Mapping of quantitative trait loci for basmati quality traits in rice (Oryza sativa L.)
Upload
independentCategory
view
3download
0
1 23
Plant Molecular Biology Reporter ISSN 0735-9640Volume 30Number 1 Plant Mol Biol Rep (2012) 30:204-213DOI 10.1007/s11105-011-0329-y
Transgenic Lines of Oryza Sativa (Basmati2000) Expressing Xa21 DemonstrateEnhanced Resistance to Bacterial Blight
Amber Afroz, Afsari Qureshi, MuznaZahur, Umer Rashid & Hamid Rashid

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer-
Verlag. This e-offprint is for personal use only
and shall not be self-archived in electronic
repositories. If you wish to self-archive your
work, please use the accepted author’s
version for posting to your own website or
your institution’s repository. You may further
deposit the accepted author’s version on a
funder’s repository at a funder’s request,
provided it is not made publicly available until
12 months after publication.

Transgenic Lines of Oryza Sativa (Basmati 2000) ExpressingXa21 Demonstrate Enhanced Resistance to Bacterial Blight
Amber Afroz & Afsari Qureshi & Muzna Zahur &
Umer Rashid & Hamid Rashid
Published online: 6 July 2011# Springer-Verlag 2011
Abstract An efficient transformation system for rice wasestablished by co-cultivating calli, derived from 21-day-oldscutellum, with Agrobacterium tumefaciens cells (OD600=0.04), maintained on filter paper, moistened with 4 mL of amedium, and supplemented with 400 μM acetosyringone,for a period of 2 days. Presence of the transgene wasconfirmed by polymerase chain reaction, and stableintegration and copy number of the transgene weredetermined by Southern blot analysis. Among seven plantsanalyzed, six possessed single T-DNA integration events,while one plant was found to have two integrated copies ofthe T-DNA. A total of 45 T0 plants were grown in thegreenhouse to obtain the T1 generation. T1 plants evaluatedfor presence of the transgene and for response to inocula-tion with the bacterial leaf blight pathogen, Xanthomonas
oryzae pv. Oryzae, exhibited Mendelian segregation (3:1)for the transgene as well as enhanced resistance to bacterialblight
Keywords Bacterial blight . Hygromycin .
Acetosyringone . Basmati . Transformation
AbbreviationsHyg HygromycinHpt Hygromycin phosphotransferaseAs AcetosyringoneYEP Yeast extract phosphateAA Amino acid mediaPCR Polymerase chain reactionBB Bacterial blightCf CefotaximeCb CarbenecillinDAI Days after inoculation
Introduction
Rice (Oryza sativa L.) is one of the major food crops of theworld. Rice production is affected by various abiotic andbiotic stresses including bacterial leaf blight (BLB) causedby Xanthomonas oryzae pv. Oryzae (Xoo) (Swings et al.1990). BLB is a vesicular disease resulting in systematicinfection that produces tanish gray to whitish lesion (attillering) that increase with plant growth, peaking at theflowering stage, and resulting in 30–50% loss in yield(Adhikari et al. 1999). Xa21 confers resistance to multiplepathogen isolates of Xoo encoding a receptor-like kinase, ismember of multigene family (Song et al. 1997; Wang et al.1996). Gene introduction for resistance to bacterial blight
Electronic supplementary material The online version of this article(doi:10.1007/s11105-011-0329-y) contains supplementary material,which is available to authorized users.
A. Afroz :U. RashidPlant Biotechnology Programme,National Agriculture Research Council,Islamabad, Pakistan
A. QureshiDepartment of Biochemistry, Faculty of Biological Sciences,Quaid-i-Azam University Islamabad,Islamabad, Pakistan
A. Afroz (*) :M. ZahurDepartment of Biochemistry Nawaz Sharif Medical College,University of Gujrat Hafiz Hayat Campus,Gujrat, Pakistane-mail: [email protected]
H. RashidDepartment of Bioinformatics, Muhammad Ali Jinnah University,Islamabad, Pakistan
Plant Mol Biol Rep (2012) 30:204–213DOI 10.1007/s11105-011-0329-y
Author's personal copy

(BB) using conventional breeding approaches is generallytedious and time consuming, resulted in linkage drags andBasmati rice to have narrow genetic base, and scarcity ofdonor parents for grain quality along with poor combiningability (Zhai et al. 2001). Recombinant DNA technologyprovides a direct approach and has opened up a plethora togenetically manipulate plants across the species barrier thusmaking it possible to transfer any gene to desired crops(Birch 1997). The severity and significance of damagecaused by BB have necessitated the development ofstrategies to control crop loss and avert an epidemic. It isdifficult to transform indica rice varieties using traditionaltransformation methods due to their poor regeneration (Linet al. 2009). Transfer of DNA to rice plants has beenaccomplished by electroporation, protoplast transformation,cell suspension of cells, and Agrobacterium-mediatedtransformation (Tada et al. 1990; Uchimiya et al. 1986;Wagiran et al. 2010; Guo et al. 2011). Agrobacteriumtumefaciens-mediated transformation was first reported injaponica rice and indica cultivars found to be recalcitrantfor A. tumefaciens-mediated transformation (Chan et al.1992; Hiei et al. 1994). Rashid et al. (1996) first reportedthe successful transformation of scutellum-derived callusesof indica cvs followed by others. Numerous donors forresistance to Xoo have been identified (Kinoshita 1995; Linet al. 1996). In many areas in Indonesia, India, China, andthe Philippines, where rice varieties with Xa4 gene havebeen widely grown, new races of the BB pathogen haveemerged. Therefore, new genes for BB resistance are beingincorporated into elite rice varieties. A dominant gene Xa21for resistance to BB was transferred from a wild species(Oryza longistaminata) to the cultivated variety IR24 (Khushet al. 1990). Xa21 confers resistance to all the known races ofXoo in India and the Philippines (Khush et al. 1990). Wang etal. (1996) transformed japonica rice variety T309 with clonedXa21, and its resistance spectrum was similar to that of thedonor line IRBB21. However, T309 is no longer cultivated,and no new commercial rice cultivars with Xa21 have beenreleased. In this study, indica rice variety Basmati 2000 wastransformed with Xa21. Research is conducted to check theresistance of Basmati 2000 to BB in lines expressing the Xa21gene. The successful production of transgenic plants of thiscultivar will open the plethora for introducing agronomicallyuseful traits into any commercially grown cultivars, providingthe best opportunity for maximizing yield.
