
Tubular and cellular localization of the cardiac L-type calcium channel in rat kidney
14
Kidney International, Vol. 61 (2002), pp. 1393–1406 ION CHANNELS – MEMBRANE TRANSPORT – INTEGRATIVE PHYSIOLOGY Tubular and cellular localization of the cardiac L-type calcium channel in rat kidney PEI-LIN ZHAO, 1 XI-TAO WANG, 1 XUE-MEI ZHANG,VALERIU CEBOTARU,LIUDMILA CEBOTARU, GANG GUO,MARCELO MORALES, and SANDRA E. GUGGINO Department of Medicine, Johns Hopkins University School of Medicine, Baltimore, Maryland, USA, and Instituto de Biophysica Carlos Chagas, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil Tubular and cellular localization of the cardiac L-type calcium and skeletal muscle, brain and endocrine organs where channel in rat kidney. they function to increase calcium influx or to transduce Background. The mRNAs of several types of calcium chan- voltage changes [1]. Six types of 1 subunit have been nels have been identified in intact rat kidney, and L-type cal- cloned including three L-type channels ( 1S , 1C , and 1D ), cium channels cause changes in intracellular calcium in primary the -conotoxin-GIVA-sensitive N-type ( 1B ), the -aga- cultures of distal tubule cells. The aim of this study was to evaluate the tubular and cellular distribution of the 1C subunit toxin-IVA-sensitive P/Q-type ( 1A ), and the resistant of the L-type calcium channel in intact kidney. R-type ( 1E ). The L-type calcium channel complex is Methods. RT-PCR and Northern blot analysis were used to composed of multiple subunits [2] but the 1 subunit assess the regional abundance of the mRNA of this channel. comprises the main conduction core, the voltage sensor Immunocytochemistry combined with confocal microscopy and and also contains the drug binding sites for dihydropyri- surface biotinylation were applied to determine the tubular and cellular localization of the protein. dine antagonists such as nitrendipine or nifedipine or Results. Northern blot and RT-PCR analysis indicated that the agonist Bay K 8644, for phenylalkylamines such as the mRNA of the 1C subunit of the cardiac L-type calcium verapamil, and for benzothiazipines such as d-cis-diltia- channel was present in whole rat kidney, kidney tubules and zem [1, 3]. When the 1 subunit is expressed alone, the kidney cell lines. Western blot of lysates from whole kidney, membrane current density is low and voltage activation kidney tubules or cell lines revealed bands of 190 kD for the 1C subunit and 60 kD for the 3 subunit. Confocal immuno- is slower than in endogenous tissue. The other subunits histochemistry indicated that the 1C subunit of this channel was ( 2 , , , and ) increase current activation and inactiva- co-expressed in cells of the distal tubule that express calbindin- tion of the conduction core. Calcium channels also are D 28K , but not in intercalated cells. The 1C subunit was also expressed in non-excitable cells [4–8] and the mRNA of highly expressed in both outer and inner medullary collecting several types of calcium channels have been identified ducts. Serial confocal microscopic images or surface biotinyla- tion experiments determined that the channel was predomi- in both the cortex and medulla of intact rat kidney [9]. nantly on the basolateral membrane but had some distribution In distal tubule cells, Barry et al [10] found that antisense on the apical membrane. to the 1D subunit inhibits a parathyroid hormone (PTH)- Conclusions. The distribution and cellular localization of the mediated calcium influx, whereas antisense to the 1C 1C subunit of cardiac L-type calcium channel suggest it is subunit inhibits the diuretic-mediated calcium influx. In probably involved in intracellular and membrane calcium sig- naling. the cortical collecting duct calcium signaling is mediated by -adrenergic receptors [11]. In medullary thick as- cending limb cells, L-type calcium channels also have The family of depolarization-gated calcium channels been implicated in increases in intracellular calcium after is distributed in excitable tissues including heart, smooth volume expansion [12]. Finally, in inner medullary col- lecting ducts, vasopressin receptors cause elevation of intracellular calcium [13]. 1 Drs. Zhao and Wang contributed equally to this project. Recently, two other calcium channels from the TRP family have been cloned and localized to the distal tubule Key words: cortical collecting duct, distal tubule, medullary collecting duct, calcium signaling, intracellular signaling. of the kidney [14, 15]. Both ECaC [14] and CAT-2 [15] are localized in the principal cells of the distal tubule Received for publication August 15, 2001 where 1,25-dihydroxyvitamin D 3 responsive transepi- and in revised form November 9, 2001 Accepted for publication November 9, 2001 thelial calcium transport occurs, and both of these pro- teins colocalize with the major 1,25-dihydroxyvitamin 2002 by the International Society of Nephrology 1393
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Tubular and cellular localization of the cardiac L-type calcium channel in rat kidney

Kidney International, Vol. 61 (2002), pp. 1393–1406
ION CHANNELS – MEMBRANE TRANSPORT – INTEGRATIVE PHYSIOLOGY
Tubular and cellular localization of the cardiac L-type calciumchannel in rat kidney
PEI-LIN ZHAO,1 XI-TAO WANG,1 XUE-MEI ZHANG, VALERIU CEBOTARU, LIUDMILA CEBOTARU,GANG GUO, MARCELO MORALES, and SANDRA E. GUGGINO
Department of Medicine, Johns Hopkins University School of Medicine, Baltimore, Maryland, USA, and Instituto deBiophysica Carlos Chagas, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil
Tubular and cellular localization of the cardiac L-type calcium and skeletal muscle, brain and endocrine organs wherechannel in rat kidney. they function to increase calcium influx or to transduce
Background. The mRNAs of several types of calcium chan- voltage changes [1]. Six types of �1 subunit have beennels have been identified in intact rat kidney, and L-type cal-cloned including three L-type channels (�1S, �1C, and �1D),cium channels cause changes in intracellular calcium in primarythe �-conotoxin-GIVA-sensitive N-type (�1B), the �-aga-cultures of distal tubule cells. The aim of this study was to
evaluate the tubular and cellular distribution of the �1C subunit toxin-IVA-sensitive P/Q-type (�1A), and the resistantof the L-type calcium channel in intact kidney. R-type (�1E). The L-type calcium channel complex is
Methods. RT-PCR and Northern blot analysis were used to composed of multiple subunits [2] but the �1 subunitassess the regional abundance of the mRNA of this channel.comprises the main conduction core, the voltage sensorImmunocytochemistry combined with confocal microscopy andand also contains the drug binding sites for dihydropyri-surface biotinylation were applied to determine the tubular
and cellular localization of the protein. dine antagonists such as nitrendipine or nifedipine orResults. Northern blot and RT-PCR analysis indicated that the agonist Bay K 8644, for phenylalkylamines such as
the mRNA of the �1C subunit of the cardiac L-type calcium verapamil, and for benzothiazipines such as d-cis-diltia-channel was present in whole rat kidney, kidney tubules andzem [1, 3]. When the �1 subunit is expressed alone, thekidney cell lines. Western blot of lysates from whole kidney,membrane current density is low and voltage activationkidney tubules or cell lines revealed bands of �190 kD for the
�1C subunit and �60 kD for the �3 subunit. Confocal immuno- is slower than in endogenous tissue. The other subunitshistochemistry indicated that the �1C subunit of this channel was (�2, �, �, and �) increase current activation and inactiva-co-expressed in cells of the distal tubule that express calbindin- tion of the conduction core. Calcium channels also areD28K, but not in intercalated cells. The �1C subunit was also
expressed in non-excitable cells [4–8] and the mRNA ofhighly expressed in both outer and inner medullary collectingseveral types of calcium channels have been identifiedducts. Serial confocal microscopic images or surface biotinyla-
tion experiments determined that the channel was predomi- in both the cortex and medulla of intact rat kidney [9].nantly on the basolateral membrane but had some distribution In distal tubule cells, Barry et al [10] found that antisenseon the apical membrane. to the �1D subunit inhibits a parathyroid hormone (PTH)-Conclusions. The distribution and cellular localization of the
mediated calcium influx, whereas antisense to the �1C�1C subunit of cardiac L-type calcium channel suggest it issubunit inhibits the diuretic-mediated calcium influx. Inprobably involved in intracellular and membrane calcium sig-
naling. the cortical collecting duct calcium signaling is mediatedby �-adrenergic receptors [11]. In medullary thick as-cending limb cells, L-type calcium channels also have
The family of depolarization-gated calcium channels been implicated in increases in intracellular calcium afteris distributed in excitable tissues including heart, smooth volume expansion [12]. Finally, in inner medullary col-
lecting ducts, vasopressin receptors cause elevation ofintracellular calcium [13].
