
TTC7B emerges as a novel risk factor for ischemic stroke through the convergence of several...
12
TTC7B emerges as a novel risk factor for ischemic stroke through the convergence of several genome-wide approaches Tiago Krug 1,2 , Joa ˜o Paulo Gabriel 3 , Ricardo Taipa 4 , Benedita V Fonseca 1,2 , Sophie Domingues-Montanari 5 , Israel Fernandez-Cadenas 5 , Helena Manso 2,6 , Liliana O Gouveia 7 , Joa ˜o Sobral 2,6 , Isabel Albergaria 6 , Gisela Gaspar 6 , Jordi Jime ´nez-Conde 8 , Raquel Rabionet 9 , Jose ´ M Ferro 1,7 , Joan Montaner 5 , Astrid M Vicente 2,6 , Ma ´rio Rui Silva 3 , Ilda Matos 10 , Gabriela Lopes 4 and Sofia A Oliveira 1,2 1 Instituto de Medicina Molecular, Faculdade de Medicina da Universidade de Lisboa, Lisboa, Portugal; 2 Instituto Gulbenkian de Cieˆncia, Oeiras, Portugal; 3 Servic¸o de Neurologia, Hospital de Sa ˜o Pedro, Vila Real, Portugal; 4 Servic¸o de Neurologia, Hospital Geral de Santo Anto ´nio, Porto, Portugal; 5 Neurovascular Research Laboratory and Neurovascular Unit, Neurology and Medicine Departments, Universitat Auto ´noma de Barcelona, Vall d’Hebron Hospital, Barcelona, Spain; 6 Instituto Nacional de Sau ´de Dr Ricardo Jorge, Lisboa, Portugal; 7 Servic¸o de Neurologia, Hospital de Santa Maria, Lisboa, Portugal; 8 NeuroMar Research Group, Stroke Unit, Department of Neurology, Hospital del Mar-IMIM, Barcelona, Spain; 9 Genes and Disease Program, Center for Genomic Regulation, Barcelona, Spain; 10 Servic¸o de Neurologia, Hospital Distrital de Mirandela, Mirandela, Portugal We hereby propose a novel approach to the identification of ischemic stroke (IS) susceptibility genes that involves converging data from several unbiased genetic and genomic tools. We tested the association between IS and genes differentially expressed between cases and controls, then determined which data mapped to previously reported linkage peaks and were nominally associated with stroke in published genome-wide association studies. We first performed gene expression profiling in peripheral blood mononuclear cells of 20 IS cases and 20 controls. Sixteen differentially expressed genes mapped to reported whole-genome linkage peaks, including the TTC7B gene, which has been associated with major cardiovascular disease. At the TTC7B locus, 46 tagging polymorphisms were tested for association in 565 Portuguese IS cases and 520 controls. Markers nominally associated in at least one test and defining associated haplotypes were then examined in 570 IS Spanish cases and 390 controls. Several polymorphisms and haplotypes in the intron 5–intron 6 region of TTC7B were also associated with IS risk in the Spanish and combined data sets. Multiple independent lines of evidence therefore support the role of TTC7B in stroke susceptibility, but further work is warranted to identify the exact risk variant and its pathogenic potential. Journal of Cerebral Blood Flow & Metabolism (2012) 32, 1061–1072; doi:10.1038/jcbfm.2012.24; published online 28 March 2012 Keywords: cerebrovascular disease; gene expression profiling; genomic convergence; PBMCs; susceptibility genes Introduction Stroke is the third leading cause of death in the developed world and is even more disabling than lethal; survival results in persistent neurological impairments and physical disabilities with a high socio-economic cost. Stroke is a complex disease, resulting from the interplay of numerous environ- mental and genetic risk factors. Current knowledge regarding the genetics of stroke is limited and identi- fication of the susceptibility genes represents the clearest path to a better understanding of its etio- pathogenic mechanisms. Received 19 September 2011; revised 22 December 2011; accepted 30 December 2012; published online 28 March 2012 Correspondence: Dr SA Oliveira, Instituto de Medicina Molecular, 1649-028 Lisboa, Portugal. E-mails: [email protected] and [email protected] This work was supported by the PTDC/SAU-GMG/64426/2006 and POCI 2010/010.6/A019/2005 Grants and a Cie ˆncia 2008 contract (SAO) from the Portuguese Fundac ¸a ˜o para a Cie ˆncia e a Tecnologia (FCT), and by the Marie Curie International Reintegration Grant 513760. TK and FM were supported by fellowships from FCT (SFRH/BD/21609/2005 and SFRH/BD/31147/2006, respectively), and LOG received support from the Fundac ¸a ˜o AstraZeneca/ Faculdade de Medicina de Lisboa. Journal of Cerebral Blood Flow & Metabolism (2012) 32, 1061–1072 & 2012 ISCBFM All rights reserved 0271-678X/12 $32.00 www.jcbfm.com
-
Upload
gulbenkian -
Category
Documents
-
view
4 -
download
0
Transcript of TTC7B emerges as a novel risk factor for ischemic stroke through the convergence of several...

TTC7B emerges as a novel risk factor for ischemicstroke through the convergence of severalgenome-wide approaches
Tiago Krug1,2, Joao Paulo Gabriel3, Ricardo Taipa4, Benedita V Fonseca1,2,Sophie Domingues-Montanari5, Israel Fernandez-Cadenas5, Helena Manso2,6,Liliana O Gouveia7, Joao Sobral2,6, Isabel Albergaria6, Gisela Gaspar6, Jordi Jimenez-Conde8,Raquel Rabionet9, Jose M Ferro1,7, Joan Montaner5, Astrid M Vicente2,6, Mario Rui Silva3,Ilda Matos10, Gabriela Lopes4 and Sofia A Oliveira1,2
1Instituto de Medicina Molecular, Faculdade de Medicina da Universidade de Lisboa, Lisboa, Portugal;2Instituto Gulbenkian de Ciencia, Oeiras, Portugal; 3Servico de Neurologia, Hospital de Sao Pedro, Vila Real,Portugal; 4Servico de Neurologia, Hospital Geral de Santo Antonio, Porto, Portugal; 5Neurovascular ResearchLaboratory and Neurovascular Unit, Neurology and Medicine Departments, Universitat Autonoma deBarcelona, Vall d’Hebron Hospital, Barcelona, Spain; 6Instituto Nacional de Saude Dr Ricardo Jorge, Lisboa,Portugal; 7Servico de Neurologia, Hospital de Santa Maria, Lisboa, Portugal; 8NeuroMar Research Group,Stroke Unit, Department of Neurology, Hospital del Mar-IMIM, Barcelona, Spain; 9Genes and DiseaseProgram, Center for Genomic Regulation, Barcelona, Spain; 10Servico de Neurologia, Hospital Distrital deMirandela, Mirandela, Portugal
We hereby propose a novel approach to the identification of ischemic stroke (IS) susceptibilitygenes that involves converging data from several unbiased genetic and genomic tools. We testedthe association between IS and genes differentially expressed between cases and controls, thendetermined which data mapped to previously reported linkage peaks and were nominally associatedwith stroke in published genome-wide association studies. We first performed gene expressionprofiling in peripheral blood mononuclear cells of 20 IS cases and 20 controls. Sixteen differentiallyexpressed genes mapped to reported whole-genome linkage peaks, including the TTC7B gene,which has been associated with major cardiovascular disease. At the TTC7B locus, 46 taggingpolymorphisms were tested for association in 565 Portuguese IS cases and 520 controls. Markersnominally associated in at least one test and defining associated haplotypes were then examined in570 IS Spanish cases and 390 controls. Several polymorphisms and haplotypes in the intron5–intron 6 region of TTC7B were also associated with IS risk in the Spanish and combined data sets.Multiple independent lines of evidence therefore support the role of TTC7B in stroke susceptibility,but further work is warranted to identify the exact risk variant and its pathogenic potential.Journal of Cerebral Blood Flow & Metabolism (2012) 32, 1061–1072; doi:10.1038/jcbfm.2012.24; published online 28 March 2012
Keywords: cerebrovascular disease; gene expression profiling; genomic convergence; PBMCs; susceptibility genes
Introduction
Stroke is the third leading cause of death in thedeveloped world and is even more disabling thanlethal; survival results in persistent neurologicalimpairments and physical disabilities with a highsocio-economic cost. Stroke is a complex disease,resulting from the interplay of numerous environ-mental and genetic risk factors. Current knowledgeregarding the genetics of stroke is limited and identi-fication of the susceptibility genes represents theclearest path to a better understanding of its etio-pathogenic mechanisms.
