
Detection of Begomovirus in chilli and tomato plants using ...

151
Expression of PR-5 Protein Thaumatin II for Improving Disease Resistance and Fruit Quality of Tomato I.V. Korneeva1, K.A. Shestibratov2, N.V. Lavrova1, A.P. Firsov2, V.G. Lebedev2, P.N. Kharchenko1 and S.V. Dolgov1,2 1Department of Genetic Engineering, All-Russia Research Institute of Agricultural Biotechnology, 127550, Timiryasevskaya 42, Moscow, Russia
2Artificial Climate Station "Biotron", Branch of Shemyakin Institute of Bioorganic Chemistry, 142290, Pushchino, Moscow Region, Russia
Keywords: tomato, transformation, Agrobacterium tumefaciens, Phytophthora infestans,
disease resistance, inheritance Abstract
A thaumatin II cDNA (thau II) driven by the CaMV 35S promoter was introduced into tomato line YALF via Agrobacterium-mediated transformation. The pBI121-based binary vector for expression of this gene was constructed using a coding sequence cloned at Leiden University. Twenty-three transformants obtained on the selective media with kanamycin were tested for the presence of the neomycin phosphotransferase (nptII) and thaumatin II (thauII) genes. The nptII gene was detected in all kanamycin-resistant plants but among them only 16 contained thauII. Transformation frequency was 9.4%. Accumulation of the foreign protein in leaves and fruits of transgenic plants was confirmed by Western blotting. The foreign gene expression in the fruits was detected organoleptically. Greenhouse testing confirmed enhanced resistance against Phytophtora infestans for two thaumatin-producing tomato lines. Inheritance of transferred genes in the T1 generation and vector design for cell compartmentalization of foreign protein expression will be discussed. INTRODUCTION
Tomato (Lycopersicon esculentum Mill.) is a leading vegetable crop in the world; moreover, tomato is a convenient object for genetic-engineering manipulations focused on producing higher quality fruit and other agronomic properties and enhancing plant resistance towards biotic and abiotic stress factors. Plant resistance genes have advantage over genes from other organisms as they do not need modification to prevent breakdown of their product by plant proteases. The thaumatin II gene from Thaumatococcus daniellii (Edens et al., 1982) codes a super sweet protein (several thousand times sweeter than sucrose) and this offers an important side benefit of improving fruit quality for Russian varieties of tomato, which are sufficiently winter hardy but have modest taste characteristics.
Several proteins belonging to the PR-5 group have been used successfully to enhance plant resistance to fungal pathogens (review: Punja, 2001). For example, Zhu et al. (1996) demonstrated that transgenic potato plants constitutively expressing high levels of an osmotin-like protein, a member of the PR-5 group, showed increased resistance to the late blight fungus, Phytophtora infestans. Transgenic rice plants with constitutive expression of rice thaumatin-like protein (TLP) also had enhanced resistance to the pathogen Rhizoctonia solani (Datta et al., 1999). Enhanced resistance of transgenic wheat plants to Fusarium graminearum due to the integration and expression of rice TLP was also demonstrated (Chen et al., 1999).
Recently, Szwacka et al. (2002) generated transgenic cucumber plants and their progeny with the thaumatin II gene, driven by the 35S promoter. The transgenic plants showed enhanced resistance against the pathogenic fungus Pseudoperonospora cubensis; moreover, 46% of the progenies in the regeneration exhibited Mendelian segregation. The phenomenon of non-Mendelian segregation was also documented (Yin et al., 2004).
The aims of the present investigation were: 1) to develop transgenic tomato plants with the gene encoding thaumatin II protein through Agrobacterium-mediated
Proc. XVth EUCARPIA Tomato Eds.: M. Cirulli et al. Acta Hort. 789, ISHS 2008

