
Transgenic Tomato Plants Expressing the Arabidopsis NPR1 Gene Display Enhanced Resistance to a...
15
Transgenic tomato plants expressing the Arabidopsis NPR1 gene display enhanced resistance to a spectrum of fungal and bacterial diseases Wan-Chi Lin 1 , Ching-Fang Lu 1 , Jia-Wei Wu 1 , Ming-Lung Cheng 1 , Yu-Mei Lin 1 , Ning-Sun Yang, Lowell Black 2 , Sylvia K. Green 2 , Jaw-Fen Wang 2 & Chiu-Ping Cheng 1, * 1 Institute of BioAgricultural Sciences, Academia Sinica, Taipei, Taiwan 115, R. O. C. 2 AVRDC-The World Vegetable Center, P.O. Box 42, Shanhua, Tainan, Taiwan 741, R. O. C. Received 26 August 2003; accepted 16 June 2004 Key words: Arabidopsis NPR1, disease resistance, PR genes, tomato Abstract Development of effective disease-resistance to a broad-range of pathogens in crops usually requires tre- mendous resources and effort when traditional breeding approaches are taken. Genetic engineering of disease-resistance in crops has become popular and valuable in terms of cost and efficacy. Due to long- lasting and broad-spectrum of effectiveness against pathogens, employment of systemic acquired resis- tance (SAR) for the genetic engineering of crop disease-resistance is of particular interest. In this report, we explored the potential of using SAR-related genes for the genetic engineering of enhanced resistance to multiple diseases in tomato. The Arabidopsis NPR1 (nonexpresser of PR genes) gene was introduced into a tomato cultivar, which possesses heat-tolerance and resistance to tomato mosaic virus (ToMV). The transgenic lines expressing NPR1 were normal as regards overall morphology and horticultural traits for at least four generations. Disease screens against eight important tropical diseases revealed that, in addition to the innate ToMV-resistance, the tested transgenic lines conferred significant level of enhanced resistance to bacterial wilt (BW) and Fusarium wilt (FW), and moderate degree of enhanced resistance to gray leaf spot (GLS) and bacterial spot (BS). Transgenic lines that accumulated higher levels of NPR1 proteins exhibited higher levels and a broader spectrum of enhanced resistance to the diseases, and enhanced disease-resistance was stably inherited. The spectrum and degree of these NPR1- transgenic lines are more significant compared to that of transgenic tomatoes reported to date. These transgenic lines may be further explored as future tomato stocks, aiming at building up resistance to a broader spectrum of diseases. Introduction Genetic engineering of disease-resistance through transferal of plant defense-related genes or patho- gen-originated genes into crops is valuable in terms of cost, efficacy and reduction of pesticide usage (Shah, 1997; Salmeron & Vernooij, 1998; Rommens & Kishore, 2000; Stuiver & Custers, 2001). Among the strategies used for the genetic engineering of disease-resistance, the employment of SAR is of special interest. In an incompatible plant-pathogen interaction, in addition to the immediate activation of a whole complex defense network and hypersensitive response blocking pathogen growth at the site of initial infection, SAR is established in uninfected parts of the plant for further resistance to a broad-spectrum of pathogens (Ryals et al., 1996). SAR is long lasting and often associated with local and systemic accu- mulation of salicylic acid (SA) (Malamy et al., 1990; Me´ traux et al., 1990; Rasmussen et al., *Author for correspondence E-mail: [email protected] Transgenic Research 13: 567–581, 2004. Ó 2004 Kluwer Academic Publishers. Printed in the Netherlands. 567
Transcript of Transgenic Tomato Plants Expressing the Arabidopsis NPR1 Gene Display Enhanced Resistance to a...

Transgenic tomato plants expressing the Arabidopsis NPR1 gene display
enhanced resistance to a spectrum of fungal and bacterial diseases
Wan-Chi Lin1, Ching-Fang Lu1, Jia-Wei Wu1, Ming-Lung Cheng1, Yu-Mei Lin1, Ning-Sun
Yang, Lowell Black2, Sylvia K. Green2, Jaw-Fen Wang2 & Chiu-Ping Cheng1,*1Institute of BioAgricultural Sciences, Academia Sinica, Taipei, Taiwan 115, R. O. C.2AVRDC-The World Vegetable Center, P.O. Box 42, Shanhua, Tainan, Taiwan 741, R. O. C.
Received 26 August 2003; accepted 16 June 2004
Key words: Arabidopsis NPR1, disease resistance, PR genes, tomato
Abstract
Development of effective disease-resistance to a broad-range of pathogens in crops usually requires tre-mendous resources and effort when traditional breeding approaches are taken. Genetic engineering ofdisease-resistance in crops has become popular and valuable in terms of cost and efficacy. Due to long-lasting and broad-spectrum of effectiveness against pathogens, employment of systemic acquired resis-tance (SAR) for the genetic engineering of crop disease-resistance is of particular interest. In this report,we explored the potential of using SAR-related genes for the genetic engineering of enhanced resistanceto multiple diseases in tomato. The Arabidopsis NPR1 (nonexpresser of PR genes) gene was introducedinto a tomato cultivar, which possesses heat-tolerance and resistance to tomato mosaic virus (ToMV).The transgenic lines expressing NPR1 were normal as regards overall morphology and horticulturaltraits for at least four generations. Disease screens against eight important tropical diseases revealedthat, in addition to the innate ToMV-resistance, the tested transgenic lines conferred significant level ofenhanced resistance to bacterial wilt (BW) and Fusarium wilt (FW), and moderate degree of enhancedresistance to gray leaf spot (GLS) and bacterial spot (BS). Transgenic lines that accumulated higherlevels of NPR1 proteins exhibited higher levels and a broader spectrum of enhanced resistance to thediseases, and enhanced disease-resistance was stably inherited. The spectrum and degree of these NPR1-transgenic lines are more significant compared to that of transgenic tomatoes reported to date. Thesetransgenic lines may be further explored as future tomato stocks, aiming at building up resistance to abroader spectrum of diseases.
Introduction
Genetic engineering of disease-resistance throughtransferal of plant defense-related genes or patho-gen-originated genes into crops is valuable interms of cost, efficacy and reduction of pesticideusage (Shah, 1997; Salmeron & Vernooij, 1998;Rommens & Kishore, 2000; Stuiver & Custers,2001). Among the strategies used for the genetic
engineering of disease-resistance, the employmentof SAR is of special interest. In an incompatibleplant-pathogen interaction, in addition to theimmediate activation of a whole complex defensenetwork and hypersensitive response blockingpathogen growth at the site of initial infection,SAR is established in uninfected parts of the plantfor further resistance to a broad-spectrum ofpathogens (Ryals et al., 1996). SAR is long lastingand often associated with local and systemic accu-mulation of salicylic acid (SA) (Malamy et al.,1990; Metraux et al., 1990; Rasmussen et al.,
*Author for correspondenceE-mail: [email protected]
Transgenic Research 13: 567–581, 2004.� 2004 Kluwer Academic Publishers. Printed in the Netherlands.
567

1991) and induced expression of a number ofgenes, including a group of pathogenesis-related(PR) genes (Ward et al., 1991; Ryals et al., 1996).Overexpression of some PR genes in transgenicplants conferred modest protection against patho-gens (Broglie et al., 1991; Alexander et al., 1993;Liu et al., 1994; Zhu et al., 1994; Jach et al.,1995). However, the protection provided by a sin-gle specific PR gene is usually very limited in itsspectrum, degree and duration, compared to thatof a native SAR response (Jach et al., 1995; Jon-gedijk et al., 1995). Therefore, more durable resis-tance to a broader spectrum of pathogens may beexpected from engineering defense reactions thatare more closely related to the natural SARdefense mechanisms employed in most incompati-ble plant-pathogen interactions.
To elucidate the mechanism of SAR and iden-tify important genes involved in this response,extensive efforts were spent on genetic screens ofArabidopsis mutants (Clarke et al., 1998). Partic-ularly, screens of Arabidopsis mutants defectivein response to the SA signal have led to identifi-cation of NPR1 (also known as NIM1 or SAI1)(Cao et al., 1994; Delaney et al., 1995; Glaze-brook et al., 1996; Shah et al., 1997). NPR1encodes a protein that contains multiple ankyrinrepeat domains that is found in some regulatoryproteins (Cao et al., 1997; Ryals et al., 1997).The NPR1 gene product is localized in thenucleus (Kinkema et al., 2000) where it physicallyand differentially interacts with Arabidopsis TGAfamily members of basic region, leucine zipper(bZIP) transcription factors (Zhang et al., 1999;Despres et al., 2000; Zhou et al., 2000; Subr-amaniam et al., 2001; Fan & Dong, 2002; Kim &Delaney, 2002). Under normal conditions, NPR1expresses at a very low level; however, upontreatment of plants with SA, 2,6-dichloroisoni-cotinic acid (INA) or benzo [1,2,3] thiadiazole-7-carbothioic acid S-methyl ester (BTH), the levelof NPR1 gene expression is enhanced (Cao et al.,1997; Ryals et al., 1997), and regulated throughSA-induced WRKY DNA binding proteins (Yuet al., 2001) and redox changes (Mou et al.,2003). In fact, it has been shown that priming ofdefense gene activation by chemical SAR induc-ers requires NPR1 (Conrath et al., 2002; Kohleret al., 2002). In addition, NPR1 is also involvedin jasmonic acid (JA)/ethylene-dependent inducedsystemic resistance (ISR) shared by some
Pseudomonas flourescens strains (Pieterse et al.,1998), and has been shown to modulate crosstalk between salicylate- and jasmonate-dependentdefense pathways (Spoel et al., 2003). Therefore,the NPR1 protein is a key regulator in the trans-duction of SA and JA/ethylene signals leading togeneral acquired resistance responses. Overex-pression of NPR1 in Arabidopsis has beenshown to lead to a non-specific resistance toPeronospora parasitica and Pseudomonas syringaepv. maculicola in a dosage-dependent fashion,without obvious detrimental effects on the plants(Cao et al., 1998). Overexpression of NPR1 alsoresults in enhanced efficacy of fungicides inArabidopsis (Friedrich et al., 2001). In addi-tion, transgenic rice plants overexpressing theArabidopsis NPR1 gene exhibit enhanced resis-tance to the rice bacterial blight pathogen Xan-thomonas oryzae pv. oryzae (Chern et al., 2001).A disease-resistance pathway similar to that ofthe Arabidopsis NPR1-mediated signaling path-way was demonstrated in rice, suggesting a con-served signal transduction pathway controllingNPR1-mediated resistance shared by monocotand dicot plants (Zhang et al., 1999; Chern et al.,2001). In addition to its key role in activatingSAR and ISR, NPR1 has also been suggested tobe involved in regulating cell growth (Vanackeret al., 2001).
