
Untitled attachment 00023
7
Strain and elongation of the human semitendinosus muscle – Tendon unit Eleftherios Kellis ⇑ , Glykeria Patsika, Evaggelos Karagiannidis Laboratory of Neuromechanics, Department of Physical Education and Sport Sciences at Serres, Aristotle University of Thessaloniki, Greece article info Article history: Received 23 February 2013 Received in revised form 4 July 2013 Accepted 15 July 2013 Available online xxxx Keywords: Ultrasonography Tendons Connective tissue Isometric contraction Skeletal muscles abstract The semitendinosus (ST) consists of a long distal tendon and it is divided in two parts by a tendinous inscription (TI). The purpose of this study was to quantify strain and elongation of the TI and the distal tendon of ST. Fourteen subjects performed ramp isometric contractions of the knee flexors at 0°, 45° and 90° of knee flexion. Two ultrasound probes were used to visualize the displacement of the distal ten- don and selected points across the TI and aponeuroses. Three-way analysis of variance designs indicated that: (a) strain and elongation of the ST distal muscle–tendon junction were higher than that of the apo- neurosis – TI junction points (p < 0.05) (b) the long arm of the TI reach strain of 49.86 ± 7.77% which was significantly (p < 0.05) higher than that displayed by the short arm (28.35 ± 0.59%) (c) Strain of tendinous and TI-aponeuroses segments significantly increased from 90° to 0° of knee flexion while the inverse was observed for the TI arm length (p < 0.05). (d) Tendon strain was significantly higher than strain of the TI- aponeuroses segments at 45° and 90° of knee flexion while the opposite was observed at 0° of knee flexion. The arrangement of TI along ST length results in differential local strains, indicating that the mechanical properties of the ST muscle are affected by tendon, aponeuroses and tendinous inscription interactions. Ó 2013 Elsevier Ltd. All rights reserved. 1. Introduction Strain and elongation of muscle fibers and tendons have an important effect on the force–velocity and force–length properties of the muscle–tendon unit. Previous studies examined the mechan- ical properties of pennated muscles such as the gastrocnemious, the tibialis anterior, the vastus lateralis or the soleus (Muramatsu et al., 2001; Bojsen-Moller et al., 2003; Arampatzis et al., 2005; De Monte et al., 2006; Magnusson et al., 2008). Therefore any conclusions regarding muscle–tendon behavior are mainly applicable to this type of muscle. However, all human muscles do not display the same architecture. The semitendinosus is characterized by long fibers, small force generation and high excursion capacity (ST) (Woodley and Mercer, 2005; Kellis et al., 2010). Most information on ST tendon mechan- ical properties is based on in vitro experiments with maximum strains ranging from 8% to 14% (Noyes et al., 1984; Lieber et al., 1991; Piedade et al., 2006; Abramowitch et al., 2010). To our knowledge, no studies have quantified strain of the ST tendon in vivo. Strain appears to vary along the length of the muscle–tendon unit; however, the direction of the difference varies among studies; some studies have reported that tendon strain is lower (Lieber et al., 1991; Maganaris and Paul, 2000), higher (Finni et al., 2003; Magnusson et al., 2003; Kongsgaard et al., 2011) or similar (Mura- matsu et al., 2001; Arampatzis et al., 2005) compared with aponeu- rosis strain. Lieber et al. (1991) reported strain values for the frog ST ranging from 2% (tendon) to 8% (aponeurosis). Whether these observations are valid for human ST tissue is not clear. The mechanical properties of muscle–tendon units have been examined when muscle length changes passively as well as during active contractions (Finni, 2006; Magnusson et al., 2008). In vivo passive change in muscle length is achieved by changing joint angular position. From an anatomical point of view, when the hip joint is maintained constant, the ST muscle lengthens as the knee extends. Previous research studies have shown that even small changes in angular position have a varying effect on strain and elongation of the aponeurosis and the tendon (Muramatsu et al., 2001; Bojsen-Moller et al., 2003; Arampatzis et al., 2005; De Monte et al., 2006). Consequently, ST mechanical properties during active contraction may differ depending on joint angular position (initial muscle length) used for testing. A factor that may affect strain along the ST muscle–tendon unit is the presence of a tendinous inscription (TI) which divides the muscle in two compartments. The TI has two ‘‘arms’’, a lateral 1050-6411/$ - see front matter Ó 2013 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.jelekin.2013.07.016 ⇑ Corresponding author. Address: Laboratory of Neuromechanics, Aristotle University of Thessaloniki, TEFAA Serres, Serres 62100, Greece. Tel./fax: +30 2310 991053. E-mail address: [email protected] (E. Kellis). Journal of Electromyography and Kinesiology xxx (2013) xxx–xxx Contents lists available at ScienceDirect Journal of Electromyography and Kinesiology journal homepage: www.elsevier.com/locate/jelekin Please cite this article in press as: Kellis E et al. Strain and elongation of the human semitendinosus muscle – Tendon unit. J Electromyogr Kinesiol (2013), http://dx.doi.org/10.1016/j.jelekin.2013.07.016
Transcript of Untitled attachment 00023

Journal of Electromyography and Kinesiology xxx (2013) xxx–xxx
Contents lists available at ScienceDirect
Journal of Electromyography and Kinesiology
journal homepage: www.elsevier .com/locate / je lek in
Strain and elongation of the human semitendinosus muscle – Tendonunit
1050-6411/$ - see front matter � 2013 Elsevier Ltd. All rights reserved.http://dx.doi.org/10.1016/j.jelekin.2013.07.016
⇑ Corresponding author. Address: Laboratory of Neuromechanics, AristotleUniversity of Thessaloniki, TEFAA Serres, Serres 62100, Greece. Tel./fax: +30 2310991053.
E-mail address: [email protected] (E. Kellis).
Please cite this article in press as: Kellis E et al. Strain and elongation of the human semitendinosus muscle – Tendon unit. J Electromyogr Kinesiolhttp://dx.doi.org/10.1016/j.jelekin.2013.07.016
Eleftherios Kellis ⇑, Glykeria Patsika, Evaggelos KaragiannidisLaboratory of Neuromechanics, Department of Physical Education and Sport Sciences at Serres, Aristotle University of Thessaloniki, Greece
a r t i c l e i n f o a b s t r a c t
Article history:Received 23 February 2013Received in revised form 4 July 2013Accepted 15 July 2013Available online xxxx
Keywords:UltrasonographyTendonsConnective tissueIsometric contractionSkeletal muscles
The semitendinosus (ST) consists of a long distal tendon and it is divided in two parts by a tendinousinscription (TI). The purpose of this study was to quantify strain and elongation of the TI and the distaltendon of ST. Fourteen subjects performed ramp isometric contractions of the knee flexors at 0�, 45�and 90� of knee flexion. Two ultrasound probes were used to visualize the displacement of the distal ten-don and selected points across the TI and aponeuroses. Three-way analysis of variance designs indicatedthat: (a) strain and elongation of the ST distal muscle–tendon junction were higher than that of the apo-neurosis – TI junction points (p < 0.05) (b) the long arm of the TI reach strain of 49.86 ± 7.77% which wassignificantly (p < 0.05) higher than that displayed by the short arm (28.35 ± 0.59%) (c) Strain of tendinousand TI-aponeuroses segments significantly increased from 90� to 0� of knee flexion while the inverse wasobserved for the TI arm length (p < 0.05). (d) Tendon strain was significantly higher than strain of the TI-aponeuroses segments at 45� and 90� of knee flexion while the opposite was observed at 0� of kneeflexion. The arrangement of TI along ST length results in differential local strains, indicating that themechanical properties of the ST muscle are affected by tendon, aponeuroses and tendinous inscriptioninteractions.