Received 19 September 2011; revised 22 December 2011; accepted30 December 2012; published online 28 March 2012
Correspondence: Dr SA Oliveira, Instituto de Medicina Molecular,1649-028 Lisboa, Portugal.E-mails: [email protected] and [email protected]
This work was supported by the PTDC/SAU-GMG/64426/2006 and
POCI 2010/010.6/A019/2005 Grants and a Ciencia 2008 contract
(SAO) from the Portuguese Fundacao para a Ciencia e a Tecnologia
(FCT), and by the Marie Curie International Reintegration Grant
513760. TK and FM were supported by fellowships from FCT
(SFRH/BD/21609/2005 and SFRH/BD/31147/2006, respectively),
and LOG received support from the Fundacao AstraZeneca/
Faculdade de Medicina de Lisboa.
Journal of Cerebral Blood Flow & Metabolism (2012) 32, 1061–1072& 2012 ISCBFM All rights reserved 0271-678X/12 $32.00
www.jcbfm.com

In the present study, we propose a novel multi-factorial approach that combines genomic profilingwith linkage and association studies to dissect thegenetic underpinnings of stroke. The usefulnessof microarray expression analysis is often greatlyhampered by the overwhelming amount of informationgenerated combined with the fact that genes with thegreatest levels of differential expression or significanceare not necessarily the most important to furtherinvestigate. The use of intersecting data derivedfrom other powerful and unbiased resources (e.g.,linkage screens and genome-wide association studies(GWAS)), represents the first step toward a moreefficient method of identifying susceptibility genes.
To our knowledge, there are no published studiesinvestigating gene expression changes in humansthat specifically increase the risk for a stroke event.The reported profiling studies in humans wereperformed either during the acute phase or in thefirst months after the stroke event (Moore et al,2005a, b; Tang et al, 2006; Baird, 2007; Xu et al, 2008;Sharp et al, 2011), and thus these studies address thestroke severity and/or recovery mechanisms morethan the risk of the stroke event.
To investigate the genetic architecture of familialstroke, three whole-genome linkage screens havepreviously been performed. The initial linkage peakon chromosome 5q12 was identified in Icelandicfamilies (Gretarsdottir et al, 2002) and was replicatedin northern Sweden (Nilsson-Ardnor et al, 2005),with additional linkage evidence for loci on 1p34,5q13, 7q35, 9q22, 9q34, 13q32, 14q32, 18p11, and20q13 (Nilsson-Ardnor et al, 2007). Follow-up of twoIcelandic studies on stroke (Gretarsdottir et al, 2003;Helgadottir et al, 2004) has sparked intense and on-going investigation and debate regarding whetherPDE4D and ALOX5P are true stroke susceptibilitygenes (Domingues-Montanari et al, 2010a).
The recent advent of GWAS has enabled genome-wide investigations of stroke, mostly in case–controldata sets. The first IS GWAS tested the association ofover 400,000 single-nucleotide polymorphisms(SNPs) in 249 Caucasian IS cases and 268 controls(Matarın et al, 2007); no SNPs reached genome-widesignificance, but the ones with the strongest associa-tions merit further investigation. A second GWASwas performed in 188 Japanese IS cases and 188controls using 52,608 gene-based tagging SNPs. Thisstudy was followed by validation in large Japanesesamples and sequencing; an association was identi-fied between lacunar infarction and SNP 1425G/A inthe protein kinase C Z (PRKCH) gene on chromosome14q22–q23 (Kubo et al, 2007). However, this SNP hasa very small minor allele frequency in Europeans andAfricans, suggesting that this association is likely tobe Asian-specific (Cheng et al, 2009). In 2008, anIS GWAS performed in Icelandic samples andvalidated in several large European cohorts, foundtwo neighboring SNPs (rs2200733 and rs10033464) onchromosome 4q25 to be strongly associated withcardioembolic stroke (Gretarsdottir et al, 2008). Single-
nucleotide polymorphism rs2200733, which waspreviously strongly implicated in atrial fibrillationsusceptibility, was also associated with all IS typescombined. Finally, the latest stroke GWAS was ajoint analysis of four large American and Europeandata sets (Ikram et al, 2009); it yielded two intergenicSNPs within 11 kb of NINJ2 (ninjurin 2) that werehighly associated with IS and atherothromboticstroke, as well as with all strokes combined. Thesefindings were not replicated in a study of 8,637 IScases and 8,733 controls of European ancestry (ISGCand WTCCC2, 2010).
The only family-based genome-wide scan for ISwas conducted on 1,345 Framingham Heart Studyparticipants from 310 pedigrees (Larson et al, 2007).Four major cardiovascular disease (CVD) outcomes(e.g., major atherosclerotic CVD, which includesmyocardial infarction, coronary heart disease, death,and stroke) were analyzed and several associationsreached P < 1� 10�5.
Given the discrepancies in the findings amongGWAS, it is clearly necessary to validate their mostsignificant results (above and below genome-widesignificance level) in independent data sets, as wellas by other approaches, such as the one proposed here(combination of genetics and genomic profiling), topinpoint the real genetic players in stroke etiology.
Materials and methods
Study Subjects
The Portuguese and Spanish stroke cases and controls usedin this study were ascertained and collected as describedpreviously (Krug et al (2010) for Portuguese samples,Montaner et al (2006) for Spanish cases, and Domingues-Montanari et al (2010c) for Spanish controls). All participantswere adults and Caucasian. Spanish patients were classifiedinto causative subtypes according to the Trial of Org 10172 inAcute Stroke Treatment classification (Adams et al, 1993).
More stringent inclusion and exclusion criteria wereapplied to individuals participating in the genomic expres-sion profiling study; IS patients were required to have sufferedonly one stroke episode, at least 6 months before the bloodcollection, and controls could not have a family history ofstroke. Participants with severe anemia or active allergieswere also excluded.
The study was approved by the ethics committees of theparticipating institutions. All participants were informedof the study and provided informed consent.
Gene Profiling Studies
Whole blood samples were obtained by venipuncture andcollected in BD Vacutainer CPT tubes (BD, Franklin Lakes,NJ, USA). These samples were centrifuged to isolateperipheral blood mononuclear cells (PBMCs), which werethen washed twice and their RNA was stabilized usingRNAlater (Qiagen, Hilden, Germany) within 3 hours aftersample collection. Total RNA was extracted using the
TTC7B and ischemic stroke riskT Krug et al
1062
Journal of Cerebral Blood Flow & Metabolism (2012) 32, 1061–1072

RNeasy Mini kit (Qiagen). High-quality total RNA, 3.5mgfrom each individual, was hybridized to a GeneChipHuman Genome U133 Plus 2.0 microarray (Affymetrix,Santa Clara, CA, USA) at the Instituto Gulbenkian deCiencia’s Affymetrix Core Facility following the manufac-turer’s protocol. Extensive quality control checks wereperformed in all steps of the process.
The generated intensity array data were analyzed togetherwith their respective CDF file from Affymetrix on thePartek software (Partek Incorporated, St Louis, MO, USA). Theimported CEL files were subjected to background correc-tion, normalization, and summarization using the robustmultichip average algorithm. Analysis of variance was usedto identify the differentially expressed genes among casesand controls, taking into account known experimental(type, sex, and age) and study design (geographic originand scan date) covariates (P value). The genes with a > 1.2-fold change and a Q value (Storey, 2002) p0.05 wereconsidered differentially expressed. All genes with P valuep0.05 also had a Q value p0.05. The false discovery ratewas determined based on Q values, as these have a higherapparent power when compared with other standardmethods (Qian and Huang, 2005).
