
Insecticide exposure impacts vector-parasite interactions in insecticide-resistant malaria vectors
Upload
eastangliaCategory
view
2download
0
Provided for non-commercial research and educational use only. Not for reproduction, distribution or commercial use.
This chapter was originally published in the book Advances in Genetics, Vol. 78, published by Elsevier, and the attached copy is provided by Elsevier for the author's benefit and for the benefit of the author's institution, for non-commercial research and educational use including without limitation use in instruction at your institution, sending it to specific colleagues who know you, and providing a copy to your institution’s administrator.
All other uses, reproduction and distribution, including without limitation commercial reprints, selling or licensing copies or access, or posting on open internet sites, your personal or institution’s website or repository, are prohibited. For exceptions, permission may be sought for such use through Elsevier's permissions site at:
http://www.elsevier.com/locate/permissionusematerial
From: Wayne G. Rostant, Nina Wedell, and David J. Hosken, Transposable Elements and Insecticide Resistance. In Stephen F. Goodwin, Theodore Friedmann and
Jay C. Dunlap editors: Advances in Genetics, Vol. 78, Burlington: Academic Press, 2012, pp. 169-201.
ISBN: 978-0-12-394394-1 © Copyright 2012 Elsevier Inc.
Academic Press

Author's personal copy
Advances inCopyright 2
2
Transposable Elements andInsecticide Resistance
Wayne G. Rostant, Nina Wedell, and David J. HoskenCentre for Ecology and Conservation, University of Exeter, Cornwall Campus,Tremough, Penryn, Cornwall, United Kingdom
I. I
Genet012, El
is
ntroduction
II. T ransposable ElementsA
.cs,ev
D
Vier
efinition and origin
B . C lassification of TEs C . T ransposition rates D . T he abundance and distribution of TEsIII. E
ffects of TEs on Host Fitness and Evolution A . T Es as selfish DNA B . N egative effects on hosts C . T E population dynamics D . N egating host fitness costs E. B eneficial effects of TEs F. C o-option/domesticationIV. I
nsecticide Resistance A . T he rate of evolution B . W hat is resistance? C . M echanisms D . C osts of resistance?V. T
Es Conferring Insecticide Resistance A . I nitial findings B . W hy are TEs so important? C . M echanisms of resistance via TEs D . T E-mediated DDT resistance in D. melanogasterand D. simulans
ol. 78 0065-2660/12 $35.00Inc. All rights reserved. http://dx.doi.org/10.1016/B978-0-12-394394-1.00002-X

170 Rostant et al.
Author's personal copy
VI. S
ex-Specific Effects of TEs Independent of DDT Resistance VII. O ngoing and Future ResearchA
cknowledgments R eferencesABSTRACT
Transposable elements (TEs) are mobile DNA sequences that are able to copythemselves within a host genome. They were initially characterized as selfishgenes because of documented or presumed costs to host fitness, but it has becomeincreasingly clear that not all TEs reduce host fitness. A good example of TEsbenefiting hosts is seen with insecticide resistance, where in a number of cases,TE insertions near specific genes confer resistance to these man-made products.This is particularly true of Accord and associated TEs in Drosophila melanogasterand Doc insertions in Drosophila simulans. The first of these insertions also hassexually antagonistic fitness effects in the absence of insecticides, and althoughthe magnitude of this effect depends on the genetic background in which Accordfinds itself, this represents an excellent example of intralocus sexual conflictwhere the precise allele involved is well characterized. We discuss this findingand the role of TEs in insecticide resistance. We also highlight areas for furtherresearch, including the need for surveys of the prevalence and fitness conse-quences of the Doc insertion and how Drosophila can be used as models toinvestigate resistance in pest species. � 2012, Elsevier Inc.
I. INTRODUCTION
The concept of an essentially stable genome, with each specific genetic elementconfined to a single locus was developed during the first few decades of lastcentury. This simple picture first came under serious challenge through the workof McClintock (1950, 1984) who, while analyzing chromosome breakage inmaize at Cornell University, first discovered what we now know as transposableelements (TEs). McClintock called these mobile elements “controllingelements,” a term which reveals her early assertion of their potential involve-ment in gene expression.
This view has turned out to be remarkably prescient. However, TEshave spent much of the time since their discovery under the monikers “junkDNA” and “selfish DNA,” revealing a general opinion that these mobilestretches of DNA played little if any part in evolution of their “hosts.” TEswere largely thought to have no influence on host genes and were interesting

2. TEs and Resistance 171
Author's personal copy
only insofar as their unique form of drive allowed them to invade host genomesand spread through populations. A recent review by Biemont (2010) gives a goodaccount of how prevailing views have come full circle to vindicate McClintock’sproposal that TEs are crucial components of genomes and drivers of theirevolution through their ability to affect gene expression. This journey fromjunk to critical agents of adaptive change has gathered pace as the sequencingof whole genomes has revealed the ubiquity and diversity of TEs.
There was an initial reluctance by many geneticists to accept that maizewas not an anomalous case. That so-called jumping genes might exist in othergenomes was difficult to reconcile with the fact that genetic maps had revealedremarkable homogeneity between individuals within species. The success ofmapping of genes to precise positions on a chromosome was incompatible withgenes moving around the genome. The isolation of bacterial TEs from Escherichiacoli (Shapiro, 1969) was the first step toward acceptance that TEs were a generalfeature of genomes.
In spite of these discoveries, it was not until the 1970s that skepticism overthe fundamental importance of TEs finally began to erode (Biemont and Vieira,2006). The reason for this was the emergence of hybrid dysgenesis—a phenomenonobservedwhen females of laboratoryDrosophilamelanogaster stockswerematedwithmales derived from natural populations. The progeny of these crosses displayedunusual germ line phenotypes including sterility, highmutation rate, and increasedfrequency of chromosomal aberration, while no such deficiencies exist in thereciprocal cross. The source of the dysgenesis turned out to be a TE called the Pelement which was present in wild strains but absent in laboratory strains.
Concurrent with a developing understanding of the ubiquity and impor-tance of TEs within genomes has been the increased use of pesticides to controlpest organisms, particularly from the 1950s onward (Wilson, 2001). This strongpervasive selection over many generations has provided the theoretical condi-tions under which adaptation by major genes might be favored, although earlymodels suggested that strong selection, while necessary, is probably insufficient tofavor major gene over polygenic adaptation (Lande, 1983). Nevertheless, theoverwhelming empirical evidence is that the evolution of pesticide resistance ismost often associated with the spread of a major mutation (Wilson, 2001), and ithas been suggested that it is not the strength of selection per se, but the amount ofphenotypic change required to achieve adaptation which determines the geneticarchitecture of the adaptive response (Macnair, 1991).
In light of these theoretical and empirical findings, it is perhaps unsur-prising that TEs are increasingly being implicated in the adaptive response oforganisms to man-made xenobiotics. In this review, we highlight the propertiesof TEs and insecticide resistance that make the former uniquely suited to thelatter adaptive response. While drawing on several putative and several well-documented examples of TE-mediated insecticide resistance from the literature,

172 Rostant et al.
Author's personal copy
we focus primarily on the striking cases of DDT resistance in D. melanogaster andDrosophila simulans which have been particularly well studied. In both instances,the resistance phenotype has been conferred by parallel insertions of TEs near acytochrome P450 gene.
II. TRANSPOSABLE ELEMENTS
A. Definition and origin
TEs, simply put, are DNA sequences that have the capacity to transpose. That is,they change their chromosomal location from one position to another within thesame genome, within a single cell (Hua-Van et al., 2011; Kidwell and Lisch,2001). They typically encode genes to promote this movement, in which casetranspositional ability is intrinsic. These TEs are said to be autonomous andcontrast with nonautonomous TEs which cannot transpose on their own, insteaddepending on the transposition machinery of other TEs (Hua-Van et al., 2011;Kidwell and Lisch, 2001; Wicker et al., 2007).
While questions concerning the origin and early evolution of TEs maynever be fully resolved, it does appear that their evolution has occurred primarilythrough the serial addition of domains, several of which seem likely to haveevolved from bacteria (Kidwell and Lisch, 2001). The question of a commonorigin for all TEs remains open (Wicker et al., 2007).
B. Classification of TEs
The first TE classification system was proposed by Finnegan (1989) and includedtwo main TE classes which were distinguished by their transposition intermedi-ate. Class I elements include those which transcribe via an RNA intermediateand, using a “copy-and-paste” mechanism, establish new copies of themselveselsewhere in a genome. Class II elements, in contrast, excise from donor sites andmove to new locations in a genome without use of an RNA intermediate, that is,they use a “cut-and-paste” method of transposition (Fig. 2.1).
To cope with an expanded array of TEs with diverse characteristics,Wicker et al. (2007) proposed a classification scheme that built on Finnegan’soriginal proposal (Table 2.1) by incorporating mechanistic and enzymatic cri-teria to the classification procedure. The original two classes were retained, andtwo subclasses within class II (DNA transposons) were formed to separate DNAtransposons which leave the donor site (excision) to reintegrate elsewhere(subclass 1) from those which copy themselves for insertion (subclass 2). Thenext hierarchical ranking (i.e., order) marks differences in the insertion mech-anism and thus organization and enzymology. Superfamilies within an order

Class I
Transcription
Reversetranscription
Excision
Insertion Insertion
RNA intermediate
Class II
Figure 2.1. A diagram of the transpositional modes of the two major transposable element classes,
class I and class II. Class I elements do not move once inserted into host DNA but use
RNA intermediates to insert additional TE copies in new genomic locations. Class II
elements can move by excision from host DNA, followed by reinsertion into a new
location.
Table 2.1. The Transposable Element Classification System Proposed by Wicker et al. (2007)
Class Order Superfamilies
Class I
(retrotransposons)
LTR (long-terminal repeat) Copia,Gypsy, Bel-Pao, Retrovirus, ERV
DIRS (Dictyostelium intermediate
repeat sequence)
DIRS, Ngaro, VIPER
PLE (Penelope-like elements) Penelope
LINE (long interspersed nuclear
element)
R2, RTE, Jockey, L1, I element
SINE (short interspersed nuclear
element)
tRNA, 7SL, 5S
Class II (DNA
transposons):
subclass 1
TIR (terminal inverted repeat) Tc1-Mariner, hAT, Mutator, Merlin,
Transib, P element, PiggyBac,
PIF-Harbinger, CACTA
Crypton Crypton
Class II (DNA
transposons):
subclass 2
Helitron Helitron
Maverick Maverick
2. TEs and Resistance 173
Author's personal copy
share a replication strategy but are distinguished by large-scale features such asthe structure of protein or noncoding regions. Families within superfamilies aredefined by DNA sequence conservation.

174 Rostant et al.
Author's personal copy
C. Transposition rates
The best data on TE transposition rates have come from laboratory experimentson D. melanogaster (Burt and Trivers, 2006). Rates are variable, ranging from2.9�10�6 per element per generation for P elements in inbred lines (Dominguezand Albornoz, 1996) to 0.25 per P element in dysgenic crosses. However, theyare typically low—estimates of 10�4 with the order of magnitude variation havebeen found for LINEs and LTR retrotransposons in two separate experiments(Maside et al., 2000; Nuzhdin and Mackay, 1995). That these estimates tend tobe higher than excision rates (of the order 10�6) implies that TEs should, ingeneral, accumulate in genomes over evolutionarily trivial timescales (Burt andTrivers, 2006). As a recent, well-cited example of this, P elements had beenshown to have invaded all known wild populations of D. melanogaster in thematter of about 50 years (Anxolabehere et al., 1988) after horizontal transferfrom Drosophila willistoni (Daniels et al., 1990).
D. The abundance and distribution of TEs
TEs have been discovered and characterized in most species that have beenadequately examined (Kidwell and Lisch, 2001). They are more ubiquitous ineukaryotes where they are present in virtually all species investigated to date,with few exceptions (Wicker et al., 2007). In prokaryotes, on the other hand,more than 20% of sequenced genomes lack TEs or their remnants (Touchon andRocha, 2007). TEs also tend to be more abundant in eukaryote genomes, makingup to 80% of the genome. For example, they comprise 60% of the maize genome(Messing and Dooner, 2006), 45% of the human genome (Cordaux and Batzer,2009; Lander et al., 2001), and 15% of the D. melanogaster genome (Dowsett andYoung, 1982), while in prokaryotes, they form only a maximum of 10% ofgenomes (Hua-Van et al., 2011).
Thomas (1971) famously coined the term “C-value paradox” to definethe then-curious lack of correlation between genome size (measured as DNAcontent or C-value) and the biological complexity of eukaryotes. Subsequently,it was found that, rather than correlating with gene content, genome size oftencorrelates with quantities of TE and TE-derived DNA. In fact, because theabundance of TEs within a genome can vary widely (Biemont and Vieira,2006), they, in addition to repetitive DNA, are major determinants of genomesize within taxa (Bennetzen, 2005). For example, the genome size of barley is 10times larger than that of rice (Argumuganathan and Earle, 1991), a related grasswith which it shows a great degree of synteny except that its genes are separatedby large clusters of retrotransposons.

