
Trl-GAGA directly interacts with lola like and both are part of the repressive complex of Polycomb...
9
Trl-GAGA directly interacts with lola like and both are part of the repressive complex of Polycomb group of genes Krishnaveni Mishra, Vivek S. Chopra, Arumugam Srinivasan, Rakesh K. Mishra * Centre for Cellular and Molecular Biology, Uppal Road, Hyderabad 500 007, India Received 11 December 2002; received in revised form 1 April 2003; accepted 1 April 2003 Abstract Epigenetic inheritance to maintain the expression state of the genome is essential during development. In Drosophila, the cis regulatory elements, called the Polycomb Response Elements (PREs) function to mark the epigenetic cellular memory of the corresponding genomic region with the help of PcG and trxG proteins. While the PcG genes code for the repressor proteins, the trxG genes encode activator proteins. The observations that some proteins may function both as PcG and trxG member and that both these group of proteins act upon common cis elements indicate at least a partial functional overlap among these proteins. Trl-GAGA was initially identified as a trxG member but later was shown to be essential for PcG function on several PREs. In order to understand how Trl-GAGA functions in PcG context, we have looked for the interactors of this protein. We identified lola like, aka batman, as a strong interactor of GAGA factor in a yeast two-hybrid screen. lolal also interacts with polyhomeotic and, like Trl, both lolal and ph are needed for iab-7 PRE mediated pairing dependent silencing of mini-white transgene. These observations suggest a possible mechanism of how Trl-GAGA plays a role in maintaining the repressed state of target genes involving lolal, which may function as a mediator to recruit PcG complexes. q 2003 Elsevier Science Ireland Ltd. All rights reserved. Keywords: Polycomb group of genes; Epigenetic inheritance; Polycomb response element; Trl-GAGA; lola like 1. Introduction During development, differential expression patterns of genes that define cell types once established, need to be epigenetically maintained. Specific DNA sequences and the protein complexes working in combination confer this cellular memory. In Drosophila, early regulatory network of genes, referred to as segmentation genes, setup the expression pattern of homeotic genes during the first 2–3 h of development. After this initial period, segmentation proteins disappear but the expression pattern of homeotic genes set up early on is maintained by a mechanism that involves the protein product of a number of genes grouped into Polycomb group (PcG) and trithorax group (trxG) of genes. Generally, these two groups of genes function in opposite directions. Mutations in the PcG genes lead to ectopic activation of homeotic genes, which suggests that their normal function is to repress genes. The trxG genes, on the other hand, have been shown to be positive regulators (Kennison, 1993; Pirrotta, 1998; Simon, 1995). Protein products of PcG and trxG genes are recruited to the target loci through common DNA sequences referred to as Polycomb Response Elements, PRE (Chan et al., 1994; Muller and Bienz, 1991; Simon et al., 1993). Although there are no significant sequence similarities among different PREs that have been functionally characterized so far, there is at least one sequence motif that is conserved (Mihaly et al., 1998) which turned out to be the binding site for a PcG member, pleiohomeotic, pho (Brown et al., 1998). Among the other sequence motifs that are present in most PREs is the GAGA factor binding site. We have shown recently that GAGA motifs are necessary for iab-7 PRE (Mishra et al., 2001). The significance of GAGA binding sites in PRE function has been demonstrated by us and others in case of different PRE by showing that GAGA protein is a part of the protein complex that is assembled at these cis elements (Busturia et al., 2001; Hagstrom et al., 1997; Hodgson et al., 2001; Horard et al., 2000; Mishra et al., 2001). These observations suggest that multiple sequence motifs, in certain combinations, contribute to PRE function, probably, 0925-4773/03/$ - see front matter q 2003 Elsevier Science Ireland Ltd. All rights reserved. doi:10.1016/S0925-4773(03)00046-7 Mechanisms of Development 120 (2003) 681–689 www.elsevier.com/locate/modo * Corresponding author. Tel.: þ91-40-2719-2658; fax: þ 91-40-2716- 0311. E-mail address: [email protected] (R.K. Mishra).
Transcript of Trl-GAGA directly interacts with lola like and both are part of the repressive complex of Polycomb...

Trl-GAGA directly interacts with lola like and both are part of the
repressive complex of Polycomb group of genes
Krishnaveni Mishra, Vivek S. Chopra, Arumugam Srinivasan, Rakesh K. Mishra*
Centre for Cellular and Molecular Biology, Uppal Road, Hyderabad 500 007, India
Received 11 December 2002; received in revised form 1 April 2003; accepted 1 April 2003
Abstract
Epigenetic inheritance to maintain the expression state of the genome is essential during development. In Drosophila, the cis regulatory
elements, called the Polycomb Response Elements (PREs) function to mark the epigenetic cellular memory of the corresponding genomic
region with the help of PcG and trxG proteins. While the PcG genes code for the repressor proteins, the trxG genes encode activator proteins.
The observations that some proteins may function both as PcG and trxG member and that both these group of proteins act upon common cis
elements indicate at least a partial functional overlap among these proteins. Trl-GAGA was initially identified as a trxG member but later was
shown to be essential for PcG function on several PREs. In order to understand how Trl-GAGA functions in PcG context, we have looked for
the interactors of this protein. We identified lola like, aka batman, as a strong interactor of GAGA factor in a yeast two-hybrid screen. lolal
also interacts with polyhomeotic and, like Trl, both lolal and ph are needed for iab-7 PRE mediated pairing dependent silencing of mini-white
transgene. These observations suggest a possible mechanism of how Trl-GAGA plays a role in maintaining the repressed state of target genes
involving lolal, which may function as a mediator to recruit PcG complexes.
q 2003 Elsevier Science Ireland Ltd. All rights reserved.
Keywords: Polycomb group of genes; Epigenetic inheritance; Polycomb response element; Trl-GAGA; lola like
1. Introduction
During development, differential expression patterns of
genes that define cell types once established, need to be
epigenetically maintained. Specific DNA sequences and the
protein complexes working in combination confer this
cellular memory. In Drosophila, early regulatory network of
genes, referred to as segmentation genes, setup the
expression pattern of homeotic genes during the first 2–3 h
of development. After this initial period, segmentation
proteins disappear but the expression pattern of homeotic
genes set up early on is maintained by a mechanism that
involves the protein product of a number of genes grouped
into Polycomb group (PcG) and trithorax group (trxG) of
genes. Generally, these two groups of genes function in
opposite directions. Mutations in the PcG genes lead to
ectopic activation of homeotic genes, which suggests that
their normal function is to repress genes. The trxG genes, on
the other hand, have been shown to be positive regulators
(Kennison, 1993; Pirrotta, 1998; Simon, 1995).