� 2013 Elsevier Ltd. All rights reserved.
1. Introduction
Strain and elongation of muscle fibers and tendons have animportant effect on the force–velocity and force–length propertiesof the muscle–tendon unit. Previous studies examined the mechan-ical properties of pennated muscles such as the gastrocnemious, thetibialis anterior, the vastus lateralis or the soleus (Muramatsu et al.,2001; Bojsen-Moller et al., 2003; Arampatzis et al., 2005; De Monteet al., 2006; Magnusson et al., 2008). Therefore any conclusionsregarding muscle–tendon behavior are mainly applicable to thistype of muscle. However, all human muscles do not display thesame architecture.
The semitendinosus is characterized by long fibers, small forcegeneration and high excursion capacity (ST) (Woodley and Mercer,2005; Kellis et al., 2010). Most information on ST tendon mechan-ical properties is based on in vitro experiments with maximumstrains ranging from 8% to 14% (Noyes et al., 1984; Lieber et al.,1991; Piedade et al., 2006; Abramowitch et al., 2010). To ourknowledge, no studies have quantified strain of the ST tendonin vivo.
Strain appears to vary along the length of the muscle–tendonunit; however, the direction of the difference varies among studies;some studies have reported that tendon strain is lower (Lieberet al., 1991; Maganaris and Paul, 2000), higher (Finni et al., 2003;Magnusson et al., 2003; Kongsgaard et al., 2011) or similar (Mura-matsu et al., 2001; Arampatzis et al., 2005) compared with aponeu-rosis strain. Lieber et al. (1991) reported strain values for the frogST ranging from 2% (tendon) to 8% (aponeurosis). Whether theseobservations are valid for human ST tissue is not clear.
The mechanical properties of muscle–tendon units have beenexamined when muscle length changes passively as well as duringactive contractions (Finni, 2006; Magnusson et al., 2008). In vivopassive change in muscle length is achieved by changing jointangular position. From an anatomical point of view, when thehip joint is maintained constant, the ST muscle lengthens as theknee extends. Previous research studies have shown that evensmall changes in angular position have a varying effect on strainand elongation of the aponeurosis and the tendon (Muramatsuet al., 2001; Bojsen-Moller et al., 2003; Arampatzis et al., 2005;De Monte et al., 2006). Consequently, ST mechanical propertiesduring active contraction may differ depending on joint angularposition (initial muscle length) used for testing.
A factor that may affect strain along the ST muscle–tendon unitis the presence of a tendinous inscription (TI) which divides themuscle in two compartments. The TI has two ‘‘arms’’, a lateral
(2013),
2 E. Kellis et al. / Journal of Electromyography and Kinesiology xxx (2013) xxx–xxx
and a medial one. Recent evidence indicates that local geometrybetween this inscription and muscle fascicles affects the way mus-cle contracts, as the longer (lateral) arm is elongated more com-pared with the shorter (medial) one (Kellis et al., 2012a,b).However, neither strain of the aponeurosis nor ST tendon elonga-tion has been examined in humans in vivo.
There is evidence that the presence of internal tendinous tissueresults in non-uniform strains along the muscle tendon unit(Pappas et al., 2002; Fiorentino et al., 2012). It is not clear whetherthis affects the susceptibility of this particular muscle to injuries.Nevertheless, the ST muscle is the only component of the ham-strings which displays a TI but it is rarely ruptured as opposed tothe rest hamstrings muscles (Koulouris and Connell, 2005). Exam-ination of the mechanical properties of the fusiform muscles, suchas the ST, may provide useful information regarding their function.For the ST muscle, in particular, the presence of the TI may affectwhole muscle–tendon behavior. Therefore, the purpose of thisstudy was to compare strain and elongation between (a) differentareas of the TI-aponeuroses intersection and the muscle–tendonjunction (b) the long and the short arm of the TI of the human STmuscle c) different angular positions of the knee joint.
2. Methods
Seven males and seven females (age 21.37 ± 1.48 years; mass75.8 ± 3.82 kg; height 1.77 ± 0.03 m) were stabilized on a Cybex(Humac Norm, CSMI, MA) dynamometer in the prone positionand performed 5 sub-maximal (warm up) and 3 maximum isomet-ric flexion contractions (MVCs) at 0�, 45� and 90� knee flexion an-gles in a randomized order. After 10 min of rest, three 10-s rampisometric contractions guided by an audiovisual signal were per-formed. Dynamometer moment and knee joint angular position(using a twin – axis goniometer, Model TSD 130B, Biopac Systems,Inc., Goleta, CA) were simultaneously recorded at 1000 Hz. The mo-ment of force was corrected for the effects of gravity whilst goni-ometer data were used to have a more precise angular positiondata due to axes alignment errors.
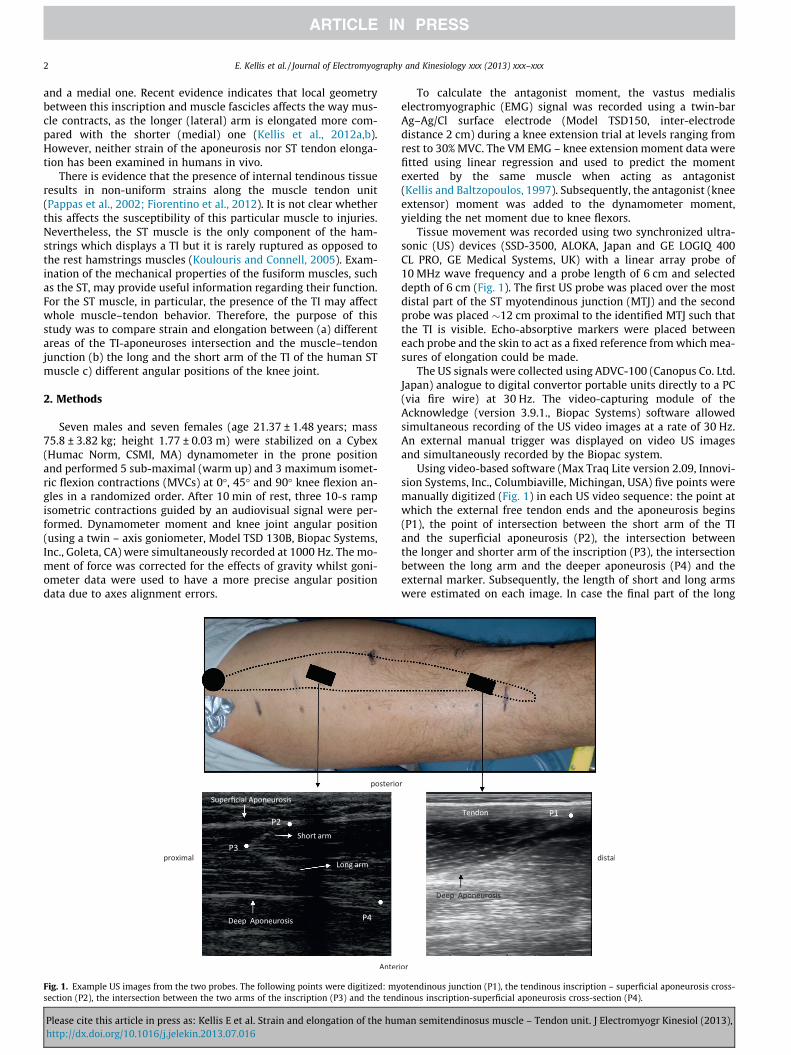
Fig. 1. Example US images from the two probes. The following points were digitized: mysection (P2), the intersection between the two arms of the inscription (P3) and the tend
Please cite this article in press as: Kellis E et al. Strain and elongation of the humhttp://dx.doi.org/10.1016/j.jelekin.2013.07.016
To calculate the antagonist moment, the vastus medialiselectromyographic (EMG) signal was recorded using a twin-barAg–Ag/Cl surface electrode (Model TSD150, inter-electrodedistance 2 cm) during a knee extension trial at levels ranging fromrest to 30% MVC. The VM EMG – knee extension moment data werefitted using linear regression and used to predict the momentexerted by the same muscle when acting as antagonist(Kellis and Baltzopoulos, 1997). Subsequently, the antagonist (kneeextensor) moment was added to the dynamometer moment,yielding the net moment due to knee flexors.
