm 18mm
Length of talon 16mm 21mm
TABLE 05: Locality - From the gem sand at Sannasgama, near Pälmädulla from a depth of 9
feet Tooth were also secured from Gonapitiya near Kuruvita from gem sand at a depth of 15 feet
(Deraniyagala, 1957).
Measurements Fossil No – F 57 – 154 – 15 Sus s. zeylonicus
Length 21+mm 29mm
Width 19mm 21mm
Depth 12mm 10mm
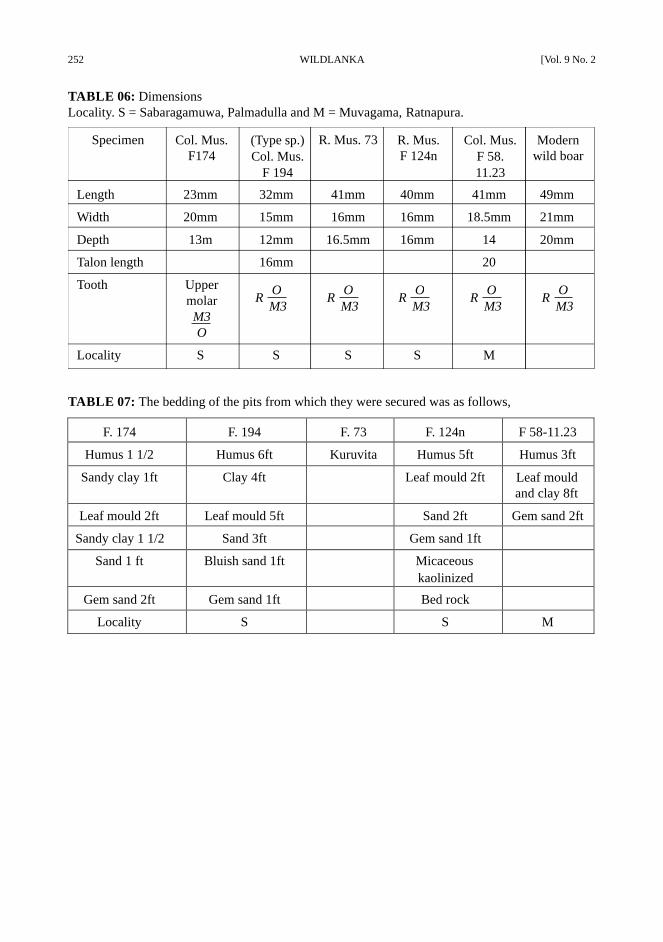
O O O O O
Specimen Col. Mus.
F174
(Type sp.)
Col. Mus.
F 194
R. Mus. 73 R. Mus.
F 124n
Col. Mus.
F 58.
11.23
Modern
wild boar
Length 23mm 32mm 41mm 40mm 41mm 49mm
Width 20mm 15mm 16mm 16mm 18.5mm 21mm
Depth 13m 12mm 16.5mm 16mm 14 20mm
Talon length 16mm 20
Tooth Upper
molar
M3
O
R
M3
R
M3
R
M3
R
M3
R
M3
Locality S S S S M
252 WILDLANKA [Vol. 9 No. 2
TABLE 06: Dimensions
Locality. S = Sabaragamuwa, Palmadulla and M = Muvagama, Ratnapura.
TABLE 07: The bedding of the pits from which they were secured was as follows,
F. 174 F. 194 F. 73 F. 124n F 58-11.23
Humus 1 1/2 Humus 6ft Kuruvita Humus 5ft Humus 3ft
Sandy clay 1ft Clay 4ft Leaf mould 2ft Leaf mould
and clay 8ft
Leaf mould 2ft Leaf mould 5ft Sand 2ft Gem sand 2ft
Sandy clay 1 1/2 Sand 3ft Gem sand 1ft
Sand 1 ft Bluish sand 1ft Micaceous
kaolinized
Gem sand 2ft Gem sand 1ft Bed rock
Locality S S M

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 253
FIGURE 70: Lower jaw_ tooth of Sus sinhaleyus | Location: Wakadura, Kuruvita | Depth: 48ft. ©
SMKA 2013. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

254 WILDLANKA [Vol. 9 No. 2
Family Bovidae | Bibos sinhaleya (extinct)
Considering the current distribution of
Gaur, a wild cattle native to tropical forests of
Southeast Asia and the Indian subcontinent.
Today, the range of the species is seriously
fragmented, and it is regionally extinct in Sri
Lanka. Frist fossil record (femur) of Bibos
gaurus sinhaleyus in Sri Lanka, discovered
close to Elapatha, Rathnapura (Deraniyagala,
1939,1951,1958). Due to present exploration in
sabaragamuwa beds have identified many parts
of fossils which belong to Bibos sinhaleya as
femurs, vertebrae, premolars, molars, humerus
metacarpals.
“A sort of beast they call Gauvera, so
much resembling a Bull, that I think it one
of that kind. His back stands up with a
sharp ridge; all his four feet white up half his
Legs. I never saw but one, which was kept
among the King’s Creatures”. This is how,
Robert Knox_ express his live scenery
regarding the Bibos sinhaleya in the book
called An Historical Relation of the Island of
Ceylon in the East Indies (Knox, 2016).
FIGURE 71: Most probably vertebra of Bibos sinhaleya | Location: Kahengama west, Kuruvita |
Depth: 35ft. © SMKA 2008. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.
June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 255
FIGURE 72: Bovine tooth | Location: Moladeniya, Paradise, Kuruvita | Depth: 30ft. © SMKA
1995. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.
256 WILDLANKA [Vol. 9 No. 2
FIGURE 73: Bovine upper molar tooth | Location: Gonapitiya, Kuruvita | Depth: 40ft. © SMKA
1997. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 257
FIGURE 74: Decay and eroded bone of Metacarpals_ Bibos sinhaleya | Location: Gonapitiya,
Kuruvita | Depth: 55ft. © SMKA 1995. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

258 WILDLANKA [Vol. 9 No. 2
FIGURE 75: 3D reconstructed paleo environment of Sabaragamuwa basin includes Bibos sinhaleya
| Location: Bubuladeniya, Kuruvita ©Aravinda Ravibhanu 2019.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 259
Family Cervidae | Muva sinhaleya and Rusa
unicolor (extinct)
Cervidae is a family of hoofed ruminant
mammals that belongs to the order of
Artiodactyla. Males are usually available
with horns cores upon the frontal bones and
supporting solid, bony, deciduous antlers that
are generally branched. Considering current
fauna statutes in Sri Lanka, there are four genus
that can be identified, which are documented
to belong to the family Cervidae like Axis
axis, Axis porcinus, Muntiacus muntjak, Rusa
unicolor.
According to Deraniyagala reports which
provided important database for this specie
identification, a small deer (Muntiacus muntjak)
was documented o be known from a single
antler base consisting of the part of the bezel
and a short length of the beam and the brow tine
which is found at the Ratnapura Museum F264.
Furthermore, based on Dr. Deraniyagala
research he have presented the first holotypic
reading of Muva sinhaleya, based on a fossils
samples found at Mimavaladeniya, Gonapitiya,
and at Kuruvita (Deraniyagala, 1958). The
scientific records of the fossils that belongs
to the genus Axis axis and Axis porcinus are
not found to be rare among the fossils of the
family Cervidae that lived previously in the
Pleistocene period that occurred in Sri Lankan
territory. These two species have been present
continuously from the Pleistocene period to the
present geological time scale. In fact, the Rusa
unicolor deer is the largest genus category of
the deer family in Sri Lanka we can found this
specie occupying different habitat at all altitudes
ranges and in all climatic diversified zones.
Still, reports and data have documented that
the fossilized horns, hooves, teeth, mandibles,
etc that belongs to the family Cervidae are
found in the gem mines associated with the
Sabaragamuwa bed (Sumanarathna et al., 2016)
(Fig. 76-81).
Further more, we estimated the encounter
of this fossils specifically at the mining sites
of the Sabaragamuwa bed perhaps due to
the fact that continues digging surveys are
conducted in these gem mines places, as well,
the digging in this gem mines are reaching a
considerable depth in which obviously have
been documented to have important fossils
findings. Based on this valuable research
records we believe that the Sabaragamuwa bed
may have still many valuable fossilized remains
of some of the Sri Lankan unique species.
Based in our interpretation of the data recorded
we have presented a 3D reconstructed paleo
environment (Figure. 81) of Sabaragamuwa
basin that includes Rusa unicolor genus, at
Bubuladeniya, Kuruvita.