Protein products of PcG and trxG genes are recruited to
the target loci through common DNA sequences referred
to as Polycomb Response Elements, PRE (Chan et al., 1994;
Muller and Bienz, 1991; Simon et al., 1993). Although there
are no significant sequence similarities among different
PREs that have been functionally characterized so far, there
is at least one sequence motif that is conserved (Mihaly
et al., 1998) which turned out to be the binding site for a PcG
member, pleiohomeotic, pho (Brown et al., 1998). Among
the other sequence motifs that are present in most PREs is
the GAGA factor binding site. We have shown recently that
GAGA motifs are necessary for iab-7 PRE (Mishra et al.,
2001). The significance of GAGA binding sites in PRE
function has been demonstrated by us and others in case of
different PRE by showing that GAGA protein is a part of the
protein complex that is assembled at these cis elements
(Busturia et al., 2001; Hagstrom et al., 1997; Hodgson et al.,
2001; Horard et al., 2000; Mishra et al., 2001). These
observations suggest that multiple sequence motifs, in
certain combinations, contribute to PRE function, probably,
0925-4773/03/$ - see front matter q 2003 Elsevier Science Ireland Ltd. All rights reserved.
doi:10.1016/S0925-4773(03)00046-7
Mechanisms of Development 120 (2003) 681–689
www.elsevier.com/locate/modo
* Corresponding author. Tel.: þ91-40-2719-2658; fax: þ91-40-2716-
0311.
E-mail address: [email protected] (R.K. Mishra).

by recruiting different subsets of proteins that form the
chromatin complex at different loci or same locus in
different cell type.
Involvement of Trl-GAGA in repressive function of PcG
proteins, however, raises a paradox as Trl-GAGA has been
identified as an activator protein, an enhancer of PEV and a
member of trxG of genes (Biggins and Tjian, 1988; Soeller
et al., 1993; Farkas et al., 1994). How does Trl-GAGA
carryout these apparently opposing functions? It is now
emerging that PcG and trxG proteins function on the same
cis elements and therefore PRE and TRE may be closely
linked or overlapping (Orlando et al., 1998; Strutt et al.,
1997; Tillib et al., 1999). The mechanism of this assembly
of chromatin structure that is capable of maintaining
expression of linked genes, is not clear yet. The GAGA
factor has been shown to be involved in ATP dependent
nucleosome remodeling (Tsukiyama et al., 1994). Recent
studies provide a direct link between chromatin remodeling
and PcG mediated maintenance of the repressive chromatin
(Czermin et al., 2002; Muller et al., 2002). Any direct link
between this remodeling and Trl-GAGA remains to be
established. On the contrary, GAGA protein was not
detected in the ESC-E(Z) protein complex capable of
histone H3 lysine 27 methylation which may be one of the
steps in remodeling process that ensures nucleosomes stay
in repressive chromatin conformation (Muller et al., 2002).
There still may, however, be a more dynamic role of
Trl-GAGA in the assembly of the PcG protein complex
(Poux et al., 2001a).
We decided to investigate the mechanism behind the
function of Trl-GAGA as a component of the repressive
Polycomb system by identifying the direct interacting
partners of this protein. We used the yeast two-hybrid
system to screen a cDNA library made from Drosophila
embryos. During this screen we identified lola like (lolal)
aka batman as one of the strong interactors of GAGA factor.
Since lolal has previously been shown to interact with
polyhomeotic, ph-p, a known PcG gene, it is likely that PRE
function of Trl-GAGA is mediated by lolal. Our genetic and
biochemical interaction studies presented here suggest that
lolal may be the effecter molecule for the Trl-GAGA
mediated PcG function.
2. Results
2.1. Identification of lolal as a potential partner for GAGA
GAGA factor contains three distinct domains, Fig. 1A
(Farkas et al., 1994). The BTB/POZ domains of several
proteins have been reported to mediate homo- and
heterodimerizations (Bardwell and Treisman, 1994) and
can be functionally swapped between two proteins (Read
et al., 2000). We reasoned that GAGA may carry out its
diverse roles by recruiting different proteins to the target
DNA sequences and the BTB domain may mediate this. We,
therefore, performed a yeast two-hybrid screen using the
BTB domain of GAGA, amino acids 1–245 (Fig. 1A) as the
bait. We screened the 0–16 h embryo cDNA library to
identify potential partners for Trl-GAGA protein. Out of the
46 adenine auxotrophs, 23 strongly activated the second
reporter gene, b-galactosidase. All the 23 positives inter-
acted specifically with the bait upon retransformation and 14
turned out to be one single gene lola like (lolal) (Fig. 1A).
All the clones had complete coding region of the gene.
LOLAL also interacts with the full length GAGA protein
(Fig. 1B). Quantitative assays showed that the interaction of
LOLAL with BTB and GAGA full-length protein was
comparable (data not shown). Indeed, in an independent
screen using full length GAGA protein, LOLAL appeared
eight times (Srinivasan, A., Mishra, K., Mishra, R.K.,
unpublished). We also isolated other BTB domain contain-
ing proteins multiple times and one already reported
interactor of Trl-GAGA, Sin3A (Espinas et al., 2000). We,
however, did not isolate GAGA factor itself in these screens,
which may suggest that BTB domain of this protein might
not form a homodimer. When tested in directed two hybrid
assay, though, we did see BTB–BTB interaction. It has
been reported that Trl-GAGA exists in several isoforms
(Benyajati et al., 1997). All the experiments reported here
have been done with the major isoform, GAGA 519. We do
not know if other isoforms can interact differently.
Furthermore, pipsqueak that binds to GAGA sequences
(Lehmann et al., 1998) and contains BTB domain has also
been shown to be essential for sequence specific targeting of
PcG protein complex (Huan et al., 2002). It is unclear,
however, how Trl-GAGA is placed in this multi-component
recruitment system working on GAGA motif of PREs.
Further studies will be needed to answer these questions.
lolal was originally identified in the Drosophila gene
disruption project (Spradling et al., 1999). Later on this
mutation was also found to enhance the homeotic phenotype
of polyhomeotic, ph, and renamed as batman (Faucheux
et al., 2001). lolal encodes a protein of 127 amino acids that
contains a BTB domain of about 90 amino acids, leaving
Fig. 1. (A) Molecular map showing the domains of GAGA factor and
LOLAL. The N-terminal region containing BTB domain of GAGA factor
was used in the yeast two hybrid screen. (B) Yeast transformants containing
the constructs indicated in the middle panel were patched on to synthetic
media lacking leucine and uracil (to select for the plasmids) and synthetic
media lacking leucine, uracil and adenine (to select for the interactors).