Tissue movement was recorded using two synchronized ultra-sonic (US) devices (SSD-3500, ALOKA, Japan and GE LOGIQ 400CL PRO, GE Medical Systems, UK) with a linear array probe of10 MHz wave frequency and a probe length of 6 cm and selecteddepth of 6 cm (Fig. 1). The first US probe was placed over the mostdistal part of the ST myotendinous junction (MTJ) and the secondprobe was placed �12 cm proximal to the identified MTJ such thatthe TI is visible. Echo-absorptive markers were placed betweeneach probe and the skin to act as a fixed reference from which mea-sures of elongation could be made.
The US signals were collected using ADVC-100 (Canopus Co. Ltd.Japan) analogue to digital convertor portable units directly to a PC(via fire wire) at 30 Hz. The video-capturing module of theAcknowledge (version 3.9.1., Biopac Systems) software allowedsimultaneous recording of the US video images at a rate of 30 Hz.An external manual trigger was displayed on video US imagesand simultaneously recorded by the Biopac system.
Using video-based software (Max Traq Lite version 2.09, Innovi-sion Systems, Inc., Columbiaville, Michingan, USA) five points weremanually digitized (Fig. 1) in each US video sequence: the point atwhich the external free tendon ends and the aponeurosis begins(P1), the point of intersection between the short arm of the TIand the superficial aponeurosis (P2), the intersection betweenthe longer and shorter arm of the inscription (P3), the intersectionbetween the long arm and the deeper aponeurosis (P4) and theexternal marker. Subsequently, the length of short and long armswere estimated on each image. In case the final part of the long
otendinous junction (P1), the tendinous inscription – superficial aponeurosis cross-inous inscription-superficial aponeurosis cross-section (P4).
an semitendinosus muscle – Tendon unit. J Electromyogr Kinesiol (2013),

E. Kellis et al. / Journal of Electromyography and Kinesiology xxx (2013) xxx–xxx 3
arm exceeded the field of view, its length was predicted using sim-ple trigonometry based on muscle thickness, the angle of the visi-ble part of the long arm with the aponeurosis and the position ofthe deep aponeurosis (Kellis et al., 2012a,b). Validity and reliabilityof the US measurements have been reported elsewhere (Kelliset al., 2009; Kellis et al., 2012a,b). In addition, two independent tes-ters have digitized 5 times, each video sequence during ramp con-tractions. The intraclass correlation coefficients were 0.85 (P1),0.93 (P2), 0.94 (P3) and 0.91 (P4) indicating acceptable reliability.
To measure elongation due to knee joint rotation additional USfootages during a passive test (from approximately 110� of kneeflexion to about 3� hyper-extension) immediately after the ramptrials was performed. Each point was then digitized during the pas-sive range of motion (Muramatsu et al., 2001). Subsequently, thedisplacement of each point during the ramping trial was calculatedas the difference of the measured and the passive (due to jointrotation) displacement of the same point at the same angularposition.
Resting length was measured at 40� of knee flexion (where thepassive moment is almost zero (Silder et al., 2007)) as the curvedpath from the ST distal origin (pes anserinus) to each of the mark-ers measured along the skin surface (Kellis et al., 2009). Kellis et al.(2009) reported good agreement between US measurement of STlength properties with cadaveric data. However, they measuredST tendon length by using a non-flexible tape from the origin toST muscle tendon junction. In the present study, to improve accu-racy further, the ST origin was identified using US and it wasmarked on the skin. Then, a flexible measurable tape was used toalong the path of the ST until the skin markers corresponding toP1, P2, P3 and P4 markers. The distance of each marker from theorigin was 184.30 mm (P1), 271.66 mm (P2), 285.21 mm (P3) and258.11 mm (P4).
For the purposes of this study, three sets of data were furtheranalyzed. First, absolute displacement at all testing conditionswere compared between the four points. Presentation of absolutedisplacement values served to show actual displacements of spe-cific points along the ST muscle–tendon unit. Second, previousstudies compared tendon with aponeuroses elongation and strain,by subtracting the absolute displacement of the tendon marker bythe absolute displacement of any randomly chosen point along theaponeurosis (Magnusson et al., 2001; Muramatsu et al., 2001). Thisis because the aponeuroses points are in line with the tendon andtheir elongation is affected by tendon elongation. This procedurewas applied in this study by subtracting tendon (P1) elongationby the elongation of each TI – aponeuroses intersection point(P2, P3, P4). The ‘‘corrected’’ elongation data were then used toestimate strain of each segment defined by the distance of eachpoint marker relative to the origin. In this occasion, however, the
Table 1Elongation (mm) of the ST tendon point (P1) and the intersection of the tendinous inscriptiointervals.
MVC% Tendon (P1)
0� 45� 90�
10 1.85 ± 0.47 1.88 ± 0.53 2.00 ± 0.5920 3.32 ± 0.97 3.51 ± 0.77 3.41 ± 0.9930 4.43 ± 1.21* 5.15 ± 1.55* 5.13 ± 1.56*
40 5.82 ± 1.67* 6.61 ± 1.95* 6.81 ± 2.34*
50 7.39 ± 2.73* 8.57 ± 2.61* 8.61 ± 2.98*
60 8.73 ± 3.21* 10.50 ± 3.42* 10.27 ± 3.56*
70 10.32 ± 4.27* 11.76 ± 3.37* 11.73 ± 3.78*
80 11.09 ± 4.52* 12.94 ± 3.06* 12.79 ± 3.62*
90 12.46 ± 4.62* 13.79 ± 3.91* 13.56 ± 3.52*
100 13.65 ± 4.80* 14.16 ± 3.82* 14.74 ± 3.74*
* Significantly different compared with values at 10% MVC.^ Values, collapsed for angular position and level of effort, higher than P1 and P4 value
Please cite this article in press as: Kellis E et al. Strain and elongation of the humhttp://dx.doi.org/10.1016/j.jelekin.2013.07.016
resting length of P2, P3 and P4 segments was defined as thedifference between the resting lengths of the tendon plusaponeuroses – TI intersection points and the tendon alone(120.07 ± 16.99 mm (P2), 131.64 ± 18.09 mm (P3) and88.13 ± 16.44 mm (P4)). In elongation and strain calculations, thedisplacement of each marker when the knee was passively movedfrom 90� to 0� and from 45� to 0� were also taken into consider-ation. Third, to examine relative movement of the TI-aponeurosesintersection points (P2, P3, P4), length and strain (expressed as per-centage of rest value at 40�) of the long and short arms of the TIwere estimated. All parameters were analyzed at 10% increments,from 0% to 100% MVC.