1 Drs. Zhao and Wang contributed equally to this project. Recently, two other calcium channels from the TRPfamily have been cloned and localized to the distal tubuleKey words: cortical collecting duct, distal tubule, medullary collecting
duct, calcium signaling, intracellular signaling. of the kidney [14, 15]. Both ECaC [14] and CAT-2 [15]are localized in the principal cells of the distal tubuleReceived for publication August 15, 2001where 1�,25-dihydroxyvitamin D3 responsive transepi-and in revised form November 9, 2001
Accepted for publication November 9, 2001 thelial calcium transport occurs, and both of these pro-teins colocalize with the major 1�,25-dihydroxyvitamin 2002 by the International Society of Nephrology
1393

Zhao et al: �1C subunit of the L-type calcium channel1394
D3-responsive component of the transepithelial calcium Eagle’s medium and Ham’s F12 medium containing 10%FBS and 50 U/mL penicillin and 50 �g/mL streptomycin.transport system, the intracellular calcium buffering pro-
tein calbindin-D28K. Evidence suggests that these chan- For extraction of RNA for Northern blot or proteins forWestern blot, cells were split in a 1:4 ratio and platednels are the entry pathway for transepithelial calcium
transport including calcium permeability, and activation onto T75 flasks without coating and grown to two dayspost-confluence. For immunocytochemistry or surfaceby hyperpolarization, inhibition by cadmium and low
pH, but little effect of blockers of L-type calcium chan- biotinylation cells were split and plated at 1:4 density onhigh pore density PET track-etched transwells (Becktonnels [15]. However, because the �1C subunit has been
implicated by Barry et al to cause changes in intracellular Dickinson Falcon, Franklin Lakes, NJ, USA) and grownthree to five days after confluence.calcium in primary cultures of distal tubule cells [10], we
wondered whether the channel also would be colocalizedNorthern blotwith other components of the transepithelial calcium-
transporting pathway in the intact kidney. Such colocali- The probe for the calcium channel was generated byRT-PCR from rat brain. Primers for �1C, bp 5851-5871zation would suggest that transepithelial calcium trans-
port mediated by ECaC or CAT-2 may be modulated and bp 6360-6341 were designed using sequence fromrbC-II �1C Accession # M67515, to give an expected PCRby calcium influx via L-type calcium channels. However,
since the mRNA of the �1C subunit was localized in the product of 510 bp. The area chosen for the probe be-tween amino acid 1864 to 2033 is relative to Accessionmedulla where the highest abundance was in the inner
medulla [9], calcium influx via this channel also may # AAA18905. This region was chosen because it sharedonly 51 amino acids with �1D and 22 amino acids withfunction to alter transport in tubule segments in this
region. In this regard, the mRNA of the pore forming �1S. Our procedures for probe preparation, RNA samplepreparation and blot hybridization have been previouslysubunit of the �1G channel (a T type channel) is distrib-
uted in high abundance in the inner medulla and in described [17, 18].IMCD tubules [16], and using immunohistochemistry
RT-PCRthis channel was localized to the membranes of connect-ing tubules cells, distal convoluted tubule cells and the Reverse transcription-polymerase chain reaction was
performed on cDNA made from RNA isolated from ratinner medullary collecting duct cells, suggesting that morethan one pathway for calcium influx exists in these cells. kidney cortex, outer medulla, and inner medulla. Rat brain
was used as a positive control. In addition RT-PCR wasOur results show that the mRNA for the �1 subunitof the cardiac L-type calcium channel is present in the performed on isolated rat kidney tubules. In order to iso-
late the tubular RNA, male Sprague-Dawley rats, aboutcortex and medulla of the intact rat kidney and in amouse inner medullary collecting duct cell line. Using 150 g, were anesthetized and then the left kidney was
perfused via the abdominal aorta using a perfusion/dis-reverse transcription-polymerase chain reaction (RT-PCR), mRNA was found in the proximal straight tubule, section solution containing 133 mmol/L NaCl, 5 mmol/L
KCl, 1 mmol/L Na2HPO4, 3 mmol/L Na acetate, 1 mmol/Lthe cortical collecting duct and in medullary collectingduct tubules. The protein was localized in cells of the CaCl2, 1.2 mmol/L MgCl2, 0.5 mmol/L HEPES and 5.5
mmol/L glucose in 0.1% diethylpyrocarbonate treateddistal tubule that also express calbindin D28K and in cellsof the medullary collecting duct where its apical and water adjusted to pH 7.4, followed by 30 mL perfusion/
dissection solution supplemented with 100 mg collagen-basolateral membrane distribution would allow it to par-ticipate in calcium signaling. The �3 subunit also is pres- ase B and a 0.1% bovine serum albumin (BSA; Boeh-
ringer Mannheim, Mannheim, Germany). Once the tu-ent in inner medullary collecting duct tubules, in themouse inner medullary collecting duct cell line, and in bules were dissected they were added to a pre-chilled
sterile 1.5 mL Eppendorf tube containing 1 mL TRIzol.a mouse medullary thick ascending limb cell line.The tubules were homogenized immediately upon dis-section using a tissue homogenizer (Kontes Glass Co,
METHODSVineland, NJ, USA) designed for Eppendorf tubes. The
Cells RNA was isolated according to the TRIzol protocol(Gibco BRL, Gaithersburg, MD, USA).The mouse medullary thick ascending limb cell line
(mTAL), derived from an SV40-transgenic mouse, was The primers for amplifying PCR products were takenfrom the nucleotide sequence of the mRNA as outlinedgrown in Dulbecco’s modified Eagle’s medium containing
10% fetal bovine serum (FBS), 50 U/mL penicillin and for the calcium channel probe used in the Northern blot.The area chosen for these primers, amino acids YQDD50 �g/mL streptomycin. The mouse inner medullary col-
lecting duct cell line (IMCD; ATCC CRL-2123; ATCC, ENR and APGRQF relative to Accession number AAA18905, is not shared by �1D, which has the sequencesRockville, MD, USA) derived from an SV-40 transgenic
mouse, was grown in a 1:1 mixture of Dulbecco’s modified TWSRQNY and NKNSDK, respectively, in the same

Zhao et al: �1C subunit of the L-type calcium channel 1395
area or �1S that has HVDKLER, and the reverse primer blots were visualized by the SuperSignal�West Dura ex-tended duration substrate (Pierce, Rockford, IL, USA).has a deleted sequence in this region. RT-PCR for �1C
subunit of L-type calcium channel was performed follow- The free peptide utilized to generate each antibody waspre-incubated with the primary antibody at 5 �g/mL toing the protocols described previously [17, 18].determine the specificity of the labeling.
Western blotImmunocytochemistryThe calcium channel �1C subunit antibody (ACC-003)
was obtained from Alomone Labs Ltd. (Jerusalem, Is- The identification of various tubular segments in thekidney was primarily based on their general anatomicalrael). This affinity-purified rabbit polyclonal antibody
was raised to the peptide sequence TTKINMDDLQP and histological features combined with co-immunolo-calization of several specific markers. The anatomicalSENEDKS corresponding to amino acids 818-835 of rat
�1C subunit (Accession # JH0426). A BLAST search at and histological features of various tubular segmentswere examined on hematoxylin and eosin (H&E) stainedthe National Center for Biotechnology Information re-
vealed that this amino acid sequence was specific for �1C. sections (data not shown) that were adjacent to the im-munostained sections. The criteria used to identify theWhen the sequences of the other L-type calcium channel
isoforms were lined up no more than seven dispersed tubular segments were done according to definitions out-lined by Tisher and Madsen [21]. In addition, we usedamino acids matched this sequence for �1S and no more
than five dispersed amino acids matched for �1D. A like calbindin-D28K as a marker for calcium transporting distaltubules. Calbindin-D28K was chosen since it is well docu-sequence was not found in any other voltage-gated cal-
cium channel including N, R or P/Q types. The antibody mented that calbindin-D28K is expressed in the distal tu-bule of many species including chicken, rat, rabbit, pigto the �3 subunit of the calcium channel (ACC-008) was
also obtained from Alomone Labs Ltd. This affinity puri- and human [22–26]. In human and rat kidney, calbindin-D28K has been immunolocalized in the distal convolutedfied rabbit polyclonal antibody was raised to the peptide
sequence DRNWQRNRPWPKDSY corresponding to tubule, the connecting tubule and the initial segmentof the cortical collecting duct. Connecting tubules andamino acids 463-477 of the rat brain �3 subunit (Acces-
sion # AAB32350.1). Rat heart [2] or rat brain [19] was cortical collecting tubules were recognized by the ap-pearance of the intermingled intercalated cells [27, 28],used as a positive control tissue for the �1C and �3 subunit
antibodies, respectively. which were identified using fluorescence staining withpeanut lectin [29, 30]. Discrimination between connect-Our procedures for protein sample preparations and
Western blot have been described previously [18, 20]. To ing tubules and cortical collecting tubules, though some-times difficult, was based on their location in the cortexevaluate the distribution of protein in individual kidney
tubules, male Sprague-Dawley rats weighing about 150 g and in particular by the superficial T branches, indicatingtransitions between these segments [31]. Furthermore,were anesthetized, then the left kidney was perfused via
the aorta using Hank’s balanced salt solution (GIBCO we co-localized �1C L-type calcium channel with Tamm-Horsfall protein using a sheep anti-Uromucoid antibodyBRL) containing a cocktail tablet of protease inhibi-
tors (Roche Diagnostics, Indianapolis, IN, USA). After (Biodesign International, Kennebunk, ME, USA) toidentify the medullary thick ascending limbs (mTALs;this, the tubules were dissected on ice in the same solu-
tion, and then the isolated kidney tubules were homoge- data not shown).The calcium channel �1C subunit antibody (ACC 003)nized in Eppendorf tubes in the RIPA buffer [18, 20].
