
Trafficking of HIV-1 RNA is Mediated by Heterogeneous Nuclear Ribonucleoprotein A2 Expression and...
17
# 2006 The Authors Journal compilation # 2006 Blackwell Publishing Ltd doi: 10.1111/j.1600-0854.2006.00461.x Traffic 2006; 7: 1177–1193 Blackwell Munksgaard Trafficking of HIV-1 RNA is Mediated by Heterogeneous Nuclear Ribonucleoprotein A2 Expression and Impacts on Viral Assembly Kathy Le ´ vesque 1,2,† , Melanie Halvorsen 1,2,† , Levon Abrahamyan 1,2 , Laurent Chatel- Chaix 1,2,3 , Viviane Poupon 1,2,4 , Heather Gordon 1,2,6 , Luc DesGroseillers 3 , Anne Gatignol 2,5,6 and Andrew J. Mouland 1,2,5,6,* 1 HIV-1 RNA Trafficking Laboratory, 3755 Co ˆ te-Ste- Catherine Road, Montre ´ al, Que ´ bec, Canada H3T 1E2 2 Lady Davis Institute for Medical Research-Sir Mortimer B. Davis Jewish General Hospital, 3755 Co ˆ te-Ste- Catherine Road, Montre ´ al, Que ´ bec, Canada H3T 1E2 3 Department of Biochemistry, Universite ´ de Montre ´ al, Montre ´ al, Que ´ bec, Canada H3C 1G4 4 Current address: Montreal Neurological Institute, McGill University, 3801 University Street, Montre ´ al, Que ´ bec, Canada H3A 2B4 5 Department of Medicine, McGill University, Montre ´ al, Que ´ bec, Canada H3A 2B4 6 Department of Microbiology & Immunology, McGill University, Montre ´ al, Que ´ bec, Canada H3A 2B4 *Corresponding author: Andrew J. Mouland, [email protected] † These authors contributed equally to this work. Few details are known about how the human immuno- deficiency virus type 1 (HIV-1) genomic RNA is trafficked in the cytoplasm. Part of this process is controlled by the activity of heterogeneous nuclear ribonucleoprotein A2 (hnRNP A2). The role of hnRNP A2 during the expression of a bona fide provirus in HeLa cells is investigated in this study. Using immunofluorescence and fluorescence in situ hybridization techniques, we show that knock- down of hnRNP A2 expression in HIV-1-expressing cells results in the rapid accumulation of HIV-1 genomic RNA in a distinct, cytoplasmic space that corresponds to the microtubule-organizing center (MTOC). The RNA exits in the nucleus and accumulates at the MTOC region as a result of hnRNP A2 knockdown even during the expres- sion of a provirus harboring mutations in the hnRNP A2- response element (A2RE), the expression of which results in nuclear retention of genomic RNA. We also demon- strate that hnRNP A2 expression is required for down- stream trafficking of genomic RNA from the MTOC in the cytoplasm. Genomic RNA localization at the MTOC that was both the result of hnRNP A2 knockdown and the overexpression of Rab7-interacting lysosomal protein had little effect on pr55 Gag synthesis but negatively influenced virus production and infectivity. These data indicate that altered HIV-1 genomic RNA localization modulates viral assembly and that the MTOC serves as a central site to which HIV-1 genomic RNA converges following its exit from the nucleus, with the host protein, hnRNP A2, playing a central role in taking it to and from this site in the cell. Key words: AIDS, assembly, HIV-1, hnRNP A2, RNA trafficking, virus–host interaction Received 23 December 2005, revised and accepted for publication 13 June 2006, published on-line 27 July 2006 HIV-1 infection is characterized by a lengthy latent period before the onset of acquired immunodeficiency syndrome (AIDS). During this period, abundant viral production is kept in check by the immune system and cells that are killed by infection are replaced. Despite mounting a strong early immune response, HIV-1 expression progressively de- pletes CD4þ T cells, a situation that leads to a progressive weakening of the immune response to infection and the onset of AIDS (1,2). HIV-1 gene transcription generates a primary 9-kilobase pair (kbp) RNA that has three fates dictated by a tight regulatory circuit and temporal activities of viral proteins. The 9-kbp RNA is multiply spliced follow- ing transcription to generate several 2-kbp RNAs that give rise to regulatory proteins Tat, Rev and Nef. Tat accumu- lates and is primarily responsible for high-level transacti- vation of the integrated HIV-1 provirus. Once a threshold level of Rev is reached, a molecular switch occurs to promote the inhibition of splicing of the primary transcript. The decreased splicing activity also produces singly- spliced RNA species (4-kbp) (3). Rev binds the Rev- responsive cis-acting element RNA (4) to promote the nuclear export of the 9-kbp and singly-spliced 4-kbp HIV-1 RNAs. The 9-kbp RNA is not only a substrate for the translation machinery to generate structural (Gag) and viral enzymes, but in addition, it is selected for encapsidation into new virus particles. The 4-kbp RNAs are translated to produce auxiliary proteins and the viral envelope protein, Env. Without Rev, both of the latter RNA species are retained in the nucleus and are presumably degraded by cellular mechanisms, and as a consequence, viral production is severely impaired. Nuclear export by Rev is also aided by the activities of multiple host cell proteins. Several proteins, including RNA-binding proteins, SR-splicing proteins, RNA helicases and other proteins such as actin, influence HIV-1 Rev activity to export RNA from the nucleus (5–7). Very little is known about HIV-1 RNA trafficking following disengage- ment of Rev in the cytosol. Recent data support the notion that additional viral and host proteins engage to promote 1177
Transcript of Trafficking of HIV-1 RNA is Mediated by Heterogeneous Nuclear Ribonucleoprotein A2 Expression and...

# 2006 The AuthorsJournal compilation # 2006 Blackwell Publishing Ltd
doi: 10.1111/j.1600-0854.2006.00461.x
Traffic 2006; 7: 1177–1193Blackwell Munksgaard
Trafficking of HIV-1 RNA isMediated by HeterogeneousNuclear Ribonucleoprotein A2 Expression and Impactson Viral Assembly
Kathy Levesque1,2,†, Melanie Halvorsen1,2,†,
Levon Abrahamyan1,2, Laurent Chatel-
Chaix1,2,3, Viviane Poupon1,2,4, Heather
Gordon1,2,6, Luc DesGroseillers3, Anne
Gatignol2,5,6 and Andrew J. Mouland1,2,5,6,*
1HIV-1 RNA Trafficking Laboratory, 3755 Cote-Ste-Catherine Road, Montreal, Quebec, Canada H3T 1E22Lady Davis Institute for Medical Research-Sir MortimerB. Davis Jewish General Hospital, 3755 Cote-Ste-Catherine Road, Montreal, Quebec, Canada H3T 1E23Department of Biochemistry, Universite de Montreal,Montreal, Quebec, Canada H3C 1G44Current address: Montreal Neurological Institute, McGillUniversity, 3801 University Street, Montreal, Quebec,Canada H3A 2B45Department of Medicine, McGill University, Montreal,Quebec, Canada H3A 2B46Department of Microbiology & Immunology, McGillUniversity, Montreal, Quebec, Canada H3A 2B4*Corresponding author: Andrew J. Mouland,[email protected]†These authors contributed equally to this work.
Few details are known about how the human immuno-
deficiency virus type 1 (HIV-1) genomic RNA is trafficked
in the cytoplasm. Part of this process is controlled by the
activity of heterogeneous nuclear ribonucleoprotein A2
(hnRNP A2). The role of hnRNP A2 during the expression
of a bona fide provirus in HeLa cells is investigated in this
study. Using immunofluorescence and fluorescence
in situ hybridization techniques, we show that knock-
down of hnRNP A2 expression in HIV-1-expressing cells
results in the rapid accumulation of HIV-1 genomic RNA
in a distinct, cytoplasmic space that corresponds to the
microtubule-organizing center (MTOC). The RNA exits in
the nucleus and accumulates at the MTOC region as
a result of hnRNP A2 knockdown even during the expres-
sion of a provirus harboring mutations in the hnRNP A2-
response element (A2RE), the expression of which results
in nuclear retention of genomic RNA. We also demon-
strate that hnRNP A2 expression is required for down-
stream trafficking of genomic RNA from the MTOC in the
cytoplasm. Genomic RNA localization at the MTOC that
was both the result of hnRNP A2 knockdown and the
overexpression of Rab7-interacting lysosomal protein
had little effect on pr55Gag synthesis but negatively
influenced virus production and infectivity. These data
indicate that altered HIV-1 genomic RNA localization
modulates viral assembly and that the MTOC serves as
a central site to which HIV-1 genomic RNA converges
following its exit from the nucleus, with the host protein,
hnRNP A2, playing a central role in taking it to and from
this site in the cell.
Key words: AIDS, assembly, HIV-1, hnRNP A2, RNA
trafficking, virus–host interaction
Received 23 December 2005, revised and accepted for
publication 13 June 2006, published on-line 27 July 2006
HIV-1 infection is characterized by a lengthy latent period
before the onset of acquired immunodeficiency syndrome
(AIDS). During this period, abundant viral production is kept
in check by the immune system and cells that are killed by
infection are replaced. Despite mounting a strong early
immune response, HIV-1 expression progressively de-
pletes CD4þ T cells, a situation that leads to a progressive
weakening of the immune response to infection and the
onset of AIDS (1,2). HIV-1 gene transcription generates
a primary 9-kilobase pair (kbp) RNA that has three fates
dictated by a tight regulatory circuit and temporal activities
of viral proteins. The 9-kbp RNA is multiply spliced follow-
ing transcription to generate several 2-kbp RNAs that give
rise to regulatory proteins Tat, Rev and Nef. Tat accumu-
lates and is primarily responsible for high-level transacti-
vation of the integrated HIV-1 provirus. Once a threshold
level of Rev is reached, a molecular switch occurs to
promote the inhibition of splicing of the primary transcript.
