
Rapid change in early Holocene environments inferred from Lake Pupuke, Auckland City, New Zealand
13
Rapid change in early Holocene environments inferred from Lake Pupuke, Auckland City, New Zealand PAUL AUGUSTINUS, 1 * NERIDA BLEAKLEY, 2 YANBIN DENG, 3 PHIL SHANE 1 and URSULA COCHRAN 4 1 School of Geography, Geology and Environmental Science, University of Auckland, Auckland, New Zealand 2 Department of Primary Industries, Hobart, Australia 3 School of Biological Sciences, University of Auckland, Auckland, New Zealand 4 Geological and Nuclear Sciences, Lower Hutt, New Zealand Augustinus, P., Bleakley, N., Deng, Y., Shane, P., and Cochran, U. 2008. Rapid change in early Holocene environments inferred from Lake Pupuke, Auckland City, New Zealand. J. Quaternary Sci., Vol. 23 pp. 435–447. ISSN 0267-8179. Received 27 February 2007; Revised 19 August 2007; Accepted 5 September 2007 ABSTRACT: Sediment cores from Lake Pupuke in Auckland City, New Zealand, contain a high- resolution millennial to centennial-scale record of changing climate and catchment hydrology spanning the past ca. 10 000 years. Here, we focus on the period between 9500 25 and 7000 155 cal. yr BP during which grain size, diatom palaeoecology, biogenic silica concentrations, sediment elemental and carbon isotope geochemistry reflect changes in sediment sources and lake conditions, with a significant event commencing at ca. 8240 cal. yr BP, commensurate with a lowering of lake level, faster erosion rates and increased sediment influx with a duration of ca. 360 yrs. However, the changes in the lake are not reflected in the terrestrial vegetation, where the pollen record indicates that podocarp forest dominated the Auckland region, with apparent environmental stability during this part of the early Holocene. The synchronous change in most of the proxies between ca. 8240 and 7880 cal. yr BP at Lake Pupuke indicates the presence of a sustained episode of relatively low lake level and concomitant increased rate of erosion in the early Holocene that appears to be at least partly coeval with the 8200 cal. yr BP meltwater event proposed for the North Atlantic region. Copyright # 2008 John Wiley & Sons, Ltd. KEYWORDS: lake sediment; organic matter geochemistry; diatoms; pollen; Lake Pupuke; New Zealand. Introduction High-resolution and well-dated terrestrial records of Lateglacial to early Holocene environments are rare from the southwest Pacific. Such records are important in the context of the present debate over the nature, distribution and timing of sustained climate perturbations such as the Antarctic Cold Reversal, Younger Dryas and 8200 cal. yr BP event (e.g., Rohling and Pa ¨like, 2005; Alloway et al., 2007). In the New Zealand context, the development of high-resolution palaeo-environ- mental records has been hindered by poor chronology, gaps in the records and the often fragmentary nature of the sequences. However, in the Auckland region these limitations are minimised because there are several maar crater lakes that provide continuous and high-resolution palaeo-environmental records that span at least the Lateglacial to early Holocene within which are found numerous rhyolitic tephra that allow the development of a robust chronology. Furthermore, the Auckland region of northern New Zealand (Fig. 1) is important in a palaeo-environmental context because it forms an ecological boundary between the far northern North Island and cooler regions to the south. The southern limit of closed forest during the Last Glacial Maximum lies somewhere in the Auckland region (Sandiford et al., 2003). Prior to human settlement of New Zealand ca. 700 cal. yr BP, the Auckland region was covered by dense podocarp–hardwood forest (McGlone, 1983; McGlone and Wilmshurst, 1999; Horrocks et al., 2005). In addition, it has an unusually dense cluster (about 50) of Quaternary volcanoes (Kermode, 1992), all within Auckland City. Lake sediments are excellent recorders of past climates and geological events via a number of proxy indicators (Battarbee, 2000), and several palaeo-environmental studies of Holocene lake sediment records from central Auckland have been produced (e.g. Newnham and Lowe, 1991; Horrocks et al., 2002, 2005; Augustinus et al., 2006). However, the Holocene JOURNAL OF QUATERNARY SCIENCE (2008) 23(5) 435–447 Copyright ß 2008 John Wiley & Sons, Ltd. Published online 20 February 2008 in Wiley InterScience (www.interscience.wiley.com) DOI: 10.1002/jqs.1153 * Correspondence to: P. Augustinus, School of Geography, Geology and Environ- mental Science, University of Auckland, PB 92019, Auckland, New Zealand. E-mail: [email protected] Contract/grant sponsor: Royal Society of New Zealand Marsden Fund; contract/ grant number: UOA0517. Contract/grant sponsor: University of Auckland Research Committee. Contract/grant sponsor: Australian Institute for Nuclear Science and Engineering; contract/grant number: AINGRA05004.
Transcript of Rapid change in early Holocene environments inferred from Lake Pupuke, Auckland City, New Zealand

Rapid change in early Holocene environmentsinferred from Lake Pupuke, Auckland City,New ZealandPAUL AUGUSTINUS,1* NERIDA BLEAKLEY,2 YANBIN DENG,3 PHIL SHANE1 and URSULA COCHRAN4
1 School of Geography, Geology and Environmental Science, University of Auckland, Auckland, New Zealand2 Department of Primary Industries, Hobart, Australia3 School of Biological Sciences, University of Auckland, Auckland, New Zealand4 Geological and Nuclear Sciences, Lower Hutt, New Zealand
Augustinus, P., Bleakley, N., Deng, Y., Shane, P., and Cochran, U. 2008. Rapid change in early Holocene environments inferred from Lake Pupuke, Auckland City,New Zealand. J. Quaternary Sci., Vol. 23 pp. 435–447. ISSN 0267-8179.
Received 27 February 2007; Revised 19 August 2007; Accepted 5 September 2007
ABSTRACT: Sediment cores from Lake Pupuke in Auckland City, New Zealand, contain a high-resolution millennial to centennial-scale record of changing climate and catchment hydrologyspanning the past ca. 10 000 years. Here, we focus on the period between 9500� 25 and7000� 155 cal. yr BP during which grain size, diatom palaeoecology, biogenic silica concentrations,sediment elemental and carbon isotope geochemistry reflect changes in sediment sources and lakeconditions, with a significant event commencing at ca. 8240 cal. yr BP, commensurate with a loweringof lake level, faster erosion rates and increased sediment influxwith a duration of ca. 360 yrs. However,the changes in the lake are not reflected in the terrestrial vegetation, where the pollen record indicatesthat podocarp forest dominated the Auckland region, with apparent environmental stability during thispart of the early Holocene. The synchronous change in most of the proxies between ca. 8240 and7880 cal. yr BP at Lake Pupuke indicates the presence of a sustained episode of relatively low lake leveland concomitant increased rate of erosion in the early Holocene that appears to be at least partlycoeval with the 8200 cal. yr BP meltwater event proposed for the North Atlantic region. Copyright #2008 John Wiley & Sons, Ltd.
KEYWORDS: lake sediment; organic matter geochemistry; diatoms; pollen; Lake Pupuke; New Zealand.
Introduction
High-resolution andwell-dated terrestrial records of Lateglacialto early Holocene environments are rare from the southwestPacific. Such records are important in the context of the presentdebate over the nature, distribution and timing of sustainedclimate perturbations such as the Antarctic Cold Reversal,Younger Dryas and 8200 cal. yr BP event (e.g., Rohling andPalike, 2005; Alloway et al., 2007). In the New Zealandcontext, the development of high-resolution palaeo-environ-mental records has been hindered by poor chronology, gaps inthe records and the often fragmentary nature of the sequences.However, in the Auckland region these limitations are
minimised because there are several maar crater lakes thatprovide continuous and high-resolution palaeo-environmentalrecords that span at least the Lateglacial to early Holocenewithin which are found numerous rhyolitic tephra that allowthe development of a robust chronology.Furthermore, the Auckland region of northern New Zealand
(Fig. 1) is important in a palaeo-environmental context becauseit forms an ecological boundary between the far northern NorthIsland and cooler regions to the south. The southern limit ofclosed forest during the Last Glacial Maximum lies somewherein the Auckland region (Sandiford et al., 2003). Prior to humansettlement of New Zealand ca. 700 cal. yr BP, the Aucklandregion was covered by dense podocarp–hardwood forest(McGlone, 1983; McGlone and Wilmshurst, 1999; Horrockset al., 2005). In addition, it has an unusually dense cluster(about 50) of Quaternary volcanoes (Kermode, 1992), all withinAuckland City.Lake sediments are excellent recorders of past climates and
geological events via a number of proxy indicators (Battarbee,2000), and several palaeo-environmental studies of Holocenelake sediment records from central Auckland have beenproduced (e.g. Newnham and Lowe, 1991; Horrocks et al.,2002, 2005; Augustinus et al., 2006). However, the Holocene
JOURNAL OF QUATERNARY SCIENCE (2008) 23(5) 435–447Copyright � 2008 John Wiley & Sons, Ltd.Published online 20 February 2008 in Wiley InterScience(www.interscience.wiley.com) DOI: 10.1002/jqs.1153
*Correspondence to: P. Augustinus, School of Geography, Geology and Environ-mental Science, University of Auckland, PB 92019, Auckland, New Zealand.E-mail: [email protected]
Contract/grant sponsor: Royal Society of New Zealand Marsden Fund; contract/grant number: UOA0517.Contract/grant sponsor: University of Auckland Research Committee.Contract/grant sponsor: Australian Institute for Nuclear Science and Engineering;contract/grant number: AINGRA05004.

records in these cores are often of low resolution and disturbed,affecting their stratigraphic continuity and reliability of the agecontrol (e.g. Horrocks et al., 2002;Woodward and Shulmeister,2005). Furthermore, none of the Holocene lake sediment seq-uences from the Auckland region published to date provides ahigh-resolution multi-proxy record of changing environmentsin the Auckland region as is required to enable reliableidentification of centennial to millennial-scale palaeoclimaticevents.We present biological (diatoms and pollen), sedimentologi-
cal, and isotopic and elemental geochemical proxies ofenvironmental change from Lake Pupuke in the centralAuckland region for the period 9500� 25 to 7000� 155 cal.yr BP. This dataset provides the only complete, laminated,high-resolution multi-proxy environmental record of the earlyHolocene from New Zealand, and is of note given the currentcontroversy over the timing, forcing mechanism and possibleglobal extent of the 8200 cal. yr BP event (e.g. Alley et al., 1997;Alley and Agustsdottir, 2005; Rohling and Palike, 2005;Kobashi et al., 2007; Thomas et al., 2007). The evidence forthe 8200 cal. yr BP event is well represented in lake sedimentrecords from the North Atlantic region (e.g. Nesje and Dahl,2001; Hammarlund et al., 2003; Veski et al., 2004) and hasrecently been shown to have inter-hemispheric significance(Ljung et al., 2007).