The differences in elongation and strain between the four seg-ments (defined by P1, P2, P3, and P4 markers) at 10 force levels,at three angular positions were checked using a three-way analysisof variance (ANOVA). Similar designs were applied for the elonga-tion and strain of the short and long arms of the TI. A two-way AN-OVA was also applied to examine point marker displacement(relative to the resting length) when the knee passively changedposition from 0� to 90�.
3. Results
3.1. Absolute displacement
The recorded (absolute) displacement values for each of themarker points are presented in Table 1 (P1 and P2) and Table 2(P3 and P4). The ANOVA showed a statistically significant main ef-fect for level of MVC (p < 0.05) and 4 location points (P < 0.05) butno effect of angular position was found (p > 0.05). Post-hoc Tukeytests indicated that displacement (collapsed across angular posi-tions) significantly increased from 20% to 100% MVC (p < 0.05). Fur-thermore, displacement was significantly higher for the superficialaponeurosis-TI intersection (P2) and apex (P3) points comparedwith the tendinous (P1) and the deep aponeurosis-TI (P4) points(p < 0.05).
3.2. Effects of angular position
Displacement of each point as the knee was passively moved tothe starting position of each ramping test is presented in Fig. 2. Theresults showed a statistically significant displacement of all pointmarkers, from 45� to 0� and from 45� to 90� (p < 0.05). Further-more, displacement was significantly higher for the P1 comparedwith P2, P3 and p4 at knee flexion angle of 90� while the oppositewas found at 0� testing position (p < 0.05).
n with the superficial aponeurosis (P2) during ramp contractions at 3 angular position
Superficial aponeuroses – TI intersection (P2)
0� 45� 90�
2.92 ± 1.15 2.00 ± 0.41 2.03 ± 0.595.07 ± 2.08 4.13 ± 0.79 3.40 ± 0.847.95 ± 2.51* 6.53 ± 1.51* 5.09 ± 1.56*
10.11 ± 3.18* 9.12 ± 2.71* 7.28 ± 2.56*
12.14 ± 3.37* 12.82 ± 3.55* 10.38 ± 3.18*
13.98 ± 4.18* 14.30 ± 3.28* 12.06 ± 4.53*
16.42 ± 4.45* 16.00 ± 3.09* 14.19 ± 4.09*
18.79 ± 5.30* 17.30 ± 3.23* 15.04 ± 4.18*
21.08 ± 5.84* 19.57 ± 4.11* 17.91 ± 4.59*
22.97 ± 6.06*^ 20.74 ± 4.03*^ 21.78 ± 5.11*^
s, see also Table 2).
an semitendinosus muscle – Tendon unit. J Electromyogr Kinesiol (2013),
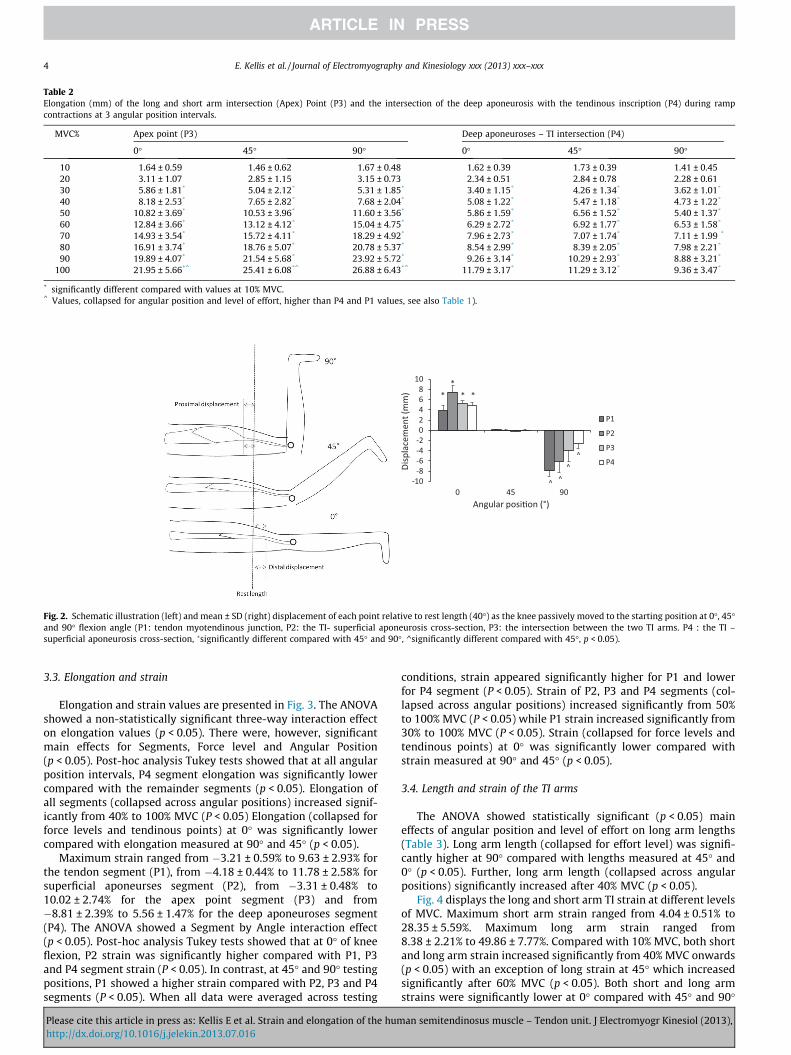
Table 2Elongation (mm) of the long and short arm intersection (Apex) Point (P3) and the intersection of the deep aponeurosis with the tendinous inscription (P4) during rampcontractions at 3 angular position intervals.
MVC% Apex point (P3) Deep aponeuroses – TI intersection (P4)
0� 45� 90� 0� 45� 90�
10 1.64 ± 0.59 1.46 ± 0.62 1.67 ± 0.48 1.62 ± 0.39 1.73 ± 0.39 1.41 ± 0.4520 3.11 ± 1.07 2.85 ± 1.15 3.15 ± 0.73 2.34 ± 0.51 2.84 ± 0.78 2.28 ± 0.6130 5.86 ± 1.81* 5.04 ± 2.12* 5.31 ± 1.85* 3.40 ± 1.15* 4.26 ± 1.34* 3.62 ± 1.01*
40 8.18 ± 2.53* 7.65 ± 2.82* 7.68 ± 2.04* 5.08 ± 1.22* 5.47 ± 1.18* 4.73 ± 1.22*
50 10.82 ± 3.69* 10.53 ± 3.96* 11.60 ± 3.56* 5.86 ± 1.59* 6.56 ± 1.52* 5.40 ± 1.37*
60 12.84 ± 3.66* 13.12 ± 4.12* 15.04 ± 4.75* 6.29 ± 2.72* 6.92 ± 1.77* 6.53 ± 1.58*
70 14.93 ± 3.54* 15.72 ± 4.11* 18.29 ± 4.92* 7.96 ± 2.73* 7.07 ± 1.74* 7.11 ± 1.99 *
80 16.91 ± 3.74* 18.76 ± 5.07* 20.78 ± 5.37* 8.54 ± 2.99* 8.39 ± 2.05* 7.98 ± 2.21*
90 19.89 ± 4.07* 21.54 ± 5.68* 23.92 ± 5.72* 9.26 ± 3.14* 10.29 ± 2.93* 8.88 ± 3.21*
100 21.95 ± 5.66*^ 25.41 ± 6.08*^ 26.88 ± 6.43*^ 11.79 ± 3.17* 11.29 ± 3.12* 9.36 ± 3.47*
* significantly different compared with values at 10% MVC.^ Values, collapsed for angular position and level of effort, higher than P4 and P1 values, see also Table 1).
