
Just One Thing Twelve Of The World's Best Investors Reveal ...
Upload
independentCategory
view
4download
0
Twelve Group I Introns in the Same Pre-rRNA Transcript of theMyxomycete Fuligo septica: RNA Processing and Evolution
Eirik W. Lundblad,* Christer Einvik,* Sissel Rønning,* Kari Haugli,* and Steinar Johansen*�*Department of Molecular Biotechnology, RNA research group, Institute of Medical Biology, University of Tromsø,Tromsø, Norway; and �Faculty of Fishery and Natural Sciences, Bodø Regional University, Bodø, Norway
The ribosomal DNA region of the myxomycete Fuligo septica was investigated and found to contain 12 group I introns(four in the small subunit and eight in the large subunit ribosomal RNAs). We have performed molecular andphylogenetic analyses to provide insight into intron structure and function, intron-host biology, and intron origin andevolution. The introns vary in size from 398 to 943 nt, all lacking detectable open reading frames. Secondary structuremodels revealed considerable structural diversity, but all, except one (subclass IB), represent the common group IC1intron subclass. In vitro splicing analysis revealed that 10 of the 12 introns were able to self-splice as naked RNA, but all12 introns were able to splice out from the precursor rRNA in vivo as evaluated by reverse transcription PCR analysis ontotal F. septica RNA. Furthermore, RNA processing analyses in vitro and in vivo showed that 10 of 12 introns performhydrolytic cleavage at the 39 splice site, as well as intron circularization. Full-length intron RNA circles were detected invivo. The order of splicing was analyzed by a reverse transcription PCR approach on cellular RNA, but no strict order ofintron excision could be detected. Phylogenetic analysis indicated that most Fuligo introns were distantly related to eachother and were independently gained in ribosomal DNA during evolution.
Introduction
All group I intron sequences known in the nucleusinterrupt the ribosomal RNA genes of protists and fungi,and more than 1,300 examples at 50 different integrationsites are known (Cannone et al. 2002; Jackson et al. 2002).Group I introns are characterized by conserved secondaryRNA structures consisting of at least 10 paired segments(P1 to P10) common to most group I ribozymes, andusually several optional segments (P11 to P17) present insubsets of introns (Lehnert et al. 1996; Einvik et al. 1998).The highly conserved catalytic core responsible for theself-splicing reaction consists of P4 to P6 and P3 to P9(Michel and Westhof 1990; Golden et al. 1998). Group Iintron RNAs have been divided into 12 subclasses withinfive main groups (IA1-3, IB1-4, IC1-3, ID, and IE), basedon conserved secondary structure, tertiary interactions, andphylogenetic analyses (see Michel and Westhof [1990] andSuh, Jones, and Blackwell [1999]). Group I intron RNAsare ribozymes catalyzing their own splicing reactions,resulting in perfectly ligated exons. The self-splicingreaction is initiated by a nucleophilic attack of anexogenous guanosine at the 59 splice site and proceedsby a second transesterification reaction between thegenerated free 39 end of the 59 exon and the 39 splicesite (Cech and Herschlag 1996). A parallel, competing,intron RNA–processing reaction, known as the introncircularization pathway, is initiated by hydrolytic cleavageat the 39 splice site (Nielsen et al. 2003). Here, the free 39hydroxyl group of the last intron residue (xG) generatedby hydrolysis attacks the 59 exon-intron junction andresults in the formation of full-length circular (FLC) intronRNAs, as well as nonligated exons. Thus, the circulariza-tion pathway generates nonfunctional rRNAs and appearsto challenge the viability of the host.
It is generally considered that nuclear group I intronsbenefit from a ribosomal DNA localization because theyare replicated as part of the host chromosomes andcotranscribed by RNA polymerase I as an integrated partof the pre-rRNA (Lin and Vogt 1998; Jackson et al. 2002).Once transcribed, the group I introns have to be preciselyspliced out to generate functional pre-rRNAs essential tohost cell viability. The excised group I intron RNAs areeither rapidly degraded (Brehm and Cech 1983), reversedspliced into cognate or related RNA sites (Roman andWoodson 1998), or, in the case of some mobile introns,further processed into homing endonuclease messengerRNAs (Lin and Vogt 1998; Vader, Nielsen, and Johansen1999; Decatur, Johansen, and Vogt 2000). The widespreaddistribution and patterns of relatedness suggest that group Iintrons are selfish genetic elements able to spread bothvertically and horizontally between evolutionarily distinctlineages and are thought to be the product of multipleinsertions and selective losses (see Goddard and Burt[1999]).
Despite the large number of nuclear group I intronsknown, very few have been subjected to experimentallytests for RNA processing activities in vitro as naked RNAand in vivo from endogenous pre-rRNAs. Among these arethe Tetrahymena introns, their mobile cognate intron fromPhysarum, and the complex twin-ribozyme introns inDidymium and Naegleria. We recently reported that allthese introns generate FLC intron RNAs when incubated invitro (Haugen, De Jonckheere, and Johansen 2002; Nielsenet al. 2003). Whereas the Tetrahymena introns do notgenerate detectable FLCs in vivo, the Didymium intronforms a significant amount of FLCs in the nucleus, thuschallenging the viability of the cell. In this report we havecharacterized the small subunit (SSU) and large subunit(LSU) ribosomal DNA (rDNA) from Fuligo septica. TherDNA was found to contain 12 group I introns, which is thehighest group I intron content reported for a singleprecursor rRNA species. Thus, F. septica serves as anattractive model system in the characterization of biologicalroles and molecular evolution of group I introns. Here, we
Key words: Fuligo, group I intron, myxomycete, ribosomal RNA,RNA splicing.
E-mail: [email protected].
Mol. Biol. Evol. 21(7):1283–1293. 2004doi:10.1093/molbev/msh126Advance Access publication March 19, 2004
Molecular Biology and Evolution vol. 21 no. 7 � Society for Molecular Biology and Evolution 2004; all rights reserved.
by guest on October 14, 2014
http://mbe.oxfordjournals.org/
Dow
nloaded from

present the results from molecular and phylogeneticanalyses of the Fuligo introns to provide new insight intofundamental questions such as intron structure andfunction, intron-host biology, and intron origin andevolution.
Materials and MethodsFuligo septica Culturing, DNA Isolation,and Sequencing
The myxomycete F. septica (NY-1 [Haugen et al.2003]) was grown on DS/2 media (Johansen et al. 1997) at268C feeding on Escherichia coli. Before DNA isolation,cells were harvested and washed twice in DS/2 media bycentrifugation at 3,000 rpm to separate amoeboid cells frombacteria. Total DNA was isolated as previously described(Haugen et al. 2003), and F. septica rDNA was amplifiedby PCR using various primers designed from knownmyxomycete rDNA sequences. Automatic sequencing wasperformed by the ABI PRISM BigDyeTerminator CycleSequencing Ready Reaction Kit (PerkinElmer) running onan ABI Prism 377 system (PerkinElmer), and manualsequencing was performed by the Thermo SequenaseRadiolabeled Terminator Cycle Sequencing kit (USBCorp.) according to the manufacturer’s instructions.