2. TEs and Resistance 175
Author's personal copy
III. EFFECTS OF TES ON HOST FITNESS AND EVOLUTION
A. TEs as selfish DNA
Selfish genetic elements (SGEs) may be defined as stretches of DNA that actnarrowly to advance their own interests at the expense of the whole organism byensuring that a disproportionate fraction of offspring carry the DNA in question(Burt and Trivers, 2006). The concept of TEs as “selfish” or “parasitic” wascodified in seminal papers by Doolittle and Sapienza (1980) and Orgel andCrick (1980), but while the view of TEs as SGEs is now widely accepted(Werren, 2011), it perhaps obscures the continuum of interactions (from extremeparasitism to obligate mutualism) between host and TE (Kidwell and Lisch,2001) that often profoundly influence host genome evolution (Biemont, 2010;Biemont and Vieira, 2006; Feschotte, 2008; Hua-Van et al., 2011; Hurst andWerren, 2001; Kidwell and Lisch, 2001). Nevertheless, it is clear that the defaultview of TEs cannot be that they are simply functional parts of the genome.Brookfield (2005), in developing an analogy first made by Kidwell and Lisch(2001), describes the interaction of TEs and their hosts (and indeed between TEswithin a host) in terms of the “ecology of the genome.” He suggests thatquestions about TE numbers, diversity, and population dynamics within genomeshave ecological parallels with species in communities, and ecology thereforeprovides insights into the biology of TEs.
While most SGEs compete for representation at a single locus,TEs accumulate by copying themselves to new genomic locations and it isthis unique aspect of their drive that lies at the heart of their influence onhost fitness and evolution. Because TEs can transpose at a frequency (typically10�5 to 10�3 per element per generation) that is often much higher thanclassical nucleotide-base substitution rates (10�9 to 10�8), they are powerfulproducers of the raw material for evolution (Biemont and Vieira, 2006). Themutations caused by TE insertion and excision are also diverse, encompassinga broad spectrum from small-scale nucleotide changes to large chromosomalrearrangements (Hua-Van et al., 2011; Kidwell and Lisch, 2001) includingTE-mediated gene duplication (Jiang et al., 2004; Xiao et al., 2008; Yanget al., 2008). The combination of these two factors means that TEs may playan especially important role in evolution as the main source of spontaneousinternal mutations (Kidwell and Lisch, 2001; Li et al., 2007). For example, thehigh rate of new insertions of Alu and LINE-1 elements (Xing et al., 2009)means that TE insertions are a significant source of mutations in humans(Cordaux and Batzer, 2009). Additionally, 50–80% of mutations in Drosophilaare the result of TE insertions (Biemont and Vieira, 2006; Finnegan, 1992;Green, 1988).

176 Rostant et al.
Author's personal copy
B. Negative effects on hosts
As with other types of mutation, TE-induced changes will tend to be eitherharmful or neutral in their fitness effects on the host. TEs harm hosts in a numberof ways. Insertions may disrupt coding sequences or cis-regulatory regions, whilerecombination between TE copies can result in deletions and rearrangements.On top of this are the costs to the host of transcription and translation of largenumbers of TEs (Charlesworth et al., 1994; Kidwell and Lisch, 2001; O’Donnelland Burns, 2010). Fitness reductions have been quantified for P element trans-position in D. melanogaster (e.g., Currie et al., 1998; Fitzpatrick and Sved, 1986;Mackay, 1986, 1989; Mackay et al., 1992) where even nonlethal inserts tend toreduce host fitness by as much as 12.2% per insert when homozygous (Eanes et al.,1988; Mackay et al., 1992).
In most cases, highly deleterious insertions will be quickly removed byselection, but areas of the genome which experience low recombination might beexpected to accumulate insertions that have even moderately harmful effects.Y chromosomes and neo-Y chromosomes, where TE fixation rates tend to be muchhigher than on X chromosomes or autosomes, present such a case—since recom-bination is suppressed, selection is expected to be less effective due to hitchhikingand other effects (Charlesworth and Charlesworth, 2000). For example, TEs haveaccumulated at a very high abundance on the Drosophila miranda neo-Y chromo-some and might have been involved in causing a loss of gene activity (Steinemannand Steinemann, 1998).
C. TE population dynamics
While there is continuing debate as to which of the various sources of harm aremore important (Burt and Trivers, 2006), it is the interplay between selection forincreased replication at the TE level, but against deleterious host fitness effectsthat is responsible for TE population dynamics. Most of the deleterious pheno-typic effects of TEs will be removed from a population over time by purifyingselection (Kidwell and Lisch, 2001). Nonetheless, a population genetics modelhas shown that TEs can produce significant deleterious effects in the host andstill spread in the population (Hickey, 1982). Other models (e.g., Brookfield andBadge, 1997) highlight the importance of host population demography on TEcopy number. In these models, factors such as small host effective population size(Ne) attenuate the power of natural selection in regulating TE copy number.Empirical evidence suggests that these factors play an important part in TE copynumber and distribution in natural populations (Charlesworth andCharlesworth, 1995; Lockton and Gaut, 2010; Lockton et al., 2008). Lynchand Conery (2003) suggest that many aspects of complex genomes such as TEabundance were indirect consequences of reduced Ne, producing less effectiveselection against mildly deleterious insertions.

2. TEs and Resistance 177
Author's personal copy
While transposition rates tend to exceed excision rates, there is strongevidence that TE copy number is regulated. For example, the most abundant TEfamily still active inD. melanogaster is the retrotransposon roo, and there are only60 full-length copies per haploid genome in the euchromatin (Kaminker et al.,2002). Charlesworth and Charlesworth (2010) list five kinds of processes whichmay be involved in regulating TE abundance: (1) self- and/or host regulationof transposition rates, (2) selection against mutations, (3) ectopic exchange,(4) direct negative fitness effects of transposition on host fitness, and (5) indirecteffects of copy number on fitness. They conclude that while each of theseprocesses is plausible, and they are not mutually exclusive, the ectopic exchangemodel seems to be most consistent with current evidence (Charlesworth andCharlesworth, 2010; Charlesworth et al., 1997). Petrov et al. (2011) come tosimilar conclusions while examining the population frequencies of 755 TEs in sixD. melanogaster populations.
D. Negating host fitness costs
1. Cost minimization at the level of TEs
The fate of a TE in its host population thus depends not only on transposition ratebut also on host fitness effects, and TEs themselves should evolve to reduce hostharm (Burt and Trivers, 2006). Germ line specificity of transpositional activity, asdemonstrated in a number of class I (e.g., I elements and gypsy) and class II (e.g., Pelements and hobo) elements in Drosophila, is one such adaptation (Burt andTrivers, 2006) since transposition within the soma does not benefit the TE butdoes damage the host (Charlesworth and Langley, 1986).
Another damage-limiting strategy adopted by TEs is to insert preferen-tially into safe sites in the genome, as seen in Ty1, Ty2, Ty3, and Ty4 retro-transposons in baker’s yeast which target intergenic regions upstream of tRNAgenes (Kim et al., 1998). Additionally, there are many TEs which integrate intogene-rich regions, but which use mechanisms that prevent the disruption ofopen-reading frames (ORFs) (Levin and Moran, 2011). One example of this isseen in D. melanogaster P elements which tend to avoid disrupting ORFs byinserting within 500 bp upstream of host gene transcription start sites (Bellenet al., 2011). Other safe haven transpositions include insertion into other TEsand preferential insertion at or near telomeric chromosome ends. Examples ofthe latter include the HeTA, TART, and TAHRE non-LTR retrotransposonswhich comprise the ends of D. melanogaster chromosomes (Biessmann et al.,1992; George et al., 2010; Levis et al., 1993).
It has also been proposed that some TEs may have evolved autoregula-tion of transposition rate to avoid the deleterious effects of uncontrolled trans-position bursts (Burt and Trivers, 2006; Hua-Van et al., 2011). Theory suggests

178 Rostant et al.
Author's personal copy
that the circumstances under which such regulation would evolve are probablycommon, although unlikely to exist in unstructured random-mating hosts(Charlesworth and Langley, 1986, 1989). Nevertheless, there are exampleswhere self-regulation appears to be the case such as the P element-encodedrepressor which represses transposition and excision (Robertson and Engels, 1989).
2. Cost minimization at the level of the host: TE suppression
Hosts are not defenseless against harmful transposition. Many organisms haveevolved complex mechanisms to deal with TEs. Small RNA-based mechanismsact to defend eukaryotic cells against TEs by posttranscriptional disruption of TEmRNA (Aravin et al., 2007; Malone and Hannon, 2009; van Rij and Berezikov,2009). Another way in which some host taxa suppress their TEs is throughepigenetic control, including methylation. In fact, it is widely thought thatepigenetics, whose processes are commonly used by metazoans in cell lineage-specific gene regulation, first evolved to defend against foreign DNA includingTEs (Hua-Van et al., 2011). This is one example of how the prolonged interac-tion of host and TE has ultimately benefited the host—it is far from the only one.
E. Beneficial effects of TEs
As with any other source of mutation, TEs can occasionally produce beneficialgenetic alterations to host genomic DNA. A beneficial insertion would beexpected to go to fixation within a population, and TE fixation has beenobserved, particularly in D. melanogaster (Gonzalez and Petrov, 2009). The Selement(s) associated with the Hsp70 (heat-shock protein) genes in D. melano-gaster is one possible example of a beneficial TE (Maside et al., 2002). While thefunctional significance of this insertion has not been elucidated, there is strongevidence of a selective sweep around it. Furthermore, the insertion apparentlyoccurs in a freely recombining region of the genome, which substantially lowersthe probability of fixation via drift.
F. Co-option/domestication
In contrast to the benefits derived from genetic alteration of host genomicsequences per se, TE sequences themselves may be co-opted for host function, aprocess which has been called “domestication” or “exaptation” when TE-codingsequence function has been appropriated for host use. There are several examplesof this in the literature, one of the most cited being the full domestication ofthe Drosophila telomeric retrotransposons HeTA and TART which function astelomerase to heal chromosome ends. Even noncoding TE sequences may be

2. TEs and Resistance 179
Author's personal copy
useful—one striking example of the fixation of a beneficial insertion which is ofparticular importance to this review is found in the evolution of DDT resistancein D. melanogaster. Here, an Accord retrotransposon-derived sequence insertedupstream of a cytochrome P450 gene has been shown to upregulate the detox-ification enzyme and increase pesticide resistance (Chung et al., 2007; Dabornet al., 2002). The remainder of the review focuses on insecticide resistance andhow TEs influence this.
IV. INSECTICIDE RESISTANCE
A. The rate of evolution
Given the evolutionary potential of TEs, perhaps it is not surprising that they playan important role in such key fitness traits as pesticide resistance. Over the past100 years, there has been an increased use of toxic chemicals to control pestorganisms, particularly from the 1950s onward (Wilson, 2001). This strong,pervasive source of selection has demonstrated the tremendous capacity of popula-tions to evolve resistance to toxins. Since the first insecticide resistance case wasreported almost a century ago (Melander, 1914), there have been thousands ofcases of resistance in hundreds of species (Georghiou and Lagunes-Tejeda, 1991;Whalon et al., 2008). Some of the most dramatic examples of microevolution inaction have come from selection for chemical resistance (Hartl and Clark, 1997),with resistance evolving in as few as 5–50 generations (May, 1985) and towardrapid global fixation in many insect pest populations (Catania et al., 2004;Schlenke and Begun, 2004; Whalon et al., 2008).
B. What is resistance?
From a functional point of view, insecticide resistance may be defined as theability of an organism to survive a dose of insecticide that is lethal to a suscep-tible one (Georghiou and Saito, 1983), and dynamically, it has also beendescribed as the microevolutionary process whereby genetic adaptation throughpesticide selection results in populations of susceptible insects being replaced byresistant ones over a period of time (Wilson, 2001). The biochemical mecha-nisms and molecular genetics underlying resistance have been well studied andhave been the subject of several books (Clark and Yamaguchi, 2002; Denholmet al., 1999; Ishaaya, 2001) and reviews (Feyereisen, 1995; ffrench-Constant et al.,2004; Oakeshott et al., 2003).
The proximate biochemical mechanisms of resistance can be divided intofour main categories (Wilson, 2001). The first of these is behavioral resistance (i.e.,avoidance of the insecticide), which may involve genetic changes, but is probably

