
Changing Drug Users’ Risk Environments: Peer Health Advocates as Multilevel Community Change Agents

Trophic level asynchrony in rates of phenological changefor marine, freshwater and terrestrial environmentsS T E P H E N J . T H A C K E R AY *, T I M O T H Y H . S PA R K S w 1 , M O R T E N F R E D E R I K S E N z, § , S A R A H
B U R T H E § , P H I L I P J . B A C O N } , J A M E S R . B E L L k, M A R C S . B O T H A M **, T O M M .
B R E R E T O N w w , PA U L W. B R I G H T zz, L A U R E N C E C A R VA L H O § , T I M C L U T T O N - B R O C K § § ,
A L I S T A I R D A W S O N § , M A R T I N E D WA R D S } } , J . M A L C O L M E L L I O T T kk, R I C H A R D
H A R R I N G T O N k, D AV I D J O H N S } } , I A N D . J O N E S *, J A M E S T . J O N E S ***, D AV I D I .
L E E C H w w w , D AV I D B . R O Y **, W. A N D Y S C O T T *, M AT T S M I T H zzz, R I C H A R D J .
S M I T H E R S § § § , I A N J . W I N F I E L D * and S A R A H WA N L E S S §
*Centre for Ecology and Hydrology Lancaster, Lancaster Environment Centre, Library Avenue, Bailrigg, Lancaster, LA1 4AP, UK,
wInstitute of Zoology, Poznan University of Life Sciences, Wojska Polskiego 71 C, 60-625 Poznan, Poland, zNERI, Aarhus
University, Frederiksborgvej 399, 4000 Roskilde, Denmark, §Centre for Ecology and Hydrology Edinburgh, Bush Estate, Penicuk,
Midlothian EH26 0QB, UK, }Marine Scotland, Freshwater Laboratory, Faskally, Pitlochry, Perthshire, PH16 5LB, UK,
kRothamsted Insect Survey, Department of Plant and Invertebrate Ecology, Rothamsted Research, Harpenden, Hertfordshire AL5
2JQ, UK, **Centre for Ecology and Hydrology Wallingford, Maclean Building, Benson Lane, Crowmarsh Gifford, Wallingford,
Oxfordshire OX10 8BB, UK, wwButterfly Conservation, Manor Yard, East Lulworth, Wareham, Dorset BH20 5QP, UK, zzSchool of
Biological Sciences, Royal Holloway University of London, Egham, Surrey TW20 0EX, UK, §§Department of Zoology, University of
Cambridge, Downing Street, Cambridge CB2 3EJ, UK, }}The Sir Alister Hardy Foundation for Ocean Science, The Laboratory,
Citadel Hill, Plymouth PL1 2PB, UK, kkThe Freshwater Biological Association, The Ferry Landing, Far Sawrey, Ambleside,
Cumbria LA22 0LP, UK, ***People’s Trust for Endangered Species, 15 Cloisters House, 8 Battersea Park Road, London SW8 4BG,
UK, wwwBritish Trust for Ornithology, The Nunnery, Thetford IP24 2PU, UK, zzzNational Pollen and Aerobiology Research Unit,
University of Worcester, Henwick Road, Worcester WR2 6AJ, UK, §§§The Woodland Trust, Autumn Park, Dysart Road, Grantham,
Lincolnshire NG31 6LL, UK
Abstract
Recent changes in the seasonal timing (phenology) of familiar biological events have been one of the most
conspicuous signs of climate change. However, the lack of a standardized approach to analysing change has
hampered assessment of consistency in such changes among different taxa and trophic levels and across freshwater,
terrestrial and marine environments. We present a standardized assessment of 25 532 rates of phenological change for
726 UK terrestrial, freshwater and marine taxa. The majority of spring and summer events have advanced, and more
rapidly than previously documented. Such consistency is indicative of shared large scale drivers. Furthermore,
average rates of change have accelerated in a way that is consistent with observed warming trends. Less coherent
patterns in some groups of organisms point to the agency of more local scale processes and multiple drivers. For the
first time we show a broad scale signal of differential phenological change among trophic levels; across environments
advances in timing were slowest for secondary consumers, thus heightening the potential risk of temporal mismatch
in key trophic interactions. If current patterns and rates of phenological change are indicative of future trends, future
climate warming may exacerbate trophic mismatching, further disrupting the functioning, persistence and resilience
of many ecosystems and having a major impact on ecosystem services.
Keywords: climate, linear mixed effects models, meta-analysis, phenology, traits, trophic mismatch
Received 9 October 2009 and accepted 9 December 2009
Introduction
Recent climate warming has dramatically altered the
distribution, abundance and population dynamics of
many aquatic and terrestrial organisms (IPCC, 2007).
Acting in concert with other ecological stressors, cli-
matic change is projected to weaken the resilience of
many ecosystems, leading to sudden re-organizations
of communities and drastic alterations to ecosystem
structure and function (Scheffer & Carpenter, 2003).
Recorded changes in the seasonal timing, or phenology,
of life history events have formed a key part of this
assessment, with spring and summer events generally
Correspondence: Stephen J. Thackeray, tel. 1 44 01524 595852, fax
1 44 01524 61536, e-mail: [email protected]
1Formerly: Centre for Ecology and Hydrology Monks Wood,
Abbots Ripton, Huntingdon, Cambridgeshire PE28 2LS, UK.
Global Change Biology (2010) 16, 3304–3313, doi: 10.1111/j.1365-2486.2010.02165.x
3304 r 2010 Blackwell Publishing Ltd

becoming earlier as temperatures have risen. Species-
specific variation in phenological responses to climate
can disrupt the synchrony of ecological interactions
(Harrington et al., 1999; Visser & Both, 2005) and poten-
tially affect community persistence. To assess the poten-
tial risk of phenological asynchrony affecting community
structure, it is crucial to determine how rates of pheno-
logical change vary among species, functional groups
and trophic levels.