Plasmid Construction
All introns from F. septica rDNA were PCR amplifiedwith a forward T7 promoter-containing primer locatedapproximately 30 nt upstream of the 59 splice site, as wellas a reverse primer located approximately 100 ntdownstream the 39 splice site. For introns closely spaced,neighbor intron sequences were included to obtain flankingregions of appropriate length. The amplified products wereinserted into pUC18 using the SureClone ligation kit(Amersham Pharmacia Biotech) and transformed into E.coli JM105 as described by Sambrook and Russel (2001).Recombinant plasmids were isolated from E. coli using theDNA purification Wizard SV plus kit (Promega) andsubsequently confirmed by sequencing. Site-specfic muta-genesis was performed by the PCR-based Quick Changesite-directed mutagenesis kit (Stratagene).
Reverse Transcription PCR Determination of IntronSplicing Order, Intron Circle, and LigatedExon Junctions
Total RNA was isolated from F. septica amoeboidcells by the TRIzol reagent (Invitrogen). The DNA wasremoved by incubating the RNA preparation with RQ1DNase (Promega) and subsequental RNA Cleanup by theRNeasy kit (Qiagen). Reverse transcription (RT) PCRreactions were performed by using the First-Strand cDNASynthesis kit (Amersham Bioscience) or Superscript IIRNaseH-Reverse Transcriptase (Invitrogen) according tothe manufacturer’s instructions. Splicing order of introns inrRNA was determined by RT-PCR on total-cellular RNAswith nested reverse primers. PCR amplifications using thesame primer sets were also performed directly on total-
cellular RNA as a control. Intron circle and ligated exonjunctions were determined by RT-PCR on purified cellularRNA, in vitro spliced RNA, and RNA purified frompolyacrylamide gels, as previously described (Haugen,DeJonckheere, and Johansen 2002). All primer sequencesused in this study are available on our Web site (seeAcknowledgments).
In vitro Transcription and Group I Intron Splicing
RNA was transcribed by T7 RNA polymerase offrestriction enzyme–linearized intron plasmid using [35S]CTP aS (10 mCi/ml [Amersham Pharmacia Biotech]) forlabeling, purified by phenol extraction and ethanol pre-cipitation, and dissolved in DEPC-treated water. RNA wassubjected to self-splicing conditions essentially as de-scribed by Johansen and Vogt (1994) at 508C for 0 to 60min in low-salt buffer (40 mM Tris-HCl pH 7.5, 200 mMKCl, 2 mM spermidine, 5 mM DTT, 10 mM MgCl2, 0.2mM GTP) or high-salt buffer (40 mM Tris-HCl pH 7.5, 1 MKCl, 2 mM spermidine, 5 mM DTT, 25 mM MgCl2, 0.2mM GTP). Splicing intermediates and products wereanalyzed by denaturing polyacrylamide gel electrophoresis(5% polyacrylamid, 7 M urea) and visualized by auto-radiografi.
Nucleic Acid Sequence and Phylogenetic Analyses
Computer analyses of DNA sequences were per-formed in general by the DNASTAR version 5 softwarepackage (DNASTAR Inc.). For the phylogenetic analyses,a nucleotide sequence alignment based on approximately100 conserved positions within the catalytic core of thegroup I introns was constructed according to thesecondary structure of the introns. ClustalX version 1.81(Thompson et al. 1997) alignments gave similar treetopologies compared with those generated by manualalignment according to secondary structure. The ClustalXalignment was used to calculate distance matrixes, p-distance method (Nei and Kumar 2000), from whichphylogenetic trees were constructed. Phylogenetic analy-ses performed with the neighbor-joining (NJ) andmaximum-parsimony (MP) methods gave similar treetopologies. A bootstrap test with 1,000 resamplings wasperformed. The phylogenetic trees were generated byMEGA version 2.1 (Kumar et al. 2001).
ResultsStructural Features of 12 Group I Introns inFuligo septica rDNA
Sequencing analysis of F. septica rDNA revealed 12group I introns at conserved regions in the SSU and LSUrRNA genes (fig. 1). Six of the rDNA sites (S516, S956,L1090, L1949, L2449, and L2500) were previously knownto contain group I introns in either nuclear or organellargenomes in some eukaryotic microorganisms (see Cannoneet al. [2002]), but others appear novel (S911, S1065, L569,L1898, L1911, and L2584). The Fuligo introns, which arenamed according to Johansen and Haugen (2001), range insize from 398 nt to 943 nt, and lack recognizable open
1284 Lundblad et al.
by guest on October 14, 2014
http://mbe.oxfordjournals.org/
Dow
nloaded from

reading frames. All introns, except Fse.L1949 andFse.L2449 (see below), represent typical group IC1 intronswith structural features resembling the well studiedTetrahymena rDNA intron (see Lehnert et al. [1996] andGolden et al. [1998]). Whereas Fse.S956 and Fse.S1065contain all base-paired segments proposed for the Tetra-hymena intron, others (Fse.S516, Fse.S911, Fse.L569,Fse.L1090, and Fse.L2584) have gained an additionalsegment in the P4 to P6 domain (P5d). Here, Fse.L569is unusual because of a 16-nt direct-repeat motif (59-CTAAGGGTTCACGGAT-39) present in 17 identicalcopies at P5d. One intron (Fse.S911) has a small one-segment P9 region, and three introns (Fse.S516, Fse.S1065,and Fse.L2584) contain the P13 tertiary base pairingbetween L2.1 and L9.1.
Two Fuligo introns possess unusual secondarystructural features. Fse.L1949 and Fse.L2449 resemblecognate introns of the related myxomycetes Physarumpolycephalum and P. flavicomum (Johansen, Johansen, andHaugli 1992; Vader et al. 1994). Fse.L1949 and Fse.L2449contain large extension sequences in L1, which includesthree copies of a nearly perfect 45-nt direct-repeat motif inFse.L1949. Furthermore, Fse.L1949 contains a U:A pair inplace of the conserved U:G pair at the 59 splice site.Fse.L2449 contains large additional extensions at periph-eral loops, as well as the unusual P5 hairpin P5.1 (fig. 2).Fse.L1949 folds into a group IB–like structure lackinga recognizable P8 segment, and Fse.L2449 folds intoa group IC–like secondary structure (fig. 2).
Ten of 12 Fuligo Introns Self-Splice AsNaked RNA In vitro
Catalytic activities of the Fuligo rDNA introns wereanalyzed from in vitro transcribed and purified RNAs,subsequently incubated at splicing conditions. All theintrons, except Fse.L1949 and Fse.L2449, were found toself-splice but at slightly different optimal conditions.Representative time-course experiments show that a signif-icant fraction of precursor RNAs was spliced after 60 minof incubation, generating free intron RNAs and ligatedexons (fig 3A). RT-PCR and sequencing analyses ofprocessed RNAs confirmed that all the self-splicing Fuligointrons perform perfect exon ligation in vitro (data notshown). Lack of self-splicing of Fse.L1949 and Fse.L2449may be assigned to their unusual sequence features (seeabove). These observations corroborate results of the
cognate introns in the myxomycetes Didymium andPhysarum, where no in vitro splicing products weredetected (Johansen, Johansen, and Haugli 1992; Vaderet al. 1994). All 12 Fuligo introns splice accurately invivo, a conclusion based on an RT-PCR and sequencingapproach using exon-specific oligo primers (figure 4).Thus, the lack of in vitro self-splicing of Fse.L1949 andFse.L2449 is probably because of the dependence onessential splicing factors (maturase-like activities) in vivo.