Gene expression profiling was conducted and reportedin accordance with the minimum information about amicroarray experiment (MIAME) criteria (Brazma et al,2001). The Gene Expression Omnibus (GEO) accessionnumber for the data is GSE22255.
With the Partek software, principal component analysiswas performed to visualize the relative position of eachindividual in a low-dimensional space, and hierarchicalclustering analyses were used to determine the expressionpatterns across the samples. As these visualization toolscannot correct for study design batch effects, we removedthe effects of the geographic origin of the participants andthe scan date of the microarrays using the batch-removetool implemented in the Partek software before visualiza-tion. Principal component analysis was performed with thecorrelation dispersion matrix and normalized eigenvectorscaling, and hierarchical clustering was performed with thecorrelation distance metric and centroid linkage method.
Gene ontology was executed using the Gene FunctionEnrichment tool from the dChip 2009 software (http://biosun1.harvard.edu/complab/dchip/), with a gene P thresholdof 0.01 and the HG-U133_Plus_2.na28.annot.csv annotationfile jointly with the most recent gene ontology structurefiles.
Quantitative real-time polymerase chain reaction con-firmation of some of the microarray results was notperformed because of an insufficient amount of RNA stillavailable for several of the controls and patients used inthis study.
Genotyping
Tagging SNPs in TTC7B and 10 kb flanking regions wereidentified in Haploview v4.0 (Mark Daly’s Lab at the BroadInstitute, Cambridge, MA, USA; Barrett et al, 2005) basedon genotypes of 30 European (CEU, CEPH Utah residentswith ancestry from northern and western Europe) familytrios (HapMap Release 21/phase II Jul06) and using the
following options: pairwise mode, r2 > 0.8, and minor allelefrequency > 0.1. A total of 61 SNPs were genotyped inthe Portuguese data set using Sequenom’s iPlex assays(Sequenom, San Diego, CA, USA) following the manufac-turer’s instructions. The primer sequences were designedusing Sequenom’s MassArray Assay Design 3.0 software(Sequenom) and are indicated in Supplementary Table 1.All genotype determinations were performed and blindedto affection status, and an extensive quality control wasperformed (e.g., eight HapMap controls of diverse ethnicaffiliation, and sample duplication within and acrossplates). Single-nucleotide polymorphisms with a < 90%call rate and SNPs out of Hardy–Weinberg equilibrium inthe control group (P < 0.05) were excluded.
For validation purposes, 10 SNPs in TTC7B (rs2343,rs12147413, rs11629065, rs942738, rs12893100, rs1742100,rs1742098, rs1535321, rs13379124, and rs7154098) weregenotyped in the Spanish sample at the Spanish NationalGenotyping Center (CeGen) using Sequenom’s iPlex assays.
Association Analyses
For all genotyped SNPs, deviations from Hardy–Weinbergequilibrium (P < 0.05) were assessed at each marker in thecase and control samples separately, using the SNPassocv.1.4-9 package (Gonzalez et al, 2007) implemented in theR freeware (http://cran.r-project.org/). Linkage disequili-brium (LD) plots were constructed with Haploview v4.0.All possible pairwise correlation coefficients (r2) in eachgene were calculated, and LD plots were constructed withthe r2 color scheme (r2 = 0: white; 0 < r2 < 1: shades of gray;r2 = 1: black).
The associations between IS risk and specific classes ofalleles/genotypes/haplotypes were tested using a standardw2 test. Haplotypes were estimated in Haploview v4.0 usingthe confidence intervals algorithm. Multivariate logisticregression with backward elimination of risk factors wasperformed using SNPassoc to adjust the association analysesfor confounding covariates. Hypertension, diabetes, andever smoking were the covariates in the Portuguese sample;hypertension, diabetes, ever smoking, and dyslipidemicstatus were the covariates in the Spanish data set. In thejoint analysis with both data sets, the covariates werehypertension, diabetes, ever smoking, and sample origin.Only a weak interaction (i) was observed among covariatesin regression models (�0.2 < i < 0.2). Results were consid-ered statistically significant below the conventional levelof 0.05. Odds ratios (ORs) and their associated 95%confidence intervals (CIs) were calculated to assess therelative disease risk conferred by a particular associatedallele/genotype. Since some of the markers were in LD andthe haplotype comparisons were not independent, we didnot perform corrections for multiple testing; uncorrectedP values are reported.
Results
Gene Expression Profiling
To investigate the gene expression differencesbetween IS cases and controls in the nonacute phase
TTC7B and ischemic stroke riskT Krug et al
1063
Journal of Cerebral Blood Flow & Metabolism (2012) 32, 1061–1072

of stroke, we compared the genetic profiles of PBMCsfrom 20 IS cases (from whom the samples werecollected at least 6 months after the first and onlystroke event) and 20 controls. The principal demo-graphic, clinical, and lifestyle characteristics of theexpression profiling study participants are shown inTable 1. The control and case groups were age- andsex-matched, and only the frequency of diabetes wassignificantly higher in IS patients than in controls(P = 0.035). Four patients had diabetes controlled bymedication and no controls had diabetes.
Total RNA from each individual was hybridized toan Affymetrix GeneChip Human Genome U133 Plus2.0 microarray and all of the hybridized arrays metquality scores. The average±s.d. of present calls andof background for all the arrays were 44.8%±1.6%and 45.3%±5.7%, respectively.
Using analysis of variance on the normalizedexpression data, 709 probe sets (representing 580genes) were found to be differentially expressedamong IS cases and controls, with a threshold of 1.2-fold change and a Q value p0.05 (SupplementaryTable 2). Downregulation was observed in 331 ofthese probe sets, representing 287 genes. The geneswith the greatest and smallest changes are indicatedin Supplementary Table 3.
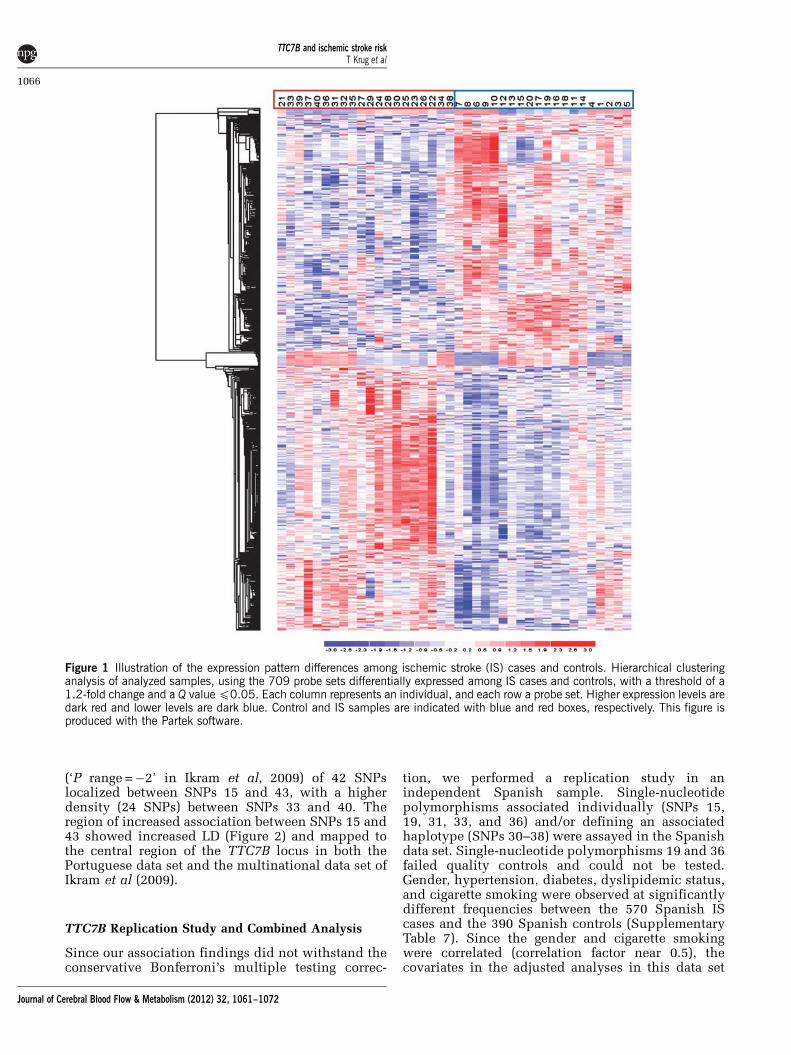
The 3D principal component analysis plot ob-tained using probe sets with P < 0.01 shows a clearseparation between cases and controls along the firstprincipal component (Supplementary Figure). Thehierarchical clustering diagram using the 709 probesets discriminates between the patients and unaf-fected individuals (Figure 1).