Tomato is an economically important cropworldwide, and is susceptible to various fungal,bacterial, viral and nematode diseases, particu-larly in the tropics. Resistance to multiple dis-eases is an important trait when developingtropically adapted varieties of tomato. However,such task requires tremendous resources andefforts when traditional breeding approaches bycrossing are taken. Therefore, this study exploredthe potential of employing SAR-related genes forthe genetic engineering of long lasting andbroad-range disease-resistance in tomato. Here,we report the generation and characterizationof transgenic tomato plants overexpressingArabidopsis NPR1. The transgenic lines generatedfrom this study exhibited significant enhancementof resistance to important bacterial and fungaldiseases, without causing deleterious effects onplant growth and development. Importantly, thespectrum and degree of these transgenic lines aremore significant compared to that of transgenictomatoes reported to date. These transgenic lines
568

could be useful tomato stocks for breeding fur-ther resistance to more diseases, or used asresource for breeding resistance to BW and FWin other tomato varieties.
Materials and methods
Transgene construction and plant transformation
The NPR1 coding sequence was isolated fromArabidopsis thaliana ecotype Col0 by polymerasechain reaction (PCR) using primers designedbased on the published sequence (Cao et al., 1997;Ryals et al., 1997) and cloned under the control ofthe 35S promoter of Cauliflower Mosaic Virus(CaMV 35S promoter). This transgene cassettewas further cloned into the binary Ti-plasmid vec-tor pCAMBIA1301 (Center for the Application ofMolecular Biology to International Agriculture,CAMBIA, Inc., Canberra, ACT, Australia),which carries kanamycin resistance gene and hy-gromycin resistance gene (hygromycin phospho-transferase) as bacterial selection and planttransformation selection genes, respectively. Theconstruct was transformed into the Agrobacteriumtumefaciens strain LBA4404 by cold-shock trans-formation (An, 1987). Agrobacterium-mediatedtransformation was performed on cotyledon ex-plants of 7–8 day old seedlings of Lycopersicon es-culentum according to Fillatti et al. (1987).Hygromycin at 20 mg/l in culture medium wasused to select transformants (defined as R0 lines),followed by staining for a GUS activity. Thetomato line used for the transformation wasCL5915-93D4-1-0-3 (CL5915), a line bred at theAVRDC-The World Vegetable Center, Taiwan.This tomato line is a small-fruit, highly heat-toler-ant line that has been cultivated in several tropicalcountries. It has been used as a good parental linefor generating heat tolerant hybrid cultivar fortropical and subtropical areas. R1 lines were prog-eny lines deriving from the R0 transgenic linesand R2 lines were progeny lines deriving from theR1 transgenic lines.
Molecular characterization of transgenic plants
For genomic Southern blot analysis, genomicDNA was isolated from plant leaves using aCTAB method described by Saghai-Maroof et al.
(1984). Five micrograms of genomic DNAdigested with the appropriate restriction enzymeswas separated in 0.8% agarose gels in 1X TBEbuffer and then transferred to NYTRAN� mem-branes (Schleicher & Schuell BioScience, Keene,NH, USA) following a procedure described bySambrook and Russell (2001). EcoRI and BamHI are restriction enzymes that cut only once inthe NPR1 expression binary vector. For northernblot analysis, total RNA was extracted fromleaves using TRIzol� Reagent (Invitrogen LifeTechnologies, Co., Carlsbad, CA, USA). Fivemicrograms of total RNA was separated in 1.0%agarose gels containing 6.6% formaldehyde andthen transferred to positively charged NylonMembranes (Roche Molecular Biochemicals,Mannheim, Germany). The probes used forSouthern and northern blot analyses were labeledwith digoxigenin (DIG), following manufacture’sinstructions (Roche Molecular Biochemicals). Theblots were probed with the appropriate labeledprobes at 42�C overnight, and then subjected to15 min washes in 2X SSC containing 0.1% SDSat room temperature twice, and in 0.5X SSC con-taining 0.1% SDS at 68�C twice. PR1b1(Y08804), PR1p6 (M69248), acidic b-1,3-glucan-ase (GLUa, M80604), basic b-1,3-glucanase(GLUb, M80608), acidic chitinase 9 (CHI9,Z15140) and basic chitinase 3 (CHI3, Z15141)were used as probes to analyze the expression ofdefense-related genes in the transgenic plants bynorthern blot analysis. In addition, reverse tran-scription PCR (RT-PCR) was performed for thedetection of transgene expression using Ready-To-Go� RT-PCR Beads (Amersham PharmaciaBiotech, Inc., Piscataway). For western blot anal-ysis, total proteins were extracted from plantleaves as described by Jinn et al. (1993), and20 lg of the total proteins was subjected to SDS-PAGE electrophoresis. After transfer to PVDFmembranes (Millipore Co., Billerica, MA, USA),the immunoblots were developed with an antise-rum raised against NPR1 recombinant proteins,following the procedure described in the manual‘‘Immuno-Blot Assay Procedure’’ (Bio-Rad Lab-oratories Inc., Hercules, CA, USA). Since wefailed to express full-length NPR1 protein inE. coli cells, the carboxy-terminal half (310 aminoacids) and the amino-terminal half (300 aminoacids) of NPR1 protein were expressed as recom-binant proteins in E. coli individually by cloning
569

the corresponding sequences into the pET28avector (Novagen, Madison, WI, USA). Thesetwo recombinant proteins were then purifiedfrom IPTG-induced E. coli cells following manu-facture’s instruction, mixed at a ratio of 1:1 andused to immunize rabbits for the production ofan antiserum.
Disease screens of transgenic plants
Several important tropical diseases of tomatowere selected as targets for evaluation of resis-tance of transgenic lines in the disease screens.The fungal diseases selected for the screens wereFusarium wilt (FW, caused by Fusarium oxyspo-rum f. sp. lycopersici), gray leaf spot (GLS, causedby Stemphylium solani), and late blight (LB,caused by Phytophthora infestans). The selectedbacterial diseases were bacterial spot (BS, causedby Xanthomonas campestris pv. vesicatoria) andbacterial wilt (BW, caused by Ralstoniasolanacearum). Viral diseases caused by Cucum-ber Mosaic Virus (CMV), ToMV, and TomatoYellow Leaf Curl Virus (TYLCV) were includedfor the disease screens. The evaluation protocolshave been routinely used at the Asian VegetableResearch and Development Center (AVRDC).Tomato varieties resistant and susceptible to eachdisease were included in each evaluation as con-trols to ensure reliability of the results. Generallyseedlings were raised in 2 inch pots with pasteur-ized potting mixture, except plants to be testedfor FW where seedlings were grown in plastictrays with cell size of 4 cm · 4 cm. Evaluationprotocols are described below. For all the evalua-tions, plants were kept in a greenhouse at a meantemperature of 28�C, unless mentioned otherwise.All evaluations were conducted at the R1 and R2stage. Some of the R2 lines were derived fromindividual plants displaying enhanced resistanceat the R1 stage.
For FW evaluation, Isolate Fol-34ssl (race 2)was used to prepare inoculum. The entire con-tents of 7-day-old potato dextrose agar cultureswere blended with 125 ml of distilled water perplate for 10 s. The slurry contained about107 conidia/ml. Tomato seedlings were uprootedand immersed in the slurry for 5 min, trans-planted to individual cells in plastic trays, andkept in a greenhouse at a mean temperature of25�C. Tomato variety MH-1 and Bonny Best
were used as resistant and susceptible controls,respectively. In total, 18 plants per line weretested in a complete randomized design (CRD).The severity score was rated 21 days post inocu-lation with 0 – no symptoms, 5 – stunted andvascular browning, 7.5 – severely stunted andvascular browning, and 10 – wilted beyondrecovery.
For LB evaluation, sporangium suspension(5 · 104 sporangia/ml) of Isolate Pi39A was pre-pared from 12 to 14 day-old rye A agar culturesand chilled at 12�C for 2–3 h for zoosporerelease. Leaves of 5-week-old tomato seedlingswere sprayed with the chilled suspension untilrun-off. A total of 24 plants per line were testedin a CRD. Tomato varieties L3708 and TS19 wereused as resistant and susceptible controls, respec-tively. Inoculated plants were kept at 20�C andrated 7–10 days after inoculation. The severityscores ranged from 0 to 6, where 0 means nosymptoms and 6 means 91–100% leaf area affectedand/or extensive stem damage or plant dead.
For GLS evaluation, conidia were harvestedfrom 7-day-old V8 cultures and made into suspen-sion (2.5 · 104 conidia/ml). Leaves of 5-week-oldtomato seedlings were sprayed with the conidiumsuspension until run-off. A total of 24 plants perline were tested in a CRD. Tomato varietiesUC82-L and Bonny Best were used as the resistantand susceptible controls, respectively. Inoculatedplants were scored 7 days after inoculation withlevels 0–4, where 0 means no symptoms, 1 meansvery few small lesions, 2 means about 10 lesionsper leaf, 3 means numerous lesions per leaf, and 4means coalescing lesions and leaf collapse.