No Essential Order of Intron Excisionand Splicing In vivo
In vitro self-splicing analysis of two cotranscribedintron pairs (Fse.L1898/Fse.L1911 and Fse.L2500/Fse.L2584) demonstrated that one of the intron partners(Fse.L1898 and Fse.L2584, respectively) performed thesplicing reaction faster than the other when incubated atsame reaction conditions (data not shown). These obser-vations suggested a preferred order of group I intronexcision in Fuligo pre-rRNA similar to what was pre-viously reported for three P. polycephalum rDNA introns(Rocheleau and Woodson 1995). However, the observed invitro splicing order for the Fuligo intron pairs appeared notto be essential. Inactivation of the most efficient splic-ing partner (Fse.L1898 or Fse.L2584) by mutating theP7 guanosine-binding site (G:C to C:G), with subsequentcotranscription and in vitro splicing, showed thatboth Fse.L1911 and Fse.L2500 were able to self-splicebefore the mutated Fse.L1898 and Fse.L2584 (data notshown).
To investigate the order of splicing in vivo,endogenous Fuligo RNA was isolated and subjected toan RT-PCR approach (see Rocheleau and Woodson[1995]). Here, splicing intermediates of isolated pre-rRNAwere detected by the use of primer pairs complementary tosequences within introns, as well as the flanking exons(fig. 4). By this approach, the presence or absence ofa given intron can be investigated in a pre-RNAintermediate that contains a second intron. Because ofthe large number of introns in Fuligo rDNA, the analysiswas restricted to neighboring intron pairs. All amplifiedproducts were confirmed by DNA sequencing.
The majority of endogenous rRNA sequencesappeared completely spliced because only intron-lackingsegments could be amplified from exon-specific primerpairs (figure 4, primer combination F1/R2). No essential
FIG. 1.—Schematic presentation of the sequence-analyzed rDNA region in F. septica. The SSU rRNA and LSU rRNA gene regions are interruptedby four and eight group I introns, respectively. The position of each intron is indicated and numbered according to E. coli rDNA (see Johansen andHaugen [2001] and Cannone et al. [2002]). ITS1 and ITS2, internal transcribed spacers 1 and 2. The sequence has been assigned the EMBL/GenBankData library accession number AJ584697.
Multiple Group I Introns in Fuligo rDNA 1285
by guest on October 14, 2014
http://mbe.oxfordjournals.org/
Dow
nloaded from
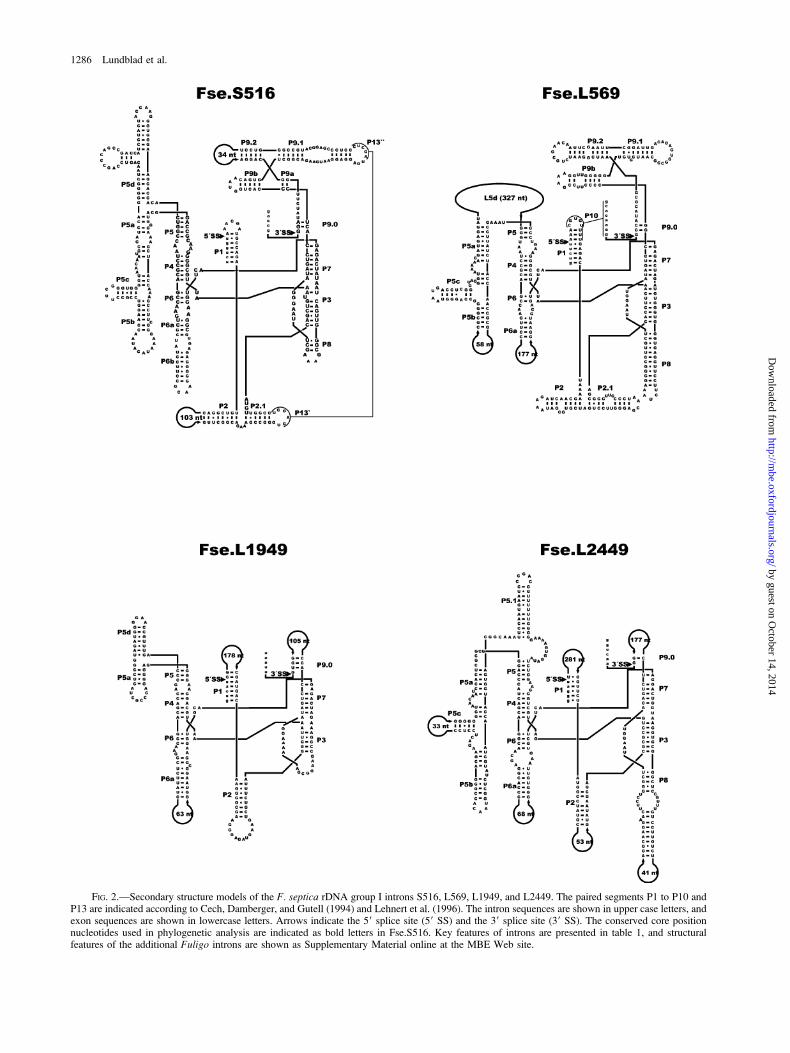
FIG. 2.—Secondary structure models of the F. septica rDNA group I introns S516, L569, L1949, and L2449. The paired segments P1 to P10 andP13 are indicated according to Cech, Damberger, and Gutell (1994) and Lehnert et al. (1996). The intron sequences are shown in upper case letters, andexon sequences are shown in lowercase letters. Arrows indicate the 59 splice site (59 SS) and the 39 splice site (39 SS). The conserved core positionnucleotides used in phylogenetic analysis are indicated as bold letters in Fse.S516. Key features of introns are presented in table 1, and structuralfeatures of the additional Fuligo introns are shown as Supplementary Material online at the MBE Web site.
1286 Lundblad et al.
by guest on October 14, 2014
http://mbe.oxfordjournals.org/
Dow
nloaded from

order of intron excision and exon splicing was detectedfrom the intron pair analysis of Fse.L1898/ Fse.L1911 andFse.L2500/ Fse.L2584. These results support the in vitromutation analysis (see above) but do not exclude thepossibility of different rates of splicing of particular intronsin vivo. Similar results were seen for all intron combina-tions investigated because all the experiments detectedprocessed RNAs lacking both introns and lacking one ofthe two introns (schematically presented in figure 4). Basedon these observations, we conclude that there is no essentialorder of intron excision and splicing in Fuligo pre-rRNA.