260 WILDLANKA [Vol. 9 No. 2
FIGURE 76: Mandible of Rusa unicolor | Location: Waladura, Kuruvita | Depth: 25ft. © SMKA
2007. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 261
FIGURE 77: Fossil of antler, Rusa unicolor | Location: Paradise, Kuruvita | Depth: 23ft. © SMKA
2005. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

262 WILDLANKA [Vol. 9 No. 2
FIGURE 78: Fossil of antler, Rusa unicolor | Location: Paradise, Kuruvita | Depth: 18ft. © SMKA
2000. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 263
FIGURE 79: Fossil of antler, Rusa unicolor | Location: Endana, Kuruvita | Depth: 30ft. © SMKA
1998. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

264 WILDLANKA [Vol. 9 No. 2
FIGURE 80: Deer fossil of antler | Location: Paradise, Kuruvita | Depth: 45ft. © SMKA 1996.
Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 265
FIGURE 81: 3D reconstructed paleo environment of Sabaragamuwa basin includes Rusa unicolor
| Location: Bubuladeniya, Kuruvita ©Aravinda Ravibhanu | Adeepa Nisal 2019.

266 WILDLANKA [Vol. 9 No. 2
Fossils and Semi Fossils shells
Paleoecology uses essentially a combination
of geological and biological pieces of evidence
from fossil deposits that will lead to investigate
deeply and to understand clearly the past
occurrence, distribution, and abundance of
different ecological units on a variety of time
scales, which provides scientific evidence for the
present and future research analysis and habitat
reconstruction scenarios. The Sabaragamuwa
Basin, in Sri Lanka provides dominant type
of natural eco-Paleo evidences (Deraniyagala,
1958). The cultural remains of early men of this
area were discovered together with their skeletal
fragments and geometric microliths.
Other detections include various types
of fauna and flora that are thought to have
formed part of the human diet, also the animal
bones, which were fossilized adjoining the
Sabaragamuwa Basin like we discussed (Fig 86-
87). As a result of the paleo-diet of Balangoda
man (Homo sapiens), who lived in the cave of
pre historic site, Batadombalena cave (38,000
BP) which was the proper Harbor life station
(Sumanarathna et al., 2016), accumulated many
snails fossil shells (Acavidae and Pleurocridae
Family), such as Acavus phoenix, Acavus
superbus, Oligospira waltoni, Paludomus
loricate, Paludomus neritoides, Paludomus
sulcatus (Fig. 82-85).
FIGURE 82: Shells of snail fossils via 3rd excavation of “Batadoba-lena Pre Historic Site”
by Dr,Nimal Perera and his team in 2005. Image © Nimal Perera 2005.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 267
FIGURE 83: [Top and Bottom] : Snail fossils_ Oligospira skinneri & Acavus superbus |
Location: Batadomba-lena, Kuruvita | Depth: 10ft. © SMKA 2005. Scale = 1cm. Image:
©Aravinda Ravibhanu 2019.

268 WILDLANKA [Vol. 9 No. 2
FIGURE 84: [Top and Bottom] : Snail fossils_ Paludomus sp. | Location: Batadomba-lena,
Kuruvita |Depth: 10ft. © SMKA 2005. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 269
FIGURE 85: A piece from foot of shell. Most probably typical formation might be a prefect tool
for apply as a fishing hook | Location: Near Booth Ella water fall, Udakada, Kuruvita | Depth: 8ft. ©
SMKA 2003. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

270 WILDLANKA [Vol. 9 No. 2
Family Cyprinidae | Tor khudree
FIGURE 86: 3D reconstructed paleo environment of Boopath Ella water fall includes Tor khudree
| Location: Kuruvita ©Aravinda Ravibhanu 2019.
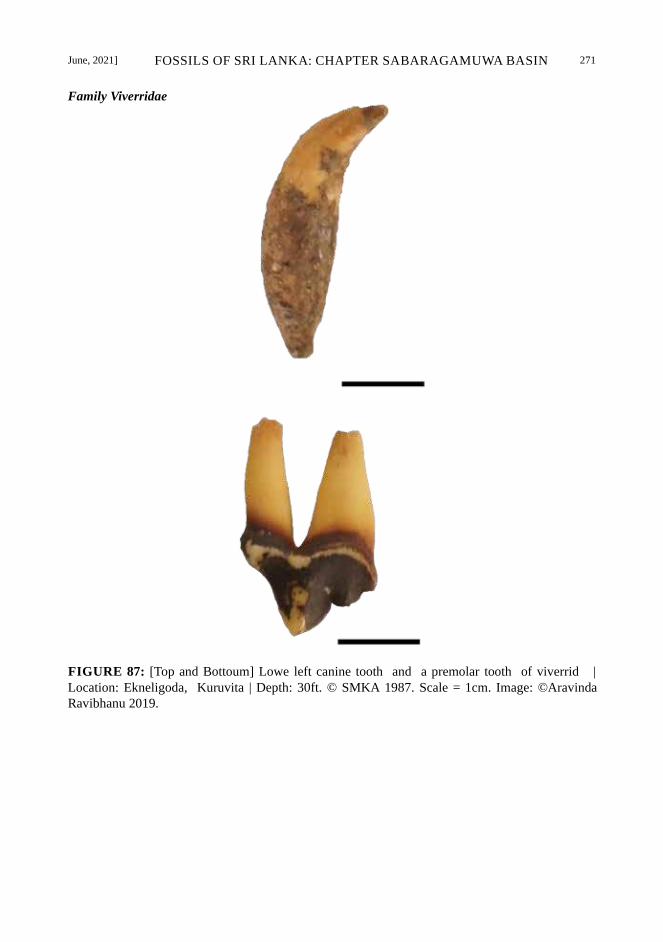
June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 271
Family Viverridae
FIGURE 87: [Top and Bottoum] Lowe left canine tooth and a premolar tooth of viverrid |
Location: Ekneligoda, Kuruvita | Depth: 30ft. © SMKA 1987. Scale = 1cm. Image: ©Aravinda
Ravibhanu 2019.

272 WILDLANKA [Vol. 9 No. 2
Paleobotany
This is the scientific study of paleo flora,
using plant fossils via process of fossilization.
These fossils can be impressions, compressions
a of the plants left on the sedimentary rock’s
surface, or “petrified” objects, such as wood,
which preserve the original plant material
in rocklike form (Fig 88-95). Additionally,
some perversion type in gastrolith can be
contaminate parts of flora seeds. Considering
the Rathnapura flora in Pleistocene Sri
Lanka follow as Alsophila zeylanica (Pini
Baru), Bambusa vulgaris Schard (Bambu),
Ochlandra stridual Thwaites (Bata), Ochlandra
fasciculata Thwaites (Katu Kithul), Caryota
urens Linne (Kithul), Elaeocarpus subvillosus
Arn (Gal Veralu), Myristica dactylodies Caertn
(Malaboda), Canarium zeylanicum (Kekuna),
Coscinium fenestrum Coleber (Veni Val)
(Deraniyagala, 1958).
The Samples (Fig. 91, Fig. 92, Fig. 94, Fig.
95) deposited in a Sabaragamuwa bed, there
were rivers flowing through it, and on the banks
of those rivers grew plants. When the water
stopped flowing, the river banks were parched
in the sun and any water in the sediments were
drawn out. In the process, minerals such as
calcium became concentrated around the roots
hardening into what is known as ‘Caliche’,
cementing the sands and preserving the form of
the roots.
FIGURE 88: A part of Artocarpus nobilis leaf | Location: Ekneligoda, Kuruvita | Depth: 55ft. ©
SMKA 2011. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 273
FIGURE 89: Canarium zeylanicum | Location: Vithanakanda, Kuruvita | Depth: 20ft. © SMKA
2005. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

274 WILDLANKA [Vol. 9 No. 2
FIGURE 90: Most probably, a part of the leaf of Elaeocarpus sp. | Location: Ekneligoda, Kuruvita
| Depth: 20ft. © SMKA 2006. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 275
FIGURE 91: Rhizoconcretions via silica and calcium carbonate replacement of root | Location:
Paradise, Kuruvita | Depth: 65ft. © SMKA 2007. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

276 WILDLANKA [Vol. 9 No. 2
FIGURE 92: Concrete roots by process of rhizoconcretions. | Location: Bubuladeniya, Kuruvita |
Depth: 85ft. © SMKA 2004. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 277
FIGURE 93: Unclassified amber | Location: Vithanakanda, Kuruvita | Depth: 55ft. © SMKA 2003.
Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

278 WILDLANKA [Vol. 9 No. 2
FIGURE 94: Root rhizoconcretions via silica and calcium carbonate replacement | Location:
Paradise, Kuruvita | Depth: 85ft. © SMKA 2003. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 279
FIGURE 95: Concrete roots by process of rhizoconcretions. | Location: Gonapitiya, Kuruvita |
Depth: 80ft. © SMKA 2002. Scale = 1cm. Image: ©Aravinda Ravibhanu 2019.