180 Rostant et al.
Author's personal copy
of minor importance even though it has been documented for a few species (Sparkset al., 1989). Reduction in the penetrative ability of the toxin is a second mecha-nism, but again this does not seem to be of major importance (Wilson, 2001).Target-site inactivation (changes in the insecticides site of action) is a veryimportant biochemical resistance mechanism (Hollingworth and Dong, 2008;Wilson, 2001). Every potent insecticide has one or more specific binding sites oncritical macromolecules, and changes in the ability of the toxin to bind must affectits impact on the insect (Hollingworth andDong, 2008). Lastly, biotransformation,the metabolic breakdown of a toxin, is a common defense against natural xenobi-otics (Li et al., 2007). It is therefore not surprising that, with the widespread use ofsynthetic organic agricultural chemicals, the enzymatic systems which originallyevolved to detoxify phytotoxins should been enlisted to defend against insecticides(Wilson, 2001). Three types of enzymes—esterases (through ester hydrolysis),cytochrome P450 monoxygenases (through oxidation), and glutathione transfer-ases (through ester hydrolysis)—are commonly used to transform insecticides intoless toxic products (Hollingworth and Dong, 2008).
When an insecticide is first introduced, the target population largelyconsists of susceptible phenotypes (Macnair, 1991; Mallet, 1989; McKenzie andBatterham, 1994; Roush andMcKenzie, 1987).Within the population, there willbe a distribution of susceptibility based on factors such as size, age, and physio-logical condition (McKenzie and Batterham, 1994), which are generally poly-genically inherited. Insecticide selection on this distribution will act via thephenotype and resistance will be polygenically inherited, combining preexistingfactors of primarily minor effect (such as size and developmental rate) (ffrench-Constant et al., 2004). This type of selection is seen in most laboratory studies(ffrench-Constant et al., 2004; McKenzie and Batterham, 1994), which explainswhy early studies of DDT resistance (e.g., Crow, 1957) determined that resis-tance evolution was a polygenic response.
This contrasts strongly with what has been found in natural popula-tions, where resistance to particular insecticides often involves one or two majorgenes (ffrench-Constant et al., 2004; Field et al., 1988; Mallet, 1989; McKenzieand Batterham, 1994; Raymond et al., 1989; Roush and McKenzie, 1987). Thismay represent detection bias, but another explanation could be that insecticidesin the field tend to occur at concentrations which favor variation outside of thenormal phenotypic distribution (i.e., rare resistant mutations of major effect).Natural populations are much larger than laboratory populations and so morelikely to contain individuals with these rare mutations. A second reason for thepreponderance of monogenic resistance in the wild may be evolutionary con-straint resulting from opposing natural selection on multiple targets. The natureof the ultimate genetic changes which lead to monogenic resistance also varieswith respect to the proximate biochemical mechanism involved (Wilson, 2001).

2. TEs and Resistance 181
Author's personal copy
C. Mechanisms
Target-site inactivation is usually effected by subtle changes in the targetprotein—it is therefore easy to understand the importance of point mutationsfor this resistance mechanism (Wilson, 2001). An altered protein must retainat least some degree of normal function while decreasing its xenobiotic sensi-tivity, which explains the highly conserved nature of such changes (ffrench-Constant, 1999; ffrench-Constant et al., 1998; Li et al., 2007; Wilson, 2001).A striking illustration of this is the parallel evolution of cyclodiene resistancein a wide range of pest species and in Drosophila, which is a result of the samesingle amino acid substitution in the chloride ion channel pore of the gamma-aminobutyric acid receptor protein (ffrench-Constant et al., 1998; Thompsonet al., 1993).
Metabolic resistance, on the other hand, tends to involve the over-expression of existing metabolic enzymes either through gene amplification(i.e., gene duplication, which results in more gene product) or alterations intheir regulatory systems, which increase transcription and/or stabilize mRNA(Hollingworth and Dong, 2008; Li et al., 2007; Wilson, 2001). Examples ofresistance through gene copy increase are seen for esterase genes in mosqui-toes and aphids, GSTs in the housefly and the aphid Nilaparvata lugens andcytochrome P450s in three dipterans including D. melanogaster and D. simu-lans and the potato aphid Myzus persicae (reviewed in Bass and Field, 2011;Devonshire and Field, 1991). A particularly striking example is provided byresistant Culex pipiens quinquefasciatus mosquitoes, where the esterase geneB1 is amplified in a tandem array as much as 250-fold, conferring highorganophosphate (OP) resistance (Karunaratne et al., 1993; Mouches et al.,1986, 1990).
Gene upregulation is the most common process involved in P450-mediated insecticide resistance, but upregulation has also been documented forthe other two major classes of detoxification enzymes already mentioned(Li et al., 2007). This is usually achieved through changes (point mutations orindels) in either cis- or trans-regulatory loci. An example of the former isprovided by the P450 Cyp6g1 gene in D. melanogaster where the insertion of adefective copy of the Gypsy-like LTR retrotransposon Accord in the 50 promoterregion results in upregulation of the enzyme and cross-resistance to DDT,imidacloprid, nitenpyram, and lufenuron (Catania et al., 2004; Daborn et al.,2002; Schlenke and Begun, 2004). As an example of the latter, overexpression ofa GST allele in the resistant Aedes egypti GG strain is due largely to a loss-of-function mutation in an unidentified trans-acting repressor that represses mRNAtranscription and/or decreases mRNA stability in the susceptible strains (Grantand Hammock, 1992).

182 Rostant et al.
Author's personal copy
D. Costs of resistance?
A central question in the evolution of resistance is the fitness of the organismcarrying a mutant allele of a resistance gene. Theory holds that, in the absence ofinsecticide, the majority of insecticide-resistant organisms should show somedifferential survival in comparison with “wild-type” organisms. That is, resis-tance should be costly (e.g., Crow, 1957). However, empirical evidence on thepleiotropic fitness effects of insecticide resistance appears to be equivocal. Thereare a few empirical studies that confirm that investment in resistance entails afitness cost (Alyokhin and Ferro, 1999; Berticat et al., 2002; Boivin et al., 2001;Carriere et al., 1994, 1995, 2001; Chevillon et al., 1997; Foster et al., 2003;Minkoff and Wilson, 1992; Rivero et al., 2011; Smith et al., 2011; Yamamotoet al., 1995). On the other hand, some authors have failed to reveal anydetrimental effects of insecticide resistance (Baker et al., 1998, 2008;Castaneda et al., 2011; Follett et al., 1993; Tang et al., 1997, 1999), and somehave demonstrated pleiotropic fitness benefits (Arnaud and Haubruge, 2002;Bielza et al., 2008; Bloch and Wool, 1994; Haubruge and Arnaud, 2001;Mason, 1998; McCart et al., 2005; Omer et al., 1992; White and Bell, 1995).
In other studies, some measures of fitness have been negatively affected,others positively (Brewer and Trumble, 1991), and this may even involve sexualantagonism, where resistance alleles have opposing fitness effects depending onwhich sex they reside. This has recently been documented for DDT resistance inD. melanogaster, where resistance confers a strong fecundity advantage to females,but a competitive mating disadvantage to males (McCart et al., 2005; Smithet al., 2011). In addition, how resistance alleles impact nonresistance-relatedfitness can depend on the strain being investigated (Chevillon et al., 1997;Hollingsworth et al., 1997; Oppert et al., 2000; Smith et al., 2011). This reflectsepistasis, where the pleiotropic fitness effect is mediated by the genotype(or genetic background) of the insect in question.
V. TES CONFERRING INSECTICIDE RESISTANCE
A. Initial findings
Wilson (1993) was the first to speculate that TEs were implicated in insecticideresistance, although the evidence was indirect—he was able to generateMethoprene-resistant alleles in D. melanogaster using P element mutagenesis(Wilson and Turner, 1992). Around the same time, Waters et al. (1992) foundan association between Drosophila strains resistant to DDT and Malathion and a17.6 TE insertion in the 30 region of a cytochrome P450 enzyme gene. In this

2. TEs and Resistance 183
Author's personal copy
case, it was found that the resistant strains lacked the insertion, suggesting thatresistance was a result of an excision of the TE. However, Delpuech et al. (1993)subsequently reported that the presence or absence of the 17.6 LTR wasuncorrelated with resistance in 31 strains of D. melanogaster and D. simulans.
Wilson (2001) was less convinced about the possibility that TEs play asignificant role in insecticide resistance in nature (notwithstanding the Pelement-induced resistance, he demonstrated in the laboratory a decade earlier)conceding that, at most, “TE mutagenesis may be important only for a few geneswhere resistance can result from severe underexpression or nonfunctional geneproduct.” However, since his review, evidence has been steadily accumulatingthat TEs do, in fact, play an important part in the evolution of insecticideresistance.
The observation that TEs are frequently found within or in closeproximity to resistance genes provides indirect evidence that TEs are involvedin resistance-related adaptive genomic changes (Li et al., 2007). This inferencewas bolstered by the findings of Chen and Li (2007) who reported that TEinsertions were enriched around and within xenobiotic-metabolizing P450genes of both Helicoverpa zea moths and D. melanogaster flies. They also foundthat TE insertions were absent from essential housekeeping P450 genes inD. melanogaster, which might be expected since mutation of essential genes ismore likely to be lethal and not simply reduce fitness. Taken together, theseresults indicate that TEs are also selectively retained within or in close proximityto xenobiotic-metabolizing P450 genes. Similarly, while a Bari-1 element inser-tion occurs downstream of the P450 gene Cyp12a4 in an Australian lufenuron-resistant D. melanogaster strain, its presence in lufenuron-susceptible strainssuggests that while the insertion may be important, it is not the main cause ofresistance (Bogwitz et al., 2005).
Recent studies provide more conclusive, direct evidence for a causativelink between resistance and TEs. For example, insertion of a 2.3-kb LTR retro-transposon Hel-1 in the putative Bt-toxin receptor gene cadherin leads to30-truncated nonfunctional cadherin protein and Bt resistance in a laboratory-selected Heliothis virescens strain (Gahan et al., 2001). Furthermore, parallelinsertions of Accord-LTR or Doc non-LTR retrotransposon into the 50-regulatoryregion of Cyp6g1 in D. melanogaster or D. simulans are associated with Cyp6g1upregulation and DDT resistance (Daborn et al., 2002; Schlenke and Begun,2004) (Section IV.D). Additionally, in D. melanogaster, insertion of a Doc1420retrotransposon into the second exon of the predicted gene CG10618 (CHKov1,a putative choline kinase gene) generates two sets of altered transcripts and anovel polypeptide (Aminetzach et al., 2005). Whether through loss of originalCHKov1 function or through function of the new protein, the Doc1420 insertionconfers moderate OP resistance (Aminetzach et al., 2005).

