
Transport of Farfantepenaeus Aztecus Postlarvae in Three Lagoon-System Inlets in the Southwestern...
13
TRANSPORT OF FARFANTEPENAEUS AZTECUS POSTLARVAE IN THREE LAGOON-SYSTEM INLETS IN THE SOUTHWESTERN GULF OF MEXICO BY SERGIO CHÁZARO-OLVERA 1,2 ), IGNACIO WINFIELD 1,3 ) and VERÓNICA CORIA-OLVERA 1,4 ) 1 ) Laboratorio de Ecología de Crustáceos, Facultad de Estudios Superiores Iztacala, Universidad Nacional Autónoma de México, Apartado Postal 314, Tlalnepantla, Estado de México 54090, Mexico ABSTRACT This study evaluates the night-time recruitment of Farfantepenaeus aztecus postlarvae and its relation with temperature, salinity, and velocity and flow direction of the current in the artificial inlet of the Camaronera lagoon of Alvarado, Veracruz, the Soto la Marina river inlet of Tamaulipas, and the Puerto Real inlet of Términos lagoon, Campeche. Sampling was done during the night. A conical 243 μm mesh net was used for capture. The total catch of F. aztecus for the three coastal systems was 95,279 postlarvae. The average temperature was 28.23 ± 1.37 ◦ C, salinity 23.54 ± 7.10 psu, and stream velocity 0.39 ± 0.18 m s −1 . During this study, the existence of recruitment peaks between 20:00 h and 02:00 h was established. The correlation coefficient between the density of postlarvae in the hours of sampling and environmental factors was significant (P< 0.05) in all three coastal systems. The highest density appeared when salinity was >23 psu. Recruitment is related to the influx of seawater towards the coastal systems. There was no significant difference in postlarval density (P> 0.05) between consecutive sampling cycles. RESUMEN En este estudio se evaluó el reclutamiento de las postlarvas de Farfantepenaeus aztecus y su relación con la temperatura, salinidad, velocidad y dirección de la corriente en la boca de comunicación artificial de la laguna Camaronera de Alvarado, Veracruz, la boca del río Soto la Marina, Tamaulipas y la boca de comunicación Puerto Real de la laguna de Términos, Campeche. Los muestreos se realizaron durante la noche. Para las capturas se utilizó una red cónica de 243 micras de abertura de malla. La abundancia total de F. aztecus para los tres sistemas fue de 95 279 postlarvas. El promedio de temperatura fue de 28,23 ± 1,37 ◦ C, salinidad de 23,54 ± 7,10 ups y velocidad de corriente de 0,39 ± 0,18 m s −1 . Durante la realización de este estudio se presentaron los picos de reclutamiento entre las 20:00 h y las 02:00. El coeficiente de correlación entre 2 ) e-mail: [email protected] 3 ) e-mail: [email protected] 4 ) e-mail: [email protected] © Koninklijke Brill NV, Leiden, 2009 Crustaceana 82 (4): 425-437 Also available online: www.brill.nl/cr DOI:10.1163/156854008X400612
Transcript of Transport of Farfantepenaeus Aztecus Postlarvae in Three Lagoon-System Inlets in the Southwestern...

TRANSPORT OF FARFANTEPENAEUS AZTECUS POSTLARVAE IN THREELAGOON-SYSTEM INLETS IN THE SOUTHWESTERN GULF OF MEXICO
BY
SERGIO CHÁZARO-OLVERA1,2), IGNACIO WINFIELD1,3) andVERÓNICA CORIA-OLVERA1,4)
1) Laboratorio de Ecología de Crustáceos, Facultad de Estudios Superiores Iztacala, UniversidadNacional Autónoma de México, Apartado Postal 314, Tlalnepantla, Estado de México 54090,
Mexico
ABSTRACT
This study evaluates the night-time recruitment of Farfantepenaeus aztecus postlarvae and itsrelation with temperature, salinity, and velocity and flow direction of the current in the artificial inletof the Camaronera lagoon of Alvarado, Veracruz, the Soto la Marina river inlet of Tamaulipas, andthe Puerto Real inlet of Términos lagoon, Campeche. Sampling was done during the night. A conical243 μm mesh net was used for capture. The total catch of F. aztecus for the three coastal systemswas 95,279 postlarvae. The average temperature was 28.23 ± 1.37◦C, salinity 23.54 ± 7.10 psu, andstream velocity 0.39 ± 0.18 m s−1. During this study, the existence of recruitment peaks between20:00 h and 02:00 h was established. The correlation coefficient between the density of postlarvaein the hours of sampling and environmental factors was significant (P < 0.05) in all three coastalsystems. The highest density appeared when salinity was >23 psu. Recruitment is related to theinflux of seawater towards the coastal systems. There was no significant difference in postlarvaldensity (P > 0.05) between consecutive sampling cycles.
RESUMEN
En este estudio se evaluó el reclutamiento de las postlarvas de Farfantepenaeus aztecus ysu relación con la temperatura, salinidad, velocidad y dirección de la corriente en la boca decomunicación artificial de la laguna Camaronera de Alvarado, Veracruz, la boca del río Soto laMarina, Tamaulipas y la boca de comunicación Puerto Real de la laguna de Términos, Campeche.Los muestreos se realizaron durante la noche. Para las capturas se utilizó una red cónica de243 micras de abertura de malla. La abundancia total de F. aztecus para los tres sistemas fue de95 279 postlarvas. El promedio de temperatura fue de 28,23±1,37◦C, salinidad de 23,54±7,10 upsy velocidad de corriente de 0,39 ± 0,18 m s−1. Durante la realización de este estudio se presentaronlos picos de reclutamiento entre las 20:00 h y las 02:00. El coeficiente de correlación entre
2) e-mail: [email protected]) e-mail: [email protected]) e-mail: [email protected]
© Koninklijke Brill NV, Leiden, 2009 Crustaceana 82 (4): 425-437Also available online: www.brill.nl/cr DOI:10.1163/156854008X400612

426 S. CHÁZARO-OLVERA, I. WINFIELD & V. CORIA-OLVERA
la densidad de postlarvas y los factores ambientales fue significativo (P < 0,05) en los tressistemas costeros. La mayor densidad se registró cuando la salinidad esta por arriba de 23 ups.El reclutamiento también se relacionó con el influjo hacia los sistemas costeros. No se encontrarondiferencias significativas (P > 0,05) entre los ciclos de muestreo consecutivos.