280 WILDLANKA [Vol. 9 No. 2
Paleoenvironment and Extreme Conditions |
Planetary Geology
Sabaragamuwa basin consisted unique
paleoenvironment that has been preserved in the
fossils and rock record in the quaternary period.
Sometimes it has been represented an extreme
environment which is a habitat that is considered
very hard to survive in due to its considerable
factors, such as temperature, accessibility to
different energy. Even the heavy tectonic actions
such as faulting, tilting, dislocation and block
hosting that occurred during the Pleistocene
caused the present association of fossils of
different ages in the gem sand by breaking down
and redepositing fossiliferous strata of different
ages doubtless affected spaces formation in Sri
Lanka (Deraniyagala, 1958).
Considering the redepositing patterns of
alluvial soil in Sabaragamuwa beds is quite
supportive to make a comparative simulation
on Exo-terrestrial landslides (Scaioni et al.,
2018; Sumanarathna 2019, 2020). Therefore,
strong climatic fluctuation also occurred during
the Pleistocene influences of certain elements
in the fauna of Sri Lanka. The Montane lizard
Cophotis which dies within a day or two
when brought from the cool montane down to
Colombo. (Deraniyagala, 1958).
Whole extreme conditions around the
Pleistocene epoch in Sri Lanka is the perfect
platform to discuss many events corresponding
to Extreme Life formation via researching
Planetary geology and Astrobiology (Fig. 96).
Hence, here we partially discuss fundamentals
analog in extreme and habitable life conditions.
Researchers often approach to debate the
question of how life began on Earth or if it
could be started elsewhere, by focusing their
efforts generally to involve attempts for a clear
understanding on how non-biological molecules
bonded, resulting in an increasingly complex
type of life, which eventually could replicate
and somehow could use sources of energy to
make biological, chemical as well as physical
reactions.
Undeniably, for life mechanism processing
in our cosmos, we needed both parameters,
which are biomass as well as energy. Scientists
have considered studying and understanding
this complex life process. Furthermore, some
have sought to make synthetic life out of both
selected components and energy. Incredible
achievements have been realized for both of
these endeavors, still many mysteries of life
concepts remain unexplained.
That what makes the origin of life on Earth
difficult to be fully understood. To
complicate the picture, the history of early Earth
documents a period of extreme heat caused
by meteorite bombardment, without
forgetting the most important event, which
is the enormous impact that happened around
4.5 billion years ago on the Mars-sized planet
that became our moon. In conclusion, many
early researchers think that our planet was
uninhabitable until about 4 billion years ago.
In fact, some researchers argue the fact that
signs of Earth life were detected in the rocks
record around 3.8 billion years ago, which
makes them suggest that life forms were indeed
present since 3.5 billion years ago on Earth.
Considering the early evolution, our planet
Earth supported only a single-cell form of
life estimated to be for several billion years
before multicellular life emerged and evolved.
However, researchers are skeptical about the
likelihood of DNA-based on life evolving
and the relatively short period between when
Earth became cool enough to support life and
the earliest evidence of actual life recorded
(Sumanarathna 2020a, 2020b) (Fig. 97). In
another hand, theories invoke panspermia and
the mechanism of life spreading via meteorites
in some cases delivered through comets around
the cosmos. Based on what we discussed till
now, an ultimate question needs to be debated,
which is whether Earth has been seeded by early
Extraterrestrial life.
Theories about the possibility of life
development in the red planet Mars become
now accepted by some scientists, some
speculate that Mars was habitable in its early
period than Earth. But panspermia inherently
could go the other way as well, or possibly even
around our solar system. Furthermore, multiple
deep questions rise in this debate. Could science
explain the process that made life been created
from non-life? Based on a concept such as Eco-

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 281
FIGURE 96: 3D reconstructed_ alluvial pan of the paleo environment in Sabaragamuwa basin |
Location: Hidurangala, Eheliyagoda ©Aravinda Ravibhanu 2019.
Astronomy research on extreme environmental
conditions that affect the Harbor life creation
cycle.
A science concept is now being forged
within the Union of the General Theory of
Eco Astronomy Mechanics and Concept. This
concept is all about studies that documented
the extreme conditions that synchronize with
evolutionary theory and the adaptational cases
through quantitative methods (Relevant to
Paleontology and Petrology) using the energetic
organism-machine analogy as a guideline.
Which seeks to reduce the complex form to
fewer generating factors and causal influences
cases.
If a function will be postulated for a structure,
then its optimum stat form, or paradigm can be
specified on mechanical grounds corresponding
to an extreme environment. The approach
about a structure to its paradigm provides the

282 WILDLANKA [Vol. 9 No. 2
FIGURE 97: 3D reconstructed_ alluvial pan of the Coprates chasma, Mars | © Giulia Magnarini/
NASA.
elusive criterion, relative efficiency that any
science of adaptation and evolution requires.
Physical laws and forces also specify that form
is adapted to the requirements of space. When
we cannot establish paradigms on deductive
criteria, an experimental approach to form is
appropriate. Idealized models are favored over
actual specimens because they can be built to
test predetermined factors.
Eco Astronomy needs to be based not
solely on descriptive science made through
observational methods but may as well adopt
the experimental techniques of explanatory
procedures filled. It is inconceivable that
each aspect of a complex form of life is an
obvious direct product of individual energetic
instruction. We can simplify the process, and
thereby understand, the extreme conditions
in which results in apparent complexity by
recognizing that physical forces directly
influence, the shape of substrates that circulate
energy and that a few simple rules can fashion
some very intricate final products.
These same rules can be programmed;
computers have simulated structures that bear
remarkable correspondence to actual forms; the
extreme condition for the harboring condition
of life need to be no more complex. Also,
these rules can be used to generate a range of
potential evolutional forms available to match
with Homo sapiens structure. The actual forms
we know fill specifically a part of the total
spectrum; their basic adaptation may be grasped
when we notice why unoccupied areas are not
used.
Among inductive studies done about harbor
life, univariate techniques display trends and
rates of change for single characters; they
have been applied to the periodic growth
lines of fossil shells, providing thereby a
paleontological input to geophysics as a
multidisciplinary interpretation stage. Anyhow,
part of synchronizing factors are relatives to the
ongoing science. Considering some properly
applied methodology used during the separation
of species and sexual dimorphs case; it’s become