Materials and Methods
O. sativa (Basmati 2000) seeds were obtained fromMonsanto Islamabad. Nine-centimeter-diameter petri plateswere used in this experiment and all the plates were sealedwith medical-permeable tape (Microspore Surgical Tape;
3 M). Seeds were surface sterilized according to the methodof Rashid et al. (1996). To remove surfactants, sterilizedseeds were rinsed three times with deionized sterile waterand blotted on sterile filter paper. Seeds were germinatedfor 2 weeks in a growth chamber in the dark and afterwardsshifted to fluorescent light (600 μmol m−2 s−1; 16 h light/8 h dark) at 25±1°C and 70% relative humidity. Seed-derived calli (18, 21, 25, 28, and 35 days old) were used forAgrobacterium-mediated genetic transformation.
A. tumefaciens Culture and Co-Cultivation with Explants
A. tumefaciens strain (EHA101) with pTCL5, containingXa21 gene, hygromycin phospho transferase gene (Hpt),and an intron-beta-glucuronidase (GUS) gene was used fortransformation (Fig. 1). A single colony of A. tumefacienscarrying the binary vector was inoculated in yeast extractphosphate (YEP) (Table 1) broth containing 50 mg L−1
kanamycin and hygromycin (hyg) and grown overnight onrotary shaker at 28±2°C. Overnight-grown bacterial pre-cultures were adjusted to OD600=0.04. Cultures werecollected by centrifugation at 4,000×g for 10 min at roomtemperature, and resuspended in co-cultivation liquidmedium (Table 1) with different concentrations of acetosyr-ingone (As). The 18–35-day-old scutellum-derived calliwere subjected to A. tumefaciens infection. Calli weresubmerged in the bacterial suspension for 15 min andblotted onto a sterile paper towel. Rice calli were co-cultured on a double filter paper according to the method ofOzawa (2009). Fifteen pieces of calli (2–4 mm in diameter)were placed on a plastic petri dish (9 cm in diameter) containingdouble filter paper moistened with 3–4 mL of liquid co-cultivation medium (AA-As; Table 1) for 1–3 days at 28±2°Cin the dark. To compare effect of As on transformationefficiency, AA along with different concentrations of As (50–400 μM) was used in co-cultivation (Table 1).
GUS Assay
Histochemical GUS assay was carried out essentially asdescribed by Jefferson (1987). Infected scutellum-derived calliwere incubated in x-gluc solution containing 1 mg 5-bromo-4-chloro-3-indolyl-β-D- glucuronide, 0.5% triton X-100, 20%methanol, and 50 mM sodium phosphate (pH 7.0). Thereaction mixture was incubated at 37°C overnight.
Selection and Shoot Regeneration
After co-cultivation, calluses were washed with washingselection media (Table 1). Calli were pre-cultured on thepetri plates with As (50 μM) on callus induction mediumwithout hyg (Table 1) for 3 days. Afterwards, calli wereshifted to selection medium, followed by shifting to
Plant Mol Biol Rep (2012) 30:204–213 205
Author's personal copy

selection regeneration medium within 7 days. After 4 weeks,regenerated shoots were transferred to the same selectionmedia for further development (Table 1). Elongated shoots(3–5 cm) were detached from primary explants andtransferred to the rooting medium (Table 1). After 2 weeks,the rooted plantlets were transferred to soil.
DNA Isolation and Polymerase Chain Reaction Analysis
Genomic DNA was extracted from leaf tissues of control andtransgenic plants according to cetyltrimethylammonium bro-mide (CTAB) method (Sambrook and Russell 2001). Forwardand reverse primers used for Xa21 gene (1.4 kb) were F 5–3'(ATA GCA ACT CAT TGC TTGG) and R 3–5' (CGA TCGCTA TAA CAG CAA AAC). Ten microliters of polymerasechain reaction (PCR) reactions contained 10 ng of thetemplate DNA, 2 μl of 10× PCR buffer, 0.6 μl of 0.1 MMgCl2, 1 μL of 10 mM dNTP mixture, 0.5 μl of 5 μM eachforward and reverse primer, 0.3 μL of Taq polymerase and4.1 μl of nanopure water. PCR reactions were run for35 cycles with an initial denaturation step for 3 min (94°C),annealing temperature (51°C for Xa21) for 2 min andextension (72°C) for 3 min, followed by 35 cycles of
denaturation (94°C) for 1 min, annealing (52°C) for 1 minand extension (72°C) for 2 min with a final extension cycle of15 min (72°C). The amplified PCR products were analyzedby agarose gel electrophoresis on 0.8% (w/v) agarose gel.