K. Mishra et al. / Mechanisms of Development 120 (2003) 681–689682

only few residues at both ends of the protein for any other
functional motif/domain. Another PcG protein, ESC,
consists almost entirely of six WD40 repeat motifs (Sathe
and Harte, 1995; Simon et al., 1995). Unlike multi-domain
proteins, the ones made of a single domain alone may
function as adaptor modules to bring together two different
molecules/complexes. Trl-GAGA is known to activate
transcription of several genes. In this context, lolal may
function to inhibit this activation role of Trl-GAGA, in a
way similar to MyoD–Id interaction (Sun et al., 1991),
further studies will be required to differentiate between
these mechanisms.
2.2. LOLAL interacts with GAGA factor in vitro and exists
in a complex that also contains PC
In order to further verify the GAGA–LOLAL inter-
action, we performed a GST pull-down assay. Full length
LOLAL was expressed as a GST fusion protein in
Escherichia coli. Nuclear extracts were prepared from
0–16 h wild-type Drosophila embryos and incubated with
either GST or GST–LOLAL. The complexes were pulled
down using glutathione Sepharose and tested for the
presence of GAGA protein by Western blotting. As shown
in Fig. 2A, GAGA protein comes down specifically with
GST–LOLAL. In the light of yeast two-hybrid interaction
studies, this assay further suggests that GAGA and LOLAL
probably interact directly. It has been shown that GAGA
factor associates with PC, PH and PSC (Poux et al., 2001a).
Since our results show a direct interaction between LOLAL
and GAGA factor, we tested if PC is also pulled down by
GST–LOLAL. As shown in Fig. 2A lower panel, GST–
LOLAL does enrich PC. We used antibodies raised against
two nuclear proteins, lamin B1 and F7BP1, to probe the
same blot to rule out any nonspecific protein–protein
interaction under these conditions. The two pull-down
experiments suggest that LOLAL is a part of the PcG
protein complex. LOLAL could be the adaptor between
GAGA and PC as the latter two coimmunoprecipitate
(Horard et al., 2000)
2.3. PH and GAGA factor interact in nuclear extracts
Earlier reports suggested that lolal enhances the sex
comb phenotype of ph (Faucheux et al., 2001). Also our
results described above suggest that, at least in vitro,
LOLAL can be associated with the GAGA containing
complex that includes PC and other PcG proteins. We
decided to test if GAGA and PH interact. The nuclear
extracts were made from 0–16 h embryo and larvae of
transgenic flies carrying FLAG-tagged PH (Shao et al.,
1999). Nuclei were isolated from the embryo or larvae and
extracts were prepared using standard protocols. To the
extracts as such or after incubation with bacterially
produced GST–LOLAL, M2 coupled agarose (Sigma)
was added. FLAG-PH bound complex isolated in this
manner was analyzed by western analysis for the presence
of GAGA. As shown in Fig. 2B, GAGA association with PH
in embryos was dependent on externally provided GST–
LOLAL, while in the larval extracts, PH was found to be
associated with GAGA factor, irrespective of whether or not
external GST–LOLAL was added.
2.4. Genetic interactions of lolal
2.4.1. Pairing dependent silencing caused by iab-7PRE is
dependent on lolal and ph
The pairing-dependent silencing or pairing sensitivity,
PS, assay is widely used for checking the PRE nature of a
given sequence and previous studies have shown that
mutations in PcG genes relieve this pairing-dependent
silencing (Chan et al., 1994; Gindhart and Kaufman, 1995;
Hagstrom et al., 1997; Kassis, 1994). We have already
shown that transgenic lines carrying iab-7 PRE next to
mini-white reporter, PRE-mw, show pairing dependent
silencing of the mini-white and that like several PcG
mutations, this silencing was affected by Trl mutations
(Hagstrom et al., 1997; Mishra et al., 2001). We used
these PRE-mw lines to test if lolal is required for the
PS function. Since lolal was shown to interact with ph
(Faucheux et al., 2001), we also checked ph in this PS
assay.
Fig. 2. (A) GST pull-down assay. Total nuclear extract (NE), equivalent to
one fourth of the input, are loaded as control for the antibody detection.
G-LOLAL is pull-down pellet loaded from the GST–LOLAL fusion
protein coupled to glutathione Sepharose, GST indicates the lane
where pull-down pellet is loaded from the GST protein coupled to
glutathione Sepharose as negative control. Western blotting with anti-
bodies against GAGA factor, POLYCOMB, LAMIN B and F7BP1 are
shown. (B) FLAG-PH coimmunoprecipitation: FLAG-PH nuclear extracts
prepared from embryos (EE) or larvae (LE) were incubated with GST-lolal
fusion protein or GST alone. The complex was then pulled down using M2
agarose. Western blot was probed with anti GAGA antibody.
K. Mishra et al. / Mechanisms of Development 120 (2003) 681–689 683
In several transgenic lines, PRE-mw/PRE-mw flies show
lighter eye color as compared to the PRE-mw/ þ flies
(Mishra et al., 2001). All the lines tested showed clear
requirement of both lolal and ph for the PS function, as the
eye color of PRE-mw/PRE-mw flies showed clear enhance-
ment when brought in lolal (ban Df(2R)311a) or ph ( ph-p)
background, Fig. 3. We got similar results with P element
insertion allele of lolal (lolal k02512), data not shown. In this
case, although there was a clear enhancement of pigmenta-
tion in PRE-mw/PRE-mw flies when tested in lolal back-
ground, we have also measured the red pigment (Ashburner,
1989), of these mutant combinations to distinguish the
increase in pigmentation from mere additive effect of the
light eye color of P element of the lolal allele. The results
show that value for PRE-mw/PRE-mw in lolal/CyO context
is 50% more than added values of PRE-mw/PRE-mw and
lolal/CyO flies (data not shown).