184 Rostant et al.
Author's personal copy
B. Why are TEs so important?
Insecticide resistance results from very strong, persistent directional selection.TE-mediated changes in regulation can lead to massive and rapid changes inexpression, responses that are potentially highly adaptive when an organism isfaced with a major, pervasive, and novel mortality agent in the environment, likean insecticide. A useful contrast which illustrates this point is the essentialabsence of TEs involved in natural xenobiotic resistance—if we consider thatmutational changes in plant allelochemicals are unlikely to bring about massivechanges in mode of action or in toxicity, then mutational change associatedwith allelochemical resistance may be acquired more slowly as a result of theaccumulation of small changes in structural genes (Li et al., 2007).
Application of insecticide tends to favor insecticide resistance, involvingsingle genes of major effect rather than polygenic resistance (ffrench-Constantet al., 2004), and it has been found that most resistant field strains showmonogenicresistance (Roush and McKenzie, 1987). Where resistance genes are alreadyinvolved in essential functions, as is often the case for metabolic enzymes, it isadvantageous to maintain the quality of mRNA to allow wild-type function tobe retained and instead regulate gene expression. TE insertion within regulatoryregions of genes which confer resistance often results in upregulation, that is,increase in the quantity of mRNA. This may be because many TEs have built-inenhancer sequences related to their transposition (Zhang and Saier, 2009)that have been co-opted by the host, but another possibility is that such spacingmay move genes further from existing regulatory sequences (Schlenke andBegun, 2004).
C. Mechanisms of resistance via TEs
ffrench-Constant et al. (2006) list four possible mechanisms whereby TE in-sertions might confer insecticide resistance. First, a TE insertion in the 50-end ofa gene may introduce a novel enhancer sequence. The Accord-LTR upstream ofthe cytochrome P450 gene Cyp6g1 in D. melanogaster is one such case (Chunget al., 2007), and the Cyp6g1 homolog in D. simulans—where the insertion is acomplete Doc element—may also be one (Schlenke and Begun, 2004). Anotherexample of this mechanism is found in the mosquitoCulex quinquefasciatus wherethe insertion of a miniature-inverted terminal repeat (MITE)-like elementupstream of another cytochrome P450 gene is associated with increasedpyrethroid resistance (Itokawa et al., 2010). The second mechanism involvesincreased mRNA stability via TE insertion in the 30-end of a gene whichincreases the final pool of translatable RNA. Third, TEs might excise a geneand move it to a different genomic location away from local repressor elementsnormally responsible for shutting off expression or to a position proximal to an

2. TEs and Resistance 185
Author's personal copy
enhancer element. This position effect was demonstrated in principle by Berradaand Fournier (1997) who used P element-mediated transposition to initiatetranscriptional overexpression of an artificially constructed acetylcholinesteraseminigene in D. melanogaster. Finally, TE insertion might alter the patternof resulting transcripts and potentially lead to a truncated gene product ofnovel function as appears to be the cases described by Gahan et al. (2001)and Aminetzach et al. (2005). One further mechanism not mentioned byffrench-Constant et al. (2006) involves gene amplification. The transpositionalmechanism of TEs may result in gene duplication through ectopic recombination(e.g., Yang et al., 2008) and consequent increase in gene product, or evolution ofnew gene function in the duplicated gene.
D. TE-mediated DDT resistance in D. melanogaster and D. simulans
Fifty years ago, studies in D. melanogaster indicated that many genes contributedto DDT resistance (Crow, 1957; Dapkus and Merrell, 1977; Hallstrom, 1985;Kikkawa, 1961) including loci on all three major chromosomes. However, workon the Hikone-R strain indicated that resistance in this strain was largelyconferred by a single dominant locus on chromosome II, and this was laterfound to be Cyp6g1 (Daborn et al., 2001). Resistant alleles were found to havea defective copy of an Accord-LTR retrotransposon inserted about 300 bpupstream of the transcription start site. Subsequently, the molecular mechanismof upregulation was identified (Chung et al., 2007) with cis-regulatory sequencesin the Accord-LTR being responsible for increased Cyp6g1 transcription.
Schlenke and Begun (2004), while investigating reduced heterozygosityaround the Cyp6g1 locus in D. melanogaster and D. simulans, found that anotherTE insertion, this time a full-length copy of the non-LTR retrotransposon Doc,occurred 200 bp upstream of the gene in Californian populations of the latterspecies. Once again, the insertion correlated with increased Cyp6g1 expressioncompared with that found in African populations lacking the insertion.In contrast to the Accord insertion in D. melanogaster which is highly degenerate(comprising only the LTR), the Doc insertion in D. simulans is of an autonomouselement, suggesting that it is a much more recent event. Selective sweeps atCyp6g1, associated with strong recent selection, were demonstrated in bothspecies (Catania et al., 2004; Schlenke and Begun, 2004).
Catania et al. (2004) conducted a survey of the Accord-LTR insertion atCyp6g1 in 673 lines from 34 populations from around the world. They found nearfixation of the Accord-LTR-inserted alleles in non-African and North andWestern African D. melanogaster populations (85–100% of chromosomessampled), with significantly lower frequencies in East African populations(32–55%). Variation in the Accord-LTR-inserted allele was also found—diagnostic PCR revealed some variability in product size, and subsequent cloning

186 Rostant et al.
Author's personal copy
and sequencing of the variants revealed an insertion of a partial P element nestedwithin the Accord-LTR in a New Delhi line (Catania et al., 2004). Furthermore,Emerson et al. (2008), using genome-wide tilling arrays, found copy numberpolymorphism in D. melanogaster at Cyp6g1, with 13 of 15 lines tested showinga duplication encompassing both Cyp6g1 and Cyp6g2.
Most recently, Schmidt et al. (2010) characterized copy number varia-tion and further allelic variation at theCyp6g1 locus. Characterization ofCyp6g1copy number variation and TE insertion complexity in the D. melanogasterRK146 strain revealed two full-length copies of Cyp6g1, named Cyp6g1-a andCyp6g1-b. A repeat unit was found between the two full-length copies thatcontained a fusion of partial copies of both Cyp6g1 and Cyp6g2, the genefound downstream of Cyp6g1-b. Both copies of Cyp6g1 were found to containthe LTR of the Accord element. Unexpectedly, a HMS-Beagle TE was foundinserted into the Accord-LTR upstream of Cyp6g1-a. Testing of other D. mela-nogaster lines revealed that the partial P element found by Catania et al. (2004)was located upstream of Cyp6g1-b.
Schmidt et al. (2010) also demonstrated an allelic progression of fivedifferent alleles, including the ancestral allele lacking any TE insertions andfour alleles that involve duplication of Cyp6g1 (Fig. 2.2). They reason thatthese alleles represent multiple adaptive steps at Cyp6g1, with increased DDTresistance being demonstrated along the allelic progression. Their survey ofD. melanogaster global populations showed that most flies in Europe, Asia, andthe United States carry the Cyp6g1 duplication and the double Accord-LTR (noP element) insertion or the Cyp6g1 duplication and combined Accord-LTRinsertion/Accord-LTR (with nested HMS-Beagle) insertion.
The question of the age of the original Accord-inserted allele remainsopen. In their survey, Schmidt et al. (2010) did not find any Cyp6g1 alleles withthe insertion that did not also represent a duplication, nor did they find any geneduplicates which did not also contain theAccord insertion. This strongly suggeststhat either the original insertion and duplication events occurred simultaneous-ly, or, more likely, the original Accord insertion preceded the duplication. If thelatter, then we are yet to find the original Accord-inserted allele.
Catania et al. (2004) suggested that the lower than expected reductionin variability in microsatellites around the Cyp6g1 locus could be explained mostparsimoniously if the Accord-LTR insertion occurred at low frequency in Africanpopulations before the species’ global expansion. This would imply that theinsertion was already part of the genetic variation at this locus well before itpermitted adaptation to insecticide and may be an example of how TEs providelatent genetic variation facilitating adaptive responses to selection.
The absence of strong DDT selection since its ban in most countriesglobally over the past 30 years has not brought about the loss of DDT-R alleles inD. melanogaster, as might be expected from theory—overexpression of P450

Ancestralallele
Accord LTR
HMS-Beagle
P elementA
A A
A
A
A
A
A
Be. P
P
Be.
Be.
Cyp6g1-a
Cyp6g1-a
Cyp6g1-a Cyp6g1-b
Cyp6g1-b
Cyp6g1-b
Incr
easi
ng D
DT
res
ista
nce
A
B
C
D
M
A?
AA
BA
BP
Figure 2.2. Allelic progression at the Cyp6g1 locus in D. melanogaster as described by Schmidt et al.
(2010). The ancestral allele (Cyp6g1-[M]) is found in DDT-susceptible strains such as
Canton-S and is the only allele found in laboratory lines established in the 1930s. The
most plausible sequence of changes to the ancestral allele are as follows: (A) Insertion of
an Accord retrotransposon 50 of Cyp6g1, followed by excision (leaving only the LTR
footprint), produces hypothetical allele Cyp6g1-[A?]. (B) Duplication of the Accord-
LTR-inserted allele produces a tandem repeat of two full-length Cyp6g1 copies (Cyp6g1-
a and Cyp6g1-b) separated by partial Cyp6g1/Cyp6g2 repeat units and resulting in the
DDT-resistant allele Cyp6g1-[AA]. (C) Insertion of anHMS-Beagle retrotransposon into
the Accord-LTR found in Cyp6g1-a of Cyp6g1-[AA] produces the DDT-resistant allele
Cyp6g1-[BA]. (D) Insertion of a P-element DNA transposon into the Accord-LTR found
in Cyp6g1-b of Cyp6g1-[BA] produces the DDT-resistant allele Cyp6g1-[BP].
2. TEs and Resistance 187
Author's personal copy
genes must have a cost, and if the resistance-to-DDT benefit to balance this costis not there, then selection should remove the resistance allele. The near fixationof Accord-inserted Cyp6g1 alleles in worldwide populations (Catania et al., 2004)and the seemingly adaptive, ongoing elaboration of these alleles through subse-quent TE insertion increasing DDT resistance (Schmidt et al., 2010) thus poses apuzzle. The most obvious explanation is that these alleles confer cross-resistanceto other pesticides and therefore remain under strong xenobiotic selection.There are other explanations, however.

188 Rostant et al.
Author's personal copy
VI. SEX-SPECIFIC EFFECTS OF TES INDEPENDENT OF DDTRESISTANCE
The strong fitness benefit conferred by an Accord-inserted Cyp6g1 allele toCanton-S strain females and pupae in the absence of pesticide (McCart et al.,2005) offers another explanation for the persistence of these resistance alleles athigh frequencies in the wild. However, this nonresistance fitness benefit shouldnot be viewed in isolation—Smith et al. (2011) very recently confirmed acompetitive mating disadvantage to males carrying the same allele and in thesame genetic background as used by McCart et al. (2005). This male fitness costalmost perfectly balances the benefit conferred to females. The presence of aDDT-R male fitness disadvantage was first suggested by Drnevich et al. (2004),who in breedingD. melanogastermales of high and lowMCRS (male competitivereproductive success) discovered that high MCRS was associated with lowexpression of Cyp6g1.
This apparent sexually antagonistic selection at Cyp6g1 is an excellentexample of intralocus sexual conflict where the precise locus has been identified.Intralocus sexual conflict occurs when an allele has positive fitness effects in onesex and negative in the other sex (Bonduriansky and Chenoweth, 2009; Hoskenet al., 2009). Sexually antagonistic selection generally helps maintain geneticvariation and has important implications for the history of the DDT-R allele.If the original Accord-inserted allele antedates insecticide use, as Catania et al.(2004) posit, sexually antagonistic selection may have accounted for its lowfrequency in populations prior to DDT’s introduction—no Accord-insertedCyp6g1 alleles have been found from lines established in the 1930s (Schmidtet al., 2010).
It should be noted that the male fitness disadvantage of the Accord-inserted allele is not consistent across different genetic backgrounds. When theallele was introgressed into another D. melanogaster strain, males were stillcompetitively disadvantaged, but not significantly so (Smith et al., 2011). Asyet, the fitness effects of the Accord insertion in females of the latter geneticbackground have not been examined. This latter strain had been established byTrudy MacKay in North Carolina, USA, from wild-caught flies in 2004 andcontrasts with the Canton-S background which represents one of the earliestestablished laboratory populations (1930s). The lack of a significant male disad-vantage in the more recently established strains may thus represent the evolutionof male-specific modifiers in populations dominated by resistant Cyp6g1 alleles(although this could also represent a statistical power issue). In this context, it ispossible that DDT (and other insecticide) selection may have prompted not onlyrapid spread of the resistance allele but also male-specific counter-adaptation toreduce pleiotropic fitness costs of resistance. Interestingly, while homozygousresistant Canton-S males were significantly smaller than their susceptible