Assessments of broad-scale taxonomic variation in
phenological change have generally involved meta-
analyses in which previously published results on
changes in the timing of biological events are extracted
from the literature and combined to provide overall
rates of change. While there has been broad unanimity
that spring events in the northern hemisphere have
become earlier, with estimated mean advances ranging
from 2.3 days per decade (Parmesan & Yohe, 2003) to
5.5 days per decade (Root et al., 2003), there has been a
lack of consensus on which taxonomic groups show the
most, and least, change. Differences in the data selection
criteria adopted by different meta-analyses have been
cited as a contributory factor to this lack of consistency
(Englund et al., 1999; Parmesan, 2007). Reliance on
published studies creates two further potential pro-
blems. Firstly, positive publication bias (Gurevitch
et al., 2001) leads to the under-representation of taxa
showing little or no phenological change. Researchers
have sought to overcome this by restricting analyses to
multispecies studies that include nonresponding spe-
cies (Parmesan & Yohe, 2003), though there is no
assurance that such assemblages are representative of
the wider community. Secondly, despite the fact that
rates (and even directions) of phenological change are
known to vary over time (Crick & Sparks, 1999; Thack-
eray et al., 2008), the lengths and periods of phenologi-
cal time series used in meta-analyses are rarely
standardized (Badeck et al., 2004; Parmesan, 2007).
Thus, apparent taxonomic differences in rates of change
can be generated by differences in time series start year,
end year and length, rather than by biologically relevant
characteristics.
To date analyses overcoming such problems by using
raw time series data and consistent time scales rather
than published values have been mainly restricted to
species comparisons within major taxonomic or func-
tional groups, such as plankton (Edwards & Richard-
son, 2004), terrestrial plants (Fitter & Fitter, 2002;
Menzel et al., 2006), insects (Roy & Sparks, 2000; Stefa-
nescu et al., 2003) and birds (Cotton, 2003; Both &
Artemyev, 2004). Here, we apply this approach across
environments to obtain the first directly comparable
estimates of phenological change for UK terrestrial,
freshwater and marine taxa between 1976 and 2005.
This has been a period of major environmental change
with factors such as eutrophication (including atmo-
spheric deposition of nutrients), acid rain, land-use
change, overfishing and, of course, climate change
impacting on natural systems (Smart et al., 2003; Mil-
lenium Ecosystem Assessment, 2005).
A number of functional traits might be expected to
influence the rate of phenological change demonstrated
by organisms. It might be expected that the mode of
thermal physiology would be particularly influential.
Developmental rates of ectotherms are, by definition,
directly affected by habitat warming (Precht et al., 1973).
One might hypothesize therefore that they have shown
the most rapid mean rates of phenological advance. It
has been argued that generation time should affect
evolutionary rates in response to selective pressures
imposed by environmental change (Rosenheim &
Tabashnik, 1991; Berteaux et al., 2004) and so we might
expect this trait to affect rates of phenological change,
assuming a role for microevolution. At the scale of
single ecosystems, differential phenological change
among trophic levels has frequently been observed
(Harrington et al., 1999; Visser & Both, 2005) though it
is not clear whether mean rates of change differ sig-
nificantly among trophic levels at larger scales of eco-
logical organization (Blenckner & Hillebrand, 2002), or
whether these patterns are idiosyncratic. Also, choice of
phenological metric (Miller-Rushing et al., 2008; Van
Buskirk et al., 2009) may affect observed rates of change.
The aims of this study were to (i) provide a fully
standardized and unbiased assessment of rates of phe-
nological change in the United Kingdom and (ii) to
resolve differences in rates of change among environ-
ments and major taxonomic and functional groups. We
explicitly considered the following functional attri-
butes; thermal physiology, generation time, environ-
ment (where each species feeds/gathers resources
during the focal phenological event) and trophic level.
The latter attribute allowed us to compute average rates
for each trophic level and thus evaluate, across the three
environments, likely risks of trophic mismatching
i.e. reduced synchrony between the seasonal occurrence
of food resources and the peak demand for those
resources.
Materials and methods
Quantifying rates of change
All rates of change were calculated from the raw observational
data for each nonoverlapping 10-year period between 1976
and 2005, and for the whole 30-year period.
We selected only records indicating the beginning or middle
of phenological events, occurring on average between January
P H E N O L O G I C A L C H A N G E A C R O S S M A J O R E N V I R O N M E N T S 3305
r 2010 Blackwell Publishing Ltd, Global Change Biology, 16, 3304–3313

and August. Thus, our emphasis was on spring and summer
events rather than those occurring in autumn or winter.
All phenological indicators were the standard metrics
used by each of the monitoring programmes represented in
the analysis (supporting information, Table S1). Linear regres-
sion was used to quantify phenological trends over each
specified time period. Annual seasonal timing (day of year)
of phenological events was analysed as a function of year
by linear regression. Slope coefficients quantified rates of
change in days per year. Regressions were performed for
each nonoverlapping 10-year period between 1976 and 2005
and for the whole 30-year period. Trends were only retained
for meta-analysis if phenological data were available for 80%
or more of the years within the respective time period. Con-
sidering all taxa, sites and time periods this yielded 25 532
rates of phenological change for 726 distinct taxa (supporting
information, Table S2) including terrestrial plants, freshwater
and marine phytoplankton and zooplankton, insects, amphi-
bians, birds, fish and mammals (supporting information, Table
S2). Phenological records included dates of flowering and
leafing, plankton population growth, insect flight periods,
births and migration. The analysis included data from across
the whole United Kingdom, spanning the latitudinal range
49.9–60.91N.
Categorization of phenological trends
For the meta-analysis, phenological trends were categorized
by major taxonomic group (plants/phytoplankton, inverte-
brates, vertebrates) and by functional attributes: environment
(marine, freshwater, terrestrial), trophic level (primary produ-
cer, primary consumer, secondary consumer), thermal physiol-
ogy (ectothermic, endothermic) and generation time (o1
week, 1 week–1 month, 1 month–1 year, 41 year). Previous
studies have shown a tendency for first events to shift more
rapidly than middle or peak events (Miller-Rushing et al., 2008;
Van Buskirk et al., 2009), and so we also included this dichot-
omy as a factor in our analyses. Species that used more than
one major environment (e.g. amphibians, dragonflies, sea-
birds) were assigned to the environment with which they
were most closely associated for feeding, during the focal
phenological event.
Differences in time series lengths among monitoringprogrammes
Some of the monitoring programmes represented could only
provide phenological data for a single decade from the 30-year
period (supporting information, Table S1). It is possible that
among-decade differences in phenological change were a
statistical artefact of having a different assemblage of monitor-
ing programmes represented in each decade. We therefore
calculated mean rates of change separately for more finely
resolved taxonomic groups that were each represented by a
single monitoring programme in all three decades. Patterns of
phenological change present in these more consistently re-
corded taxa were used to confirm patterns of change observed
in the whole dataset.