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 283
a standard tool of quantitative description
process of classification. In fact, multivariate
methods are based on the more satisfactory
premise that an organism grows and evolves
as a set of interacting parts all together; these
interactions should be considered together, not
abstracted as pairs and do not consider how
energy has sleeted exact harbor niche.
In the Energy transaction mood, the
detection of interrelated character clusters,
reduces the high dimensionality of a system
to few interpretable directions of variation,
and eliminates redundant variables. In energy
stabilizing mood, they provide an objective
picture of phenetic differences among samples
and specify how the measured characters
produce these differences. In a dimensional
state, it is represented by some identical
factors from paleontology and petrology.
The importance of the energetic flow can be
gauged by its impact on ideas of life’s history
and it correspondence to extreme condition.
A functional and quantitative science of form
suggests that parallelism and convergence are
dominant well observed phenomena, not mere
taxonomic nuisances.
Fossils Reconstruction and Virtual reality
It is widely believed that fossils are
fundamentally important in the understanding
of a specific habitat and the environmental
based on the reconstructing scenarios via
different valuable methodologies. for example,
some researchers argued that fossils provide the
soundest basis for evolutionary classification.
Although the virtual reality and the 3D printing
of our groups of fossils have been reconstructed
based on using morphological or chemical
characters of extant Sri Lankan organisms
alone, it is often noted that fossils would have
been of great use in clarifying relationships
between population of some extinct species
and that type of conclusions are impressively
tenuous in their absence. Therefore, to ensure
the understanding of the relationship that this
extant organism use to have in their habitat
we are trying to introduce some technological
applications which support the development
actions of the 3D reconstruction of fossils.
Digitalization of Fossils
Digitalization of fossils means creating
a computer-understandable version of them.
In technical terms, it’s called 3D modeling.
During the past decade, a number of innovative
applications of 3D technology in multiple
domains have immerged. A case study from
“Frammix” is presented here to apply the state-
of-the-art computer and 3D technology for the
multidisciplinary field of paleontology.
Frammix is a technology solution provider
based in Sri Lanka who carries out research
and development on merging cutting-edge
computer technology for real-world cases. They
have developed a 3D scanning, 3D printing,
and 3D visualization technique for the field of
paleontology with the purpose of supporting
scientific research, formal education, and
informal outreach in this domain. Depending on
the application, different techniques are utilized
to create 3D models of physical fossils and they
can be categorized as follows.
Manual 3D Modeling
This is a simple and low-cost technique. The
3D model is created based on the measurements
and the scaled drawings of the physical object
or the fossil. 3D designing software is used to
create the 3D model manually based on the
measurements. This is a time and labor-intensive
process but no special equipment is needed.
The objects with complex shapes require a lot
of time and effort in order to create 3D models
using this technique (Fig. 98).
The surface scan of fossils is an easy and
relatively faster method of creating 3D models
of physical objects. This technique uses a light
beam projected on the surface of the object for
scanning. Mostly laser or LED light beam is
used as the light source. As in fig 98, a camera
or a sensor monitors how the light beam bends
according to the shape of the surface and
generates the 3D model using the software. The
software output the 3D model in .obj or .fbx
format.
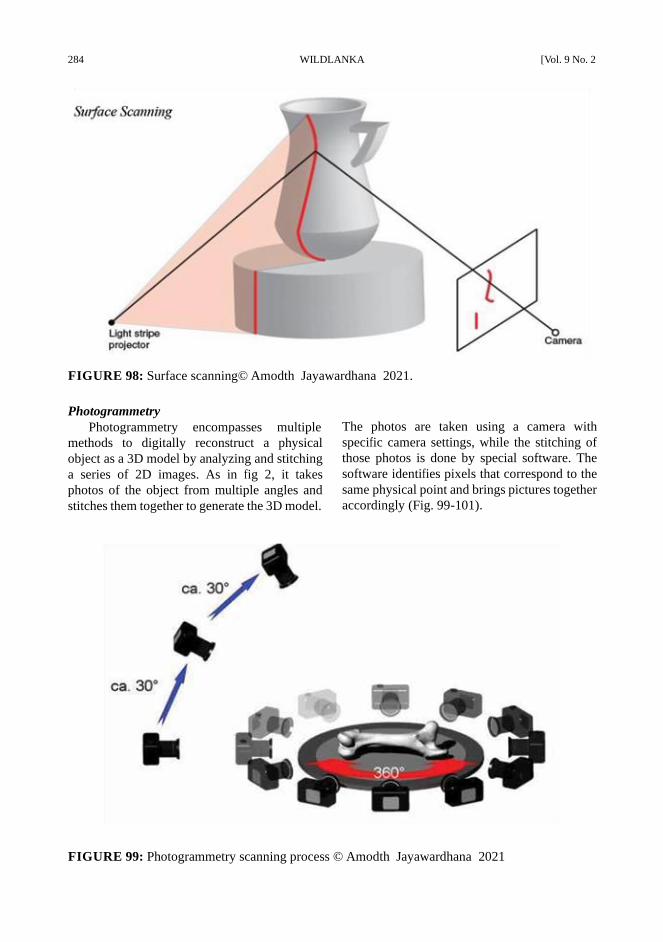
284 WILDLANKA [Vol. 9 No. 2
FIGURE 98: Surface scanning© Amodth Jayawardhana 2021.
Photogrammetry
Photogrammetry encompasses multiple
methods to digitally reconstruct a physical
object as a 3D model by analyzing and stitching
a series of 2D images. As in fig 2, it takes
photos of the object from multiple angles and
stitches them together to generate the 3D model.
The photos are taken using a camera with
specific camera settings, while the stitching of
those photos is done by special software. The
software identifies pixels that correspond to the
same physical point and brings pictures together
accordingly (Fig. 99-101).
FIGURE 99: Photogrammetry scanning process © Amodth Jayawardhana 2021

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 285
FIGURE 100: (A) An image of the actual fossil. (B). Orientations and the number of the images
taken of the fossil (C) A photo of the 3D model generated using photogrammetry© Michael
Ziegler 2020.
FIGURE 101: Photogrammetry processing for physical object (Left) for 3D modeling item (Right)
© Amodth Jayawardhana 2021.

286 WILDLANKA [Vol. 9 No. 2
Another unique benefit of Photogrammetry
is its capability of creating 3D models of objects
on a different scale. This becomes really useful
because objects ranging from micro-level,
like microfossils which are only visible under
electron microscope up to entire geologic
locations, can be digitalized based on this
technique. Because it uses photos of the object
to create the 3D model.
Figure 101 shows a 3D model generated
based on the Photogrammetry technique for a
fossil and Fig.4 shows a 3D model generated for
a physical replica of the lion statue in Yapahuwa
using the same technique.
Digital Fossil Repositories
Once the 3D models are created, they can be
made available for digital download through the
internet. That enables researchers all around the
world to get easy access to these digital versions
of fossils to carry out their studies. There are
online repositories where these 3D models are
digitally available. MorphoSource, Sketchfab,
myFOSSIL are few examples of online 3D
repositories.
Make Use of Digital Fossils
Once the fossils are digitized, they can
be easily shared digitally with anyone from
anywhere in the world. There are multiple ways
to make use of these digital fossils and they are
discussed in the following section.
3D printing
Having a physical replica of the fossil may
be beneficial sometimes for some studies than
the digital one. 3D printing is advantageous
for paleontologists who wish to replicate and
modify fossils. Specifically, printing enables
investigators to remove and reform fossils
from their matrix and permits the manipulation
of taphonomically distorted specimens.
Additionally, the ability to alter existing taxa or
create new, hypothetical forms allows the study
of non-existent morphologies, optimization,
and evolutionary constraint (Fig 102).
3D printing is the process of making three-
dimensional solid objects from a digital file. The
creation of a 3D printed object is achieved using
additive processes, where the object is created
by laying down successive layers of material
until the object is created. Each of these layers
can be seen as a thinly sliced cross-section of the
object. 3D printing is the opposite of subtractive
manufacturing which is cutting out / hollowing
out a piece of metal or plastic with for instance
a milling machine. This enables the production
of complex shapes using less material than
traditional manufacturing methods.
Once the model is ready for 3D printing, a
printing material must be chosen. There are a
variety of materials available for 3D printing
including plastics, steel, aluminum, sandstone,
thermoplastic polyurethane, photopolymer
resin, and composite powders. A special
machine is used for 3D printing.
Immersive technology is a state-of-the-art
computer technology which generates a distinct
experience by merging the physical world
with digital or simulated reality. As in fig 6.
Augmented Reality (AR), Virtual Reality (VR),
and Mixed Reality (MR) are the three main
Immersive Technologies (Fig 103).
Augmented Reality blends computer-
generated digital information into the user’s
real environment and enhances it. Whereas in
Virtual Reality, the user’s real environment gets
completely replaced with a simulated world.
Mixed Reality is quite similar to AR where it
also blends computer-generated information
onto the real-user environment, but the real-
user environment changes depending on user
interaction with the virtual objects. These
three realities together are called immersive
technologies.
June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 287
FIGURE 102: 3D printed model of T. rex | Eco Astronomy Inc- Reconstruction program in China.
Image © Majda Aouititen 2021.
Visualize with the use of Immersive Technologies
FIGURE 103: Immersive Technologies © Amodth Jayawardhana 2021.
Augmented Reality for Fossil Visualization
Frammix is carrying out research to find
the possibility of merging the immersive
technology with multiple domains. Based on
the findings of this research, an Augmented
Reality platform for Paleontology is developed.
It allows virtually the place and visualization
of real scale digitized 3D fossils in the real
environment using smartphones (Fig 104 -109).
As shown in Fig 104, digitized 3D fossils
can be virtually placed in real space and interact
with them through smartphones. This allows
paleontologists to study fossils as they are in
their own space.