The decreased splicing activity also produces singly-
spliced RNA species (4-kbp) (3). Rev binds the Rev-
responsive cis-acting element RNA (4) to promote the nuclear
export of the 9-kbp and singly-spliced 4-kbp HIV-1 RNAs.
The 9-kbp RNA is not only a substrate for the translation
machinery to generate structural (Gag) and viral enzymes,
but in addition, it is selected for encapsidation into new virus
particles. The 4-kbp RNAs are translated to produce auxiliary
proteins and the viral envelope protein, Env. Without Rev,
both of the latter RNA species are retained in the nucleus
and are presumably degraded by cellular mechanisms, and
as a consequence, viral production is severely impaired.
Nuclear export by Rev is also aided by the activities of
multiple host cell proteins. Several proteins, including
RNA-binding proteins, SR-splicing proteins, RNA helicases
and other proteins such as actin, influence HIV-1 Rev
activity to export RNA from the nucleus (5–7). Very little
is known about HIV-1 RNA trafficking following disengage-
ment of Rev in the cytosol. Recent data support the notion
that additional viral and host proteins engage to promote
1177

trafficking to sites of translation or assembly, identifying
roles for the cellular proteins, Sam68 and the human Rev-
interacting protein (hRIP) in these processes (5,8,9). Viral
Gag and motor proteins like KIF-4 also appear to play a role,
but the mechanisms of action remain uncharacterized
(10,11). We implicated heterogeneous nuclear ribonucleo-
protein A2 (hnRNP A2) in this step in a previous study in
which we identified HIV-1 RNA trafficking sequences in
several HIV-1 RNAs (known as the hnRNP A2-responsive
elements or A2RE) (12). We also showed that the interac-
tion of hnRNP A2 with the HIV-1 RNA via these sequences
was important for HIV-1 RNA trafficking from the nucleus,
whereby a single point mutation in the A2RE resulted in
nuclear sequestration of genomic RNA. Blocking this inter-
action also impacted on HIV-1 RNA, Gag and Vpr expression
patterns and the packaging of viral genomic RNA into virions
(13). Because RNA compartmentalization in the nucleus
only occurred late in the replication cycle, comparatively
normal levels of intracellular Gag levels accumulated. This
work suggested that hnRNP A2 acts at a specific time that
coincides with a late stage of the replication cycle.
In this study, we identify the roles of hnRNP A2 in the fate
of HIV-1 genomic RNA from the nucleus and into the
cytosol. We used small interfering (si) RNA to knockdown
hnRNP A2 gene expression in HIV-1-expressing HeLa
cells. Near-complete knockdown of hnRNP A2 had a sig-
nificant impact on HIV-1 RNA localization. The phenotype
was characterized by an accumulation of HIV-1 genomic
RNA at the microtubule-organizing center (MTOC), as
identified by fluorescence in situ hybridization (FISH) and
immunofluorescence (IF) analyses. This was also ob-
served in the context of the expression of an A2RE mutant
provirus that normally results in nuclear sequestration of
genomic RNA due to loss of association of hnRNP A2 to
the RNA. These new data reveal a nuclear retention
function for hnRNP A2. Kinetic studies and studies that
follow the fate of cytoplasm-localized HIV-1 RNA reveal that
the accumulation of RNA occurs very rapidly at the MTOC
and that HIV-1 genomic RNA egress within the cytoplasm
is dependent on hnRNP A2 expression. Gag expression
was not affected by hnRNP A2 knockdown and its traffick-
ing can be physically separated from that of the genomic
RNA by promoting the recruitment of dynein/dynactin
complex to the MTOC. This work represents the first study
to demonstrate that the fate of the HIV-1 genomic RNA
during proviral gene expression is mediated directly by
expression levels of the host cell protein, hnRNP A2.
Results
Efficient knockdown of hnRNP A2 expression is
achieved and has little effect on Gag synthesis or
HIV-1 RNA splicing
HeLa cells have been used in studies on viral RNA egress
(14,15), and successful depletion of heterogeneous
nuclear ribonucleoproteins (hnRNPs) by siRNA has
recently been documented in this cell line (16). To study
the involvement of hnRNP A2 in HIV-1 RNA trafficking, we
used 21-bp siRNA duplexes to specifically target its
expression. Cells were first transfected with siRNA du-
plexes and then cotransfected with proviral DNA, HxBRU
and the same siRNA duplexes 24 h later. The siRNA
duplexes were designed to target hnRNP A2 (siA2) and
hnRNP A1 (siA1). A nonsilencing siRNA (siNS) was also
included. At 30 h post-HxBRU transfection, protein expres-
sion was assessed byWestern analyses. To determine the
efficacy of the siRNA-mediated knockdown, hnRNPs were
identified using a pan-specific hnRNP polyclonal antibody
that recognizes predominantly hnRNP A1 and hnRNP A2
(16). Glyceraldehyde-3-phosphate dehydrogenase
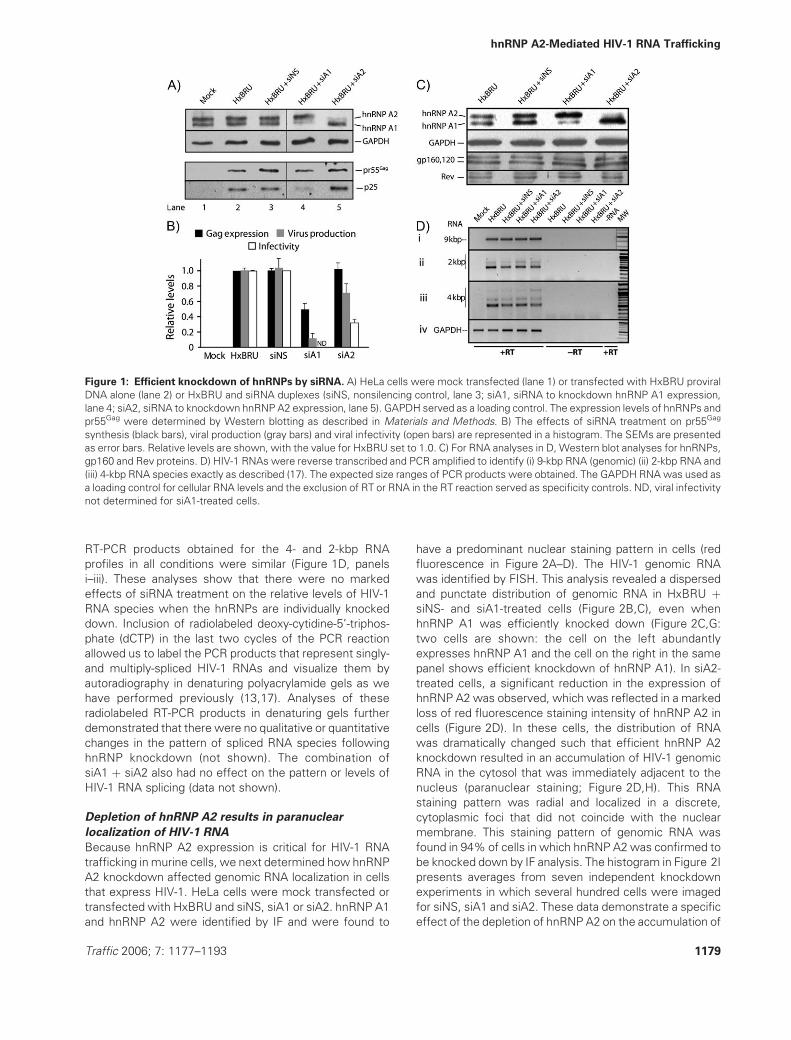
(GAPDH) was used as a loading control (Figure 1A).
Whereas in mock, HxBRU or HxBRU þ siNS-transfected
cells, hnRNP A2 and A1 expression levels were constant
(Figure 1A, lanes 1–3); cells transfected with siA1 resulted
in an 85% reduction in hnRNP A1 expression (Figure 1A,
lane 4). When hnRNP A2 was targeted, it was also
specifically knocked down by 90% (lane 5). The siA2
treatment did not have an impact on the 55-kDa precursor
Gag (pr55Gag) synthesis in cells, whereas siA1 had a signifi-
cant negative impact on pr55Gag expression and viral
production when this siRNA was used alone or in combi-
nation with siA2 (Figure 1A, lane 4; data not shown). Viral
production levels were reduced by 29% (�11%, SEM) in
siA2-treated cells, and furthermore, a striking decrease in
infectivity was found in siA2-treated cells (65 � 6%, SEM;
Figure 1B). The results shown in Figure 1B represent
averages from 10 experiments each (pr55Gag synthesis
and viral production) and four experiments (viral infectivity).
There were no general effects on cell cycle distribution
when hnRNP A1 and hnRNP A2 were individually knocked
down [not shown; (16)].