When analyzing the 580 differentially expressedgenes for their function (Supplementary Table 4), wefound a significant overrepresentation (1.0� 10�9 <P < 8.8� 10�4) of genes related to antigen binding (13genes), immune and inflammatory responses (38 and16 genes, respectively), platelet a granule membrane(five genes), response to virus (11 genes), oxido-reductase activity (eight genes), and response to DNAdamage stimulus (17 genes). These findings suggestthat the PBMCs (from which the total RNA wasextracted) of the IS patients have an active role in thecomplex immune and homeostatic responses to thevascular injuries that cause the ischemic attacks.
Convergence of Expression with Linkage and Genome-Wide Association Studies
To prioritize genes to be tested for association withstroke susceptibility, we intersected our expressionresults with those from published whole-genomelinkage screens and GWAS for stroke. We found that16 differentially expressed genes (Table 2) repre-sented by specific probe sets (‘_at’ suffix) mapped topreviously reported linkage peaks on chromosomes1p34, 5q12, 9q22, 9q34, 13q32, 14q32, and 20q13(Gretarsdottir et al, 2002; Nilsson-Ardnor et al, 2007).One of these 16 prioritized genes was TTC7B
(ENSG00000165914), which was also a top hit formajor CVD in the GWAS for CVDs of the FraminghamHeart Study 100K project (Larson et al, 2007). In thatproject, the major CVD phenotype included myocar-dial infarction, coronary insufficiency, coronary heartdisease death, and atherothrombotic stroke diseases.The emergence of this gene from our multifactorialapproach emphasizes its likely role in IS andsupports our expression findings.
TTC7B Association Study
To investigate the role of TTC7B in IS, we performeda two-phase association study. In the first stage, 46tagging SNPs in the gene or in its 10 kb flankingregions (Supplementary Tables 1 and 5) were testedfor association with IS in a Portuguese case–controldata set. In the second stage, SNPs that were asso-ciated at a low-stringency significance (Pp0.05) andthose that defined associated haplotypes in the firstphase were tested for association in a Spanishsample.
The principal demographic and clinical character-istics of the Portuguese study participants (565unrelated IS patients and 520 unrelated healthyindividuals) are shown in Supplementary Table 6.As expected, male-to-female ratio, and frequenciesof hypertension, diabetes, ever smoking, and everdrinking were significantly higher in IS patients thanin controls. The age-at-examination was deliberatelysignificantly higher in controls relative to patients tominimize misclassification biases. Since the sex,ever smoking, and ever drinking were correlated(correlation factors near 0.5), only the hypertension,diabetes, and ever smoking were included in theanalyses adjusted for covariates.
The top plot in Figure 2 depicts the allelic,genotypic (crude and adjusted), and top haplotypeassociation results from the Portuguese data set(represented by open circles connected by lines).The pairwise LD plot (bottom picture of Figure 2)shows little LD between most of the 46 SNPs studied.Three polymorphisms (SNPs 15, 31, and 36) demon-strated allelic associations with IS, while six SNPswere associated with IS in unadjusted (SNPs 5, 15,19, 33, and 36) and/or adjusted (SNPs 5, 7, 19, 33,and 36) genotypic tests (recessive model). Only SNP36 was associated in all tests performed, withP < 0.0095. Furthermore, the G allele of SNP 36 thatwas associated with increased stroke risk (OR (95%CI) = 1.33 (1.10–1.61)), also drives the associationbetween IS and the TACGTC haplotype defined bySNPs 33_34_35_36_37_38 (P = 0.012, top plot inFigure 2). The AGG haplotype defined by SNPs30_31_32 was also associated with IS (P = 0.015, topplot in Figure 2).
We analyzed 259 polymorphisms investigated inthe IS GWAS conducted by Ikram et al (2009) thatwere identified in the TTC7B gene region (secondplot in Figure 2). There was a modest association
TTC7B and ischemic stroke riskT Krug et al
1064
Journal of Cerebral Blood Flow & Metabolism (2012) 32, 1061–1072

Table 1 Characterization of the samples used in the gene profiling study
Affection status Sex ID AAE (years) AAO (years) Clinical/lifestyle characteristics Geographic origina Scan dateb
Control F 1 68 — Mirandela AF 2 68 — Hypertension Mirandela AF 3 73 — Hypertension Mirandela BF 4 65 — Hypertension, hypercholesterolemia Mirandela CF 5 73 — Hypertension, hypercholesterolemia Porto CF 6 45 — Mirandela AF 7 48 — Ever drinker Mirandela AF 8 47 — Hypertension Mirandela AF 9 47 — Hypertension, ever smoker Porto DF 10 48 — Hypercholesterolemia, ever drinker Lisboa FM 11 69 — Ever smoker, ever drinker Vila Real CM 12 72 — Ever drinker Vila Real CM 13 66 — Hypertension, hypercholesterolemia, ever smoker, ever drinker Porto FM 14 68 — Hypercholesterolemia, ever drinker Lisboa FM 15 69 — Lisboa FM 16 45 — Vila Real FM 17 52 — Hypertension, hypercholesterolemia, ever drinker Vila Real EM 18 53 — Ever smoker Vila Real EM 19 48 — Ever smoker Lisboa FM 20 50 — Hypercholesterolemia, ever smoker, ever drinker Lisboa F
Mean±s.d. 58.7±11.0
Case F 21 71 65 Porto FF 22 65 65 Hypertension Vila Real FF 23 73 70 Vila Real BF 24 70 69 Hypertension, hypercholesterolemia Vila Real BF 25 65 65 Hypertension, hypercholesterolemia Porto FF 26 53 45 Hypertension, hypercholesterolemia, ever smoker, ever drinker Porto BF 27 54 49 Hypertension, diabetes, hypercholesterolemia, ever drinker Vila Real AF 28 47 43 Hypercholesterolemia Vila Real FF 29 54 50 Hypertension Vila Real AF 30 49 48 Diabetes, hypercholesterolemia, ever smoker, ever drinker Porto FM 31 69 65 Hypertension, diabetes, hypercholesterolemia, ever drinker Mirandela BM 32 74 72 Hypertension, ever drinker Mirandela CM 33 74 73 Hypertension, hypercholesterolemia, ever drinker Mirandela CM 34 68 66 Ever drinker Mirandela CM 35 71 65 Hypertension, ever smoker, ever drinker Porto CM 36 54 53 Diabetes, hypercholesterolemia, ever drinker Mirandela CM 37 52 48 Hypercholesterolemia, ever smoker, ever drinker Mirandela CM 38 46 45 Hypertension, ever drinker Mirandela BM 39 49 47 Ever smoker, ever drinker Mirandela BM 40 46 45 Hypertension, ever drinker Vila Real A
Mean±s.d. 60.2±10.6 57.4±10.8
The principal demographic, clinical, and lifestyle characteristics of the study subjects are shown, including the geographic origin of the participants, the microarray hybridization group of the samples, and the meanvalues and s.d. of the age-at-examination (AAE) and age-at-onset (AAO), where applicable.aName of the city/town where the samples were collected.bThe microarrays of these samples were performed and scanned in six groups, coded here as A through F.
TTC7Band
ischemic
strokerisk
TKrug
etal
1065
JournalofCerebralBloodFlow
&M
etabolism(2012)
32,1061–1072
(‘P range =�2’ in Ikram et al, 2009) of 42 SNPslocalized between SNPs 15 and 43, with a higherdensity (24 SNPs) between SNPs 33 and 40. Theregion of increased association between SNPs 15 and43 showed increased LD (Figure 2) and mapped tothe central region of the TTC7B locus in both thePortuguese data set and the multinational data set ofIkram et al (2009).