Fig. 2. Schematic illustration (left) and mean ± SD (right) displacement of each point relative to rest length (40�) as the knee passively moved to the starting position at 0�, 45�and 90� flexion angle (P1: tendon myotendinous junction, P2: the TI- superficial aponeurosis cross-section, P3: the intersection between the two TI arms. P4 : the TI –superficial aponeurosis cross-section, �significantly different compared with 45� and 90�, ^significantly different compared with 45�, p < 0.05).
4 E. Kellis et al. / Journal of Electromyography and Kinesiology xxx (2013) xxx–xxx
3.3. Elongation and strain
Elongation and strain values are presented in Fig. 3. The ANOVAshowed a non-statistically significant three-way interaction effecton elongation values (p < 0.05). There were, however, significantmain effects for Segments, Force level and Angular Position(p < 0.05). Post-hoc analysis Tukey tests showed that at all angularposition intervals, P4 segment elongation was significantly lowercompared with the remainder segments (p < 0.05). Elongation ofall segments (collapsed across angular positions) increased signif-icantly from 40% to 100% MVC (P < 0.05) Elongation (collapsed forforce levels and tendinous points) at 0� was significantly lowercompared with elongation measured at 90� and 45� (p < 0.05).
Maximum strain ranged from �3.21 ± 0.59% to 9.63 ± 2.93% forthe tendon segment (P1), from �4.18 ± 0.44% to 11.78 ± 2.58% forsuperficial aponeurses segment (P2), from �3.31 ± 0.48% to10.02 ± 2.74% for the apex point segment (P3) and from�8.81 ± 2.39% to 5.56 ± 1.47% for the deep aponeuroses segment(P4). The ANOVA showed a Segment by Angle interaction effect(p < 0.05). Post-hoc analysis Tukey tests showed that at 0� of kneeflexion, P2 strain was significantly higher compared with P1, P3and P4 segment strain (P < 0.05). In contrast, at 45� and 90� testingpositions, P1 showed a higher strain compared with P2, P3 and P4segments (P < 0.05). When all data were averaged across testing
Please cite this article in press as: Kellis E et al. Strain and elongation of the humhttp://dx.doi.org/10.1016/j.jelekin.2013.07.016
conditions, strain appeared significantly higher for P1 and lowerfor P4 segment (P < 0.05). Strain of P2, P3 and P4 segments (col-lapsed across angular positions) increased significantly from 50%to 100% MVC (P < 0.05) while P1 strain increased significantly from30% to 100% MVC (P < 0.05). Strain (collapsed for force levels andtendinous points) at 0� was significantly lower compared withstrain measured at 90� and 45� (p < 0.05).
3.4. Length and strain of the TI arms
The ANOVA showed statistically significant (p < 0.05) maineffects of angular position and level of effort on long arm lengths(Table 3). Long arm length (collapsed for effort level) was signifi-cantly higher at 90� compared with lengths measured at 45� and0� (p < 0.05). Further, long arm length (collapsed across angularpositions) significantly increased after 40% MVC (p < 0.05).
Fig. 4 displays the long and short arm TI strain at different levelsof MVC. Maximum short arm strain ranged from 4.04 ± 0.51% to28.35 ± 5.59%. Maximum long arm strain ranged from8.38 ± 2.21% to 49.86 ± 7.77%. Compared with 10% MVC, both shortand long arm strain increased significantly from 40% MVC onwards(p < 0.05) with an exception of long strain at 45� which increasedsignificantly after 60% MVC (p < 0.05). Both short and long armstrains were significantly lower at 0� compared with 45� and 90�
an semitendinosus muscle – Tendon unit. J Electromyogr Kinesiol (2013),
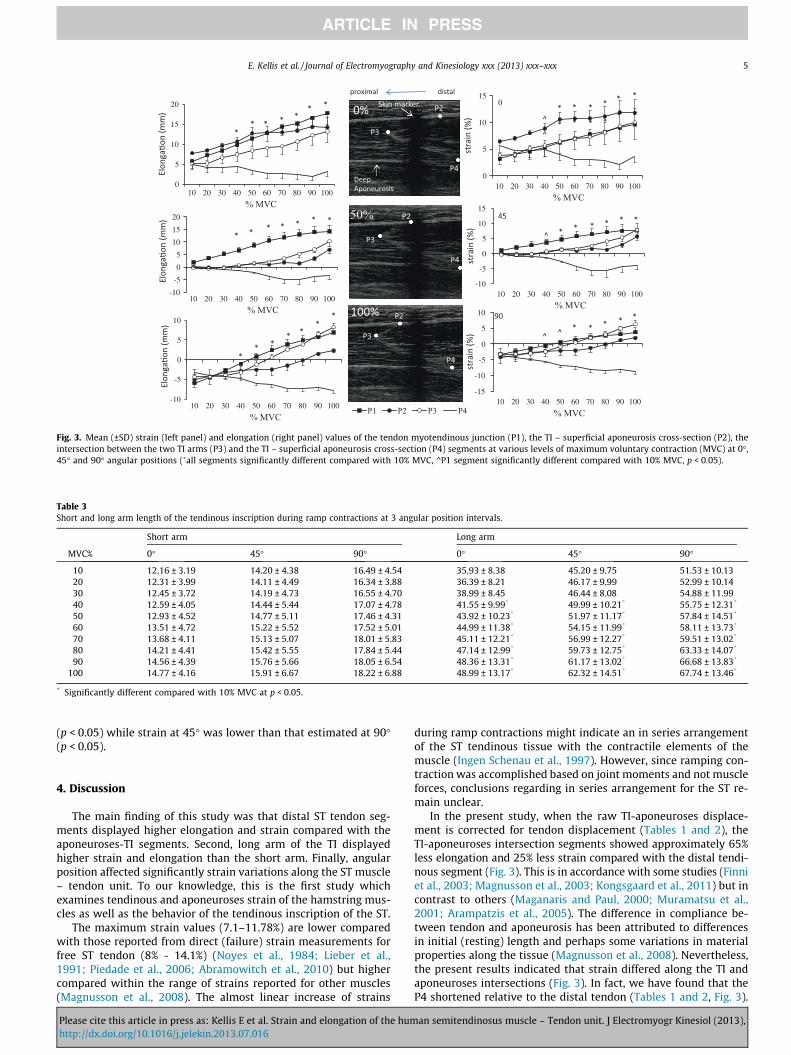
Fig. 3. Mean (±SD) strain (left panel) and elongation (right panel) values of the tendon myotendinous junction (P1), the TI – superficial aponeurosis cross-section (P2), theintersection between the two TI arms (P3) and the TI – superficial aponeurosis cross-section (P4) segments at various levels of maximum voluntary contraction (MVC) at 0�,45� and 90� angular positions (�all segments significantly different compared with 10% MVC, ^P1 segment significantly different compared with 10% MVC, p < 0.05).