INTRODUCTION
The transport of penaeid postlarvae is a complex process. Suggested mech-anisms include selective tidal transport cued by synchronized salinity changes(Hughes, 1969a, b), endogenous rhythms (Hughes, 1972), or hydrostatic pressure(Forbes & Benfield, 1986; Rothlisberg et al., 1995). Another transport hypothesissuggests that changes in coastal water temperature, salinity, and direction of cur-rents in combination with diel movements of postlarvae facilitate their transport(Hughes, 1969b; Rogers et al., 1993). The interaction of diel vertical migrationwith tidal and wind-forced currents has also been suggested as a possible mode ofrecruitment (Rothlisberg et al., 1983; Wenner et al., 1998).
Several studies have shown evidence that penaeid post-larvae migrate verticallyas a means to enhance recruitment to estuaries (Staples & Vance, 1985; Forbes &Benfield, 1986; Dall et al., 1990). Tidally based endogenous rhythms with peaksof movement around flood tide have been found for several species (Copeland &Truitt, 1966; Garcia & Le Reste, 1981; Mair et al., 1982; Forbes & Benfield, 1986).The position in the water column strongly affects their onshore transport (Blantonet al., 1995). Postlarval shrimp may use tidal transport to facilitate movements intothe estuary, by moving up in the water column during flood tide and by sinking tonear bottom depths at ebb tide, where flow is weakest because of bottom friction,thereby avoiding the stronger ebb currents (Hill, 1991).
Among penaeid shrimp, Hughes (1972) found that vertical movement of post-larval pink shrimp, Farfantepenaeus duorarum (Burkenroad, 1939), was synchro-nized with the tides; however, no evidence of a tidally mediated rhythm could befound for F. aztecus (Ives, 1891) or Litopenaeus setiferus (Linnaeus, 1767) duringstatic salinity gradient experiments (Keiser & Aldrich, 1976).
Our knowledge of larval transport and supply has made considerable progressduring the past two decades, due to the development of sampling gear designed tocapture pre-settlement stages entering the inshore habitats, and to the integration ofbiological and oceanographic data in multidisciplinary studies. Relatively little isknown about the larval supply of tropical fishes and invertebrates of the CaribbeanSea and Gulf of Mexico (GOM) (Criales et al., 2002).
The brown shrimp, Farfantepenaeus aztecus, is a decapod crustacean withsignificant commercial value in Mexico. Brown shrimp occur from Martha’sVineyard, Massachusetts, to the Florida Keys and then westward and southward

FARFANTEPENAEUS AZTECUS POSTLARVAE TRANSPORT 427
in the GOM from Apalachicola Bay, Florida, to the northwestern coast of theYucatan Peninsula. Recruitment of postlarvae occurs during spring through fall(Zimmerman & Minello, 1984; Rozas & Reed, 1993). The objectives of thisstudy were to identify the relationship between F. aztecus post-larval densities andsalinity, temperature, and velocity of currents during their ingress to three estuarineinlets in three Mexican states in the southwestern portion of the Gulf of Mexicoduring autumn.
MATERIAL AND METHODS
Three estuarine inlets in three Mexican states from the southwestern portionof the Gulf of Mexico were sampled in the autumn of 2007. The localitiessurveyed were: Camaronera Lagoon, Alvarado (CV), Veracruz State (18◦50′-18◦52′N 95◦54′-95◦58′W) on 29 and 30 September; Soto la Marina River (SM),Tamaulipas State (23◦45′55′′-23◦46′27′′N 97◦44′05′′-97◦44′21′′W) on 15 and 16October; and Puerto Real, Términos Lagoon (PR), Campeche State (18◦30′-18◦40′N 92◦00′-92◦50′W) on 1 and 2 November (fig. 1). These areas constitutesubsystems transitional between marine and brackish environments, exerting astrong influence on the migration and distribution of organisms and on biodiversitylevels inside the lagoon-estuary systems. The SM inlet is located in the LagunaMadre System, with a mean depth of 1.0 m. This system has a sand barrier islandparallel to the coast that, in conjunction with land drainage, influences the sedimentcomposition of the estuary. The CV inlet is located at the Camaronera lagoonof the Alvarado Lagoonal System, and was built during the 1980’s. It includestwo artificial pipes, which allow for water exchange between the GOM and thelagoon system by simple hydrodynamic effects. This lagoon has a mean depth of2.2 m, and extensive meadows of Ruppia maritima (Linnaeus). The PR inlet, oneof several inlets of Términos lagoon, has a mean depth of 2.0 m and is located atthe eastern portion of the lagoon system. It is a natural channel and the bottom iscovered with Thalassia testudinum Banks & Solander ex Koenig and Halodulebeaudettei (Den Hartog) Den Hartog. The most common sediments are sandylimestone (Cházaro-Olvera et al., 2002).