The study site
Lake Pupuke (368 47.250 S, 1758 46.250 E) occupies a ca.260 000-year-old basaltic explosion crater (maar) (Hall and
York, 1984) on the North Shore of Auckland City (Fig. 1). Thearea, including the lake catchment, is mainly residential, withthe eastern margin of the lake lying ca. 200m from the sea andca. 5m above sea level. It is surrounded by a tuff cone, whichhas protected the freshwater lake from erosion and seawaterinflux. The tuff ring of Pupuke crater consists of a mixture ofjuvenile and accidental ejecta deposited by base surgesassociated with phreatomagmatic explosions. Part of theseaward rim is also supported by lava flows. The lake isclosed hydrologically and occupies 57% of its 1.9 km2
catchment. Mean annual rainfall is 1119mm (Tomlinson andSansom, 1994a), and is distributed evenly throughout the year.Mean daily air temperature is 15.68C and the average dailyrange is 6.08C (Tomlinson and Sansom, 1994b).
The limnology of Lake Pupuke has been described by Barker(1970), Cassie (1989) and Holmes (1994). Rainfall into thecatchment accounts for the bulk of the water entering the lakeand there are no significant inflows or outflows, although anadjacent coastal spring originates from the lake. The surfacewaters have a pH of ca. 8.36, suggesting low levels ofdissolved CO2 and a predominance of bicarbonate. The surfacewaters are separated from the anoxic hypolimnion by awell-defined thermocline, with the depth to its lower boundaryranging from 21 to 25m (Holmes, 1994).
Pollen records from Lake Pupuke indicate that, apart from anexpansion of Agathis australis suggesting increasingly dryconditions from ca. 7000 cal. yr BP (Newnham and Lowe,1991; Horrocks et al., 2005), the pre-human Holocene was aperiod of relative environmental stability (Horrocks et al., 2005;Augustinus et al., 2006). Dramatic vegetation change onlyoccurred over the last ca. 700 cal. yr as a result of large-scalePolynesian deforestation by fire, and appearance of pollen ofintroduced Pinus and Plantago lanceolata associated withEuropean settlement of Auckland in the early 1840s (Augus-tinus et al., 2006; Hayward et al., 2006).
Methods
A 5.7m long core (core P5) was obtained from Lake Pupuke(Fig. 1) using a Mackereth corer (Mackereth, 1966; Neale andWalker, 1996) with 65mm diameter PVC tubes. The sedimentsretrieved were subjected to analysis for a range of proxies(described below) at intervals ranging from 1 to 2 cm dependingon the proxy used.
Grain size analysis was undertaken at 1 cm intervals. Thesamples were treated with H2O2 to remove organic matter,diatoms dissolved in 2M Na2CO3, dispersed with sodiumhexametaphosphate and sonified before measurement with aMalvern Mastersizer 2000 laser particle sizer. Grain size isgiven as mean size and percentage greater than 32mm. Thelatter grain size component is used as an indicator of clasticsediment influx into the lake.
Loss-on-ignition (LOI) analyses were carried out on oven-dried samples combusted at 5508C for 24 h using the procedureof Gale and Hoare (1991). Total organic carbon (TOC), totalnitrogen (TN) and d13C analysis of bulk organic matter firstinvolved acidification with 1N HCl to remove any carbonate.Samples were then washed three times with deionised water,freeze-dried and crushed to a powder. The TOC and TNanalyses were undertaken at the Isotope Laboratory at theUniversity ofWaikato, Hamilton, NewZealand, using aDumaselemental analyser (Europa Scientific ANCA-SL) interfaced toan isotope mass spectrometer (Europa Scientific 20-20 stableisotope analyser) for d13C measurement.
Figure 1 Location map showing Lake Pupuke bathymetry (inset).Shading indicates elevated regions, with darker shading indicatingincreasing elevation
Copyright � 2008 John Wiley & Sons, Ltd. J. Quaternary Sci., Vol. 23(5) 435–447 (2008)DOI: 10.1002/jqs
436 JOURNAL OF QUATERNARY SCIENCE

Biogenic silica (BGS) estimation was undertaken every 2 cmusing the method of Mortlock and Froelich (1989), with Siconcentration measurement by ICP-OES at GNS Science,Wairakei, New Zealand.Accelerator mass spectrometry (AMS) 14C age determi-
nations were carried out on bulk sediment samples from 520,530, 540 and 550 cm depths in the core on the tandemaccelerator at the Australian Nuclear Science and TechnologyOrganisation, Sydney, Australia. The samples received stan-dard acid–alkali–acid pretreatments, with radiocarbon agecalibration undertaken using CALIB 5.0.2 using the INTCAL04dataset (Reimer et al., 2004). Radiocarbon years (before AD1950) are reported as 14C yr BP. Calibrated ages are reportedas cal. yr BP and expressed with 1 sigma errors.Macroscopic tephra layers were sampled for geochemical
characterisation and correlation studies. The glass shards wereanalysed using a Jeol JXA-840 probe fitted with a PGT Prism2000 EDS detector at the University of Auckland. Absorbedcurrent of 1.5 nA at 15 kV and a beam defocused to 20mmwasused. The analyses were normalised to 100% to account forvariablemeteoric hydration and to facilitate correlation to othersites. New Zealand Quaternary tephrochronology is furtherdescribed in Shane (2000) and Shane and Hoverd (2002).Samples for pollen analysis were selected at 2 cm intervals
except between 590 and 560 cm and between 515 and 507 cmdepths. The samples were prepared by the standard acetylationand hydrofluoric acid method and bleached (Moore et al.,1991). The pollen sum was at least 250 grains, excludingLeptospermum type, herbaceous swamp plants and ferns. Apercentage pollen diagram was prepared using the TILIAprogramme (E. Grimm, Illinois State Museum), with zonationundertaken using CONISS.Diatom analysis was carried out on subsamples of
approximately 1 cm3 taken at 2 cm intervals from 510 cm to570 cm depth in Pupuke core P5. Diatom samples wereprepared for analysis by oxidation with dilute hydrogenperoxide (10%) in a water bath at 708C for 2 h, or untilobvious organic residue had been removed. Samples were thenrinsed by repeated washing with distilled water. Subsamples ofprocessed sediment were mounted using Naphrax1.At least 300 and at times up to 900 diatom valves were
counted from each sample using an Olympus compoundmicroscope at�1000magnification (oil immersion) with phasecontrast. Taxonomy was based on Foged (1979), Krammer andLange-Bertalot (1986, 1988, 1991a,b, 2000), Round et al.(1990) and Sonneman et al. (1999). Diatom species abun-dances were expressed as percentages, calculated using totalnumbers of valves recorded for each sample. Species with lessthan 1% abundance in any sample, and/or occurring in fewerthan three samples, were eliminated from the analyses. Specieswere grouped together according to their habitat preferences asderived from Round et al. (1990) and Van Dam et al. (1994).The percentage diatom diagram was prepared using the TILIAprogramme (E. Grimm, Illinois State Museum), with zonationundertaken using CONISS.
Results
Stratigraphy
Core P5 comprises highly organic muds laminated atsub-millimetre scale (Fig. 2), with the section of the corebetween the Tuhua and Rotoma tephras containing sedimentnoticeably darker and denser than the sediment above this
depth (Horrocks et al., 2005). Tephra-filled cracks in the lami-nated muds were encountered immediately below the Rotomatephra and are infilled with the latter (Fig. 2). These features areclearly depositional and not an artefact of the coring process asthey appear at the same location in all of the cores obtainedfrom Lake Pupuke. Furthermore, the sediments into which thefractures penetrate are plane-laminated and otherwise undis-turbed.Mean grain sizes in the section of core P5 between 574 and
507 cm depth are variable but display a trend to fining upwards,with distinct mean grain size maxima of 12.1mm at 535 cm(Fig. 3). The distinct peak in mean grain size commences at537 cm and terminates at 530 cm depth, abovewhich grain sizeis significantly finer and uniform with a range of only6.6–7.5mm (Fig. 3). The percentage of grains greater than32mm displays a similar trend to that of mean grain size, withvariable but high percentages of coarser than medium silt(34.7% to 53.1%) from 570 to 537 cm, above which the peak ofcoarser clastic sediment occurs between 537 and 529 cmdepth. Above 529 cm depth a marked reduction in percentage(27.4% to 10% >32mm) of the coarser clastic sedimentsoccurs, mirroring the mean grain size trend (Fig. 3).
Dating
Tephrochronology
Two macroscopic tephra beds were observed in core P5 in thesection between 574 and 505 cm depth. A 2.5 cm thick,normal-graded, fine to medium ash occurs at 574 cm depth(Fig. 2). This tephra bed contains the ferromagnesian mineralcummingtonite and minor hornblende. Only three tephra bedscontain cummingtonite in high abundance: ca. 50 000 yrRotoehu tephra, ca. 9500 cal. yr BP Rotoma tephra, and5500 cal. yr BP Whakatane tephra, all from Okataina VolcanicCentre (e.g., Shane, 2000). The glass chemistry of this tephrabed (Table 1; Horrocks et al., 2005) supports correlation withRotoma tephra. Rotoma tephra is dated at 9500� 25 cal. yr BPby wiggle-match on a peat sequence against IntCal04 viaOxCal (Lowe et al., 2008). At 507 to 505 cm depth, a greycoarse ash bed occurs. The composition of the glass isperalkaline rhyolite (Table 1; Horrocks et al., 2005). TuhuaVolcanic Centre (Mayor Island) is the only peralkaline volcanothat has been active during the Holocene (Shane, 2000). TheTuhua tephra from this volcano has been previously identifiedat Auckland sites (Newnham and Lowe, 1991; Sandiford et al.,2001) and is the only known widespread Holocene tephrafrom Tuhua volcano (e.g. Lowe et al., 2008). It is dated at7000� 155 cal. yr BP by wiggle-match on a peat sequenceagainst IntCal04 via OxCal (Lowe et al., 2008).