Southern Blot Analysis
Genomic DNA samples used for Southern blot analysiswere extracted and purified from young leaves of trans-genic and non-transgenic plants using the CTAB method(Sambrook and Russell 2001). For this purpose, genomicDNA (10 μg) of selected lines was digested with Pst I andincubated at 37°C overnight. The digested DNA wasfractionated on 0.8% agarose gel supplemented with 10 gmL−1 ethidium bromide at 40 V in TAE buffer for 5–6 h.DNA was depurinated, denatured, neutralized, and wastransferred onto nylon membrane (Hybond-Amersham) bycapillary method using 10× SSC (150 mM sodium citrateand 1.5 M NaCl) as transfer buffer for 20 h. The membranewas removed from gel and crosslinked in UV crosslinker(CL-1000 Ultraviolet Crosslinker-UVP) at 120 mJ cm−2
energy. To block the attachment of probe to non-specificnucleic acid binding sites, membrane was treated with
Fig. 1 Schematic diagram of a part of the T-DNA region oftransformation vector pTCL5 (RB) right border, (LB) left border,(35S) cauliflower mosaic virus 35S promoter, ß-glucuronidase coding
region (GUS), first intron of the caster bean catalase gene (INT),hygromycin phosphotransferase (Hpt)
Table 1 Media used for transformation protocol and regeneration of transformed calli. All medium except (Wakimotos and inoculation medium)contain 30 gL−1 sucrose, 4 gL−1 gelrite, except liquid media and pH is adjusted to 5.7–5.8
Media Abbreviation Composition
Wakimoto's WAM Peptone (5 g), Ca (No3)2 (0.5 g), sodium hydrogen phosphate (0.82 g),ferrous sulfate (0.05 g), agar (15 g)
Inoculation YEP Yeast extract (10 g), bactopeptone (10 g), NaCL (5 g), pH adjusted to 7.2
Callus induction CIM (solid) N6 salts and vitamins (Chu et al. 1975), 2,4-D (2 mg L−1)
Co-cultivation CIM (liquid)+As N6 salts and vitamins, 2, 4-D (2 mg L−1), sorbitol (30 gL−1), casein hydrolysate(2 gL−1), As (50–400 μM)
AA+As AA media (Toriyama and Hinata 1985), 2, 4-D (2 mg L−1), As (50–400 μM)
Washing WM N6 salts and vitamins, 2, 4-D (2 mg L−1), Cf (500 mg L−1)
Pre-selection CIM+Cf N6 salts and vitamins+2, 4-D (2 mg L−1), sorbitol (30 gL−1), casein hydrolysate(2 gL−1), Cf (1,000 mg L−1)
Selection RM+hyg+Cf N6 salts and vitamins, 2, 4-D (2 mg L−1), hyg (50 mg L−1), Cf (1000 mg L−1),sorbitol (30 gL−1), casein hydrolysate (2 gL−1)
Selection–regeneration RM+hyg+Cf MS salts and vitamins (Murashige and Skoog 1962), NAA (1 mg L−1), BAP(5 mg L−1), sorbitol (30 gL−1), casein hydrolysate (2 gL−1), hyg (50 mg L−1),Cf (1,000 mg L−1)
Rooting RM MS salts and vitamins (Murashige and Skoog 1962), IAA (0.5 mg L−1), hyg (50 mg L−1)
2, 4 D 2, 4-dichlorophenoxyacetic acid; IAA indole acetic acid; NAA naphthalene acetic acid; BAP benzyl aminopurine
206 Plant Mol Biol Rep (2012) 30:204–213
Author's personal copy

0.2 mL cm−2 pre-hybridization solution. Probing was donewith biotin labeled Xa21 (879 bp) fragments using theBiotin Deca- Label DNA Labeling kit (Fermentas, Ger-many) for labeling purified DNA products of specificgenes, following the manufacturer's instructions.
Stable Transformation Frequency and Data Analyses
The systematic optimization of various parameters enabledus to obtain putative transgenic plants. Transformation
frequency was defined as the percentage of regeneratedcalli which were PCR and Southern blot positive.
Xoo Inoculation
Transgenic T1 plants were grown in Plant BiotechnologyProgramme containment greenhouse at 29°C and 85%humidity. At the maximum tillering stage, all T1 plantswere inoculated with Philippine strain race 6 (PXo55) byscissor dip method (Song et al. 1995). Strain was revived
Fig. 2 a, b, c, d Response ofcalli of rice (Oryza sativa L. cv.Basmati 2000) at different con-centrations of hyg (25, 35, 50,75 mg L−1). Calli on selectionmedium stained with X-Glucsolution. e GUS expression ofthe calli 2 weeks after selection.f Leaf section of To plant (mi-croscopic view). g To leaf. Calliproliferation with different con-centrations of As on selectionmedium h 100 μM, i 200 μM,and j 400 μM
Fig. 3 a Response of 21-day-old calli 7 days after culturingon different hyg containing me-dia. White, gray, and black barshowing number of calli showsbrowning, necrosis, and growth,respectively. b Effect of differentages of calli on GUS expression.c Effect of co-cultivation periodwith A. tumefaciens strainEHA101 (pTCL5) on GUS ex-pression in rice. d Effect ofdifferent doses of As on GUSexpression
Plant Mol Biol Rep (2012) 30:204–213 207
Author's personal copy
on WAM medium and isolated colony was picked formaking dilution (Fig. 7 a, b). Top two to three expandedleaves of each tiller were inoculated; lesion length wasmeasured from inoculated leaves. Plant reaction to PXo55was scored 14 days after inoculation (DAI).