The proteins were separated by 5% sodium dodecyl used for immunocytochemistry was the same as describedabove for the Western blot. Tissue fixation, section prep-sulfate-polyacrylamide gel electrophoresis (SDS-PAGE;
�1C subunit) or 7.5% SDS-PAGE (�3 subunit). Then pro- aration and immunostaining were performed followingroutine protocols as described previously [18, 20]. Ratteins were transferred to polyvinylidene difluoride (PVDF)
membranes (Bio-Rad, Hercules, CA, USA) in a Tris/ kidney and brain were fixed by perfusion through theaorta with 4% paraformaldehyde in 0.1 mol/L phosphateglycine transfer buffer (Bio-Rad) containing 5% metha-
nol and 0.007% SDS (�1C subunit) or 10% methanol (�3 buffer, followed by a post-fixation in the same solutionovernight. Both frozen sections and paraffin sectionssubunit). Membranes were blocked with 5% nonfat milk
in Tris buffered saline containing 0.05% Tween 20 (TTBS) were used for immunostaining. To determine specificityof the staining, 1 �g of the antigenic peptide was addedfor one hour, and then were incubated overnight with pri-
mary antibody at 1.5-3 �g/mL (�1C subunit) or 0.3 �g/mL per microgram of primary antibody (3 or 6 �g/mL) in athree hour preincubation. The subcellular distribution of(�3 subunit) diluted with blocking buffer. After washes,
membranes were incubated with horseradish peroxidase- this subunit was analyzed with a confocal scanning unit(LSM 410; Carl Zeiss, Oberkochen, Germany) by evalu-conjugated secondary antibody (Amersham Life Sci-
ence, Arlington Heights, IL, USA) diluted 1:10,000 with ating at least 10 tubule segments in a series of x-y cuts(z sections) at 1 �m through the tubules.blocking buffer for one hour at room temperature. The

Zhao et al: �1C subunit of the L-type calcium channel1396
The mouse monoclonal antibody to calbindin-D28K ob- from which 1/20 was taken as an estimate of the input,and the rest was incubated with 200 �L streptavidin-tained from Sigma (St. Louis, MO, USA) was generated
using purified chicken gut calbindin-D28K as the immuno- agarose beads (Pierce). To ensure the complete recoveryof biotinylated protein, the extraction with streptavidin-gen. This antibody reacts specifically with calbindin-D28K
in brain and kidney tissues from human, monkey, rabbit, agarose beads was performed two times. The first extrac-tion was performed overnight and the second for tworat, mouse, chicken. Rat cerebral cortex was used as pos-
itive control because the easily identified Purkinje cells hours. After washing the beads with RIPA buffer fivetimes, 50 �L of 2 � Laemmli sample buffer (Bio-Rad)(data not shown) are well established to have calbin-
din-D28K. This antibody was used at a dilution of 1:500 containing 200 mmol/L dithiothreitol (DTT) was addedand incubated at 37�C for one hour to elute the biotinyl-with a secondary FITC-conjugated donkey anti-mouse
IgG at a dilution of 1:200 (Jackson Immunoresearch, ated protein. The elution process was also performedtwo or three times to ensure complete recovery. AfterWest Grove, PA, USA). The tubular colocalization of
the antibodies to calbindin-D28K and to the �1C subunit centrifugation at 14,000 � g for two minutes, the eluateswere collected for Western blot analysis using the �1Cwere done by scoring about 2000 tubules in coronal sec-
tions of rat kidney cortex. The cellular colocalization of subunit antibody (ACC 003) as described above and amonoclonal antibody to the �1 subunit of Na,K-ATPasethe antibodies to calbindin-D28K and to the �1C subunit
was determined by scoring over 500 random cells in the (Upstate Biotechnology, Lake Placid, NY, USA). Thedensitometry analysis was performed using the Scioncortex. The fluorescein-labeled, affinity-purified, peanut
agglutinin was obtained from Biomeda (Foster City, CA, Image (Scion Corporation, Frederick, MD, USA).USA) and used at a dilution of 1:300. The tubular andcellular colocalization of peanut lectin and the �1C sub-
RESULTSunit were preformed as described above for calbindin-
mRNA of �1C in kidney and kidney cell linesD28K and the �1C subunit. Peanut lectin and calbindin-D28K
were expressed in tubules together 87% of the time. Hybridization by Northern blot probe specific to the�1C subunit of the L-type calcium channel showed twoDouble fluorescence sections were analyzed with an in-
verted microscope coupled to a confocal scanning unit bands one at 9.5 kb and another at about 12 kb comparedto the top molecular weight marker at 9.5 kb and the(LSM 410; Carl Zeiss).lower molecular marker at 7.5 kb (Fig. 1A, lane M).
Cell surface biotinylation Hybridization of these bands was weak in the cortex(Fig. 1A, lanes C and C), stronger in the outer medullaInner medullary collecting duct (IMCD) cells were
plated on six-well cell culture inserts (Beckton Dickinson (Fig. 1A, lane OM) and the strongest kidney expressionwas in the inner medulla (Fig. 1A, IM). Inner medullaryFalcon). The cells were analyzed three to five days after
confluence. The cell surface biotinylation assay was car- collecting duct cells expressed comparable mRNA to thekidney cortex (Fig. 1A, lanes IMCD cells). The heartried out essentially as previously described [32]. The
filter-grown monolayers were placed on the ice and expressed similar mRNA when the total RNA wasgreatly diluted compared to that used in the kidney lanes.washed three times with ice-cold Dulbecco’s phosphate
buffer saline (Gibco BRL) containing 1.36 mmol/L CaCl2 The mRNA for the �1C subunit was previously reportedin both cortex and medulla of the kidney with a lesserand 1 mmol/L MgCl2 (DPBS-Ca-Mg). Then the cells
were incubated with 1 mg/mL NHS-SS-biotin (Pierce) amount in the cortex and a larger amount in the innermedulla [9]. Figure 1B, top panel, shows RT-PCR prod-in DPBS-Ca-Mg for 30 minutes at 4�C with gentle shak-
ing. For apical biotinylation, 1 mL biotin solution was ucts of the �1C subunit of the L-type calcium channelfrom rat brain, kidney cortex, outer medulla and inneradded to the apical chamber, and the basolateral com-
partment received DPBS-Ca-Mg lacking NHS-SS-biotin. medulla. The size of the product was 510 bp as expectedfrom the primer sequences and sequencing the productsFor basolateral biotinylation, 2 mL biotin solution was
added to the basal chamber, and the apical compartment in both directions determined that the sequence wasthat of rat �1C. RT-PCR products from tubules segmentsreceived DPBS-Ca-Mg lacking NHS-SS-biotin. After Bi-
otinylation, the cells were rinsed once and washed twice isolated from rat kidney are shown in Figure 1B, bottompanel. The lane labeled M shows the 100 bp ladder. Thewith 100 mmol/L glycine in DPBS-Ca-Mg to quench
the reaction. After an additional two washes with cold brightest molecular weight marker in the middle of theblot is 600 bp. The RT-PCR products ran near the 500DPBS-Ca-Mg, the monolayers were excised from the
filter cup with a razor blade. The monolayers were then bp marker as expected. The product was present in alltubule segments tested, including the glomeruli, proxi-scraped and solubilized with RIPA buffer containing the
protease inhibitors as used for protein isolated for the mal convoluted tubules (PCT), proximal straight tubules(PST), medullary thick ascending limb (mTAL), corticalWestern blots. The lysates were centrifuged at 10,000 � g
for 10 minutes at 4�C. The supernatants were collected, collecting ducts (CCD), outer medullary collecting ducts

Zhao et al: �1C subunit of the L-type calcium channel 1397
Fig. 1. (A) Northern blot of �1C in rat heart,rat kidney, and kidney cell lines using a cDNAprobe. The probe was made from bp 5851-6360 relative to Accession #M67515. The toppanel shows the Northern blot hybridizationshowing two hybridization bands at 9.5 kb andanother above 12 kb. The molecular makerson the left (M) are 7.5 and 9.6 kb. In thebottom panel the RNA is stained with methy-lene blue to show the 28S and 18S bands. Inthis lower blot the molecular marker on theleft (M) shows the 28 S band at about 4.4 kband the 18S band between 2.4 kb and 1.8 kb.Abbreviations are: C, rat kidney cortex; OM,rat outer medulla; IM, rat inner medulla;IMCD cells, the mouse inner medullary col-lecting duct cell line; H, rat heart total RNA.There is much less mRNA for the �1C subunitin kidney when compared to heart becausethe hybridization was weaker in kidney com-pared to heart when less total mRNA wasloaded. Forty micrograms of total RNA wasloaded in each lane of kidney RNA whereasthe four lanes of rat heart (H) had 2, 5, 10and 20 �g of RNA loaded. (B) Distributionof mRNA of �1C in whole kidney and tubulesegments of the kidney as demonstrated byRT-PCR. (Top) PCR products were amplifiedfrom all areas of the kidney. Abbreviationsare: M, molecular weight marker showing thatthe PCR product runs at 510 bp as expected;WB, the PCR reaction run with water insteadof cDNA as a negative control; Br, brain; C,cortex; OM, outer medulla; IM, inner medulla.(Lower) PCR products were amplified fromeach tubule segment of isolated tubules of therat kidney and also from rat brain (Br). Abbre-viations are: M, molecular weight marker (thebright band in the middle of the gel is the 600bp marker); WB, water blank (this PCR wasrun with water rather than cDNA as a negativecontrol); Br, rat brain as a positive control; G,glomerulus; PCT, proximal convoluted tubule;PST, proximal straight tubule; mTAL, medul-lary thick ascending limb; CC, cortical collect-ing duct; OMC, outer medullary collectingduct; IMC, inner medullary collecting duct.