Because hnRNPs have been shown to be involved in RNA
splicing in vitro, we therefore performed RNA splicing
analyses to determine if the knockdown of hnRNPs had
effects at this level. Cells were mock transfected or
transfected with HxBRU alone, HxBRU and siNS, and
siA2 or siA1. The knockdown was verified by Western
blot analyses (Figure 1C) and compared with the loading
control, GAPDH. Representative HIV-1 proteins encoded
by 4- and 2-kbp RNAs were also quantitated by Western
analyses, and this analysis showed that encoded proteins
gp160 and Rev levels were unaffected, indicating that the
expression of their RNAs was not affected in knockdown
conditions (Figure 1C). The RNA was isolated from cells
and first assessed by Northern analyses. There were no
differences between the relative levels of 9-, 4- and 2-kbp
RNAs in siNS and siRNA targeting hnRNP expression (data
not shown). In other experiments, RNA was subjected to
reverse transcription–polymerase chain reaction (RT-PCR)
to amplify 9-, 4- and 2-kbp HIV-1 RNA species using
specific sets of oligonucleotides as described in Materials
and Methods. The relative levels of genomic 9-kbp RNA
were virtually constant and the quantity and profiles of the
1178 Traffic 2006; 7: 1177–1193
Levesque et al.
RT-PCR products obtained for the 4- and 2-kbp RNA
profiles in all conditions were similar (Figure 1D, panels
i–iii). These analyses show that there were no marked
effects of siRNA treatment on the relative levels of HIV-1
RNA species when the hnRNPs are individually knocked
down. Inclusion of radiolabeled deoxy-cytidine-5’-triphos-
phate (dCTP) in the last two cycles of the PCR reaction
allowed us to label the PCR products that represent singly-
and multiply-spliced HIV-1 RNAs and visualize them by
autoradiography in denaturing polyacrylamide gels as we
have performed previously (13,17). Analyses of these
radiolabeled RT-PCR products in denaturing gels further
demonstrated that there were no qualitative or quantitative
changes in the pattern of spliced RNA species following
hnRNP knockdown (not shown). The combination of
siA1 þ siA2 also had no effect on the pattern or levels of
HIV-1 RNA splicing (data not shown).
Depletion of hnRNP A2 results in paranuclear
localization of HIV-1 RNA
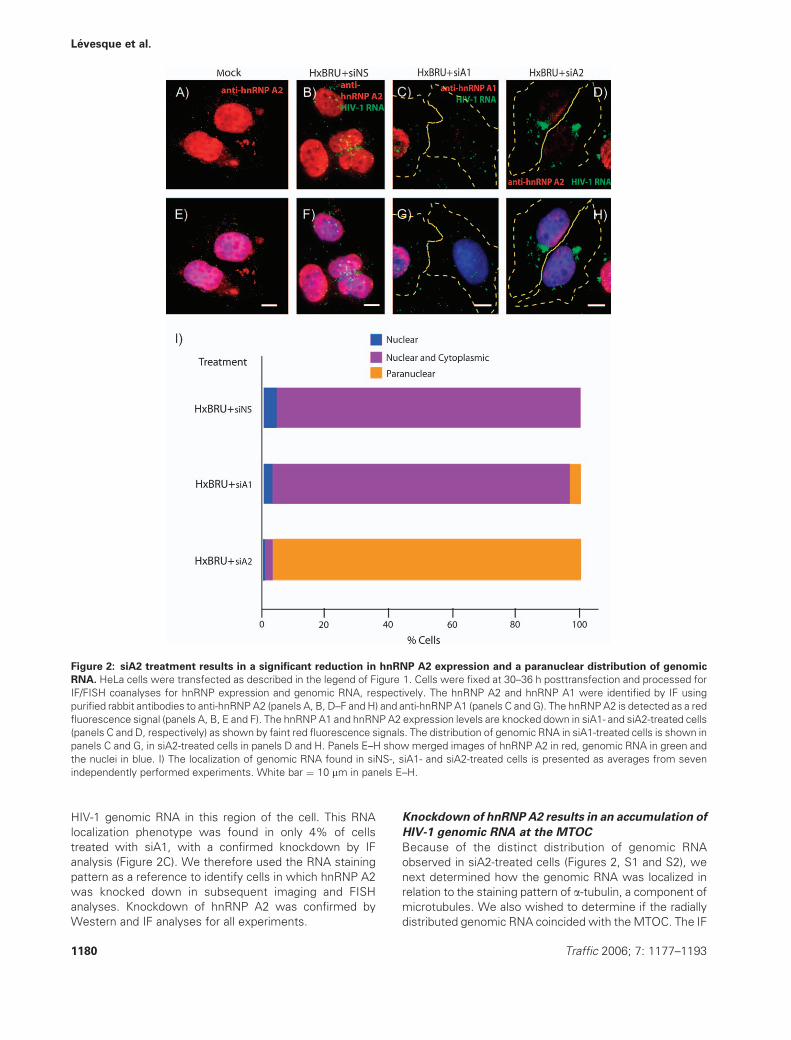
Because hnRNP A2 expression is critical for HIV-1 RNA
trafficking in murine cells, we next determined how hnRNP
A2 knockdown affected genomic RNA localization in cells
that express HIV-1. HeLa cells were mock transfected or
transfected with HxBRU and siNS, siA1 or siA2. hnRNP A1
and hnRNP A2 were identified by IF and were found to
have a predominant nuclear staining pattern in cells (red
fluorescence in Figure 2A–D). The HIV-1 genomic RNA
was identified by FISH. This analysis revealed a dispersed
and punctate distribution of genomic RNA in HxBRU þsiNS- and siA1-treated cells (Figure 2B,C), even when
hnRNP A1 was efficiently knocked down (Figure 2C,G:
two cells are shown: the cell on the left abundantly
expresses hnRNP A1 and the cell on the right in the same
panel shows efficient knockdown of hnRNP A1). In siA2-
treated cells, a significant reduction in the expression of
hnRNP A2 was observed, which was reflected in a marked
loss of red fluorescence staining intensity of hnRNP A2 in
cells (Figure 2D). In these cells, the distribution of RNA
was dramatically changed such that efficient hnRNP A2
knockdown resulted in an accumulation of HIV-1 genomic
RNA in the cytosol that was immediately adjacent to the
nucleus (paranuclear staining; Figure 2D,H). This RNA
staining pattern was radial and localized in a discrete,
cytoplasmic foci that did not coincide with the nuclear
membrane. This staining pattern of genomic RNA was
found in 94% of cells in which hnRNP A2was confirmed to
be knocked down by IF analysis. The histogram in Figure 2I
presents averages from seven independent knockdown
experiments in which several hundred cells were imaged
for siNS, siA1 and siA2. These data demonstrate a specific
effect of the depletion of hnRNP A2 on the accumulation of
Figure 1: Efficient knockdown of hnRNPs by siRNA. A) HeLa cells were mock transfected (lane 1) or transfected with HxBRU proviral
DNA alone (lane 2) or HxBRU and siRNA duplexes (siNS, nonsilencing control, lane 3; siA1, siRNA to knockdown hnRNP A1 expression,
lane 4; siA2, siRNA to knockdown hnRNP A2 expression, lane 5). GAPDH served as a loading control. The expression levels of hnRNPs and
pr55Gag were determined by Western blotting as described in Materials and Methods. B) The effects of siRNA treatment on pr55Gag
synthesis (black bars), viral production (gray bars) and viral infectivity (open bars) are represented in a histogram. The SEMs are presented
as error bars. Relative levels are shown, with the value for HxBRU set to 1.0. C) For RNA analyses in D, Western blot analyses for hnRNPs,
gp160 and Rev proteins. D) HIV-1 RNAs were reverse transcribed and PCR amplified to identify (i) 9-kbp RNA (genomic) (ii) 2-kbp RNA and
(iii) 4-kbp RNA species exactly as described (17). The expected size ranges of PCR products were obtained. The GAPDH RNAwas used as
a loading control for cellular RNA levels and the exclusion of RT or RNA in the RT reaction served as specificity controls. ND, viral infectivity
not determined for siA1-treated cells.
Traffic 2006; 7: 1177–1193 1179
hnRNP A2-Mediated HIV-1 RNA Trafficking
HIV-1 genomic RNA in this region of the cell. This RNA
localization phenotype was found in only 4% of cells
treated with siA1, with a confirmed knockdown by IF
analysis (Figure 2C). We therefore used the RNA staining
pattern as a reference to identify cells in which hnRNP A2
was knocked down in subsequent imaging and FISH
analyses. Knockdown of hnRNP A2 was confirmed by
Western and IF analyses for all experiments.
Knockdown of hnRNP A2 results in an accumulation of
HIV-1 genomic RNA at the MTOC
Because of the distinct distribution of genomic RNA
observed in siA2-treated cells (Figures 2, S1 and S2), we
next determined how the genomic RNA was localized in
relation to the staining pattern of a-tubulin, a component of
microtubules. We also wished to determine if the radially
distributed genomic RNA coincided with theMTOC. The IF
Figure 2: siA2 treatment results in a significant reduction in hnRNP A2 expression and a paranuclear distribution of genomic
RNA. HeLa cells were transfected as described in the legend of Figure 1. Cells were fixed at 30–36 h posttransfection and processed for
IF/FISH coanalyses for hnRNP expression and genomic RNA, respectively. The hnRNP A2 and hnRNP A1 were identified by IF using
purified rabbit antibodies to anti-hnRNP A2 (panels A, B, D–F and H) and anti-hnRNPA1 (panels C and G). The hnRNPA2 is detected as a red
fluorescence signal (panels A, B, E and F). The hnRNP A1 and hnRNP A2 expression levels are knocked down in siA1- and siA2-treated cells
(panels C and D, respectively) as shown by faint red fluorescence signals. The distribution of genomic RNA in siA1-treated cells is shown in
panels C and G, in siA2-treated cells in panels D and H. Panels E–H show merged images of hnRNP A2 in red, genomic RNA in green and
the nuclei in blue. I) The localization of genomic RNA found in siNS-, siA1- and siA2-treated cells is presented as averages from seven
independently performed experiments. White bar ¼ 10 mm in panels E–H.