Two sampling cycles each were run at CV, SM, and PR, to yield a total of sixsampling cycles. Collections were made over a 13 h period with a 1.0-m lengthand 0.5-m diameter conical plankton net constructed of 243 μm mesh. Collectionstarted at 18:00 h and ended at 6:00 h of the following day, as for postlarvaeof decapods the largest recruitment density occurs during the night (Epifanio etal., 1984). Collections were filtered for 15 min. every hour and samples wereplaced in labelled plastic bags with 4% formalin solution. The depth of sampling

428 S. CHÁZARO-OLVERA, I. WINFIELD & V. CORIA-OLVERA
Fig. 1. The locations of the three sampling sites in the Gulf of Mexico.
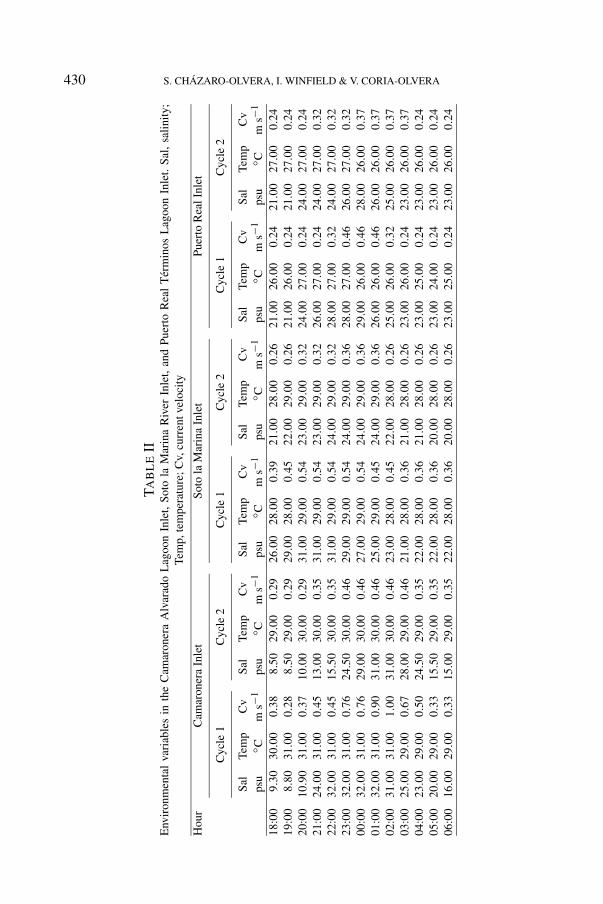
was of 0.85 ± 0.15 m in CV, 1.0 ± 0.20 m in SM, and 0.80 ± 0.10 m in PR.Simultaneously, we recorded water temperature with a Brannan thermometer of−10 to 110◦C, salinity (psu) with a salinometer model YSI 33, and current velocity(m s−1) with a Forestry 2030r6 flowmeter.
Postlarvae were identified according to the descriptions by Chuensri (1968),Ringo & Zamora (1968), and Rocha-Ramírez et al. (1996). Postlarval densitywas standardized as number/1000 m3 following the norms of APHA-AWWA-AWRA (1998), and the homogeneity of variance and normality assumptions ofANOVA were tested for these data and, unless noted, were met prior to each test.Correlations between density vs. water temperature, salinity, or current velocitywere tested with Pearson’s correlation analysis and multiple regression analysis.We conducted two separate one-way ANOVAs; the first comparison was for log10
density between two consecutive sampling days at each site, and the second forlog10 density between the samples of the three inlets (Sokal & Rohlf, 1995). Allstatistics were calculated with SigmaStat for Windows (Ver. 3.1, Jandel Corp.). Allcomparisons were considered significant when P < 0.05.
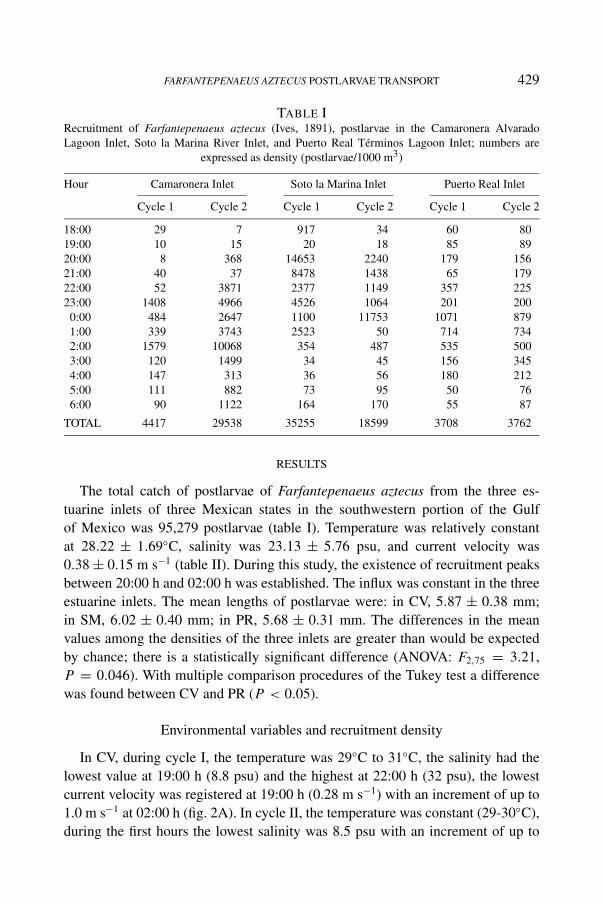
FARFANTEPENAEUS AZTECUS POSTLARVAE TRANSPORT 429
TABLE IRecruitment of Farfantepenaeus aztecus (Ives, 1891), postlarvae in the Camaronera AlvaradoLagoon Inlet, Soto la Marina River Inlet, and Puerto Real Términos Lagoon Inlet; numbers are
expressed as density (postlarvae/1000 m3)
Hour Camaronera Inlet Soto la Marina Inlet Puerto Real Inlet
Cycle 1 Cycle 2 Cycle 1 Cycle 2 Cycle 1 Cycle 2
18:00 29 7 917 34 60 8019:00 10 15 20 18 85 8920:00 8 368 14653 2240 179 15621:00 40 37 8478 1438 65 17922:00 52 3871 2377 1149 357 22523:00 1408 4966 4526 1064 201 200