288 WILDLANKA [Vol. 9 No. 2
FIGURE 104: Visualization of 3D models of dinosaur skeletons placed in the real environment
with use of AR technology © Amodth Jayawardhana 2021.
Teleport with Virtual Reality
The Augmented reality supplements the
real world with computer-generated contents to
create what we call a virtual environment that
we can navigate through. In this case, Virtual
Reality completely replaces the real world with
a simulated world based on two different tasks:
first “the cognitive process of determining
a path based on visual cues, knowledge of
the environment, and aids such as maps or
compasses”, and the second task is “the control
of the user’s viewpoint motion in the three-
dimensional environment” (Bowman et al.,
1998).
Unlike watching a video on a 2D screen,
with virtual reality, the viewer can be completely
teleported to a different environment while
providing an immersive experience. Travel is
“one of the most basic and universal interactions
found in VE applications”, so it received
considerable attention since the ‘90s (Bowman
et al., 1998; Slater et al., 1995; Bowman et al.,
1997; Templeman et al., 1999).
To achieve this task, a special device called
a VR headset or closed head-mounted display
needs to be used. Recently, new virtual reality
(VR) head-mounted displays (HMDs) for
consumers have been released, and different
authors started to study travel using these new
HMDs (Christou et al., 2017; Frommel et al.,
2017; Hashemian et al., 2017; Kitson et al.,
2017; Langbehn et al., 2018).
The advantage of teleport extends the
results found that showed a better performance
of teleport over visiting or watching the real
place or subject. Interestingly, by using this
technology we can both gain time and the high-
quality knowledge we are processing during the

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 289
experience we are conducting using Teleported
Virtual Reality.
Much more, using this technology will help
in the conservation process of the subject we
are proposing for our audience to discovered
through these devices, either we are representing
fossils in a virtual reality; or by making our
audience visit virtually the escalation place and
processing.
We are planning in the upcoming future
to use the Virtual Reality (VR) in our projects
of education (Fig. 105, Fig. 106, Fig. 107,
Fig. 108, Fig. 109) concerning specifically of
highlighting the Sri Lankan fossils. We estimate
that by using this technology, we can give the
opportunity for all those who are interested
to discover the Sri Lankan unique fossils,
nationally and internationally speaking. As well
by using the VR we will be sure that our fossils
will not be damaged during the lectures or
visits. That because we are presenting a virtual
version of these fossils that we have scanned
and reconstructed them in an augmented virtual
reality.
FIGURE 105: A virtual reality demo at the Communic Asia show at Marina Bay Sands on May 23.
Image © Desmond Koh.

290 WILDLANKA [Vol. 9 No. 2
FIGURE 106: Spinosaurus sp. | Synchronizing and floating trail of Augmented reality in Sri Lanka.
| Location: CCC | Image ©Aravinda Ravibhanu 2021.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 291
FIGURE 107: Tyrannosaurus rex. | Synchronizing and floating trail of Augmented reality in Sri
Lanka. | Location: CCC | Image ©Aravinda Ravibhanu 2021.

292 WILDLANKA [Vol. 9 No. 2
FIGURE 108: Dinosaur fighting. | Synchronizing and floating trail of Augmented reality in Sri
Lanka. | Location: CCC | Image ©Aravinda Ravibhanu 2021.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 293
FIGURE 108: Paluxysaurus sp | Synchronizing and floating trail of Augmented reality in Sri
Lanka. | Location: CCC | Image ©Aravinda Ravibhanu 2021.

294 WILDLANKA [Vol. 9 No. 2
DISCUSSION AND CONCLUSION
The last ice age ended about 12,500 to
10,300 years ago as it was reported by many
research papers based on strong facts and
evidence. However, the occurrence of these
changes cannot be certainly correlated in a way
or another with the Earth axis movements or
was related to the Earth’s precession. In fact, we
find it important to mention that the Earth’s axis
rotates (called processes) could be described
just as the rotation of a spinning top which has
the period of one complete cycle of precession
which is about 26,000 years, which means
approximately 1° every 72 years. Therefore,
geographically speaking the North Celestial
Pole will not always be pointing towards the
same star field, this precession is caused by the
gravitational pull of the Sun and the Moon on
the Earth axis. As a result, the earth’s precession
tends to stimulate the massive variables in
abiotic factors, especially during the period
of the LGM. Last Glacial Maximum (LGM)
coverage was between 26,500 and dominated
20,000-19,000 years ago. The sea level then fell
slowly until 21,000 and 16,000 yr B.P reaching
a maximum depth of about -130m (Katupotha et
al., 2021). According to our last study, sea-level
variations occurred mainly between 12,500 ±
1,500 YBP to 11,000± 1,500 YBP, indicating
that 25m ± 5m recorded to be the lower sea level
documented than the current sea level found
around the South Asian region. (Sumanarathna
et al., 2021).
Even in the paleoenvironment of the
Sabaragamuwa basin, it has been found to
support intensive habitation around 18ka and
12ka periods. The environmental fluctuations
in 18ka might be caused by the last glacial
maximum (LGM); in fact, during the12ka it
was partially affected by younger drya’s impact.
Hence, all of these events can open the doors
for extreme environmental conditions that could
trigger the extinction of many vulnerable species
that couldn’t adapt to these extreme conditions.
Hence, during the quaternary period, climate
changes resulted in the extinction of several
animals that have been proved through the
study of some fossils of this period that have
been fossilized in the alluvial beds. Much more,
this extinction wave did not stop at the end of
the Pleistocene, because the records show that
this extinction phenomenon has continued
due to the sea-level fluctuations, especially on
isolated islands in the Holocene epoch. Also,
the excavation data records of Batadomba-
lena (1982/2005) documents the existence of
the forager occupation of the rainforests of Sri
Lanka from around 40000 years ago until the
late Holocene. However, based on our records
(Fig. 92, Fig. 94, Fig. 95) of concrete roots by
the process of Rhizoconcretions were found
to represent a drought climate around a depth
of 60ft to 80ft from the current surface of the
Sabaragamuwa beds.
During our surveys, we have noticed
that the fossils abundance depth found in the
Sabaragamuwa bed is around 30ft to 200ft, and
we are currently working on a different set of
depts such as SEC -A: 20ft to 70ft, SEC -B: 70ft
to 120ft, SEC -C:120ft to 170ft, SEC -D: 170ft
to 220ft.
The continental drift theory explained that
during the Late Jurassic Period accrued Sri
Lanka was positioned within 67oS - 65oS and
32oE - 36oE in the southern hemisphere and
by the end of the Miocene Period Sri Lanka
location has changed to be between 4oN -
8oN and 77oE - 79oE situated in the northern
hemisphere (Katupotha, 2013 and 2020).
Due to these impressive facts, we found
that the Jurassic and the Miocene fossils from
Sri Lanka are very significant to be compared
with other fossils from different locations of the
world, and these types of fossils are very useful
to conduct studies of the evolutionary stages
through the climate changes of Sri Lanka.
Accordingly, we need to mention as well
that Sri Lanka has been subjected to Quaternary
glacial cycles due to the advancing and
retreating continental glaciers movements; due
as well to the warmer, the cooler, and the dry
climatic conditions; the evolution of hominids
and associated cultures was indeed part of
these cycles, and also the extinction of the
megafauna; the deposition of terrestrial and
marine sediments, and the development of soil
conditions (Sumanarathna et al., 2017).