Radiocarbon ages
Four horizons between the Tuhua and Rotoma tephra weredated using AMS 14C (Table 1). Bulk carbon extracted fromthe sample from 520 cm depth (OZH877) provided an age of6540� 80 14C yr BP, while the samples collected at 530, 540and 550 cm depth provided ages in stratigraphic order rangingfrom 7900� 60 to 8470� 60 14C yr BP, but with a significantage offset from the expected calibrated age assuming near-constant sedimentation rates between the Tuhua and Rotomatephra (Fig. 4). This constant sedimentation rate assumption isreasonable given the well-laminated nature of the sediments
Copyright � 2008 John Wiley & Sons, Ltd. J. Quaternary Sci., Vol. 23(5) 435–447 (2008)DOI: 10.1002/jqs
RAPID CHANGE IN EARLY HOLOCENE ENVIRONMENTS 437

and apparent constant lamina thickness (Fig. 2). Furthermore,there is a striking difference between the d13C of the bulkcarbon in samples from 520 cm depth compared with thosefrom 530–550 cm depths (Table 1), with d13C enrichmentapparent in the lower three samples.On the assumption of a constant sedimentation rate between
the 9500� 25 cal. yr BP Rotoma tephra and the 7000� 155 cal.yr BP Tuhua tephra, there is an apparent age offset of the AMS14C ages from the constant sedimentation rate line ranging fromca. 790 yr at 530 cm depth to ca. 700 yr at 550 cm depth in coreP5 (Table 1, Fig. 4). Interestingly, the 520 cm depth sampledisplays significant d13C depletion and plots close to theconstant sedimentation rate line.
Organic matter elemental andisotopic geochemistry
LOI and TOC
The organic matter elemental and isotopic geochemistry variesconsiderably throughout the 574 to 507 cm depth section ofcore P5 (Fig. 3). LOI values increase gradually to peaks of 48%at 556 cm and 47% at 534 cm depth, with aminimum of 30% at538 cm depth. A clear LOI peak of 47% occurs at 534 cm, afterwhich the values reduce to a constant level of between 26%
and 31%up to the level of the Tuhua tephra (Fig. 3). This patternis mirrored by TOC, although the peak in TOC values between538 and 530 cm depth is more distinct and the values above530 cm are much lower and vary between 11% and 14%.
TOC/TN ratios and d13C
The TOC/TN ratio displays a similar pattern to that of TOCand LOI. TOC/TN values are variable and mostly rangebetween 20 and 26 from 570 to 538 cm depth, above which theratio increases rapidly to a peak of 32 at 534 cm depth (Fig. 3).Above 530 cm the values are significantly lower and vary onlyslightly between 13 and 16.
d13C resembles the trend in TOC/TN, TOC and LOI (Fig. 3).From 574 to 562 cm there is a pattern of d13C enrichment,above which the values oscillate about a mean of ca. �21%up to 538 cm depth. As is the case with the other organicmatter-derived proxies, a marked enrichment phase occursin d13C between 538 and 530 cm depth, with a maximumenrichment of �20% attained at 534 cm depth. Above thisdepth a marked depletion occurs until a minimum of �26% isreached at 530 cm. d13C values for the remainder of the core aredepleted and vary only slightly about a mean of ca. �25.5%(Fig. 3).
1.5
1.0
0.5
0.0
4.0
3.5
3.0
2.5
5.5
5.0
4.5
2.0
m
Laminated muds
Tephra-filled tension cracks
Tuhua tephra(7000 ± 150 cal. yr BP)
Rotoma tephra(9500 ± 25 cal. yr BP)
Rangitoto tephra (ca. 550 cal. yr BP)
Taupo tephra (ca. 1720 cal. yr BP)C
lay
Silt Fine sa
nd
Med sand
34.5 cm (3696-3580 cal yr BP)95 cm (1886-1735 & 2141-2005 cal yr BP)
325 cm (5283-5155 cal yr BP)
520 cm (7562-7335 cal. yr BP)
530 cm (8949-8600 cal. yr BP)
540 cm (8995-8775 cal. yr BP)
550 cm (9532-9460 cal. yr BP)
Lake Pupuke core P5
Figure 2 Stratigraphic log of Lake Pupuke core P5 with photographs of the laminated sediment between the Rotoma and Tuhua tephra
Copyright � 2008 John Wiley & Sons, Ltd. J. Quaternary Sci., Vol. 23(5) 435–447 (2008)DOI: 10.1002/jqs
438 JOURNAL OF QUATERNARY SCIENCE
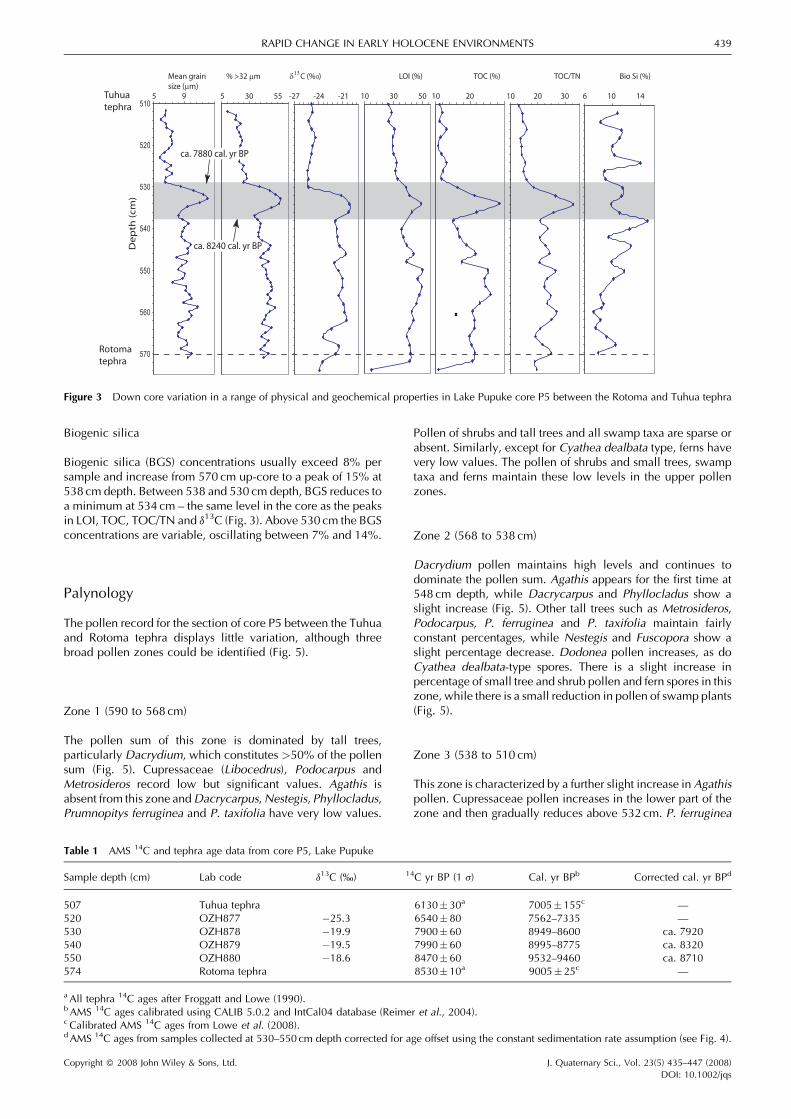
Biogenic silica
Biogenic silica (BGS) concentrations usually exceed 8% persample and increase from 570 cm up-core to a peak of 15% at538 cm depth. Between 538 and 530 cm depth, BGS reduces toa minimum at 534 cm – the same level in the core as the peaksin LOI, TOC, TOC/TN and d13C (Fig. 3). Above 530 cm the BGSconcentrations are variable, oscillating between 7% and 14%.
Palynology
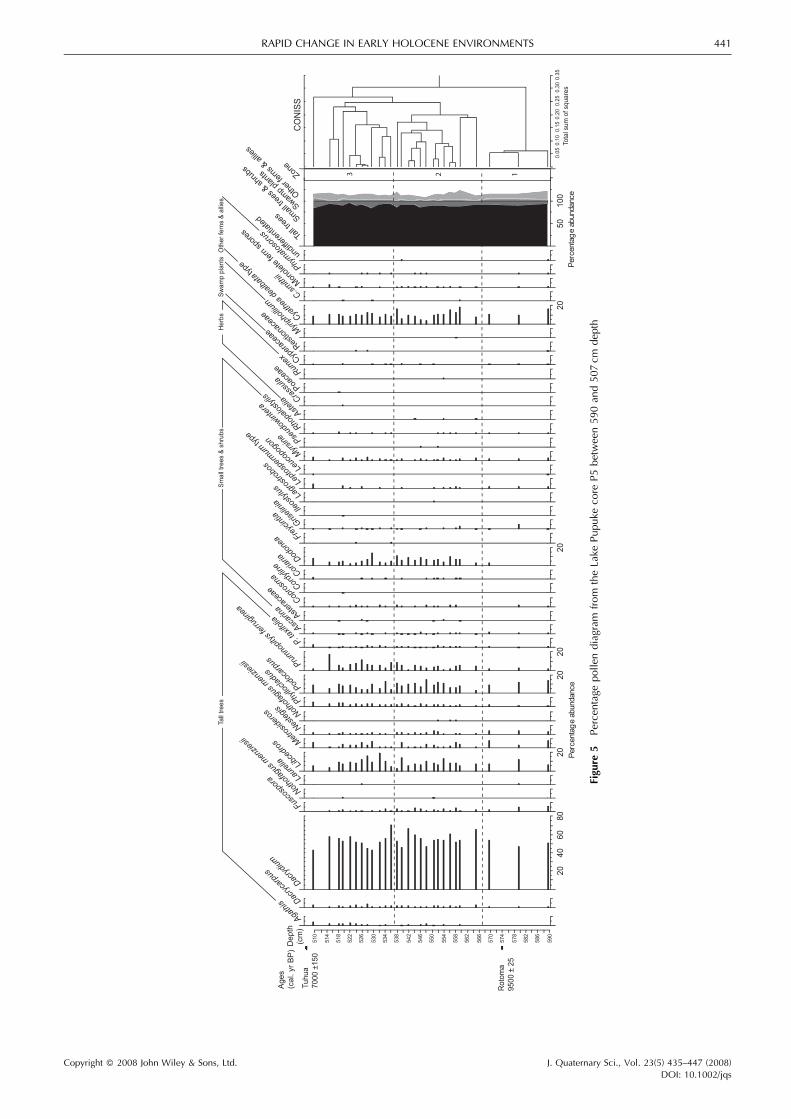
The pollen record for the section of core P5 between the Tuhuaand Rotoma tephra displays little variation, although threebroad pollen zones could be identified (Fig. 5).
Zone 1 (590 to 568 cm)
The pollen sum of this zone is dominated by tall trees,particularly Dacrydium, which constitutes >50% of the pollensum (Fig. 5). Cupressaceae (Libocedrus), Podocarpus andMetrosideros record low but significant values. Agathis isabsent from this zone andDacrycarpus,Nestegis, Phyllocladus,Prumnopitys ferruginea and P. taxifolia have very low values.