Summarizing rates of change
For each major taxonomic group and habitat type, both the
mean rate of change and variability in rates of change were
calculated. To test whether trends have on balance shifted
either towards earliness or lateness, two-tailed binomial tests
were used to test the null hypothesis that trends were symme-
trically distributed about zero. Differences in trend variability
among groups were assessed using Levene’s tests, after An-
derson–Darling tests showed that data were not normally
distributed.
Linear mixed effects modelling
Linear mixed effects models were used to test for significant
differences in mean rates of change among taxonomic groups,
and with organism functional traits. This was carried out in
SAS 9.1 using PROC MIXED. The following random effects were
specified: data source, species nested in data source, site
nested in data source, and brood/generation nested in data
source. The collated datasets, each of which focussed on a
particular taxonomic or functional group, varied in size. By
including data source as a random effect, each dataset was
given equal weight in the analysis so that results were not
unduly influenced by any one, well represented, group. In
addition, this explicitly allowed nonindependence among
trends from closely related organisms, recorded using the
same sampling protocols. Data points (i.e. regression slopes)
were weighted by inverse variance. Tested fixed effects were
major taxonomic group, metric type (beginning or middle
event) and organism functional attributes. Denominator
degrees of freedom for fixed effects were estimated using
Satterthwaite’s approximation. All significance levels given
are from type III tests. Figures show marginal least-squares
means � 1 SE.
Results
Widespread phenological change across environments
Rapid climate change occurred over the study period:
all three environments experienced very similar warm-
ing trends (0.04–0.05 1C yr�1), with temperatures fre-
quently above the 1976–2005 mean since the late 1980s
(Fig. 1). During this period, the seasonal timing of
biological events in all major taxonomic groups in UK
terrestrial, freshwater and marine environments ad-
vanced, on average, by 0.39 d yr�1 (equivalent to 11.7
days over the whole period). Overall, 83.8% of pheno-
logical trends were towards earlier seasonal timing. In
all groups the proportion of negative trends (i.e. to-
wards earliness) was significantly greater than expected
if trends were symmetrical about zero (Fig. 2a) and the
percentage of statistically significant phenological
trends exceeded that expected by chance alone (5%) at
the P 5 0.05 significance level.
3306 S . J . T H A C K E R AY et al.
r 2010 Blackwell Publishing Ltd, Global Change Biology, 16, 3304–3313

A taxonomically resolved analysis showed that mean
rates of change were significantly different among
taxonomic groups, though not among environments
(linear mixed effects model, LME, n 5 3419, interaction:
F4,71.8 5 1.47, P 5 0.22, environment: F2,152 5 0.34, P 5 0.71,
taxonomic group: F2,73.4 5 3.58, P 5 0.033). Leafing,
flowering and fruiting dates of terrestrial plants showed
the most rapid mean rate of change (0.58 d yr�1, Fig. 2b)
and the highest percentage of advancing trends (92.5%,
Fig. 2a). Freshwater plants (phytoplankton bloom tim-
ings) had the slowest rate of response (0.23 d yr�1 ear-
lier, Fig. 2b) and showed the lowest percentage of
advancing trends (62.2%, Fig. 2a). There was also
significant heterogeneity of variance in phenological
trends among taxa (Levene’s test, W8,3411 5 39.73,
Po0.001) with marine and freshwater plants and in-
vertebrates exhibiting much higher variability in rates
of change (SD range 0.74–0.98) than other groups (SD
range 0.30–0.53).
Decadal variation in rates of change
Examination of phenological changes at a finer tempor-
al resolution indicated that at the decadal scale, trends
varied markedly among taxonomic groups and envir-
onments (Fig. 3). For many taxa, rates of phenological
change have accelerated during recent decades. In gen-
eral, timing advanced least rapidly before 1986 except
for marine plants (phytoplankton bloom timings) and
marine vertebrates (migration and spawning/egg lay-
ing in marine fish and marine birds). In some cases
there was a reversal of phenological trends between the
1976–1985 and 1986–1995 periods i.e. a period of phe-
nological delay was followed by a period of phenolo-
gical advance (marine and freshwater invertebrates,
freshwater plants). For some groups mean rates of
advance were consistently higher during 1986–2005
than during 1976–1985 (e.g. freshwater and terrestrial
plants, marine and freshwater invertebrates), for others
rates of advance declined considerably post 1996 (e.g.
terrestrial vertebrates). These results were robust when
only analysing taxa that were consistently recorded
throughout the whole 30-year period (Table 1). Results
Fig. 1 January–August mean temperatures from 1976 to 2005
for the United Kingdom. Presented are UK mean air tempera-
tures (blue line), mean sea surface temperature 101W–51E, 501N–
601N (red line) and mean lake surface water temperatures
(brown line). All series presented as anomalies from the 1976
to 2005 mean. Air temperatures provided by the UK Met Office
and sea surface anomalies by the UK Met Office Hadley Centre.
Lake temperatures are means of those recorded at the sites
included in this study: the northern and southern basins of
Windermere, Esthwaite Water and Loch Leven.
Fig. 2 Phenological change for the UK flora and fauna from
1976 to 2005. (a) percentages of advancing (below horizontal)
and delaying (above horizontal) trends for each taxon–environ-
ment combination. Statistically significant advancing and delay-
ing trends are indicated by black shading. Nonsignificant trends
are indicated by white shading. The number of trends analysed
for each taxon-environment combination (n) is given above each
bar. Also shown is the significance level (P) of a two-tailed
binomial test of the null hypothesis that negative and positive
trends are equally likely. (b) Mean � SEM rates of change for
plants/phytoplankton (plant; green bars), invertebrates (invert;
yellow bars) and vertebrates (vert; blue bars) in marine, fresh-
water and terrestrial environments. All mean trends are nega-
tive, indicating an advance of phenological events. The taxa
included in each taxonomic group-environment combination,
and the number of trends per taxon, are given in supporting
information, Table S3.
P H E N O L O G I C A L C H A N G E A C R O S S M A J O R E N V I R O N M E N T S 3307
r 2010 Blackwell Publishing Ltd, Global Change Biology, 16, 3304–3313

based on these taxa confirmed the acceleration of phe-
nological change observed in the full dataset: rates of
advancing phenology were again lowest during 1976–
1985, with higher rates of advance occurring during
1986–1995 (e.g. butterfly first flight and bird arrival
dates) or 1986–2005 (e.g. freshwater plankton and ter-
restrial plants). Flowering dates and aphid flight dates
advanced particularly rapidly, by more than 1.0 d yr�1
(10.0 days per decade) after 1986, as did moth flight
dates during 1986–1995. The major exception to this
overall trend was marine vertebrates, which showed
little phenological change after 1986.