2. TEs and Resistance 189
Author's personal copy
counterparts, the converse was true for males of the wild-type genetic back-ground (Smith et al., 2011), hinting that amelioration of the male fitness costmay involve loci influencing male body size.
Cohan et al. (1994) list two general ways in which pleiotropic fitnesscosts of an adaptive mutation may be ameliorated. The epistatic male DDT-Rcost effect seen by Smith et al. (2011) may be an example of the “compensatory”mode, in which natural selection favors modifiers (at other loci) that compensatefor the deleterious effects of the mutant allele (Fisher, 1928). The other mode,known as “replacement,” describes the case where there are multiple mutationswhich confer the same adaptation, but which vary in their pleiotropic fitnesscosts such that the original mutation is replaced by one which confers the sameadaptive benefit at lower cost (Haldane, 1932). Interestingly, this mode has beeninvoked to explain temporal allele replacement observed in the insecticideresistance gene Ester in the mosquito C. pipiens in southern France (Labbeet al., 2009). A similar scenario may exist in the Cyp6g1 allelic progressiondescribed by Schmidt et al. (2010).
VII. ONGOING AND FUTURE RESEARCH
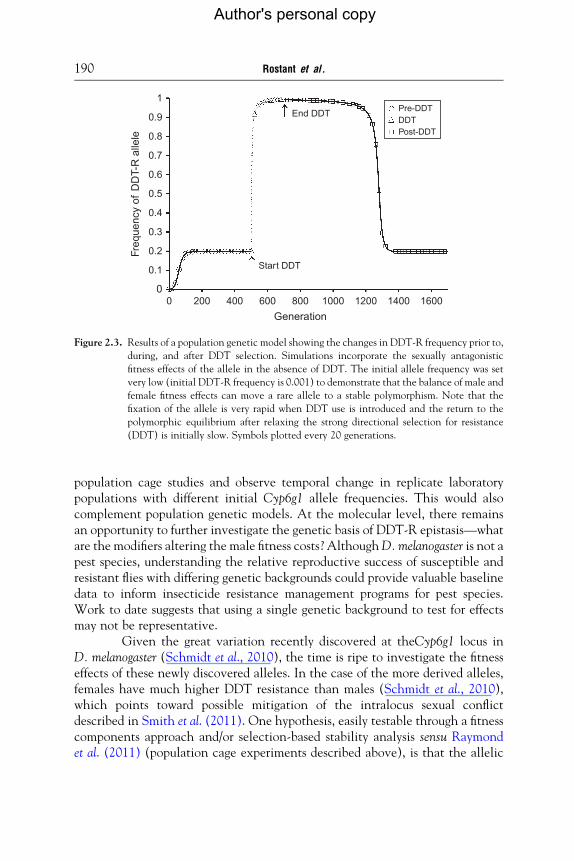
In spite of observed female fitness benefits in the absence of DDT (McCart et al.,2005), recent simple models (our unpublished data) demonstrate that an Accord-inserted Cyp6g1 (i.e., DDT-R) allele could have been kept at low frequency inpopulations before DDT was introduced, through male-associated fitness costs(Smith et al., 2011). The net effect of male and female pleiotropic fitnesscomponents, with parameter values based on empirical work by Daborn et al.(2001), McCart et al. (2005), and Smith et al. (2011), was a very slow return to astable polymorphism at the Cyp6g1 locus (Fig. 2.3). These results are thereforeconsistent with continued high levels of DDT-R several years after the removalof DDT selection, a phenomenon which has been previously explained byinvoking cross-resistance, a lack of fitness cost and low migratory rates(Catania et al., 2004; McCart et al., 2005).
Further work is required to determine the range of fitness effects ofDDT-R in different genetic backgrounds and to further explore the populationgenetics of these alleles. We also need to investigate the underlying reasons forthe competitive disadvantage observed in resistant males of Canton-S back-ground—is it simply size mediated or are there other mechanisms (behavioraland/or physiological) which affect male fitness? Related to this, the demonstra-tion of epistasis with regard to male fitness effects begs a more thorough inves-tigation of the distribution of pleiotropic fitness effects in both sexes. Howcommon is the DDT-R-associated male competitive disadvantage? How com-mon is the female fitness benefit? In this respect, it would be useful to run
Start DDT
End DDT
Generation
00
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
200 400 600 800 1000 1200 1400 1600
Pre-DDTDDTPost-DDT
Freq
uenc
y of
DD
T-R
alle
le
Figure 2.3. Results of a population genetic model showing the changes in DDT-R frequency prior to,
during, and after DDT selection. Simulations incorporate the sexually antagonistic
fitness effects of the allele in the absence of DDT. The initial allele frequency was set
very low (initial DDT-R frequency is 0.001) to demonstrate that the balance of male and
female fitness effects can move a rare allele to a stable polymorphism. Note that the
fixation of the allele is very rapid when DDT use is introduced and the return to the
polymorphic equilibrium after relaxing the strong directional selection for resistance
(DDT) is initially slow. Symbols plotted every 20 generations.
190 Rostant et al.
Author's personal copy
population cage studies and observe temporal change in replicate laboratorypopulations with different initial Cyp6g1 allele frequencies. This would alsocomplement population genetic models. At the molecular level, there remainsan opportunity to further investigate the genetic basis of DDT-R epistasis—whatare the modifiers altering the male fitness costs? AlthoughD. melanogaster is not apest species, understanding the relative reproductive success of susceptible andresistant flies with differing genetic backgrounds could provide valuable baselinedata to inform insecticide resistance management programs for pest species.Work to date suggests that using a single genetic background to test for effectsmay not be representative.
Given the great variation recently discovered at theCyp6g1 locus inD. melanogaster (Schmidt et al., 2010), the time is ripe to investigate the fitnesseffects of these newly discovered alleles. In the case of the more derived alleles,females have much higher DDT resistance than males (Schmidt et al., 2010),which points toward possible mitigation of the intralocus sexual conflictdescribed in Smith et al. (2011). One hypothesis, easily testable through a fitnesscomponents approach and/or selection-based stability analysis sensu Raymondet al. (2011) (population cage experiments described above), is that the allelic

2. TEs and Resistance 191
Author's personal copy
succession may be partially driven by the “replacement” mode of amelioration.However, intralocus conflict resolution like this presumably requires a change togenetic architecture, which makes resolution more difficult than it may appear(e.g., Harano et al., 2010).
It is unknown whether gene amplification or TE-induced cis-actingmutation has the greater effect on DDT resistance and associated pleiotropicfitness effects in D. melanogaster—dissecting the respective contributions toresistance/fitness would require single-copy TE-inserted Cyp6g1 alleles, andthese are yet to be found (Schmidt et al., 2010). The universal presence of TEinsertions in both copies of all DDT-R alleles thus found suggests that theinsertion occurred prior to, or concurrently with, the duplication event.
This parallels pyrethroid resistance in the mosquito C. quinquefasciatus.Here, resistance is associated with overexpression of another cytochrome P450gene, Cyp9m10 (Hardstone et al., 2010; Itokawa et al., 2010, 2011). As withD. melanogaster DDT-R, the constitutive upregulation occurs in haplotypes thathave an upstream insertion of a TE (in this case a truncated copy of the MITETE, CuRE1; Itokawa et al., 2010). Moreover, one of the resistant haplotypes alsoconsists of a tandem repeat of the TE-inserted sequence (Itokawa et al., 2011).Unlike the D. melanogaster DDT-R system, the relative contributions of the TEinsertion and gene amplification to resistance (and for that matter pleiotropicfitness) can easily be parsed out, since there are haplotypes that possessthe former but not the latter. Itokawa et al. (2011) suggest that, based on thenonlinear resistance efficacy to Cyp9m10 expression, the resistance phenotypeis disproportionately stronger as a result of the cis-acting mutation (the TEinsertion) occurring before the duplication event, than if the duplication hadpreceded the insertion. Such eerily similar stories for two different enzymes,conferring resistance to two different insecticides in two distantly related species,underline the usefulness of intensive study of model insect systems. They alsohint at a general pattern—tandem repeats, which are difficult to detect, could becommonly associated with TE insertion-induced insecticide resistance. Thisremains to be established.
Compared with the extensive work done on the Accord-insertedCyp6g1 in D. melanogaster, little is known about Doc-inserted Cyp6g1 inD. simulans. It remains to be seen whether this mutation has a significant andconsistent effect on resistance across different strains/genetic backgrounds.Furthermore, no work has been done to examine potential pleiotropic fitnesseffects of this insertion, much less the presence of epistatic interactions or thepossibility of intralocus sexual conflict, as has been demonstrated for D. mela-nogaster. A good first step may be to perform a worldwide survey akin to that ofthe Accord-LTR insertion by Catania et al. (2004). This would provide someindication of the geographic range of the Doc-inserted allele. Given the evi-dence for ongoing and rapid adaptation at Cyp6g1 in D. melanogaster, it may

192 Rostant et al.
Author's personal copy
well be worth having a closer look at the variation which exists at this locus inD. simulans. Just how similar the responses of the two species are to similarselection also remains to be seen.
Another avenue of research involves gene by environment interaction asit relates to fitness costs of resistance in these model systems. The laboratory-basedfitness component approach cannot fully encompass the full diversity of environ-ments in which wild populations face selection, and this may be a reason why costsare not always detected—environmental factors such as natural enemies, resourcelimitation, overwintering, and different host plant have all been shown to increaseresistance costs in various taxa (Carriere et al., 2001; Janmaat and Myers, 2005;Raymond et al., 2005, 2007, 2011). Moving population cage experiments outdoorscould increase the reality of the stability-selection approach, giving a betterreflection of how well resistance genotypes perform under natural conditions.
Just as the genetic background provided by the rest of the genomerepresents a genetic “environment” in which resistance alleles act, so does thepresence of extragenomic DNA, including cytoplasmic endosymbionts. Wolba-chia is a maternally transmitted intracellular bacterium found in a wide range ofarthropods and nematodes (Stouthamer et al., 1999; Werren, 1997). Its relation-ship with its host ranges from parasitic to symbiotic. At the parasitic end of thespectrum, it can have profound effects on host reproduction, displaying a range ofphenotypes from male killing to feminization to cytoplasmic incompatibility(Stouthamer et al., 1999; Werren, 1997). These strategies increase its transmis-sion within a population, often at the expense of its host’s fitness—the hallmarkof an SGE. Wolbachia is found not only in Drosophila (where it has undergone avery recent expansion to near fixation in many populations), but also in manyother insects including pest species—one recent estimate is that more than 66%of arthropod species harbor Wolbachia infections (Hilgenboecker et al., 2008).Given its ubiquity and potentially profound effect on host fitness, Wolbachiacannot be ignored when examining pleiotropic effects of resistance. For example,Wolbachia has been implicated in directly modifying the cost of insecticideresistance in mosquitoes (Duron et al., 2006). Where insecticide resistance isconferred by a TE, we may find that intergenomic interactions (akin to epistasis)between the TE and intracellular endosymbionts are critical to the populationgenetics of insecticide resistance alleles.
Although D. melanogaster is not a pest species, understanding therelative fitness of susceptible and resistant flies with differing genetic backgroundsand under different environments could provide valuable insights to informinsecticide resistance management programs for pest species. To this end, weurge the use of multiple avenues of investigation that include the laboratory-based, sex-specific fitness component approach, stability-selection experiments,and mathematical modeling to increase our understanding of insecticideresistance dynamics in natural populations.