Results
Influence of Various Parameters on Transformation
The aim of present study was to standardize transformationprotocol for enhanced resistance of Basmati 2000 to BB.Conditions such as suitable starting material, age of callus, hygconcentration, antibiotics selection, and As concentration inco-cultivation were optimized for transformation. Four con-
centrations of hyg were tested to determine the selection lethaldose (25, 35, 50, and 75 mg L−1). At 25 mg L−1, 26% calliturned brown and 60% calli exhibited growth (Fig. 3 a).While at 50 mg L−1, 65% calli turned brown, 35% undergoesnecrosis and callus growth was not observed (Figs. 2 a, b, c,d, and 3a). At higher hyg concentration (75 mg L−1), twothirds calli (75%) turned black without growth. hyg(50 mg L−1) was optimized as lethal dose; as with this dose,no callus growth was observed.
Co-cultivation period was tested for 1, 2, 3, and 4 days,keeping other factors constant (pH 5.8 and As (50 μM);Fig. 3 c). Seven days of growth following co-cultivation onselection medium, each callus was incubated in X-Glucsolution and the number of foci showing GUS stainingactivity (GUS+) was counted. The highest GUS expression(73%) was observed when co-cultivation period was keptfor 2 days with normal bacterial growth. GUS expressionwas decreased to 30% when co-cultivation period wasincreased to 3 days with excessive bacterial growth. Thelowest GUS percentage (10%) and minimum bacterialgrowth was observed with 1 day of co-cultivation. GUSexpression was not observed in cultures co-cultivated for4 days (Fig. 3 c). Scutellum-derived calli (18, 21, 25, 28,32, and 35 day old) were co-cultivated for 2 days withbacterial strain. Ten co-cultured calli were incubated withGUS substrate after washing and the remaining weretransferred to selection media (Table 1). The highest GUSexpression (80%) was observed from 21-day-old callifollowed by 25-day-old calli (60%). No GUS expressionwas observed with 35-day-old calli (Fig. 3 b).
Rice calli immersed in Agrobacterium suspension(EHA101, Agrobacterium concentration of OD600=0.04)were co-cultured on three pieces of filter paper moistenedwith 3–4 mL of liquid co-cultivation medium with differentAs concentrations at 25°C for 2 days in the dark (Table 1).When As was omitted from co-cultivation media, no GUSexpression was observed. Maximum GUS expression(70%) was observed with 300 and 400 μM As (Figs. 3dand 2e, f). Different doses of cefotaxime/carbenecillin (Cb/Cf) were optimized to control bacteria and maximize callus
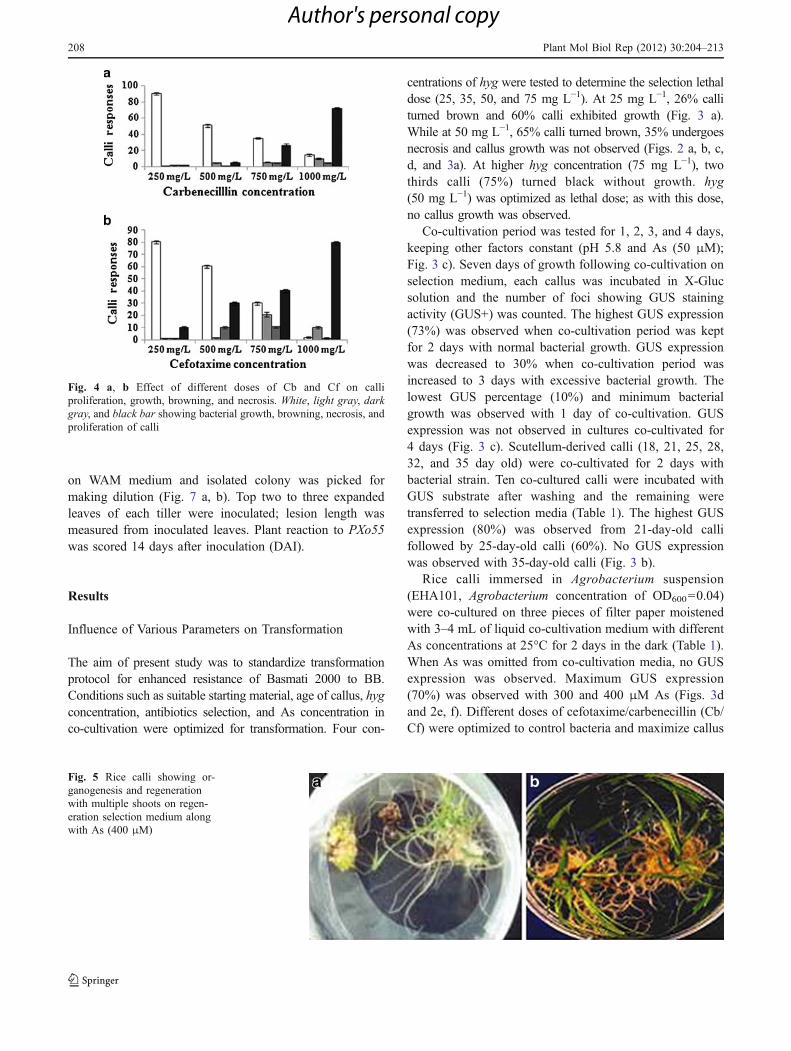
Fig. 4 a, b Effect of different doses of Cb and Cf on calliproliferation, growth, browning, and necrosis. White, light gray, darkgray, and black bar showing bacterial growth, browning, necrosis, andproliferation of calli
Fig. 5 Rice calli showing or-ganogenesis and regenerationwith multiple shoots on regen-eration selection medium alongwith As (400 μM)
208 Plant Mol Biol Rep (2012) 30:204–213
Author's personal copy

growth (250, 500, 750, and 1,000 mg L−1) (Fig. 4 a, b). At250 and 500 mg L−1, bacterial growth was not controlled.At 750 mg L−1, 35% bacterial growth with Cb and 30%with Cf was observed (Fig. 4 a, b). With 1,000 mg L−1, Cfbacterial was growth controlled completely, with 80% calligrowth and proliferation. With 1,000 mg L−1, Cb 15%bacterial growth and 70% calli proliferation was observed(Fig. 4 a, b). Cf (1,000 mg L−1) was selected fortransformation experiments (Fig. 5).