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 295
Due to this evolutionary process, some
species’ ancestors of former geologic periods
were extinct, some were adapted and others
were newly evolved after an important nature
selection phenomenon.
The eminent groups of paleontologists,
zoologists, and also artists, such as Deraniyagala
P.E.P and Deraniyagala S.U that have
specialized in fauna and human fossils was been
the first scientists to study and document and to
represent the Indian subcontinent. They have
provided great data that we are using nowadays
to develop our knowledge based on their work
as they have discovered many unique fossils.
Considering as well the primary data
collected and recorded by our Research Unite Eco
Astronomy Inc and © SMKA, which presented
important details about the faunal habitat (with
human attachment) in Sabaragamuwa bed,
we found that the habitat fauna in this area
belongs to the families and species mentioned
as follow: Acavidae family, Pleuroceridae
family, Freshwater crabs (Prebrinckia sp. and
Ceylonthelphusa sp.), Cyprinidea family,
Bagridae family, Siluridea family, Clariidae
family, Heteropneustidae family, Channidea
family, Angulillidea family, Bufonidae family,
Trionychidae family, Geoemydidae family,
Testudinae family, Varanidae family, Agamidae
family, Gekkonodea family, Scincidae family,
Boidea family, Viperidea family, Phasianidea
family, Cercopithecidea family, Lorisidae
family, Pteropodidae family, Hipposiderdae
family, Pteromyidae family, Hystricidae family,
Muridae family, Leporidea family, Soricidea
family, Manidae family, Herpestidae family,
Mustelifae family, Viverridae family, Felidae
family,Canidae family, Ursidae family, Suidae
family, Ehephantidae family, Bovidae family,
Cervidae family, Tragulidae family (Perera,
2010 and 2011).
Due to our analysis of cross sections in a few
gem pits of Sabaragamuwa basin have shown,
the actual land form around the region is about
20m-30m below than today. Because the land is
heavily eroded, affected by extreme geological
and environmental conditions. Hence proper
survey via soil carbon or carbon sequestration
analysis will provide more informative data of
Sabaragamuwa-paleoenvironment. As a part of
studying the extreme environmental conditions
in SL, we are working on a few projects
regarding the Digital Conservation of samples
of fossils via AR\VR. Upgrading further
explorations and research in Sabaragamuwa
beds will provide a much more sustainable point
of view in conservation of Ratnapura fauna.
ACKNOWLEDGEMENT
We would like to express our special thanks
of gratitude to Ms. Uma Savindini Dissanayake
(Undergraduate at Physical Education, SUSL)
for assisting to develop the draft and documents.
As well as very special thanks to Mr. Adeepa
Nisal (Undergraduate at NSBM - Plymouth
University) and Mr. Prasanna Jayathilaka
(Research assistant at Eco Astronomy Inc) for
the outstanding support via reconstructing 3D
environments. Secondly, we would also like to
thank Mr Amodth Jayawardhana and Madura
Dissanayaka whom are AR\VR engineers at
Frammix for providing data for develop the
section Fossils Reconstruction and Virtual
reality.
Abbreviations used: ©SMKA – SM Kamal
Abyewardhana’s collection (1985- 2020).
REFERENCES
Adikari, A. G. M. (1998). Aspects of the
prehistory of the Sigiriya-Dambulla
region (Doctoral dissertation).
Bandaranayake, S. (1994). Further studies in the
settlement Archaeology of the Sigiriya-
Dambulla Region.
Bowman, D. A., D. Koller, and L. F. Hodges,
(1997). Travel in immersive virtual
environments: An evaluation of
viewpoint motion control technique.
In Proceedings of IEEE 1997 Annual
International Symposium on Virtual
Reality (pp. 45-52). IEEE.
Bowman, D. A., D. Koller, and L. F. Hodges,
(1998). A methodology for the evaluation
of travel techniques for immersive
virtual environments. Virtual reality,
3(2): 120-131.

296 WILDLANKA [Vol. 9 No. 2
Bucher, W. H. (1953). Fossils in metamorphic
rocks: a review. Geological Society of
America Bulletin, 64(3): 275-300.
Christou, C. G., and P. Aristidou, (2017).
Steering versus teleport locomotion for
head mounted displays. In International
conference on augmented reality, virtual
reality and computer graphics (pp. 431-
446). Springer, Cham.
Cooray P.G. and J. Katupotha, (1991).
Geological evolution of the coastal
zone of Sri Lanka. Proc., Symposium
on “Causes of Coastal Erosion in Sri
Lanka”. CCD/GTZ, Colombo, Sri
Lanka, 9-11, Feb. 1991, 5-26
Dávid, Árpád, (2011). The Book Paleontology,
Szerzői jog © 2011 EKF.
Deraniyagala P.E.P. (1914). Hypselephag hi/
esudricus Sinhaleyus , Spolia Zeylanica,
Vol. 24, pp. 45, 46 (type 100. Kuruvita,
type in Colombo Museum, paratype in
Brit. Mug).
Deraniyagala, P.E.P. (1936). Some vertebrate
fossils from Ceylon. Geological
Magazine, 73(7): 316-318.
Deraniyagala, P.E.P. (1936)., Palae.olocodorv
“namadicus sin.haleyucl, Asia, Soc.
(Ceylon) X,XXIIIE No. 88, fig. 2 and
Spolia zoylanica 8X, pl. VI.
Deraniyagala, P.E.P. (1936)., Rhinoceros
sinhalcyus Journal Royal Asiatic Society
(Ceylon vol. XXXIII, No. 88 (116).
Deraniyagala, P.E.P. (1937)., Rhinoceros
Sivalensis, Spolia Zeylanica, vol. XX, p.
196, Pl. VI.
Deraniyagala, P.E.P. (1937). Tiger or Lion,
Loris, Vol. I, No. 3, p. 147, fig. 2.
Deraniyagala, P. E. P. (1937). Some Miocene
and Upper Siwalik vertebrates from
Ceylon.
Deraniyagala, P. E. P. (1938). Some Fossil
Animals from Ceylon. Part II. The
Journal of the Ceylon Branch of the
Royal Asiatic Society of Great Britain &
Ireland, 34(91): 231-239.
Deraniyagala,P.E.P (1939)., Rhinoceros
sivalcnsis sinhaleyus, Journal Royal
Asiatic Society (Ceylon Branch,) vol.
XXXIV, No. 91, Fig. 2, Science and
Culture Vil, p. 94, Fig. 2. (116)
Deraniyagala, P. E. P. (1939). The Stone Age
and cave men of Ceylon. The Journal of
the Ceylon Branch of the Royal Asiatic
Society of Great Britain & Ireland,
34(92): 351-373.
Deraniyagala, P.E.P. (1939). Crocodyloidea,
Tetrapod Reptiles of Ceylon, vol 1, p,
310. (51).
Deraniyagala, P.E.P. (1942). Hupselephas
hysudricus SSP, Proc. Ind. Sci. Congress,
pt. 111. pp. 114, 115.
Deraniyagala, P.E.P. (1944). Hecaprotodon
Sinhaleyus, Spolia Zeylanica, vol. 24,
p. 37, Pl. Vr, text figs. .7—9, 10b and
c, 1946 Ibid, vol. 24, p. 168, pls. XXII
and XXIII text fig. 3, 41947 Journ.
Royal As. soc. (Ceylon Branch) vol. 37,
p. 230, pl. I, figs. 3—5, 1948 Ceylon
Administration Reports, part 4, July p. F.
8, 1955 Spolia Zeyla,nica, vol. 27, pt. Il,
pp. 223, 224, 1955 Geological Magazine
Vol. XCII No, 1 pp. 50—52 ; Some
Extinct Elephants, Their Relatives and
the Two Living Species, pp. 109, 110.
Deraniyagala, P. E. P. (1944). A new Cnemaspis
gecko from Ceylon. The Journal of the
Ceylon Branch of the Royal Asiatic
Society of Great Britain & Ireland,
36(97): 226-227.
Deraniyagala, P. E. P. (1945). Some odontoceti
from Ceylon. Spolia Zeylanica, 24(2):
112-120.
Deraniyagala, P. E. P. (1947). A new genus for
the Chinese alligator. In Proceedings of
the Third Annual Sessions of the Ceylon
Association of Science (Vol. 2, p. 12).
Deraniyagala, P. E. P. (1947). The names of the
Water monitors of Ceylon, the Nicobars
and Malaya. Proceedings of Third
annual session of Ceylon Association of
Science.
Deraniyagala, P.E.P. (1947). Sus Sinhaleyus,
Journal Royal Asiatio Soc. (Ceylon Br.)
Vol. XXXVII, NO. 104, p. 224, Pl. 1, 1,
2.