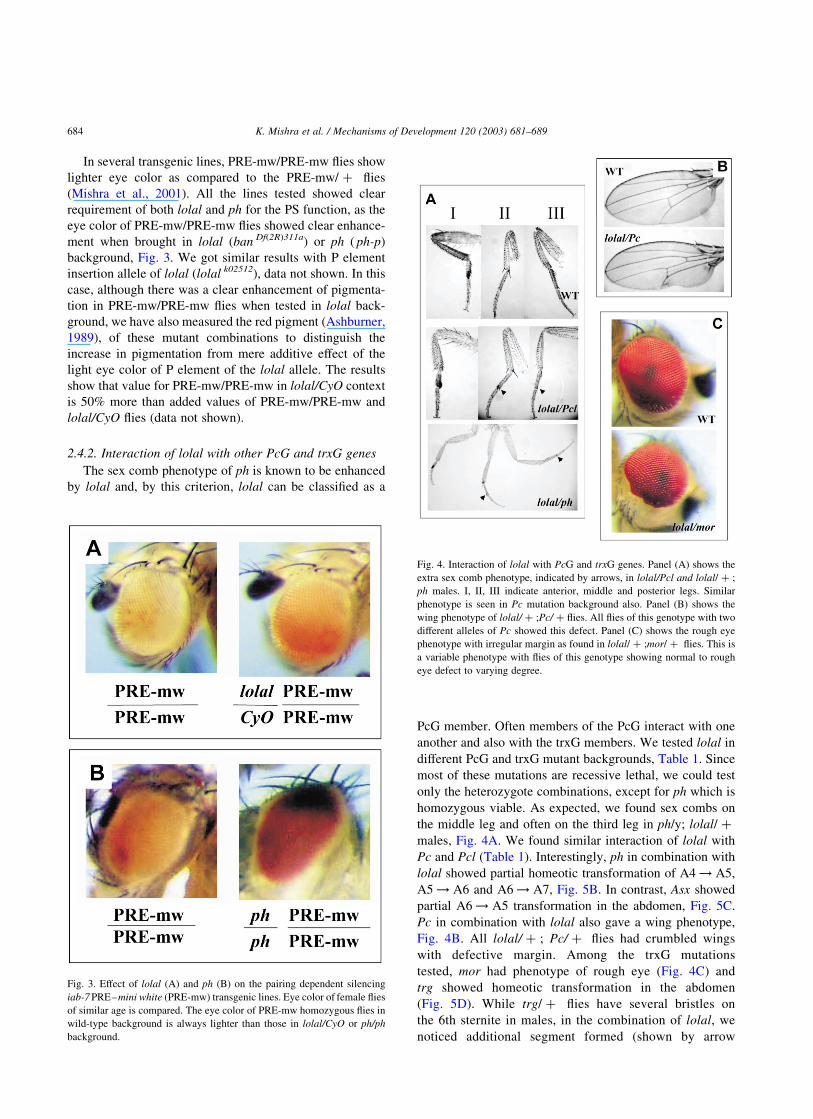
2.4.2. Interaction of lolal with other PcG and trxG genes
The sex comb phenotype of ph is known to be enhanced
by lolal and, by this criterion, lolal can be classified as a
PcG member. Often members of the PcG interact with one
another and also with the trxG members. We tested lolal in
different PcG and trxG mutant backgrounds, Table 1. Since
most of these mutations are recessive lethal, we could test
only the heterozygote combinations, except for ph which is
homozygous viable. As expected, we found sex combs on
the middle leg and often on the third leg in ph/y; lolal/ þ
males, Fig. 4A. We found similar interaction of lolal with
Pc and Pcl (Table 1). Interestingly, ph in combination with
lolal showed partial homeotic transformation of A4 ! A5,
A5 ! A6 and A6 ! A7, Fig. 5B. In contrast, Asx showed
partial A6 ! A5 transformation in the abdomen, Fig. 5C.
Pc in combination with lolal also gave a wing phenotype,
Fig. 4B. All lolal/ þ ; Pc/ þ flies had crumbled wings
with defective margin. Among the trxG mutations
tested, mor had phenotype of rough eye (Fig. 4C) and
trg showed homeotic transformation in the abdomen
(Fig. 5D). While trg/ þ flies have several bristles on
the 6th sternite in males, in the combination of lolal, we
noticed additional segment formed (shown by arrow
Fig. 3. Effect of lolal (A) and ph (B) on the pairing dependent silencing
iab-7 PRE–mini white (PRE-mw) transgenic lines. Eye color of female flies
of similar age is compared. The eye color of PRE-mw homozygous flies in
wild-type background is always lighter than those in lolal/CyO or ph/ph
background.
Fig. 4. Interaction of lolal with PcG and trxG genes. Panel (A) shows the
extra sex comb phenotype, indicated by arrows, in lolal/Pcl and lolal/ þ ;
ph males. I, II, III indicate anterior, middle and posterior legs. Similar
phenotype is seen in Pc mutation background also. Panel (B) shows the
wing phenotype of lolal/ þ ;Pc/ þ flies. All flies of this genotype with two
different alleles of Pc showed this defect. Panel (C) shows the rough eye
phenotype with irregular margin as found in lolal/ þ ;mor/ þ flies. This is
a variable phenotype with flies of this genotype showing normal to rough
eye defect to varying degree.
K. Mishra et al. / Mechanisms of Development 120 (2003) 681–689684

mark in Fig. 5D) indicating partial anteriorization,
transformation of A7 to A6.
Contrary to the expectations based on the biochemical
studies discussed above, we did not notice any apparent
interaction of lolal with Trl 13C in heterozygous condition.
Similarly, among the mutations that we tested, osa, trx,
ash1, Psc, Scm did not show any interaction with lolal.
Interestingly, it has been shown recently that Trl also
enhances ph phenotype in a fashion similar to what we see
in case of lolal and ph (Hodgson et al., 2001). This raises the
possibility that there may be a networking of Trl-lolal-ph
and, perhaps, other PcG proteins in the PRE mediated
repression mechanisms. These interactions of lolal with ph
and Pc suggest that association of lolal may be the key step
of PcG function of Trl-GAGA.
3. Discussion
Chromatin organization is a vital component of the
mechanism regulating gene expression in eukaryotes. The
molecular mechanisms of how these regulatory processes
take place – how active and inactive regions of chromatin
are established during development and maintained through
a number of cell divisions – is still far from being
understood. In Drosophila, extensive genetic and biochemi-
cal studies on the homeotic gene regulation has implicated
Polycomb and trithorax group of genes in the maintenance
of expressed state at the level of higher order chromatin
organization (Pirrotta, 1997; Simon, 1995). It is emerging
that high molecular weight complexes consisting of few
‘core’ PcG proteins and other variable components exist
(Hodgson et al., 2001; Horard et al., 2000; Poux et al.,
2001b; Saurin et al., 2001; Simon et al., 1993). The variable
members may determine stage specific or locus specific
function. For example, pho has been shown to be more
crucial for late larval and pupal stages as compared to the
early embryonic development (Poux et al., 2001b).
One of the key steps in the PcG/trxG mediated
maintenance is the recruitment of the multi-protein complex
of correct composition onto the PRE. Recent studies have
Table 1
Interaction of lolal with PcG and trxG genesa
Mutation Phenotypeb Comments
Pcl Sex comb on the 2nd leg All male flies, sex comb teeth per leg 4 ^ 1
Sex comb on the 3rd leg All male flies, sex comb teeth per leg 3 ^ 1
Pc1 Sex comb on the 2nd leg All male flies, sex comb teeth per leg 5 ^ 1
Sex comb on the 3rd leg All male flies, sex comb teeth per leg 2 ^ 1
Curved wings with posterior margin defect All flies show the phenotype
ph410 Sex comb on the 2nd leg All male flies, sex comb teeth per leg 3 ^ 2
Sex comb on the 3rd leg 12.5% male flies, sex comb teeth per leg 3 ^ 1
Segmental transformation All male flies show reduction in 6th segment
Asx[XF23] Bristle on the 6th male sternite 26.3% male flies
mor1 Rough eye (In females) This phenotype is quite variable with flies showing very mild to strong
defects
trg Segmental transformation 69.2% males show partial A7 to A6 transformation (47% males show this
phenotype when trg is brought paternally)
a Psc1, ScmR5.13B, ash1 B1, trxred, osa00090 phoCV and Trl13C did not show any interaction in double heterozygous conditions based on such clear visible
phenotypic criteria.b Phenotype shown is the most prominent one seen with the corresponding allele in lolal/ þ background. Phenotype reported for Asx[XF23], Pcl and ph
were seen when the PcG mutation was brought maternally. In case of Pc1, the Pc1/ þ ; þ /CyO siblings also showed 5 ^ 1 sex comb teeth per 2nd leg, while
this genotype did not show any sex comb teeth on the 3rd leg.