0:00 484 2647 1100 11753 1071 8791:00 339 3743 2523 50 714 7342:00 1579 10068 354 487 535 5003:00 120 1499 34 45 156 3454:00 147 313 36 56 180 2125:00 111 882 73 95 50 766:00 90 1122 164 170 55 87
TOTAL 4417 29538 35255 18599 3708 3762
RESULTS
The total catch of postlarvae of Farfantepenaeus aztecus from the three es-tuarine inlets of three Mexican states in the southwestern portion of the Gulfof Mexico was 95,279 postlarvae (table I). Temperature was relatively constantat 28.22 ± 1.69◦C, salinity was 23.13 ± 5.76 psu, and current velocity was0.38 ± 0.15 m s−1 (table II). During this study, the existence of recruitment peaksbetween 20:00 h and 02:00 h was established. The influx was constant in the threeestuarine inlets. The mean lengths of postlarvae were: in CV, 5.87 ± 0.38 mm;in SM, 6.02 ± 0.40 mm; in PR, 5.68 ± 0.31 mm. The differences in the meanvalues among the densities of the three inlets are greater than would be expectedby chance; there is a statistically significant difference (ANOVA: F2,75 = 3.21,P = 0.046). With multiple comparison procedures of the Tukey test a differencewas found between CV and PR (P < 0.05).
Environmental variables and recruitment density
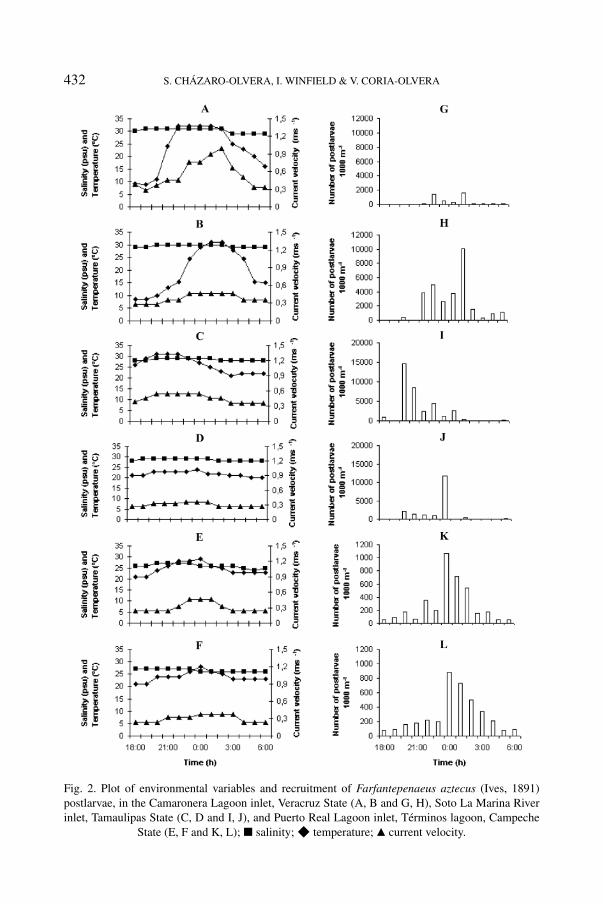
In CV, during cycle I, the temperature was 29◦C to 31◦C, the salinity had thelowest value at 19:00 h (8.8 psu) and the highest at 22:00 h (32 psu), the lowestcurrent velocity was registered at 19:00 h (0.28 m s−1) with an increment of up to1.0 m s−1 at 02:00 h (fig. 2A). In cycle II, the temperature was constant (29-30◦C),during the first hours the lowest salinity was 8.5 psu with an increment of up to
430 S. CHÁZARO-OLVERA, I. WINFIELD & V. CORIA-OLVERA
TA
BL
EII
Env
iron
men
tal
vari
able
sin
the
Cam
aron
era
Alv
arad
oL
agoo
nIn
let,
Soto
laM
arin
aR
iver
Inle
t,an
dPu
erto
Rea
lT
érm
inos
Lag
oon
Inle
t.Sa
l,sa
linity
;Te
mp,
tem
pera
ture
;Cv,
curr
entv
eloc
ity
Hou
rC
amar
oner
aIn
let
Soto
laM
arin
aIn
let
Puer
toR
ealI
nlet
Cyc
le1
Cyc
le2
Cyc
le1
Cyc
le2
Cyc
le1
Cyc
le2
Sal
Tem
pC
vSa
lTe
mp
Cv
Sal
Tem
pC
vSa
lTe
mp
Cv
Sal
Tem
pC
vSa
lTe
mp
Cv
psu
◦ Cm
s−1
psu
◦ Cm
s−1
psu
◦ Cm
s−1
psu
◦ Cm
s−1
psu
◦ Cm
s−1
psu
◦ Cm
s−1
18:0
09.
3030
.00
0.38
8.50
29.0
00.
2926
.00
28.0
00.
3921
.00
28.0
00.
2621
.00
26.0
00.
2421
.00
27.0
00.
2419
:00
8.80
31.0
00.
288.
5029
.00
0.29
29.0
028
.00
0.45
22.0
029
.00
0.26
21.0
026
.00
0.24
21.0
027
.00
0.24
20:0
010
.90
31.0
00.