Table 3Short and long arm length of the tendinous inscription during ramp contractions at 3 angular position intervals.
Short arm Long arm
MVC% 0� 45� 90� 0� 45� 90�
10 12.16 ± 3.19 14.20 ± 4.38 16.49 ± 4.54 35.93 ± 8.38 45.20 ± 9.75 51.53 ± 10.1320 12.31 ± 3.99 14.11 ± 4.49 16.34 ± 3.88 36.39 ± 8.21 46.17 ± 9.99 52.99 ± 10.1430 12.45 ± 3.72 14.19 ± 4.73 16.55 ± 4.70 38.99 ± 8.45 46.44 ± 8.08 54.88 ± 11.9940 12.59 ± 4.05 14.44 ± 5.44 17.07 ± 4.78 41.55 ± 9.99* 49.99 ± 10.21* 55.75 ± 12.31*
50 12.93 ± 4.52 14.77 ± 5.11 17.46 ± 4.31 43.92 ± 10.23* 51.97 ± 11.17* 57.84 ± 14.51*
60 13.51 ± 4.72 15.22 ± 5.52 17.52 ± 5.01 44.99 ± 11.38* 54.15 ± 11.99* 58.11 ± 13.73*
70 13.68 ± 4.11 15.13 ± 5.07 18.01 ± 5.83 45.11 ± 12.21* 56.99 ± 12.27* 59.51 ± 13.02*
80 14.21 ± 4.41 15.42 ± 5.55 17.84 ± 5.44 47.14 ± 12.99* 59.73 ± 12.75* 63.33 ± 14.07*
90 14.56 ± 4.39 15.76 ± 5.66 18.05 ± 6.54 48.36 ± 13.31* 61.17 ± 13.02* 66.68 ± 13.83*
100 14.77 ± 4.16 15.91 ± 6.67 18.22 ± 6.88 48.99 ± 13.17* 62.32 ± 14.51* 67.74 ± 13.46*
* Significantly different compared with 10% MVC at p < 0.05.
E. Kellis et al. / Journal of Electromyography and Kinesiology xxx (2013) xxx–xxx 5
(p < 0.05) while strain at 45� was lower than that estimated at 90�(p < 0.05).
4. Discussion
The main finding of this study was that distal ST tendon seg-ments displayed higher elongation and strain compared with theaponeuroses-TI segments. Second, long arm of the TI displayedhigher strain and elongation than the short arm. Finally, angularposition affected significantly strain variations along the ST muscle– tendon unit. To our knowledge, this is the first study whichexamines tendinous and aponeuroses strain of the hamstring mus-cles as well as the behavior of the tendinous inscription of the ST.
The maximum strain values (7.1–11.78%) are lower comparedwith those reported from direct (failure) strain measurements forfree ST tendon (8% - 14.1%) (Noyes et al., 1984; Lieber et al.,1991; Piedade et al., 2006; Abramowitch et al., 2010) but highercompared within the range of strains reported for other muscles(Magnusson et al., 2008). The almost linear increase of strains
Please cite this article in press as: Kellis E et al. Strain and elongation of the humhttp://dx.doi.org/10.1016/j.jelekin.2013.07.016
during ramp contractions might indicate an in series arrangementof the ST tendinous tissue with the contractile elements of themuscle (Ingen Schenau et al., 1997). However, since ramping con-traction was accomplished based on joint moments and not muscleforces, conclusions regarding in series arrangement for the ST re-main unclear.
In the present study, when the raw TI-aponeuroses displace-ment is corrected for tendon displacement (Tables 1 and 2), theTI-aponeuroses intersection segments showed approximately 65%less elongation and 25% less strain compared with the distal tendi-nous segment (Fig. 3). This is in accordance with some studies (Finniet al., 2003; Magnusson et al., 2003; Kongsgaard et al., 2011) but incontrast to others (Maganaris and Paul, 2000; Muramatsu et al.,2001; Arampatzis et al., 2005). The difference in compliance be-tween tendon and aponeurosis has been attributed to differencesin initial (resting) length and perhaps some variations in materialproperties along the tissue (Magnusson et al., 2008). Nevertheless,the present results indicated that strain differed along the TI andaponeuroses intersections (Fig. 3). In fact, we have found that theP4 shortened relative to the distal tendon (Tables 1 and 2, Fig. 3).
an semitendinosus muscle – Tendon unit. J Electromyogr Kinesiol (2013),
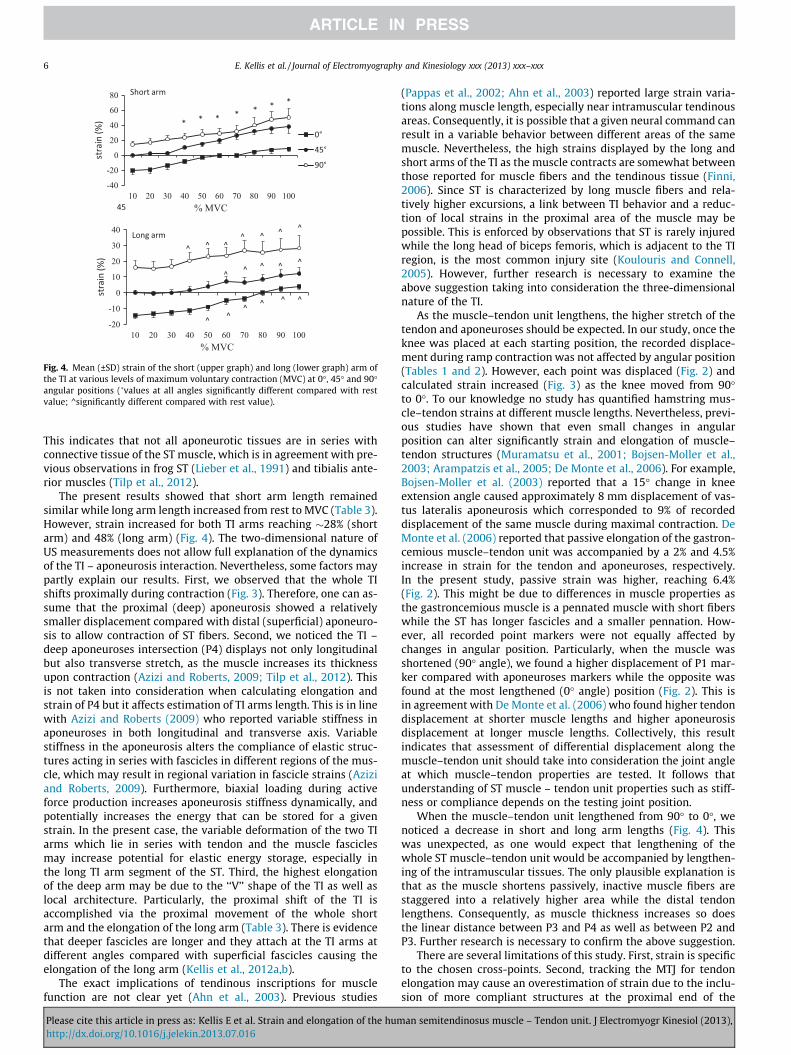
Fig. 4. Mean (±SD) strain of the short (upper graph) and long (lower graph) arm ofthe TI at various levels of maximum voluntary contraction (MVC) at 0�, 45� and 90�angular positions (�values at all angles significantly different compared with restvalue; ^significantly different compared with rest value).