1180 Traffic 2006; 7: 1177–1193
Levesque et al.

and FISH coanalyses were performed to identify the
tubulin-based cytoskeleton and genomic RNA in control
siNS-, siA1- and siA2-treated cells. In siNS-treated cells
and likewise in siA1-treated cells, genomic RNAwas found
to be distributed throughout the cells in a punctate staining
pattern as shown above (Figure 2). In siA2-treated cells,
the genomic RNA staining coincided with intense tubulin
staining in the pericentriolar region (Figure 3A–D). In siA2-
treated cells, genomic RNAwas also observed to reside on
microtubule tracks either emanating from the MTOC or at
distal sites near the periphery of the cell (data not shown).
Dual color analyses and black and white representations of
the IF and FISH images demonstrate that the area at which
the genomic RNA accumulates corresponds to the region
that overlaps the MTOC, a structure from which micro-
tubules polymerize (Figure 3D). A reduction in hnRNP A2
expression, but not that of the related hnRNP A1, results in
an accumulation of genomic RNA at this cellular site,
indicating that the HIV-1 genomic RNA may traffic to and
from the MTOC region during the gene expression stages
of virus replication.
We also explored whether the microtubule-disrupting
agent, nocodazole, altered the localization of genomic
RNA in siA2-treated cells. Cells were briefly treated with
nocodazole for 30 min before fixation to disrupt micro-
tubules but withminimal effects on the localization of other
organelles such as the Golgi (18). In these cells, nocoda-
zole disrupted the microtubule cytoskeleton as shown by
the resultant disaggregation of microtubule streaming in
cells (Figure 3E,H). This treatment resulted in a diffusion of
the genomic RNA from the MTOC region consistent with
a dependence on an intact microtubule cytoskeleton for
the anchoring of the genomic RNA at this region (Figure 3
E–H). To demonstrate this more clearly, hnRNP A2 expres-
sion in cells was identified by IF in mock- and siA2-treated
HxBRU-expressing cells (Figure 3I). hnRNP A2 was abun-
dantly expressed in mock-treated cells (Figure 3I, top
panel). The siA2 treatment resulted in a marked loss of
hnRNP A2 staining and a redistribution of genomic RNA to
the MTOC region (Figure 3I, middle panel). Again, brief
nocodazole treatment resulted in a loss in the MTOC-
localized genomic RNA and this was clearly observed in
hnRNP A2-depleted cells (Figure 3I, bottom panel), show-
ing that anchoring to the MTOC region depends on
polymerized microtubules. However, nocodazole treat-
ment also promotes a diffusion of late endosomes/lyso-
somes from the MTOC region (19), so these results may
also point to these vesicular organelles as sites of accu-
mulation of HIV-1 genomic RNA.
We next confirmed the localization of the genomic RNA by
costaining with an MTOC-resident protein and nucleator of
microtubules, g-tubulin. For this analysis, laser scanning
confocal microscopy was necessary to accurately localize g-tubulin and genomic RNA staining in cells. Similar to the
staining pattern shown in Figure 2, genomic RNA staining
was punctate throughout the cell (one representative cell is
shown in Figure 4, panels A–D). The g-tubulin staining was
discreet and characterized by a distinctive and intense
staining adjacent to the nucleus in most cells (indicated by
arrows in Figure 4). A black and white rendition is shown in
Figure 4C to provide better visualization of the g-tubulin/MTOC staining. A merged image of g-tubulin and HIV-1
genomic RNA in siNS-treated cells is shown in panel D. In
siA2-treated cells, the genomic RNA was predominantly
found in the distinct cytoplasmic region adjacent to the
nucleus (one representative cell is shown in Figure 4E). The
Figure 3: HIV-1 genomic RNA localizes to the MTOC in siA2-treated cells. Cells were transfected with HxBRU and siNS or siA2, and
at 30–36 h posttransfection, the cells were fixed and analyzed for microtubule staining using an anti-a-tubulin anti-serum and the
localization of genomic RNA (by FISH as green fluorescence). A–D) a-tubulin (red) and genomic RNA (green) staining in siA2-treated cells.
The MTOC is highlighted by the inset in panel D. Cells were treated with nocodazole for 30 min prior to fixation (panels E–H) and then
stained for a-tubulin as red fluorescence in all cells in panels E and G or more vividly in black and white in panel H. I) In complementary
experiments, hnRNP A2 expression (red) was analyzed by IF and the HIV-1 RNA by FISH (in green) in mock-transfected cells (top panel) or
cells transfectedwith HxBRUþ siA2 (middle panel), followed by nocodazole treatment (bottom panel) asmentioned above.White bar¼ 10
mm in panels D and H and panels presented in I.
Traffic 2006; 7: 1177–1193 1181
hnRNP A2-Mediated HIV-1 RNA Trafficking

MTOCwas identified by g-tubulin staining in red fluorescence(panel F) and a black and white rendition is shown in panel G.
In siA2-treated HIV-1-expressing cells, laser scanning confo-
cal imaging shows that the genomic RNAwas localized in the
direct vicinity of theMTOC as shown by a radial RNA staining
pattern emanating from the MTOC, identified by the distinct
and central g-tubulin staining in these cells [Figure 4, panel H
(and see inset showing magnified MTOC region)].
HIV-1 RNA accumulation occurs rapidly at the MTOC
A kinetic study was performed to determine how fast the
HIV-1 genomic RNA accumulated at the MTOC in siA2-
treated cells. Cells were transfected with either siNS or
siA2 and then with HxBRU. At 12, 20 and 30 h later, cells
were fixed and stained for hnRNP A2 and genomic RNA. At
the first time-point tested (12 h) in siNS-treated cells, the
genomic RNA was detectable at low levels and hnRNP A2
was abundantly expressed (Figure 5A). At the following
time-points in siNS-treated cells, genomic RNA became
more abundant and was distributed in a punctate staining
pattern (Figure 5B,C). When hnRNP A2 and genomic RNA
were identified by FISH/IF coanalyses in siA2-treated cells
(panels D–I), MTOC-localized genomic RNA was apparent
at 12 h in cells that were efficiently knocked down for
hnRNP A2 (identified by an arrow in Figure 5D). A set of
overexposed images in red, green and blue channels is
presented in order to first delineate the outline of the
nucleus of cells and second, to demonstrate the efficient
knockdown of hnRNP A2 as indicated by the faint red
fluorescence signal for hnRNP A2 in the HIV-1 RNA-
expressing cells shown at all time-points (Figure 5D–F).
The next set of panels represent normal exposed images,
and DAPI staining is included to characterize the distribu-
tion of HIV-1 RNA in relation to the nucleus (Figure 5G–I).
These results demonstrate again that in cells in which
hnRNP A2 expression is significantly knocked down, the
genomic RNA accumulates at the MTOC. The rapidity with
which this occurs suggests that hnRNP A2 serves key
roles in nuclear retention of RNA and genomic RNA
trafficking to the cytosol; moreover, these results indicate
that genomic RNA traffic might be blocked at this site in
these conditions.
The hnRNP A2 is required to traffic HIV-1 RNA from the
MTOC and maintain localization in the cytoplasm
Our previous work showed that hnRNP A2 expression is
necessary for trafficking of A2RE-containing HIV-1 RNAs
from the perikaryon into the dendrites of oligodendrocytes.
Furthermore, antisense-DNA-mediated suppression of
Figure 4: HIV-1 genomic RNA and g-tubulin staining patterns coincide in siA2-treated, HIV-1-expressing cells. HeLa cells were
mock transfected or transfected with HxBRU alone or with either siNS (panels A–D) or siA2 (panels E–H). At 30–36 h later, cells were
harvested forWestern blot analysis (top panel) and fixed and costained for HIV-1 RNA by FISH (panels A and E) and g-tubulin by IF (B and F).
Black and white renditions of the g-tubulin staining are shown in panels C and G and merged color images are shown in panels D and H.
Western blot analyses for hnRNPs and g-tubulin was performed to verify knockdown of hnRNP A2 (top panel). The g-tubulin was used as
a loading control. The inset in panel H magnifies the MTOC region in a cell (boxed). White bar ¼ 10 mm in panels C and G.
1182 Traffic 2006; 7: 1177–1193
Levesque et al.

hnRNP A2 expression in these cells blocked trafficking
to distal dendrite regions of these cells (A.J.M., E. Barbar-
ese, E. A. Cohen, J. Carson, unpublished). Therefore, in
order to determine if hnRNP A2 functions to promote HIV-
1 RNA transport from the MTOC in provirus-expressing
cells, an experiment was designed to examine the depen-
dence of hnRNP A2 expression on the fate of HIV-1 RNA in
the cytosol. To do this, cells were first transfected with
proviral DNA and expression of HIV-1 was allowed to
progress for 24 h. At this time-point, one set of cells was
fixed and stained for genomic RNA (Figure 6A). The
distribution of RNA was punctate in both nuclear and
cytoplasmic compartments. Cells were then treated with
siRNA (siNS or siA2), and 4 h later, the cells were treated
with Leptomycin B (LMB) to block nuclear RNA export.