(OMCD), and inner medullary collecting ducts (IMCD). molecule [2]. These two molecular weights also corre-sponded to the two molecular weight mRNAs in theThe assay was not optimized to be quantitative.Northern blot, suggesting that there is some full length
�1C subunit in heart, kidney and kidney cell lines protein in the IMCD and mTAL cells resulting in thehigher molecular weight protein. Lanes 9 to 14 show thatImmunoblots of rat kidney tubules and rat heart ly-the peptide to which the antibody was raised, completelysates revealed a band about 190 kD (Fig. 2A). The inten-displaced the bands near 190 and 240 kD, suggestingsity of the band in rat mTAL tubules (lane 1) and ratthat the antibody was specific.CCD tubules (lanes 2, 3) was much less than that of
IMCD tubules (lanes 4, 5). Mouse mTAL cells grown �3 subunit in brain, kidney and kidney cell linesin culture (lane 6) and mouse IMCD cells grown in cul- Figure 2B is a Western immunoblot of the �3 subunitture (lane 7) also have less intense bands, whereas rat of the L-type calcium channel. This antibody stained aheart (lane 8) had the most intense band even though band at about 60 kD in IMCD tubules (lane 1), mTALhalf the amount of protein compared to the kidney was cultured cells (lane 2), IMCD cultured cells (lane 3), andloaded. The most prominent band was about 190 kD in rat brain as a positive control (lane 4) [19]. The bandsall preparations and the second upper band above the stained by the anti-�3 antibody were displaced by the220 kD marker was consistent with a molecular weight peptide to which the antibody was raised in mTAL cells
(lane 5), IMCD cells (lane 6) and brain tissue (lane 7).of about 240 kD, which corresponded to the full length

Zhao et al: �1C subunit of the L-type calcium channel1398
Fig. 2. (A) Identification of the �1C subunitof the L-type calcium channel in an immu-noblot of rat kidney tubules and kidney celllines and rat heart using an antibody to theintracellular loop between domains II and III(ACC-003). Lanes 1 and 9, medullary thickascending limb (mTAL) tubules. Lanes 2, 3and 10, cortical collecting duct (CCD) tubules.Lanes 4, 5 and 11, inner medullary collectingduct (IMCD) tubules. Lanes 6 and 12, mTALcells. Lanes 7 and 13 are IMCD cells. Lanes8 and 14, rat heart (15 �g per lane) as a control.All other lanes had 30 �g of protein addedper lane. As shown in lanes 1 through 8, the�1C protein is about 200 kD, and there wasmore expression of the �1C subunit in IMCDtubules and IMCD cultured cells. Lanes 9through 14 show that the peptide, to which theantibody was raised, completely displaced thebands near 200 kD. (B) Western blot of the cal-cium channel �3 subunit protein in rat IMCDtubules and cultured mTAL and IMCD cells.Lane 1, the affinity purified anti-�3 subunitantibody stained a 60 kD protein in rat IMCDtubules. The antibody stained a similar molec-ular weight protein in mTAL cells in Lane 2and in IMCD cells in Lane 3. Lane 4, rat braintissue used as a positive control for the �3 sub-unit. This band is about 60 kD as previouslyshown. There was stronger expression in IMCDcells than in mTAL cells. The bands stainedby the anti-�3 subunit antibody were displacedby the peptide to which the antibody wasraised in mTAL cells (lane 5), IMCD cells(lane 6) and brain tissue (lane 7). Each lanehad 10 �g of total protein added.
Immunocytochemical localization of the �1C subunit II). The staining in the cortical thick ascending limbs wasin kidney comparable to that in PTs and appeared intracellular. In
the medulla (Fig. 3B), the bright staining was in variousImmunocytochemical localization of the �1C subunitsegments of the OMCD (panel V) and the IMCD (panelof the L-type calcium channel in the kidney indicates aVI), which were identified as tubules with a thin cyto-distribution to tubule segments in both the cortex andplasm and a large inner tubular diameter. The stainingmedulla (Fig. 3). To confirm the identity of the variousin the medullary thick ascending limb was weak andtubule segments, H&E staining (not shown) was per-distributed in the cytoplasm. Little staining was foundformed in the adjacent sections to the immunofluores-in the thin limbs of Henle’s loop, but it could be thatcent section. Two types of immunostaining for the �1Cthe fluorescence was below the limits of detectability insubunit were easily identified. The first type of stainingthese thin segments.was bright, intense, and sharp and closely associated with
Double labeling of �1C and calbindin-D28K on the samemembranes (Fig. 3A, II, III, IV; Fig. 3B, V, VI), whereaskidney section showed that most of the cortical brightthe second type of staining was weak and distributedtubules that stained for �1C colocalized with tubules thatevenly in the cytoplasm (Fig. 3A, II). Both the brightexpress calbindin-D28K. It is well known that calbindin-and diffuse staining was completely displaced by peptideD28K is expressed in the distal tubule of many speciespre-absorption (Fig. 3B, VII), which suggests that theseincluding chicken, rat, rabbit, pig and human [22–25].two types of staining were specific for the �1C subunit.Murine calbindin-D28K mRNA has been localized to theIn the kidney cortex, the bright staining was localizeddistal convoluted tubule, the cortical connecting tubule,to the distal convoluted tubules (DCT), the connectingand the cortical collecting ducts [26]. In human and rattubule (CT) or the cortical collecting duct (CCD; Fig.kidney, calbindin-D28K has been immuno-localized in the3A, II, III, IV). In these cells, the staining was mainlydistal convoluted tubule, the connecting tubule and thelocalized to the basolateral membrane. but some wasinitial segment of the cortical collecting duct. Rat calbin-also present in the apical membrane. The weaker, moredin-D28K was previously localized, in addition to the distaldiffuse staining in the cortex was primarily distributed
in the cytoplasm of the proximal tubules (PT; Fig. 3A, tubule of the kidney, to cerebellum Purkinje cells and

Zhao et al: �1C subunit of the L-type calcium channel 1399
to dentate gyrus granule cells [33]. As a positive control this distribution in a more quantitative manner using sur-face biotinylation. In order to determine the percentagefor the calbindin-D28K antibody used in this study, we
localized staining to Purkinje cells (not shown). Of all of the channel that is targeted to the plasma membranescompared to that inside IMCD cells, surface biotinylationthe �1C bright-stained tubules in the cortex 71% were
colocalized with calbindin-D28K staining (Fig. 4A) and was performed on both the apical and basolateral mem-branes of polarized, cultured IMCD cells grown to fivethe remaining 29% of the red staining for the �1C subunit
was alone, mainly distributed in the medullary rays (Fig. days post-confluence on permeable transwell supports.Surface biotinylation of IMCD cells showed that although4A, DCT). Of the total population of calbindin-D28K-
labeled tubules, 48% were not colocalized with the bright some of the �1C subunit was distributed in the membranefractions a major portion of the channel protein was�1C stained tubules. These tubules that stained only for
calbindin-D28K were mainly distributed in the cortical present inside the cell (Fig. 5A). Figure 5A lanes 2 and 3show 1/20 of the total cellular �1C subunit in the apicallabyrinth. These results suggest that in the cortex, the
bright �1C staining was localized to the late portion of and basolateral biotinylation experiments, respectively.Figure 5A lanes 4 and 6 show the total apical biotinylatedthe DCT, the CT and most of the length of the CCD.