2. TEs and Resistance 193
Author's personal copy
Acknowledgments
N. W. is funded by a Royal Society Wolfson Research Merit Award and D. J. H. is funded by NERC.
We thank Judith Mank for comments on a previous version of this chapter.
References
Alyokhin, A. V., and Ferro, D. N. (1999). Relative fitness of Colorado potato beetle (Coleoptera:
Chrysomelidae) resistant and susceptible to the Bacillus thuringiensis Cry3A toxin. J. Econ.
Entomol. 92, 510–515.Aminetzach, Y. T., Macpherson, J. M., and Petrov, D. A. (2005). Pesticide resistance via transposition-
mediated adaptive gene truncation in Drosophila. Science 309, 764–767.Anxolabehere, D., Kidwell, M., and Periquet, G. (1988). Molecular characteristics of diverse
populations are consistent with the hypothesis of a recent invasion of Drosophila melanogaster by
mobile P elements. Mol. Biol. Evol. 5, 252–269.Aravin, A. A., Hannon, G. J., and Brennecke, J. (2007). The Piwi-piRNA pathway provides an
adaptive defense in the transposon arms race. Science 318, 761–764.Argumuganathan, K., and Earle, E. D. (1991). Nuclear DNA content of some important plant
species. Plant Mol. Biol. Rep. 9, 208–218.Arnaud, L., and Haubruge, E. (2002). Insecticide resistance enhances male reproductive success in a
beetle. Evolution 56, 2435–2444.Baker, J. E., Perezmendoza, J., Beeman, R. W., and Throne, J. E. (1998). Fitness of a malathion-
resistant strain of the parasitoid Anisopteromalus calandrae (Hymenoptera: Pteromalidae). J. Econ.
Entomol. 91, 50–55.Baker, M. B., Dastur, S. R., Jaffe, B. D., and Wong, T. (2008). Mating competition in Colorado
potato beetles (Coleoptera: Chrysomelidae) does not show a cost of insecticide resistance. Ann.
Entomol. Soc. Am. 101, 371–377.Bass, C., and Field, L. M. (2011). Gene amplification and insecticide resistance. Pest Manag. Sci. 67,
886–890.
Bellen, H. J., Levis, R.W., He, Y. C., Carlson, J. W., Evans-Holm, M., Bae, E., Kim, J., Metaxakis, A.,
Savakis, C., Schulze, K. L., et al. (2011). The Drosophila gene disruption project: Progress using
transposons with distinctive site-specificities. Genetics 188, 731–743.Bennetzen, J. (2005). Transposable elements, gene creation and genome rearrangement in flowering
plants. Curr. Opin. Genet. Dev. 15, 621–627.Berrada, S., and Fournier, D. (1997). Transposition-mediated transcriptional overexpression as a
mechanism of insecticide resistance. Mol. Gen. Genet. 256, 348–354.Berticat, C., Boquien, G., Raymond, M., and Chevillon, C. (2002). Insecticide resistance genes
induce a mating competition cost in Culex pipiens mosquitoes. Genet. Res. 79, 41–47.Bielza, P., Quinto, V., Gravalos, C., Abellan, J., and Fernandez, E. (2008). Lack of fitness costs of
insecticide resistance in the western flower thrips (Thysanoptera: Thripidae). J. Econ. Entomol.
101, 499–503.Biemont, C. (2010). A brief history of the status of transposable elements: From junk DNA to major
players in evolution. Genetics 186, 1085–1093.Biemont, C., and Vieira, C. (2006). Genetics—Junk DNA as an evolutionary force. Nature 443,
521–524.
Biessmann, H., Valgeirsdottir, K., Lofsky, A., Chin, C., Ginther, B., Levis, R. W., and Pardue, M. L.
(1992). HeTA, a transposable element specifically involved in “healing” broken chromosome
ends in Drosophila melanogaster. Mol. Cell. Biol. 12, 3910–3918.

194 Rostant et al.
Author's personal copy
Bloch, G., and Wool, D. (1994). Methidathion resistance in the sweet potato whitefly (Aleyrodidae:
Homoptera) in Israel: Selection, heritability, and correlated changes of esterase activity. J. Econ.
Entomol. 87, 1147–1156.
Bogwitz, M. R., Chung, H., Magoc, L., Rigby, S., Wong, W., O’Keefe, M., McKenzie, J. A.,
Batterham, P., and Daborn, P. J. (2005). Cyp12a4 confers lufenuron resistance in a natural
population of Drosophila melanogaster. Proc. Natl. Acad. Sci. U.S.A. 102, 12807–12812.
Boivin, T., Chabert d’Hieres, C., Bouvier, J. C., Beslay, D., and Sauphanor, B. (2001). Pleiotropy of
insecticide resistance in the codling moth, Cydia pomonella. Entomol. Exp. Appl. 99, 381–386.
Bonduriansky, R., and Chenoweth, S. F. (2009). Intralocus sexual conflict. Trends Ecol. Evol. 24,280–288.
Brewer, M. J., and Trumble, J. T. (1991). Inheritance and fitness consequences of resistance to
fenvalerate in Spodoptera exigua (Lepidoptera, Noctuidae). J. Econ. Entomol. 84, 1638–1644.
Brookfield, J. F. Y. (2005). The ecology of the genome—Mobile DNA elements and their hosts.Nat.
Rev. Genet. 6, 128–136.
Brookfield, J., and Badge, R. (1997). Population genetics models of transposable elements. Genetica
100, 281–294.
Burt, A., and Trivers, R. (2006). Genes in Conflict: The Biology of Selfish Genetic Elements. The
Belknap Press of Harvard University Press, London.
Carriere, Y., Deland, J.-P., Roff, D. A., and Vincent, C. (1994). Life-history costs associated with the
evolution of insecticide resistance. Proc. R. Soc. Lond. B Biol. Sci. 258, 35–40.
Carriere, Y., Roff, D. A., and Deland, J. P. (1995). The joint evolution of diapause and insecticide
resistance—A test of an optimality model. Ecology 76, 1497–1505.
Carriere, Y., Ellers-Kirk, C., Patin, A. L., Sims, M. A., Meyer, S., Liu, Y. B., Dennehy, T. J., and
Tabashnik, B. E. (2001). Overwintering cost associated with resistance to transgenic cotton in the
pink bollworm (Lepidoptera: Gelechiidae). J. Econ. Entomol. 94, 935–941.
Castaneda, L. E., Barrientos, K., Cortes, P. A., Figueroa, C. C., Fuentes-Contreras, E., Luna-
Rudloff, M., Silva, A. X., and Bacigalupe, L. D. (2011). Evaluating reproductive fitness and
metabolic costs for insecticide resistance in Myzus persicae from Chile. Physiol. Entomol. 36,253–260.
Catania, F., Kauer, M. O., Daborn, P. J., Yen, J. L., ffrench-Constant, R. H., and Schlotterer, C.
(2004). World-wide survey of an Accord insertion and its association with DDT resistance in
Drosophila melanogaster. Mol. Ecol. 13, 2491–2504.
Charlesworth, D., and Charlesworth, B. (1995). Transposable elements in inbreeding and outbreed-
ing populations. Genetics 140, 415–417.
Charlesworth, B., and Charlesworth, D. (2000). The degeneration of Y chromosomes. Philos. Trans.
R. Soc. Lond. B Biol. Sci. 355, 1563–1572.
Charlesworth, B., and Charlesworth, D. (2010). Elements of Evolutionary Genetics. Roberts and
Company Publishers, Greenwood Village, Colorado.
Charlesworth, B., and Langley, C. H. (1986). The evolution of self-regulated transposition of
transposable elements. Genetics 112, 359–383.
Charlesworth, B., and Langley, C. H. (1989). The population genetics of Drosophila transposable
elements. Annu. Rev. Genet. 23, 251–287.
Charlesworth, B., Sniegowski, P., and Stephan, W. (1994). The evolutionary dynamics of repetitive
DNA in eukaryotes. Nature 371, 215–220.
Charlesworth, B., Langley, C., and Sniegowski, P. (1997). Transposable element distributions in
Drosophila. Genetics 147, 1993–1995.
Chen, S., and Li, X. C. (2007). Transposable elements are enriched within or in close proximity to
xenobiotic-metabolizing cytochrome P450 genes. BMC Evol. Biol. 7, 13.

2. TEs and Resistance 195
Author's personal copy
Chevillon, C., Bourguet, D., Rousset, F., Pasteur, N., and Raymond, M. (1997). Pleiotropy of
adaptive changes in populations: Comparisons among insecticide resistance genes inCulex pipiens.
Genet. Res. 70, 195–203.
Chung, H., Bogwitz, M. R., McCart, C., Andrianopoulos, A., ffrench-Constant, R. H., Batterham, P.,
and Daborn, P. J. (2007). Cis-regulatory elements in the Accord retrotransposon result in tissue-
specific expression of the Drosophila melanogaster insecticide resistance geneCyp6g1.Genetics 175,1071–1077.
Clark, J. M., and Yamaguchi, I. (eds.) (2002). In “Agrochemical Resistance: Extent, Mechanism, and
Detection”In ACS Symposium Series No. 808. American Chemical Society, Washington, DC.
Cohan, F., King, E., and Zawadzki, P. (1994). Amelioration of the deleterious pleiotropic effects of an
adaptive mutation in Bacillus subtilis. Evolution 48, 81–95.
Cordaux, R., and Batzer, M. A. (2009). The impact of retrotransposons on human genome evolution.
Nat. Rev. Genet. 10, 691–703.
Crow, J. F. (1957). Genetics of insect resistance to chemicals. Annu. Rev. Entomol. 2, 227–246.
Currie, D. B., Mackay, T. F., and Partridge, L. (1998). Pervasive effects of P element mutagenesis on
body size in Drosophila melanogaster. Genet. Res. 72, 19–24.
Daborn, P., Boundy, S., Yen, J., Pittendrigh, B., and ffrench-Constant, R. (2001). DDT resistance in
Drosophila correlates with Cyp6g1 over-expression and confers cross-resistance to the neonicoti-
noid imidacloprid. Mol. Genet. Genomics 266, 556–563.
Daborn, P. J., Yen, J. L., Bogwitz, M. R., Le Goff, G., Feil, E. S., Jeffers, S., Tijet, N., Perry, T.,
Heckel, D., Batterham, P., Feyereisen, R., Wilson, T. G., et al. (2002). A single P450 allele
associated with insecticide resistance in Drosophila. Science 297, 2253–2256.
Daniels, S. B., Peterson, K. R., Strausbaugh, L. D., Kidwell, M. G., and Chovnick, A. (1990).
Evidence for horizontal transmission of the P-transposable element between Drosophila species.
Genetics 124, 339–355.
Dapkus, D., and Merrell, D. J. (1977). Chromosomal analysis of DDT-resistance in a long-term
selected population of Drosophila melanogaster. Genetics 87, 685–697.
Delpuech, J.-M., Aquadro, C. F., and Roush, R. T. (1993). Noninvolvement of the long terminal
repeat of transposable element 17.6 in insecticide in Drosophila. Proc. Natl. Acad. Sci. U.S.A. 90,5643–5647.
Denholm, I., Pickett, J. A., and Devonshire, A. L. (eds.) (1999). In “Insecticide Resistance: From
Mechanisms to Management.” CAB International Publishing, Wallingford, UK.
Devonshire, A. L., and Field, L. M. (1991). Gene amplification and insecticide resistance.Annu. Rev.
Entomol. 36, 1–23.
Dominguez, A., and Albornoz, J. (1996). Rates of movement of transposable elements in Drosophila
melanogaster. Mol. Gen. Genet. 251, 130–138.
Doolittle, W. F., and Sapienza, C. (1980). Selfish genes, the phenotype paradigm and genome
evolution. Nature 284, 601–603.
Dowsett, A., and Young, M. (1982). Differing levels of dispersed repetitive DNA among closely
related species of Drosophila. Proc. Natl. Acad. Sci. U.S.A. 79, 4570–4574.
Drnevich, J. M., Reedy, M. M., Ruedi, E. A., Rodriguez-Zas, S., and Hughes, K. A. (2004).
Quantitative evolutionary genomics: Differential gene expression and male reproductive success
in Drosophila melanogaster. Proc. R. Soc. Lond. B Biol. Sci. 271, 2267–2273.
Duron, O., Labbe, P., Berticat, C., Rousset, F., Guillot, S., Raymond, M., andWeill, M. (2006). High
Wolbachia density correlates with cost of infection for insecticide resistant Culex pipiens mosqui-
toes. Evolution 60, 303–314.
Eanes, W. F., Wesley, C., Hey, J., Houle, D., and Ajioka, J. W. (1988). The fitness consequences of P
element insertion in Drosophila melanogaster. Genet. Res. 52, 17–26.