3710
.00
30.0
00.
2931
.00
29.0
00.
5423
.00
29.0
00.
3224
.00
27.0
00.
2424
.00
27.0
00.
2421
:00
24.0
031
.00
0.45
13.0
030
.00
0.35
31.0
029
.00
0.54
23.0
029
.00
0.32
26.0
027
.00
0.24
24.0
027
.00
0.32
22:0
032
.00
31.0
00.
4515
.50
30.0
00.
3531
.00
29.0
00.
5424
.00
29.0
00.
3228
.00
27.0
00.
3224
.00
27.0
00.
3223
:00
32.0
031
.00
0.76
24.5
030
.00
0.46
29.0
029
.00
0.54
24.0
029
.00
0.36
28.0
027
.00
0.46
26.0
027
.00
0.32
00:0
032
.00
31.0
00.
7629
.00
30.0
00.
4627
.00
29.0
00.
5424
.00
29.0
00.
3629
.00
26.0
00.
4628
.00
26.0
00.
3701
:00
32.0
031
.00
0.90
31.0
030
.00
0.46
25.0
029
.00
0.45
24.0
029
.00
0.36
26.0
026
.00
0.46
26.0
026
.00
0.37
02:0
031
.00
31.0
01.
0031
.00
30.0
00.
4623
.00
28.0
00.
4522
.00
28.0
00.
2625
.00
26.0
00.
3225
.00
26.0
00.
3703
:00
25.0
029
.00
0.67
28.0
029
.00
0.46
21.0
028
.00
0.36
21.0
028
.00
0.26
23.0
026
.00
0.24
23.0
026
.00
0.37
04:0
023
.00
29.0
00.
5024
.50
29.0
00.
3522
.00
28.0
00.
3621
.00
28.0
00.
2623
.00
25.0
00.
2423
.00
26.0
00.
2405
:00
20.0
029
.00
0.33
15.5
029
.00
0.35
22.0
028
.00
0.36
20.0
028
.00
0.26
23.0
024
.00
0.24
23.0
026
.00
0.24
06:0
016
.00
29.0
00.
3315
.00
29.0
00.
3522
.00
28.0
00.
3620
.00
28.0
00.
2623
.00
25.0
00.
2423
.00
26.0
00.
24

FARFANTEPENAEUS AZTECUS POSTLARVAE TRANSPORT 431
31 psu at 01:00 h, the current velocity registered was 0.29 m s−1 at 18:00 h and0.46 m s−1 at 23:00 h (fig. 2B). One recruitment peak was obtained in each cycle at18:00 and 22:00 h, and a total of 33,955 post-larvae was obtained. During cycle I,the lowest density of Farfantepenaeus aztecus was 8 postlarvae/1000 m3 at 20:00 h,whereas the highest value was 1579 postlarvae/1000 m3 at 02:00 h (fig. 2G).In cycle II, the lowest density was found at 18:00 h with 7 postlarvae/1000 m3,and the highest density was registered at 02:00 h with 10,068 postlarvae/1000 m3
(fig. 2H). A significant, multiple correlation was found for both cycle I (r2 =0.91, P < 0.05) and cycle II (r2 = 0.81, P < 0.05). On the other hand,variance analysis (ANOVA) used to compare density between both cycles revealedsignificant differences as well (F1,24, P < 0.05).
In SM, during cycle I, the temperature was constant (28-29◦C), the lowestsalinity was registered at 03:00 h (21 psu) and the highest was registered at20:00 h (31 psu), current velocity was 0.36 m s−1 as lowest value and there wasa high of 0.54 m s−1 (fig. 2C). In cycle II, the temperature was 28◦C to 29◦C, thelowest salinity was registered at 05:00 h with 20 psu and the highest at around22:00 h with 24 psu, the current velocity was 0.32-0.36 m s−1, diminishing to0.26 m s−1 at 04:00 h (fig. 2D). One recruitment peak was obtained in eachcycle, and a total of 53,854 postlarvae was obtained. During cycle I, the lowestdensity was 20 postlarvae/1000 m3 at 19:00 h, whereas the highest value was14,653 postlarvae/1000 m3 at 20:00 h (fig. 2I). In cycle II, the lowest densitywas found at 19:00 h with 18 postlarvae/1000 m3 and the highest density wasrecorded at 00:00 h with 11,753 postlarvae/1000 m3 (fig. 2J). A significant,multiple correlation was found for both cycle I (r2 = 0.86, P < 0.05) and cycle II(r2 = 0.86, P < 0.05). On the other hand, variance analysis (ANOVA) used tocompare both cycles did not evidence significant differences (F1,24, P > 0.34).
In PR, during cycle I, the highest temperature was 27◦C at 20:00 h, and thelowest 24◦C at around 05:00 h; salinity showed an increasing tendency, with thelowest of 21 psu between 18:00 and 19:00 h, whereas the highest value of 29 psuwas recorded at 00:00 h. Initially, the current velocity recorded was 0.24 m s−1
from 18:00 h to 21:00 h, showing an increment starting at 22:00 h with 0.32 m s−1
(fig. 2E). In cycle II, the temperature was from 26◦C to 27◦C, the lowest salinitywas registered at 18:00 h with 21 psu and the highest at around 00:00 h with29 psu, the current velocity was 0.24 m s−1 from 18:00 to 21:00 h and 0.32-0.46 m s−1 from 22:00 at 02:00, diminishing to 0.24 m s−1 at 03:00 h (fig. 2F).One recruitment peak was recorded in each cycle, and a total of 7,470 postlarvaewas obtained. During cycle I, the lowest density was 50 postlarvae/1000 m3 at05:00 h, whereas the highest value was 1,071 postlarvae/1000 m3 at 00:00 h(fig. 2K). In cycle II, the lowest density was found at 05:00 h with 76 post-larvae/1000 m3 and the highest density was recorded at 00:00 h with 879 post-larvae/1000 m3 (fig. 2L). A significant, multiple correlation was found for both
432 S. CHÁZARO-OLVERA, I. WINFIELD & V. CORIA-OLVERA
Fig. 2. Plot of environmental variables and recruitment of Farfantepenaeus aztecus (Ives, 1891)postlarvae, in the Camaronera Lagoon inlet, Veracruz State (A, B and G, H), Soto La Marina Riverinlet, Tamaulipas State (C, D and I, J), and Puerto Real Lagoon inlet, Términos lagoon, Campeche
State (E, F and K, L); 2 salinity; F temperature; Q current velocity.