6 E. Kellis et al. / Journal of Electromyography and Kinesiology xxx (2013) xxx–xxx
This indicates that not all aponeurotic tissues are in series withconnective tissue of the ST muscle, which is in agreement with pre-vious observations in frog ST (Lieber et al., 1991) and tibialis ante-rior muscles (Tilp et al., 2012).
The present results showed that short arm length remainedsimilar while long arm length increased from rest to MVC (Table 3).However, strain increased for both TI arms reaching �28% (shortarm) and 48% (long arm) (Fig. 4). The two-dimensional nature ofUS measurements does not allow full explanation of the dynamicsof the TI – aponeurosis interaction. Nevertheless, some factors maypartly explain our results. First, we observed that the whole TIshifts proximally during contraction (Fig. 3). Therefore, one can as-sume that the proximal (deep) aponeurosis showed a relativelysmaller displacement compared with distal (superficial) aponeuro-sis to allow contraction of ST fibers. Second, we noticed the TI –deep aponeuroses intersection (P4) displays not only longitudinalbut also transverse stretch, as the muscle increases its thicknessupon contraction (Azizi and Roberts, 2009; Tilp et al., 2012). Thisis not taken into consideration when calculating elongation andstrain of P4 but it affects estimation of TI arms length. This is in linewith Azizi and Roberts (2009) who reported variable stiffness inaponeuroses in both longitudinal and transverse axis. Variablestiffness in the aponeurosis alters the compliance of elastic struc-tures acting in series with fascicles in different regions of the mus-cle, which may result in regional variation in fascicle strains (Aziziand Roberts, 2009). Furthermore, biaxial loading during activeforce production increases aponeurosis stiffness dynamically, andpotentially increases the energy that can be stored for a givenstrain. In the present case, the variable deformation of the two TIarms which lie in series with tendon and the muscle fasciclesmay increase potential for elastic energy storage, especially inthe long TI arm segment of the ST. Third, the highest elongationof the deep arm may be due to the ‘‘V’’ shape of the TI as well aslocal architecture. Particularly, the proximal shift of the TI isaccomplished via the proximal movement of the whole shortarm and the elongation of the long arm (Table 3). There is evidencethat deeper fascicles are longer and they attach at the TI arms atdifferent angles compared with superficial fascicles causing theelongation of the long arm (Kellis et al., 2012a,b).
The exact implications of tendinous inscriptions for musclefunction are not clear yet (Ahn et al., 2003). Previous studies
Please cite this article in press as: Kellis E et al. Strain and elongation of the humhttp://dx.doi.org/10.1016/j.jelekin.2013.07.016
(Pappas et al., 2002; Ahn et al., 2003) reported large strain varia-tions along muscle length, especially near intramuscular tendinousareas. Consequently, it is possible that a given neural command canresult in a variable behavior between different areas of the samemuscle. Nevertheless, the high strains displayed by the long andshort arms of the TI as the muscle contracts are somewhat betweenthose reported for muscle fibers and the tendinous tissue (Finni,2006). Since ST is characterized by long muscle fibers and rela-tively higher excursions, a link between TI behavior and a reduc-tion of local strains in the proximal area of the muscle may bepossible. This is enforced by observations that ST is rarely injuredwhile the long head of biceps femoris, which is adjacent to the TIregion, is the most common injury site (Koulouris and Connell,2005). However, further research is necessary to examine theabove suggestion taking into consideration the three-dimensionalnature of the TI.
As the muscle–tendon unit lengthens, the higher stretch of thetendon and aponeuroses should be expected. In our study, once theknee was placed at each starting position, the recorded displace-ment during ramp contraction was not affected by angular position(Tables 1 and 2). However, each point was displaced (Fig. 2) andcalculated strain increased (Fig. 3) as the knee moved from 90�to 0�. To our knowledge no study has quantified hamstring mus-cle–tendon strains at different muscle lengths. Nevertheless, previ-ous studies have shown that even small changes in angularposition can alter significantly strain and elongation of muscle–tendon structures (Muramatsu et al., 2001; Bojsen-Moller et al.,2003; Arampatzis et al., 2005; De Monte et al., 2006). For example,Bojsen-Moller et al. (2003) reported that a 15� change in kneeextension angle caused approximately 8 mm displacement of vas-tus lateralis aponeurosis which corresponded to 9% of recordeddisplacement of the same muscle during maximal contraction. DeMonte et al. (2006) reported that passive elongation of the gastron-cemious muscle–tendon unit was accompanied by a 2% and 4.5%increase in strain for the tendon and aponeuroses, respectively.In the present study, passive strain was higher, reaching 6.4%(Fig. 2). This might be due to differences in muscle properties asthe gastroncemious muscle is a pennated muscle with short fiberswhile the ST has longer fascicles and a smaller pennation. How-ever, all recorded point markers were not equally affected bychanges in angular position. Particularly, when the muscle wasshortened (90� angle), we found a higher displacement of P1 mar-ker compared with aponeuroses markers while the opposite wasfound at the most lengthened (0� angle) position (Fig. 2). This isin agreement with De Monte et al. (2006) who found higher tendondisplacement at shorter muscle lengths and higher aponeurosisdisplacement at longer muscle lengths. Collectively, this resultindicates that assessment of differential displacement along themuscle–tendon unit should take into consideration the joint angleat which muscle–tendon properties are tested. It follows thatunderstanding of ST muscle – tendon unit properties such as stiff-ness or compliance depends on the testing joint position.
When the muscle–tendon unit lengthened from 90� to 0�, wenoticed a decrease in short and long arm lengths (Fig. 4). Thiswas unexpected, as one would expect that lengthening of thewhole ST muscle–tendon unit would be accompanied by lengthen-ing of the intramuscular tissues. The only plausible explanation isthat as the muscle shortens passively, inactive muscle fibers arestaggered into a relatively higher area while the distal tendonlengthens. Consequently, as muscle thickness increases so doesthe linear distance between P3 and P4 as well as between P2 andP3. Further research is necessary to confirm the above suggestion.
There are several limitations of this study. First, strain is specificto the chosen cross-points. Second, tracking the MTJ for tendonelongation may cause an overestimation of strain due to the inclu-sion of more compliant structures at the proximal end of the
an semitendinosus muscle – Tendon unit. J Electromyogr Kinesiol (2013),

E. Kellis et al. / Journal of Electromyography and Kinesiology xxx (2013) xxx–xxx 7
tendon. Third, soft tissue deformation might have an effect on therecorded moment, but such deformation is very difficult to preventor control (Arampatzis et al., 2005). Further, US analysis refers totwo-dimensional displacements while the tendinous – musclemorphology is three-dimensional.
5. Conclusion
The ST tendon segment displays a higher strain than segmentsalong the aponeuroses–tendinous inscription intersection. As themuscle contracts, the long arm shows almost double strain thanthe shorter arm and both arms show higher strain than the apo-neuroses and tendinous segments. These results indicate that themechanical properties of the ST muscle are affected by tendon,aponeuroses and tendinous inscription interactions.
Conflict of Interest Statement
None.
References
Abramowitch SD, Zhang X, Curran M, Kilger R. A comparison of the quasi-staticmechanical and non-linear viscoelastic properties of the humansemitendinosus and gracilis tendons. Clin Biomech (Bristol, Avon)2010;25:325–31.