196 Rostant et al.
Author's personal copy
Emerson, J. J., Cardoso-Moreira, M., Borevitz, J. O., and Long, M. (2008). Natural selection shapes
genome-wide patterns of copy-number polymorphism in Drosophila melanogaster. Science 320,1629–1631.
Feschotte, C. (2008). Transposable elements and the evolution of regulatory networks. Nat. Rev.
Genet. 9, 397–405.Feyereisen, R. (1995). Molecular biology of insecticide resistance. Toxicol. Lett. 82, 83–90.ffrench-Constant, R. H. (1999). Target site mediated insecticide resistance: What questions remain?
Insect Biochem. Mol. Biol. 29, 397–403.ffrench-Constant, R. H., Pittendrigh, B., Vaughan, A., and Anthony, N. (1998). Why are there so
few resistance-associated mutations in insecticide target genes? Philos. Trans. R. Soc. Lond. B Biol.
Sci. 353, 1685–1693.ffrench-Constant, R. H., Daborn, P. J., and Le Goff, G. (2004). The genetics and genomics of
insecticide resistance. Trends Genet. 20, 163–170.ffrench-Constant, R., Daborn, P., and Feyereisen, R. (2006). Resistance and the jumping gene.
Bioessays 28, 6–8.Field, L. M., Devonshire, A. L., and Forde, B. G. (1988). Molecular evidence that insecticide
resistance in peach-potato aphids (Myzus persicae Sulz.) results from amplification of an esterase
gene. Biochem. J. 251, 309–312.Finnegan, D. J. (1989). Eukaryotic transposable elements and genome evolution. Trends Genet. 5,
103–107.
Finnegan, D. J. (1992). Transposable elements. In “The Genome of Drosophila melanogaster”
(D. L. Lindsley and G. G. Zimm, eds.), pp. 1096–1107. Academic Press, San Diego.
Fisher, R. A. (1928). The possible modification of the responses of wild type to recurrent mutations.
Am. Nat. 62, 115–126.Fitzpatrick, B., and Sved, J. (1986). High-levels of fitness modifiers induced by hybrid dysgenesis in
Drosophila melanogaster. Genet. Res. 48, 89–94.Follett, P. A., Gould, F., and Kennedy, G. G. (1993). Comparative fitness of 3 strains of Colorado
potato beetle (Coleoptera, Chrysomelidae) in the field—Spatial and temporal variation in
insecticide selection. J. Econ. Entomol. 86, 1324–1333.Foster, S. P., Young, S., Williamson, M. S., Duce, I., Denholm, I., and Devine, G. J. (2003).
Analogous pleiotropic effects of insecticide resistance genotypes in peach-potato aphids and
houseflies. Heredity 91, 98–106.Gahan, L. J., Gould, F., and Heckel, D. G. (2001). Identification of a gene associated with Bt
resistance in Heliothis virescens. Science 293, 857–860.George, J. A., Traverse, K. L., DeBaryshe, P. G., Kelley, K. J., and Pardue, M. L. (2010). Evolution of
diverse mechanisms for protecting chromosome ends by Drosophila TART telomere retrotranspo-
sons. Proc. Natl. Acad. Sci. U.S.A. 107, 21052–21057.Georghiou, G. P., and Lagunes-Tejeda, A. (1991). The Occurrence of Resistance to Pesticides in
Arthropods. An Index of Cases Reported Through 1989. Food and Agriculture Organization,
Rome.
Georghiou, G. P., and Saito, T. (eds.) (1983). In “Pest Resistance to Pesticides” Plenum Press,
New York.
Gonzalez, J., and Petrov, D. A. (2009). The adaptive role of transposable elements in the Drosophila
genome. Gene 448, 124–133.Grant, D. F., and Hammock, B. D. (1992). Genetic and molecular evidence for a trans-acting
regulatory locus controlling glutathione stransferase-2 expression in Aedes aegypti. Mol. Gen.
Genet. 234, 169–176.Green, M. (1988). Mobile DNA elements and spontaneous gene mutation. In “Eukaryotic Trans-
posable Elements as Mutagenic Agents” (M. Lambert, J. McDonald, and I. Weinstein, eds.),
pp. 41–50. Cold Spring Harbor Laboratory Press, Cold Spring Harbor.

2. TEs and Resistance 197
Author's personal copy
Haldane, J. B. S. (1932). The Causes of Evolution. Harper, New York, NY.
Hallstrom, I. (1985). Genetic regulation of the cytochrome P-450 system in Drosophila melanogaster.
II. Localization of some genes regulating cytochrome P-450 activity. Chem. Biol. Interact. 56,173–184.
Harano, T., Okada, K., Nakayama, S., Miyatake, T., and Hosken, D. J. (2010). Intralocus sexual
conflict unresolved by sex-limited trait expression. Curr. Biol. 20, 2036–2039.Hardstone, M. C., Komagata, O., Kasai, S., Tomita, T., and Scott, J. G. (2010). Use of isogenic
strains indicates CYP9M10 is linked to permethrin resistance in Culex pipiens quinquefasciatus.
Insect Mol. Biol. 19, 717–726.Hartl, D. L., and Clark, A. G. (1997). Principles of Population Genetics. Sinauer Associates,
Sunderland, Massachusetts.
Haubruge, E., and Arnaud, L. (2001). Fitness consequences of malathion-specific resistance in red
flour beetle (Coleoptera: Tenebrionidae) and selection for resistance in the absence of malathion.
J. Econ. Entomol. 94, 552–557.Hickey, D. A. (1982). Selfish DNA: A sexually-transmitted nuclear parasite. Genetics 101, 519–531.Hilgenboecker, K., Hammerstein, P., Schlattmann, P., Telschow, A., andWerren, J. H. (2008). How
many species are infected with Wolbachia? A statistical analysis of current data. FEMS Microbiol.
Lett. 281, 215–220.Hollingsworth, R. G., Tabashnik, B. E., Johnson, M. W., Messing, R. H., and Ullman, D. E. (1997).
Relationship between susceptibility to insecticides and fecundity across populations of cotton
aphid (Homoptera: Aphididae). J. Econ. Entomol. 90, 55–58.Hollingworth, R. M., and Dong, K. (2008). The biochemical and molecular genetic basis of
resistance to pesticides in arthropods. In “Global Pesticide Resistance in Arthropods”
(M. E. Whalon, D. Mota-Sanchez, and R. M. Hollingworth, eds.), pp. 40–89. CABI, Oxfordshire,
UK.
Hosken, D. J., Stockley, P., Tregenza, T., and Wedell, N. (2009). Monogamy and the battle of the
sexes. Annu. Rev. Entomol. 54, 361–378.Hua-Van, A., Le Rouzic, A., Boutin, T. S., Filee, J., and Capy, P. (2011). The struggle for life of the
genome’s selfish architects. Biol. Direct 6, 19.Hurst, G. D. D., and Werren, J. H. (2001). The role of selfish genetic elements in eukaryotic
evolution. Nat. Rev. Genet. 2, 597–606.Ishaaya, I. (ed.) (2001). In “Biochemical Sites of Insecticide Action and Resistance” Springer, New
York.
Itokawa, K., Komagata, O., Kasai, S., Okamura, Y., Masada, M., and Tomita, T. (2010). Genomic
structures of Cyp9m10 in pyrethroid resistant and susceptible strains of Culex quinquefasciatus.
Insect Biochem. Mol. Biol. 40, 631–640.Itokawa, K., Komagata, O., Kasai, S., Masada, M., and Tomita, T. (2011). Cis-acting mutation and
duplication: History of molecular evolution in a P450 haplotype responsible for insecticide
resistance in Culex quinquefasciatus. Insect Biochem. Mol. Biol. 41, 503–512.Janmaat, A. F., and Myers, J. H. (2005). The cost of resistance to Bacillus thuringiensis varies with the
host plant of Trichoplusia ni. Proc. R. Soc. Lond. B Biol. Sci. 272, 1031–1038.Jiang, N., Bao, Z. R., Zhang, X. Y., Eddy, S. R., and Wessler, S. R. (2004). Pack�MULE transposable
elements mediate gene evolution in plants. Nature 431, 569–573.Kaminker, J. S., Bergman, C. M., Kronmiller, B., Carlson, J., Svirskas, R., Patel, S., Frise, E.,
Wheeler, D. A., Lewis, S. E., Rubin, G. M., Ashburner, M., and Celniker, S. E. (2002). The
transposable elements of the Drosophila melanogaster euchromatin: A genomics perspective.
Genome Biol.3RESEARCH0084.Karunaratne, S. H., Jayawardena, K. G., Hemingway, J., and Ketterman, A. J. (1993). Characteriza-
tion of a B-type esterase involved in insecticide resistance from the mosquito Culex quinquefascia-
tus. Biochem. J. 294, 575–579.

198 Rostant et al.
Author's personal copy
Kidwell, M., and Lisch, D. (2001). Perspective: Transposable elements, parasitic DNA, and genome
evolution. Evolution 55, 1–24.
Kikkawa, H. (1961). Genetical studies on the resistance to parathion in Drosophila melanogaster.
Annu. Rep. Sci. Works Fac. Sci. Osaka Univ. 9, 1–20.
Kim, J., Vanguri, S., Boeke, J. D., Gabriel, A., and Voytas, D. F. (1998). Transposable elements and
genome organization: A comprehensive survey of retrotransposons revealed by the complete
Saccharomyces cerevisiae genome sequence. Genome Res. 8, 464–478.
Labbe, P., Sidos, N., Raymond, M., and Lenormand, T. (2009). Resistance gene replacement in the
mosquito Culex pipiens: Fitness estimation from long-term cline series. Genetics 182, 303–312.
Lande, R. (1983). The response to selection on major and minor mutations affecting a metrical trait.
Heredity 50, 47–65.
Lander, E. S., Linton, L. M., Birren, B., Nusbaum, C., Zody, M. C., Baldwin, J., Devon, K., Dewar, K.,
Doyle, M., FitzHugh, W., et al. (2001). Initial sequencing and analysis of the human genome.
Nature 409, 860–921.
Levin, H. L., and Moran, J. V. (2011). Dynamic interactions between transposable elements and
their hosts. Nat. Rev. Genet. 12, 615–627.
Levis, R. W., Ganesan, R., Houtchens, K., Tolar, L. A., and Sheen, F. M. (1993). Transposons in
place of telomeric repeats at a Drosophila telomere. Cell 75, 1083–1093.
Li, X., Schuler, M. A., and Berenbaum, M. R. (2007). Molecular mechanisms of metabolic resistance
to synthetic and natural xenobiotics. Annu. Rev. Entomol. 52, 231–253.
Lockton, S., and Gaut, B. S. (2010). The evolution of transposable elements in natural populations of
self-fertilizing Arabidopsis thaliana and its outcrossing relative Arabidopsis lyrata. BMC Evol. Biol.
10, 10.
Lockton, S., Ross-Ibarra, J., and Gaut, B. S. (2008). Demography and weak selection drive patterns of
transposable element diversity in natural populations of Arabidopsis lyrata. Proc. Natl. Acad. Sci.
U.S.A. 105, 13965–13970.
Lynch, M., and Conery, J. (2003). The origins of genome complexity. Science 302, 1401–1404.
Mackay, T. F. (1986). Transposable element-induced fitness mutations in Drosophila melanogaster.
Genet. Res. 48, 77–87.
Mackay, T. F. (1989). Transposable elements and fitness in Drosophila melanogaster. Genome 31,284–295.
Mackay, T. F., Lyman, R. F., and Jackson, M. S. (1992). Effects of P element insertions on
quantitative traits in Drosophila melanogaster. Genetics 130, 315–332.
Macnair, M. R. (1991). Why the evolution of resistance to anthropogenic toxins normally involves
major gene changes: The limits to natural selection. Genetica 84, 213–219.
Mallet, J. (1989). The evolution of insecticide resistance: Have the insects won? Trends Ecol. Evol. 4,336–340.
Malone, C. D., and Hannon, G. J. (2009). Small RNAs as guardians of the genome. Cell 136,656–668.
Maside, X., Assimacopoulos, S., and Charlesworth, B. (2000). Rates of movement of transposable
elements on the second chromosome of Drosophila melanogaster. Genet. Res. 75, 275–284.
Maside, X., Bartolome, C., and Charlesworth, B. (2002). S-element insertions are associated with the
evolution of the Hsp70 genes in Drosophila melanogaster. Curr. Biol. 12, 1686–1691.
Mason, P. L. (1998). Selection for and against resistance to insecticides in the absence of insecticide:
A case study of malathion resistance in the saw-toothed grain beetle, Oryzaephilus surinamensis
(Coleoptera: Silvanidae). Bull. Entomol. Res. 88, 177–188.
May, R. (1985). Population biology—Evolution of pesticide resistance. Nature 315, 12–13.
McCart, C., Buckling, A., and ffrench-Constant, R. H. (2005). DDT resistance in flies carries no
cost. Curr. Biol. 15, R587–R589.