Pollen of shrubs and tall trees and all swamp taxa are sparse orabsent. Similarly, except for Cyathea dealbata type, ferns havevery low values. The pollen of shrubs and small trees, swamptaxa and ferns maintain these low levels in the upper pollenzones.
Zone 2 (568 to 538 cm)
Dacrydium pollen maintains high levels and continues todominate the pollen sum. Agathis appears for the first time at548 cm depth, while Dacrycarpus and Phyllocladus show aslight increase (Fig. 5). Other tall trees such as Metrosideros,Podocarpus, P. ferruginea and P. taxifolia maintain fairlyconstant percentages, while Nestegis and Fuscopora show aslight percentage decrease. Dodonea pollen increases, as doCyathea dealbata-type spores. There is a slight increase inpercentage of small tree and shrub pollen and fern spores in thiszone, while there is a small reduction in pollen of swamp plants(Fig. 5).
Zone 3 (538 to 510 cm)
This zone is characterized by a further slight increase in Agathispollen. Cupressaceae pollen increases in the lower part of thezone and then gradually reduces above 532 cm. P. ferruginea
De
pth
(cm
)
510
520
530
540
550
560
570
Mean grain % >32 µm δ C (% ) LOI (%) TOC (%) TOC/TN Bio Si (%) size (µm)
5 9 5 30 55 -27 -24 -21 10 30 50 10 20 10 20 30 6 10 14
130
ca. 8240 cal. yr BP
ca. 7880 cal. yr BP
Rotomatephra
Tuhuatephra
Figure 3 Down core variation in a range of physical and geochemical properties in Lake Pupuke core P5 between the Rotoma and Tuhua tephra
Table 1 AMS 14C and tephra age data from core P5, Lake Pupuke
Sample depth (cm) Lab code d13C (%) 14C yr BP (1 s) Cal. yr BPb Corrected cal. yr BPd
507 Tuhua tephra 6130� 30a 7005� 155c —520 OZH877 �25.3 6540� 80 7562–7335 —530 OZH878 �19.9 7900� 60 8949–8600 ca. 7920540 OZH879 �19.5 7990� 60 8995–8775 ca. 8320550 OZH880 �18.6 8470� 60 9532–9460 ca. 8710574 Rotoma tephra 8530� 10a 9005�25c —
aAll tephra 14C ages after Froggatt and Lowe (1990).b AMS 14C ages calibrated using CALIB 5.0.2 and IntCal04 database (Reimer et al., 2004).c Calibrated AMS 14C ages from Lowe et al. (2008).d AMS 14C ages from samples collected at 530–550cm depth corrected for age offset using the constant sedimentation rate assumption (see Fig. 4).
Copyright � 2008 John Wiley & Sons, Ltd. J. Quaternary Sci., Vol. 23(5) 435–447 (2008)DOI: 10.1002/jqs
RAPID CHANGE IN EARLY HOLOCENE ENVIRONMENTS 439
levels are slightly higher than in the lower pollen zones, whileall other tall tree pollen maintains similar percentages to thosein Zone 2. Small tree and shrub pollen also maintains similarlevels to that observed in Zone 2, while swamp taxa continue tobe sparse (Fig. 5).
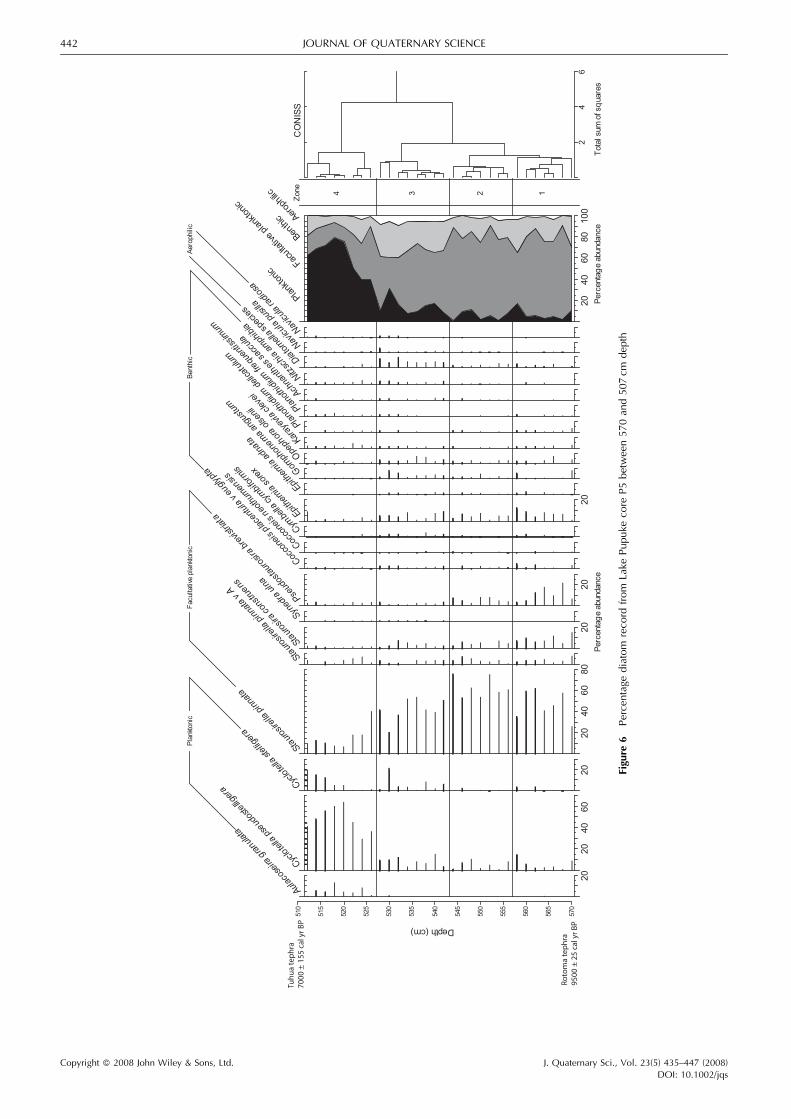
Diatom record
A total of 86 diatom taxa were identified in core P5. Theproportion of the total number of diatoms at each depth isshown in Fig. 6 for the 23most abundant species. The dominantpattern seen in core P5 is the high proportion of smallfacultative planktonic species of the family Fragilariaceae(dominated by Staurosirella pinnata) at the base of the section,and the abundance of planktonic species Cyclotella pseudos-telligera, Cyclotella stelligera and Aulacoseira granulata at thetop of the section from 526 to 512 cm depth (Fig. 6). Benthicand aerophilic taxa occur in small proportions throughout thesection, with a peak in abundance from 543 to 527 cm depth.The diatom record for the section of core P5 between theRotoma and Tuhua tephra displays four distinct zonesdetermined by cluster analysis (Fig. 6).
Zones 1 and 2 (570 to 543 cm)
These zones contain high proportions of the facultativeplanktonic diatoms Staurosirella pinnata, Staurosira construensand Pseudostaurosira brevistriata. Staurosirella pinnata isdominant and in Zone 2 it reaches its greatest abundance
of 45–75% of the total assemblage. There are moderateproportions of benthic species such as Epithemia sorex andCocconeis placentula throughout this section of core P5.Planktonic species (mainly Cyclotella pseudostelligera) gener-ally form less than 10% of the assemblage and aerophilicspecies only occur in trace amounts (Fig. 6).
Zone 3 (543 to 527 cm)
This zone is also dominated by Staurosirella pinnata and theother facultative planktonic species present in the previouszones. However, this zone is differentiated from Zones 1 and 2by higher proportions of planktonic, benthic and aerophilicspecies. The increase in aerophilic species such as Navicellapusilla,Navicula radiosa and species of the genusDiatomella isparticularly notable because such species usually only occur intrace amounts in lake sediments, if at all (Fig. 6).
Zone 4 (527 to 510 cm)
This zone contains a distinctly different diatom flora, and isdistinguished by the dominance of the planktonic speciesCyclotella pseudostelligera and Aulacoseira granulata (Fig. 6).Cyclotella stelligera is also common in the top five samples.Facultative planktonic species are significantly lower inabundance than in the rest of the core section and aerophilicspecies have all but disappeared.
Discussion
Dating
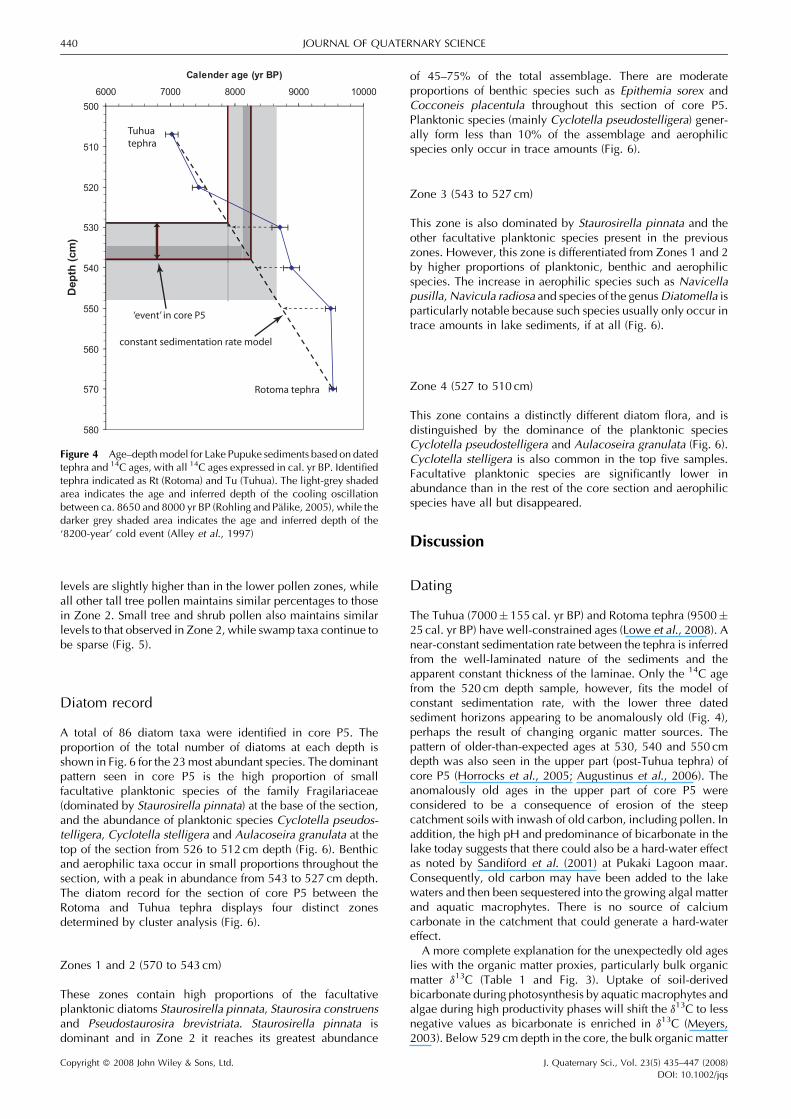
The Tuhua (7000� 155 cal. yr BP) and Rotoma tephra (9500�25 cal. yr BP) have well-constrained ages (Lowe et al., 2008). Anear-constant sedimentation rate between the tephra is inferredfrom the well-laminated nature of the sediments and theapparent constant thickness of the laminae. Only the 14C agefrom the 520 cm depth sample, however, fits the model ofconstant sedimentation rate, with the lower three datedsediment horizons appearing to be anomalously old (Fig. 4),perhaps the result of changing organic matter sources. Thepattern of older-than-expected ages at 530, 540 and 550 cmdepth was also seen in the upper part (post-Tuhua tephra) ofcore P5 (Horrocks et al., 2005; Augustinus et al., 2006). Theanomalously old ages in the upper part of core P5 wereconsidered to be a consequence of erosion of the steepcatchment soils with inwash of old carbon, including pollen. Inaddition, the high pH and predominance of bicarbonate in thelake today suggests that there could also be a hard-water effectas noted by Sandiford et al. (2001) at Pukaki Lagoon maar.Consequently, old carbon may have been added to the lakewaters and then been sequestered into the growing algal matterand aquatic macrophytes. There is no source of calciumcarbonate in the catchment that could generate a hard-watereffect.