Full-Length Intron RNA Circles Are Generated BothIn vitro and In vivo
Gel analysis of in vitro self-splicing intron RNAsidentified slow-migrating circular RNA species in allsamples (fig. 3A). An RT-PCR and sequencing approachof gel-purified circular RNAs (see Einvik et al. [1997])confirmed that the majority of Fuligo rDNA intronsgenerate full-length circle (FLC) RNAs during incubation(table 1). FLCs are generated from the circularizationpathway initiated by hydrolytic cleavage at the 39 splicesite, a reaction distinct from the intron-splicing pathway(figure 3B and Nielsen et al. [2003]). A representativeexample from the analysis of ligated exons and FLCs isshown for the Fse.L1090 intron (fig. 3C). Similar resultswere obtained when the RT-PCR sequencing approach wasapplied on endogenous Fuligo pre-rRNA (table 1). All theself-splicing Fuligo introns were found to generate FLCseither in vitro or in vivo. Furthermore, additional intronRNA circles were observed for six introns in vitro and twointrons in vivo (table 1). These circles are truncated at theinternal guide sequence (IGS) and thus lack nucleotideresidues corresponding to the 59 end of the introns.
Multiple Origin of Fuligo Group I Introns inNuclear Ribosomal DNA
A comparison of the Fuligo introns with otherrepresentative nuclear group I introns (table 2) was basedon conserved sequence positions within the ribozyme core.All intron cores were folded according to establishedgroup I intron RNA secondary structure models (Cech,Damberger, and Gutell 1994; Lehnert et al. 1996) beforealignment analysis. The p-distance method (Nei andKumar 2000) was used to generate the distance matrixbecause the number of introns included in the analysis wasrelatively high, and few positions of each intron could beunambiguously aligned. The tree topology was found to besimilar for both NJ (fig. 5) and MP analyses.
The overall pattern seen from the NJ tree (fig. 5)corroborates previous reports on group I intron evolution(Bhattacharya et al. 1994; Shinohara, LoBuglio, andRogers 1996; Nishida, Tajiri, and Sugiyama 1998; Suh,Jones, and Blackwell 1999; Jackson et al. 2002; Haugen etal. 2003). Here, group IE introns are found to be distantlyrelated to the group IC1 introns, and introns at the samerDNA site from distantly related hosts tend to be moresimilar than introns at different sites in closely relatedhosts. An illustrating example is given in figure 5 by the
highly similar introns present at position 1925 in LSUrDNA (L1925) of the ciliate Tetrahymena and the distantlyrelated myxomycete Physarum. Features relevant to theorigin and evolution of Fuligo introns are seen from the NJtree in figure 5. First, most Fuligo introns are interspersedamong nuclear group I introns present in distantly relatedorganisms. Second, introns located within the same gene(SSU or LSU rRNA gene) appear not more closely relatedthan introns between the genes. Third, some of the intronsites (S516, S956, L1949, and L2449) are known to containgroup I introns in other myxomycetes that are more closelyrelated to those in Fuligo (Johansen, Johansen, and Haugli1992; Vader et al. 1994; Haugen et al. 2003). Finally, threeof the 12 Fuligo introns (Fse.L1898, Fse.L1911, andFse.L2500) seem more related to each other than to anyother group I intron in the database (Cannone et al. 2002)and may have originated by intron transfer between sites inFuligo rRNA (intron transposition). We conclude that theFuligo group I introns represent a complex evolutionaryhistory and that most introns have been independentlyacquired in the Fuligo rDNA. Thus, both horizontal andvertical intron transfers have to be considered to explain themultiple origins and high number of group I introns inFuligo rDNA.
Discussion
Posttranscriptional processing of pre-rRNA in eukar-yotes consists of a complex pathway of RNA cleavages,digestions, and covalent modifications performed by a largenumber of trans-acting protein or ribonucleoprotein factors(e.g., Venema and Tollervey [1995]). The existence of 12group I introns in F. septica rDNA, which have to beperfectly removed from the pre-rRNA either cotranscrip-tionally or posttranscriptionally to render the rRNAsfunctional, is an additional challenge to the host cells.Thus, it is surprising that the multiple Fuligo rDNA intronsare not removed in a strict order. The relaxed splicing orderimplies that exon sequence contexts are not critical for theremoval of individual introns, a situation similar to whatfound in the multiple group I intron–containing gene psbAof the Chlamydomonas chloroplast (Deshpande, Bao, andHerrin 1997). However, ordered splicing of nuclear group Iintrons has previously been reported in Physarum poly-cephalum pre-rRNA, where a mobile, self-splicing intronwas removed before two obligate conserved myxomyceteintrons (Rocheleau and Woodson 1995). A very differentcase of ordered splicing in pre-rRNA was observed inDidymium iridis. Here, the spliceosomal intron foundinserted into a group I self-splicing intron was removedsubsequently to the group I intron (Vader, Nielsen, andJohansen 1999). When the group I intron RNA isinactivated by endonucleolytic cleavage, the spliceosomalintron remains unspliced in vivo (Vader, Johansen, andNielsen 2002). Finally, a highly preferred, but not obligateorder of splicing was reported in a multi-intron mammalianpre-mRNA transcript (Kessler, Jiang, and Chasin 1993).Ordered spliceosomal intron splicing is the result ofindividual differences in the splicing kinetics betweenintrons of the same transcripts (Audibert, Weil, and Dautry2002).