TTC7B Replication Study and Combined Analysis
Since our association findings did not withstand theconservative Bonferroni’s multiple testing correc-
tion, we performed a replication study in anindependent Spanish sample. Single-nucleotidepolymorphisms associated individually (SNPs 15,19, 31, 33, and 36) and/or defining an associatedhaplotype (SNPs 30–38) were assayed in the Spanishdata set. Single-nucleotide polymorphisms 19 and 36failed quality controls and could not be tested.Gender, hypertension, diabetes, dyslipidemic status,and cigarette smoking were observed at significantlydifferent frequencies between the 570 Spanish IScases and the 390 Spanish controls (SupplementaryTable 7). Since the gender and cigarette smokingwere correlated (correlation factor near 0.5), thecovariates in the adjusted analyses in this data set
Figure 1 Illustration of the expression pattern differences among ischemic stroke (IS) cases and controls. Hierarchical clusteringanalysis of analyzed samples, using the 709 probe sets differentially expressed among IS cases and controls, with a threshold of a1.2-fold change and a Q value p0.05. Each column represents an individual, and each row a probe set. Higher expression levels aredark red and lower levels are dark blue. Control and IS samples are indicated with blue and red boxes, respectively. This figure isproduced with the Partek software.
TTC7B and ischemic stroke riskT Krug et al
1066
Journal of Cerebral Blood Flow & Metabolism (2012) 32, 1061–1072
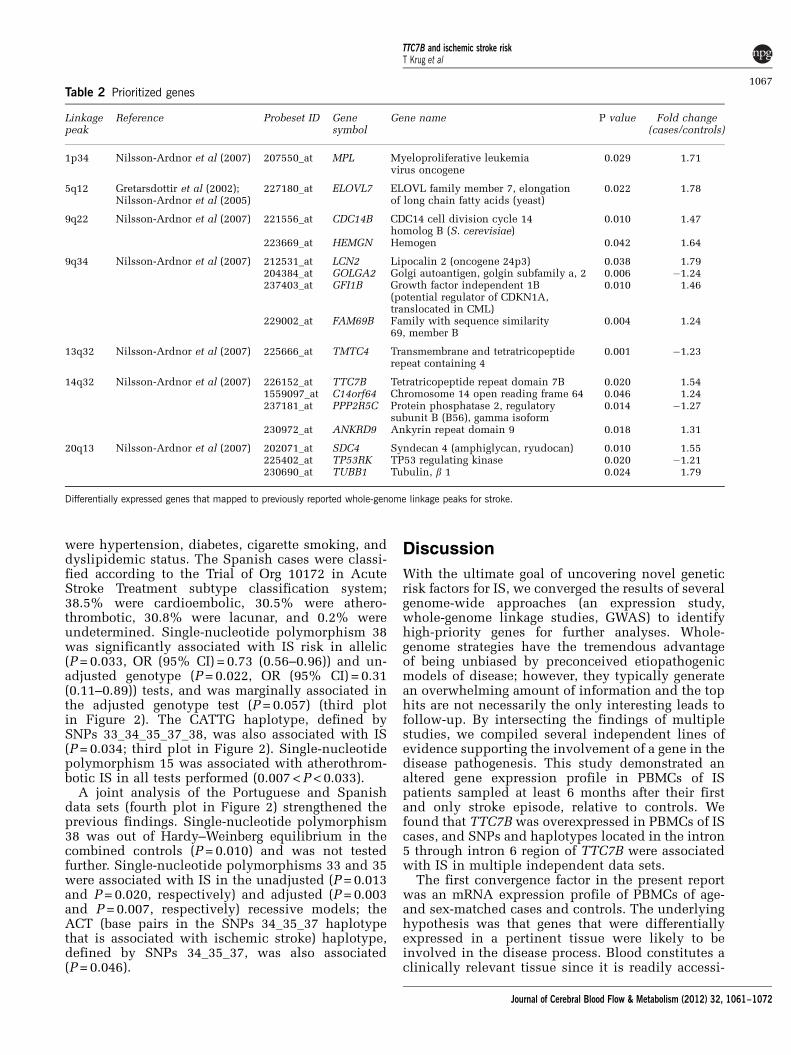
were hypertension, diabetes, cigarette smoking, anddyslipidemic status. The Spanish cases were classi-fied according to the Trial of Org 10172 in AcuteStroke Treatment subtype classification system;38.5% were cardioembolic, 30.5% were athero-thrombotic, 30.8% were lacunar, and 0.2% wereundetermined. Single-nucleotide polymorphism 38was significantly associated with IS risk in allelic(P = 0.033, OR (95% CI) = 0.73 (0.56–0.96)) and un-adjusted genotype (P = 0.022, OR (95% CI) = 0.31(0.11–0.89)) tests, and was marginally associated inthe adjusted genotype test (P = 0.057) (third plotin Figure 2). The CATTG haplotype, defined bySNPs 33_34_35_37_38, was also associated with IS(P = 0.034; third plot in Figure 2). Single-nucleotidepolymorphism 15 was associated with atherothrom-botic IS in all tests performed (0.007 < P < 0.033).
A joint analysis of the Portuguese and Spanishdata sets (fourth plot in Figure 2) strengthened theprevious findings. Single-nucleotide polymorphism38 was out of Hardy–Weinberg equilibrium in thecombined controls (P = 0.010) and was not testedfurther. Single-nucleotide polymorphisms 33 and 35were associated with IS in the unadjusted (P = 0.013and P = 0.020, respectively) and adjusted (P = 0.003and P = 0.007, respectively) recessive models; theACT (base pairs in the SNPs 34_35_37 haplotypethat is associated with ischemic stroke) haplotype,defined by SNPs 34_35_37, was also associated(P = 0.046).
Discussion
With the ultimate goal of uncovering novel geneticrisk factors for IS, we converged the results of severalgenome-wide approaches (an expression study,whole-genome linkage studies, GWAS) to identifyhigh-priority genes for further analyses. Whole-genome strategies have the tremendous advantageof being unbiased by preconceived etiopathogenicmodels of disease; however, they typically generatean overwhelming amount of information and the tophits are not necessarily the only interesting leads tofollow-up. By intersecting the findings of multiplestudies, we compiled several independent lines ofevidence supporting the involvement of a gene in thedisease pathogenesis. This study demonstrated analtered gene expression profile in PBMCs of ISpatients sampled at least 6 months after their firstand only stroke episode, relative to controls. Wefound that TTC7B was overexpressed in PBMCs of IScases, and SNPs and haplotypes located in the intron5 through intron 6 region of TTC7B were associatedwith IS in multiple independent data sets.
The first convergence factor in the present reportwas an mRNA expression profile of PBMCs of age-and sex-matched cases and controls. The underlyinghypothesis was that genes that were differentiallyexpressed in a pertinent tissue were likely to beinvolved in the disease process. Blood constitutes aclinically relevant tissue since it is readily accessi-
Table 2 Prioritized genes
Linkagepeak
Reference Probeset ID Genesymbol
Gene name P value Fold change(cases/controls)
1p34 Nilsson-Ardnor et al (2007) 207550_at MPL Myeloproliferative leukemiavirus oncogene
0.029 1.71
5q12 Gretarsdottir et al (2002);Nilsson-Ardnor et al (2005)
227180_at ELOVL7 ELOVL family member 7, elongationof long chain fatty acids (yeast)
0.022 1.78
9q22 Nilsson-Ardnor et al (2007) 221556_at CDC14B CDC14 cell division cycle 14homolog B (S. cerevisiae)
0.010 1.47
223669_at HEMGN Hemogen 0.042 1.64
9q34 Nilsson-Ardnor et al (2007) 212531_at LCN2 Lipocalin 2 (oncogene 24p3) 0.038 1.79204384_at GOLGA2 Golgi autoantigen, golgin subfamily a, 2 0.006 �1.24237403_at GFI1B Growth factor independent 1B
(potential regulator of CDKN1A,translocated in CML)
0.010 1.46
229002_at FAM69B Family with sequence similarity69, member B
0.004 1.24
13q32 Nilsson-Ardnor et al (2007) 225666_at TMTC4 Transmembrane and tetratricopeptiderepeat containing 4
0.001 �1.23
14q32 Nilsson-Ardnor et al (2007) 226152_at TTC7B Tetratricopeptide repeat domain 7B 0.020 1.541559097_at C14orf64 Chromosome 14 open reading frame 64 0.046 1.24237181_at PPP2R5C Protein phosphatase 2, regulatory
subunit B (B56), gamma isoform0.014 �1.27
230972_at ANKRD9 Ankyrin repeat domain 9 0.018 1.31
20q13 Nilsson-Ardnor et al (2007) 202071_at SDC4 Syndecan 4 (amphiglycan, ryudocan) 0.010 1.55225402_at TP53RK TP53 regulating kinase 0.020 �1.21230690_at TUBB1 Tubulin, b 1 0.024 1.79
Differentially expressed genes that mapped to previously reported whole-genome linkage peaks for stroke.