A more complete explanation for the unexpectedly old ageslies with the organic matter proxies, particularly bulk organicmatter d13C (Table 1 and Fig. 3). Uptake of soil-derivedbicarbonate during photosynthesis by aquatic macrophytes andalgae during high productivity phases will shift the d13C to lessnegative values as bicarbonate is enriched in d13C (Meyers,2003). Below 529 cm depth in the core, the bulk organic matter
500
510
520
530
540
550
560
570
580
6000 7000 8000 9000 10000
Calender age (yr BP)D
ep
th (
cm
)
Tuhuatephra
constant sedimentation rate model
‘event’ in core P5
Rotoma tephra
Figure 4 Age–depthmodel for Lake Pupuke sediments based on datedtephra and 14C ages, with all 14C ages expressed in cal. yr BP. Identifiedtephra indicated as Rt (Rotoma) and Tu (Tuhua). The light-grey shadedarea indicates the age and inferred depth of the cooling oscillationbetween ca. 8650 and 8000 yr BP (Rohling and Palike, 2005), while thedarker grey shaded area indicates the age and inferred depth of the‘8200-year’ cold event (Alley et al., 1997)
Copyright � 2008 John Wiley & Sons, Ltd. J. Quaternary Sci., Vol. 23(5) 435–447 (2008)DOI: 10.1002/jqs
440 JOURNAL OF QUATERNARY SCIENCE
510
514
518
522
526
530
534
538
542
546
550
554
558
562
566
570
574
578
582
586
590
Depth
(cm
)T
uh
ua
70
00
±1
50
Ro
tom
a
95
00
± 2
5
Agathi
s Dacryc
arpus 2
040
60
80
Dacryd
ium
Fusco
spora
Nothofa
gus m
enzie
sii
Laure
lia
20
Libce
dros Metr
oside
ros
Nesteg
is Nothofa
gus m
enzie
sii
Phyllo
cladu
s
20
Podoc
arpus
20
Prumno
pitys
ferru
ginea
P. tax
ifolia
Ascari
na Aster
acea
e
Copros
maCord
yline
Coriari
a 20
Dodon
eaFrey
cintia Gris
elinia Ile
ostyl
us Lagro
strob
os
Lepto
sperm
um ty
pe
Leuc
opog
onMyrs
ine Pseud
owint
era
Rhopa
lostyl
is
Astelia Crassu
laPoa
ceae Rum
ex Cyp
erac
eae
Res
tiona
ceae
Myriph
ollium 2
0
Cyathe
a dea
lbata
type
C.smith
iiM
onol
ete
fern
spo
res
Phymato
sorus
undi
ffere
ntia
ted
50
100
Tall
trees Sm
all t
rees
& s
hrub
s
Swam
p pl
ants
Oth
er fe
rns
& allie
s
Sw
am
p p
lan
tsO
the
r fe
rns &
alli
es
He
rbs
Sm
all
tre
es &
sh
rub
sTa
ll tr
ee
s
0.0
50.1
00.1
50.2
00.2
50.3
00.3
5
To
tal su
m o
f sq
ua
res
CO
NIS
S
Zone
3 2 1
Ages
(cal. y
r B
P)
Perc
enta
ge a
bundance
Perc
enta
ge a
bundance
Figu
re5
Perce
ntage
pollen
diagram
from
theLa
kePupuke
core
P5betwee
n590an
d507cm
dep
th
Copyright � 2008 John Wiley & Sons, Ltd. J. Quaternary Sci., Vol. 23(5) 435–447 (2008)DOI: 10.1002/jqs
RAPID CHANGE IN EARLY HOLOCENE ENVIRONMENTS 441
510
515
520
525
530
535
540
545
550
555
560
565
570
Depth (cm)
20
Aulaco
seira
granula
ta
20
40
60
Cyclotel
la ps
eudos
telligera
20
Cyclotel
la ste
lligera 2
040
60
80
Staurosir
ella p
innata
Staurosir
ella p
innata
v A
20
Staurosir
a con
strue
ns
Syned
ra uln
a20
Pseud
ostau
rosira
brev
istria
ta
Cocco
neis
placen
tula v
eugly
pta
Cocco
neis
neothu
mensis
Cymbe
lla cy
mbiform
is
20
Epithe
mia so
rex
Epithe
mia ad
nata
Gomph
onema a
ngus
tum
Navicu
la rad
iosa
Opeph
ora ol
senii
Navicu
la pu
silla
Karaye
via cl
evei
Planoth
idium
delica
tulum
Planoth
idium
frequ
entis
simum
Achna
nthes
sacc
ula
Nitzsc
hia am
phibi
a
Diatomell
a spe
cies
20
40
60
80
100
Plank
toni
cFac
ultativ
e pl
ankt
onic
Benth
ic Aerop
hilic
Zone
4 3 12
Aero
philic
Facultative
pla
nkt
onic
Pla
nkt
onic
Perc
enta
ge a
bundance
24
6
Tota
l sum
of sq
uare
s
CO
NIS
S
Roto
ma
tep
hra
9500
± 2
5 ca
l yr B
P
Tuh
ua
tep
hra
7000
± 1
55 c
al y
r BP
Perc
enta
ge a
bundance
Benth
ic
Figu
re6
Percentage
diatom
reco
rdfrom
Lake
Pupuke
core
P5betwee
n570an
d507cm
dep
th
Copyright � 2008 John Wiley & Sons, Ltd. J. Quaternary Sci., Vol. 23(5) 435–447 (2008)DOI: 10.1002/jqs
442 JOURNAL OF QUATERNARY SCIENCE

displays d13C enrichment, which is consistent with high levelsof in situ lacustrine productivity and photosynthetic CO2
uptake in excess of available dissolved CO2. Consequently, thealgae and aquatic macrophytes are forced to use bicarbonatedissolved in the lake water as their CO2 source, with thebicarbonate probably sourced from the soils on the maar cratertuff ring surrounding the crater lake. This model is supported bythe influx of high levels of silt size and coarser (>32mm) clasticsand high TOC/TN ratios indicative of terrigenous organicmatter influx. Hence it appears that we are seeing a systematicoffset in ages introduced by the addition of soil-derivedbicarbonate to the lake. The ‘corrected’ calibrated ages for thesediments from 530 to 550 cm depth produced by shifting theircalibrated ages to the constant sedimentation rate line (Fig. 5)range from ca. 8670 to 7950 cal. yr BP (Table 1). Although thisage correction must be used with caution, it provides ages forthe apparent climatic perturbation between 538 and 529 cmdepth of ca. 8260 to 7900 cal. yr BP, which is coeval with theperiod of cooling lasting several hundred years recognisedacross the North Atlantic region between 8650 and 8000 cal. yrBP (Rohling and Palike, 2005), and the pronounced 8250 to8150 cal. yr BP ‘8200-year event’ (Alley et al., 1997; Alley andAgustsdottir, 2005).
Organic matter geochemistry
The proportion of sedimentary organic matter derived fromterrestrial plants and that derived from within a lake can beidentified from the difference between TOC/TN ratios ofvascular land plants and algae (Meyers, 1994, 2003). Increasedinflux of land-plant derived organic matter is recorded asincreased TOC/TN ratios, because vascular land plants haveTOC/TN ratios >20 whereas algae typically have TOC/TNratios between 4 and 10. Values between 10 and 20 usuallyrelate to a mixed terrestrial–aquatic source for the organicmatter. This difference is likely to be a consequence of theabsence of cellulose in algae and its abundance in vascularplants (Meyers, 1994; Meyers and Lallier-Verges, 1999).Consequently, increased dominance of autochthonous algal-derived organic matter would result in reduced TOC/TN ratios,while elevated TOC/TN ratios would indicate an increasedproportion of allochthonous terrestrial organic matter.The TOC/TN ratios of >20 between 570 and 538 cm depth
in core P5 are indicative of the flux of organic matterpredominantly from a terrestrial source (Fig. 3). The markedspike in terrestrial organic matter flux between 538 and 528 cmdepth most likely reflects erosion on the crater rim. Periods ofincreased runoff or storminess would simultaneously delivermagnified pulses of clastic sediment components, organicmatter derived from land plants, and nitrates, phosphates andbicarbonate washed out of soil to the lake. Hence, the largeTOC/TN spike between 538 and 528 cm in Fig. 3 appears to bea valid indicator of an episode of increased inwash of land plantmaterial that was probably associated with a significant butshort-lived period of storminess. Furthermore, the spike ofTOC/TN at 538 to 529 cm corresponds to a layer containingclastic sediment and elevated TOC percentages (Fig. 3),suggesting that influx of land-plant (C3) material contributedto the enhanced TOC concentrations. The lower TOCpercentages above 528 cm depth are commensurate withTOC/TN values of 12–15 that indicate a mixed terrestrial-aquatic organic matter source. The low but stable TOC/TNratios suggest a high level of lake stability over this period ofsedimentation.Sedimentary organic matter d13C values are affected by rates
of algal production in lakes which are controlled by dissolved
nutrient and CO2 availability (Meyers and Lallier-Verges,1999). Runoff of soil-derived nitrates and phosphates enhancesaquatic productivity, which selectively removes 12C-richdissolved inorganic carbon and leads to enriched d13C valuesin the organic matter produced by the remaining inorganiccarbon (Meyers, 2003). Hence, the zone of d13C enrichmentbetween 570 and 538 cm and the shift to further enrichmentfrom 538 to 529 cm depth – after which d13C depletion occurs –could reflect phases of enhanced algal productivity as aconsequence of erosion-induced enhanced nutrient flux to thelake. However, the factors that influence the d13C values oflacustrine organic matter are complex, with productivity onlyone of several possible controls that could be involved. Othercontrols include: (1) proportion of organic matter from differentsource organisms; (2) d13C of the carbon substrate used by eachsource; and (3) post-depositional d13C changes (Street-Perrottet al., 2004). Furthermore, lacustrine organic matter is acomplex mix of compounds synthesized by terrestrial plants,aquatic macrophytes, algae and bacteria, all of which maydiffer substantially in their d13C values.Consequently, bulk organic matter d13C analysis on its own is
limited diagnostically because it does not distinguish betweendifferent sources of organic matter entering a lake, so that otherorganic indicators such as TOC/TN or pollen analysis areneeded to identify the organic matter sources (Lamb et al.,2004). In the case of Lake Pupuke, the elevated TOC/TN ratiosbetween 570 and 538 cm, followed by a sustained peak inTOC/TN between 538 and 528 cm depth, indicate thatcatchment erosion has induced increased input of terrigenousorganic matter and sediment into the lake. Such an influxshould be accompanied by increased nutrient delivery to thelake system and enhanced algal growth, with the latter forcingthe utilization of soil-derived bicarbonate in the lake waters togenerate the d13C enrichment during the high aquaticproductivity phases.However, this simple model is complicated by changes in
biogenic silica (BGS) concentrations in the sediment whichprovide an index of algal productivity in the lake. Between 570and 538 cm depth, BGS concentrations are variable butgenerally increasing with a peak at 538 cm. Subsequently,BGS decreases between 538 and 529 cm with a minimum at534 cm, coincident with maxima in the other organic matterproxies and grain size (Fig. 3). The trend in BGS concentrationup-core suggests that changing aquatic productivity is not theonly driver of the d13C enrichment of the bulk organic carbon.Rather, the d13C signal between 529 and 538 cm depth iscontrolled largely by the d13C composition of terrestrialmacrophytes, an interpretation supported by the TOC/TNratios of >20 for all samples below 530 cm depth.