2. TEs and Resistance 199
Author's personal copy
McClintock, B. (1950). The origin and behavior of mutable loci in maize. Proc. Natl. Acad. Sci. U.S.A.
36, 344–355.McClintock, B. (1984). The significance of responses of the genome to challenge. Science 226,
792–801.
McKenzie, J. A., and Batterham, P. (1994). The genetics, molecular and phenotypic consequences of
selection for insecticide resistance. Trends Ecol. Evol. 9, 166–169.Melander, A. L. (1914). Can insects become resistant to sprays? J. Econ. Entomol. 7, 167–173.Messing, J., and Dooner, H. (2006). Organization and variability of the maize genome. Curr. Opin.
Plant Biol. 9, 157–163.Minkoff, C., and Wilson, T. G. (1992). The competitive ability and fitness components of the
Methoprene-tolerant (Met) Drosophila mutant resistant to juvenile hormone analog insecticides.
Genetics 131, 91–97.Mouches, C., Pasteur, N., Berge, J. B., Hyrien, O., Raymond, M., de Saint Vincent, B. R.,
de Silvestri, M., and Georghiou, G. P. (1986). Amplification of an esterase gene is responsible
for insecticide resistance in a California Culex mosquito. Science 233, 778–780.Mouches, C., Pauplin, Y., Agarwal, M., Lemieux, L., Herzog, M., Abadon, M., Beyssat-Arnaouty, V.,
Hyrien, O., de Saint Vincent, B. R., Georghiou, G. P., and Pasteur, N. (1990). Characterization of
amplification core and Esterase-b1 gene responsible for insecticide resistance in Culex. Proc. Natl.
Acad. Sci. U.S.A. 87, 2574–2578.Nuzhdin, S., and Mackay, T. (1995). The genomic rate of transposable element movement in
Drosophila melanogaster. Mol. Biol. Evol. 12, 180–181.O’Donnell, K. A., and Burns, K. H. (2010). Mobilizing diversity: Transposable element insertions in
genetic variation and disease. Mob. DNA 1(1), 21.Oakeshott, J. G., Horne, I., Sutherland, T. D., and Russell, R. J. (2003). The genomics of insecticide
resistance. Genome Biol. 4, 202.Omer, A. D., Leigh, T. F., and Granett, J. (1992). Insecticide resistance of greenhouse-whitefly
(Homoptera, Aleyrodidae) and fitness on plant hosts relative to the San-Joaquin valley (Califor-
nia) cotton agroecosystem. J. Appl. Entomol. 113, 244–251.Oppert, B., Hammel, R., Throne, J. E., and Kramer, K. J. (2000). Fitness costs of resistance to Bacillus
thuringiensis in the Indian meal moth, Plodia interpunctella. Entomol. Exp. Appl. 96, 281–287.Orgel, L. E., and Crick, F. H. (1980). Selfish DNA: The ultimate parasite. Nature 284, 604–607.Petrov, D. A., Fiston-Lavier, A.-S., Lipatov, M., Lenkov, K., and Gonzalez, J. (2011). Population
genomics of transposable elements in Drosophila melanogaster. Mol. Biol. Evol. 28, 1633–1644.Raymond, M., Beyssat-Arnaouty, V., Sivasubramanian, N., Mouches, C., Georghiou, G. P., and
Pasteur, N. (1989). Amplification of various esterase B’s responsible for organophosphate resis-
tance in Culex mosquitoes. Biochem. Genet. 27, 417–423.Raymond, B., Sayyed, A. H., andWright, D. J. (2005). Genes and environment interact to determine
the fitness costs of resistance to Bacillus thuringiensis. Proc. R. Soc. Lond. B Biol. Sci. 272,1519–1524.
Raymond, B., Sayyed, A. H., Hails, R. S., and Wright, D. J. (2007). Exploiting pathogens and their
impact on fitness costs to manage the evolution of resistance to Bacillus thuringiensis. J. Appl. Ecol.
44, 768–780.Raymond, B., Wright, D. J., and Bonsall, M. B. (2011). Effects of host plant and genetic background
on the fitness costs of resistance to Bacillus thuringiensis. Heredity 106, 281–288.Rivero, A., Magaud, A., Nicot, A., and Vezilier, J. (2011). Energetic cost of insecticide resistance in
Culex pipiens mosquitoes. J. Med. Entomol. 48, 694–700.Robertson, H. M., and Engels, W. R. (1989). Modified P elements that mimic the P cytotype in
Drosophila melanogaster. Genetics 123, 815–824.Roush, R. T., and McKenzie, J. A. (1987). Ecological genetics of insecticide and acaricide resistance.
Annu. Rev. Entomol. 32, 361–380.

200 Rostant et al.
Author's personal copy
Schlenke, T. A., and Begun, D. J. (2004). Strong selective sweep associated with a transposon
insertion in Drosophila simulans. Proc. Natl. Acad. Sci. U.S.A. 101, 1626–1631.
Schmidt, J. M., Good, R. T., Appleton, B., Sherrard, J., Raymant, G. C., Bogwitz, M. R., Martin, J.,
Daborn, P. J., Goddard, M. E., Batterham, P., et al. (2010). Copy number variation and transpos-
able elements feature in recent, ongoing adaptation at the Cyp6g1 locus. PLoS Genet. 6,e1000998.
Shapiro, J. A. (1969). Mutations caused by the insertion of genetic material into the galactose operon
of Escherichia coli. J. Mol. Biol. 40, 93–105.
Smith, D. T., Hosken, D. J., Rostant, W. G., Yeo, M., Griffin, R. M., Bretman, A., Price, T. A. R.,
ffrench-Constant, R. H., andWedell, N. (2011). DDT resistance, epistasis and male fitness in flies.
J. Evol. Biol. 24, 1351–1362.
Sparks, T. C., Lockwood, J. A., Byford, R. L., Graves, J. B., and Leonard, B. R. (1989). The role of
behavior in insecticide resistance. Pestic. Sci. 26, 383–399.
Steinemann, M., and Steinemann, S. (1998). Enigma of Y chromosome degeneration: Neo-Y and
Neo-X chromosomes of Drosophila miranda a model for sex chromosome evolution. Genetica 102,409–420.
Stouthamer, R., Breeuwer, J., and Hurst, G. (1999). Wolbachia pipientis: Microbial manipulator of
arthropod reproduction. Annu. Rev. Microbiol. 53, 71–102.
Tang, J. D., Gilboa, S., Roush, R. T., and Shelton, A. M. (1997). Inheritance, stability, lack of fitness
costs of field-selected resistance to Bacillus thuringiensis in diamondback moth (Lepidoptera:
Plutellidae) from Florida. J. Econ. Entomol. 90, 732–741.
Tang, J. D., Collins, H. L., Roush, R. T., Metz, T. D., Earle, E. D., and Shelton, A. M. (1999).
Survival, weight gain, and oviposition of resistant and susceptible Plutella xylostella (Lepidoptera:
Plutellidae) on broccoli expressing CrylAc toxin of Bacillus thuringiensis. J. Econ. Entomol. 92,47–55.
Thomas, C. (1971). Genetic organization of chromosomes. Annu. Rev. Genet. 5, 237–256.
Thompson, M., Steichen, J. C., and ffrench-Constant, R. H. (1993). Conservation of cyclodiene
insecticide resistance associated mutations in insects. Insect Mol. Biol. 2, 149–154.
Touchon, M., and Rocha, E. P. C. (2007). Causes of insertion sequences abundance in prokaryotic
genomes. Mol. Biol. Evol. 24, 969–981.
van Rij, R. P., and Berezikov, E. (2009). Small RNAs and the control of transposons and viruses in
Drosophila. Trends Microbiol. 17, 163–171.
Waters, L. C., Zelhof, A. C., Shaw, B. J., and Ch’ang, L.-Y. (1992). Possible involvement of the long
terminal repeat of transposable element 17.6 in regulating expression of an insecticide resistance-
associated P450 gene in Drosophila. Proc. Natl. Acad. Sci. U.S.A. 89, 4855–4859.
Werren, J. H. (1997). Biology of Wolbachia. Annu. Rev. Entomol. 42, 587–609.
Werren, J. H. (2011). Selfish genetic elements, genetic conflict, and evolutionary innovation. Proc.
Natl. Acad. Sci. U.S.A. 108, 10863–10870.
Whalon, M. E., Mota-Sanchez, D., and Hollingworth, R. M. (eds.) (2008). In “Global Pesticide
Resistance in Arthropods.” CAB International Publishing, Wallingford, UK.
White, N. D. G., and Bell, R. J. (1995). A malathion resistance gene associated with increased life-
span of the rusty grain beetle, Cryptolestes ferrugineus (Coleoptera, Cucujidae). J. Gerontol. A Biol.
Sci. Med. Sci. 50, B9–B13.
Wicker, T., Sabot, F., Hua-Van, A., Bennetzen, J. L., Capy, P., Chalhoub, B., Flavell, A., Leroy, P.,
Morgante, M., Panaud, O., et al. (2007). A unified classification system for eukaryotic transposable
elements. Nat. Rev. Genet. 8, 973–982.
Wilson, T. G. (1993). Transposable elements as initiators of insecticide resistance. J. Econ. Entomol.
86, 645–651.
Wilson, T. G. (2001). Resistance of Drosophila to toxins. Annu. Rev. Entomol. 46, 545–571.

2. TEs and Resistance 201
Author's personal copy
Wilson, T., and Turner, C. (1992). Molecular analysis of methoprene-tolerant, a gene in Drosophila
involved in resistance to juvenile-hormone analog insect growth-regulators. In “Molecular Mech-
anisms of Insecticide Resistance: Diversity Among Insects” (C. A. Mullin and J. G. Scott, eds.),
ACS Symposium Series No. 505, pp. 99–112. American Chemical Society, Washington, DC.
Xiao, H., Jiang, N., Schaffner, E., Stockinger, E. J., and van der Knaap, E. (2008). A retrotransposon-
mediated gene duplication underlies morphological variation of tomato fruit. Science 319,1527–1530.
Xing, J. C., Zhang, Y. H., Han, K., Salem, A. H., Sen, S. K., Huff, C. D., Zhou, Q., Kirkness, E. F.,
Levy, S., Batzer, M. A., et al. (2009). Mobile elements create structural variation: Analysis of a
complete human genome. Genome Res. 19, 1516–1526.Yamamoto, A., Yoneda, H., Hatano, R., and Asada, M. (1995). Influence of hexythiazox resistance
on life-history parameters in the citrus red mite, Panonychus citri (McGregor). J. Pestic. Sci. 20,521–527.
Yang, S., Arguello, J. R., Li, X., Ding, Y., Zhou, Q., Chen, Y., Zhang, Y., Zhao, R., Brunet, F.,
Peng, L., et al. (2008). Repetitive element-mediated recombination as a mechanism for new gene
origination in Drosophila. PLoS Genet. 4, e3.Zhang, Z., and Saier, M. H. (2009). A novel mechanism of transposon-mediated gene activation.
PLoS Genet. 5, e1000689.
Copyright © 2022 FDOKUMEN




