Multiple Group I Introns in Fuligo rDNA 1287
by guest on October 14, 2014
http://mbe.oxfordjournals.org/
Dow
nloaded from
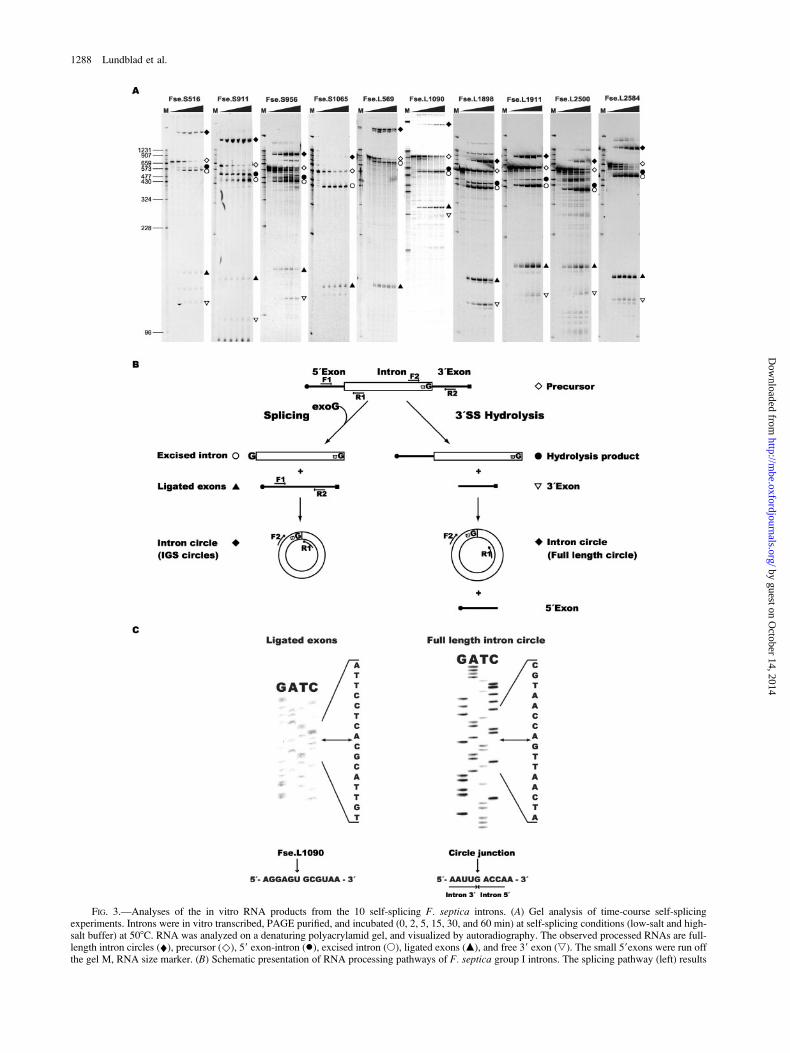
FIG. 3.—Analyses of the in vitro RNA products from the 10 self-splicing F. septica introns. (A) Gel analysis of time-course self-splicingexperiments. Introns were in vitro transcribed, PAGE purified, and incubated (0, 2, 5, 15, 30, and 60 min) at self-splicing conditions (low-salt and high-salt buffer) at 508C. RNA was analyzed on a denaturing polyacrylamid gel, and visualized by autoradiography. The observed processed RNAs are full-length intron circles (¤), precursor ()), 59 exon-intron (�), excised intron (s), ligated exons (m), and free 39 exon (,). The small 59exons were run offthe gel M, RNA size marker. (B) Schematic presentation of RNA processing pathways of F. septica group I introns. The splicing pathway (left) results
1288 Lundblad et al.
by guest on October 14, 2014
http://mbe.oxfordjournals.org/
Dow
nloaded from

What are the costs and benefits for host cells of havinggroup I introns in the essential and heavily transcribed rRNAgenes? Early studies in Tetrahymena pigmentosa concludedthat the presence of a group I intron in rDNA appears neutralwith no observed phenotypic feature compared withcorresponding cells lacking an intron (Nielsen and Engberg1985; Nielsen, Simon, and Engberg 1992). This observationmay be explained by the fact that group I introns, such as theTetrahymena intron, splice out very efficiently and at thesame time are self-sufficient by encoding the ribozyme-based splicing activity (Brehm and Cech 1983). However,recent studies have unveiled several group I intron activitiesthat may challenge the host. First, the excised linear introncould reverse splice into cellular ribosomal RNA sequences,or perhaps other endogenous RNAs, and, thus, interfere withcellular gene expression (Roman, Rubin, and Woodson1999). Second, internal intron RNA processing could renderthe host RNA nonfunctional. During starvation-inducedencystment in Didymium, internal processing of a mobilerDNA group I intron results in a partial processed pre-rRNAtranscript incompatible with the formation of functionalrRNAs (Vader, Johansen, and Nielsen 2002). Finally, wehave recently shown that the formation of full-lengthcircular (FLC) intron RNAs from nuclear group I intronsis common in vivo (Haugen, De Jonckheere, and Johansen2002; Nielsen et al. 2003). FLC-formation is dependent ona processing pathway different from splicing, and initiatedby hydrolytic cleavage at the 39 splice site of the intron(figure 3B and Nielsen et al. [2003]). Similar to the internalprocessing event described above, FLC formation resultsin nonfunctional rRNAs from the same transcript. The
observation that 10 of the 12 Fuligo introns are able toform FLCs, and that FLCs were detected from eight intronsin vivo, is consistent with the idea of an ongoing competitionbetween FLC formation and splicing in Fuligo nuclei.
In contrast, there are few examples of group I intronswith reported advantageous effects to the host. Splicing of
FIG. 4.—Analysis of splicing orders of F. septica rDNA group I introns. A possible strict order of excision among neighboring introns wasassessed by an RT-PCR approach designed to detect the presence or absence of introns in endogenous RNA transcripts. A schematic presentation ofneighboring introns (A and B) in precursor rRNA (pre-rRNA) and detected processed RNA are shown. Total RNA isolated from growing F.septica cells was subjected to RT-PCR using the primer combinations F1/R2, F2/R2, and F1/R1, and corresponding products were gel purified andDNA sequenced. Intron pairs analyzed were S516/S911, S911/S956, S956/S1065, L569/L1090, L1090/L1898, L1898/L1911, L1911/L1949,L1949/L2449, L2449/L2500, and L2500/L2584. The three variants of processed RNAs were detected for all intron pairs. F1 and F2 are forwardprimers. R1 and R2 are reverse primers. The primer sequences used in this experiment are listed as Supplementary Material online at the MBE Web site.
in exon ligation, intron excision, and truncated IGS intron circles. The hydrolysis pathway (right) results in full-length intron circles and free, nonligatedexons (see Nielsen et al. [2003]). Location of primers used in the RT-PCR approach for analyzing ligated exons and intron circle junctions areindicated. (C) Representative experiments of ligated exon (left) and full-length intron circle (right) junction analyses from Fse.L1090. RNA sequence ofjunctions corresponding to ligated exon and full-length intron circle are presented below.
Table 1Key Features and Summary of Functional Analyses ofGroup I Introns Present in F. septica rDNA
IntronName
Size(bp)
In vitroSelf-Splicing
In vitro39SS
Hydrolysis
In vitroRNACircle
In vivoRNACircle
Fse.S516 534 1 1 1a 1a
Fse.S911 457 1 1 1b 1a
Fse.S956 436 1 1 1a NDc
Fse.S1065 419 1 1 1a 1a
Fse.L569 907 1 1 1b 1a
Fse.L1090 592 1 1 1a 1a
Fse.L1898 424 1 1 1a,b 1a
Fse.L1911 432 1 1 1a,b 1b
Fse.L1949 531 2 2 NDc NDc
Fse.L2449 943 2 2 NDc NDc
Fse.L2500 398 1 1 1b 1a
Fse.L2584 504 1 1 1b 1a,b
NOTE.—In vitro self-splicing, 39SS hydrolysis and intron circle in vitro were
detected by PAGE analyses of pre-RNA incubated at splicing conditions and by
RT-PCR of ligated exon and inverse RT-PCR of intron circle junction. Intron circle
in vivo was detected by sequencing of inverse RT-PCR product of intron circle
junction from total RNA isolated from F. septica.a Full-length circles.b Truncated circles.c ND indicates not detected.