Transformation efficiency is expressed as number of hygresistant and GUS-positive calli/total number of calli inocu-lated (Table 2). A 19.3% regeneration frequency was observedin the average of three experiments (Table 2). Afterregeneration, different parts of transgenic plants were histo-chemically tested for GUS (Fig. 2f, g). A total of 45 T0 plantswere obtained, grown in a glasshouse to produce seeds. All ofthem were normal in development and morphology, withflowers and fertile seeds. The transgenic plants were assessedfor their growth by the following parameters, i.e., plant height,length of panicle, and number of seeds per panicle (Table 3).Transformed rice plants displayed slight decrease in plantheight and panicle length compared with control plants ofBasmati 2000 grown from seeds.
Analysis of Transgenic Plants
Integration of T-DNA, in putative transgenic plants wasconfirmed by PCR and Southern blot analyses. Fourindependent, hyg resistant transgenic rice plants of the
Basmati 2000 were randomly selected for PCR analysis.Plasmid DNA (pTCL5) was used as a positive control. PCRanalysis was conducted while using the primer sets of theXa21 gene, and it showed the expected band of 1.4 kb forXa21gene in all four plants. No amplified band wasdetected in non-transgenic plants (Fig. 6a). Although PCRanalysis is a convenient method for the initial screening ofputative transgenic plants, it does not prove the stableintegration of transgenes because detection by PCR canreflect the survival of A. tumefaciens in host plant tissue.Therefore, when the transgenic plants were 20–30 cm inheight, Southern blot analysis was carried out on genomicDNA (isolated from PCR-positive transgenic plants andnon-transformed control plants) to provide informationrelated to pattern of Xa21 integration. Genomic DNA fromsix randomly selected PCR-positive transgenic lines ofBasmati 2000 were digested with Pst I, which has a uniquerestriction site at the 5′ end of the integrated sequence ofXa21, and hybridized with a transgene (Xa21) specificprobe. Results confirmed the presence of one to two copiesof the transgene per genome in T0 lines (Fig. 6 b).
Inheritance of the Xa21 gene in selfed T1 generation wasanalyzed in term of inoculation studies (bacterial leaf blightresistance). Disease symptoms first appeared 3 DAI inBasmati rice in control plants. The initial symptoms wereleaf curling near the cut-off portion; 21 DAI vascularbundles in the meristem region got filled with bacteria.Through artificial inoculation, it was observed that the levelof resistance of the transgenic plants to PXo45 race 4 washigher than control plants of Basmati 2000 (Table 3, Fig. 7
Table 2 Regeneration frequency of transgenic rice Oryza sativa L Basmati 2000
Sample number Number of callico-cultivated
Number ofcalli selected
Number ofcalli proliferating
Green spotformation
Plantletformation
PCR and Southernblot positive
TE%
Experiment no. 1 150 72 40 32 36 36 24
Experiment no. 2 200 77 43 33 30 30 15
Experiment no. 3 200 85 52 41 38 38 19
Average 183.3 78 45 35.3 34.6 34.6 19.3
TE transformation efficiency, PCR polymerase chain reaction
Table 3 Response of transgenic and control plants to strain PX055 15 days after inoculation & glasshouse assessment of plants
T1 seeds Height (cm) Length of panicle (cm) No of grains per panicle Lesion length (cm) Host response
Control Transgenic Control Transgenic Control Transgenic Control Transgenic Control Transgenic
T10-2 133 130 18 16 105 88 70 55 S MS
T11-3 134 100 18 17 120 90 89 15 S R
T21-5 128 122 14 12 108 100 90 25 S R
Plant Mol Biol Rep (2012) 30:204–213 209
Author's personal copy

c). Lesion length of all transgenic plants was less thancontrol plants. Segregation ratio of 3:1 (resistant: suscepti-ble) was observed in inoculation analysis (Table 4).
Discussion
Xa21 gene cloned in rice through genetic engineering hadbeen shown to confer resistance to over 30 distinct strainsof Xoo for BB resistance (Song et al. 1995; Wang et al.1996; Hao et al. 2009; Park et al. 2010a, b, c). Broad-termresistance caused by Xa21 gene had opened the plethora fordicot in addition to monocots for bacterial diseases (Afrozet al. 2011; Mendes et al. 2009; Park et al. 2010a, b; Peng etal. 2008; Wang et al. 1996). The objective of the presentstudy was to investigate the role of Xa21 in developing BBstress tolerance in Basmati 2000 via Agrobacterium-mediated transformation.