For BW evaluation, suspensions of StrainPss4 (race 1, biovar 3) were prepared to give anOD600 equal to 0.3 (about 108 cfu/ml) andpoured over the pot soil surface of 3-week-oldseedlings as described previously (Wang et al.,2000). A randomized complete block design(RCBD) was used with three replications and 12plants per replication. Tomato cultivarsHawaii7996 (H7996) and L390 were used as theresistant and susceptible controls, respectively.The response of the inoculated plants was evalu-ated as the percentage of wilted plants 28 daysafter inoculation.
For BS evaluation, suspension of StrainXVP28 (race P1) was prepared from overnight523 cultures to give an OD600 equal to 0.3 (about
570

108 cfu/ml). Prior to inoculation, 0.025% (v/v) ofSilwet L-77 surfactant was added to the bacterialsuspension. The entire upper ground part of3-week-old seedlings were dipped into the suspen-sion for 30 s. Tomato cultivars Hawaii 7998 andBonny Best were used as the resistant and sus-ceptible controls, respectively. A total of 12 plantsper line were tested in a CRD. The response ofthe inoculated plants was evaluated with Barrat-Horsfall scores 10–12 days post inoculation.Score 0 means no symptom development; score 1means less than 3% of leaf area showing symp-toms; score 2 means 3–6% of leaf area showingsymptoms; score 3 means 3–6% of leaf area show-ing symptoms; score 4 means 3–6% of leaf areashowing symptoms.
For ToMV evaluation, the inoculum wasprepared by grinding leaves of ToMV-infectedNicotiana tabacum Samsun in 0.03 M Na-phos-phate buffer (pH 7.0) amended with 0.5% (w/v)Na2SO3, 0.17% (w/v) Na2-DIECA and 0.01%(w/v) charcoal. The leaf to buffer ratio (w/v) was1 g to 4 ml. Tomato seedlings were inoculated atthe 3–4-leaf stage by spreading the inoculum onleaves in the presence of carborundum. ELISAtests were carried out two weeks after inoculationto detect the virus in the inoculated plants.Tomato variety TK70 was used as the susceptiblecontrol. A total of 24 plants per line were testedin a CRD.
For CMV evaluation, the inoculum was pre-pared by grinding leaves of CMV-infectedNicotiana glutinosa in the same buffer and ratioas for ToMV. Seedling inoculation was con-ducted as above, except that a second inoculationwas conducted one week after the first inocula-tion. ELISA tests were carried out two weeksafter the second inoculation to detect the virus inthe inoculated plants. Tomato variety TK70 wasused as the susceptible control. A total of 24plants per line were tested in a CRD.
For TYLCV evaluation, tomato seedlings atthe 1–2-leaf stage were exposed to viruliferouswhiteflies (Bemisia tabaci). Visual symptoms wererecorded once a week until clear symptomsappeared on all plants of the susceptible control(normally 1–1.5 months). Tomato variety TK70was used as the susceptible control. To confirmvisual symptom observations, all plants were sub-jected to nucleic acid hybridizations with a1.4 kb probe of the TYLCV-TW (GenBank No.
U88692) (Shih et al., 1995). A total of 24 plantsper line were tested in a CRD.
To confirm the enhanced resistance to BW,an R2 (26–36, derived from R1 Line 26) and anR3 (9-BW54-1, derived from R2 Line 9-BW54)line were selected for a time-course study. Theinoculation method and experiment design wereconducted as described above, except the percentwilted plants were recorded every 3–7 days until28 days after inoculation. R2 Lines 26–36 andR3 Line 9-BW94-BW8 (derived from R2 Line9-BW54) were used to determine the colonizationdegree by Fusarium oxysorum f.sp. lycopersiciover time. RCBD was used with 40 plants perreplication, and the inoculation was performed asdescribed above. Stem segments (about 0.2 cm inlength) were removed from collar and mid-stem(mid-point of stem) regions at 3, 7, 10, 14, and21 days after inoculation. A total of five plantswere randomly selected from each replication ateach time point. Each segment was split into foursections. Pathogen colonization frequency wasdetermined as the percentage of colonized sec-tions, in which the result of isolating FW patho-gen from the stem was positive.
Two statistical methods were used to checkwhether the transgenic lines performed differentlyfrom the wild-type plants, CL5915, or other con-trol varieties. For BW screening and the FW col-onization experiment, the percentage data wastransformed by arcsin square root for ANOVAand mean comparison by LSD. For the otherdiseases with rating scores, the Mann Whitneynon-parametric U test was conducted to test thenull hypothesis that the ratio of CL5915 (wild-type) plants across the rating scores was equal tothe ratio observed for the transgenic lines.
Results
Generation and identification of stabletomato transgenic lines
To express Arabidopsis NPR1 in tomato, a trans-gene expression plasmid was constructed, in whichthe NPR1 coding region was driven by a CaMV35S promoter and gusA was used as the reportergene. This plasmid was introduced into a tomatoline, namely CL5915-93D4-1-0-3 (CL5915), via anAgrobacterium-mediated transformation method.
571
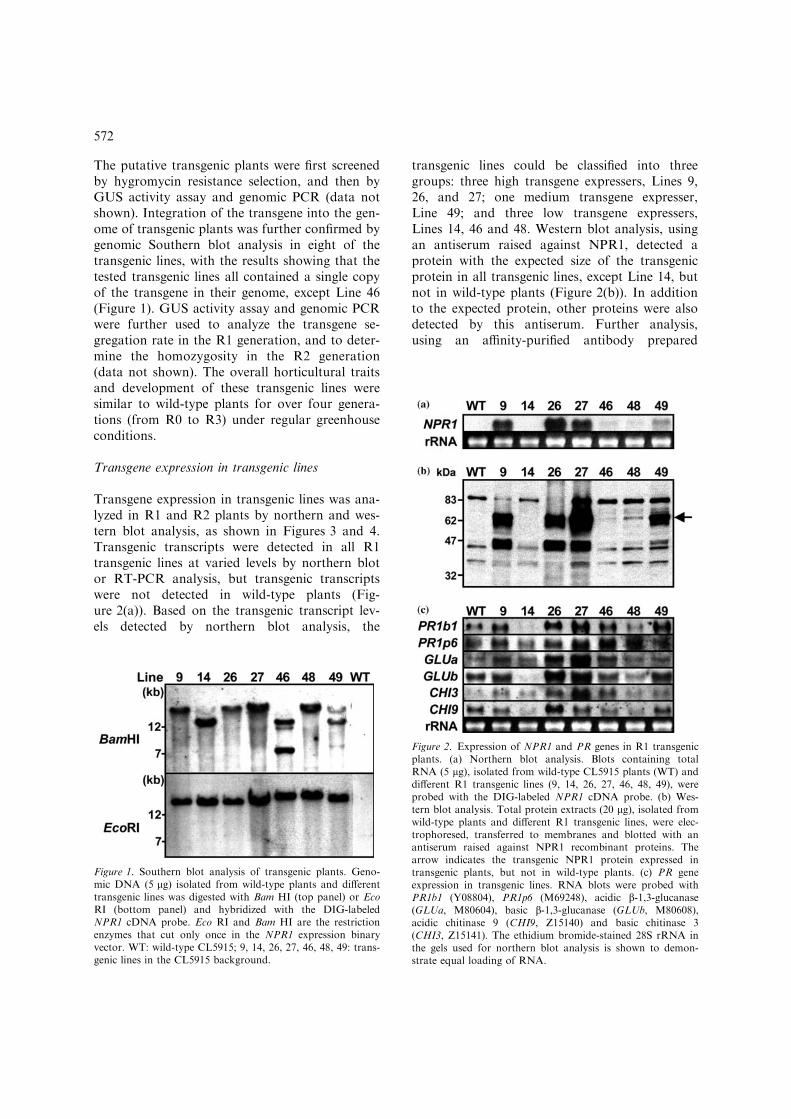
The putative transgenic plants were first screenedby hygromycin resistance selection, and then byGUS activity assay and genomic PCR (data notshown). Integration of the transgene into the gen-ome of transgenic plants was further confirmed bygenomic Southern blot analysis in eight of thetransgenic lines, with the results showing that thetested transgenic lines all contained a single copyof the transgene in their genome, except Line 46(Figure 1). GUS activity assay and genomic PCRwere further used to analyze the transgene se-gregation rate in the R1 generation, and to deter-mine the homozygosity in the R2 generation(data not shown). The overall horticultural traitsand development of these transgenic lines weresimilar to wild-type plants for over four genera-tions (from R0 to R3) under regular greenhouseconditions.
Transgene expression in transgenic lines
Transgene expression in transgenic lines was ana-lyzed in R1 and R2 plants by northern and wes-tern blot analysis, as shown in Figures 3 and 4.Transgenic transcripts were detected in all R1transgenic lines at varied levels by northern blotor RT-PCR analysis, but transgenic transcriptswere not detected in wild-type plants (Fig-ure 2(a)). Based on the transgenic transcript lev-els detected by northern blot analysis, the
transgenic lines could be classified into threegroups: three high transgene expressers, Lines 9,26, and 27; one medium transgene expresser,Line 49; and three low transgene expressers,Lines 14, 46 and 48. Western blot analysis, usingan antiserum raised against NPR1, detected aprotein with the expected size of the transgenicprotein in all transgenic lines, except Line 14, butnot in wild-type plants (Figure 2(b)). In additionto the expected protein, other proteins were alsodetected by this antiserum. Further analysis,using an affinity-purified antibody prepared
Figure 2. Expression of NPR1 and PR genes in R1 transgenicplants. (a) Northern blot analysis. Blots containing totalRNA (5 lg), isolated from wild-type CL5915 plants (WT) anddifferent R1 transgenic lines (9, 14, 26, 27, 46, 48, 49), wereprobed with the DIG-labeled NPR1 cDNA probe. (b) Wes-tern blot analysis. Total protein extracts (20 lg), isolated fromwild-type plants and different R1 transgenic lines, were elec-trophoresed, transferred to membranes and blotted with anantiserum raised against NPR1 recombinant proteins. Thearrow indicates the transgenic NPR1 protein expressed intransgenic plants, but not in wild-type plants. (c) PR geneexpression in transgenic lines. RNA blots were probed withPR1b1 (Y08804), PR1p6 (M69248), acidic b-1,3-glucanase(GLUa, M80604), basic b-1,3-glucanase (GLUb, M80608),acidic chitinase 9 (CHI9, Z15140) and basic chitinase 3(CHI3, Z15141). The ethidium bromide-stained 28S rRNA inthe gels used for northern blot analysis is shown to demon-strate equal loading of RNA.