Multiple Group I Introns in Fuligo rDNA 1289
by guest on October 14, 2014
http://mbe.oxfordjournals.org/
Dow
nloaded from
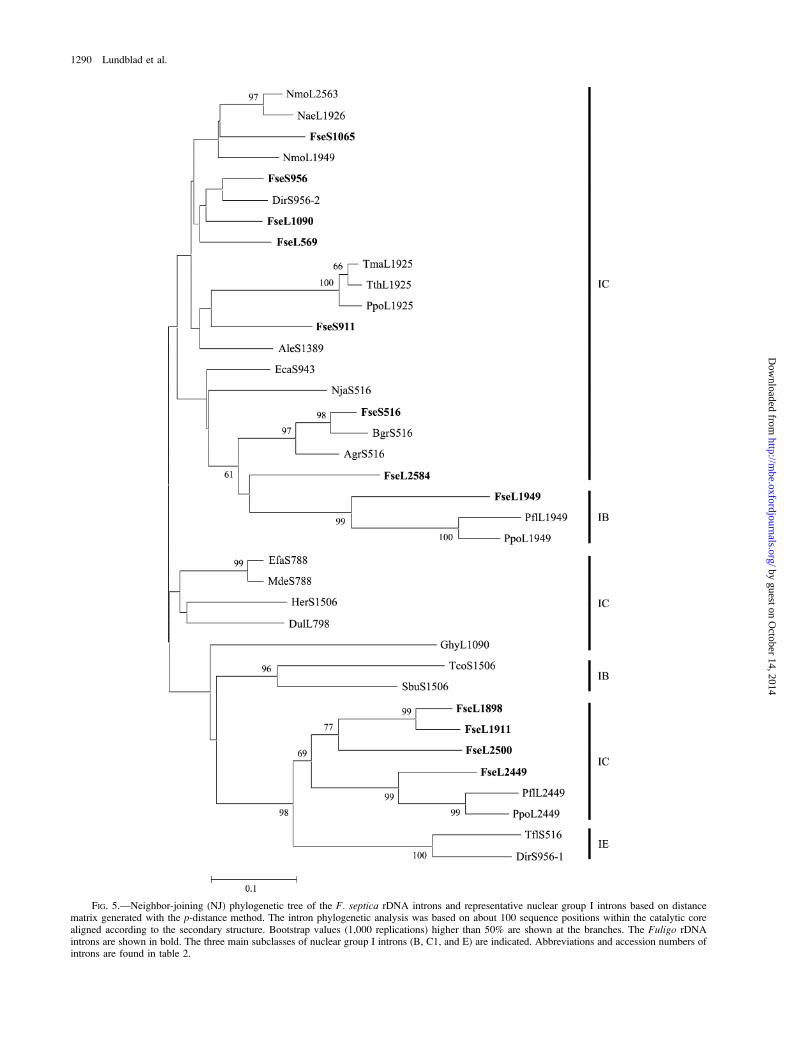
FIG. 5.—Neighbor-joining (NJ) phylogenetic tree of the F. septica rDNA introns and representative nuclear group I introns based on distancematrix generated with the p-distance method. The intron phylogenetic analysis was based on about 100 sequence positions within the catalytic corealigned according to the secondary structure. Bootstrap values (1,000 replications) higher than 50% are shown at the branches. The Fuligo rDNAintrons are shown in bold. The three main subclasses of nuclear group I introns (B, C1, and E) are indicated. Abbreviations and accession numbers ofintrons are found in table 2.
1290 Lundblad et al.
by guest on October 14, 2014
http://mbe.oxfordjournals.org/
Dow
nloaded from

introns in the psbA gene of Chlamydomonas chloroplastsis light dependent and regulates the expression of the psbAencoded D1 protein (Deshpande, Bao, and Herrin 1997).A different case of an advantageous effect for the host isthat the presence of introns makes the DNA resistant tohoming endonucleases that cleave in the vicinity of theintron insertion site. Double-strand breaks generated bysuch endonucleases (homing endonucleases) are fatal tothe host unless rapidly repaired. Homing endonucleasesare known to be present in a variety of microorganismsand with specificities that overlap several of the FuligorDNA intron insertion sites (Johansen et al. 1997; Elde,
Willassen, and Johansen 2000; Chevalier and Stoddard2001).
The intron phylogeny supports an independent andmultiple gain of group I introns by Fuligo rDNA. A similarconclusion was obtained by Cummings, Michel, andMcNally (1989), who studied 16 introns (14 group Iintrons) present in the CO I gene of the Podosporamitochondrial genome. The fact that introns with identicalinsertion sites tend to cluster together regardless of the hostspecies indicates lateral intron transfer during evolution.The actual mechanism for lateral transfer is not known, butboth RNA-based and DNA-based pathways have beenproposed (Lambowitz and Belfort 1993). The observationthat many fungi and protists harbor viruses (Schuster andDunnebacke 1986; Ghabrial 1998; Van Etten and Meints1999) and that group I introns have been detected in someof these (Nishida et al. 1998) make viruses interestingas possible intron vectors. A common feature amongorganisms harboring related group I introns is that theyshare natural niches. Intron transfers may be facilitatedthrough predation, endosymbiosis, endoparasitism, orinterspecies hybridization (Nishida and Sugiyama 1995;Adams, Clements, and Vaughn 1998). Nikoh and Fukatsu(2001) noted that organisms possessing multiple group Iintrons seem to be highly specialized for symbiotic orparasitic life styles. They proposed that the slow-growingendosymbionts/parasites are particularly well suited formultiple introns. The negative fitness effect of multipleinsertions in rDNA would be much less for these organismsthan for free-living organisms where optimal fitnessthrough growth rate and nutrient utilization is critical forsurvival and reproduction. Indeed, lichen-forming fungi areknown to contain multiple group I introns in nuclear rDNA(Gargas, De Priest, and Taylor 1995; Bhattacharya, Friedl,and Helms 2000). In contrast, F. septica is neitherendosymbiotic nor slow growing, and, unlike most otherknown myxomycetes, it feeds on almost all kinds ofmicroorganisms at the forest floor. Perhaps the largenumber of introns accumulated in Fuligo rDNA reflects itsnatural behavior and promiscuous feeding habits and, thus,is a case in point of the phrase ‘‘you become what you eat.’’
Supplementary Material
Secondary structural features of the eight additionalFuligo introns are presented as supplementary material tofigure 2, and primer sequences and specificities areincluded as supplementary materials to figure 4.
Acknowledgments
We thank Anna Vader for initiating PCR and DNAsequencing analyses and Volker M. Vogt for providing F.septica spores. This work was supported by grants fromThe Norwegian Research Council, The Norwegian CancerSociety, The Aakre Foundation for Cancer Research, andSimon Fougner Hartmanns Foundation.