Parameters for inoculation and co-cultivation of Agro-bacterium, including concentration of Agrobacterium, days
to co-cultivation, and hyg and As concentration in themedia were optimized. hyg (50 mg L−1) was optimized aslethal dose used to select transformed calli from non-transformed, extensively used in rice transformation (Shrawatand Good 2010; Ozawa 2009; Rajesh et al. 2008). Severalnew native plant genes have been implemented included theanthranilate synthase alpha-subunit gene, mannose selection,and GFP reporter gene (Todd and Tague 2001; Hashizume etal. 1999). These selection systems resulted in lowselection efficiency of transformed cells as compared toantibiotic resistance genes. Pre-culturing of calli in pre-selection following selection was found to be effective inimprovement of transformation efficiency (Guo et al.2011). Agrobacterium concentration (OD600=0.04), 21-day-old calli and 1000 mg L−1 Cf was optimized for transforma-tion (Ratnayake and Hettiarachchi 2010). Young plant portionsare more suitable targets for transformation as they areactively dividing (Toki et al. 2006). Two days of co-cultivation was optimized, while co-cultivation which lastedfor 3 days for maximum GUS activity is reported before(Ratnayake and Hettiarachchi 2010).
Paper bridges support medium was used for cell growth andproliferation of Agrobacterium was controlled by adjusting theamount of medium (4 mL AA medium) on the paper filter(Tables 1 and 2), whereas conditions using 5.0 and 3.0 mLmedia were not different from those using solid media alone(Howe et al. 2006; Ozawa 2009). Co-cultivation of explantswith Agrobacterium in the presence of As had been reportedin rice (Guo et al. 2011; Wagiran et al. 2010; Ozawa 2009).Signal molecules diffused well in medium containing 4 mL ofliquid AA medium along with 400 μM As. Averagetransformation of 19.3% was observed. Guo et al. (2011)reported drying of mature embryo-derived calli following co-cultivation was effective in higher transformation efficiency. Inthis experiment, also drying seed-derived calli was effective incontrolling bacterial growth following co-cultivation. A totalof 45 To plants obtained were grown in a glasshouse. All werenormal in development and morphology, with flowers andfertile seeds. Transformed rice plants displayed decrease inplant height and panicle length compared with control plantsof Basmati 2000 grown from seeds. A 1.4 kb band of Xa21was observed from the transgenic T0 plants. Baisakh et al.(2000) reported presence of 1.4-kb fragment in some Xa21-resistant transgenic rice plants by using the same primersequence. Southern blot analysis confirmed one to two copiesof gene per transformed rice genome. In general, direct genetransfer methods resulted in multiple insertions and rearrangedfragments in transgenic plants in comparison to Agrobacte-rium-mediated transformation (Krasnyanski et al. 1999).Transgenic plants with multiple copies of the integratedDNA, at one or more chromosomal locations, have beenshown to be more likely to exhibit transgene “silencing”affecting the level and stability of gene expression (Tang et al.
Fig. 6 Molecular analysis of transgenic and non-transgenic rice plantsa PCR amplification of the 1.4-kb fragment corresponding to theCaMV 35S promoter and the nested fragment of the insert in thetransgenic plants (see Fig. 1); lane M molecular ladder, lane (+tive)pTCL5, lane (−tive) non-transgenic plant, lanes 1–4 transgenic plants.b Southern blot analysis of genomic DNA from six PCR-positive Torice plants. Fifteen micrograms of genomic DNA per lane isolatedfrom the leaves of transgenic and non-transgenic rice plants anddigested with PstI. Digested DNA was separated on 0.8% agarose,transferred to membrane and hybridized to a biotin-labeled probeXa21 (879 bp); lanes 2–7 transformed, lane 1 positive control(plasmid pTCL5)
210 Plant Mol Biol Rep (2012) 30:204–213
Author's personal copy

2007). OsHRL (Oryzae sativa HR-like lesion inducing) geneis reported for improvement of BB resistance in rice recently,which is enhanced in response to bacterial infection (Park etal. 2010c). The artificial inoculation of the T1 plants(expressing Xa21 gene) with PXo45 gave the higher level ofresistance than the control plants and mostly has single copyof gene. Mendelian segregation ratio (3:1) was observed inmost of the progeny of the T0 plants. There are not too manyreports for the inheritance studies for the bacterial blight aftertransformation. Most of the reports discussed about theoptimization of the transformation protocols. Lin et al.(2009) reported in planta transformation and did not succeed
in getting the 3:1 ratio for introduced bar gene for Bastaresistance in indica rice. PCR analysis demonstrated that bargene was integrated in to the rice genome DNA. Shrawat andGood (2010) and Saika and Toki (2010) obtained Mendeliansegregation in T1 rice plants with alanine aminotransferaseand luciferase gene. Inoculation tests displayed that transgenicPCR-positive T1 plants were highly resistant to bacterial blightdisease. Since inactivation of the transgene Xa21 in thetransgenic T1 generation occurs at a lower rate, a largerprogeny was obtained for selecting a good homozygous linewith a consistently higher level of resistance to the BBpathogen.