Vegetation history
The pollen evidence indicates that the regional vegetation ofLake Pupuke in the early to middle Holocene was denseconifer–hardwood forest. The canopy emergents were mainlyDacrydium cupressinum, with some Libocedrus and Podocar-pus. Other conifer emergents such as Dacrycarpus dacryoides,Phyllocladus, Prumnopitys taxifolia, P. ferruginea and Agathisaustralis were uncommon, although Agathis, Dacrycarpus andP. ferruginea increase up-core through Zones 2 and 3. Thehardwood component of the forest canopy was mainlyMetrosideroswith someNestegis. The importance of Fuscoporareduces up-core through Zones 1–3, with the low valuesindicating that it was a very minor part of the vegetation, as it istoday. Cyathea tree ferns and Dodonea comprised the major
Copyright � 2008 John Wiley & Sons, Ltd. J. Quaternary Sci., Vol. 23(5) 435–447 (2008)DOI: 10.1002/jqs
RAPID CHANGE IN EARLY HOLOCENE ENVIRONMENTS 443

part of the understorey, with all other understorey taxa poorlyrepresented. These regional trends are similar to those recordedat Lake Waiatarua (Newnham and Lowe, 1991), thoughDacrydium was less dominant at Waiatarua and the other talltree taxa better represented.The early Holocene dominance of Dacrydium is common in
New Zealand pollen records and suggests that climate waswetter than at present (McGlone, 1988; Newnham et al., 1989).Since Agathis australis has tropical affinities, the delay inAgathis expansion when New Zealand forests were rapidlyexpanding as a result of postglacial warming (McGlone, 1988)may have been driven bymoist, cloudy summer conditions thatare not suitable for Agathis growth (Ogden et al., 1992).From 9500� 25 to 7000� 155 cal. yr BP, the forest margins
near the shoreline of Lake Pupuke were fringed with small treesof Dodonea. The very low values for swamp taxa suggest theabsence of a significant local swamp community whichcontinued throughout the Holocene (Horrocks et al., 2005).This absence is probably due to the local topography where thesteep crater rim and small catchment size combined to limit thearea of substrate suitable for wetland plants.The pollen evidence for generally uniform warm, humid
conditions between 9500� 25 and 7000� 155 cal. yr BPcontrasts markedly with the variability in organic productivityevident in TOC, LOI, TOC/TN, biogenic silica, d13C andsediment flux to the lake over this period (Fig. 3).
Diatom palaeoecology
Zone 1 (9500� 25 to ca. 8980 cal. yr BP) is likely to represent arelatively shallow phase in the history of Lake Pupuke. Zones 1,2 and 3 are dominated by Staurosirella pinnata, a very smalldiatom that is common worldwide in both freshwater andbrackish environments (Campeau et al., 1999). Its environ-mental preferences are not well defined, possibly because it istolerant of a wide range of conditions. There is debate aboutwhat factors cause this species to dominate an assemblage, butit is often associated with periods of continuous environmentalchange in core sediment records (e.g., Haworth, 1976; Stabell,1985). In modern lake samples, Gell et al. (2002) found thatS. pinnata occurred in moderately turbid lakes that maintainedwater depth of between 2 and 3m in southeast Australia. InNew Zealand, S. pinnata has been found in very shallow wateraround lake margins (Cochran, 2002). It is classified here as‘facultative planktonic’ because it is generally considered tolive attached to sediment grains, but it is often present inshallow and/or turbulent environments (Vos and de Wolf,1988; Round et al., 1990). In summary, the zones dominated bythis species (Zones 1, 2 and 3) are likely to represent a veryshallow, turbulent and turbid water body that existed at a timeof variable environmental conditions. Most of the abundantdiatoms throughout the core section are recorded to haveenvironmental preferences for water with a pH> 7.5, butsometimes >9 (Vos and de Wolf, 1993; Van Dam et al., 1994;Sonneman et al., 1999), implying that the water body remainedalkaline throughout this time. This is also reflected in the diatomtransfer function-derived pH reconstructions for the sedi-ment record spanning the past 200 years from Lake Pupuke(Augustinus et al., 2006).Zone 2 (ca. 8980 to 8430 cal. yr BP) displays an increase in
the proportion of Staurosirella pinnata, and a reduction in theproportions of Staurosira construens and Pseudostaurosirabrevistriata compared with Zone 1. Although these small
species of the family Fragilariaceae generally occupy similarecological niches, it has been shown that they have distinctenvironmental preferences. For example, Staurosira con-struens is recorded as having a lower and narrower toleranceof trophic conditions (Van Dam et al., 1994) and of salinity(Sonneman et al., 1999). Since all the other diatoms in theassemblage are consistent with freshwater conditionsthroughout this section of core, it is likely that the minordifference between Zones 1 and 2 was the result of a smallincrease in trophic status during the time of Zone 2 rather thanan increase in salinity.
Zone 3 (ca. 8430 to ca. 7790 cal. yr BP) represents a waterbody that was shallow, turbulent and variable, as discussedabove for Zones 1 and 2. However, it is differentiated by therelatively high proportion of aerophilic taxa. Naviculla pusilla,N. radiosa and Diatomella sp. live in wet, moist andtemporarily dry terrestrial (Round et al., 1990; Van Damet al., 1994). This habitat is not consistent with the remainder ofthe shallow but open water diatom assemblage, so it is likelythat these diatoms were transported from the lake margins and/or catchment. This allochthonous input of diatoms indicateseither an increase in wetland/moist soil environments aroundthe lake or an increase in the transport of catchment-derivedsediment to the central lake basin at this time. The increase inclastic sediment influx and TOC/TN-based inference ofincreased terrigenous organic matter inwash occurs over asimilar depth range (Fig. 3) and supports the second explanationfor the high proportion of aerophilics. The highest proportion ofaerophilics occurs in Zone 3 between 543 and 527 cm depth,which correlates with the zone of greatest variability in thephysical and geochemical proxies (Fig. 3), although the changein the diatoms appears to commence below the physical andchemical changes in the sediment and terminates just above.
Zone 4 (ca. 7790 to 7000� 155 cal. yr BP) represents a deep,freshwater lake environment dominated by the planktonicdiatomsCyclotella pseudostelligera andC. stelligera, which arecommon in many New Zealand lakes (Reid, 2005). Haworthand Hurley (1994) proposed that C. pseudostellera is a varietyof C. stelligera. The size difference may be driven byenvironmental conditions because C. stellera is also dominantin oligotrophic–mesotrophic environments (Van Dam et al.,1994). Aulacoseira granulata is a planktonic species thattypically occurs in slow-moving lowland rivers (Gell et al.,2005). A. granulata is also recorded in lakes and rivers withrelatively high silica loads and more turbulent environments(Sonneman et al., 1999). It also occurs in eutrophicenvironments with higher levels of the nutrients P and N(Vos and de Wolf, 1993; Van Dam et al., 1994). This trend fitswell with the lower TOC/TN and reduced TOC and LOI valuesin this zone.
The higher proportion of planktonic species in this zonemakes it more indicative of a typical lake diatom assemblagethan Zones 1, 2 and 3. However, the combined proportions ofbenthic, facultative planktonic and aerophilic species are stillrelatively high, suggesting that the water body was transitioningfrom a shallow-water phase, or that transport from shallowmargins of the lake was effective throughout this time.