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 297
Deraniyagala, P. E. P. (1951). Elephas maximus,
the elephant of Ceylon (Vol. 1). National
Museums of Ceylon.
Deraniyagala, P. E. P. (1953). A Colored Atlas
of Some Vertebrates from Ceylon:
Tetrapod Reptilia. Volume Two. Ceylon
Government Press.
Deraniyagala, P. E. P. (1955). A Colored Atlas
of Some Vertebrates from Ceylon:
Serpentoid Reptilia (Vol. 3). Ceylon
Government Press.
Deraniyagala, P. E. P. (1955). Some Extinct
Éléphants, their Relatives and the Two
Living Species. Type Palaeoloøodon
Magtumoto 1925.
Deraniyagala, P.E.P. (1955). Crocodylus porosus
minikanna, spolia Zeylanicay Vol. 27, p,
277 (type in Colombo Museum Ceylon).
(22)
Deraniyagala, P. E. P. (1957). The races of
the Stone Age and of the Ferrolithic
of Ceylon. The Journal of the Ceylon
Branch of the Royal Asiatic Society of
Great Britain & Ireland, 5(1): 1-23.
Deraniyagala, P.E.P. (1957a). Homo sinhaleyus,
Journal Royal Society (Ceylon Brmch),
Vol. V, New Seriæ, p. 7. Holotype in
Ratnapura Museum.
Deraniyagala, P.E.P. (1957b). Üdupiyan
Udupiyan Cave and culture the upper
phase culture, Journal Royal Asiatic
eleven Soc. miles new south-west series,
Vol. of Diavinna V, pt. l, in p. the13
Balangoda district. Type collection in
Colombo National Mugeumo. (64).
Deraniyagala, P. E. P. (1958). Three new
cyprinoids, a new cat fish and variation
among some cyprinoids and an
anabantoid of Ceylon.
Deraniyagala, P. E. P. (1958). The Pleistocene
of Ceylon. (Ceylon: Department of
National Museums.; Natural history
series. ed.). Ceylon, Printed at the Govt.
Press.
Deraniyagala, P. E. P. (1963). Prehistoric
archaeology in Ceylon. Asian
Perspectives, 7(1/2): 189-192.
Deraniyagala, S. U. (1988). The prehistory of
Sri Lanka: an ecological perspective.
Harvard University.
Deraniyagala, S. U. (1992). The Prehistory of
Sri Lanka. Department of Archaeological
Survey.
Dissanayake, C. B., and M. S. Rupasinghe,
(1995). Classification of gem deposits
of Sri Lanka. Geologie en Mijnbouw,
74:79-79.
Dissanayake, C. B., and R. Chandrajith,
(1999). Sri Lanka–Madagascar
Gondwana Linkage: Evidence for a Pan‐
African Mineral Belt. The Journal of
Geology, 107(2): 223–235. https://doi.
org/10.1086/314342
Dutta, S. K., and P. Mohanty-Hejmadi, (1976).
Breeding and life history of the Indian
bull frog, Rana tigerina (Daudin).
Prakruti Utkal Univ. J. Sci, 13(1&3):
51-59.
Emmel, B., F. Lisker, and T. Hewawasam,
(2012). Thermochronological dating
of brittle structures in basement rocks:
A case study from the onshore passive
margin of SW Sri Lanka. Journal
of Geophysical Research: Solid
Earth, 117(B10): 1–17. https://doi.
org/10.1029/2012jb009136
Embery, G., A.C. Milner, R.J. Waddington, R.C.
Hall, M.S. Langley, and A.M. Milan,
(2003). Identification of proteinaceous
material in the bone of the dinosaur
Iguanodon. Connective tissue research,
44(1): 41-46.
Falconer, H. (1859). Descriptive Catalogue of
the Fossil Remains of Vertebrata from
the Sewalik Hills, the Nerbudda, Perim
Island, &c. in the Museum of the Asiatic
Society of Bengal. CB Lewis.
Flynn, L. J., D. Pilbeam, J. C. Barry, M. E.
Morgan, and S.M. Raza, (2016). Siwalik
synopsis: A long stratigraphic sequence
for the later Cenozoic of South Asia.
Comptes Rendus Palevol, 15(7): 877-
887.
Franz, G., V. Mosbruggr, and R. Menge,
(1991). Carbo-Permian pteridophyll leaf

298 WILDLANKA [Vol. 9 No. 2
fragments from an amphibolite facies
basement, Tauern Window, Austria.
Terra Nova, 3(2): 137-141.
Frommel, J., S. Sonntag and M. Weber, (2017).
Effects of controller-based locomotion
on player experience in a virtual reality
exploration game. In Proceedings of the
12th international conference on the
foundations of digital games (pp. 1-6).
Grimaldi, D. (2009). Pushing back amber
production. Science, 326(5949): 51-52.
Hashemian, A. M., and B. E. Riecke, (2017).
Leaning-based 360 interfaces:
Investigating virtual reality navigation
interfaces with leaning-based-translation
and full-rotation. In International
Conference on Virtual, Augmented and
Mixed Reality (pp. 15-32). Springer,
Cham.
Hemmer, H. (1987). The phylogeny of the tiger
(Panthera tigris). Tigers of the World,
28-35.
Hendavitharana, W., S. Dissanayake, M. De
Silva and C. Santiapillai, (1994). The
survey of elephants in Sri Lanka. Gajah,
12: 1-30.
Hill, M. L. (1985). Remarkable fossil locality:
Crinoid stems from migmatite of
the Coast Plutonic Complex, British
Columbia. Geology, 13(11): 825-826.
https://doi.org/10.1016/j.lithos.2019.04.016
Ingicco, T., J. de Vos, and O. F. Huffman, (2014).
The oldest gibbon fossil (Hylobatidae)
from insular Southeast Asia: Evidence
from Trinil,(East Java, Indonesia),
Lower/Middle Pleistocene. PloS one,
9(6), e99531.
Katupotha, J. (1994). Quaternary Research
in Sri Lanka. In: Recent Advances in
the Earth Sciences of Sri Lanka - P.W.
Vitanage, Felicitation Volume. Jour of
the Geol. Soc. of Sri Lanka, (5): 141-
152, 1994
Katupotha, J. (1995): Evolution and geological
significance of Holocene emerged shell
beds on the southern coastal zone of Sri
Lanka. Journal of Coastal Research, 11
(4): 1042-1061.
Katupotha, J. and P. Dias, (2001). The Geological
Evolution Correlated to the Stratigraphy
of theKalpitiya Peninsula. Journal of
Indian Association of Sedimentalogists.
20(1): 21-37.
Katupotha, J. (2013). Palaeoclimate change
during Glacial Periods: Evidence from
Sri Lanka Katupotha /Journal of Tropical
Forestry and Environment Vol. 3: No. 01
(2013) 42-54
Katupotha, J. (2020). Wandering of Sri Lanka
from Precambrian to Recent. Wildlanka,
8(3): pp. 128 - 151: 2020. Copyright 2020
Department of Wildlife Conservation,
Sri Lanka.
Katupotha, J., A.R. Sumanarathna and Sachich
Gamage, (2021). LAST GLACIAL
MAXIMUM, THE FLANDRIAN
TRANSGRESSION AND THE
CONFIGURATION OF SRI LANKA
AND ITS SURROUNDINGS.
Department of Wildlife Conservation,
Proceedings of the Annual Session -2020,
Abstractshttps://www.researchgate.net/
publication/353347992.
Kitchener, A. C., and A. J. Dugmore, (2000).
Biogeographical change in the tiger,
Panthera tigris. In Animal Conservation
forum, 3(2): pp. 113-124). Cambridge
University Press.
Kitson, A., A. M. Hashemian, E. R. Stepanova,
E. Kruijff, and B.E. Riecke, (2017).
Comparing leaning-based motion cueing
interfaces for virtual reality locomotion.
In 2017 IEEE Symposium on 3d user
interfaces (3DUI) (pp. 73-82). IEEE.
Knox, R. (2016). A historical relation of the
Island Ceylon in the East Indies (1681).
In English Siege and Prison Writings
(pp. 19-43). Routledge India.
Laborda-Lopez, C., J. Aguirre, and S.K.
Donovan, (2015). Surviving
metamorphism: taphonomy of fossil
assemblages in marble and calc-silicate
schist. Palaios, 30(9): 668-679.
Langbehn, E., P. Lubos, and F. Steinicke, (2018).
Evaluation of locomotion techniques for
room-scale vr: Joystick, teleportation,
and redirected walking. In Proceedings