Fig. 7 a Pure culture of Xan-thomonas oryzae pv. Oryzae(PXo55). b 48 h old culture ofPXo55 on the slants used forinoculum preparation. c BB re-action of Basmati 2000 toPXo55. Susceptibility/resistanceof transgenic (T1) lines and thecontrol lines of Basmati 2000after PX055 inoculation. Trans-genic plants showed very littleor no symptoms 2 weeks afterinoculation (c 1–13) and controllines (c 14–15)
Test lines of rice Number of T1 seedling Resistant Susceptible Chi square (χ2) P value
2 12 9 3 0 1.65
4 17 12 5 0.04 1.58
6 8 6 2 0 1.64
7 13 9 4 0.05 1.56
11 12 9 3 0 1.64
15 10 8 2 0.03 1.59
21 24 15 9 0.5 1.28
27 20 16 4 0.06 1.55
32 16 12 4 0 1.64
39 17 11 6 0.24 1.41
Table 4 Levels of resistance/susceptibility/resistance of T1
transgenic plants of Basmati2000 lines observed after PXo45inoculation (expected ratio 3:1)
3:1 ratio indicates integration oftransgene at single locus
Plant Mol Biol Rep (2012) 30:204–213 211
Author's personal copy

References
Adhikari TB, Mew TW, Leach JE (1999) Genotypic and pathotypicdiversity in Xanthomonas oryzae pv. oryzae in Nepal. Phytopa-thology 89:687–694
Afroz A, Chaudhry Z, Rashid U, Muhammad Ali G, Nazir F, Iqbal J,Khan MR (2011) Enhanced resistance against bacterial wilt intransgenic tomato (Lycopersicon esculentum) lines expressing theXa21 gene. Plant Cell Tissue Organ Cult 104:227–237
Baisakh N, Datta K, Rashid H, Oliva N, Datta SK (2000) Agro-bacterium tumefaciens-mediated transformation of an elite indicarice maintainer line IR68899B with a reconstructed T-DNAcarrying multiple genes. Rice Genet Newslett 17:122–125
Birch RG (1997) Plant transformation: problems and strategies forpractical application. Annu Rev Plant Physiol Plant Mol Biol48:297–326
Chan M-T, Lee T-M, Chang H-H (1992) Transformation of indica rice(Oryza sativa L.) mediated by Agrobacterium tumefaciens. PlantCell Physiol 33:577–583
Chu CC, Wang CS, Sun CC, Hsu C, Yin KC, Chu CY (1975)Establishment of an efficient medium for anther culture of ricethrough comparative experiments on the nitrogen sources.Scientia Sinica Sci Sinica 18:659–668
Guo G, Yu J, Zhao D (2011) Rapid acquirement of transgenic riceplants derived from callus of mature embryos transformed byAgrobacterium mediation. Mol Plant Breed 2(2):8–13
Hao ZN, Wang J, Wang LP, Tao RX (2009) Influences of the diseaseresistance conferred by the individual transgenes, Pi-d2, Pi-d3and Xa21, on the transgenic rice plants in yield and grain quality.Afr J Biotechnol 8(19):4845–4848
Hashizume F, Tsuchiya T, Ugaki M, Niwa Y, Tachibana N, KowyamaY (1999) Efficient Agrobacterium-mediated transformation andthe usefulness of a synthetic GFP reporter gene in leadingvarieties of japonica rice. Plant Biotech J 16(5):397–401
Hiei Y, Ohta S, Komari T, Kumashiro T (1994) Efficient transforma-tion of rice (Oryza sativa L.) mediated by Agrobacterium andsequence analysis of the boundaries of the T-DNA. Plant J 6:271–282
Howe A, Sato S, Dweikat I, Fromm M, Clemente T (2006) Rapid andreproducible Agrobacterium-mediated transformation of sor-ghum. Plant Cell Rep 25(8):784–791
Jefferson RA (1987) Assaying chimeric genes in plants: the GUS genefusion system. Plant Mol Biol Reptr 5:387–405
Khush GS, Bacalangco E, Ogawa T (1990) A new gene for resistanceto bacterial blight from O. longistaminata. Rice Genet Newslett7:121–122
Kinoshita T (1995) Report of committee on gene symbolization,nomenclature and linkage group. Rice Genet Newslett 12:9–153
Krasnyanski S, May RA, Loskutov A, Ball TM, Sink KC (1999)Transformation of the limonene synthase gene into peppermint(Mentha piperita L.) and preliminary studies on the essential oilprofiles of single transgenic plants. Theor Appl Genet 99:676–682
Lin XH, Zhang DP, Xie YF, Gao HP, Zhang QF (1996) Identifying andmapping a new gene for bacterial blight resistance in rice basedon RFLP marker. Phytopathology 86:1156–1159
Lin J, Zhou B, Yang Y, Mei J, Zhao X, Guo X, Huang X, Tang D, LiuX (2009) Piercing and vacuum infiltration of the mature embryo:a simplified method for Agrobacterium-mediated transformationof indica rice. Plant Cell Rep 28:1065–1074
Mendes BMJ, Cardoso SC, Boscariol-Camargo RL, Cruz RB, MouraoFilho FAA, Bergamin Filho A (2009) Reduction in susceptibilityto Xanthomonas axonopodis pv. citri in transgenic Citrus sinensisexpressing the rice Xa21 gene. Plant Pathol 59(1):68–75
Murashige T, Skoog F (1962) A revised medium for rapid growth andbioassay with tobacco tissue culture. Plant Physiol 15:472–493