Ahn AN, Monti RJ, Biewener AA. In vivo and in vitro heterogeneity of segmentlength changes in the semimembranosus muscle of the toad. J Physiol2003;549:877–88.
Arampatzis A, Stafilidis S, DeMonte G, Karamanidis K, Morey-Klapsing G,Bruggemann GP. Strain and elongation of the human gastrocnemius tendonand aponeurosis during maximal plantarflexion effort. J Biomech2005;38:833–41.
Azizi E, Roberts TJ. Biaxial strain and variable stiffness in aponeuroses. J Physiol2009;587:4309–18.
Bojsen-Moller J, Hansen P, Aagaard P, Kjaer M, Magnusson SP. Measuringmechanical properties of the vastus lateralis tendon–aponeurosis complexin vivo by ultrasound imaging. Scand J Med Sci Sports 2003;13:259–65.
De Monte G, Arampatzis A, Stogiannari C, Karamanidis K. In vivo motiontransmission in the inactive gastrocnemius medialis muscle–tendon unitduring ankle and knee joint rotation. J Electromyogr Kinesiol 2006;16:413–22.
Finni T. Structural and functional features of human muscle–tendon unit. Scand JMed Sci Sports 2006;16:147–58.
Finni T, Hodgson JA, Lai AM, Edgerton VR, Sinha S. Nonuniform strain of humansoleus aponeurosis–tendon complex during submaximal voluntary contractionsin vivo. J Appl Physiol 2003;95:829–37.
Fiorentino NM, Epstein FH, Blemker SS. Activation and aponeurosis morphologyaffect in vivo muscle tissue strains near the myotendinous junction. J Biomech2012;45:647–52.
Ingen Schenau GJ, Bobbert MF, de Haan A. Does elastic energy enhance work andefficiency in the stretch-shortening cycle. J Appl Biomech 1997;13:389–415.
Kellis E, Baltzopoulos V. The effects of antagonist moment on the resultant kneejoint moment during isokinetic testing of the knee extensors. Eur J Appl PhysiolOccup Physiol 1997;76:253–9.
Kellis E, Galanis N, Natsis K, Kapetanos G. Validity of architectural properties of thehamstring muscles: Correlation of ultrasound findings with cadavericdissection. J Biomech 2009;42:2549–54.
Kellis E, Galanis N, Natsis K, Kapetanos G. Muscle architecture variations along thehuman semitendinosus and biceps femoris (long head) length. J ElectromyogrKinesiol 2010;20:1237–43.
Kellis E, Galanis N, Kapetanos G, Natsis K. Architectural differences between thehamstring muscles. J Electromyogr Kinesiol 2012.
Kellis E, Galanis N, Natsis K, Kapetanos G. In vivo and in vitro examination of thetendinous inscription of the human semitendinosus muscle. Cells TissuesOrgans 2012b;195:365–76.
Kongsgaard M, Nielsen CH, Hegnsvad S, Aagaard P, Magnusson SP. Mechanicalproperties of the human Achilles tendon, in vivo. Clin Biomech (Bristol, Avon)2011;26:772–7.
Koulouris G, Connell D. Hamstring muscle complex: an imaging review.Radiographics 2005;25:571–86.
Lieber RL, Leonard ME, Brown CG, Trestik CL. Frog semitendinosis tendon load–strain and stress–strain properties during passive loading. Am J Physiol1991;261:C86–92.
Maganaris CN, Paul JP. Load-elongation characteristics of in vivo human tendon andaponeurosis. J Exp Biol 2000;203:751–6.
Please cite this article in press as: Kellis E et al. Strain and elongation of the humhttp://dx.doi.org/10.1016/j.jelekin.2013.07.016
Magnusson SP, Aagaard P, Dyhre-Poulsen P, Kjaer M. Load–displacement propertiesof the human triceps surae aponeurosis in vivo. J Physiol 2001;531:277–88.
Magnusson SP, Hansen P, Aagaard P, Brond J, Dyhre-Poulsen P, Bojsen-Moller J, et al.Differential strain patterns of the human gastrocnemius aponeurosis and freetendon, in vivo. Acta Physiol Scand 2003;177:185–95.
Magnusson SP, Narici MV, Maganaris CN, Kjaer M. Human tendon behaviour andadaptation, in vivo. J Physiol 2008;586:71–81.
Muramatsu T, Muraoka T, Takeshita D, Kawakami Y, Hirano Y, Fukunaga T.Mechanical properties of tendon and aponeurosis of human gastrocnemiusmuscle in vivo. J Appl Physiol 2001;90:1671–8.
Noyes FR, Butler DL, Grood ES, Zernicke RF, Hefzy MS. Biomechanical analysis ofhuman ligament grafts used in knee-ligament repairs and reconstructions. JBone Joint Surg Am 1984;66:344–52.
Pappas GP, Asakawa DS, Delp SL, Zajac FE, Drace JE. Nonuniform shortening in thebiceps Brachii during elbow flexion. J Appl Physiol 2002;92:2381–9.
Piedade SR, Dal Fabbro IM, Mischan MM. Cyclic-loading of the human gracilis andsemitendinosus muscle tendons: study of young adult cadavers. Artif Organs2006;30:680–5.
Silder A, Whittington B, Heiderscheit B, Thelen DG. Identification of passive elasticjoint moment-angle relationships in the lower extremity. J Biomech2007;40:2628–35.
Tilp M, Steib S, Herzog W. Length changes of human tibialis anterior centralaponeurosis during passive movements and isometric, concentric, and eccentriccontractions. Eur J Appl Physiol 2012;112:1485–94.
Woodley SJ, Mercer SR. Hamstring muscles: architecture and innervation. CellsTissues Organs 2005;179:125–41.
Eleftherios Kellis is an associate professor in SportKinesiology at the Department of Physical Educationand Sports Sciences at Serres, Aristotle University ofThessaloniki, Greece. He obtained his B.Ed. in PhysicalEducation and Sport Sciences, at the Aristotle Universityof Thessaloniki, Greece (1993) and his Ph.D. at theDepartment of Movement Sciences and Physical Edu-cation, University of Liverpool, United Kingdom (1996).He has many research publications while he authored abook entitled ‘‘Neuromechanical principles of humanmuscle strength assessment’’ in 2008. He is a co-foun-der of the Laboratory of Neuromechanics at Serres,Greece and his main research interests include ham-
string muscle modeling and function, joint mechanics and clinical electromyogra-phy applications.
Glykeria Patsika completed her B.Ed. in Physical Edu-cation and Sport Sciences at the Aristotle University ofThessaloniki and received her Master degree in Kinesi-ology (2009) from the same Department at Serres,Greece. She is currently a Doctoral student in Kinesiol-ogy at the Aristotle University of Thessaloniki and hermain research interests include muscle activation andmechanics of knee joint in people with knee osteoar-thritis.
Evaggelos Karagiannidis completed his B.Ed. in Physi-cal Education and Sport Sciences at the Aristotle Uni-versity of Thessaloniki and received a Master degree inPhysical Activity and Quality of Life (2008) and Kinesi-ology (2009) from the same university. He is currently aDoctoral student in Kinesiology at Department ofPhysical Education and Sport Science at Serres. His mainresearch interests include muscle activation andmechanics of knee joint in people who underwentanterior cruciate ligament reconstruction.
an semitendinosus muscle – Tendon unit. J Electromyogr Kinesiol (2013),




