Figure 1. Southern blot analysis of transgenic plants. Geno-mic DNA (5 lg) isolated from wild-type plants and differenttransgenic lines was digested with Bam HI (top panel) or EcoRI (bottom panel) and hybridized with the DIG-labeledNPR1 cDNA probe. Eco RI and Bam HI are the restrictionenzymes that cut only once in the NPR1 expression binaryvector. WT: wild-type CL5915; 9, 14, 26, 27, 46, 48, 49: trans-genic lines in the CL5915 background.
572
against the NPR1 recombinant proteins, indi-cated that some of the detected proteins may beproteins derived from the transgenic proteins,while some may be endogenous gene products ofNPR1-related homologue(s) (data not shown). Ingeneral, the amounts of the transgenic proteinsaccumulated in individual R1 lines were consis-tent with the levels of transgenic transcripts (Fig-ure 2(b)). The patterns of transgene expression inR2 homozygous plants deriving from three R1lines were also consistent with that in the parentR1 lines at the transcriptional level (Figure 3(a)).R2 lines deriving from high transgene expressers(Lines 9 and 26) accumulated high levels of tran-scripts, whereas the transcript levels in R2 linesderiving from the low transgene expresser (Line14) were too low to be detected by northern blotanalysis. However, despite the levels of transgenictranscripts being similar among cognate R2 lines,the amounts of transgenic proteins varied(Figure 3(b)). These results indicated that trans-
gene expression was heritable at transcriptionallevel, but that transgene silencing may haveoccured in some individuals.
Defense gene expression in transgenic lines
As induction of NPR1 expression leads to theactivation of a group of PR genes (Cao et al.,1998), we further assessed expression of severaltomato PR genes in R1 and R2 transgenic plantsby northern blot analysis (Figures 2(c) and 3(c)).Results showed that these PR genes were consti-tutively expressed at different levels in wild-typeplants. In R1 transgenic lines, although theexpression patterns of these test PR genes didnot apparently correlate with the level oftransgene expression, higher levels of transgeneexpression in some of the high transgene express-ers (i.e. lines 26 and 27) led to more significantdegree of enhanced expression of a few PR genes(e.g., GLUa, GLUb and CHI3) (Figure 2(c)).However, no clear correlation between PR geneexpression and transgene expression wasobserved in R2 transgenic lines (Figure 3(c)).
Disease screens of transgenic lines
For future cultivation of the promising trans-genic tomato plants in the tropics, we evaluated
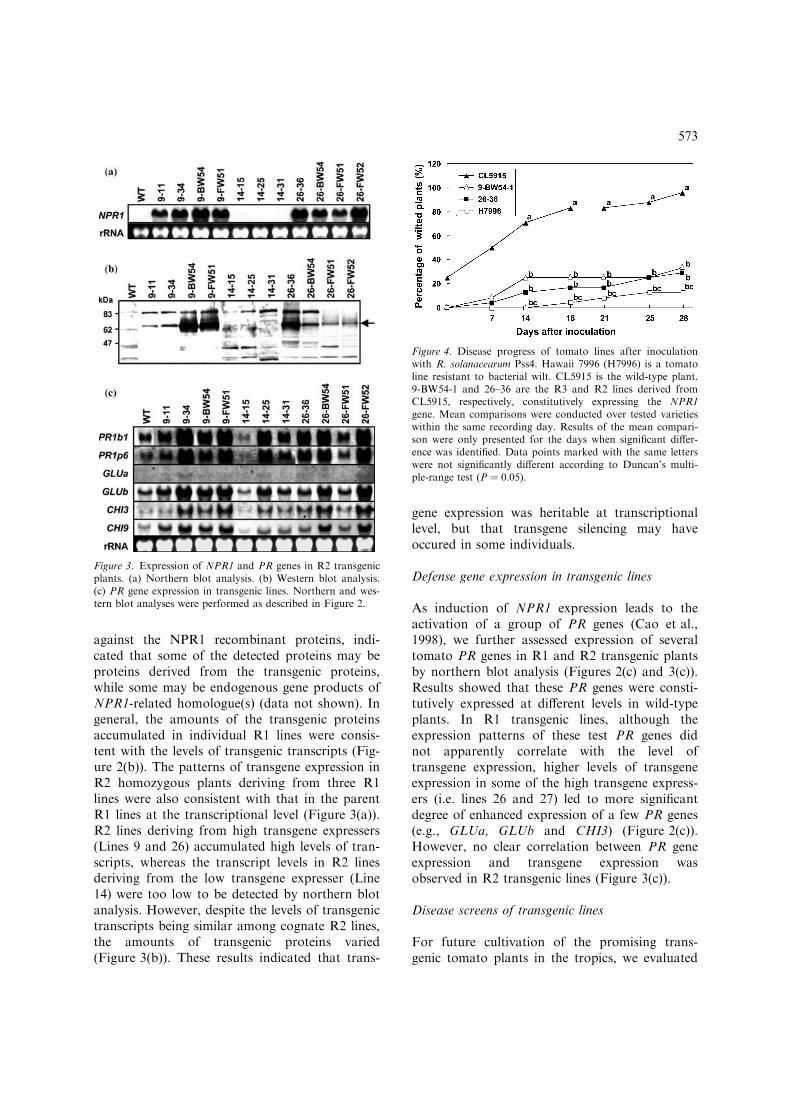
Figure 3. Expression of NPR1 and PR genes in R2 transgenicplants. (a) Northern blot analysis. (b) Western blot analysis.(c) PR gene expression in transgenic lines. Northern and wes-tern blot analyses were performed as described in Figure 2.
Figure 4. Disease progress of tomato lines after inoculationwith R. solanacearum Pss4. Hawaii 7996 (H7996) is a tomatoline resistant to bacterial wilt. CL5915 is the wild-type plant.9-BW54-1 and 26–36 are the R3 and R2 lines derived fromCL5915, respectively, constitutively expressing the NPR1gene. Mean comparisons were conducted over tested varietieswithin the same recording day. Results of the mean compari-son were only presented for the days when significant differ-ence was identified. Data points marked with the same letterswere not significantly different according to Duncan’s multi-ple-range test (P ¼ 0.05).
573

resistance of the transgenic lines to eight impor-tant tropical diseases. The diseases selected forthis evaluation included leaf-localized, systemicand vascular diseases caused by fungal, bacterialand viral pathogens. Seven transgenic lines at R1stage and three transgenic lines (derived fromlines 9, 14 and 26) at R2 stage were subjected todisease screens to determine whether the trans-genic lines displayed increased levels of resistanceto the chosen diseases compared to wild-typeCL5915 plants.
Tomato CL5915 innately processes resistanceto ToMV, and all of the tested R1 and R2 trans-genic lines remained resistant to ToMV (data notshown). When challenged with TYLCV, CMVand the pathogens of LB, all R1 lines exhibitedsusceptibility similar to wild-type plants (data notshown). However, R1 transgenic lines exhibitedvarious levels of enhanced resistance to FW, BW,BS and GLS. Particularly, significant NPR1-induced enhancement of resistance to FW andBW was observed (Table 1). For FW, the three
high NPR1-expressers (lines 9, 26, and 27) showedlower disease severity than wild-type plants. Par-ticularly, lines 26 and 27 displayed low diseaseseverity symptoms similar to the FW-resistantcontrol MH-1, which processes a single dominantresistance gene, I2 (Crill et al., 1971). Transgeniclines with medium or low levels of NPR1 expres-sion, however, showed similar or slightly higherseverity than wild-type plants. These data suggestthat enhanced FW-resistance may require athreshold level of NPR1 expression in transgenictomato plants. For BW, R1 lines 9, 14, 26 and 49showed significantly lower disease severity symp-toms than wild-type plants (Table 1). However,the final disease severity symptoms of these trans-genic lines was still higher than that of H7996, atomato cultivar resistant to BW (Wang et al.,1998). In addition, enhanced resistance to BWobserved in R1 transgenic lines did not correlatewith the level of transgene expression. Forinstance, among the high NPR1-expressers, lines9 and 26 displayed enhanced resistance to BW,
Table 1. Evaluation of transgenic tomato plants expressing NPR1 on their responses to FW, GLS, BW and BS pathogens. Datapresented are maximum severity symptoms of each disease
R1 lines FWb R1/CL5915 BWc R1/CL5915 R2 lines FWb GLSb BWc BW2c BSb
9 2.2/7.6** 50.0/91.7* 9–11 3.2** 3.0** 25.0** 33.3** 3.2ns
14 7.1/7.6ns 16.7/91.7* 9–34 2.1** 2.8** 58.3ns n.t.d 3.5ns
26 0.6/7.2** 33.3/100.0* 9-FW51 3.0** 2.6** 33.3* 25.0** 2.3**
27 0.3/7.2** 75.0/100.0ns 9-BW54 1.0** 2.4** 25.0** 33.3**. 2.7ns
46 8.3/6.5** 66.7/91.7ns 14–15 3.9* 3.4** 41.7ns n.t. 3.2ns
48 6.4/6.5ns 100.0/91.7ns 14–25 2.8** 3.8ns 41.7ns n.t. 3.2ns
49 6.5/6.5ns 33.3/91.7* 14–31 5.2ns 3.7ns 50.0ns n.t. 3.3ns
Ra 0.0 11.3 14–32 2.6** 2.9** 41.7ns n.t. 2.7ns
Sa 9.4 100.0 14-BW54 3.2** 3.2** 25.0** 29.2** 3.2ns
26–36 0.6** 3.2** 25.0** 29.2** 3.2ns
26-FW51 2.5** 3.4** 25.0** 58.3** 3.3ns
26-FW52 2.6** 3.3** 33.3* 33.3** 2.8ns
26-BW54 3.8* 3.5** 58.4ns 45.8** 3.0ns
CL5915a 5.7 3.9 58.3 95.8 3.3
Ra 0.2 0.4 8.3 12.5 1.0
Sa 8.5 4.0 100.0 100.0 3.3
a Varieties resistant (R) and susceptible (S) to each disease were used as controls. They were MH1 (R) and Bonny Best (S) for FW,UC82-L (R) and Bonny Best (S) for GLS, Hawaii 7996 (R) and L390 (S) for BW, and Hawaii 7998 (R) and Bonny Best (S) for BS.Data of the control varieties in the R1 evaluations were mean over two or three trials.b Data presented were mean severity scores. Results of the MannWhitney U non-parametric test for testing the null hypothesis that theratio of plants of CL5915 across the rating scores is equal to the ratio observed for the transgenic lines were marked as non-significant(ns) and significantly different at 0.05 (*) and 0.01 level (**). Comparison of each R1 line was made with CL5915 in the specific trial.c Data presented were mean percent wilted plant. The percentage data was transformed with arcsin square root for ANOVA and theresults of mean comparison by LSD were marked as non-significant (ns) and significantly different at 0.05 (*) or 0.01 (**) level. For theBW2 trial, 9-BW54-1 (R3 line) was tested instead of it R2 line due to seed shortage.d ‘‘n.t.’’ means ‘‘not tested’’.