The LMB will block genomic RNA export because this is
CRM1 dependent but will not affect hnRNP shuttling (20).
In this way, we examined the fate of the genomic RNA in
the cytoplasmic compartment following hnRNP A2 knock-
down. After 12 h of LMB treatment, cells were fixed and
stained for hnRNP A2 by IF to identify siA2 knockdown
cells and for genomic RNA by FISH. DAPI staining was
used to identify the nucleus. In all siNS-treated cells, HIV-1
RNA was nuclear following LMB treatment and RNA
staining was absent in the cytosol (Figure 6B,C). Presum-
ably at this time-point, most of the genomic RNA that had
accumulated in the cytosol during the first 24 h of HIV-1
expression had been trafficked out of the cell into new
virions, for instance, and/or had been turned over. In cells
exhibiting a significant knockdown of hnRNP A2 expres-
sion (Figure 6D,E), the genomic RNA focalized in a cyto-
plasmic region reminiscent of that observed in siA2-
treated cells. This genomic RNA distribution was observed
as early as at 8 h post-LMB treatment (data not shown). The
distribution of genomic RNA differed from the siA2 pheno-
type shown in Figures 2–6 such that genomic RNA also
accumulated in the nucleus demonstrating that there was
an efficient block to nuclear RNA export in LMB- and siA2-
treated cells. This analysis reveals that in LMB-treated
cells in which nuclear RNA export is blocked, the genomic
RNA found in theMTOC is derived from the cytosol. These
results also show that the MTOC region might and that
hnRNP A2 expression is critical for trafficking of the
genomic RNA away from the MTOC.
Figure 5: Time course of the accumulation of genomic RNA at the MTOC in siA2-treated cells. HeLa cells were transfected with
either siNS or siA2 and then with HxBRU. At 12, 20 and 30 h, the cells were fixed and processed for IF/FISH. The hnRNP A2 expression
levels are shown as red fluorescence signals in cells. Genomic RNA expression levels and distribution in siNS-treated cells are shown
(panels A–C). In siA2-treated cells, the localization of genomic RNA at theMTOCwas apparent at 12 h. Overexposed images are presented
in panels D–F to highlight the weak signals for hnRNP A2. Panels G–I show normal exposed merged images of genomic RNA and hnRNP
A2 and DAPI staining in blue. White bar ¼ 10 mm in panels C, F and I.
Traffic 2006; 7: 1177–1193 1183
hnRNP A2-Mediated HIV-1 RNA Trafficking

Recruitment of HIV-1 genomic RNA to the MTOC by
Rab7-interacting lysosomal protein expression
The MTOC region has been shown to play host to late
endosomes, lysosomes and other cellular organelles.
These organelles are trafficked intracellularly by motor
proteins and adaptor proteins. Our previous work impli-
cated kinesin motor proteins in the translocation of HIV-1
RNAs. It remains unclear, however, whether these motor
proteins act to propel HIV-1 RNA in a vesicle-/membrane-
bound state or as naked ribonucleic protein (RNP) com-
plexes. Translocating RNP complexes (or RNA trafficking
granules) are known to contain RNAs, translation factors,
motor proteins and usually both plus-end- and minus-end-
directed motor proteins, kinesin and dynein (21). We were
interested to determine if the disruption of motor activities
was responsible for HIV-1 RNA trafficking in HeLa cells.
Because the expression of the Rab7-interacting lysosomal
protein (RILP) recruits dynein–dynactin motor complexes
to the MTOC region (19), we evaluated the localization of
genomic RNA following the expression of RILP in HIV-1-
expressing cells. Cells were transfected with HIV-1 proviral
DNA with or without a myc-tagged RILP-expression vector
(19) and processed for IF and FISH coanalyses. In this set
of experiments, cells were also treated with the complete
series of siRNAs that is listed in the headings of the ex-
pression blots shown in Figure 7A to determine if knock-
down conditions would affect the RILP-mediated effects.
The knockdown of hnRNP expression was very effective,
and both hnRNP A1 and hnRNP A2 were barely detectable
in the respective knockdown conditions (Figure 7A, lanes
3, 6 and 7). The myc-RILP was abundantly expressed in
cells (Figure 7A, lanes 4–7). The effects of siA1 on pr55Gag
synthesis are demonstrated here again showing that siA1
treatment resulted in a three-fold decrease in pr55Gag
synthesis levels (Figure 7A, lane 6 and Figure 1A). Equal
loading in the lanes is shown by the invariant levels of
GAPDH and g-tubulin expression in cell extracts (Figure 7
A). In siNS-transfected cells, the distribution of Gag and
Figure 6: hnRNP A2 is required for cytoplasmic trafficking of genomic RNA from the MTOC in HIV-1-expressing cells. HeLa cells
were transfected with HxBRU and HIV-1 expression proceeded from 0–24 h as shown in the scheme in the top panel. At this time, cells
were fixed and processed for FISH analyses (panel A) and other cells were transfected with either siNS (or siA1; not shown) as control or
siA2 to specifically target hnRNP A2 expression. Four hours later (time ¼ 28 h), cells were treated with LMB for 12 h and were then fixed
and processed for IF/FISH coanalyses (40 h). A) HIV-1 genomic RNA at 24 h. Following siRNA and LMB treatments, hnRNP A2 and
genomic RNA localization were assessed in cells. B,C) siNS-treated cells. D,E) siA2-treated cells. Knockdown was confirmed by Western
blot analysis (not shown). White bar ¼ 10 mm in panels A, C and E.
1184 Traffic 2006; 7: 1177–1193
Levesque et al.
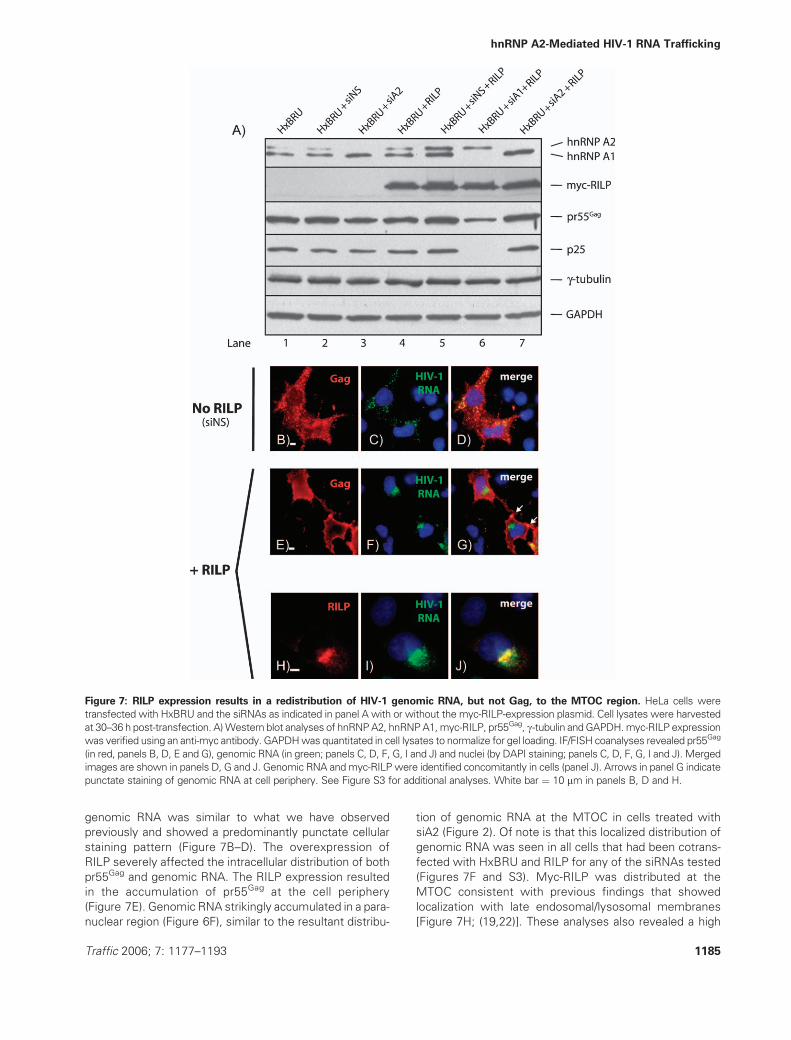
genomic RNA was similar to what we have observed
previously and showed a predominantly punctate cellular
staining pattern (Figure 7B–D). The overexpression of
RILP severely affected the intracellular distribution of both
pr55Gag and genomic RNA. The RILP expression resulted
in the accumulation of pr55Gag at the cell periphery
(Figure 7E). Genomic RNA strikingly accumulated in a para-
nuclear region (Figure 6F), similar to the resultant distribu-
tion of genomic RNA at the MTOC in cells treated with
siA2 (Figure 2). Of note is that this localized distribution of
genomic RNA was seen in all cells that had been cotrans-
fected with HxBRU and RILP for any of the siRNAs tested
(Figures 7F and S3). Myc-RILP was distributed at the
MTOC consistent with previous findings that showed
localization with late endosomal/lysosomal membranes
[Figure 7H; (19,22)]. These analyses also revealed a high
Figure 7: RILP expression results in a redistribution of HIV-1 genomic RNA, but not Gag, to the MTOC region. HeLa cells were
transfected with HxBRU and the siRNAs as indicated in panel A with or without the myc-RILP-expression plasmid. Cell lysates were harvested
at 30–36 h post-transfection. A)Western blot analyses of hnRNPA2, hnRNPA1,myc-RILP, pr55Gag, g-tubulin andGAPDH.myc-RILP expression
was verified using an anti-myc antibody. GAPDHwas quantitated in cell lysates to normalize for gel loading. IF/FISH coanalyses revealed pr55Gag
(in red, panels B, D, E and G), genomic RNA (in green; panels C, D, F, G, I and J) and nuclei (by DAPI staining; panels C, D, F, G, I and J). Merged
images are shown in panels D, G and J. Genomic RNA andmyc-RILP were identified concomitantly in cells (panel J). Arrows in panel G indicate
punctate staining of genomic RNA at cell periphery. See Figure S3 for additional analyses. White bar ¼ 10 mm in panels B, D and H.