Fig. 5. Cuticular structures of the adult male abdomen. Anterior is top and
posterior is bottom. The abdomen was split mid-dorsally and flattened.
Cuticle preparation of differential mutant combinations with lolal is shown.
(A) is wild-type fly; (B) is lolal/ þ ;ph showing strong reduction in the size
of 6th tergite (indicated by black arrowhead) and additional pigmentation of
the 4th tergite (indicated by hatched arrow). (C) is lolal/ þ ;Asx/ þ
abdomen with few additional bristles on the 6th sternite (indicated by
arrow), wild-type male cuticle is devoid of bristles in this region. We almost
always found two bristles in these flies. The rest of the cuticle appears
normal. (D) is lolal/ þ ;trg/ þ cuticle preparation. These flies show
development of a small tergite (indicated by arrowheads). These
preparations have several bristles in the 6th sternite, which is not due to
lolal as these bristles are present in similar numbers in trg/ þ flies.
K. Mishra et al. / Mechanisms of Development 120 (2003) 681–689 685

shown that more than one or perhaps several recruiting
processes take place in concert. It is likely that different
recruitment possibilities provide the necessary variation that
is needed for the establishment and maintenance of varying
transcriptional states at hundreds of different loci. Our
studies identify a new member in this process. Trl-GAGA
bound to specific sites on the PREs recruits LOLAL, which
in turn, through direct or indirect means, incorporates PH
into the complex. This raises a question whether the other
recruiting agents like pho, zeste, etc. (Americo et al., 2002;
Brown et al., 1998) function in cooperation or competition
with each other. Also, it is not clear if the complex is
assembled de novo on the PREs, a pre-assembled complex
is recruited or partly assembled sub-complexes are
recruited. Since large complexes of PcG proteins can be
isolated, we can conclude that such structures, once
assembled are stable. It is not clear though if these
complexes are stable during cell division or they assemble
each time a cell divides.
As most of the PREs contain Trl-GAGA binding sites, it
is likely that at least some PcG complexes are recruited by
GAGA factor through its direct interaction with LOLAL.
These findings establish a molecular link between Trl-
GAGA and the PcG complex. Since Trl-GAGA functions in
several other processes that do not seem to be directly linked
to the PRE function, it is likely that there are several
interactors of Trl-GAGA. We are studying such interactors
and initial observations suggest that a large number of
proteins can interact with Trl-GAGA with a potential to
target a repressive or activator function to different loci. It is
not clear though how the selection of appropriate partner is
made. Is it in the context in which Trl-GAGA is bound or
different heterodimers preexist in the nucleus and these are
then recruited to appropriate loci? New assays will have to
be designed to appropriately answer these questions.
Our genetic interaction studies (Table 1) show that lolal
interacts with a variety of PcG and trxG mutations, Figs. 4
and 5. This underscores the important role of this protein in
the regulation of developmental genes. Interestingly, we
find that lolal interactions with ph mutation leads to
transformation of 2nd (and some times 3rd) leg to 1st leg,
an apparent anteriorization type of homeotic transformation
in thoracic but in the abdominal region same combination
leads to posteriorization type of homeotic transformation,
pigmentation of A4 (A4 ! A5) reduction in the size of A6
(A6 ! A7). However, appearance of sex comb in 2nd and
3rd legs is also known to be due to derepression of Scr in
posterior segments thereby explaining this phenotype as due
to loss of the repression function of the PcG proteins.
Furthermore, trxG and PcG mutations upon interaction with
lolal can give a similar phenotype. In lolal context, Asx and
trg both show partial A6 ! A5 transformation in abdominal
region. Pc is involved in pairing dependent silencing
complex recruited by iab-7 PRE. In this study we show
that ph is also involved in the PS function of iab-7 PRE.
While it was known that lolal enhances the homeotic
phenotype of ph, we demonstrate that both ph and lolal are
involved in establishing the repressive complex at the
iab-7 PRE. This indicates that lolal and ph function in
coordination to set up a repressive complex. Taken together,
these results suggest that lolal may be acting along with
Trl-GAGA or with other partners in different complexes in a
locus or stage specific manner. Depending on the context it
could be an activator or repressor function. Since not only
‘ON’ or ‘OFF’ but also several ‘levels of expression states’
for a given hox gene or indeed other regulated loci are
maintained, it is likely that a unique combination of trxG
and PcG proteins may be needed for each varying levels of
expression state of a given locus.
Affinity pull-down experiments, Fig. 2, show that
GAGA, LOLAL, PH and PC proteins coexist in a complex.
This is also in agreement with the genetic interaction
studies, where we found interaction of lolal with ph and Pc.
Genetic and biochemical studies also suggest that, like Trl,
lolal could also be not specifying the kind of complex to be
assembled. It is likely, therefore, that specificity of the
GAGA partner, PcG or trxG member, does not come from
the lolal. It is even possible that lolal could also be a
multifunctional adapter of Trl-GAGA in assembling multi
protein complexes. The specificity may come from yet
another factor or even from the transcriptional activity
around the locus. Recent observations that transcription
process itself may contribute to the cellular memory may
support this view (Bender and Fitzgerald, 2002; Hogga and
Karch, 2002; Rank et al., 2002). This might bring together
the ability of GAGA factor to support transcription and
recruitment of multi-protein complexes and nucleosome
remodeling activity in one mechanistic context.
Trl-GAGA has been suggested to be involved in creating
a nucleosome free region (Hagstrom et al., 1997). The first
step in establishing a PcG complex may be this Trl-GAGA
mediated nucleosome remodeling of the chromatin on the
PRE region to create a more accessible (DNase I
hypersensitive site). The recruitment of a protein complex
to the accessible region may take place through GAGA
factor or by other factors that can anchor the complex on to
DNA. As the next step LOLAL could mediate recruitment
of initial complex, for example, ESC-E(Z) protein complex,
which modifies nearby histone tails to covalently mark the
region for the recruitment of another complex, like PRC1.