Synthesis
The multi-proxy palaeo-environmental record presented fromcore P5 is complex and provides a good example of the value ofusing multiple and independent proxies for assessing changingenvironments. Although the pollen record suggests generallymild, wet and stable early to mid-Holocene conditions,
Copyright � 2008 John Wiley & Sons, Ltd. J. Quaternary Sci., Vol. 23(5) 435–447 (2008)DOI: 10.1002/jqs
444 JOURNAL OF QUATERNARY SCIENCE

possibly with some drying after ca. 7800 cal. yr BP, all of theother proxies indicate significant change in the lake andcatchment over the same period. The interpretation of thevariability in the diatom taxa is complex, but largely reflectschanging lake hydrology, particularly lake level, and phases ofclastic sediment influx. Planktonic diatoms are at low levelsbetween 9000� 25 and ca. 7800 cal. yr BP, while benthic andfacultative planktonic diatoms such as Staurosirella pinnatadominate the diatom flora. Facultative planktonics andbenthics reduce in importance after ca. 7800 cal. yr BP andplanktonic diatoms such as Cyclotella species dominate up tothe time of the deposition of the Tuhua tephra. This pattern isconsistent with a model of relatively low lake levels prior to ca.7800 yr BP accompanied by transport of diatoms derived fromthe lake margins and catchment, with maximum allochthonousinput between ca. 8430 and 7790 cal. yr BP, followed byrapidly rising lake levels with the establishment of a deep andstable lake just prior to the deposition of the Tuhua tephra at7000� 155 cal. yr BP.The diatom-based model of lake level and hydrological
change is supported by the grain size and organic mattergeochemical proxies, whereby clastic sediment influx between9500� 25 and ca. 7800 cal. yr BP is significantly greater thanafter 7800 cal. yr BP and peaks between ca. 8240 and 7880 cal.yr BP. This sediment influx is consistent with a model ofincreased storminess and erosion of the catchment soils.Increased erosion in the catchment at this time is also supportedby the TOC/TN and TOC records: TOC/TN ratios of >20 areindicative of a dominantly terrestrial source of organic matter,while organic matter d13C and low biogenic silica concen-trations between ca. 8240 and 7880 cal. yr BP suggest stress onboth aquatic algae and terrestrial macrophytes during photo-synthesis. The relatively low phytoplankton productivity duringthis low lake level phase may thus be a consequence of bothgrowth limitation due to the restricted availability ofdissolved CO2 required for photosynthesis in relatively shallowand warmer lake waters, and limitations on the availability ofdissolved silica sourced from the erosion of the relativelysilica-poor basaltic soils on the crater rim.The pollen record suggests that moist and mild conditions
dominated throughout the period 9500� 25 to 7000� 155 cal.yr BP with tall trees dominant. Indicators of disturbance such asPoaceae and Pteridium are absent or at very low levelsthroughout the sequence, suggesting little in the way of forestdisturbance as would be expected with the inferred increase instorminess between ca. 8240 and 7880 cal. yr BP. The increasein Agathis after ca. 7800 cal. yr BP is consistent with increase inregional dryness at that time, but is at odds with the otherproxies that all indicate an increase in lake depth and lakestability accompanied by reduced erosion in the catchment.This apparent insensitivity of the pollen record to changingenvironments in the lake and catchment reflects minimal forestdisturbance during the proposed erosional events during theearly Holocene.The overall model of Lake Pupuke palaeo-environments in
the early Holocene is one of warm, mild conditions withrelatively low lake levels between 9505� 25 and ca. 7880 cal.yr BP. A peak in erosion was associated with overall drynessaccompanied by increased storminess between ca. 8240 and7880 cal. yr BP, after which mild but moister conditionscommensurate with higher lake levels dominated for the nextca. 880 years. Such a view of early Holocene climates in theAuckland region is not evident from the pollen record andindicates the relative insensitivity of this proxy in the lowland/coastal Auckland region to all but major climate perturbations.The period of enhanced dryness accompanied by increased
erosion and storminess apparent in the Lake Pupuke multi-
proxy record is broadly coeval with the 8200-year event fromthe North Atlantic region but is of much longer duration. Thediatom evidence for low lake level had a duration of ca640 years (ca. 8340 to 7790 cal. yr BP), while the inferredepisode of increased erosion and storminess was of shorterduration (ca. 360 years) between ca. 8240 and 7880 cal. yr BP.This is similar to the timing and duration of the general climateanomaly in the North Atlantic region starting at ca 8500 cal. yrBP and with a duration of ca. 600 years (Rohling and Palike,2005) rather than the more restricted 8200-year event of150–160 years duration (Kobashi et al., 2007; Thomas et al.,2007). A recent model–data comparison for the 8200-year BPevent suggests that weakening of the thermohaline circulationforced by a freshwater pulse into the Labrador Sea would causedry conditions in the western subtropical Pacific, and onlyslight warming with a total duration of ca. 280 years (Wiersmaand Renssen, 2006). This pattern is consistent with theinterpretation of the multi-proxy Lake Pupuke record asindicating a sustained drier, stormier episode at this time andemphasises the importance of obtaining high-resolution multi-proxy terrestrial palaeo-environmental records from theSouthern Hemisphere that span the early–mid Holocene. Thisenlarged database is essential to allow improvement of ourunderstanding of the global distribution of the 8200-year eventand the nature of the global climate response.
Conclusions
On the basis of the pollen record alone, the early Holoceneenvironmental record from Lake Pupuke appears to be one ofenvironmental stability. However, the high-resolution, multi-proxy record spanning 9500� 25 to 7000� 155 cal. yr BPpresented here indicates that the real situation is likely to be oneof considerable environmental variability. The section of theLake Pupuke sequence we focus on here records warm andrelatively conditions between 9500� 25 and ca. 8240 cal. yrBP, followed by an interval of intensified erosion andstorminess that lasted for ca. 360 years. This climatic per-turbation is most evident in the grain size and organic mattergeochemical proxies, and the diatom flora also record a clearshift to taxa indicative of a lower lake level and increasederosion at this time.An age range of ca. 8240 to 7880 cal. yr BP for the per-
turbation was estimated by assuming constant sedimentationrates between tephra marker beds (Tuhua and Rotoma tephras)of known age. The age and duration remain uncertain at thistime, however, because of an age offset of ca. 570–590 years,probably because the bulk organic matter at these depths in thecore is dominated by terrestrial and aquatic macrophytes.Hence, the production of amore precisely dated sequence fromthis site is problematic without a comprehensive 14C datingprogramme to allow a reliable chronological model for lakesedimentation to be constructed.Nevertheless, the event recorded in Lake Pupuke is of ca.
360 years duration and is consistent with a stormier and drierearly Holocene interval that appears to be coeval with thewell-known ‘8200-year event’ in the North Atlantic region(Alley et al., 1997; Alley and Agustsdottir, 2005; Rohling andPalike, 2005; Kobashi et al., 2007; Thomas et al., 2007).Furthermore, well-dated sections of laminated maar lakesediments from nearby Onepoto maar (Pepper et al., 2004)provided evidence for an El Nino Southern Oscillation signal inthe sedimentary record during the Lateglacial and Last GlacialMaximum, and the erosion signals present in core P5 of Lake
Copyright � 2008 John Wiley & Sons, Ltd. J. Quaternary Sci., Vol. 23(5) 435–447 (2008)DOI: 10.1002/jqs
RAPID CHANGE IN EARLY HOLOCENE ENVIRONMENTS 445

Pupuke between 9500� 25 and ca. 7880 cal. yr BP may reflectan intensification of ENSO at this time. Nevertheless, theproduction of high-resolution Holocene multi-proxy recordsfrom multiple sites in New Zealand is required to enableassessment of whether this is a local effect or of widerpalaeoclimatic significance.
Acknowledgements We acknowledge the Royal Society of NewZealandMarsden Fund (contract UOA0517, the University of AucklandResearch Committee, and the Australian Institute for Nuclear Scienceand Engineering (AINGRA05004) for funding support. Assistance withthe lake sediment coring from Jim Neale, David Jenkinson and JamieShulmeister is gratefully acknowledged.
References
Alley RB, Agustsdottir AM. 2005. The 8k event: cause and con-sequences of a major Holocene abrupt climate change. QuaternaryScience Reviews 24: 1123–1149.
Alley RB, Mayewski PA, Sowers T, Stuiver M, Taylor KC, Clark PU.1997. Holocene climatic instability: a prominent, widespread event8200 yr BP. Geology 25: 483–486.
Alloway BV, Lowe DJ, Barrell DJA, Newnham RM, Almond PC,Augustinus PC, Bertler NAN, Litchfield NI, McGlone MS, Shulme-ister J, Vandergoes MJ, Williams PW. 2007. Towards a climate eventstratigraphy for New Zealand over the past 30,000 years(NZ-INTIMATE project). Journal of Quaternary Science 22: 9–35.
Augustinus PC, Reid M, Andersson S, Deng Y, Horrocks M. 2006.Biological and geochemical record of anthropogenic impacts inrecent sediments from Lake Pupuke, Auckland City, New Zealand.Journal of Paleolimnology 35: 789–805.
Barker MA. 1970. Physico-chemical features of Lake Pupuke, Auck-land. New Zealand Journal of Marine and Freshwater Research 13:406–430.
Battarbee RW. 2000. Palaeolimnological approaches to climaticchange, with special regard to the biological record. QuaternaryScience Reviews 19: 107–124.
Campeau S, Pienitz R, Hequette A. 1999. Diatoms from the BeaufortSea coast, southern Arctic Ocean (Canada): modern analogues forreconstructing Late Quaternary environments and relative sea levels.Bibliotheca Diatomologica, Vol. 42 J. Cramer: Berlin.
Cassie V. 1989. Micro-algae of Lake Pupuke, Auckland, New Zealand.New Zealand Natural Sciences 16: 39–50.
Cochran UA. 2002. Detection of late Holocene earthquakes in thesedimentary record of Wellington, New Zealand, using diatomanalysis PhD thesis, Victoria University of Wellington, Wellington.
Foged N. 1979. Diatoms in New Zealand, the North Island. BibliothecaPhycologica 47: 1–224.
Froggatt PC, Lowe DJ. 1990. A review of the late Quaternary silicic andsome other tephra formations from New Zealand: their stratigraphy,nomenclature, distribution, volume and age.New Zealand Journal ofGeology and Geophysics 33: 89–109.
Gale SJ, Hoare PJ. 1991. Quaternary Sediments. Belhaven Press:London.
Gell P, Sluiter IR, Fluin J. 2002. Seasonal and inter-annual variations indiatom assemblages in Murray River-connected wetlands in north-west Victoria, Australia. Marine and Freshwater Research 53:981–992.
Gell P, Tibby J, Fluin J, Leahy P, Reid M, Adamson K, Bulpin S,MacGregor A, Wallbrink P, Hancock G, Walsh B. 2005. Accessinglimnological change and variability using fossil diatom assemblages,South-East Australia. River Research and Applications 21: 257–269.
Hall CM, York D. 1984. The applicability of 40Ar/39Ar dating to youngvolcanics. In Quaternary Dating Methods, Mahaney WC (ed.).Elsevier: New York; 67–74.
Hammarlund D, Bjorck S, Buchardt B, Israelson C, Thomsen CT. 2003.Rapid hydrological changes during the Holocene revealed by stable
isotope records of lacustrine carbonates from Lake Ogeljon, southernSweden. Quaternary Science Reviews 22: 353–370.
Haworth EY. 1976. Two late-glacial (late Devensian) diatom assem-blage profiles from Northern Scotland. New Phytologist 77:227–256.
Haworth EY, Hurley MA. 1994. Comparison of the stelligeroid taxa ofthe centric diatom genus Cyclotella. In Proceedings of the 8thInternational Diatom Symposium, Renesse 1992, Van Dam H(ed.). Kluwer: Dordrecht; 49–56.