Trophic and functional variation in rates of change
Phenological trends varied significantly among trophic
levels over the whole 30-year period (LME, n 5 3419,
F2,63.7 5 4.16, P 5 0.020). In particular, events associated
with secondary consumers advanced less rapidly than
those for both primary producers and primary consu-
mers (Fig. 4a). The nonsignificant interaction between
trophic level and environment (LME, n 5 3419,
F4,139 5 0.99, P 5 0.42) would suggest that this has been
a general phenomenon in marine, freshwater and ter-
restrial ecosystems. Analysis of decadal phenological
trends confirmed the difference in rates of change
among trophic levels (LME, n 5 22113, F2,116 5 7.46,
P 5 0.0009). There was a significant acceleration in over-
all rates of advance post 1986 (LME, n 5 22113,
F2,16*103 5 37.7, Po0.0001) but this was less pronounced
Fig. 3 Decadal variations in rates of phenological change. Dec-
adal mean � SEM rates of change for plants/phytoplankton
(plant; green bars), invertebrates (invert; yellow bars) and verte-
brates (vert; blue bars) in marine, freshwater and terrestrial
environments. For each taxon–environment combination, dec-
ades appear chronologically: 1976–1985 (1), 1986–1995 (2) and
1996–2005 (3). Negative trends indicate advance, and positive
trends delaying, of phenological events. The number of trends
analysed for each taxon–environment combination, in each
decade, is given above each bar.T
ab
le1
Mea
n�
SE
Mtr
end
s(s
amp
lesi
ze)
for
var
iou
sm
on
ito
rin
gsc
hem
esan
dp
hen
olo
gic
alm
etri
cs.
Tre
nd
sar
esu
mm
ariz
edfo
rea
cho
fth
e10
-yea
rp
erio
ds
1976
–198
5,19
86–
1995
and
1996
–200
5,an
do
ver
all
for
the
30-y
ear
per
iod
1976
–200
5
Rec
ord
ing
sch
eme
Ph
eno
log
ical
met
ric
1976
–198
5(d
yr�
1)
1986
–199
5(d
yr�
1)
1996
–200
5(d
yr�
1)
1976
–200
5(d
yr�
1)
RIS
:A
ph
ids
Fir
stfl
igh
t0.
10�
0.18
(114
)�
2.35�
0.18
(114
)�
1.64�
0.19
(114
)�
0.87�
0.04
(114
)
5th
per
cen
tile
cum
ula
tiv
ean
nu
alca
tch
0.53�
0.14
(114
)�
1.51�
0.14
(114
)�
1.28�
0.12
(114
)�
0.59�
0.05
(114
)
UK
BM
S:
Bu
tter
flie
sF
irst
flig
ht
�0.
58�
0.08
(104
6)�
0.98�
0.05
(227
9)�
0.58�
0.03
(541
0)�
0.42�
0.02
(115
6)
Mea
nfl
igh
td
ate
�0.
11�
0.05
(104
4)�
0.85�
0.03
(227
2)�
0.51�
0.02
(539
1)�
0.35�
0.01
(115
4)
BT
O:
Bir
ds
Mea
nfi
rst
egg
dat
e0.
05�
0.10
(51)
�0.
47�
0.09
(63)
�0.
27�
0.11
(55)
�0.
19�
0.03
(56)
Co
un
tyb
ird
rep
ort
sF
irst
arri
val
�0.
26�
0.07
(238
)�
0.64�
0.07
(254
)�
0.10�
0.06
(250
)�
0.31�
0.02
(239
)
CE
H:
Fre
shw
ater
pla
nk
ton
50%
max
.m
on
thly
mea
nab
un
dan
ce0.
42�
0.40
(114
)�
0.13�
0.30
(150
)�
0.60�
0.38
(130
)�
0.23�
0.09
(126
)
Blo
om
cen
tre
of
gra
vit
y0.
75�
0.34
(94)
�0.
38�
0.27
(129
)�
0.48�
0.32
(120
)�
0.22�
0.09
(108
)
RIS
:M
oth
sD
ayo
fm
edia
nca
tch
0.14�
0.09
(39)
�1.
01�
0.13
(39)
�0.
53�
0.09
(38)
�0.
33�
0.04
(39)
SA
HF
OS
:M
arin
ep
lan
kto
nB
loo
mce
ntr
eo
fg
rav
ity
�0.
20�
0.31
(101
)�
0.54�
0.26
(152
)�
0.51�
0.26
(164
)�
0.38�
0.07
(136
)
UK
PN
:P
lan
tsF
irst
flo
wer
ing
�0.
02�
0.07
(270
)�
1.31�
0.09
(300
)�
1.75�
0.22
(105
)�
0.57�
0.07
(50)
UK
BM
S:
UK
Bu
tter
fly
Mo
nit
ori
ng
Sch
eme,
BT
O:
Bri
tish
Tru
stfo
rO
rnit
ho
log
y,C
EH
:C
entr
efo
rE
colo
gy
and
Hy
dro
log
y,R
IS:
Ro
tham
sted
Inse
ctS
urv
ey,
SA
HF
OS
:S
irA
list
er
Har
dy
Fo
un
dat
ion
for
Oce
anS
cien
ce,
UK
PN
:U
KP
hen
olo
gy
Net
wo
rk
3308 S . J . T H A C K E R AY et al.
r 2010 Blackwell Publishing Ltd, Global Change Biology, 16, 3304–3313
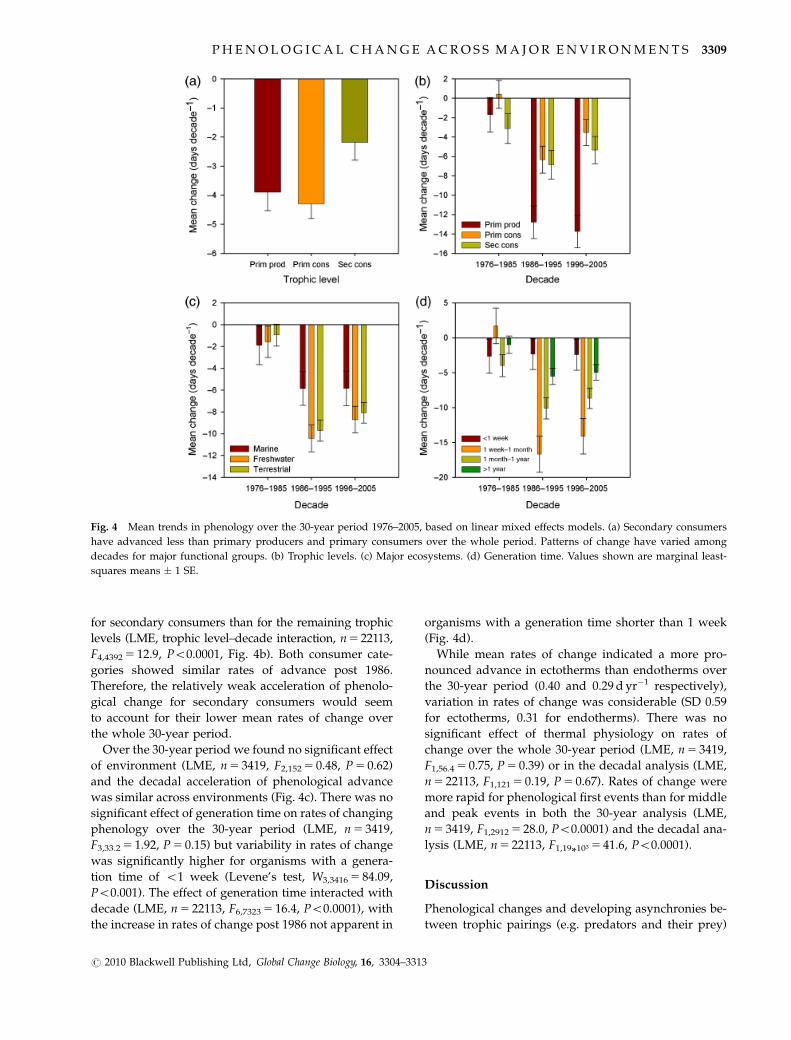
for secondary consumers than for the remaining trophic
levels (LME, trophic level–decade interaction, n 5 22113,
F4,4392 5 12.9, Po0.0001, Fig. 4b). Both consumer cate-
gories showed similar rates of advance post 1986.
Therefore, the relatively weak acceleration of phenolo-
gical change for secondary consumers would seem
to account for their lower mean rates of change over
the whole 30-year period.
Over the 30-year period we found no significant effect
of environment (LME, n 5 3419, F2,152 5 0.48, P 5 0.62)
and the decadal acceleration of phenological advance
was similar across environments (Fig. 4c). There was no
significant effect of generation time on rates of changing
phenology over the 30-year period (LME, n 5 3419,
F3,33.2 5 1.92, P 5 0.15) but variability in rates of change
was significantly higher for organisms with a genera-
tion time of o1 week (Levene’s test, W3,3416 5 84.09,
Po0.001). The effect of generation time interacted with
decade (LME, n 5 22113, F6,7323 5 16.4, Po0.0001), with
the increase in rates of change post 1986 not apparent in
organisms with a generation time shorter than 1 week
(Fig. 4d).
While mean rates of change indicated a more pro-
nounced advance in ectotherms than endotherms over
the 30-year period (0.40 and 0.29 d yr�1 respectively),
variation in rates of change was considerable (SD 0.59
for ectotherms, 0.31 for endotherms). There was no
significant effect of thermal physiology on rates of
change over the whole 30-year period (LME, n 5 3419,
F1,56.4 5 0.75, P 5 0.39) or in the decadal analysis (LME,
n 5 22113, F1,121 5 0.19, P 5 0.67). Rates of change were
more rapid for phenological first events than for middle
and peak events in both the 30-year analysis (LME,
n 5 3419, F1,2912 5 28.0, Po0.0001) and the decadal ana-
lysis (LME, n 5 22113, F1,19*103 5 41.6, Po0.0001).
Discussion
Phenological changes and developing asynchronies be-
tween trophic pairings (e.g. predators and their prey)
Fig. 4 Mean trends in phenology over the 30-year period 1976–2005, based on linear mixed effects models. (a) Secondary consumers
have advanced less than primary producers and primary consumers over the whole period. Patterns of change have varied among
decades for major functional groups. (b) Trophic levels. (c) Major ecosystems. (d) Generation time. Values shown are marginal least-
squares means � 1 SE.
P H E N O L O G I C A L C H A N G E A C R O S S M A J O R E N V I R O N M E N T S 3309
r 2010 Blackwell Publishing Ltd, Global Change Biology, 16, 3304–3313

have been linked to reductions in individual fitness and
declines in the population size of focal species (Platt
et al., 2003; Winder & Schindler, 2004; Both et al., 2006;
M�ller et al., 2008), increasing the risk of population
extinctions and biodiversity loss. Such changes are also
of great economic and societal importance due to the
influence of phenological synchrony on processes such
as pollination (Elzinga et al., 2007), fisheries production
(Cushing, 1990) and herbivory by agricultural pests
(Harrington et al., 2007). Our analysis of the most
comprehensive UK phenological data collection to date
has shown that the timing of spring and summer events
has become earlier for the majority of taxa, and at a
more rapid rate than previously reported. The phenom-
enon is widespread across the terrestrial, freshwater
and marine environments and rates of change differ
significantly among trophic levels. Our results therefore
suggest a heightened risk that phenological asynchrony
may disrupt the stability and functioning of aquatic and
terrestrial ecosystems, and the delivery of key ecosys-
tem services.
Previous analyses, restricted to taxa showing signifi-
cant trends in the northern temperate zone, suggest
mean rates of advance of 4.4–5.5 days per decade (Root
et al., 2003; Root et al., 2005). A recent synthesis (Parme-
san, 2007) calculated an ‘unbiased’ estimated mean
phenological advance of 2.8 days per decade by analys-
ing published rates of change from taxa showing both
significant and nonsignificant phenological trends. Our
unbiased, standardized analysis reports a substantially
higher rate of advance. This comes contrary to expecta-
tion: our avoidance of potentially biased published
results, and inclusion of trends from ‘nonresponding’
taxa, should result in a lower mean rate of change. The
discrepancy probably arises due to the standardization
of time series lengths, and the focus on a period with a
consistent warming trend. In previous meta-analyses
some taxa were represented over longer time scales,
including periods of stable and decreasing mean tem-
peratures. Since warming trends are projected to inten-
sify (IPCC, 2007), we believe our results to be indicative
of a period of continuing, possibly increasing, phenolo-
gical change.