In those tubules where calbindin-D28K and �1C colo- �1C subunit and lanes 5 and 7 show the total basolateralbiotinylated �1C subunit. In this experiment about 1% ofcalized, 92% of the cells had staining for both calbindin-
D28K and the �1C subunit (Fig. 4B, C and D), suggesting the total �1C subunit is on the apical membrane whereas5.4% is on the basolateral membrane. (The quantitativethat �1C was mainly distributed in the calbindin-D28K posi-
tive cells. The calbindin in these cells buffers the hor- details of the densitometry analysis are shown in thelegend to Fig. 5). Figure 5B lanes 2 and 3 show 1/20 of themone-regulated calcium entry that is part of active trans-
epithelial calcium transport [14]. As shown in Figure C total cellular Na,K-ATPase in the apical and basolateralbiotinylation experiments, respectively. Figure 5B lanesand D, the subcellular localization of calbindin-D28K and
the �1C subunit was different. Whereas calbindin-D28K is 4 and 6 show the total apical biotinylated Na,K-ATPaseand lanes 5 and 7 show the total basolateral biotinylatedknown to be primarily in the cytoplasm [22–25], the �1C
subunit was localized more in the plasma membrane. Na,K-ATPase. In this experiment about 4.1% of the totalNa,K-ATPase is on the apical membrane whereas 36%The distribution of the �1C subunit was mainly in the
basolateral membrane but also in the apical membrane; is on the basolateral membrane. The densitometry analy-sis of immunoblots from three separate experiments re-however, as revealed in multiple x-z sections that were
analyzed by confocal microscopy, the �1C subunit ap- vealed that 5.3 0.8% (mean SE) of the total cellularcontent of the �1C subunit resided in either the apical orpeared to be most often in basolateral membranes. Fur-
ther confirmation of the cell type where the �1C subunit basolateral plasma membranes, whereas 40 1.8% ofthe total Na,K-ATPase resided in the membrane frac-resides was illustrated by double-labeling the �1C subunit
with peanut lectin. On the same section, only 20% of tions. Of the �1C subunit that was inserted into the plasmamembrane, 17 1.1% resided at the apical membranethe cells showed colocalization of both the �1C subunit
and peanut lectin. Peanut lectin has been used previously whereas 83 1.1% was inserted into the basolateralmembrane. In the same three experiments, 6.7 3.6%as a marker of intercalated cells in the cortical collecting
duct [29, 30]. In the rat kidney, these intercalated cells of the Na,K-ATPase was in the apical membrane and93 2.9% was on the basolateral membrane, suggestingare present in the later portion of the distal convoluted
tubule, the connecting tubules and the cortical collecting that the surface biotinylation assay itself has a small finiteleak or error because it is well documented that the Na,duct [27, 28]. Of the total tubules that stained for peanut
lectin, 89% were colocalized in tubules that also con- K-ATPase is predominantly in the basolateral mem-brane [34]. The estimate of the apical membrane distri-tained �1C staining (Fig. 4 E, F). The fact that there was
a higher percentage of tubular, although not cellular, bution of the Na,K-ATPase using the surface biotinyla-tion assay was probably an overestimate for two reasons.colocalization of the �1C subunit with peanut lectin as
compared to calbindin-D28K also supports the idea that First, because the density of the Na,K-ATPase is so highon the basolateral membrane not all of the basolateralthe �1C staining is localized in the later portion of distal
convoluted tubule, the connecting tubule and most of Na,K-ATPase was removed in the three elutions (notshown), although this efficiency is greater than other re-the cortical collecting duct in rat kidney cortex.
The staining of �1C subunit of the calcium channel in ports of 5% efficiency [35], probably resulting in an un-derestimate of the membrane fraction of Na,K-ATPasethe outer and inner medullary collecting duct cells was
very bright and appeared to be both intracellular and in compared to total Na,K-ATPase. This also causes anoverestimate of the apical distribution compared to baso-the plasma membrane. Because the staining in the IMCD
cells appeared to be both in the membranes and inside lateral distribution. In comparison, all of the �1C subunitprotein was removed by the second elution (Fig. 5, lanesthe cell and the intracellular staining could be a result of
fluorescence bleeding in these small cells, we investigated 6, 7) and all of the apical Na,K-ATPase was removed

Zhao et al: �1C subunit of the L-type calcium channel1400
Fig. 3. See legend next page.

Zhao et al: �1C subunit of the L-type calcium channel 1401
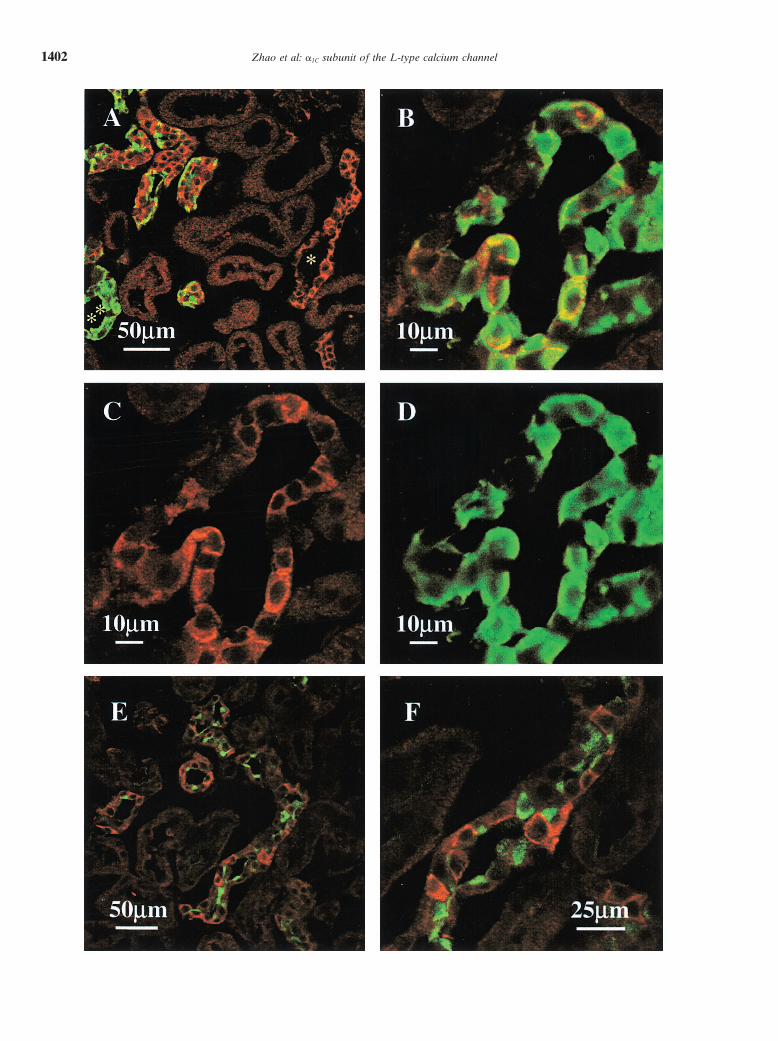
Fig. 3. (A) Immunohistochemical localization of the �1C subunit of the L-type calcium channel in whole rat kidney. The primary antibody, apeptide purified rabbit polyclonal antibody (ACC-003) to �1C (Alomone, Israel) was diluted 1:100 (3 �g/mL) and the secondary antibody goatanti-rabbit IgG cy3 (red) was diluted 1:200. The nuclei are counterstained with Hoechst 33342 (green). (panel I) Localization of the �1C subunitof the L-type calcium channel in the kidney cortex at a low-magnification illustrates that positive staining of the �1C subunit antibody distributesin the cortical labyrinth (**) around the corpuscles and in the medullary ray (). The boxes in Fig. 3A panel I that are labeled II, III, and IVare shown in higher magnification in Fig. 3A panels II, III, IV, below. (Panel II) The bright staining of �1C was in the cortical labyrinth in isolatedshort tubule segments of small diameter near the glomerulus indicating the distal convoluted tubule (*). This staining was mainly distributed inthe basolateral membranes as analyzed by X-Z sections (not shown). The staining in tubules of larger diameter that indicated proximal tubules(**) was less intense, diffuse and located in the cytoplasm. G denotes a glomerulus that also shows diffuse staining. (Panel III) The bright stainingof �1C was localized mainly in the plasma membrane of the tubules of the cortical connecting tubule (�) that were recognized by the position inthe cortex and by the superficial T branch indicating transition from connecting tubules to cortical collecting ducts. The staining also was localizedprimarily in basolateral membranes as analyzed by X-Z sections (not shown). (Panel IV) In the medullary rays, bright staining was distributed intubules with a large lumen diameter suggesting tubules of the cortical collecting duct. The lumen is marked (��). (B) High power magnificationof tubules from the medulla of the rat kidney. (Panel V) Tubules of the OMCD and (panel VI) tubules of IMCD. In the medulla, the brighteststaining was in various segments of the medullary collecting duct. The staining in IMCD was most intense and distributed not only in the membranefraction but also in the cytoplasm. (Panel VII) The peptide to which the antibody was raised displaced all of the bright staining for �1C in thecortex and the medulla including all of the weaker staining in the proximal tubules.