Consistent histone modifications and remodeling may be
needed to maintain the chromatin conformation. Our studies
would place LOLAL as the factor binding to the DNA
bound GAGA even when the rest of the complex is not
recruited and therefore help in subsequent recruitment steps.
In this context, the exact function of proteins like LOLAL
becomes very important. Further studies will be required to
clarify these issues.
3.1. Note
While this manuscript was under reviewing process, lolal
K. Mishra et al. / Mechanisms of Development 120 (2003) 681–689686

(batman) was reported to interact with PcG and trxG genes
and also was found in a complex containing GAGA factor
(Faucheux et al., 2003).
4. Experimental procedures
4.1. Strains and plasmids
The full-length and BTB-containing GAGA fragments
were amplified using polymerase chain reaction (PCR)
primers and cloned in frame with the bait vector,
pGBDCU2. GST fusion constructs were made by sub
cloning full-length lolal from the pACT2-LOLAL obtained
from the screen into pGEX5x-2. E. coli strain XL1 blue was
used routinely. HB101 was used for amplifying yeast
plasmids and BL21 for expression of protein. All yeast
media were prepared according to Rose et al. (1990). PJ69-
4A (Mata trp1-901 leu2-3,112 ura3-52 his3-200 gal4 gal80
LYS2::GAL1-HIS3 GAL2-ADE2 met2::GAL7-lacZ) was
used for the screen. All yeast transformations were done by
the high efficiency LiAc method (Gietz and Schiestl, 1995).
Library transformations were scaled up 10-fold.
4.2. Yeast two-hybrid screen
The system developed by James et al. (1996) was used
for the screen. The bait, BTB containing region of GAGA
factor (amino acids 1–245), was cloned in frame with the
Gal4 DNA binding domain of pGBDUC2. The strain
PJ69-4a was transformed with pGBDUC2-BTB and
checked to see if it turned on any of the reporters. The
bait activated the HIS3 reporter weakly and therefore the
screen was carried out using the ADE2 reporter. The strain
containing the bait was transformed with the Drosophila
embryonic cDNA library (embryo MATCH MAKER
cDNA library, Clonetech) and plated on to SC-Leu-Ura-
Ade selective plates. Out of about 50 000 transformants
screened, 46 adenine auxotrophs were obtained. Out of
these, 23 also activated the b-galactosidase reporter. The
GAL4 AD plasmid was extracted from yeast, amplified in
E. coli (HB101) and retransformed into PJ69 24A either
with the bait or with full length GAGA factor in the bait
vector or empty bait vector. All the 23 interacted
specifically with GAGA factor but did not activate the
reporter genes with the vector alone. Interactions were also
quantified using liquid b-galactosidase assays.
4.3. GST pull-down assay and coimmunoprecipitation
The pull-down assay was done essentially as described
(Sambrook and Russell, 2001). Briefly, 0–16 h embryos
were collected and nuclear extract was prepared as
described (Han et al., 1993). GST and GST–LOLAL fusion
protein were expressed in bacteria, BL21 and bound to
glutathione Sepharose. The Sepharose gel was washed
several times with PBS, resuspended as a 50% suspension
and stored at 4 8C. A small aliquot was checked on SDS
polyacrylamide gel for size and concentration of proteins.
Gel containing equal amounts of protein (30 ml) was added
to nuclear extract (100 mg of protein per reaction);
incubated for 2 h at 4 8C, washed several times with PBS
and finally resuspended in 30 ml of Laemmli buffer. The
samples were run on SDS polyacrylamide gel, transferred
onto Immobilon membrane and blotted with anti-GAGA
antibodies (a gift from Carl Wu) or anti-PC antibodies (a gift
from Renato Paro). The blots were developed using the
Chemiluminescence kit from NEN Life Science Products.
As negative controls, we also probed these blots with anti-
Lamin B (a gift from Veena Parnaik) and anti-F7BP1
antibodies. The anti-Lamin B is polyclonal rat lamin B1
antibody that recognizes DmO, the Drosophila lamin B
homolog (Jagatheesan et al., 1999). The anti-F7BP1 is
polyclonal antibody was raised against F7BP1, a protein
identified in a screen to isolate Fab-7 boundary binding
protein (R.K. Mishra, F. Karch, in preparation).
Overnight collection of FLAG-PH embryos was done
and nuclear extract was prepared as described above.
Similarly, 2nd and 3rd instar larvae were collected,
homogenized to isolate nuclei and nuclear extract was
prepared. These extracts were incubated with M2 affinity
agarose for 4–6 h at 4 8C, spun down and washed several
times in PBS. Finally, the agarose matrix was boiled in
loading buffer and separated on SDS–polyacrylamide gel
electrophoresis for Western analysis.
4.4. Fly strains and mutants
FLAG-PH expressing fly was supplied by Robert Kingston,
Trl mutations and iab-7 PRE lines were from Francois Karch,
lolal deletion mutation (ban Df(2R)311a) was from Laurent
Theodore and trg was supplied by Ruth Lehmann. Following
mutations used in this study were supplied by Bloomington
Drosophila stock center: Psc 1/CyO, Scm R5-13B/TM3,
Asx[XF23]/CyO, esc 2CyO/DpGYL(esc 2), Pcl/CyO,
Pc 1/TM6, Sb, Tb, In(1)ph410, ph-p[410] w[1],
ash1 B1/TM6C, cu 1 Sb 1 ca 1, brm 2 e s ca 1/TM6B, Sb 1
Tb 1, trx/TM6, ry 506 P{ry þt7.2 ¼ PZ}osa 00090/TM3,
mor 1/TM6B, Tb 1, lolal (y1 w67c23; P {w þ mC ¼ lacW}
lolalk02512/CyO).
4.5. Pairing sensitivity lines in lolal (batman) and ph
background
To examine the role of lolal in pairing dependent
silencing we tested lolal/ þ in the context of homozygous
and heterozygous PRE-mw, transgenic lines carrying P
element with iab-7 PRE juxtaposed to mini-white reporter.
Briefly, lolal/CyO males were crossed to w;CyO,TM3,Sb/
Xasta virgins. From the progeny of this cross, we selected
lolal/CyO; þ /TM3 males and crossed them to w;pin/CyO,
TM6 virgins. Another cross consisting of PRE-mw
K. Mishra et al. / Mechanisms of Development 120 (2003) 681–689 687

insertions on the third chromosome and w;CyO,
TM3,Sb/Xasta virgins was set up. From these crosses we
collected lolal/CyO;TM3/TM6 males and virgin females and
crossed to w 2 ; þ /CyO; PRE/TM3 flies. From the progeny
of these crosses we selected lolal/CyO;PRE-mw/TM3 males
and virgin females for checking the pairing sensitivity of the
PRE in lolal background. For examining the PRE-mw in
polyhomeotic ( ph) background we took In (1) ph 410,
ph-p 410 w 1. From this stock, virgins were crossed to
w;CyO, TM3, Sb/Xasta males. From the progeny ph/Y; TM3,
Sb/Xasta males were crossed to ph-p homozygous virgins.