For the first time we have explicitly demonstrated
that there has been systematic variation in rates of
phenological change among trophic levels, and that this
has been apparent across marine, freshwater and ter-
restrial environments in the United Kingdom. Specifi-
cally, phenological advances have been slowest for
secondary consumers over the 30-year period. This
appears to result from a slower acceleration of pheno-
logical advance for this group, compared with the lower
trophic levels. Numerous potential trophic interactions
exist between the primary and secondary consumers
represented in our analysis e.g. amphibians and fish
consume both plankton and aquatic invertebrate larvae
such as dragonflies and alderflies, adult dragonflies can
consume members of the Lepidoptera and Hymeno-
ptera, many of the terrestrial bird species provision
their chicks on Lepidoptera, Hemiptera and Coleoptera
while marine fish and birds utilize larval fish such as
sandeels Ammodytes marinus. Our analysis would there-
fore suggest an increasing likelihood of trophic mis-
matching. Although developing trophic asynchronies
have been observed within single communities (Har-
rington et al., 1999; Visser & Both, 2005), generalizations
regarding rates of change among trophic levels have
proven elusive. Patterns of change within food webs
have appeared idiosyncratic, with different trophic
levels showing the most rapid rates of change in differ-
ent communities and ecosystems (Visser & Both, 2005).
This has prohibited the detection of systematic variation
in the effects of climate on phenological change at
different trophic levels (Blenckner & Hillebrand, 2002).
Our standardized, unbiased analysis has made
such detection possible. We also show for the first time
that accelerating rates of change, previously identified
for single species or small groups of species (Crick &
Sparks, 1999; Thackeray et al., 2008), have been cohe-
rent across ecosystems and have potentially widened
existing trophic asynchronies. The timing of accelera-
tion corresponds with a transition between periods
when temperatures were frequently below, and fre-
quently above, the 1976–2005 mean. This suggests
that this increase in rates of phenological advance has
been a response to climate warming over the 30-year
period.
There is much variability in the direction and magni-
tude of phenological trends. This occurs because taxa/
populations (i) differ in the extent to which life history
events are able to accelerate with warming, (ii) experi-
ence different warming trends due to variations in
mean seasonal timing and microhabitat use, (iii)
vary in the extent to which their phenological responses
are driven/constrained by factors other than increases
in temperature and (iv) may respond to changing
climate in other ways, e.g. distributional changes.
Alternative drivers of phenological change will vary
regionally and include atmospheric nutrient deposition
and precipitation (Penuelas et al., 2002; Cleland et al.,
2006), nutrient enrichment of aquatic ecosystems
(Thackeray et al., 2008) and variations in popula-
tion size (Langvatn et al., 2004; Miller-Rushing et al.,
2008) and age structure (Langvatn et al., 2004; Gillet &
Dubois, 2007). Resolution of the mechanisms under-
pinning variations in rates of change is crucial to
projecting impacts of these changes. We adopted a
functional approach to this problem, though the sources
3310 S . J . T H A C K E R AY et al.
r 2010 Blackwell Publishing Ltd, Global Change Biology, 16, 3304–3313

of variation outlined above and correlations among
functional traits render mechanistic understanding a
major challenge. We examined expectations that rates
of change would be highest for ectotherms and short
generation time organisms; attributes which should
enhance the potential for phenological advance with
warming.
Ectotherms had a higher mean rate of change than
endotherms, as might be expected. The phenology of
ectotherm life history events will be directly influenced
by warming while endotherms will respond more in-
directly, via ecosystem effects. The difference will be
exacerbated because birds, the most well-represented
endotherms in the analysis, demonstrate constrained
phenological plasticity due to photoperiodic induction
of gonad maturation and migration, especially for long-
distance migrants (Dawson, 2008). Despite the apparent
difference in rates of change between ectotherms and
endotherms, there was considerable variability around
the mean rates of change for each thermal physiology
category and the difference was not significant. Varia-
bility was highest for phytoplankton, which also
showed the highest incidences of phenological delays.
Despite having the shortest generation times of the
analysed taxa, the phytoplankton showed compara-
tively low mean rates of change over the 30-year period.
While individual phytoplankton taxa can show very
rapid phenological shifts, great trend variability ap-
pears to have prevented mean rates of change from
being exceptional. Additionally, although phytoplank-
ton growth is directly affected by temperature, bloom
timings reflect population dynamics that are also influ-
enced by light and nutrient availability, grazing and
sedimentation (Thackeray et al., 2008) and interspecific
interactions that have potentially unpredictable out-
comes (Beninca et al., 2008).
In contrast to previous meta-analyses (Parmesan,
2007), we found that terrestrial plants have demon-
strated the fastest mean phenological advance and the
highest proportion of phenological advances. This per-
haps indicates the primacy of temperature as a limiting
factor for plant growth at mid northern latitudes (Ba-
deck et al., 2004) and that temperature change has not
been great enough to induce delaying effects such as
heat stress in summer (Sherry et al., 2007) or insufficient
winter chilling (Thompson & Clark, 2008). However,
warming is also believed to be the predominant driver
of change for other phenological metrics, such as butter-
fly flight periods (Roy & Sparks, 2000). The rapid and
consistent phenological advances for terrestrial plants
cannot therefore be explained by the fact that they
respond the least to alternative drivers. Plant develop-
ment may simply accelerate more rapidly with warm-
ing than developmental rates for other taxa. It is also
possible that, since individual plants are sessile, they
cannot modulate the warming they experience by mov-
ing among microhabitats. This might result in stronger
and more consistent phenological trends for this
group.
It is possible that effects of thermal physiology and
generation time were masked by correlations among
functional traits. Trophic level and thermal physiology
are, to an extent, correlated. Endotherms were repre-
sented only in the consumer categories, such that the
fastest rates of phenological change and acceleration
occurred in the only trophic level occupied solely by
ectotherms. It is plausible that consumers showed lower
rates of advance and acceleration because of the influ-
ence of endotherms, but that the independent effect of
thermal physiology could not be resolved in the analy-
sis. Generation time covaries strongly with trophic level
in aquatic ecosystems, generally increasing higher up
the food web, but it would seem that this trait had a
limited influence on rates of change. The taxonomic
group demonstrating the most rapid change, terrestrial
plants, did not have the shortest generation times.