FARFANTEPENAEUS AZTECUS POSTLARVAE TRANSPORT 433
cycle I (r2 = 0.78, P < 0.05) and cycle II (r2 = 0.93, P < 0.05). On the otherhand, variance analysis (ANOVA) used to compare both cycles did not evidencesignificant differences (F1,24, P > 0.97).
DISCUSSION
Salinity, temperature, and current velocity change during the flood and ebbtides in the night, and greatly influenced the densities of Farfantepenaeus aztecuspostlarvae according to time of sampling in the three inlets along the southwesternGulf of Mexico during the period of our study. The shoreward transport of post-larvae occurs via shoreward propagating surface convergences; hence, transportwill only occur when the larvae are at the surface. Around the time of darknessof the new moon, larvae that migrate to the ocean surface may be transportedshoreward in convergences. In contrast, around the time of the full moon shrimppostlarvae may avoid the more illuminated surface waters, in which case theywould not be transported shoreward (Shanks, 1998). On September 29 and 30,illuminated fractions were 0.826, with 3.1 days before the last quarter, and 0.727,with 2.1 days before the last quarter, respectively; on October 15 and 16, thehighest density was recorded, and the phase of the moon was 0.213 illuminatedfraction with 3.1 days before the first quarter, and 0.295 illuminated fraction, with2.1 days before the first quarter, respectively; on November 1 and 2 illuminatedfractions were 0.450, with 0.4 days after the last quarter, and 0.351, with 1.4 daysafter the last quarter, respectively.
The relationship between water temperature and the ingress of post-larvalpenaeids has been noted for both Farfantepenaeus aztecus and Litopenaeussetiferus along the coasts of the Gulf of Mexico. The arrival of F. aztecus toGulf coast estuaries has been found to occur at sustained temperatures >20◦Cin Louisiana (St. Amant et al., 1963) and between 18 and 22◦C in Galveston Bay(Baxter & Renfro, 1966). Rogers et al. (1993) proposed that a complex behaviouralresponse of F. aztecus to the cues of temperature and salinity facilitates movementto the inlet, pooling of larvae, and ingress to the estuary. Similar interactions ofbehaviour and temperature, in conjunction with wind and tidal currents, appear toaid postlarval transport of Litopenaeus vannamei (Boone, 1931) into estuaries ofthe southeastern U.S.A. (Muncy, 1984; Wenner et al., 2005). At an inlet in SouthCarolina, postlarval ingress occurred only when temperatures exceeded 20◦C, withpeak ingress from 25 to 30◦C (DeLancey et al., 1994). Wenner & Beatty (1993)also noted that the peak occurrence of white shrimp in estuarine nursery areas ofSouth Carolina coincided with July temperatures >26◦C. Cool water temperatureshave been shown to elicit reduced activity (at 15-17◦C) of F. aztecus (cf. Aldrich et

434 S. CHÁZARO-OLVERA, I. WINFIELD & V. CORIA-OLVERA
al., 1968). Data from this study and from others (Williams, 1955; Bearden, 1961;Christmas et al., 1966; DeLancey et al., 1994; Wenner et al., 2005) support the ideathat flood tides and salinity are factors that influence postlarval recruitment.
Interactions of temperature and salinity have been suspected of triggeringvertical movement of shrimp postlarvae, whose position in the water columnstrongly affects their onshore transport (Hughes, 1969b; Blanton et al., 1995).Rogers et al. (1993) hypothesized that if they encountered low salinity waterfrom ebb flows in the near-shore coastal zone, F. aztecus postlarvae would movedownward in the water column to a point where transport is minimal, or drop ontothe substrate itself. It has been reported that, in crab megalopae, the rate of salinitychange is a cue for behaviour (DeVries et al., 1994).
This study suggests that postlarval shrimp may use tidal transport to facilitatemovements into the estuary, by moving up in the water column during flood tideand sinking to near bottom depths at ebb tide, where flow is weakest because ofbottom friction, thereby avoiding the stronger ebb currents (Hill, 1991). The resultsof our study in the three inlets indicate a relationship between F. aztecus postlarvaldensity and salinity, as well as current velocity.
Densities of brown shrimp postlarvae were high in the north, and smaller valueswere found in the south, coinciding with reports by Castro (1990) for SM, Molina(1994) for CV, and Signoret (1974) for PR in the Gulf of Mexico.
Large volume exchanges can occur when continental shelf waves pass throughthe system and cause an influx and efflux into and from estuaries by means ofwater level change (Schwing et al., 1988). In this study, we found that 100% of thepostlarvae ingress during the influx period.
Wenner et al. (2005) reported that the density of L. setiferus ranged from130 postlarvae/1000 m3 to 2990 postlarvae/1000 m3 in surface nets at night-timeflooding tides. We found, in our study, densities, ranging from 7 postlarvae/1000m3 to 14,653 postlarvae/1000 m3.