152
transformation; 2) to characterize and evaluate the resistance to Phytophtora infection of transgenic plants expressing the thaumatin II gene; and 3) to study the inheritance of kanamycin resistance in self-pollinated T1 progenies. MATERIALS AND METHODS
Cotyledons from 2- to 3-week-old tomato seedlings line YALF were cultivated in vitro on standard MS medium with 30 g L-1 sucrose and 6–8 g L-1 agar and used as explants for Agrobacterium-mediated transformation experiments.
The binary plasmid vector pBIThau35 was constructed by replacing the β-glucuronidase uidA gene of pBI121 with the 1.0 kb DNA fragment of preprothaumatin II cDNA from plasmid pUR528, which had been kindly provided by Dr. A.M. Ledeboer (Edens et al., 1982). This vector was transferred into Agrobacterium tumefaciens strain CBE21 (Revenkova et al., 1993; Patent application, 2003).
Tomato plants were transformed in accordance with McCormic and Niedermeyer’s (1986) modified protocol.
The presence of the thauII gene was detected by polymerase chain reaction (PCR) as described by Haymes and Davis (1998) both in kanamycin-resistant shoots and in greenhouse plants. DNA was isolated from leaves according to Edwards et al. (1991).
The expression of thauII was evaluated by Western blotting according to the standard protocol (Тowbin et al., 1979).
The evaluation of the thaumatin II expression effect on tomato disease resistance to P. infestans was carried out for transgenic lines both with the nptII and thauII genes, the nptII gene only and non-transgenic control plants. The plants were cultivated in a greenhouse until August and then subjected to the resistance test (Stroikov, 1993).
The segregation ratio was analysed in seedlings by chi-square test. The seeds were grown on MS medium with 150 mg L-1 kanamycin. RESULTS AND DISCUSSION
A number of experiments defined optimal growth regulator concentrations (0.1 mg L-1 IAA, 5 mg L-1 BA) and these resulted in regeneration of 170 putative transgenic tomato lines (Fig. 1). The regeneration efficiency was 74%. Twenty-three transformants were selected on the medium with kanamycin (100 mg L-1) (Fig. 2).
Kanamycin-resistant regenerants were tested for the presence of the neomycin phosphotransferase (nptII) and thaumatin II (thauII) genes. The single nptII gene was detected in all kanamycin-resistant plants, but only 16 lines contained both genes (thauII and nptII) (Fig. 3). In the latter case transformation frequency was 9.4%.
Accumulation of foreign protein in leaves and fruits of transgenic lines was confirmed by two kinds of assays: 1) Western blotting (Fig. 4 and 5), and 2) sensory evaluation (organoleptically). The thaumatin II gene expression level differed in various lines: moreover, transgenic tomatoes were much sweeter than control samples and possessed a specific aftertaste. Szwacka et al. (2002) had produced transgenic cucumber fruit (Cucumis sativus L.), which exhibited a sweet phenotype. There was a positive correlation between thaumatin accumulation levels and sweet taste intensity.
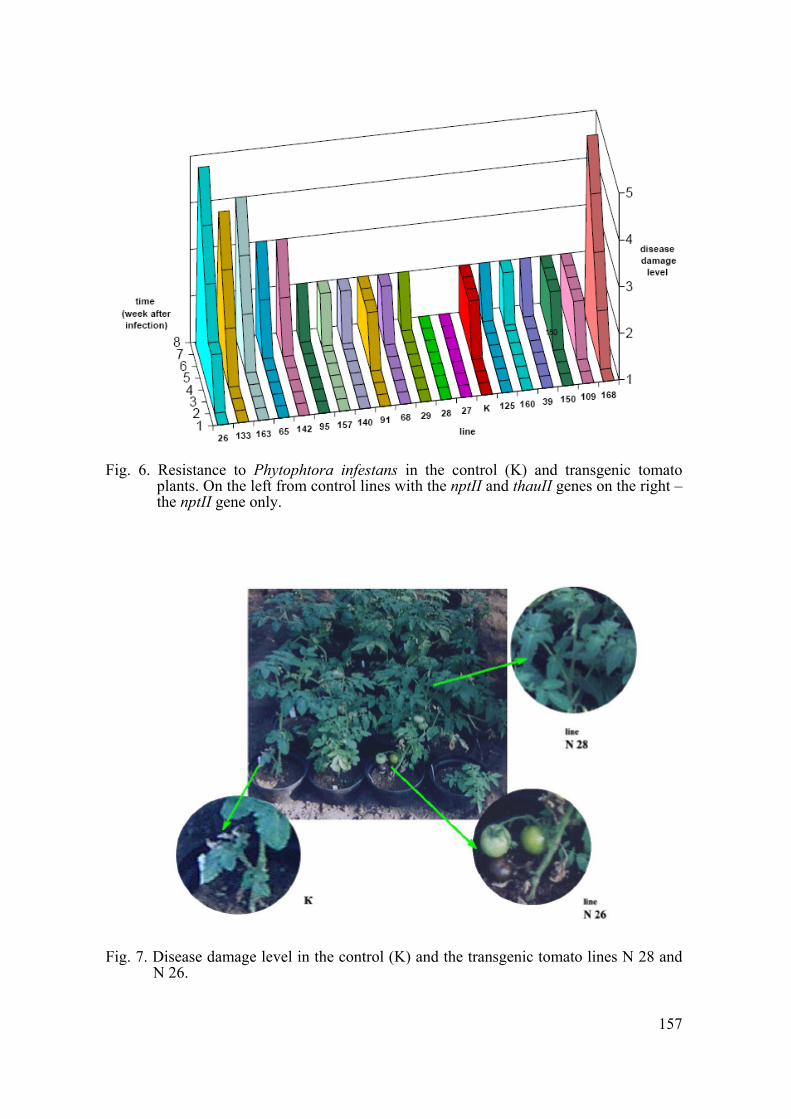
Resistance to P. infestans for greenhouse-grown transgenic lines both with the nptII and thauII genes, the nptII gene only and non-transgenic control plants was evaluated in accordance with Stroikov’s (1993) test. They were infected by Phytophtora strucked plants (cotyledon stage) during one week. Disease damage level was evaluated within the next 8 weeks on a 1–5 scale. Greenhouse testing confirmed high disease susceptibility at the early stage for control plants and enhanced resistance against P. infestans for two thaumatin-producing tomato lines - N 27 and N 28 (Fig. 6 and 7).
Segregation of kanamycin-resistance was analysed in T1 seedlings from 10 transgenic lines both in vitro on kanamycin-containing medium and in vivo in kanamycin water solution (Fig. 8). The selection concentration was 150 mg L-1.
About half of them exhibited a non-Mendelian segregation (Table 1). This is a notorious fact for Agrobacterium-mediated transformed plants with a low copy number.