Furthermore, evidence for rapid microevolution in re-
sponse to warming is equivocal, with phenotypic plas-
ticity playing a dominant role in phenological
adjustment (Berteaux et al., 2004; Charmantier et al.,
2008). Interestingly, the expected decrease in rates of
change with increasing generation time appears to be
maintained post 1986, during the period of most rapid
overall phenological change, and for organisms with
generation times in excess of 1 week. Given the correla-
tions among functional traits, a major challenge for
phenology research is to determine the traits responsi-
ble for rates of change and to establish the ecological
scale at which independent effects of these traits can be
disentangled.
Climate change has been identified as one of the most
critical threats to the maintenance of global biodiversity.
Our findings indicating earlier occurrence of many
spring and summer biological events across a wide
range of terrestrial, freshwater and marine taxa should
aid current and future assessment of the impacts of
climate change. Although the precise consequences of
phenological changes on ecosystem functioning are not
clear, our study highlights the potential risk of desyn-
chronizing trophic linkages between primary and sec-
ondary consumers. Such disruption could have
catastrophic environmental consequences and affect
delivery of important ecosystem services, particularly
food production. Given the broad taxonomic and en-
vironmental scales at which our analyses were con-
ducted, it is reasonable to expect that our findings
may generalize to other mid latitude regions and have
relevance at a global level.
P H E N O L O G I C A L C H A N G E A C R O S S M A J O R E N V I R O N M E N T S 3311
r 2010 Blackwell Publishing Ltd, Global Change Biology, 16, 3304–3313

Acknowledgements
We thank the following for collecting and allowing access todata: Miss Ursula Allitt, Belfast HSC Trust, Cardiff School ofHealth Sciences UWIC, Prof. Jean Emberlin, Environmental andPublic Protection Offices Islington, the Freshwater BiologicalAssociation, Lancashire Immunology Service (Royal PrestonHospital), the National Dormouse Monitoring Programme, Uni-versity of Plymouth School of Geography. We thank the follow-ing organizations for funding: BBSRC, Countryside Council forWales, Defra, Forestry Commission, Joint Nature ConservationCommittee (JNCC), The Lawes Agricultural Trust, Natural Eng-land, NERC, Northern Ireland Environment Agency, ScottishNatural Heritage. The Nest Record Scheme is supported by theJNCC/BTO partnership. We thank Suzanne Clark and PeterRothery for assistance with coding statistical models, and BjornBeckmann for assistance with data extraction. We are indebted tothe many thousands of volunteer recorders who have contrib-uted records to many of the datasets we have analysed. Theauthors have no conflicts of interest to declare. We thank Prof.Mark Bailey and Dr Alex Elliott for their comments on an earlierversion of the manuscript. This analysis was funded by theCentre for Ecology and Hydrology Environmental Change In-tegrating Fund project SPACE (Shifting Phenology: AttributingChange across Ecosystems).
References
Badeck FW, Bondeau A, Bottcher K, Doktor D, Lucht W, Schaber J, Sitch S (2004)
Responses of spring phenology to climate change. New Phytologist, 162, 295–309.
Beninca E, Huisman J, Heerkloss R et al. (2008) Chaos in a long-term experiment with a
plankton community. Nature, 451, 822–825.
Berteaux D, Reale D, Mcadam AG, Boutin S (2004) Keeping pace with fast climate
change: can arctic life count on evolution? Integrative and Comparitive Biology, 44,
140–151.
Blenckner T, Hillebrand H (2002) North Atlantic Oscillation signatures in aquatic and
terrestrial ecosystems - a meta-analysis. Global Change Biology, 8, 203–212.
Both C, Artemyev AV (2004) Large-scale geographical variation confirms that climate
change causes birds to lay earlier. Proceedings of the Royal Society of London Series B-
Biological Sciences, 271, 1657–1662.
Both C, Bouwhuis S, Lessells CM, Visser ME (2006) Climate change and population
declines in a long-distance migratory bird. Nature, 441, 81–83.
Charmantier A, McCleery RH, Cole LR, Perrins C, Kruuk LEB, Sheldon BC (2008)
Adaptive phenotypic plasticity in response to climate change in a wild bird
population. Science, 320, 800–803.
Cleland EE, Chiariello NR, Loarie SR, Mooney HA, Field CB (2006) Diverse responses
of phenology to global changes in a grassland ecosystem. Proceedings of the National
Academy of Sciences of the United States of America, 103, 13740–13744.
Cotton PA (2003) Avian migration phenology and global climate change. Proceedings of
the National Academy of Sciences of the United States of America, 100, 12219–12222.
Crick HQP, Sparks TH (1999) Climate change related to egg-laying trends. Nature, 399,
423–424.
Cushing DH (1990) Plankton production and year class strength in fish populations –
an update of the match mismatch hypothesis. Advances in Marine Biology, 26,
249–293.
Dawson A (2008) Control of the annual cycle in birds: endocrine constraints and
plasticity in response to ecological variability. Philosophical Transactions of the Royal
Society B-Biological Sciences, 363, 1621–1633.
Edwards M, Richardson AJ (2004) Impact of climate change on marine pelagic
phenology and trophic mismatch. Nature, 430, 881–884.
Elzinga JA, Atlan A, Biere A, Gigord L, Weis AE, Bernasconi G (2007) Time after time:
flowering phenology and biotic interactions. Trends in Ecology and Evolution, 22,
432–439.
Englund G, Sarnelle O, Cooper SD (1999) The importance of data-selection criteria:
meta-analyses of stream predation experiments. Ecology, 80, 1132–1141.
Fitter AH, Fitter RSR (2002) Rapid changes in flowering time in British plants. Science,
296, 1689–1691.
Gillet C, Dubois JP (2007) Effect of water temperature and size of females on the timing
of spawning of perch Perca fluviatilis L. in Lake Geneva from 1984 to 2003. Journal of
Fish Biology, 70, 1001–1014.
Gurevitch J, Curtis PS, Jones MH (2001) Meta-analysis in ecology. Advances in Ecological
Research, 32, 199–247.
Harrington R, Clark SJ, Welham SJ et al. (2007) Environmental change and the
phenology of European aphids. Global Change Biology, 13, 1550–1564.