574

while line 27 did not. Furthermore, R1 plants ofline 49 (a medium NPR1-expresser) and line 14 (alow NPR1-expresser) also exhibited enhancedBW-resistance. For GLS, only line 49 showedenhanced resistance with a mean severity symp-toms score of 1.3 vs. that of 3.9 for wild-typeplants. Minor levels of increased resistance to BSwere observed in three R1 lines. Lines 9 and 14had a mean severity symptoms score of 2.8 and2.3, respectively, vs. that of 3.3 for wild-typeplants, and line 49 had a mean severity symptomsscore of 1.4 vs. that of 2.0 for wild-type plants.
Selected R2 lines, derived from the R1 linesexhibiting enhanced resistance to BW and FW,were evaluated for their response to the sameeight diseases. The results were very similar tothat observed in the parent R1 lines (Table 1),where significant enhancement of resistance toFW and BW, moderate enhancement of the resis-tance to GLS and BS, and no enhanced resis-tance to LB, TYLCV and CMV were observed(data not shown). Since the disease severity ofwild-type CL5915 plants in the first BW evalua-tion was lower than the usual level (trial BW ofR2 lines shown in Table 1), selected lines exhibit-ing enhanced BW-resistance were subjected tothe evaluation again, and the results were similarto that observed in the first screening (trial BW2for R2 lines shown in Table 1). Some of the R2lines were derived from individual plants thatexhibited resistance to specific diseases tested inthe R1 screening, and thus with the respectivedisease abbreviations in their line codes. R2 lineswere also collected from individual R1 plantswithout prior selection for enhanced resistance tothe diseases. No significant difference among R2lines collected by these two approaches wasobserved. However, not all the R2 lines derivedfrom the same R1 line exhibited similar patternof enhanced resistance. For example, only one ofthe 14-derived R2 lines showed significantenhanced resistance to BW; while, surprisingly,most of the 14-derived R2 lines showed signifi-cant enhancement of FW-resistance, which wasnot observed in the parent R1 line. In addition,R2 transgenic lines that accumulated higherlevels of NPR1 proteins apparently may exhibitbetter performance in terms of the degree and/orspectrum of enhanced resistance to the diseasestested. For instance, lines 9-BW54 and 26–36,which expressed high levels of NPR1 proteins,
exhibited FW-resistance at a level similar to theFW-resistant control MH-1, in addition toenhanced resistance against BW and GLS. Line9-BW51, which also expressed NPR1 proteins ata high level, displayed not only enhanced broad-range resistance to FW, GLS and BW, but alsoincreased resistance to BS.
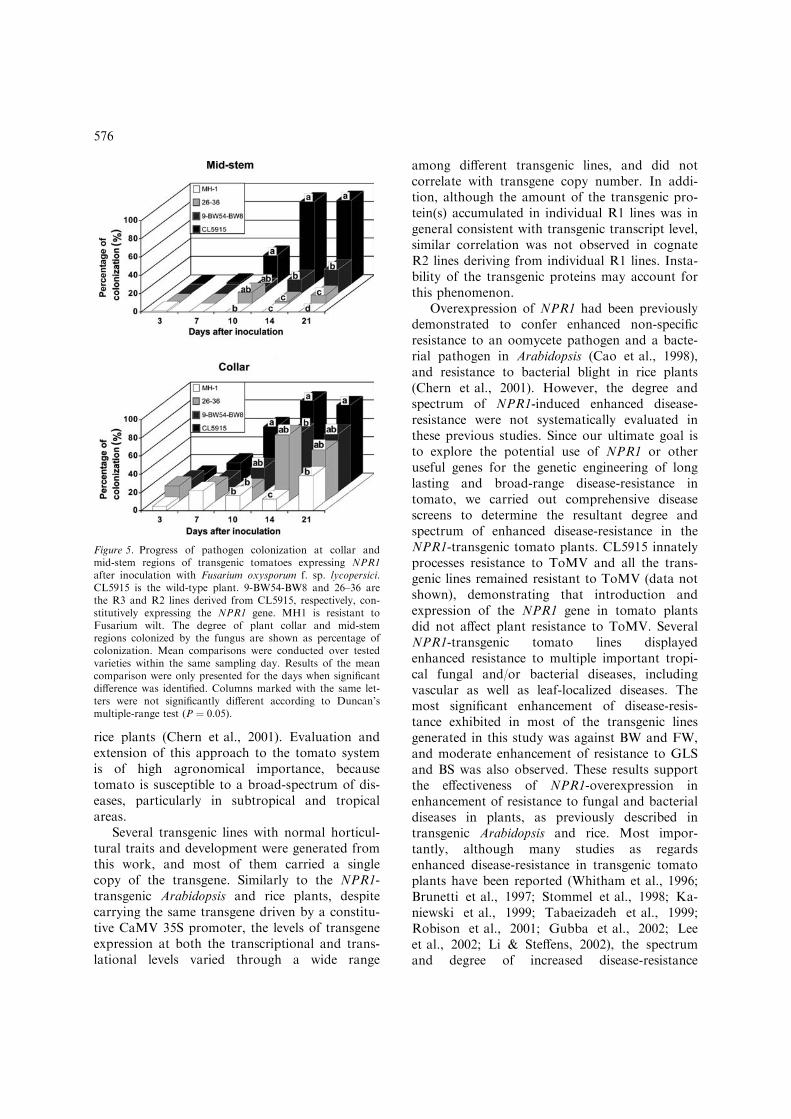
To elucidate a defense mechanism possiblyresponsible for enhanced resistance to BW andFW observed in NPR1-transgenic tomato plants,transgenic lines exhibiting high levels of enhancedresistance to BW and FW were selected for fur-ther analysis. Disease progress was monitored inselected R2 or R3 transgenic lines, in comparisonwith wild-type plants and resistant control vari-eties. For BW, R2 lines 26–36 and R3 line9-BW54-1 exhibited higher disease incidence overtime than the resistant control H7996 did, but thedifference was not statistically significant(Figure 4). In addition, H7996 had longer latentperiod of symptom appearance than the twotransgenic lines (Figure 4). Furthermore, the colo-nization progress of the pathogen of FWwas monitored. R2 lines 26–36 and R3 line9-BW54-BW8 showed similar levels of coloniza-tion with the resistant control MH1 up to 10 daysafter inoculation (Figure 5). Although FW patho-gen colonization at the collar region of transgenicplants then reached a similar level to that foundin wild-type plants, pathogen colonization at themid-stem region of transgenic plants was still sig-nificantly lower than that in wild-type plants.Therefore, the enhanced FW resistance in trans-genic lines was mostly due to the suppression ofthe upward movement of the pathogen. While inMH1, the resistance was associated with the sup-pression of both pathogen colonization andmovement.
Discussion
In this study, we explored the potential use ofSAR-related genes for the genetic engineeringof enhanced resistance/tolerance to a spectrumof diseases in tomato. Arabidopsis NPR1 waschosen as the target transgene for generationof transgenic tomato plants, as its effectiveness inconferring an enhanced degree of non-specific dis-ease-resistance has been previously demonstratedin transgenic Arabidopsis (Cao et al., 1998) and
575
rice plants (Chern et al., 2001). Evaluation andextension of this approach to the tomato systemis of high agronomical importance, becausetomato is susceptible to a broad-spectrum of dis-eases, particularly in subtropical and tropicalareas.
Several transgenic lines with normal horticul-tural traits and development were generated fromthis work, and most of them carried a singlecopy of the transgene. Similarly to the NPR1-transgenic Arabidopsis and rice plants, despitecarrying the same transgene driven by a constitu-tive CaMV 35S promoter, the levels of transgeneexpression at both the transcriptional and trans-lational levels varied through a wide range
among different transgenic lines, and did notcorrelate with transgene copy number. In addi-tion, although the amount of the transgenic pro-tein(s) accumulated in individual R1 lines was ingeneral consistent with transgenic transcript level,similar correlation was not observed in cognateR2 lines deriving from individual R1 lines. Insta-bility of the transgenic proteins may account forthis phenomenon.