TTC7B and ischemic stroke riskT Krug et al
1067
Journal of Cerebral Blood Flow & Metabolism (2012) 32, 1061–1072

ble. Additionally, it is biologically relevant for strokesince the complex immune and homeostatic re-sponses to the vascular injury that cause the strokeevent are likely to be reflected in the expressionprofiles of circulating PBMCs (Sharp et al, 2011). It is
important to note that the some of the publishedwhole-genome expression studies conducted forstroke in humans (Moore et al, 2005a, b), in animals(Tang et al, 2001), and the current study were per-formed on PBMCs, while most of the other published
Figure 2 Association results and pairwise linkage disequilibrium (LD) among all genotyped polymorphisms for TTC7B. The topdiagram schematically represents the introns and exons of the TTC7B gene (ENST00000357056 transcript) relative to thegenotyped polymorphisms. The first, third, and fourth plots display the association results in the Portuguese, Spanish, and combineddata sets, respectively. Allelic (gray squares), crude (light gray discs), and adjusted (black triangles) genotypic (recessive model)association results are shown. The second plot depicts the association (‘P range’) of all single-nucleotide polymorphisms (SNPs) inthis region that were studied in the Ikram et al (2009) genome-wide association study (GWAS). Stars in this second plot indicatepolymorphisms that were investigated in the Portuguese sample. The LD plot at the bottom represents the LD between genotypedSNPs in the Portuguese data set, with white-to-black gradient shading proportional to the magnitude of LD using the pairwisestatistic r2.
TTC7B and ischemic stroke riskT Krug et al
1068
Journal of Cerebral Blood Flow & Metabolism (2012) 32, 1061–1072

studies have been performed on whole blood (Tanget al, 2006; Zhan et al, 2010; Barr et al, 2010) usingPAXgene tubes and at times Nugen amplificationmethods. The use of PBMCs rather than whole bloodhas the advantage of increased detection sensitivitydue to the very high levels of globin mRNA inerythrocytes, which represent B95% of all bloodcells.
Previous studies analyzed the expression profilesfrom PBMCs collected during the acute or convalescentphases of IS to understand the cascade of eventsprecipitated by a stroke and its recovery (Moore et al,2005a, b; Tang et al, 2006; Larson et al, 2007). Here, wesearched for genetic factors that predispose individualsto IS, and therefore our cases were sampled at least 6months after the first and only IS. Several studiessuggest that neurological and functional recoveryafter stroke, even in patients with severe and verysevery strokes, reaches a plateau after 6 months(Jørgensen et al, 1995; Toschke et al, 2010). Sixmonths was therefore estimated to be a sufficienttime window to allow the PBMCs to return to their‘resting’ expression profiles. One drawback of thisstrategy is that the expression of genes that renderpatients more prone to stroke may be normalizedby the use of secondary prevention drugs, such asantiplatelet agents and statins.
Hierarchical clustering and principal componentanalyses (Figure 1; Supplementary Figure, respec-tively) showed a very good separation between casesand controls, based on differentially expressed genes.After a sensitivity analysis, we selected genes forfollow-up studies based on a low threshold 1.2-foldchange, which was deemed to be appropriate for alate-onset disease such as stroke, where small changesin expression over a long period of time are expectedto result in the phenotype. Some of the differentiallyexpressed genes (e.g., SELP, F13A1, and TUBB1)were previously tested and found associated withstroke in candidate gene association studies (Zeeet al, 2004; Pruissen et al, 2008; Navarro-Nunez et al,2007). Conversely, some highly investigated strokesusceptibility genes such as PDE4D and ALOX5AP(Domingues-Montanari et al, 2010a), which wereoriginally associated with stroke after linkage studiesand fine-mapping, and genes that associated withstroke in GWAS such as NINJ2 (Ikram et al, 2009),were not differentially expressed. One limitation ofthis type of expression profiling study is that,although differentially expressed genes may consti-tute good indicators of affection status and down-stream disease mechanisms, they are not necessarilysusceptibility genes that directly account for theinitial phenotype under investigation. One strategyto address this issue is to study pathways that aresignificantly overrepresented among the list ofdifferentially expressed genes. The alternativeroute proposed here is to prioritize differentiallyexpressed genes by converging the expressionprofiling results with those from published linkagescreens and GWAS.
The gene TTC7B emerged from our study usingthis approach. It was subsequently tested in detail forassociation with IS in Iberian data sets. Polymorph-isms in this gene were found to be associated with ISin Portuguese, Spanish, and the combined samples.Population stratification in these data sets does notappear to be of major concern since the mitochon-drial haplogroup distribution was similar in Portu-guese cases and controls (Rosa et al, 2008), and thesamples from the Spanish data set originated fromone hospital in Barcelona. Furthermore, since popu-lation structure was corrected for in the TTC7Bassociations identified in the GWAS (Ikram et al,2009; Larson et al, 2007), it is unlikely that theconsistent TTC7B associations are false positivescaused by stratification.
Even though our results did not retain significancefollowing the conservative Bonferroni’s correctionfor multiple testing, our association findings werestrengthened by validation in multiple independentdata sets. The recurrent nonreplication of genome-wide significant association findings in the strokegenetics field (ISGC and WTCCC2, 2010) suggeststhat there may be false positives, and that associationresults below the genome-wide significance levelmust be evaluated cautiously if they have not beenreplicated; corroboration of results in other studiesmay lead to more reproducible conclusions. Forexample, several of the most significant findings inthe only IS GWAS that attained no genome-widesignificant results have been replicated (Domingues-Montanari et al, 2010b; Ding et al, 2010). Wetherefore consider that the modest association ofTTC7B with IS (Ikram et al, 2009) and its associationwith major CVD (P = 5.23� 10�5) (Larson et al, 2007)reinforce our findings in the Portuguese and Spanishsamples. Even though the allele frequencies of SNPsthat were associated with IS individually or as part ofa haplotype in the joint analysis of the Portugueseand Spanish data set (33, 34, 35, and 37) varyappreciably across populations (http://www.ncbi.nlm.nih.gov/projects/SNP/), these SNPs are poly-morphic in all tested HapMap populations ofEuropean, Asian, and African ancestries, suggestingthat our positive association findings may be testedin other races.
Although the association signals clustered to thecentral region of the gene, no single SNP or haplotypein TTC7B emerged as consistently associated in alldata sets; thus, the true susceptibility variant(s) inthis gene remains elusive. The observed hetero-geneity of the nominally associated SNPs in TTC7Bhighlights the need for a more detailed studyof this gene, possibly involving the investigationof other types of genetic markers (e.g., insertion/deletion and copy number variations) or lesscommon non-tagging variants using next-genera-tion sequencing.
TTC7B is a member of the TPR (tetratricopeptiderepeat) gene family. Tetratricopeptide repeats consistof tandem arrays of highly degenerate 34-amino
TTC7B and ischemic stroke riskT Krug et al
1069
Journal of Cerebral Blood Flow & Metabolism (2012) 32, 1061–1072

acid repeats that are predicted to form extendedsuperhelical arrangements. These TPR domainsfunction as protein–protein interaction modules formacromolecular complexes involved in numerouscellular processes, including transcriptional regulation,mRNA processing, protein folding, and translocation(Krachler et al, 2010). However, there are no reportsto date on the function of TTC7B.