ACKNOWLEDGMENTS
Funding for this research was provided by CONACYT and the Sistema Nacionalde Investigadores, PAPCA 2006-2007 and PAPIME EN203804 projects of theUNAM. We acknowledge numerous students at the UNAM-Iztacala campus fortheir assistance in this project.
REFERENCES
ALDRICH, D. V., C. E. WOOD & K. N. BAXTER, 1968. An ecological interpretation of lowtemperature responses in Penaeus aztecus and P. setiferus postlarvae. Bull. mar. Sci., 18(1):61-71.

FARFANTEPENAEUS AZTECUS POSTLARVAE TRANSPORT 435
APHA-AWWA-AWRA, 1998. Standard methods for the examination of water and wastewater(20th ed.): 1-1090. (American Public Health Association, Washington, DC).
BAXTER, K. N. & W. C. RENFRO, 1966. Seasonal occurrence and size distribution of postlarvalbrown and white shrimp near Galveston, Texas, with notes on species identification. Fish. Bull,U.S. Fish Wildlife Serv., 66: 148-158.
BEARDEN, C. M., 1961. Notes on post-larvae of commercial shrimp (Penaeus) in South Carolina.Contrib. Bears Bluff Lab., 33: 1-8.
BLANTON, J. O., E. L. WENNER, F. E. WERNER & D. M. KNOTT, 1995. Effects of wind-generated coastal currents on the transport of blue crab megalopae on a shallow watercontinental shelf. Bull. mar. Sci., 57: 739-752.
CASTRO, M. R. G., A. M. MEDELLÍN, E. C. BONILLA, T. E. ROSAS, N. R. ORTA &G. E. CONDE, 1990. Incidencia de postlarvas de camarón café Penaeus aztecus, en la Lagunade Almagre, Tamaulipas y la relación con las poblaciones de alta mar en el noreste del Golfode México. Ciencia Pesquera, 7: 121-141.
CHÁZARO-OLVERA, S., I. WINFIELD, M. ORTIZ & F. ÁLVAREZ, 2002. Peracarid crustaceansfrom three inlets in the southwestern Gulf of Mexico: new records and range extensions.Zootaxa, 123: 1-16.
CHUENSRI, C., 1968. A morphometric and meristic study of postlarval brown shrimp, Penaeusaztecus Ives, pink shrimp, P. duorarum Burkenroad, and white shrimp, P. setiferus (Linnaeus):1-108. (M. Sc. Thesis, Univ. Miami, Coral Gables, Florida).
CHRISTMAS, J. Y., G. GUNTER & P. MUSGRAVE, 1966. Studies of annual abundance of postlarvalpenaeid shrimp in estuarine waters of the Mississippi, as related to subsequent commercialcatches. Gulf Res. Rep., 2: 177-212.
COPELAND, B. J. & M. V. TRUITT, 1966. Fauna of the Aransas Pass Inlet, Texas. I. Penaeid shrimppost-larvae. Texas Journ. Sci., 18: 65-74.
CRIALES, M. M., C. YEUNG, F. AMAYA, A. C. LOPEZ, D. L. JONES & W. J. RICHARDS, 2002.Larval supply of fishes, shrimps, and crabs into the nursery ground of the Ciénaga Grande deSanta Marta, Colombian Caribbean. Caribbean Journ. Sci., 38(1-2): 52-65.
DALL, W., B. J. HILL, P. C. ROTHLISBERG & D. S. STAPLES, 1990. The biology of the Penaeidae.Advances in Marine Biology, 27: 1-489. (Academic Press, San Diego, California).
DELANCEY, L. B., J. E. JENKINS & J. D. WHITAKER, 1994. Results of long-term, seasonalsampling for Penaeus post-larvae at Breach Inlet, South Carolina. Fish. Bull., U.S., 92: 633-640.
DEVRIES, M. C., R. A. TANKERSLEY, R. B. FORWARD, JR., W. W. KIRBY-SMITH &R. A. LUETTICH, JR., 1994. The abundance of estuarine crab larvae is associated with hy-drologic variables. Mar. Biol., Berlin, 118: 403-413.
EPIFANIO, C. E., C. C. VALENTI & A. E. PEMBROKE, 1984. Dispersal and recruitment of bluecrab larvae in Delaware Bay, U.S.A. Estuarine, Coastal and Shelf Science, 18(1): 1-12.
FORBES, A. T. & M. C. BENFIELD, 1986. Tidal behaviour of postlarval penaeid prawns (Crustacea:Decapoda: Penaeidae) in a southeast African estuary. Journ. exp. mar. Biol. Ecol., 102: 23-34.
GARCIA, S. & L. LE RESTE, 1981. Life cycles, dynamics, exploitation and management of coastalpenaeid shrimp stocks. FAO Fish. Rep., 203: 1-215.
HILL, A. E., 1991. Vertical migration in tidal currents. Mar. Ecol. Prog. Ser., 75: 39-54.HUGHES, D. A., 1969a. On the mechanisms underlying tide-associated movements of Penaeus
duorarum Burkenroad. FAO Fish. Rep., 57: 867-874.— —, 1969b. Responses to salinity change as a tidal transport mechanism of pink shrimp, Penaeus
duorarum. Biol. Bull., Woods Hole, 136: 45-53.— —, 1972. On the endogenous control of tide-associated displacements of pink shrimp, Penaeus
duorarum Burkenroad. Biol. Bull., Woods Hole, 142: 271-280.

436 S. CHÁZARO-OLVERA, I. WINFIELD & V. CORIA-OLVERA
KEISER, R. K., JR. & D. V. ALDRICH, 1976. Salinity preference of postlarval brown and whiteshrimp (Penaeus aztecus and P. setiferus) in gradient tanks. Department of Wildlife FisheriesScientific Publication, Texas Agricultural Experimental Station, Texas (TAMU-SG-75–208).