Overexpression of NPR1 had been previouslydemonstrated to confer enhanced non-specificresistance to an oomycete pathogen and a bacte-rial pathogen in Arabidopsis (Cao et al., 1998),and resistance to bacterial blight in rice plants(Chern et al., 2001). However, the degree andspectrum of NPR1-induced enhanced disease-resistance were not systematically evaluated inthese previous studies. Since our ultimate goal isto explore the potential use of NPR1 or otheruseful genes for the genetic engineering of longlasting and broad-range disease-resistance intomato, we carried out comprehensive diseasescreens to determine the resultant degree andspectrum of enhanced disease-resistance in theNPR1-transgenic tomato plants. CL5915 innatelyprocesses resistance to ToMV and all the trans-genic lines remained resistant to ToMV (data notshown), demonstrating that introduction andexpression of the NPR1 gene in tomato plantsdid not affect plant resistance to ToMV. SeveralNPR1-transgenic tomato lines displayedenhanced resistance to multiple important tropi-cal fungal and/or bacterial diseases, includingvascular as well as leaf-localized diseases. Themost significant enhancement of disease-resis-tance exhibited in most of the transgenic linesgenerated in this study was against BW and FW,and moderate enhancement of resistance to GLSand BS was also observed. These results supportthe effectiveness of NPR1-overexpression inenhancement of resistance to fungal and bacterialdiseases in plants, as previously described intransgenic Arabidopsis and rice. Most impor-tantly, although many studies as regardsenhanced disease-resistance in transgenic tomatoplants have been reported (Whitham et al., 1996;Brunetti et al., 1997; Stommel et al., 1998; Ka-niewski et al., 1999; Tabaeizadeh et al., 1999;Robison et al., 2001; Gubba et al., 2002; Leeet al., 2002; Li & Steffens, 2002), the spectrumand degree of increased disease-resistance
Figure 5. Progress of pathogen colonization at collar andmid-stem regions of transgenic tomatoes expressing NPR1after inoculation with Fusarium oxysporum f. sp. lycopersici.CL5915 is the wild-type plant. 9-BW54-BW8 and 26–36 arethe R3 and R2 lines derived from CL5915, respectively, con-stitutively expressing the NPR1 gene. MH1 is resistant toFusarium wilt. The degree of plant collar and mid-stemregions colonized by the fungus are shown as percentage ofcolonization. Mean comparisons were conducted over testedvarieties within the same sampling day. Results of the meancomparison were only presented for the days when significantdifference was identified. Columns marked with the same let-ters were not significantly different according to Duncan’smultiple-range test (P ¼ 0.05).
576

observed in the transgenic tomato plantsreported in these previous studies were not as sig-nificant as that of our NPR1-transgenic lines.The only exception may be the transgenic tomatoplants overexpressing a tomato single dominantdisease-resistance gene, Pto, as reported by Tanget al. (1999). However, even in this study, onlyenhanced level of resistance to three leaf-localizeddiseases was demonstrated in the Pto-transgenictomato plants, while our NPR1-transgenic linesexhibited enhanced resistance to leaf-localized aswell as vascular bacterial and fungal diseases.Results from our study demonstrate the effective-ness of SAR-based approaches, particularly theoverexpression of NPR1 in plants, for the geneticengineering of disease-resistance, as regards thespectrum and degree of enhanced disease-resis-tance.
Overexpression of NPR1 in Arabidopsis andrice leads to enhanced non-specific disease-resis-tance in a dosage-dependent fashion (Cao et al.,1998; Chern et al., 2001). Consistent with thesereports, in this study, we showed that transgenictomato lines that accumulated higher levels ofNPR1 proteins apparently exhibited higher levelsand a broader spectrum of enhanced resistance tothe test diseases, as demonstrated in R2 lines 9-FW51, 9-BW54 and 26–36. Additionally, ourresults indicate that stable inheritance ofenhanced disease-resistance to BW and FW mayrequire a minimal or threshold level of NPR1expression in transgenic tomato plants, as no con-sistent enhancement of resistance to BW and FWwas observed in line 14, a low NPR1-expresserline, at R1 generation and R2 generation. How-ever, the expression pattern of PR genes in ourtransgenic tomato plants was not consistent withthat previously reported. In transgenic Arabidop-sis plants overexpressing NPR1, INA-induced PRgene expression was induced in an NPR1 dosage-dependent manner and correlated with the resul-tant enhanced resistance to bacterial and fungalpathogens (Cao et al., 1998). In contrast,although expression of a few PR genes wasenhanced in some of our transgenic tomato lines,the degree of induced PR gene expression did notclearly correlate with the levels of NPR1 trans-gene expression, nor with the spectrum anddegree of resultant enhanced resistance to the testdiseases we tested. Previously, it had been shownthat PR gene expression could be either NPR1-
dependent or -independent (Zhang & Shapiro,2002), and be uncoupled from bacterial and viralresistance in Arabidopsis (Clarke et al., 1998;Wong et al., 2002). Therefore, the enhancedbroad-range disease-resistance observed in NPR1-trangenic tomato plants may involve the activa-tion of other defense-related genes that were notanalyzed in this study. Alternatively, some of thePR genes we tested may be involved, but differen-tially expressed in different tissues. Note that thetransgenic tomato lines exhibited significantenhanced resistance to BW and FW (two soil-borne vascular diseases), while displayed only lim-ited enhanced resistance to GLS and BS (twoleaf-localized diseases). However, analysis ofNPR1 and PR gene expression was performedonly on leaves of transgenic lines. Therefore, ifthe latter argument sustains, the expression pat-tern of the involved PR genes may correlate withthat of NPR1 transgene and the resultant spec-trum and degree of enhanced disease-resistance invascular tissues, particularly in roots and collars.Further analysis of spatial expression patterns ofthese genes and other defense-related genes wouldhelp elucidate the relationship between NPR1-enhanced resistance to the diseases we tested anddefense gene expression.
The enhanced resistance to BW observed in R2line 26–36 and R3 line 9-BW54-BW8 was similarto that in H7996. Previously, it has been demon-strated that resistance to BW in H7996 is relatedto the internal pathogen multiplication, ratherthan root invasion or upward movement (Wang &Lin, 2003). Our preliminary data also indicated asimilar suppression of bacterial growth in R2 line26–36 and R3 line 9-BW54-BW8 (data notshown). However, the incidence of BW in ourtransgenic lines was still higher than that inH7996. These results suggest that the enhancedBW resistance induced by NPR1 overexpression intransgenic tomato may have resulted from stimu-lation of either a portion of the defense system(s)responsible for the resistance conferred in H7996,or via activation of a defense pathway differentfrom that employed by H7996. However, since themechanism of BW-resistance conferred in H7996and the involved signal transduction pathway(s)have not been elucidated at a molecular level thisfar, further studies are necessary to elucidate andcompare the nature of BW-resistance in H7996and that in NPR1-transgenic tomato plants.
577

In a few NPR1-transgenic lines, the degree ofenhanced resistance to FW was similar to thatconferred by the single dominant gene I2 in MH1.However, detailed analysis of pathogen coloniza-tion over time revealed that the enhanced FW-resistance in the transgenic lines was mostly dueto the suppression of the upward movement ofthe pathogen, whereas the resistance in MH1 wasassociated with suppression of both pathogen col-onization and movement. These results indicatethat the enhanced FW-resistance seen in theNPR1-transgenic plants may have resulted from apartial activation of the defense system(s) medi-ated by I2. Alternatively, overexpression of NPR1may activate an I2-independent defense mecha-nism(s) and lead to the enhanced resistance intransgenic tomato lines by suppressing theupward movement of the pathogen. Because themechanism of I2-mediated resistance to FW andthe related signal transduction pathway(s) havenot been elucidated at a molecular level, we cur-rently do not have any evidence for or againsteither of these hypotheses. Previously, NPR1 wasshown to be involved in resistance of Arabidopsisto some diseases mediated by single dominant di-sease-resistance genes (Cao et al., 1994; Delaneyet al., 1994, 1995; McDowell et al., 2000; Liuet al., 2002), but did not play a role in disease-resistance mediated by other single dominant dis-ease-resistance genes (Bittner-Eddy & Beynon,2001; Li et al., 2001; Rairdan & Delaney, 2002;Shirano et al., 2002). However, the relationshipbetween I2 and NPR1 has not been studied intomato this far. Further studies on this aspect areexpected to expand our understanding on FW-resistance in tomato.
In addition to significant enhancement ofresistance to BW and FW, overexpression ofNPR1 was also effective on enhancement of mod-erate resistance to GLS and BS in high NPR1-expressing lines. No information concerningresistance to GLS at molecular level in tomato isavailable so far and genetically engineered resis-tance to GLS had not been reported before. Inaddition, the only reported cases of enhancedresistance to BS in tomato were achieved byoverexpression of the pepper disease-resistancegene Bs2 or Pto (Tai et al., 1999; Tang et al.,1999). Therefore, our transgenic lines are of highagronomical value for their disease-resistance toa broad-spectrum of important fungal and
bacterial diseases. Our present study hence opensup new possibilities for future investigations oncross-talk mechanisms involved in tomato resis-tance to a broad-spectrum of pathogens.
Except for the original ToMV-resistance car-ried in the CL5915 genome, no enhancement ofresistance to other tested tomato viral pathogens,such as CMV and TYLCV, was observed in theNPR1-transgenic tomato lines (data not shown).Resistance to viral pathogens was not reportedor tested in previous studies on transgenic Ara-bidopsis and rice plants overexpressing NPR1(Cao et al., 1998; Chern et al., 2001). However,studies have shown that, unlike resistance to fun-gal and bacterial pathogens, SA-induced antiviralresistance is independent of NPR1 and PR geneexpression (Chivasa & Carr, 1998; Wong et al.,2002). Accumulating evidence demonstrated intobacco and Arabidopsis further suggested thatSA-induced antiviral defense pathways could bedistinct from the defense pathways leading toresistance to bacterial and fungal pathogens(Murphy et al., 1999, 2001; Kachroo et al.,2000). Therefore, overexpression of NPR1 inplants may not be effective in enhancing resis-tance to viral pathogens, as we demonstrated inthis study.