Hayward BW, Grenfell HR, Sabaa AT, Morley MS, Horrocks M. 2006.Effect and timing of increased freshwater runoff into shelteredharbour environments around Auckland City, New Zealand.Estuaries and Coasts 29: 165–182.
Holmes C. 1994. Aspects of the limnology of Lake Pupuke with specialreference to the phytoplankton. MSc thesis, University of Auckland,New Zealand.
Horrocks M, Augustinus PC, Deng Y, Shane P, Andersson S. 2005.Holocene vegetation, environment and tephra recorded from LakePupuke, Auckland, New Zealand. New Zealand Journal of Geologyand Geophysics 48: 85–94.
Horrocks M, Deng Y, Nichol SL, Shane PA, Ogden J. 2002.A palaeo-environmental record of natural and human changefrom the Auckland Isthmus, New Zealand, during the LateHolocene. Journal of the Royal Society of New Zealand 32: 337–353.
Kermode LO. 1992. Geology of the Auckland Urban Area, 1:50 000.Institute of Geological and Nuclear Sciences: Lower Hutt, NewZealand.
Kobashi T, Severinghaus JP, Brook EJ, Barnola J-M, Grachev AM. 2007.Precise timing and characterization of abrupt climate change8200 years ago from air trapped in polar ice. Quaternary ScienceReviews 26: 1212–1222.
Krammer K, Lange-Bertalot H. 1986. Susswasserflora vonMitteleuropa.Bacillariophyceae i Teil Naviculaceae. Gustav Fischer: Stuttgart.
Krammer K, Lange-Bertalot H. 1988. Susswasserflora vonMitteleuropa.Bacillariophyceae ii Teil Bacillariaceae, Epithemiaceae, Surirella-ceae. Gustav Fischer: Stuttgart.
Krammer K, Lange-Bertalot H. 1991a. Susswasserflora von Mitteleur-opa. Bacillariophyceae iii Teil Centrales, Fragilariaceae, Eunotia-ceae. Gustav Fischer: Stuttgart.
Krammer K, Lange-Bertalot H. 1991b. Susswasserflora von Mitteleur-opa. Bacillariophyceae iv Teil Achnanthaceae. Gustav Fischer:Stuttgart.
Krammer K, Lange-Bertalot H. 2000. Bacillariophyceae: English andFrench translation of the keys. In Susswasserflora von Mitteleuropa,Vol. 2/5. Spektrum Akademischer: Heidelberg.
Lamb A, Leng MJ, MohammedMU, Lamb HF. 2004. Holocene climateand vegetation change in the Main Ethiopian Rift Valley, inferredfrom the composition (C/N and d13C) of lacustrine organic matter.Quaternary Science Reviews 23: 881–891.
Ljung K, Bjorck S, Renssen H, Hammarlund D. 2007. South Atlanticisland record reveals a South Atlantic response to the 8.2 kyr event.Climate of the Past Discussions 3: 729–753.
Lowe DJ, Shane PAR, Alloway BV, Newnham RM. 2008. Fingerprintsand age models for widespread New Zealand tephra marker bedserupted since 30,000 years ago: a framework for NZ-INTIMATE.Quaternary Science Reviews 27: 95–126.
McGlone MS. 1983. Polynesian deforestation of New Zealand: apreliminary synthesis. Archaeology in Oceania 18: 1–10.
McGloneMS. 1988. Glacial and Holocene vegetation history –20 ky topresent: New Zealand. In Vegetation History, Huntley B, Webb T(eds). Kluwer: Dordrecht; 557–602.
McGlone MS, Wilmshurst JM. 1999. Dating initial Maori impact inNew Zealand. Quaternary International 59: 17–26.
Mackereth FJH. 1966. Some chemical observations on post-glacial lakesediments. Philosophical Transactions of the Royal Society B250:165–213.
Meyers PA. 1994. Preservation of source identification of sedimentaryorganic matter during and after deposition. Chemical Geology 144:289–302.
Meyers PA. 2003. Evidence of mid-Holocene climate in stability fromvariations in carbon burial in Seneca Lake, New York. Journal ofPaleolimnology 28: 237–244.
Copyright � 2008 John Wiley & Sons, Ltd. J. Quaternary Sci., Vol. 23(5) 435–447 (2008)DOI: 10.1002/jqs
446 JOURNAL OF QUATERNARY SCIENCE

Meyers PA, Lallier-Verges E. 1999. Lacustrine sedimentary organicmatter records of Late Quaternary paleoclimates. Journal of Paleo-limnology 21: 345–372.
Moore PD, Webb JA, Collinson ME. 1991. Pollen Analysis, (2nd edn)Blackwell Scientific: London.
Mortlock RA, Froelich PN. 1989. A simple method for the rapiddetermination of biogenic opal in pelagic marine sediments. DeepSea Research 36: 1415–1426.
Neale JL, Walker D. 1996. Sampling sediment under warm deep water.Quaternary Science Reviews 15: 581–590.
Nesje A, Dahl SO. 2001. The Greenland 8200 cal yr BP event detectedin loss-on-ignition profiles in Norwegian lacustrine sedimentsequences. Journal of Quaternary Science 16: 155–166.
Newnham RM, Lowe DJ. 1991. Holocene vegetation and volcanicactivity, Auckland Isthmus, New Zealand. Journal of QuaternaryScience 6: 177–193.
Newnham RM, Lowe DJ, Green JD. 1989. Palynology, vegetation andclimate of Waikato Lowlands, North Island, New Zealand, since c.18000 years ago. Journal of the Royal Society of New Zealand 19:127–150.
Ogden J, Wison A, Hendy C, Newnham RW. 1992. The Late Qua-ternary history of kauri (Agathis australis) in New Zealand andits climatic significance. Journal of Biogeography 19: 611–622.
Pepper AC, Shulmeister J, NobesDC, Augustinus P. 2004. ENSO signalsduring the Last Glacial Maximum, the Antarctic Cold Reversal andthe early Holocene fromNewZealand.Geophysical Research Letters31: L15206, doi: 10.1029/2004GL020236.
Reid M. 2005. Diatom-based models for reconstructing past waterquality and productivity in New Zealand lakes. Journal of Paleolim-nology 33: 13–38.
Reimer PJ, Baillie MGL, Bard E, Bayliss A, Beck JW, Bertrand CJH,Blackwell PG, Buck CE, Burr GS, Cutler KB, Damon PE, Edwards RL,Fairbank RG, Friedrich M, Guilderson TP, Hogg AG, Hughen KA,Kromer B. 2004. IntCal04 terrestrial radiocarbon age calibration,0–26 cal kyr BP. Radiocarbon 46: 1029–1058.
Rohling EJ, Palike H. 2005. Centennial-scale climate cooling witha sudden cold event around 8200 years ago. Nature 434: 975–979.
Round FE, Crawford RM, Mann DG. 1990. The Diatoms: Biologyand Morphology of the Genera. Cambridge University Press:Cambridge, UK.
Sandiford A, Alloway BJ, Shane P. 2001. A 28,000-6,600 cal. yr recordof local and distal volcanism preserved in a paleolake, Auckland,New Zealand.New Zealand Journal of Geology and Geophysics 44:323–336.
Sandiford A, Newnham RM, Alloway BV, Ogden J. 2003. A 28000-6000 cal. yr BP pollen record of vegetation and climate change from
Pukaki Crater, northern New Zealand. Palaeogeography, Palaeocli-matology, Palaeoecology 201: 235–247.
Shane P. 2000. Tephrochronology: a New Zealand case study. EarthScience Reviews 49: 223–259.
Shane P, Hoverd J. 2002. Distal record of multi-sourced tephra inOnepoto basin, Auckland: implications for volcanic chronology,frequency and hazards. Bulletin of Volcanology 64: 441–454.
Sonneman JA, Sincock A, Fluin J, Reid M, Newall P, Tibby J, Gell P.1999. An Illustrated Guide to Common Stream Diatom Species fromTemperate Australia, Identification Guide No. 33. CooperativeResearch Centre for Freshwater Ecology: Albury, Australia.
Stabell B. 1985. The development and succession of taxa with thediatom genus Fragilaria Lyngbye as a response to basin isolation fromthe sea. Boreas 14: 273–286.
Street-Perrott FA, Ficken KJ, Huang Y, Eglinton G. 2004. Late Qua-ternary changes in carbon cycling on Mt Kenya, East Africa: anoverview of the d13C record in lacustrine organic matter.QuaternaryScience Reviews 23: 861–879.
Thomas ER, Wolf EW, Mulvaney R, Steffensen JP, Johnsen SJ, Arrow-smith C, White JWC, Vaughn B, Popp T. 2007. The 8.2 ka eventfrom Greenland ice cores. Quaternary Science Reviews 26:70–81.
Tomlinson AI, Sansom J. 1994a. Rainfall Normals for New Zealand forthe Period 1961 to, 1990. National Institute of Water and Atmos-pheric Research: Wellington, New Zealand.
Tomlinson AI, Sansom J. 1994b. Temperature Normals for NewZealand for the Period 1961 to 1990. National Institute of Waterand Atmospheric Research: Wellington, New Zealand.
Van Dam H, Mertens A, Sinkeldam J. 1994. A coded checklistand ecological indicator values of freshwater diatoms from theNetherlands. Netherlands Journal of Aquatic Ecology 28: 117–133.
Veski S, Seppa H, Ojala AEK. 2004. Cold event at 8200 yr BP recordedin annually laminated lake sediments in eastern Europe.Geology 32:681–684.
Vos PC, de Wolf H. 1988. Methodological aspects of paleo-ecologicaldiatom research in coastal areas of the Netherlands. Geologie enMijnbouw 67: 31–40.
Vos PC, de Wolf H. 1993. Diatoms as a tool for reconstruc-ting sedimentary environments in coastal wetlands: methodologi-cal aspects. Hydrobiologia 269/270: 285–296.
Wiersma AP, Renssen H. 2006. Model–data comparison for the 8.2 ka BPevent: confirmation of a forcingmechanism by catastrophic drainage ofLaurentide Lakes. Quaternary Science Reviews 25: 63–88.
Woodward CA, Shulmeister J. 2005. A Holocene record of humaninduced and natural environmental change from Lake Forsyth(Te Wairewa), New Zealand. Journal of Paleolimnology 34: 481–501.
RAPID CHANGE IN EARLY HOLOCENE ENVIRONMENTS 447
Copyright � 2008 John Wiley & Sons, Ltd. J. Quaternary Sci., Vol. 23(5) 435–447 (2008)DOI: 10.1002/jqs




