Traffic 2006; 7: 1177–1193 1185
hnRNP A2-Mediated HIV-1 RNA Trafficking

degree of colocalization between RILP and the HIV-1
genomic RNA (from 78 to 93% as a range from four ex-
periments, calculated as described in Materials and Meth-
ods; Figure 7I,J). A small proportion of the HIV-1 genomic
RNA was also consistently observed in punctate staining
at the cell periphery with pr55Gag (Figure 7G, arrows; see
Discussion). The overexpression of RILP also resulted in a
60–75% reduction in virus production that was not
enhanced by any of the siRNA treatments (data not shown).
Discussion
hnRNP A2: Multiple roles in HIV-1 post-transcriptional
regulation
In this study, we demonstrate that hnRNP A2 plays a role
in HIV-1 genomic RNA trafficking using detailed imaging
and biochemical techniques. Several new conceptual
advances on HIV-1 egress and viral assembly are sup-
ported by this work. hnRNP A2, a bona fide RNA trafficking
protein in both lower and higher eukaryotes (23–25),
appears to be a dominant player at multiple steps of the
RNA trafficking pathway. In this study, we demonstrate
that the expression levels of hnRNP A2 play a central role
in mediating HIV-1 RNA trafficking out of the nucleus, to
and then from the MTOC (Figures 2–6). First, hnRNP A2
must bind the A2REs in the nucleus to contribute to HIV-1
RNA nucleocytoplasmic trafficking (13). Second, the
expression level of hnRNP A2 appears to mediate this
event and represents either a retention signal or a dominant
signal for nucleocytoplasmic RNA export (Figure S4). Third,
hnRNP A2 must be expressed to sufficient levels to allow
the trafficking of the genomic RNA from the MTOC region
in the cytoplasm (Figure 6). This last role does not appear
to be dependent on any particular late step of the HIV-1
replication cycle as genomic RNA accumulation at the
MTOC occurs as early as 12 h and as late as 36 h post-
transfection (Figure 5). These observations suggest that
hnRNP A2 is an engagement factor for genomic RNA
trafficking, with hnRNP A2 and its cognate cis-sequence
being central components in the regulation of this path-
way. The data presented here are consistent with the
concept that the fate of a messenger RNA (mRNA) is
determined by the proteins with which it interacts through-
out its travels in the nucleus and cytoplasm (26–28).
Predominantly, nuclear proteins such as hnRNP A2 are
also localized in the cytoplasm (29) and this explains their
described roles in cytosolic RNA trafficking (30,31).
The hnRNP A2 RNA trafficking pathway involves the
passage of RNA via the MTOC and this likely represents
one of the first points of convergence of genomic RNA
following its exit from the nucleus. RNA traffic via this
structure must be transient and rapid as there is little
evidence for accumulation of RNA at the MTOC in cells
that express hnRNP A2. Interestingly, genomic RNA
staining can be found to localize closely with g-tubulin in
cells (laser scanning confocal images presented in Figure 4
A–D). For type D retroviruses such as Mason�Pfizer
monkey virus, capsid assembly occurs at this site imme-
diately following Gag synthesis before the capsid is
trafficked through the cytoplasm and then out of the cell
at the plasma membrane (32,33). Interestingly, HIV-1 as
well as other retroviruses use this site in an early step
immediately following infection at which viral capsids or
RNP complexes (the preintegration complex in the case of
HIV-1) converge before nuclear entry (34,35). These re-
sults support the concept that theMTOC is a site via which
HIV-1 gets in and out of the nucleus (35). Nevertheless,
future work will be necessary to unequivocally identify
genomic RNA trafficking to and from the MTOC using
refined molecular and live cell imaging techniques.
hnRNP A2, but not hnRNP A1, functions in HIV-1 RNA
trafficking
The specificity of the RNA trafficking phenotype between
that of hnRNP A2 and hnRNP A1 is another interesting
aspect of this work. Whereas hnRNP A1 knockdown
significantly affects pr55Gag synthesis without any effect
on RNA localization (Figures 1 and 2), knockdown of
hnRNP A2 has virtually no effect on pr55Gag synthesis
but has a major impact on RNA localization and trafficking.
Our splicing data also demonstrate that there are no
marked effects on HIV-1 RNA levels or on RNA splicing
patterns in both conditions as demonstrated in two types
of assays (Figure 1 and data not shown) despite in vitro
and in vivo evidence that support a role for these hnRNPs
in splicing regulation (36–38). This is the first study to
demonstrate that compromised levels of hnRNP A1 neg-
atively affect pr55Gag synthesis that is not the result of
effects at the levels of splicing or RNA expression or
localization (Figures 1, 2 and 7). This reproducible effect on
pr55Gag synthesis might therefore be the result of an
influence on the HIV-1 internal ribosome entry site (IRES)
(39,40). Consistently, hnRNP A1 expression positively
influences translation initiation via an IRES in the fibroblast
growth factor mRNA (41). However, the degree to which
pr55Gag synthesis is blunted cannot be completely attribut-
able to an effect of hnRNP A1 on HIV-1 RNA IRES function.
The involvement of hnRNP A1 at multiple levels of retro-
viral posttranscriptional regulation is supported by several
studies (42,43), and further work will be required to define
how the depletion of hnRNP A1 leads to compromised
pr55Gag expression. Thus, the functions of hnRNP A1 and
hnRNP A2 during HIV-1 replication are now being resolved
to better define how their functions overlap and differ.
hnRNP A2: A role in genomic RNA retention in the
nucleus?
Our previous work revealed that the hnRNP A2–A2RE
interaction in the nucleus influences nuclear export of
genomic RNA (Figure 8, step 1a). Two sets of data shown
here suggest that hnRNP A2 has a role in nuclear retention
of genomic RNA. First, upon hnRNP A2 knockdown, we
generally observe lower levels of nuclear genomic RNA
(Figure 8, step 1b). This observation suggests that the
1186 Traffic 2006; 7: 1177–1193
Levesque et al.

reduction of hnRNP A2 expression results in rapid nuclear
export of the genomic RNA to theMTOC (Figures 2 and 5).
These results might point to a role of hnRNP A2 expression
in the balance between nuclear retention and export of
RNA. In cases when hnRNP A2 is knocked down during
expression of proviral gene (HxBRU or A2RE-2/A8G),
nuclear retention of the genomic RNA is overcome to
become cytosolically localized at the MTOC region
[Figures 2–6, S4 and 8 (steps 1b and 1c)]. Thus, in the
presence of an intact or mutated A2RE, the depletion of
hnRNP A2 serves as a dominant signal for nuclear export.
This even appears to be the case in a Rev-background
where the RNA is released into the cytoplasm when
hnRNP A2 is depleted by siRNA (A. J. M., manuscript in
preparation). The depletion of hnRNP A2 somehow alters
the nuclear HIV-1 RNP to overcome RNA retention in
the nucleus that is achieved by a variety of signals. It is
not clear at present if hnRNP A2 depletion has any
impact on CRM1-mediated RNA export or confers
CRM1-independent export of genomic RNA, but the data
presented in Figure 6 suggest that it does not as RNA
remains in the nucleus in the presence of LMB.
When the hnRNP A2–HIV-1 RNA interaction is prevented
by A2RE mutation, it is retained in the nucleus despite the
expression of Rev (Figure 8, step 1d), the key viral
mediator of nucleocytoplasmic RNA trafficking. This effect
is also coincident with the late expression phase at which
a threshold of Rev is reached and a switch between
splicing and nuclear export of unspliced RNA occurs
(13,44). The association of hnRNP A2 with the A2RE or
its expression levels appear to influence the nuclear export
of the Rev-dependent genomic RNA, a conclusion that
was hinted to in our previous study (13). One possibility is
that this could be via either a direct action by binding to Rev
or interaction with CRM1, as was shown for hRIP and the
RNA helicase, DDX3 (5,9,45). Alternatively, hnRNPs have
been shown to synergize with Rev (43), interact with the
Rev-responsive RNA element of HIV-1 (46) and interfere
with Rex RNA binding in HTLV-1 (47). The hnRNPs can also
interact with known regulators of retroviral RNA nucleo-
cytoplasmic export such as Sam68 (48). In this study, the
depletion of hnRNP A2may somehowmodulate the nuclear
HIV-1 RNP, making the genomic RNA more competent for
export. Nevertheless, it is clear from this work that hnRNP
A2 expression levels are important determinants in mediat-
ing genomic RNA export from the nucleus.
Nuclear export to the MTOC: Via dynein-dependent
translocation on microtubules?