June, 2021] FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN 299
of the Virtual Reality International
Conference-Laval Virtual (pp. 1-9).
Manamendra-Arachchi, K., R. Pethiyagoda, R.
Dissanayake and M. Meegaskumbura.
2005. A second extinct big cat from the
late Quaternary of Sri Lanka. In: Yeo, D.
C. J., K. L. Ng and R. Pethiyagoda (eds.),
Contributions to biodiversity exploration
and research in Sri Lanka. The Rafles
Bulletin of Zoology, Supplement No. 12:
423-434 pp.
Manamendra-Arachchi, K. (2010). The extinct
big cats (felidae) of Sri Lanka. Sirinimal
Lakdusinghe felicitation volume, (eds.
Gunawardhana, P., G. Adikari and R. A.
E. Coningham), Neptune Publication,
137-143 pp.
Mazak, V. (1981). Panthera tigris. Mammalian
species, (152), 1-8.
Perera, H. N. (2010). Prehistoric Sri Lanka:
Late Pleistocene rockshelters and an
open-air site. Archaeopress.
Perera, N., N. Kourampas, I. A. Simpson,
S. U. Deraniyagala, D. Bulbeck, J.
Kamminga, J. Perera, D. Q. Fuller, K.
Szabó, and N.V. Oliveira, (2011). People
of the ancient rainforest: late Pleistocene
foragers at the Batadomba-lena
rockshelter, Sri Lanka. Journal of human
evolution, 61(3): 254–269. https://doi.
org/10.1016/j.jhevol.2011.04.001
Pilgrim, G. E. (1931). Pontian Carnivora
of Europe. British Museum Natural
History, London.
Pocock, R. I. (1930). The panthers and ounces
of Asia.
Prasad, K. N. (1996). Pleistocene cave fauna
from peninsular India. Journal of Caves
and Karst Studies, 58: 30-34.
Premathilake, R., and J. Risberg, (2003). Late
Quaternary climate history of the Horton
Plains, central Sri Lanka. Quaternary
Science Reviews, 22(14): 1525–
1541. https://doi.org/10.1016/s0277-
3791(03)00128-8
Prothero, D. R. (2013). Bringing fossils to
life: An introduction to paleobiology.
Columbia University Press.
Roberts, P., N. Perera, O, Wedage, S.
Deraniyagala, J. Perera, S. Eregama,
A. Gledhill, M.D. Petraglia, and J.A.
Lee-Thorp, (2015). Direct evidence for
human reliance on rainforest resources in
late Pleistocene Sri Lanka. Science (New
York, N.Y.), 347(6227): 1246–1249.
https://doi.org/10.1126/science.aaa1230
Scaioni, M., M. Marsella, M. Crosetto, V.
Tornatore, and J. Wang, (2018).
Geodetic and remote-sensing sensors for
dam deformation monitoring. Sensors,
18(11): 3682.
Schweitzer, M. H., J. L. Wittmeyer, J. R.,
Horner and J. K. Toporski, (2005). Soft-
tissue vessels and cellular preservation in
Tyrannosaurus rex. Science, 307(5717):
1952-1955.
Schweitzer, M.H., W. Zheng, T.P. Cleland,
M.B. Goodwin, E. Boatman, E., Theil.
and S.C. Fakra, (2014). A role for iron
and oxygen chemistry in preserving
soft tissues, cells and molecules
from deep time. Proceedings of the
Royal Society B: Biological Sciences,
281(1775):20132741.
Shaw, C. S. (2019). Metamorphic mineral
assemblages in fossils in the contact
metamorphic rocks of the Portrush Sill,
Northern Ireland. Lithos, 338–339, 99–
110.
Shute, C.H., and C.J. Cleal, (1987). Palaeobotany
in museums. The geological curator,
4(9): 553-559.
Slater, M., M. Usoh, M., and A. Steed, (1995).
Taking steps: the influence of a walking
technique on presence in virtual reality.
ACM Transactions on Computer-Human
Interaction (TOCHI), 2(3): 201-219.
Sumanarathna, A.R., (2021, September 14).
An Introduction to Paleontology and
Fossils (mdIdKN+; úoHdj" f*dis, iy f*dis,lrKh) [Slides]. Eco Astronomy
Sri Lanka. https://www.researchgate.
ne t /pu bl i ca t ion /35 449 29 62_ An _
Introduction_to_Paleontology_and_
Fossils

300 WILDLANKA [Vol. 9 No. 2
Sumanarathna, A.R, J. Katupotha, and
M. Aouititen, (2020). Comparative
Systematic Analysis of Milankovitch
Cycles to Identify Variations of Glaciers
and Interglacial Periods of Late
Pleistocene in South Asia. TRIVALENT
;%sixhqc: Journal of Archaeology,
Tourism & Anthropology, 1(1): 1–12.
https://doi.org/10.4038/tjata.v1i1.24
Sumanarathna, A.R., (2020). Dynamics
of Himachal Pradesh: Upgrading
Prometheus of Paleo Himachal.
Ice Stupa Project, The Trail Report
-Section one Eco Astronomy Sri
Lanka. https://www.researchgate.net/
publication/340087847_Dynamics_
of_Himachal_Pradesh_Upgrading_
Prometheus_of_Paleo_Himachal /
citations
Sumanarathna, A. R. (2019). Comparative
Systematic Analysis of Proxy to
Indicate Younger Dryas Cooling in Late
Pleistocene in Sri Lanka. Retrieved
December 10, 2019.
Sumanarathna, A.R., B., Madurapperuma,
J. Kuruppuarachchi, et al. (2016).
Morphological Variation and Speciation
of Acavidae Family: A Case Study
from Fossil and Living Species of
Batadombalena Cave Pre-Historic Site
in Sri Lanka. Annals of Valahi University
of Targoviste, Geographical Series,
16(2): pp. 59-68. Retrieved 7 Nov. 2016,
from doi:10.1515/avutgs-2016-0005
Sumanarathna, A.R., et al., (2017),
EXTINCTION OF QUATERNARY
MAMMALIAN HABITATS OF
MEGAFAUNA IN SABARAGAMU
BASIN, SRI LANKA, Journal
of Eco Astronomy 01(01): PP
16- 31 https://www.researchgate.
n e t / p u b l i c a t i o n / 3 2 3 4 1 4 3 1 7 _
EXTINCTION_OF_QUATERNARY_
MAMMALIAN_HABITATS_OF_
MEGAFAUNA_IN_SABARAGAMU_
BASIN_SRI_LANKA
Sumanarathna, A.R. (2018). Union of General
Theory - Eco Astronomy Mechanics &
Concepts. Journal of Astronomy.
Taylor, P. D. (1990). Preservation of soft-bodied
and other organisms by bioimmuration―a
review. Palaeontology, 33(1): 1-17.
Templeman, J. N., P. S. Denbrook, and L.E.
Sibert, (1999). Virtual locomotion:
Walking in place through virtual
environments. Presence, 8(6): 598-617.
Tong, H., and A.M. Moigne, (2000). Quaternary
rhinoceros of China. Acta anthropologica
sinica, 19(supp.), 257-263.
Wedage, O., A. Picin, J. Blinkhorn, K. Douka, S.
Deraniyagala, N. Kourampas, N. Perera,
I. Simpson, N. Boivin, M. Petraglia, and
P. Roberts, (2019). Microliths in the
South Asian rainforest ~45-4 ka: New
insights from Fa-Hien Lena Cave, Sri
Lanka. PloS one, 14(10): e0222606.
h t tp s : / / d o i . o rg /1 0 .1 3 71/ jo u rn a l .
pone.0222606
Wilson, M. A., T. J. Palmer, and P.D. Taylor,
(1994). Earliest preservation of soft‐
bodied fossils by epibiont bioimmuration:
Upper Ordovician of Kentucky. Lethaia,
27(3): 269-270.
Wright, A. J., and E.D. Ghent, (1973). A
metamorphosed coral in an olivine-
bearing hornfels from New South Wales.
Journal of the Geological Society of
Australia, 20(1): 79-84.
Zylberberg, L., and M. Laurin, (2011).
Analysis of fossil bone organic matrix
by transmission electron microscopy.
Comptes Rendus Palevol, 10(5-6): 357-
366.
Received Date : 12th March 2021
Accepted Date : 20th May 2021
Cite this article as
Suamanarathna, A. R., Abewardhana, K.,
Katupotha, J., & Aouititen, M. (2021).
FOSSILS OF SRI LANKA: CHAPTER SABARAGAMUWA BASIN. Journal of
Wild Lanka, 09(02), 173–300.




