In conclusion, we have demonstrated thatexpression of a heterologous NPR1 in tomatowas effective in activating certain defense signaltransduction pathways, leading to enhanced resis-tance to a broad-range of important fungal andbacterial pathogens. Furthermore, despite exhib-iting enhanced resistance to multiple diseases, noobvious deleterious effects on growth and devel-opment were observed in transgenic tomatoplants for over four generations (R0–R3). Partic-ularly, enhanced resistance to FW and BW in afew NPR1-transgenic tomato plants was verysimilar to, but not as complete as, the resistanceconferred by single dominant disease-resistancegenes. This important feature could potentiallyreduce the selection pressure for more virulentpathogens, which may overcome the resistanceconferred in the transgenic lines, and thus wouldbe particularly desirable for the genetic engineer-ing of enhanced resistance. In addition to thesignificantly enhanced resistance to FW and BW,the moderately enhanced resistance to GLS andBS in a few transgenic lines is expected to benefittomato crop production. Moreover, possible
578

further enhancement of resistance to these as wellas to other important diseases may also beachieved by appropriate application of chemicalinducers or fungicides (Friedrich et al., 2001).The transgenic tomato lines we have generatedcould thus become an important resistanceresource for breeding resistance to BW and FW,as well as be utilized as a genetic background infurther breeding for resistance to a broader spec-trum of diseases. Similar SAR-based strategiesmay also aid other important crop breedingprograms for broad-spectrum disease-resistance,particularly in combination with a genetic back-ground conferring natural disease-resistance.
Acknowledgements
We thank Dr. Kenrick Deen for his critical reviewof this manuscript and Drs. Pierre de Wit and Janvan Kan for providing the tomato PR geneclones. We also thank Hsien-Ling Liu for the gen-eration of transgenic lines and the technical assis-tance of Chiou-Fen Hsu and Hsueh-Hsiang Liaufor propagating successive generations of thetransgenic tomato. This work was supported bythe National Science and Technology Programfor Agricultural Biotechnology, Taiwan, Republicof China (Grant #: 90S201912S to C.-P. C.)
References
Alexander D, Goodman RM, Gut-Rella M, Glascock C,Weymann K, Friedrich L, Maddox D, Ahl-Goy P, LuntzT, Ward E and Ryals J (1993) Increased tolerance to twooomycete pathogens in transgenic tobacco expressing path-ogenesis-related protein 1a. Proc Natl Acad Sci USA 90:7327–7331.
An G (1987) Binary Ti vectors for plant transformation andpromoter analysis. Methods Enzymol 153: 292–305.
Bittner-Eddy PD and Beynon JL (2001) The Arabidopsisdowny mildew resistance gene, RPP13-Nd, functions inde-pendently of NDR1 and EDS1 and does not require theaccumulation of salicylic acid. Mol Plant Microbe Interact14: 416–421.
Broglie K, Chet I, Holliday M, Cressman R, Biddle P,Knowlton S, Mauvais CJ and Broglie R (1991) Transgenicplants with enhanced resistance to the fungal pathogen.Rhizoctonia solani. Science 254: 1194–1197.
Brunetti A, Tavazza M, Noris E, Tavazza R, Caciagli P,Ancora G, Crespi S and Accotto GP (1997) High expres-sion of truncated viral rep protein confers resistance totomato yellow leaf curl virus in transgenic tomato plants.Mol Plant Microbe Interact 10: 571–579.
Cao H, Bowling SA, Gordon AS and Dong X (1994) Charac-terization of an Arabidopsis mutant that is nonresponsive toinducers of systemic acquired resistance. Plant Cell 6: 1583–1592.
Cao H, Glazebrook J, Clarke JD, Volko S and Dong X(1997) The Arabidopsis NPR1 gene that controls systemicacquired resistance encodes a novel protein containingankyrin repeats. Cell 88: 57–63.
Cao H, Li X and Dong X (1998) Generation of broad-spec-trum disease resistance by overexpression of an essentialregulatory gene in systemic acquired resistance. Proc NatlAcad Sci USA 95: 6531–6536.
Chern MS, Fitzgerald HA, Yadav RC, Canlas PE, Dong Xand Ronald PC (2001) Evidence for a disease-resistancepathway in rice similar to the NPR1-mediated signalingpathway in Arabidopsis. Plant J 27: 101–113.
Chivasa S and Carr JP (1998) Cyanide restores N gene-medi-ated resisance to tobacco mosaic virus in transgenic tobaccoexpressing salicylic acid hydroxylase. Plant Cell 10: 1489–1498.
Clarke JD, Liu Y, Klessig DF and Dong X (1998) Uncou-pling PR gene expression from NPR1 and bacterial resis-tance: characterization of the dominant Arabidopsis cpr6-1mutant. Plant Cell 10: 557–569.
Conrath U, Pieterse CM and Mauch-Mani B (2002) Primingin plant-pathogen interactions. Trends Plant Sci 7: 210–216.
Crill P, Strobel JW, Burgis DS, Bryan HH, John CA, EverettPH, Bartz JA, Hayslip NC and Deen WW (1971) FloridaMH-1, Florida’s First Machine Harvest Fresh MarketTomato (p. 12), Circular S-212. Agricultural ExperimentStation. University of Florida.
Delaney TP, Friedrich L, Kessmann H, Uknes S, Vernooij B,Ward E, Weymann K and Ryals J (1994) The molecularbiology of systemic acquired resistance. Curr Plant Sci Bio-technol Agric 21: 339–347.
Delaney TP, Friedrich L and Ryals JA (1995) Arabidopsis sig-nal transduction mutant defective in chemically and biologi-cally induced disease resistance. Proc Natl Acad Sci USA92: 6602–6606.
Despres C, DeLong C, Glaze S, Liu E and Fobert PR (2000)The Arabidopsis NPR1/NIM1 protein enhances the DNAbinding activity of a subgroup of the TGA family of bZIPtranscription factors. Plant Cell 12: 279–290.
Fan W and Dong X (2002) In vivo interaction between NPR1and transcription factor TGA2 leads to salicylic acid-medi-ated gene activation in Arabidopsis. Plant Cell 14: 1377–1389.
Fillatti JJ, Kiser J, Rose R and Comai L (1987) Efficienttransfer of a glyphosate tolerance gene into tomato using abinary Agrobacterium tumefaciens vector. Bio/Technology 5:726–730.
Friedrich L, Lawton K, Dietrich R, Willits M, Cade R and Ry-als J (2001) NIM1 overexpression in Arabidopsis potentiatesplant disease resistance and results in enhanced effectivenessof fungicides. Mol Plant Microbe Interact 14: 1114–1124.
Glazebrook J, Rogers EE and Ausubel FM (1996) Isolationof Arabidopsis mutants with enhanced disease susceptibilityby direct screening. Genetics 143: 973–982.
Gubba A, Gonsalves C, Stevens MR, Tricoli DM and Gonsal-ves D (2002) Combining transgenic and natural resistance toobtain broad resistance to tospovirus infection in tomato(Lycopersicon esculentum mill). Mol Breeding 9: 13–23.
Jach G, Gornhardt B, Mundy J, Logemann J, Pinsdorf E,Leah R, Schell J and Maas C (1995) Enhanced quantitative
579

resistance against fungal disease by combinatorial expres-sion of different barley antifungal proteins in transgenictobacco. Plant J 8: 97–109.
Jinn TL, Wu SH, Yeh CH, Hsieh MH, Yeh YC, Chen YMand Lin CY (1993) Immunological kinship of class I lowmolecular weight heat shock proteins and thermostabiliza-tion of soluble proteins in vitro among plants. Plant CellPhysiol 34: 1055–1062.
Jongedijk E, Tigelaar H, van Roekel JS, Bres-Vloemans SA,Dekker I, van den Elzen PJ, Cornelissen BJ and MelchersLS (1995) Synergistic activity of chitinases and b-1,3-gluca-nases enhances fungal resistance in transgenic tomatoplants. Euphytica 85: 173–180.
Kachroo P, Yoshioka K, Shah J, Dooner HK and KlessigDF (2000) Resistance to turnip crinkle virus in Arabidopsisis regulated by two host genes and is salicylic acid depen-dent but NPR1, ethylene, and jasmonate independent. PlantCell 12: 677–690.
Kaniewski W, Ilardi V, Tomassoli L, Mitsky T, Layton J andBarba M (1999) Extreme resistance to cucumber mosaicvirus (CMV) in transgenic tomato expressing one or twoviral coat proteins. Mol Breeding 5: 111–119.
Kim HS and Delaney TP (2002) Over-expression of TGA5,which encodes a bZIP transcription factor that interactswith NIM1/NPR1, confers SAR-independent resistance inArabidopsis thaliana to Peronospora parasitica. Plant J 32:151–163.
Kinkema M, Fan W and Dong X (2000) Nuclear localizationof NPR1 is required for activation of PR gene expression.Plant Cell 12: 2339–2350.
Kohler A, Schwindling S and Conrath U (2002) Benzothi-adiazole-induced priming for potentiated responses to path-ogen infection, wounding, and infiltration of water intoleaves requires the NPR1/NIM1 gene in Arabidopsis. PlantPhysiol 128: 1046–1056.
Lee TJ, Coyne DP, Clemente TE and Mitra A (2002) Partialresistance to bacterial wilt in transgenic tomato plantsexpressing antibacterial Lactoferrin gene. J Amer Soc HortSci 127: 158–164.
Li L and Steffens JC (2002) Overexpression of polyphenoloxidase in transgenic tomato plants results in enhanced bac-terial disease resistance. Planta 215: 239–247.