Future work must be directed toward replicatingthese findings in other stroke samples as well as inother related vascular phenotypes. Additionally,studies should focus on elucidating the biochemicalfunctions of TTC7B and functional consequences ofits associated polymorphisms.
Acknowledgements
The authors are deeply grateful to all study partici-pants, to the Affymetrix Core Facility and genotypingunit at the Instituto Gulbenkian de Ciencia, and tothe neurologists and nurses of the Stroke andLaboratory Units of Vall d’Hebron Hospital for theircontributions. The Neurovascular Research Labora-tory takes part in the Spanish stroke genetics cons-ortium (GeneStroke), the international stroke genet-ics consortium (ISGC), and in the Spanish strokeresearch network (RENEVAS RD06/0026/0010).
Disclosure/conflict of interest
The authors declare no conflict of interest.
References
Adams Jr HP, Bendixen BH, Kappelle LJ, Biller J, Love BB,Gordon DL, Marsh III EE (1993) Classification of subtypeof acute ischemic stroke. Definitions for use in amulticenter clinical trial. TOAST. Trial of Org 10172in Acute Stroke Treatment. Stroke 24:35–41
Baird AE (2007) Blood genomics in human stroke. Stroke38:694–8
Barr TL, Conley Y, Ding J, Dillman A, Warach S, SingletonA, Matarin M (2010) Genomic biomarkers and cellularpathways of ischemic stroke by RNA gene expressionprofiling. Neurology 75:1009–14
Barrett JC, Fry B, Maller J, Daly MJ (2005) Haploview:analysis and visualization of LD and haplotype maps.Bioinformatics 21:263–5
Brazma A, Hingamp P, Quackenbush J, Sherlock G, Spell-man P, Stoeckert C, Aach J, Ansorge W, Ball CA, CaustonHC, Gaasterland T, Glenisson P, Holstege FC, Kim IF,Markowitz V, Matese JC, Parkinson H, Robinson A,Sarkans U, Schulze-Kremer S, Stewart J, Taylor R, Vilo J,Vingron M (2001) Minimum information about amicroarray experiment (MIAME)-toward standards formicroarray data. Nat Genet 29:365–71
Cheng H, Wang F, Ding X, Ding H, Song X (2009)Association of PRKCH gene with lacunar infarction ina local Chinese Han population. Neurosci Lett 464:146–149
Ding H, Xu Y, Bao X, Wang X, Cui G, Wang W, Hui R, WangDW (2010) Confirmation of genomewide associationsignals in Chinese Han population reveals risk loci forischemic stroke. Stroke 41:177–80
Domingues-Montanari S, Fernandez-Cadenas I, del Rio-EspinolaA, Corbeto N, Krug T, Manso H, Gouveia L, Sobral J,Mendioroz M, Fernandez-Morales J, Alvarez-Sabin J,Ribo M, Rubiera M, Obach V, Martı-Fabregas J, Freijo M,Serena J, Ferro JM, Vicente AM, Oliveira SA, Montaner J(2010a) Association of a genetic variant in the ALOX5-AP gene with higher risk of ischemic stroke. A case-control, meta-analysis and functional study. Cerebro-vasc Dis 29:528–37
Domingues-Montanari S, Fernandez-Cadenas I, Del Rıo-Espinola A, Mendioroz M, Fernandez-Morales J, CorbetoN, Delgado P, Ribo M, Rubiera M, Obach V, Martı-Fabregas J, Freijo M, Serena J, Montaner J (2010b)KCNK17 genetic variants in ischemic stroke. Athero-sclerosis 208:203–9
Domingues-Montanari S, Fernandez-Cadenas I, Del Rıo-Espinola A, Mendioroz M, Ribo M, Obach V, Marti-Fabregas J, Freijo M, Serena J, Corbeto N, Chacon P,Alvarez-Sabin J, Montaner J (2010c) The I/D polymorph-ism of the ACE1 gene is not associated withischaemic stroke in Spanish individuals. Eur J Neurol17:1390–2
Gonzalez JR, Armengol L, Sole X, Guino E, Mercader JM,Estivill X, Moreno V (2007) SNPassoc: an R package toperform whole genome association studies. Bioinfor-matics 23:644–5
Gretarsdottir S, Sveinbjornsdottir S, Jonsson HH, JakobssonF, Einarsdottir E, Agnarsson U, Shkolny D, Einarsson G,Gudjonsdottir HM, Valdimarsson EM, Einarsson OB,Thorgeirsson G, Hadzic R, Jonsdottir S, Reynisdottir ST,Bjarnadottir SM, Gudmundsdottir T, GudlaugsdottirGJ, Gill R, Lindpaintner K, Sainz J, Hannesson HH,Sigurdsson GT, Frigge ML, Kong A, Gudnason V,Stefansson K, Gulcher JR (2002) Localization of a sus-ceptibility gene for common forms of stroke to 5q12. AmJ Hum Genet 70:593–603
Gretarsdottir S, Thorleifsson G, Manolescu A, Styrkarsdot-tir U, Helgadottir A, Gschwendtner A, Kostulas K,Kuhlenbaumer G, Bevan S, Jonsdottir T, Bjarnason H,Saemundsdottir J, Palsson S, Arnar DO, Holm H,Thorgeirsson G, Valdimarsson EM, SveinbjornsdottirS, Gieger C, Berger K, Wichmann HE, Hillert J,Markus H, Gulcher JR, Ringelstein EB, Kong A,Dichgans M, Gudbjartsson DF, Thorsteinsdottir U,Stefansson K (2008) Risk variants for atrial fibrillationon chromosome 4q25 associate with ischemic stroke.Ann Neurol 64:402–9
Gretarsdottir S, Thorleifsson G, Reynisdottir ST, ManolescuA, Jonsdottir S, Jonsdottir T, Gudmundsdottir T,Bjarnadottir SM, Einarsson OB, Gudjonsdottir HM,Hawkins M, Gudmundsson G, Gudmundsdottir H,Andrason H, Gudmundsdottir AS, Sigurdardottir M,Chou TT, Nahmias J, Goss S, Sveinbjornsdottir S,Valdimarsson EM, Jakobsson F, Agnarsson U, GudnasonV, Thorgeirsson G, Fingerle J, Gurney M, GudbjartssonD, Frigge ML, Kong A, Stefansson K, Gulcher JR (2003)The gene encoding phosphodiesterase 4D confers risk ofischemic stroke. Nat Genet 35:131–8
Helgadottir A, Manolescu A, Thorleifsson G, GretarsdottirS, Jonsdottir H, Thorsteinsdottir U, Samani NJ,Gudmundsson G, Grant SF, Thorgeirsson G, Sveinb-jornsdottir S, Valdimarsson EM, Matthiasson SE,Johannsson H, Gudmundsdottir O, Gurney ME,
TTC7B and ischemic stroke riskT Krug et al
1070
Journal of Cerebral Blood Flow & Metabolism (2012) 32, 1061–1072

Sainz J, Thorhallsdottir M, Andresdottir M, Frigge ML,Topol EJ, Kong A, Gudnason V, Hakonarson H, GulcherJR, Stefansson K (2004) The gene encoding 5-lipoxy-genase activating protein confers risk of myocardialinfarction and stroke. Nat Genet 36:233–9
Ikram MA, Seshadri S, Bis JC, Fornage M, DeStefano AL,Aulchenko YS, Debette S, Lumley T, Folsom AR, vanden Herik EG, Bos MJ, Beiser A, Cushman M, Launer LJ,Shahar E, Struchalin M, Du Y, Glazer NL, RosamondWD, Rivadeneira F, Kelly-Hayes M, Lopez OL, Coresh J,Hofman A, DeCarli C, Heckbert SR, Koudstaal PJ, YangQ, Smith NL, Kase CS, Rice K, Haritunians T, Roks G, deKort PL, Taylor KD, de Lau LM, Oostra BA, UitterlindenAG, Rotter JI, Boerwinkle E, Psaty BM, Mosley TH,van Duijn CM, Breteler MM, Longstreth Jr WT, Wolf PA(2009) Genomewide association studies of stroke.N Engl J Med 360:1718–28