Ozawa K (2009) Establishment of a high ef.ciency Agrobacterium-mediated transformation system of rice (Oryza sativa L.). PlantSci 176:522–527
Park C-J, Bart R, Chern M, Canlas PE, Bai W, Ronald PC (2010a)Overexpression of the endoplasmic reticulum chaperoneBiP3 regulatesXa21-mediated innate immunity in rice. PLoS One 5(2):e9262
Park C-J, Lee S-W, Chern M, Sharma R, Canlas PE, Song M-Y, JeonJ-S, Ronald PC (2010b) Ectopic expression of rice Xa21overcomes developmentally controlled resistance to Xanthomo-nas oryzae pv. Oryzae. Plant Sci 179(5):466–471
Park SR,Moon SJ, ShinD,KimMG,HwangD-J, Bae S-C,Kim J-G,Yi BY,Byun MO (2010c) Isolation and characterization of rice OsHRL generelated to bacterial blight resistance. Plant Pathol J 26(4):417–420
Peng Y, Bartley LE, Chen X, Dardick C, Chern M, Ruan R, CanlasPE, Ronald PC (2008) OsWRKY62 is a negative regulator ofbasal and Xa21-mediated defense against Xanthomonas oryzaepv. oryzae in Rice. Mol Plant 1(3):446–458
Rajesh S, Krishnaveni S, Sudhakar D, RaveendranM, Sivakumar P, GnanamR, Manickam A (2008) Agrobacterium-mediated transformation ofindica Rice (Oryza sativa L.), IR64 with mungbean LEA protein genefor water-stress tolerance. Am J Plant Physiol 3:101–110
Rashid H, Yokoi S, Toriyama K, Hinata K (1996) Transgenic plantproduction mediated by Agrobacterium in indica rice. Plant CellRep 15:727–730
Ratnayake RMLK, Hettiarachchi GHCM (2010) Development of anefficient Agrobacterium mediated transformation protocol forSrilankan rice variety—Bg 250. Tropical Agricultural Research22(1):45–53
Saika H, Toki S (2010) Mature seed-derived callus of the model indicarice variety Kasalath is highly competent in Agrobacterium-mediated transformation. Plant Cell Rep 29:1351–1364
Sambrook J, Russell D (2001) Molecular cloning. A laboratorymanual, 3rd edn. Cold Spring Harbor Laboratory Press, NewYork, pp 6.33–6.62
Shrawat AK, Good AG (2010) A high-throughput Agrobacteriumtumefaciens-mediated transformation system for molecularbreeding and functional genomics of rice (Oryza sativa L.).Plant Biotech 27:47–58
Song WY, Wang GL, Chen LL, Kim HS, Pi LY, Holsten TE, Gardner J,Wang BG, Zhai WX, Zhu LH, Fauquet CM, Ronald PC (1995) Areceptor kinase-like protein encoded by the rice disease resistancegene, Xa21. Science 270:1804–1806
Song W-Y, Pi L-Y, Wang G-L, Gardner J, Holsten T, Ronald PC (1997)Evolution of the Rice Xa2I Disease Resistance Gene Family.Plant Cell 9:1279–1287
Swings J, Van den Mooter M, Vauterin L, Hoste B, Gillis M, Mew TW,Kersters K (1990) Reclassification of the causal agents ofbacterial blight (Xanthomonas campestris pv. oryzae) andbacterial leaf streak (Xanthomonas campestris pv. oryzicola) ofrice as pathovars of Xanthomonas oryzae (ex Ishiyama 1922) sp.nov., nom. Int J Syst Bacteriol 40:309–311
Tada T, Sakamoto M, Fujimura T (1990) Efficient gene introduction intorice by electroporation and analysis of transgenic plants: use ofelectroporation buffer lacking chloride ions. Theor Appl Genet80:475–480
Tang W, Newton RJ, Weidner DA (2007) Genetic transformation andgene silencing mediated by multiple copies of a transgene ineastern white pine. J Exp Bot 58:545–554
Todd R, Tague BW (2001) Phosphomannose Isomerase: A versatileselectable marker for Arabidopsis thaliana germ-line transforma-tion. Plant Mol Biol Reptr 19:307–319
Toki S, Hara N, Onodera H, Tagiri A, Oka S, Tanaka H (2006) Earlyinfection of scutellum tissue with Agrobacterium allows high-speed transformation of rice. Plant J 47:969–976
Toriyama K, Hinata K (1985) Cell suspension and protoplast culture inrice. Plant Sci 41:179–183
212 Plant Mol Biol Rep (2012) 30:204–213
Author's personal copy

Uchimiya H, Fuskimi T, Hashimoto H, Harda H, Syono K, Sugawara Y(1986) Expression of a foreign gene in callus derived from DNA-treated protoplasts of rice (Oryza sativa L.). Mol Gen Genet16:204–207
Wagiran A, Ismail I, Radziah C, Zain CM, Abdullah R (2010)Agrobacterium tumefaciens-mediated transformation of the iso-pentenyltransferase gene in japonica rice suspension cell culture.Aus J Crop Sci 4(6):421–429
Wang GL, Song WY, Ruan DL, Sideris S, Ronald P (1996) The clonedgene, Xa21, confers resistance to multiple Xanthomonas oryzae pv.oryzae isolates in transgenic plants. Mol Plant Microbe Interact9:850–855
Zhai W, Wang W, Zhou Y, Li X, Zheng X, Zhang Q, Wang G, Zhu L(2001) Breeding bacterial blight-resistant hybrid rice with thecloned bacterial blight resistance gene Xa21. Mol Breeding8:285–293
Plant Mol Biol Rep (2012) 30:204–213 213
Author's personal copy
Copyright © 2022 FDOKUMEN