in two elutions (not shown). Secondly, using confocal subunit of the calcium channel in the apical membraneis greater than that of the Na,K-ATPase, it suggests thatmicroscopy we confirmed that the distribution of the
Na,K-ATPase is solely in the basolateral membrane with there is a finite distribution of the calcium channel inthe apical membrane of IMCD cells. Gottardi, Dunbarno apical membrane distribution (cultured kidney cells
grown one week post-confluence on permeable trans- and Caplan previously showed that biotinylation effi-ciency sometimes depends on pH and that alkaline pHwells). In this case, the immunofluorescent staining using
the same antibody to the Na,K-ATPase used in these often improved biotinylation efficiency, but that someproteins were better biotinylated at neutral pH [32].biotinylation experiments was analyzed using confocal
microscopy (data not shown). Under the same growth Given that the biotinylation of basolateral Na,K-ATPasewas not completely efficient, the main focus of biotinyla-conditions (almost one week post-confluence grown on
transwells) IMCD cells also showed apical tight junc- tion in this study was to determine the percentage of the�1C subunit of the L-type calcium channel protein presenttions as indicated by ZO antibody staining visualized
by confocal microscopy (data not shown). These addi- on the apical versus basolateral surface, rather than thetotal amount of �1C subunit in the plasma membrane.tional confocal experiments suggest that the cells grown
in culture have established a normal apical and baso- While the total amount of �1C subunit or the Na,K-ATPase in the plasma membranes might be underesti-lateral polarity and complete basolateral distribution of
the Na,K-ATPase. Therefore, the apical distribution of mated, the ratio of �1C subunit on the apical and basolat-eral plasma membrane would not change with pH. Bythe Na,K-ATPase measured in the biotinylation experi-
ment is probably a result of the limits of the biotinyla- comparing the ratio of apical versus basolateral biotiny-lation of �1C with that of Na,K-ATPase on the sametion assay itself and similar apical biotinylation results
for the Na,K-ATPase has been observed before [32]. cells (Fig. 5B), we could determine whether there wasa significant fraction of �1C on the apical membrane.However, because the amount of distribution of the �1C
Zhao et al: �1C subunit of the L-type calcium channel1402

Zhao et al: �1C subunit of the L-type calcium channel 1403
In summary, the �1C subunit of the cardiac isoform of nel mRNA transcripts (Dmca1D and Dmca1A) havethe L-type calcium channel in rat kidney was distributed been found by RT-PCR in Drosophila melanogaster Mal-through all kidney tubules except the thin limbs of pighian tubules, suggesting that L-type calcium channelsHenle’s loop. The staining in glomerulus, proximal tu- must have functional significance in many species [36].bule and thick ascending limb of Henle was weak and The unique distribution of the mRNA for each calciumdiffuse, whereas the staining in the late distal convoluted channel isoform in the kidney suggests that each calciumtubule, the connecting tubule, the cortical collecting duct, channel � subunit isoform is probably coupled to uniqueouter and inner medullary collecting ducts was bright, calcium signaling pathways that in turn affect epithelialintense and mainly membrane associated. The most in- function or blood flow.tense staining was seen in the IMCD, where surface Sequencing of a subcloned transcript from rabbit prox-biotinylation of IMCD cells indicates a greater basolat- imal tubules indicates that the mRNA of the �1C subuniteral than apical membrane distribution. is identical to the rabbit cardiac calcium channel �1C
except for a 33 base pair deletion in the motif 4 S3-S4linker region [37]. Investigations by O’Neil et al indicateDISCUSSIONhigh affinity [3H]dihydropyridine binding sites in apical,
Yu et al showed that rat kidney expresses mRNAbasolateral and microsomal membrane preparations of
transcripts of the three �1 isoforms of the L-type calciumcultured rabbit renal proximal tubules [38], which wouldchannel as well as the P-type calcium channel, but thatbe consistent with the distribution found in proximal tu-the distribution of each of these is different [9]. Thebules by the immunostaining. Cell swelling of rabbit proxi-channel mRNA was located primarily in the cortex andmal tubules induced by hypo-osmotic medium causes anby PCR it showed a distribution in the distal convolutedincrease in intracellular calcium that is inhibited by thetubule. The mRNA of the �1D isoform was equally dis-dihydropyridine or verapamil classes of calcium channeltributed in the kidney cortex, and in the inner and outerblockers [39, 40], and this increase in calcium in responsemedulla. Yu et al also found very small amounts of theto hypo-osmotic medium is also inhibited by antisense�1S subunit mRNA in the inner medulla [9]. They foundoligo-nucleotides to the �1C subunit [37]. Pertinent to thisthat the �1C isoform mRNA was located primarily inobservation, we found that the staining for the protein ofthe medulla with the greatest hybridization in the innerthe �1C subunit in the proximal tubule is mainly intracel-medulla. The Northern blot of �1C mRNA in rat kidneylular. This raises the interesting possibility that the distri-(Fig. 1) also suggests that the major distribution of thebution of the channel could change from intracellular tomRNA for the �1C subunit is in the inner medulla, butmembrane associated, for example, with hypo-osmoticthat this amount of mRNA is significantly less than thatmedium or that the channel could have a heretofore un-in the heart. Using RT-PCR, we also found that the �1C
expected intracellular function. Cell swelling of mTALsubunit calcium channel mRNA is present in cortex, outercells caused by hypo-osmotic medium results in an in-medulla and inner medulla as well as along each tubulecrease in intracellular calcium and a compensatory volumesegment and in the glomerulus. However, based on theregulatory decrease that are inhibited by the presence ofNorthern blot the amount of mRNA in the glomerulus,the calcium channel blockers verapamil or nitrendipine, orproximal tubule, and mTAL is probably much less than in
the inner medulla. Interestingly, two L-type calcium chan- by removal of extracellular calcium or by buffering intra-
�
Fig. 4. Double immunofluorescent staining of the �1C subunit of the L-type calcium channel and calbindin-D28K or peanut lectin in rat kidney.The rabbit polyclonal antibody to �1C was recognized by an anti-rabbit cy3-conjugated secondary antibody that produced the red staining. Themouse monoclonal antibody to calbindin-D28K was recognized by a FITC-conjugated secondary antibody to produce a green stain. Overlap of thetwo stains produces yellow. (A) This view shows a low magnification of �1C-labeled tubules in the rat kidney cortex. Of the tubules that werebrightly stained for the �1C subunit, 71% were colocalized with calbindin-D28K staining and 29% of the tubules stained for the �1C subunit alone(*). Of all the calbindin-D28K positive tubules in cortex, about 48% of the tubules expressed calbindin-D28K alone (**) whereas 52% of the tubuleshad calbindin-D28K colocalized with the �1C subunit. This colocalization suggests that many tubules that contain high amounts of �1C in the kidneycortex are located in the distal tubule. (B) High power view of the cellular distribution of �1C and calbindin-D28K together. This shows that 92%of the cells stained for both �1C and calbindin-D28K. �1C is localized primarily to the basolateral membrane (C ) whereas calbindin-D28K is mainlylocalized in the intracellular compartment (D). Overlap of the two antibodies produces a yellow stain. (C ) The same view as B showing the cellulardistribution of �1C is mainly in the membranes of the cells (the green channel is subtracted). (D) The same view as B, showing the cellular dis-tribution of calbindin-D28K is mainly in the cytoplasm of the cells as previously shown (the red channel is subtracted). (E and F ) Doubleimmunofluorescence staining for the �1C subunit of the L-type calcium channel and peanut lectin in rat kidney. The rabbit polyclonal antibody to�1C was recognized by the cy3-conjugated secondary antibody producing a red stain. A FITC-conjugated peanut lectin produces a green stain. Inall of the tubules stained by �1C subunit, 88% are colocalized in the tubules that show peanut lectin staining. Panel F is a high magnification ofpanel E. The peanut lectin staining was located mainly in the apical region of the intercalated cells. Of the double-positive stained tubules, littleof the staining was in the same cells as noted by the lack of yellow stain, which would indicate overlap. The staining of each color was quitedistinct. This adds an additional indication that the calcium channel was distributed in the principal cells.