International Stroke Genetics Consortium, Wellcome TrustCase-Control Consortium 2 (2010) Failure to validateassociation between 12p13 variants and ischemicstroke. N Engl J Med 362:1547–50
Jørgensen HS, Nakayama H, Raaschou HO, Vive-Larsen J,Støier M, Olsen TS (1995) Outcome and time course ofrecovery in stroke. Part II: Time course of recovery. TheCopenhagen Stroke Study. Arch Phys Med Rehabil76:406–12
Krachler AM, Sharma A, Kleanthous C (2010) Self-association of TPR domains: lessons learned from adesigned, consensus-based TPR oligomer. Proteins78:2131–43
Krug T, Manso H, Gouveia L, Sobral J, Xavier JM,Albergaria I, Gaspar G, Correia M, Viana-Baptista M,Simoes RM, Pinto AN, Taipa R, Ferreira C, Fontes JR,Silva MR, Gabriel JP, Matos I, Lopes G, Ferro JM,Vicente AM, Oliveira SA (2010) Kalirin: a novelgenetic risk factor for ischemic stroke. Hum Genet127:513–23
Kubo M, Hata J, Ninomiya T, Matsuda K, Yonemoto K,Nakano T, Matsushita T, Yamazaki K, Ohnishi Y,Saito S, Kitazono T, Ibayashi S, Sueishi K, Iida M,Nakamura Y, Kiyohara Y (2007) A nonsynonymous SNPin PRKCH (protein kinase C eta) increases the risk ofcerebral infarction. Nat Genet 39:212–7
Larson MG, Atwood LD, Benjamin EJ, Cupples LA,D0Agostino Sr RB, Fox CS, Govindaraju DR, Guo CY,Heard-Costa NL, Hwang SJ, Murabito JM, Newton-ChehC, O0Donnell CJ, Seshadri S, Vasan RS, Wang TJ, WolfPA, Levy D (2007) Framingham Heart Study 100Kproject: genome-wide associations for cardiovasculardisease outcomes. BMC Med Genet 8:S5
Matarın M, Brown WM, Scholz S, Simon-Sanchez J, FungHC, Hernandez D, Gibbs JR, De Vrieze FW, Crews C,Britton A, Langefeld CD, Brott TG, Brown Jr RD,Worrall BB, Frankel M, Silliman S, Case LD, SingletonA, Hardy JA, Rich SS, Meschia JF (2007) A genome-wide genotyping study in patients with ischaemicstroke: initial analysis and date release. Lancet Neurol6:414–20
Montaner J, Fernandez-Cadenas I, Molina CA, Ribo M,Huertas R, Rosell A, Penalba A, Ortega L, Chacon P,Alvarez-Sabın J (2006) Poststroke C-reactive protein is apowerful prognostic tool among candidates for throm-bolysis. Stroke 37:1205–10
Moore DF, Li H, Jeffries N, Wright V, Cooper Jr RA,Elkahloun A, Gelderman MP, Zudaire E, Blevins G, YuH, Goldin E, Baird AE (2005a) Using peripheral bloodmononuclear cells to determine a gene expression
profile of acute ischemic stroke: a pilot investigation.Circulation 111:212–21
Moore DF, Wright V, Yu H, Cooper RA, Elkahloun A,Goldin E, Baird AE (2005b) Gene expression changesduring recovery from ischemic stroke: a longitudinalstudy and data analysis. Ann Neurol 58:S42–3
Navarro-Nunez L, Lozano ML, Rivera J, Corral J, Roldan V,Gonzalez-Conejero R, Iniesta JA, Montaner J, Vicente V,Martınez C (2007) The association of the b1-tubulinQ43P polymorphism with intracerebral hemorrhage inmen. Haematologica 92:513–8
Nilsson-Ardnor S, Janunger T, Wiklund PG, Lackovic K,Nilsson AK, Lindgren P, Escher SA, Stegmayr B,Asplund K, Holmberg D (2007) Genome-wide linkagescan of common stroke in families from northernSweden. Stroke 38:34–40
Nilsson-Ardnor S, Wiklund PG, Lindgren P, Nilsson AK,Janunger T, Escher SA, Hallbeck B, Stegmayr B,Asplund K, Holmberg D (2005) Linkage of ischemicstroke to the PDE4D region on 5q in a Swedishpopulation. Stroke 36:1666–71
Pruissen DM, Slooter AJ, Rosendaal FR, van der Graaf Y,Algra A (2008) Coagulation factor XIII gene variation,oral contraceptives, and risk of ischemic stroke. Blood111:1282–6
Qian HR, Huang S (2005) Comparison of false discoveryrate methods in identifying genes with differentialexpression. Genomics 86:495–503
Rosa A, Fonseca BV, Krug T, Manso H, Gouveia L,Albergaria I, Gaspar G, Correia M, Viana-Baptista M,Simoes RM, Pinto AN, Taipa R, Ferreira C, Fontes JR,Silva MR, Gabriel JP, Matos I, Lopes G, Ferro JM, VicenteAM, Oliveira SA (2008) Mitochondrial haplogroup H1 isprotective for ischemic stroke in Portuguese patients.BMC Med Genet 9:57
Sharp FR, Jickling GC, Stamova B, Tian Y, Zhan X, Liu D,Kuczynski B, Cox CD, Ander BP (2011) Molecularmarkers and mechanisms of stroke: RNA studies ofblood in animals and humans. J Cereb Blood Flow Metab31:1513–31
Storey JD (2002) A direct approach to false discovery rate.JR Stat Soc B 64:479–98
Tang Y, Xu H, Du X, Lit L, Walker W, Lu A, Ran R, Gregg JP,Reilly M, Pancioli A, Khoury JC, Sauerbeck LR,Carrozzella JA, Spilker J, Clark J, Wagner KR, JauchEC, Chang DJ, Verro P, Broderick JP, Sharp FR (2006)Gene expression in blood changes rapidly in neutro-phils and monocytes after ischemic stroke in humans: amicroarray study. J Cereb Blood Flow Metab 26:1089–1102
Tang Y, Lu A, Aronow BJ, Sharp FR (2001) Blood genomicresponses differ after stroke, seizures, hypoglycemia,and hypoxia: blood genomic fingerprints of disease.Ann Neurol 50:699–707
Toschke AM, Tilling K, Cox AM, Rudd AG, HeuschmannPU, Wolfe CD (2010) Patient-specific recovery pat-terns over time measured by dependence in activitiesof daily living after stroke and post-stroke care: theSouth London Stroke Register (SLSR). Eur J Neurol17:219–25
Xu H, Tang Y, Liu DZ, Ran R, Ander BP, Apperson M, LiuXS, Khoury JC, Gregg JP, Pancioli A, Jauch EC, WagnerKR, Verro P, Broderick JP, Sharp FR (2008) Geneexpression in peripheral blood differs after cardioem-bolic compared with large-vessel atherosclerotic stroke:biomarkers for the etiology of ischemic stroke. J CerebBlood Flow Metab 28:1320–8
TTC7B and ischemic stroke riskT Krug et al
1071
Journal of Cerebral Blood Flow & Metabolism (2012) 32, 1061–1072

Zee RY, Cook NR, Cheng S, Reynolds R, Erlich HA,Lindpaintner K, Ridker PM (2004) Polymorphism inthe P-selectin and interleukin-4 genes as determinantsof stroke: a population-based, prospective geneticanalysis. Hum Molec Genet 13:389–96
Zhan X, Ander BP, Jickling G, Turner R, Stamova B, XuH, Liu D, Davis RR, Sharp FR (2010) Brief focalcerebral ischemia that simulates transient ischemicattacks in humans regulates gene expression in ratperipheral blood. J Cereb Blood Flow Metab 30:110–8
Supplementary Information accompanies the paper on the Journal of Cerebral Blood Flow & Metabolism website (http://www.nature.com/jcbfm)
TTC7B and ischemic stroke riskT Krug et al
1072
Journal of Cerebral Blood Flow & Metabolism (2012) 32, 1061–1072




