Another cross was set up where ph-p homozygous virgins
were crossed to PRE-mw (II and III chromosome lines)
males. From these crosses ph/ph;CyO/ þ virgins were
crossed to ph/Y;PRE-mw/ þ males, in the progeny
homozygous and heterozygous PRE-mw were observed in
the ph background. Six different PRE-mw lines were used
for these experiments. We also used colorimetric estimation
to quantitate the intensity of the eye color. This was
particularly useful in the case of those lolal experiments
where lolal allele was a P-element insertion mutation with a
light eye color of its own.
Acknowledgements
We are thankful to Robert Kingston for the FLAG-PH fly
strain, Laurent Theodore for lolal (ban) deletion mutations,
Vincenzo Pirrotta and Carl Wu for the Trl-GAGA
antibodies, Renato Paro for the Pc antibody, Veena Parnaik
for Lamin B antibody and David Shore for yeast strains and
plasmids. We thank Mohammad Idris, Ch. Sabarinadh, R.U.
Pathak, T. Jayashree and S. Subramanian for help. We also
thank Vincenzo Pirrotta and Francois Karch for critical
reading of the manuscript. This work was supported by a
‘Young Investigator Grant’ from Human Frontier Science
Program to R.K.M. (RGY0316/2001-M) and a research
fellowship to V.S.C. by the Council of Scientific and
Industrial Research, India.
References
Americo, J., Whiteley, M., Brown, J.L., Fujioka, M., Jaynes, J.B., Kassis, J.,
2002. A complex array of DNA-binding proteins required for pairing-
sensitive silencing by a Polycomb group response element from the
Drosophila engrailed gene. Genetics 160, 1561–1571.
Ashburner, M., 1989. Drosophila: A Laboratory Manual, Cold Spring
Harbor Laboratory Press, Cold Spring Harbor, NY.
Bardwell, V., Treisman, R., 1994. The POZ domain: A conserved protein–
protein interaction motif. Genes Dev. 8, 1664–1677.
Bender, W., Fitzgerald, D.P., 2002. Transcription activates repressed
domains in the Drosophila bithorax complex. Development. 129,
4923–4930.
Benyajati, C., Mueller, L., Xu, N., Pappano, M., Gao, J., Mosammaparast,
M., Conklin, D., Granok, H., Craig, C., Elgin, S., 1997. Multiple
isoforms of GAGA factor, a critical component of chromatin structure.
Nucleic Acids Res. 25, 3345–3353.
Biggins, M.D., Tjian, R., 1988. Transcription factors that activate the
Ultrabithorax promoter in developmentally staged extracts. Cell 53,
699–711.
Brown, J.L., Mucci, D., Whitely, M., Dirksen, M., Kassis, J.A., 1998. The
Drosophila Polycomb Group gene pleohomeotic encodes a DNA
binding protein with homology to the transcription factorYY1. Mol.
Cell 1, 1057–1064.
Busturia, A., Lloyd, A., Bejarano, F., Zavortink, M., Xin, H., Sakonju, S.,
2001. The MCP silencer of the Drosophila Abd-B gene requires both
Pleiohomeotic and GAGA factor for the maintenance of repression.
Development 128, 2163–2173.
Chan, C.S., Rastelli, L., Pirrotta, V., 1994. A Polycomb response element in
the Ubx gene that determines an epigenetically inherited state of
repression. EMBO J. 13, 2553–2564.
Czermin, B., Melfi, R., McCabe, D., Seitz, V., Imhof, A., Pirrotta, V., 2002.
Drosophila enhancer of Zeste/ESC complexes have a histone H3
methyltransferase activity that marks chromosomal Polycomb sites.
Cell 111, 185–196.
Espinas, M.L., Canudas, S., Fanti, L., Pimpinelli, S., Casanova, J., Azorin,
F., 2000. The GAGA factor of Drosophila interacts with SAP18, a
Sin3-associated polypeptide. EMBO Rep. 1, 253–259.
Farkas, G., Gauz, J., Galloni, M., Reuter, G., Gyrkovics, H., Karch, F.,
1994. The Trithorax-like gene encodes the Drosophila GAGA factor.
Nature 371, 806–808.
Faucheux, M., Netter, S., Bloyer, S., Moussa, M., Boissonneau, E.,
Lemeunier, F., Wegnez, M., Theodore, L., 2001. Advantages of a
P-element construct containing MtnA sequences for the identification of
patterning and cell determination genes in Drosophila melanogaster.
Mol. Genet. Genomics 265, 14–22.
Faucheux, M., Roignant, J.-Y., Netter, S., Charollais, J., Antoniewski, C.,
Theodore, L., 2003. batman interacts with Polycomb and trithorax
group genes and encodes a BTB/POZ protein that is included in a
complex containing GAGA factor. Mol. Cell. Biol. 23, 1181–1195.
Gietz, R.D., Schiestl, R.H., 1995. Transforming yeast with DNA. Methods
Mol. Cell. Biol. 5, 255–269.
Gindhart, J.G. Jr, Kaufman, T.C., 1995. Identification of Polycomb and
trithorax group responsive elements in the regulatory region of the
Drosophila homeotic gene Sex combs reduced. Genetics 139, 797–814.
Hagstrom, K., Muller, M., Schedl, P., 1997. A Polycomb and GAGA
dependent silencer adjoins the Fab-7 boundary in the Drosophila
bithorax complex. Genetics 146, 1365–1380.
Han, W., Yu, Y., Alton, N., Pick, L., 1993. Multiple proteins interact with
the fushi tarazu proximal enhancer. Mol. Cell. Biol. 13, 5549–5559.
Hodgson, J.W., Argiropoulos, B., Brock, H.W., 2001. Site-specific
recognition of a 70-base-pair element containing d(GA)n repeats
mediates bithoraxoid Polycomb group response element-dependent
silencing. Mol. Cell. Biol. 21, 4528–4543.
Hogga, I., Karch, F., 2002. Transcription through the iab-7 cis-regulatory
domain of the bithorax complex interferes with maintenance of
Polycomb-mediated silencing. Development 129, 4915–4922.
Horard, B., Tatout, C., Poux, S., Pirrotta, V., 2000. Structure of a Polycomb
response element and in vitro binding of Polycomb group complexes
containing GAGA factor. Mol. Cell. Biol. 20, 3187–3197.
Huan, H., Chang, Y., Yang, C., Pan, I., King, B., 2002. pipsqueak encodes a
factor essential for sequence-specific targeting of a Polycomb Group
Protein Complex. Mol. Cell. Biol. 22, 626–6271.