Li X, Clarke JD, Zhang Y and Dong X (2001) Activation ofan EDS1-mediated R-gene pathway in the snc1 mutantleads to constitutive, NPR1-independent pathogen resis-tance. Mol Plant Microbe Interact 14: 1131–1139.
Liu D, Raghothama KG, Hasegawa PM and Bressan RA(1994) Osmotin overexpression in potato delays develop-ment of disease symptoms. Proc Natl Acad Sci USA 91:1888–1892.
Liu Y, Schiff M, Marathe R and Dinesh-Kumar SP (2002)Tobacco Rar1, EDS1 and NPR1/NIM1 like genes arerequired for N-mediated resistance to tobacco mosaic virus.Plant J 30: 415–429.
Malamy J, Carr JP, Klessig DF and Raskin I (1990) Salicylicacid: a likely endogenous signal in the resistance responseof tobacco to viral infection. Science 250: 1002–1004.
McDowell JM, Cuzick A, Can C, Beynon J, Dangl JL andHolub EB (2000) Downy mildew (Peronospora parasitica)resistance genes in Arabidopsis vary in functional require-ments for NDR1, EDS1, NPR1 and salicylic acid accumu-lation. Plant J 22: 523–529.
Metraux JP, Signer H, Ryals J, Ward E, Wyss-Benz M,Gaudin J, Raschdorf K, Schmid E, Blum W and Inver-
ardi B (1990) Increase in salicylic acid at the onset ofsystemic acquired resistance in cucumber. Science 250:1004–1006.
Mou Z, Fan W and Dong X (2003) Inducers of plant sys-temic acquired resistance regulate NPR1 function throughredox changes. Cell 113: 935–944.
Murphy AM, Chivasa S, Singh DP and Carr JP (1999) Sali-cylic acid-induced resistance to viruses and other pathogens:a parting of the ways? Trends Plant Sci 4: 155–160.
Murphy AM, Gilliland A, Wong CE, West J, Singh DP andCarr JP (2001) Signal transduction in resistance to plantviruses. European Journal of Plant Pathology 107: 121–128.
Pieterse CM, van Wees SC, van Pelt JA, Knoester M, LaanR, Gerrits H, Weisbeek PJ and van Loon LC (1998) Anovel signaling pathway controlling induced systemic resis-tance in Arabidopsis. Plant Cell 10: 1571–1580.
Rairdan GJ and Delaney TP (2002) Role of salicylic acid andNIM1/NPR1 in race-specific resistance in Arabidopsis.Genetics 161: 803–811.
Rasmussen JB, Hammerschmidt R and Zook MN (1991) Sys-temic induction of salicylic acid accumulation in cucumberafter inoculation with Pseudomonas syringae pv. syringae.Plant Physiol 97: 1342–1347.
Robison MM, Shah S, Tamot B, Pauls KP, Moffatt BA andGlick BR (2001) Reduced symptoms of Verticillium wilt intransgenic tomato expressing a bacterial ACC deaminase.Mol Plant Pathology 2: 135–145.
Rommens CM and Kishore GM (2000) Exploiting the fullpotential of disease-resistance genes for agricultural use.Curr Opin Biotechnol 11: 120–125.
Ryals J, Weymann K, Lawton K, Friedrich L, Ellis D, SteinerHY, Johnson J, Delaney TP, Jesse T, Vos P and Uknes S(1997) The Arabidopsis NIM1 protein shows homology tothe mammalian transcription factor inhibitor IjB. PlantCell 9: 425–439.
Ryals JA, Neuenschwander UH, Willits MG, Molina A, Stei-ner HY and Hunt MD (1996) Systemic acquired resistance.Plant Cell 8: 1809–1819.
Saghai-Maroof MA, Soliman KM, Jorgensen RA and AllardRW (1984) Ribosomal DNA spacer-length polymorphismsin barley: mendelian inheritance, chromosomal location,and population dynamics. Proc Natl Acad Sci USA 81:8014–8018.
Salmeron JM and Vernooij B (1998) Trnasgenic approachesto microbial disease resistance in crop plants. Curr OpinPlant Biology 1: 347–352.
Sambrook J and Russell DW (2001) Molecular Cloning. ALaboratory Manual (3rd edition). Cold Spring Harbor Lab-oratory Press, Cold Spring Harbor, New York.
Shah DM (1997) Genetic engineering for fungal and bacterialdisease. Curr Opin Biotechnol 8: 208–214.
Shah J, Tsui F and Klessig DF (1997) Characterization of asalicylic acid-insensitive mutant (sai1) of Arabidopsis thali-ana, identified in a selective screen utilizing the SA-induc-ible expression of the tms2 gene. Mol Plant MicrobeInteract 10: 69–78.
Shih SL, Wang JT, Chiang BT and Green SK (1995) Distri-bution of tomato leaf curl virus in Taiwan and its hostrange. Plant protection Bulletin, R.O.C. 37: 445.
Shirano Y, Kachroo P, Shah J and Klessig DF (2002) Again-of-function mutation in an Arabidopsis toll interleu-kin1 receptor-nucleotide binding site-leucine-rich repeattype R gene triggers defense responses and results inenhanced disease resistance. Plant Cell 14: 3149–3162.
580

Spoel SH, Koornneef A, Claessens SM, Korzelius JP, vanPelt JA, Mueller MJ, Buchala AJ, Metraux JP, Brown R,Kazan K, van Loon LC, Dong X and Pieterse CM (2003)NPR1 modulates cross-talk between salicylate- and jasmo-nate-dependent defense pathways through a novel functionin the cytosol. Plant Cell 15: 760–770.
Stommel JR, Tousignant ME, Wai T, Pasini R and M. KJ(1998) Viral satellite RNA expression in transgenic tomatoconfers field tolerance to cucumber mosaic virus. Plant Dis-ease 82: 391–396.
Stuiver MH and Custers JH (2001) Engineering disease resis-tance in plants. Nature 411: 865–868.
Subramaniam R, Desveaux D, Spickler C, Michnick SW andBrisson N (2001) Direct visualization of protein interactionsin plant cells. Nature Biotechnol 19: 769–772.
Tabaeizadeh Z, Agharbaoui Z, Harrak H and Poysa V (1999)Transgenic tomato plants expressing a Lycopersicon chilensechitinase gene demonstrate improved resistance to Verticil-lium dahliae race 2. Plant Cell Rep 19: 197–202.
Tai TH, Dahlbeck D, Clark ET, Gajiwala P, Pasion R, Wha-len MC, Stall RE and Staskawicz BJ (1999) Expression ofthe Bs2 pepper gene confers resistance to bacterial spot dis-ease in tomato. Proc Natl Acad Sci USA 96: 14153–14158.
Tang X, Xie M, Kim YJ, Zhou J, Klessig DF and Martin GB(1999) Overexpression of Pto activates defense responses andconfers broad resistance. Plant Cell 11: 15–30.
Vanacker H, Lu H, Rate DN and Greenberg JT (2001) A rolefor salicylic acid and NPR1 in regulating cell growth inArabidopsis. Plant J 28: 209–216.
Wang JF, Hanson P and Barnes JA (1998) Worldwide evalua-tion of an international set of resistance sources to bacterialwilt in tomato. in: Prior P, Allen C and Elphinstone J(eds.), Bacterial Wilt Disease. Molecular and EcologicalAspects (pp. 269–275) Springer-Verlag Press, Berlin.
Wang JF and Lin CH (2004) Colonization capacity of Ralsto-nia solanacearum tomato strains differing in aggressivenesson tomatoes and weeds. In: Allen C, Prior P and HaywardAC (eds). Bacterial Wilt: The Disease and the Ralstonia so-lanaceraum Species Complex APS Press, St Paul (in press).
Wang JF, Olivier J, Thoquet P, Mangin B, Sauviac L andGrimsley NH (2000) Resistance of tomato line Hawaii7996to Ralstonia solanacearum Pss4 in Taiwan is controlledmainly by a major strain-specific locus. Mol Plant MicrobeInteract 13: 6–13.
Ward ER, Uknes SJ, Williams SC, Dincher SS, WiederholdDL, Alexander DC, Ahl-Goy P, Metraux JP and Ryals JA(1991) Coordinate gene activity in response to agents thatinduce systemic acquired resistance. Plant Cell 3: 1085–1094.
Whitham S, McCormick S and Baker B (1996) The N gene oftobacco confers resistance to tobacco mosaic virus in trans-genic tomato. Proc Natl Acad Sci USA 93: 8776–8781.
Wong CE, Carson RA and Carr JP (2002) Chemicallyinduced virus resistance in Arabidopsis thaliana is indepen-dent of pathogenesis-related protein expression and theNPR1 gene. Mol Plant Microbe Interact 15: 75–81.
Yu D, Chen C and Chen Z (2001) Evidence for an importantrole of WRKY DNA binding proteins in the regulation ofNPR1 gene expression. Plant Cell 13: 1527–1539.
Zhang C and Shapiro AD (2002) Two pathways act in anadditive rather than obligatorily synergistic fashion toinduce systemic acquired resistance and PR gene expres-sion. BMC Plant Biol 2: 9.
Zhang Y, Fan W, Kinkema M, Li X and Dong X (1999)Interaction of NPR1 with basic leucine zipper protein tran-scription factors that bind sequences required for salicylicacid induction of the PR-1 gene. Proc Natl Acad Sci USA96: 6523–6528.
Zhou JM, Trifa Y, Silva H, Pontier D, Lam E, Shah Jand Klessig DF (2000) NPR1 differentially interacts withmembers of the TGA/OBF family of transcription factorsthat bind an element of the PR-1 gene required forinduction by salicylic acid. Mol Plant Microbe Interact13: 191–202.
Zhu Q, Maher EA, Masoud S, Dixon RA and Lamb C(1994) Enhanced protection against fungal attack by con-stitutive co-expression of chitinase and glucanase genes intransgenic tobacco. Bio/Technology 12: 807–812.
581




