153
Interpretation of the inheritance and stability of the newly introduced transgenes is great importance for determining the value and application of genetically engineered organisms in agriculture (Yin et al., 2004). Literature Cited Chen, W.P., Chen, P.D., Liu, D.J., Kynast, R., Friebe, B., Velazhahan, R.,
Muthukrishnan, S. and Gill, B.S. 1999. Development of wheat scab symptoms is delayed in transgenic wheat plants that constitutively express a rice thaumatin-like protein gene. Theor. Appl. Genet. 99:755–760.
Datta, K., Velazhahan, R., Oliva, N., Ona, I., Mew, T., Khush, G.S., Muthukrishnan, S. and Datta, S.K. 1999. Overexpression of cloned rice thaumatin-like protein (PR-5) gene in transgenic rice plants enhances environmental friendly resistance to Rhizoctonia solani causing sheath blight disease. Theor. Appl. Genet. 98:1138–1145.
Edens, L., Heslinga, L., Klok, R., Ledeboer, A.M., Maat, J., Toonen, M.Y., Visser, C. and Verrips, C.T. 1982. Cloning of cDNA encoding the sweet-tasting plant protein thaumatin and its expression in Escherichia coli. Gene 18(1):1–12.
Edwards, K., Johnstone, C. and Thompson, C. 1991. A simple and rapid method for the preparation of plant genomic DNA for PCR analysis. Nucleic Acids Res. 19:1349.
Haymes, K.M. and Davis, T.M. 1998. Agrobacterium-mediated transformation of Alpine Fragaria vesca, and transmission of transgenes to R1 progeny. Plant Cell Rep. 17:279–283.
McCormic, S. and Niedermeyer, J. 1986. Leaf disk transformation of cultivated tomato. Plant Cell Rep. 5:81–84.
Patent application PCT/RU 2003/00043 15.10.2003. Punja, Z.K. 2001. Genetic engineering of plants to enhance resistance to fungal
pathogens: a review of progress and future prospects. Can. J. Plant Pathol. 23:6–235. Revenkova, E.V., Kraev, A.S. and Skryabin, K.G. 1993. Construction of a disarmed
derivative of the supervirulent Ti plasmid pTiBo542. p.67–76. In: K.G. Skryabin (ed.), Plant biotechnology and molecular biology. Pushchino Research Centre, Moscow.
Stroikov, Y.M. 1986. Methods of infection background usage for the evaluation of plant resistance to diseases, Moscow: Timiryazevsk. S-kh. Akad.
Szwacka, M., Krzymowska, M., Osuch, A., Kowalczyk, M.E. and Malepszy, S. 2002. Variable properties of transgenic cucumber plants containing the thaumatin II gene from Thaumatococcus daniellii. Acta Physiol. Plant. 24(2):173–185.
Towbin, H., Staehelin, T. and Gordon, J. 1979. Electrophoretic transfer of proteins from acrilamide gels to nitrocellulose sheets: procedure and some applications. Proc. Natl. Acad. Sci. U.S.A. 76:4350–4354.
Yin, Z., Plader, W. and Malepszy, S. 2004. Transgene inheritance in plants. J. Appl. Genet. 45(2):127–144.
Zhu, B., Chen, T.H.H. and Li, P.H. 1996. Analysis of late-blight disease resistance and freezing tolerance in transgenic potato plants expressing sense and antisense genes for an osmotin-like protein. Planta 198:70–77.