Zhao et al: �1C subunit of the L-type calcium channel1404
cellular calcium with BAPTA, suggesting that calciuminflux via L-type calcium channels is a necessary compo-nent of the volume regulatory machinery [12]. L-typecalcium channels also have been identified in the mesan-gial cells of the glomerulus using patch clamp recordingsand fura 2 fluorescence [41], but because the staining ofthe �1C subunit protein was quite weak in the glomerulusperhaps another isoform may account for L-type calciumchannel activity in these cells.
Investigations by Barry et al suggested that the activityof the �1D subunit is correlated to increases in intracellu-lar calcium caused by parathyroid hormone (PTH) [10].On the other hand, antisense oligonucleotides directedto the �1C subunit inhibited the elevation of intracellularcalcium caused by chlorothiazide. These authors hypoth-esized that the two isoforms of the L-type calcium chan-nel cause calcium entry in the distal tubule. However, itis well established that both PTH and thiazides causehyperpolarization of the apical membrane of the distaltubule. Since the L-type calcium channel is not stimu-lated by membrane hyperpolarization but rather bymembrane depolarization, if this channel were in the
Fig. 5. (A) Western blot analyses of a representative biotinylation ex-apical membrane, calcium influx there would be inhib-periment examining the amount of apical and basolateral membrane-
biotinylated �1C subunit of the L-type calcium channel in IMCD cells. ited by PTH and chlorothiazide. By contrast, the recentlyThese results show one representative experiment of three separate identified ECaC1/CAT channels are activated by hyper-experiments performed. Lane 1, 20 �g rat heart homogenate shown as
polarization and are located in the apical membranecontrol; lane 2, 1/20 of input, 114 �g of total IMCD protein for theapical biotinylation experiment (density 2306); lane 3, 1/20 of input, 121 of distal tubules where calbindin-D28K and the sodium�g of total IMCD protein for the basolateral biotinylation experiment calcium exchanger exist [42]. Using Z cuts through many(density 2335); lane 4, apical biotinylated �1C from the first elution and
tubules, we found that the protein of the �1C subunit offirst extraction (density 297); lane 5, basolateral biotinylated �1C fromthe first elution and first extraction (density 1579); lane 6, apical biotinyl- the L-type calcium channel is primarily located on theated �1C from the second elution and first extraction (density 131). The lateral and basal membranes, but some of the stainingdensity of the third elution (not shown) is 15. Lane 7, basolateral
also resides in the apical membrane of the distal tubule.biotinylated �1C from the second elution and first extraction (density832); the density of the third elution (not shown) is 82. The calculated Based on this distribution, this channel would cause thetotal input density of �1C for this apical biotinylation experiment is greatest calcium influx when the basolateral membrane46,120 arbitrary units and the amount of apical biotinylated �1C subunit
of the distal tubule is depolarized. Because intracellulardensity from the three elutions is 443 arbitrary units. About 1% of thetotal �1C subunit found in the apical membrane of these IMCD cells. The calcium causes inactivation of the ECaC channel [42],calculated total input density of �1C subunit for this basal biotinylation calcium influx via the �1C subunit of the L-type calciumexperiment is 46,700 arbitrary units. The total amount of basolateral
channel could actually block calcium entry and transepi-density of the �1C subunit from the three elutions is 2493 arbitrary units.About 5.4% of the total �1C subunit of the calcium channel is found at thelial calcium transport in this tubule segment. Becausethe basolateral membrane of IMCD cells. (B) Western blot analyses cyclic AMP (cAMP) is known to phosphorylate the car-of a representative biotinylation experiment examining the amount of
diac �1C subunit of the L-type calcium channel enhancingapical and basolateral membrane-biotinylated Na,K-ATPase in IMCDcells. This is the same blot as used for the experiment above but it has channel opening, in the presence of PTH a depolarizingbeen stripped and reprobed with the antibody to the Na,K-ATPase. membrane potential at the basolateral membrane wouldLane 1, 20 �g rat heart homogenate shown as control; Lane 2, 1/20 of
enhance opening of the cardiac �1C subunit of the L-typeinput, 114 �g of total IMCD protein for the apical biotinylation experi-ment (density 3651). Lane 3, 1/20 of input, 121 �g of total IMCD protein calcium channel causing even greater increases in intra-for the basolateral biotinylation experiment (density 3758), Lane 4, cellular calcium. Similarly in the cortical collecting duct,apical biotinylated Na,K-ATPase from the first elution and first extrac-
calcium influx via the �1C subunit of the L-type calciumtion (density 2501). Lane 5, basolateral biotinylated Na,K-ATPase fromthe first elution and first extraction (density 12338). Lane 6, apical channel would inhibit the amiloride-sensitive sodiumbiotinylated Na,K-ATPase from the second elution and first extraction(density 573). The density of the third elution (not shown) is 15. Lane7, basolateral biotinylated Na,K-ATPase from the second elution andfirst extraction (density 8309). The total amount of apical Na,K-ATPasedensity from the three elutions is 3089 arbitrary units and comparingthis with the input density of 73,202 arbitrary units means that about 75,160 arbitrary units. The total amount of basolateral Na,K-ATPase4.1% of the total Na,K-ATPase is found in the apical membrane of from the three elutions is 26,626 arbitrary units. About 35.4% of thethese IMCD cells in this experiment. The calculated total input density total cellular Na,K-ATPase is found at the basolateral membrane ofis for the Na,K-ATPase in this basolateral biotinylation experiment is IMCD cells in this experiment.

Zhao et al: �1C subunit of the L-type calcium channel 1405
activation of �1D isoform by parathyroid hormone. Proc Natl Acadchannel, ENaC, by blocking at the intracellular face ofSci USA 92:10914–10918, 1995
this channel [43]. Likewise, the flow-dependent calcium- 9. Yu AS, Herbert SC, Brenner BM, Lytton J: Molecular character-ization and nephron distribution of a family of transcripts encodingactivated potassium channel in the apical membrane ofthe pore-forming subunit of Ca2� channels in the kidney. Procthe cortical collecting duct [44] would have increasedNatl Acad Sci USA 89:10494–10498, 1992
gating upon elevation of intracellular calcium via the �1C 10. Barry ELR, Gesek FA, Yu ASL, et al: Distinct calcium channelisoforms mediate parathyroid hormone and chlorothiazide-stimu-subunit of the L-type calcium channel situated in thelated calcium entry in transporting epithelial cells. J Membr Biolprincipal cells of this segment.161:55–64, 1998
The major distribution of the protein of the �1C subunit 11. Mandon B, Siga E, Champigneulle A, et al: Molecular analysisof beta-adrenergic receptor subtypes in rat collecting duct: Effectsof the L-type calcium channel is in the cells of the outeron cell camp and Ca2� levels. Am J Physiol 268:F1070–F1080, 1995and inner medullary collecting ducts. In these cells the
12. Montrose-Rafizadeh C, Guggino WB: Role of intracellular cal-V2 vasopressin receptor is known to mobilize calcium in cium in volume regulation by rabbit medullary thick ascending
limb cells. Am J Physiol 260:F402–F409, 1991addition to cAMP [13]. Star et al found that vasopressin13. Ecelbarger CA, Chou CL, Lolait SJ, et al: Evidence for dualincreases calcium in the terminal part of the IMCD, but
signaling pathways for V2 vasopressin receptor in rat inner medul-that the change in cAMP—not calcium—is responsible lary collecting duct. Am J Physiol 270:F623–F633, 1996
14. Hoenderop JGJ, van der Kemp AWCM, Hartog A, et al: Molecu-for the changes in urea permeability [45]. In addition,lar identification of the apical Ca2� channel in 1,25-dihydroxyvita-Mooren et al found that calcium influx was increased by min D3-reponsive epithelia. J Biol Chem 274:8375–8378, 1999
hypoosomolality in IMCD cells [46]. On the other hand, 15. Peng JB, Chen XZ, Berger UV, et al: A rat kidney-specific cal-cium transporter in the distal nephron. J Biol Chem 275:28186–the calcium influx caused by membrane associated L-type28194, 2000calcium channels may not cause global increases in intra- 16. Andreasen D, Jenssen BL, Hansen PB, et al: The �1G-subunit
cellular calcium, but these increases in calcium may be of a voltage-dependent Ca2� channel is localized in rat distal neph-ron and collecting duct. Am J Physiol (Renal Physiol) 279:F997–localized to near the membrane where the activity of theF1005, 2000calcium-activated potassium channel [47] or other cal- 17. Qiu W, Lee B, Lancaster M, et al: Cyclic nucleotide-gated cation
cium-mediated membrane events could be stimulated. channels mediate sodium and calcium influx in rat colon. Am JPhysiol (Cell Physiol) 278:C336–C343, 2000Given the technical limits of immunolocalization in small
18. Wang X-T, Nagaba S, Nagaba Y, et al: Cardiac L-type calciumcells and the inefficiency of biotinylation, we conclude channel �1-subunit is increased by cyclic adenosine monophos-that IMCD cells have both a membrane and intracellular phate: Messenger RNA and protein expression in intact bone.