MAIR, J. MCD., J. L. WATKINS & D. I. WILLIAMSON, 1982. Factors affecting the immigrationof postlarval penaeid shrimp into a Mexican lagoon system. In: Proceedings of an InternationalSymposium on Coastal Lagoons. SCOR/IABO/UNESCO, Bordeaux, France. OceanologicaActa, (Special Suppl.) 4: 339-345.
MOLINA, B. H. M., 1994. Distribución espacio — temporal de la fase postlarvaria del géneroPenaeus spp. asociadas a Ruppia marítima en el Sistema Lagunar de Alvarado, Veracruz: 1-54.(Professional Thesis of Biology, UNAM).
MUNCY, R. J., 1984. Species profiles: life histories and environmental requirements of coastalfishes and invertebrates (South Atlantic) — white shrimp. U.S.A. Fish and Wildlife Service,FWS/OBS-82/11.27. U.S.A. Army Corps of Engineers, TR EL-82-4: 1-19.
RINGO, R. D. & G. ZAMORA, JR., 1968. A penaeid postlarval character of taxonomic value. Bull.mar. Sci., 18: 471-476.
ROCHA-RAMÍREZ, A., S. CHÁZARO-OLVERA, R. ROMÁN-CONTRERAS & H. MOLINA, 1996.Clave de identificación para estadios de zoea, mysis, postlarvas (Caridea y Penaidea) ymegalopas (Anomura y Brachyura) de la Laguna de Alvarado, Veracruz México. Revista deZoología, Universidad Nacional Autónoma de México, (Numero Especial) 1: 1-22.
ROGERS, B. D., R. F. SHAW, W. H. HERKE & R. H. BLANCHET, 1993. Recruitment of postlarvaland juvenile brown shrimp (Penaeus aztecus Ives) from offshore to estuarine waters of thenorthwestern Gulf of Mexico. Estuar. coast. Shelf Sci., 36: 377-394.
ROTHLISBERG, P. C., J. A. CHURCH & C. B. FANDRY, 1995. A mechanism for near-shoreconcentration and estuarine recruitment of post-larval Penaeus plebejus Hess (Decapoda,Penaeidae). Estuar. coast. Shelf Sci., 40: 115-138.
ROTHLISBERG, P. C., J. A. CHURCH & A. M. G. FORBES, 1983. Modeling the advection ofvertically migrating shrimp larvae. Journ. mar. Res., 41: 511-554.
ROZAS, L. P. & D. J. REED, 1993. Nekton use of marsh-surface habitats in Louisiana (U.S.A.)deltaic salt marshes undergoing submergence. Mar. Ecol. Prog. Ser., 96: 147-157.
SCHWING, F. B., L. Y. OEY & J. O. BLANTON, 1988. Evidence for non-local forcing along thesoutheastern United States during a transitional wind regime. Journ. Geol. Res., 93: 8221-8228.
SHANKS, A. L., 1998. Abundance of post-larval Callinectes sapidus, Peneaus spp., Uca spp., andLibinia spp. collected at an outer coastal site and their cross-shelf transport. Mar. Ecol. Prog.Ser., 168: 57-69.
SIGNORET, M., 1974. Abundancia, tamaño y distribución de camarones (Crustacea, Penaeidae) dela Laguna de Términos, Campeche y su relación con algunos factores hidrológicos. Anales delInstituto de Biología, UNAM, (Zoologia), 45(1): 119-140.
SOKAL, R. R. & F. J. ROHLF, 1995. Biometry (3rd ed.): 1-887. (W. H. Freeman and Company, NewYork).
ST. AMANT, L. S., K. C. CORKUM & J. G. BROOM, 1963. Studies on growth dynamics of thebrown shrimp, Penaeus aztecus, in Louisiana waters. Ann. Session Gulf Caribb. Fish. Inst.Proc., 15: 14-26.
STAPLES, D. J. & D. J. VANCE, 1985. Short-term and longterm influences on the immigrationof postlarval banana prawns Penaeus merguiensis, into a mangrove estuary of the Gulf ofCarpentaria, Australia. Mar. Ecol. Prog. Ser., 23: 15-29.
WENNER, E. L. & H. R. BEATTY, 1993. Utilization of shallow estuarine habitats in South Carolina,USA, by postlarval and juvenile stages of Penaeus spp. (Decapoda: Penaeidae). Journ. Crust.Biol., 13: 280-295.
WENNER, E. L., D. M. KNOTT, C. A. BARANS, S. WILDE, J. O. BLANTON & J. AMFT, 2005.Key factors influencing transport of white shrimp (Litopenaeus setiferus) post-larvae into theOssabaw Sound system, Georgia, USA. Fish. Oceanography, 14(3): 175-194.

FARFANTEPENAEUS AZTECUS POSTLARVAE TRANSPORT 437
WENNER, E. L., D. M. KNOTT, J. O. BLANTON, C. A. BARANS & J. AMFT, 1998. Roles oftidal and wind-generated currents in transporting white shrimp (Penaeus setiferus) post-larvaethrough a South Carolina (USA) inlet. Journ. Plankton Res., 20: 2333-2356.
WILLIAMS, A. B., 1955. A contribution to the life histories of commercial shrimps (Penaeidae) inNorth Carolina. Bull. mar. Sci., 5: 116-146.
ZIMMERMAN, R. J. & T. J. MINELLO, 1984. Densities of Penaeus aztecus, P. setiferus and othernatant macrofauna in a Texas salt marsh. Estuaries, 7: 421-433.
First received 17 March 2008.Final version accepted 8 August 2008.




