154
Tables Table 1. Segregation of kanamycin-resistance in tomato transgenic lines. Plant line
Total number of seedlings
Kanamycin-resistant (KmR)
Kanamycin-sensitive (KmS) χ2 Segregation
ratio 26 125 96 29 0.21 3:1 28 125 92 33 0.13 3:1 39 112 78 34 0.45 2:1 53 101 77 24 0.08 3:1 65 137 110 27 0.006 4:1 68 133 103 30 0.43 3:1 91 166 130 36 0.97 3:1 95 112 75 37 0.004 2:1 133 86 28 58 1:2 145 123 93 30 0.03 3:1 Figures
Fig. 1. Shoot regeneration from cotyledon explants on standard MS medium with optimal
growth regulators concentrations (0.1 mg L-1 IAA, 5 mg L-1 BA).

155
Fig. 2. Tomato transgenic lines selected on the medium with kanamycin (100 mg L-1).
Fig. 3. PCR analysis of DNA samples from tomato transgenic plants. a) npt II; b) thau II.

156
Fig. 4. Western blot analysis of proteins from tomato transgenic plants.
Fig. 5. Western blot analysis. Different level of the thaumatin II gene expression. (1-6 –
protein thaumatin II; 31,2; 62,5; 125,0; 250,0; 500,0; 1000,0 ng accordingly; 22 - 250 ng thaumatin II; 7 – nontransgenic plant; 8-21 – transgenic plants, lines NN 26, 27, 28, 53, 68, 91, 95, 109, 115, 140, 145, 157, 163, 168, respectively).
157
Fig. 6. Resistance to Phytophtora infestans in the control (K) and transgenic tomato
plants. On the left from control lines with the nptII and thauII genes on the right – the nptII gene only.
Fig. 7. Disease damage level in the control (K) and the transgenic tomato lines N 28 and
N 26.

158
Fig. 8. Selection of kanamycin-resistance transgenic lines of tomato in kanamycin water
solution.
Copyright © 2022 FDOKUMEN




