J Bone Miner Res 15:1275–1285, 2000localization of �1C, suggesting that the L-type calcium19. Ludwig A, Flockerzi V, Hofmann F: Regional expression andchannel may play an additional intracellular function as cellular localization of the �1 and � subunit of high voltage-acti-
well as apical and basolateral membrane calcium signaling. vated calcium channels in rat brain. J Neurosci 17:1339–1349, 199720. Wang X-T, Nagaba Y, Cross HS, et al: The mRNA of L type
calcium channel elevated in colon cancer: Protein distribution inACKNOWLEDGMENT normal and cancerous colon. Am J Pathol 157:1549–1562, 2000
21. Tisher CC, Madsen KM: Anatomy of the kidney, in Brenner &This work was funded by NIH DK 43423 to S.E.G.Rector’s The Kidney (vol 1, 5th ed), edited by Brenner BM, Phila-delphia, WB Saunders Company, 1996, pp 1–71Reprint requests to Sandra E. Guggino, Ph.D., Department of Medi-
22. Roth J, Brown D, Norman AW, Orci L: Localization of thecine, 929 Ross Research Bldg., 720 Rutland Avenue, Baltimore, Mary-vitamin D-dependent calcium-binding protein in mammalian kid-land 21205, USAney. Am J Physiol 243:F243–F252, 1982E-mail: [email protected]
23. Taylor AN, McIntosh JE, Bourdeau JE: Immunocytochemicallocalization of vitamin D dependent calcium-binding protein inrenal tubules of rabbit, rat, and chick. Kidney Int 21:765–773, 1982REFERENCES
24. Schreiner DS, Jande SS, Parkes CO, et al: Immunocytochemical1. Catterall WA: Structure and function of voltage-gated ion chan- demonstration of two vitamin D-dependent calcium binding pro-
nels. Ann Rev Biochem 64:493–531, 1995 teins in mammalian kidney. Acta Anat 117:1–14, 19832. Perez-Reyes E, Schneider T: Molecular biology of calcium chan- 25. Kumar R, Schaefer J, Grande JP, Roche PC: Immunolocalization
nels. Kidney Int 48:1111–1124, 1995 of calcitriol receptor, 24-hydroxylase cytochrome P-450, and calbin-3. Henry PD: Comparative pharmacology of calcium antagonists: Ni- din D28K in human kidney. Am J Physiol 266:F477–F485, 1994
fedipine, verapamil and diltiazem. Am J Cardiol 46:1047–1058, 1981 26. Liu L, Khastgir A, McCauley JM, et al: RT-PCR microlocaliza-4. Yamaguchi DT, Hahn TJ, Iida-Klein, et al: Parathyroid hormone- tion of mRNAs for calbindin-D28K and vitamin D receptor in the
activated calcium channels in an osteoblast-like clonal osteosar- murine nephron. Am J Physiol 270:F677–F681, 1996coma cell line. CAMP-dependent and cAMP-independent calcium 27. Reilly RF, Ellison DH: Mammalian distal tubule: Physiol-channels. J Biol Chem 262:7711–7718, 1987 ogy, pathophysiology and molecular anatomy. Physiol Rev 80:277–
5. Lieberherr M: Effects of vitamin D3 metabolites on cytosolic free 313, 2000calcium in confluent mouse osteoblasts. J Biol Chem 262:13168– 28. Dorup J: Ultrastructure of three-dimensionally localized distal13173, 1987 nephron segments in superficial cortex of the rat kidney. J Ultrast
6. Guggino SE, Lajeunesse D, Wagner J, Snyder SH: Bone remod- Mol Struct Res 99:169–187, 1988eling signaled by a dihydropyridine and phenylalkylamine-sensitive 29. Fejes-Toth G, Naray-Fejes-Toth A: Isolated principal and inter-calcium channel. Proc Natl Acad Sci USA 86:2957–2960, 1989 calated cells: Hormone responsiveness and Na�-K�-ATPase activ-
7. Yukihiro S, Posner GH, Guggino SE: Vitamin D3 analogs stimu- ity. Am J Physiol (Renal Physiol) 25:F742–F750, 1989late calcium currents in rat osteosarcoma cells. J Biol Chem 269: 30. Hoenderop JG, Hartog A, Stuiver M, et al: Localization of the23889–23893, 1994 epithelial Ca(2�) channel in rabbit kidney and intestine. J Am
8. Barry ELR, Gesek FA, Froehner SC, Friedman PA: Multiple Soc Nephrol 11:1171–1178, 200031. Morel F, Chebardes D, Imbert M: Functional segmentation of thecalcium channel transcripts in rat osteosarcoma cells: Selective

Zhao et al: �1C subunit of the L-type calcium channel1406
rabbit distal tube by microdetermination of hormone-dependent 40. McCarty NA, O’Neil RG: Calcium-dependent control of volumeregulation in renal proximal tubule cells: II. Roles of dihydropyri-adenylate cyclase activity. Kidney Int 9:264–277, 1976dine-sensitive and -insensitive Ca2� entry pathways. J Membr Biol32. Gottardi CJ, Dunbar LA, Caplan MJ: Biotinylation and assess-123:161–170, 1991ment of membrane polarity: caveats and methodological concerns.
41. Hall DA, Carmines PK, Sansom SC: Dihydropyridine-sensitiveAm J Physiol (Renal Physiol) 37:F285–F295, 1995calcium channels in human glomerulosa mesangial cells. Am J33. Sequier JM, Hunziker W, Richards G: Localization of calbindinPhysiol (Renal Physiol) 278:F97–F103, 2000D28 mRNA in tissues by in situ hybridization. Neurosc Lett 86:155–
42. Hoenderop JG, van der Kemp AW, Hartog A, et al: The epithe-160, 1988lial calcium channel, ECaC, is activated by hyperpolarization and34. Takada T, Yamamoto A, Omori K, Tashior Y: Quantitative immu-regulated by cytosolic calcium. Biochem Biophys Res Communnogold localization of Na,K-ATPase along rat nephron. Histo-261:488–492, 1999chemistry 98:183–197, 1992
43. Ishikawa T, Muranaka Y, Rotin D: Electrophysiological char-35. Gottardi CJ, Caplan MJ: Cell surface biotinylation in the determi-acterization of the rat epithelial Na channel (rENaC) expressednation of epithelial membrane polarity. J Tiss Cult Meth 14:173–in MDCK cells. Effects of Na� and Ca2�. J Gen Physiol 111:825–180, 1992846, 199836. MacPherson MR, Pollock VP, Broderick KE, et al: Model organ-
44. Woda CB, Bragin A, Kleyman TR, Satlin LM: Flow-depen-isms: New insights into ion channel and transporter function. L-type dent K� secretion in the cortical collecting duct is mediated by acalcium channels regulate epithelial fluid transport in Drosophila maxi-K channel. Am J Physiol (Renal Physiol) 49:F786–F793, 2001melanogaster. Am J Physiol (Cell Physiol) 280:C394–C407, 2001 45. Star RA, Nonoguchi H, Balaban R, Knepper MA: Calcium and
37. Zhang MI, O’Neil RG: Molecular characterization of rabbit renal cyclic adenosine monophosphate as second messengers for vaso-epithelial calcium channel. Biochem Biophys Res Comm 280:435– pressin in the rat inner medullary collecting duct. J Clin Invest439, 2001 81:1879–1888, 1988
38. O’Neil RG, Reid JM, Williams RL, Karin NJ: Expression of 46. Mooren FC, Kinne RK: Intracellular calcium in primary culturesdihydropyridine binding sites in renal epithelial cells. Biochem Bio- of rat renal inner medullary collecting duct cells during variationsphys Res Commun 237:41–45, 1997 of extracellular osmolality. Pflugers Arch 427:463–472, 1994
39. McCarty NA, O’Neil RG: Calcium-dependent control of volume 47. Taniguchi J, Imai M: Flow dependent activation of Maxi-K� chan-regulation in renal proximal tubule cells: I. Swelling-activated Ca2� nels in apical membrane of rabbit connecting tubule. J Membr
Biol 164:35–45, 1998entry and release. J Membr Biol 123:149–160, 1991



