Jagatheesan, G., Thanumalayan, S., Muralikrishna, Bh., Rangaraj, N.,
Karande, A.A., Parnaik, V.K., 1999. Colocalization of intranuclear
lamin foci with RNA splicing factors. J. Cell Sci. 112, 4651–4661.
James, P., Halladay, J., Craig, E.A., 1996. Genomic libraries and a host
strain designed for highly efficient two-hybrid selection in yeast.
Genetics 144, 1425–1436.
Kassis, J.A., 1994. Unusual properties of regulatory DNA from the
Drosophila engrailed gene: three ‘pairing sensitive’ sites within a 1.6
kb region. Genetics 136, 1025–1038.
Kennison, J.A., 1993. Transcriptional activation of Drosophila homeotic
genes from distant regulatory elements. Trends Genet. 9, 75–79.
K. Mishra et al. / Mechanisms of Development 120 (2003) 681–689688

Lehmann, M., Siegmund, T., Lintermann, K., Korge, G., 1998. The
pipsqueak protein of Drosophila melanogaster binds to GAGA
sequences through a novel DNA-binding domain. J. Biol. Chem. 273,
28504–28509.
Mihaly, J., Mishra, R.K., Karch, F., 1998. A conserved sequence motif in
Polycomb-response elements. Mol. Cell 1, 1065–1066.
Mishra, R.K., Mihaly, J., Barges, S., Spirer, A., Karch, F., 2001. The iab-7
Polycomb Response Element maps to a nucleosome-free region of
chromatin and requires both GAGA and pleiohomeotic for silencing
activity. Mol. Cell. Biol. 21, 1311–1318.
Muller, J., Bienz, M., 1991. Long range repression conferring boundaries of
Ultrabithorax expression in the Drosophila embryo. EMBO J. 10,
3147–3155.
Muller, J., Hart, C.M., Francis, N.J., Vargas, M.L., Sengupta, A., Wild, B.,
Miller, E.L., O’Connor, M., Kingston, R.E., Simon, J.A., 2002. Histone
methyltransferase activity of a Drosophila Polycomb group repressor
complex. Cell 111, 197–208.
Orlando, V., Jane, E.P., Chinwalla, V., Harte, P.J., Paro, R., 1998. The
binding of Trithorax and Polycomb proteins to the bithorax complex:
dynamic changes during early Drosophila embryogenesis. EMBO J. 17,
5141–5150.
Pirrotta, V., 1997. Chromatin-silencing mechanisms in Drosophila
maintain patterns of gene expression. Trends Genet. 13, 314–318.
Pirrotta, V., 1998. Polycombing the genome: PcG, trxG, and chromatin
silencing. Cell 93, 333–336.
Poux, S., McCabe, D., Pirrotta, V., 2001a. Recruitment of components of
Polycomb Group chromatin complexes in Drosophila. Development
128, 75–85.
Poux, S., Melfi, R., Pirrotta, V., 2001b. Establishment of Polycomb
silencing requires a transient interaction between PC and ESC. Genes
Dev. 15, 2509–2514.
Rank, G., Prestel, M., Paro, R., 2002. Transcription through intergenic
chromosomal memory elements of the Drosophila bithorax complex
correlates with an epigenetic switch. Mol. Cell. Biol. 22, 8026–8034.
Read, D., Butte, M.J., Dernburg, A.F., Frasch, M., Kornberge, T.B., 2000.
Functional studies of the BTB domain in the Drosophila GAGA and
Mod(mdg4) proteins. Nucleic Acids Res. 28, 3864–3870.
Rose, M.D, Winston, F., Hieter, P., 1990. Methods in Yeast Genetics, Cold
Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
Sambrook, J., Russell, D.W., 2001. Molecular Cloning: A Laboratory
Manual, 3rd Edition, Cold Spring Harbor Laboratory Press, Cold Spring
Harbor, NY.
Sathe, S.S., Harte, P.J., 1995. The Drosophila extra sex combs protein
contains WD motifs essential for its function as a repressor of homeotic
genes. Mech. Dev. 52, 77–87.
Saurin, A.J., Shao, Z., Erdjument-Bromage, H., Tempst, P., Kingston, R.E.,
2001. A Drosophila Polycomb group complex includes Zeste and
dTAFII proteins. Nature 412, 655–660.
Shao, Z., Raible, F., Mollaaghababa, R., Guyon, J.R., Wu, C., Bender, W.,
Kingston, R.E., 1999. Stabilization of chromatin structure by PRC1, a
Polycomb complex. Cell 98, 37–46.
Simon, J., 1995. Locking in stable states of gene expression: transcriptional
control during Drosophila development. Curr. Opin. Cell Biol. 7,
376–385.
Simon, J., Chiang, A.C., Bender, W., Shimell, M.J., O’Connor, M., 1993.
Elements of the Drosophila bithorax complex that mediate repression
by Polycomb group products. Dev. Biol. 158, 131–144.
Simon, J., Bornemann, D., Lunde, K., Schwartz, C., 1995. The extra sex
combs product contains WD40 repeats and its time of action implies a
role distinct from other Polycomb group products. Mech. Dev. 53,
197–208.
Soeller, W.C., Oh, C.E., Kornberg, T.B., 1993. Isolation of cDNAs
encoding the Drosophila GAGA transcription factor. Mol. Cell. Biol.
13, 7961–7970.
Spradling, A.C., Stern, D., Beaton, A., Rhem, E.J., Laverty, T., Mozden, N.,
Misra, S., Rubin, G.M., 1999. The Berkeley Drosophila genome project
gene disruption project: single P-element insertions mutating 25% of
vital Drosophila genes. Genetics 153, 135–177.
Strutt, H., Cavalli, G., Paro, R., 1997. Co-localization of Polycomb protein
and GAGA factor on regulatory elements responsible for the
maintenance of homeotic gene expression. EMBO J. 16, 3621–3632.
Sun, X.H., Copeland, N.G., Jenkins, N.A., Baltimore, D., 1991. Id proteins
Id1 and Id2 selectively inhibit DNA binding by one class of helix-loop-
helix proteins. Mol. Cell. Biol. 11, 5603–5611.
Tillib, S., Petruk, S., Sedkov, Y., Kuzin, A., Fujioka, M., Gotto, T., Mazo,
A., 1999. Trithorax and Polycomb-group response elements within an
Ultrabithorax transcription unit consist of closely situated but separable
sequences. Mol. Cell. Biol. 19, 5189–5202.
Tsukiyama, T., Becker, P.B., Wu, C., 1994. ATP-dependent nucleosome
disruption at a heat-shock promoter mediated by binding of GAGA
transcription factor. Nature 367, 525–532.
K. Mishra et al. / Mechanisms of Development 120 (2003) 681–689 689


