Literature Cited
Adams, K. L., M. J. Clements, and J. C. Vaughn. 1998. ThePeperomia mitochondrial coxI group I intron: timing of
Table 2Group I Introns Included in the Phylogenetic Analysis
Species Intron SubgroupAccession
number
Myxomycota [Protist]
Badhamia gracilis Bgr.S516 C1 AJ555451Didymium iridis Dir.S956-1 E X71792D. iridis Dir.S956-2 C1 unpublishedFuligo septica Fse.S516 C1 AJ584697F. septica Fse.S911 C1 AJ584697F. septica Fse.S956 C1 AJ584697F. septica Fse.S1065 C1 AJ584697F. septica Fse.L569 C1 AJ584697F. septica Fse.L1090 C1 AJ584697F. septica Fse.L1898 C1 AJ584697F. septica Fse.L1911 C1 AJ584697F. septica Fse.L1949 B-like AJ584697F. septica Fse.L2449 C-like AJ584697F. septica Fse.L2500 C1 AJ584697F. septica Fse.L2584 C1 AJ584697Physarum flavicomum Pfl.L1949 B-like X78959P. flavicomum Pfl.L2449 C-like X78959P. polycephalum Ppo.L1925 C1 L03183P. polycephalum Ppo.L1949 B-like X60211P. polycephalum Ppo.L2449 C-like X60211
Rhizopoda [Protist]
Acanthamoeba griffini Agr.S516 C1 S81337A. lenticulata Ale.S1389 C1 U94734
Discomitochondria [Protist]
Naegleria jamiesoni Nja.S516 C1 U80250N. morganensis Nmo.L1949 C1 AJ001314N. morganensis Nmo.L2563 C1 AJ001315Naegleria sp. NG874 Nae.L1926 C1 AJ271406
Ciliophora [Protist]
Tetrahymena malaccensis Tma.L1925 C1 X03105T. thermophila Tth.L1925 C1 V01416
Basidiomycota [Fungi]
Tilletiopsis flava Tlf.S516 E D82819
Ascomycota [Fungi]
Dumontinia ulmariae Dul.L798 C1 AJ226079Encoelia fascicularis Efa.S788 C1 Z81379Exophiala calicioides Eca.S943 C1 AB007685Gyalecta hypoleuca Ghy.L1090 C1 AF465453Hymenoscyphus ericae Her.S1506 C1 U06868Myriosclerotinia dennisii Mde.S788 C1 AJ226072Symbiotaphrina buchneri Sbu.S1606 B D49657Teloschistes
contortuplicatus Tco.S1506 B AJ421258
NOTE.—Species are grouped into phyla of Protist or Fungi according to the
classification of Margulis and Schwartz (1998). Introns are named according to
abbreviation of host scientific name and insertion site in SSU or LSU rRNA
(Johansen and Haugen 2001).
Multiple Group I Introns in Fuligo rDNA 1291
by guest on October 14, 2014
http://mbe.oxfordjournals.org/
Dow
nloaded from

horizontal transfer and subsequent evolution of the intron.J. Mol. Evol. 46:689–696.
Audibert, A., D. Weil, and F. Dautry. 2002. In vivo kinetics ofmRNA splicing and transport in mammalian cells. Mol. Cell.Biol. 22:6706–67018.
Bhattacharya, D., T. Friedl, and G. Helms. 2002. Verticalevolution and intragenic spread of lichen-fungal group Iintrons. J. Mol. Evol. 55:74–84.
Bhattacharya, D., B. Surek, M. Rusing, S. Damberger, and M.Melkonian. 1994. Group I introns are inherited throughcommon ancestry in the nuclear-encoded rRNA of Zygnema-tales (Charophyceae). Proc. Natl. Acad. Sci. USA 91:9916–9920.
Brehm, S. L., and T. R. Cech. 1983. Fate of an interveningsequence ribonucleic acid: excision and cyclization of theTetrahymena ribosomal ribonucleic acid intervening sequencein vivo. Biochemistry 22:2390–2397.
Cannone, J. J., S. Subramanian, M. N. Schnare et al. (14 co-authors) 2002. The Comparative RNA Web (CRW) Site: anonline database of comparative sequence and structureinformation for ribosomal, intron, and other RNAs. BMCBioinform. 3:1–31.
Cech, T. R., S. H. Damberger, and R. R. Gutell. 1994.Representation of the secondary and tertiary structure ofgroup I introns. Nat. Struct. Biol. 1:273–280.
Cech, T., and D. Herschlag. 1996. Group I ribozymes: substraterecognition, catalytic strategies, and comparative mechanisticanalysis. Pp. 1–17 in F. Eckstein and D. M. J. Lilley, eds.Catalytic RNA: nucleic acids and molecular biology, Vol 10.Springer Verlag, New York.
Chevalier, B. S., and B. L. Stoddard. 2001. Homing endonu-cleases: structural and functional insight into the catalysts ofintron/ intein mobility. Nucleic Acids Res. 18:3757–3774.
Cummings, D. J., F. Michel, and K. L. McNally. 1989. DNAsequence analysis of the 24.5 kilobase pair cytochrome oxidasesubunit I mitochondrial gene from Podospora anserina: a genewith sixteen introns. Curr. Genet. 16:381–406.
Decatur, W. A., S. Johansen, and V. M. Vogt. 2000. Expressionof the Naegleria intron endonuclease is dependent ona functional group I self-cleaving ribozyme. RNA 6:616–627.
Deshpande, N. N., Y. Bao, and D. L. Herrin. 1997. Evidence forlight/redox-regulated splicing of psbA pre-RNAs in Chlamy-domonas chloroplasts. RNA 3:37–48.
Einvik, C., W. A. Decatur, T. M. Embley, V. M. Vogt, and S.Johansen. 1997. Naegleria nucleolar introns contain twogroup I ribozymes with different functions in RNA splicingand processing. RNA 3:710–720.
Einvik, C., H. Nielsen, E. Westhof, F. Michel, and S. Johansen.1998. Group I-like ribozymes with a novel core organizationperform obligate sequential hydrolytic cleavages at twoprocessing sites. RNA 4:530–541.
Elde, M., N. P. Willassen, and S. Johansen. 2000. Functionalcharacterization of isoschizomeric His-Cys box homingendonucleases from Naegleria. Eur. J. Biochem. 267:7257–7266.
Gargas, A., P. T. DePriest, and J. W. Taylor. 1995. Positions ofmultiple insertions in SSU rDNA of lichen-forming fungi.Mol. Biol. Evol. 12:208–218.
Ghabrial, S. A. 1998. Origin, adaptation and evolutionary path-ways of fungal viruses. Vir. Genes 16:119–131.
Goddard, M. R., and A. Burt. 1999. Recurrent invasion andextinction of a selfish gene. Proc. Natl. Acad. Sci. USA96:13880–13885.
Golden, B. L., A. R. Gooding, E. R. Podell, and T. R. Cech.1998. A preorganized active site in the crystal structure of theTetrahymena ribozyme. Science 282:259–264.
Haugen, P., D. H. Coucheron, S. B. Rønning, K. Haugli, andS. Johansen. 2003. The molecular evolution and structuralorganization of self-splicing group I introns at position 516 innuclear SSU rDNA of myxomycetes. J. Euk. Microbiol.50:283–292.
Haugen, P., J. F. De Jonckheere, and S. Johansen. 2002.Characterization of the self-splicing products of two complexNaegleria LSU rDNA group I introns containing homingendonuclease genes. Eur. J. Biochem. 269:1641–1649.
Jackson, S., J. Cannone, J. Lee, R. Gutell, and S. Woodson. 2002.Distribution of rRNA introns in the three-dimensionalstructure of the ribosome. J. Mol. Biol. 323:35–52.