Harrington R, Woiwod I, Sparks T (1999) Climate change and trophic interactions.
Trends in Ecology and Evolution, 14, 146–150.
IPCC. (2007) Summary for policymakers. In: Climate Change 2007: Impacts, Adaptation
and Vulnerability. Contribution of working group II to the Fourth Assessment Report of the
Intergovernmental Panel on Climate Change (eds Parry ML, Canziani OF, Palutikof JP,
Van Der Linden PJ, Hanson CE), Cambridge University Press, Cambridge.
Langvatn R, Mysterud A, Stenseth NC, Yoccoz NG (2004) Timing and synchrony of
ovulation in red deer constrained by short northern summers. American Naturalist,
163, 763–772.
Menzel A, Sparks TH, Estrella N et al. (2006) European phenological response to
climate change matches the warming pattern. Global Change Biology, 12, 1969–1976.
Millenium Ecosystem Assessment. (2005) Ecosystems and human well-being: synthesis.
Island Press, Washington, DC.
Miller-Rushing AJ, Inouye DW, Primack RB (2008) How well do first flowering dates
measure plant responses to climate change? The effects of population size and
sampling frequency. Journal of Ecology, 96, 1289–1296.
M�ller AP, Rubolini D, Lehikoinen E (2008) Populations of migratory bird species
that did not show a phenological response to climate change are declining.
Proceedings of the National Academy of Sciences of the United States of America, 105,
16195–16200.
Parmesan C (2007) Influences of species, latitudes and methodologies on estimates of
phenological response to global warming. Global Change Biology, 13, 1860–1872.
Parmesan C, Yohe G (2003) A globally coherent fingerprint of climate change impacts
across natural systems. Nature, 421, 37–42.
Penuelas J, Filella I, Comas P (2002) Changed plant and animal life cycles from 1952 to
2000 in the Mediterranean region. Global Change Biology, 8, 531–544.
Platt T, Fuentes-Yaco C, Frank KT (2003) Spring algal bloom and larval fish survival.
Nature, 423, 398–399.
Precht H, Christophersen J, Hensel H, Larcher W (1973) Temperature and life. Springer-
Verlag, Berlin.
Root TL, MacMynowski DP, Mastrandrea MD, Schneider SH (2005) Human-modified
temperatures induce species changes: joint attribution. Proceedings of the National
Academy of Sciences of the United States of America, 102, 7465–7469.
Root TL, Price JT, Hall KR, Schneider SH, Rosenzweig C, Pounds JA (2003) Finger-
prints of global warming on wild animals and plants. Nature, 421, 57–60.
Rosenheim JA, Tabashnik BE (1991) Influence of generation time on the rate of
response to selection. American Naturalist, 137, 527–541.
Roy DB, Sparks TH (2000) Phenology of British butterflies and climate change. Global
Change Biology, 6, 407–416.
Scheffer M, Carpenter SR (2003) Catastrophic regime shifts in ecosystems: linking
theory to observation. Trends in Ecology and Evolution, 18, 648–656.
Sherry RA, Zhou XH, Gu SL et al. (2007) Divergence of reproductive phenology under
climate warming. Proceedings of the National Academy of Sciences of the United States of
America, 104, 198–202.
Smart SM, Clarke RT, Van De Poll HM, Robertson EJ, Shield ER, Bunce RGH, Maskell
LC (2003) National-scale vegetation change across Britain; an analysis of sample-
based surveillance data from the Countryside Surveys of 1990 and 1998. Journal of
Environmental Management, 67, 239–254.
Stefanescu C, Penuelas J, Filella I (2003) Effects of climatic change on the phenology of
butterflies in the northwest Mediterranean Basin. Global Change Biology, 9,
1494–1506.
Thackeray SJ, Jones ID, Maberly SC (2008) Long-term change in the phenology of
spring phytoplankton: species-specific responses to nutrient enrichment and
climatic change. Journal of Ecology, 96, 523–535.
Thompson R, Clark RM (2008) Is spring starting earlier? Holocene, 18, 95–104.
Van Buskirk J, Mulvihill RS, Leberman RC (2009) Variable shifts in spring and autumn
migration phenology in North American songbirds associated with climate change.
Global Change Biology, 15, 760–771.
Visser ME, Both C (2005) Shifts in phenology due to global climate change: the need for
a yardstick. Proceedings of the Royal Society of London Series B-Biological Sciences, 272,
2561–2569.
Winder M, Schindler DE (2004) Climate change uncouples trophic interactions in an
aquatic ecosystem. Ecology, 85, 2100–2106.
3312 S . J . T H A C K E R AY et al.
r 2010 Blackwell Publishing Ltd, Global Change Biology, 16, 3304–3313

Supporting Information
Additional Supporting Information may be found in the online version of this article:
Table S1. Data sources for the meta-analysis of changing phenology. The column ‘data period’ indicates the longest analytical time
period to which each scheme contributed data. Where this is a multi-decadal block, the scheme also contributed data to each of the
constituent decades.
Table S2. Taxa and other phenological indicators included in meta-analyses of changing phenology. The functional categorizations
given to each taxon in the meta-analysis are shown as (Environment, Trophic level, Thermal physiology, Generation time). En-
vironment 5 M, marine; F, freshwater; T, terrestrial. Trophic level 5 PP, Primary producer; PC, Primary consumer; SC, Secondary
consumer. Thermal physiology 5 En, Endotherm; Ec, Ectotherm. Generation time 5 ow, o1 week; w-m, 1 week-1 month; m-y, 1
month-1year; 4y, 41 year. Taxa have been assigned to Orders according to the Catalogue of Life, 2009 annual checklist (http://
www.catalogueoflife.org).
Table S3. Taxa and number of trends per taxon for the analysis of phenological change from 1976–2005, categorized by environment
and major taxonomic group. NA is given where a taxon does not have a common name, or a phenological indicator does not have an
appropriate scientific name.
Please note: Wiley-Blackwell are not responsible for the content or functionality of any supporting materials supplied by the authors.
Any queries (other than missing material) should be directed to the corresponding author for the article.
P H E N O L O G I C A L C H A N G E A C R O S S M A J O R E N V I R O N M E N T S 3313
r 2010 Blackwell Publishing Ltd, Global Change Biology, 16, 3304–3313
Copyright © 2022 FDOKUMEN




