Following nuclear export, we propose that HIV-1 genomic
RNA makes its way to the MTOC for subsequent rapid
trafficking toward the periphery. Mechanistically, this
could be by simple diffusion or the genomic RNA could
be propelled along microtubules toward the minus end of
themicrotubules in a dynein-dependent manner, leading to
efficient MTOC localization (? in Figure 8). The latter idea
appears to be the more probable, and in normal cells, this
Figure 8: Model for HIV-1 RNA traffickingmediated by hnRNPA2. Step 1: The association of hnRNP A2 to the HIV-1 RNA via the intact
A2RE acts to facilitate nuclear export of the RNA to the cytosol (step 1a). Knockdown of hnRNP A2 leads to enhanced nuclear export (step
1b, this article), even in the presence of a mutated A2RE (A2RE/A8G, step 1c). Mutation of the A2RE leads to a block in nuclear export (step
1d) perhaps by influencing Rev function or interference of the RRE. Following nucleocytoplasmic export (step 2), the RNA is possibly
trafficked toward the MTOC by a microtubule minus-end-directed motor, dynein (?). The RNA can be translated to make Gag (step 3) and
a small proportion can be concomitantly selected for encapsidation at or near the MTOC (step 4), followed by a rapid shuttling via Rab9-
containing vesicles (step 5) to the plasmamembrane by way of the TGN (step 6) at which Env can interact and rapidly shuttle the Gag–RNA
complex to sites of virus assembly (step 7) via vesicles that possess specific Rab proteins. In cases when hnRNP A2 is knocked down by
siRNA (steps 1b and 1c), the genomic RNA is released from the nucleus and accumulates at the MTOC region in the cytosol. hnRNP A2
expression is necessary for subsequent trafficking via an RNP complex that likely contains hnRNP A2 and adaptor proteins such as TOG
(step 5a). This complex likely translocates on microtubules because the recruitment of the dynein–dynactin complex by RILP expression
leads to sequestration of the majority of the genomic RNA to theMTOC region, as well as Rab7-containing LE. The RNA transported in this
complex is likely needed as the principal cofactor for virus assembly (step 7). LE, late endosomes.
Traffic 2006; 7: 1177–1193 1187
hnRNP A2-Mediated HIV-1 RNA Trafficking

would allow directed outbound trafficking of the genomic
RNA, implicating a switch in the motor activity from dynein
to kinesin-directed motoring toward the plus ends of
microtubules that might rely, for instance, on Gag synthe-
sis (see below). The observed accumulation of genomic
RNA at the MTOC in hnRNP A2-depleted cells indicates
that this first trafficking event is not disrupted but down-
stream trafficking is (see below). Further work will be
required to decipher the details surrounding these initial
trafficking events following nuclear export.
pr55Gag synthesis and genomic RNA selection occur
near the MTOC
The data presented in this study lead us to the conclusion
that the genomic RNA is translated in the vicinity of the
MTOC in HeLa cells (Figure 8, step 3). This conclusion is
supported by the data that show that pr55Gag synthesis
levels are similar in mock-, siA2- and RILP-treated cells,
despite almost complete MTOC localization of the geno-
mic RNA in the latter conditions (Figures 7 and S3). Further
support for localized synthesis of pr55Gag at the MTOC is
the published report that shows the return of pr55Gag to
late endosomal compartments when the expression of the
small GTPase, Rab9, is knocked down, a treatment that
prevents late endosome to trans-Golgi network (TGN)
pr55Gag trafficking (49). In addition, recent data point toward
the involvement of the TGN in sorting pr55Gag to the plasma
membrane by a TGN-associated E3 ubiquitin ligase named
human POSH (50). The results of this last study implied
that newly synthesized pr55Gag is quickly routed to the
TGN in wait of Env recycling from the plasma membrane.
This would be consistent with observations that a number
of transmembrane proteins such as M6PR, furin and
TGN38 traffic between late endosomes and the TGN and
are rapidly recycled to the plasma membrane (51).
Recent work has also identified the MTOC region as the
site at which, following synthesis, pr55Gag interacts and
selects its cognate genomic RNA (52) (Figure 8, step 4).
This report indicated that this is an infrequent event, and
overexpression was required to detect the Gag–RNA
interaction in the MTOC region. Consistently, we did not
detect any appreciable colocalization between RNA and
pr55Gag in this region in siA2 and RILP conditions. Because
these conditions enhance genomic RNA encapsidation
(Figure S5 and data not shown), our work also implicates
the MTOC region in genomic RNA encapsidation. This
might very well be due to the resultant abundance or
concentration of genomic RNA in this region, which would
favor genomic RNA capture by newly synthesized pr55Gag
(Figures S3 and S5). Genomic RNA trafficking and pr55Gag
synthesis must be linked at the molecular level and
spatially in the cell at some point (at the MTOC) because
the genomic RNA is translated to produce pr55Gag and
then must bind its own RNA for encapsidation (52–55).
The trafficking of genomic RNA and pr55Gag is character-
ized by a rapid transit to assembly domains, although each
of these viral components appears to be differentially
treated in cells. Following pr55Gag synthesis on cytosolic
polyribosomes in the MTOC region for instance, pr55Gag
could transit to the TGN in a Rab9-dependent manner,
followed by a rapid transit to the plasmamembrane (49,50)
(Figure 8, steps 5–7). This is consistent with the findings
that pr55Gag is found at the plasmamembrane immediately
following synthesis in HeLa cells (56). While this route may
be exploited by HIV-1, one significant difference exists
between our observations for RNA and pr55Gag trafficking
in this respect. If pr55Gag and the RNA are trafficked
together, it would be expected that they would exhibit
the same behavior with a given treatment. When we
overexpress RILP, however, pr55Gag and RNA are found
in completely opposite positions, with pr55Gag at the
periphery and its RNA at the MTOC (Figures 7 and S3).
While this finding supports a late endosome to TGN sorting
of the genomic RNA because dynein activity is required for
this sorting event (57), pr55Gag trafficking does not appear
to depend on dynein and is found at the plasma membrane
in these conditions. Nevertheless, only a small proportion
of the genomic RNA is selected for by pr55Gag in the
MTOC region as suggested (52), indicating that only
a minor proportion of the RNA co-traffics with pr55Gag to
reach assembly sites at the plasma membrane. Consis-
tently, calculations of the colocalization coefficient (total
amount of pr55Gag versus the amount found with genomic
RNA) indicate that only 8–25% of the pr55Gag signal is
found with genomic RNA in cells (e.g. Figure 7; calculated
range from 10 experiments). This is also reflected in our
imaging results that show that very little of the genomic
RNA is found to colocalize with pr55Gag at the plasma
membrane in normal (Figure 7C) and siA2 conditions
(Figure 7F) [see also Poole et al. (52)]. HIV-1 may co-opt
more than one sorting pathway to ensure pr55Gag and RNA
assembly, perhaps guaranteeing this by relying on inherent
membrane-binding signal of pr55Gag or on its ability to
directly associate to kinesin proteins during egress, for
example. Because viral assembly occurs at the plasma
membrane in HeLa cells (15,56), the data support impor-
tant roles for both rapid pr55Gag and RNA trafficking events
to the plasma membrane for assembly.
Now that pr55Gag is made and the genomic RNA is
selected for in the MTOC region, what would be the need
for subsequent hnRNP A2-dependent RNA trafficking from
the MTOC in the cytosol? Part of the answer lies in the
requirement for RNA in assembly. Numerous studies now
demonstrate the requirement for RNA for pr55Gag dimer-
ization and subsequent steps of assembly (58–60). Thus,
hnRNP A2-mediated RNA trafficking to distal sites is likely
needed later for RNA-dependent assembly, leading to
virion morphogenesis and capsid formation (61,62) and
this relies on the expression levels of hnRNP A2, dynein
and other auxiliary microtubule-binding proteins (23) (Fig-
ure 8, step 5a). Although HIV-1 and other retroviruses may
be able to use cellular or ribosomal RNAs as scaffolds for
assembly (59,63), the genomic RNA may be the ideal one
1188 Traffic 2006; 7: 1177–1193
Levesque et al.

for HIV-1. Consistently, while cellular pr55Gag synthesis is
not compromised in siA2- or RILP-overexpressing condi-
tions, viral production and infectivity are markedly reduced
in these two conditions (Figure 1B and data not shown).
The aberrant distribution of pr55Gag that results when the
hnRNP A2–A2RE interaction is blocked might also be
explained by the lack of genomic RNA availability in the
cytosol late in the replication cycle (13).
Conserved RNA trafficking mechanisms used by
eukaryotic and viral RNAs
A number of reports now indicate that hnRNP A2-
mediated RNA trafficking is selective and conserved from
Drosophila to man. Hrp48, the closest orthologue of
hnRNP A2 in vertebrates, mediates oskar mRNA traffick-
ing toward the plus end of microtubules during oogenesis
(24,25). Modulation of expression of hrp48 or the intro-
duction of germ line missense mutations in the gene do
not result in altered splicing activity (24,25), consistent
with our observations (Figure 1D). RNA localization also
requires the interdependent activities of kinesin and dynein
motor proteins. Not only do these motors share the same
binding site onmicrotubules (64) but also the kinesin-based
translocation depends on dynein activity in mRNA traffick-
ing in Drosophila (65). This is also true for hnRNP A2-
mediated RNA trafficking as these RNA granules contain
both kinesin and dynein motors (66–69). Consistently, the
recruitment of the dynein/dynactin complex by RILP abro-
gates HIV-1 genomic RNA trafficking to the periphery in all
conditions tested (Figures 7 and S3). While RILP expres-
sion causes the recruitment of late endosomes/lysosomes
to the MTOC region (19), this might suggest that HIV-1
RNA is trafficked to distal regions on these vesicles
(Figure 7). However, even though we do observe some
overlap of genomic RNA staining with Lamp-1 late endo-
somal membranes in siA2 knockdown conditions, in siNS-
treated cells, the genomic RNA shows little costaining with
this compartment (Figures S1 and S2). Our data do not rule
out the possibility that RNA trafficking is achieved in RNP
granules for which dynein activity is essential (65). Work in
murine leukemia virus also suggests that RNA is recruited
onto endosomes from a cytoplasmic compartment (70),
implicating both phenomena in retroviral RNA trafficking.