Johansen, S., M. Elde, A. Vader, P. Haugen, K. Haugli, and F.Haugli. 1997. In vivo mobility of a group I twintron in nuclearribosomal DNA of the myxomycete Didymium iridis. Mol.Microbiol. 24:737–745.
Johansen, S., and P. Haugen. 2001. A new nomenclature ofgroup I introns in ribosomal DNA. RNA 7:935–936.
Johansen, S., T. Johansen, and F. Haugli. 1992. Structure andevolution of myxomycete nuclear group I introns: a model forhorizontal transfer by intron homing. Curr. Genet. 22:297–304.
Johansen, S., and V. M. Vogt. 1994. An intron in the nuclearribosomal DNA of Didymium iridis codes for a group Iribozyme and a novel ribozyme that cooperate in self-splicing.Cell 76:725–734.
Kessler, O., Y. Jiang, and L. A. Chasin. 1993. Order of intronremoval during splicing of endogenous adenine phosphori-bosyltransferase and dihydrofolate reductase pre-mRNA. Mol.Cell. Biol. 13:6211–6222.
Kumar, S., K. Tamura, I. B. Jakobsen, and M. Nei. 2001.MEGA2: molecular evolutionary genetics analysis softaware.Bioinformatics 17:1244–1245.
Lambowitz, A. M., and M. Belfort. 1993. Introns as mobilegenetic elements. Annu. Rev. Biochem. 62:587–622.
Lehnert, V., L. Jaeger, F. Michel, and E. Westhof. 1996. New loop-loop tertiary interactions in self-splicing introns of subgroup ICand ID: a complete 3D model of the Tetrahymena thermophilaribozyme. Chem. Biol. 3:993–1009.
Lin, J., and V. M. Vogt. 1998. I-PpoI, the endonuclease encodedby the group I intron PpLSU3, is expressed from an RNApolymerase I transcript. Mol. Cell. Biol. 18:5809–5817.
Margulis, L., and K. V. Schwartz. 1998. Five kingdoms: anillustrated guide to the phyla of life in earth. 3rd edition.Freeman, New York.
Michel, F., and E. Westhof. 1990. Modelling of the threedimensional architecture of group I catalytic introns based oncomparative sequence analysis. J. Mol. Biol. 216:585–610.
Nei, M., and S. Kumar. 2000. Molecular evolution and phylo-genetics. Oxford University Press, New York.
Nielsen, H., and J. Engberg. 1985. Functional intron1 and intron-rDNA in the same macronucleus of the ciliate Tetrahymenapigmentosa. Biochim. Biophys. Acta 825:30–38.
Nielsen, H., T. Fiskaa, A. B. Birgisdottir, P. Haugen, C. Einvik,and S. Johansen. 2003. The ability to form full-length intronRNA circles is a general property of nuclear group I introns.RNA 9:1464–1475.
Nielsen, H., E. M. Simon, and J. Engberg. 1992. Inheritance ofthe group I rDNA intron in Tetrahymena pigmentosa. Dev.Genet. 13:133–142.
Nikoh, N., and T. Fukatsu. 2001. Evolutionary dynamics ofmultiple group I introns in nuclear ribosomal RNA genes ofendoparasitic fungi of the genus Cordyceps. Mol. Biol. Evol.18:1631–1642.
Nishida, H., and J. Sugiyama. 1995. A common group I intronbetween a plant parasitic fungus and its host. Mol. Biol. Evol.12:883–886.
1292 Lundblad et al.
by guest on October 14, 2014
http://mbe.oxfordjournals.org/
Dow
nloaded from

Nishida, H., Y. Tajiri, and J. Sugiyama. 1998. Multiple origins offungal group I introns located in the same position of nuclearSSU rRNA gene. J. Mol. Evol. 46:442–448.
Nishida, K., S. Suzuki, Y. Kimura, N. Nomura, M. Fujie, and T.Yamada. 1998. Group I introns found in Chlorella viruses:biological implications. Virology 242:319–326.
Rocheleau, G. A., and S. A. Woodson. 1995. Enhanced self-splicing of Physarum polycephalum intron 3 by a secondgroup I intron. RNA 1:183–193.
Roman, J., M. N. Rubin, and S. A. Woodson. 1999. Sequencespecificity of in vivo reverse splicing of the Tetrahymenagroup I intron. RNA 5:1–13.
Roman, J., and S. A. Woodson. 1998. Integration ofthe Tetrahymena group I intron into bacterial rRNA by reversesplicing in vivo. Proc. Natl. Acad. Sci. USA 95:2134–39.
Sambrook, J., and D. W. Russel. 2001. Molecular cloning,a laboratory manual. Cold Spring Harbor Laboratory Press,Cold Spring Harbor, New York.
Schuster, F. L. and T. H. Dunnebacke. 1986. Virus-like particlesin a soil ameboflagellate, Naegleria gruberi. Pp. 349–362 inR. B. Wickner, A. Hinnebusch, A. M. Lambowitz, et al., eds.Extrachromosomal elements in lower eukaryotes. PlenumPress. New York.
Shinohara, M. L., K. F. LoBuglio, and S. O. Rogers. 1996.Group-I intron family in the nuclear ribosomal RNA smallsubunit genes of Cenococcum geophilum isolates. Curr.Genet. 29:377–387.
Suh, S. O., K. G. Jones, and M. Blackwell. 1999. A group Iintron in the small subunit rRNA gene of Cryptendoxyla
hypophloia, an ascomycetous fungus: evidence for a new
major class of group I introns. J. Mol. Evol. 48:493–500.Thompson, J. D., T. J. Gibson, F. Plewniak, F. Jeanmougin, and
D. G. Higgins. 1997. The CLUSTAL_X Windows interface:
flexible strategies for multiple sequence alignment aided by
quality analysis tools. Nucleic Acids Res. 25:4876–4882.Vader, A., S. Johansen, and H. Nielsen. 2002. The group I-like
ribozyme DiGIR1 mediates alternative processing of pre-
rRNA transcripts in Didymium iridis. Eur. J. Biochem.
269:5804–5812.Vader, A., J. Naess, K. Haugli, F. Haugli, and S. Johansen. 1994.
Nucleolar introns from Physarum flavicomum contain inser-
tion elements that may explain how mobile group I introns
gained their open reading frames. Nucleic Acids Res. 22:
4553–4559.Vader, A., H. Nielsen, and S. Johansen. 1999. In vivo expression
of the nucleolar group I intron-encoded I-DirI homing
endonuclease involves the removal of a spliceosomal intron.
EMBO J. 18:1003–1013.Van Etten, J. L., and R. H. Meints. 1999. Giant viruses infecting
algae. Annu. Rev. Microbiol. 53:447–494.Venema, J., and D. Tollervey. 1995. Processing of pre-ribosomal
RNA in Saccharomyces cerevisiae. Yeast 11:1629–1650.
Laura Katz, Associate Editor
Accepted February 17, 2004
Multiple Group I Introns in Fuligo rDNA 1293
by guest on October 14, 2014
http://mbe.oxfordjournals.org/
Dow
nloaded from
Copyright © 2022 FDOKUMEN




