The combined activities of distinct sets of trans-acting
RNA-binding proteins, transport vesicles and motor pro-
teins will relay a given RNA to its final destination (28, 71–
73). HIV-1 appears to have co-opted conserved cellular
mechanisms in the trafficking of its genomic RNA that rely
in large part on hnRNP A2 expression.
Materials and Methods
Antibodies, genetic clones and reagentsMouse monoclonal and rabbit polyclonal anti-hnRNP A2 and anti-hnRNP A1
antisera were generous gifts from William Rigby (Dartmouth Medical
School, NH, USA), and a pan-specific hnRNP antiserum was generously
provided by Benoit Chabot (Universite de Sherbrooke, Sherbrooke, Que-
bec, Canada) (13); rabbit anti-M6PR was a generous gift from Paul Luzio
(Cambridge, UK); rabbit anti-Lamp-1 and anti-Lamp-2 were generous gifts
from Minora Fukuda (University of Medicine and Dentistry, NJ, USA) (74);
rabbit anti-Calnexin was purchased from Stressgen (Ann Arbor, MI, USA);
rat anti-a-tubulin was obtained from Abcam (Cambridge, MA, USA); rabbit
anti-myc was obtained from TechniScience (Montreal, Quebec, Canada)
and rabbit anti-g-tubulin, LMB and nocodazole were purchased from Sigma-
Aldrich. The proviral DNAs, HxBRU and A2RE A8G (harboring a single point
mutation in the second A2RE) have been described elsewhere (75,76).A
myc-epitope-tagged RILP expressor (pRILP-myc) was generously provided
by Dr Markus Thali (University of Vermont) (14) and was identified by IF and
Western analyses using the anti-myc tag monoclonal antibody 12CA5
(Roche, Montreal, Quebec, Canada) or rabbit anti-myc tag polyclonal
antiserum from TechniScience.
Cell culture and transfectionsHeLa cells were grown in Dulbecco’s modified Eagle’s medium (Invitrogen,
Burlington, ON, USA) supplemented with 10% fetal bovine serum and 1%
of Pen/Strep antibiotics at 378C in a humidified atmosphere containing 5%
CO2. The siA2 50-AAGCTTTGAAACCACAGAAGA and siA1 50-AATGGG-
GAACGCTCACGGACT siRNA duplexes were used at 25 nM to knockdown
hnRNP gene expression as described (16). These and the siNS siRNA
(50-AATTCTCCGAACGTGTCACGA) duplexes were synthesized by Qiagen-
Xeragon (Flanders, NJ, USA). The siRNA transfections were performed
exactly as described in Chatel-Chaix et al. (17), except HeLa cells were
used. Exponentially growing HeLa cells were trypsinised, and 3 � 105 to
3.5 � 105 cells were seeded into six-well plates. Cells were also seeded
onto glass cover slips in six-well plates for imaging studies. Twenty-four
hours later or when the cells reached 70% confluency, siRNA duplexes
(25 nM) were transfected using Lipofectamine 2000 (Invitrogen) according
to the manufacturer’s instructions (17). At 24 h post-transfection, a second
transfection was performed using 0.5 mg HxBRU. Thirty hours later, the
cells were washed with ice-cold PBS and lysed in NTEN buffer (100 mM
NaCl, 10 mM Tris, pH 7.5, 1 mM ethylene-diamine-tetraacetic acid, 0.5%
Nonidet P-40) for protein analyses or fixed as described below for imaging
analyses. In experiments using myc-RILP, 0.5 mg of pRILP-myc (14) was
added to transfections with HxBRU (or at the time of the second trans-
fection) to cause recruitment of the dynein/dynactin complex to the MTOC
region and to disrupt kinesin-driven RNA trafficking and processed for IF/
FISH coanalyses 30 h later. In some experiments, nocodazole was used to
disrupt microtubules. This was added to 5 mM 30 min before cell fixation and
IF and FISH analyses. This treatment time was chosen to disrupt micro-
tubules and not Golgi integrity (18). In studies using LMB (Sigma-Aldrich),
cells were first transfected with HxBRU for 24 h to allow expression and
accumulation of genomic RNA in cytosol and then transfected with either
siNS or siA2 duplexes as described (17). Four hours later, LMB was added
to 2 nM for an additional 8 or 12 h after which cells were processed for
Western and IF/FISH analyses. This treatment completely blocked HIV-1
genomic RNA nuclear export as determined by visualization in empirical
FISH experiments while maintaining cell integrity and viability (not shown).
Western blot analysesAt 30 h post-transfection, cells were washed with ice-cold PBS and lysed in
NTEN buffer. Cytosolic extracts were quantified for protein content by the
micro Bradford assay (BioRad, Mississauga, ON, USA). Equal quantities of
protein were loaded onto gels and hnRNPs, pr55Gag, GAPDH and g-tubulinlevels were assessed by Western blot analyses.
Quantitation of virus production and infectivity
studiesVirus production was quantitated on neat harvested supernatant or purified
and concentrated virus by p24 enzyme-linked immunosorbent assay and
infectivity determinations were performed on equal quantities of virus using
CEM–LTR–GFP indicator cells as described (13,17).
Traffic 2006; 7: 1177–1193 1189
hnRNP A2-Mediated HIV-1 RNA Trafficking

IF and FISH analysesThe IF and FISH analyses have been recently described in detail elsewhere
(13). Briefly, HeLa cells were fixed at the specified time-points posttrans-
fection depending on the experiment (in 4% paraformaldehyde in PBS for
20 min, followed by permeabilization with 0.2% Trition-X-100 for 10 min).
Cells were washed with PBS and blocked with 10% dry milk in PBS and
incubated with each of the antibodies described above. Secondary fluo-
rophore-conjugated antisera (Alexa Fluor 488 and 564) were obtained from
Molecular Probes (Eugene, OR, USA). For FISH/IF coanalyses experiments,
the FISH analysis was performed first. Following fixation and permeabiliza-
tion, cells were treated with deoxyribonuclease I (DNase I; Invitrogen) and
washed in PBS. The digoxigenin-labeled RNA probe to identify genomic
RNAwas prepared as described previously (13). In several experiments, the
nucleic acid stain 4’,6-diamidino-2-phenylindole (DAPI) (Molecular Probes,
OR, USA) was used at 1:500 following FISH analysis.
Imaging analysisMost of the imaging of cells was performed on an Olympus BX-51
fluorescence microscope equipped with an UPlanFI 100X oil objectif
(Center Valley, PA, USA). Alexa Fluor 488 nm, 594 nm and DAPI images
were obtained using 460–490 nm, 510–550 nm and 330–385 nm bandpass
emission filters, respectively. Red, green and DAPI images were sequen-
tially captured in black and white with the Spot camera (Diagnostics
Instruments, Sterling Heights, MI, USA) using Spot Advanced Software
and Image-Pro-Plus version 4.0.1. Images were pseudo-colored using
Adobe Photoshop CS (Adobe Systems, San Jose, CA, USA) in RGB mode
and then merged. Phase contrast images were captured to examine cell
morphology and cell intactness and were obtained with the same micro-
scope by transmitted visible light. Images were captured at a resolution of
512 pixels. All merged digitized images were imported into Adobe Illustrator
CS for figure montage shown in this article. In several experiments, the
subcellular distribution of RNA and proteins was confirmed by laser
scanning confocal microscopy using a Carl-Zeiss (Mississauga, ON, USA)
LSM5 Pascal microscope exactly as described before (13). For all micro-
scopy analyses, at least 50 cells per experimental condition in each
experiment were examined and representative cells are shown here.
Settings for image capture were kept constant in each channel when
comparing staining intensities between siNS- and siA2- or siA1-treated
cells. In knockdown cells in particular, the gain was adjusted in order to
balance the signal intensities from knockdown and nonknockdown cells in
the same microscope field of vision and then the slides containing cells
from other transfection conditions were examined using the identical
microscope settings. In many of the images presented in this article,
silenced and nonsilenced cells (for hnRNPs) are presented in the same field
of vision. Most experiments involving extensive image analysis were
performed at least seven times. To provide an index of colocalization
between viral, cellular antigens and genomic RNA in cells, we used
Colocalization Pro Software (Boise, IO, USA) to calculate the overlap
coefficient according to Manders. This was performed in several experi-
ments as indicated in the text in at least 100 cells per treatment to obtain an
average estimation. Colocalization coefficients were also calculated to
determine the relative amount of colocalized RNA and pr55Gag in relation
to the total pr55Gag signal in cells. Size bars were precisely calculated using
Carl-Zeiss LSM Image Browser software (Mississauga, Ontario, Canada).
RNA extraction and RT-PCR analysisHeLa cells were transfected asmentioned above, and total RNAwas isolated
for splicing, RT-PCR and denaturing gel analyses. Cells were washed with
PBS and lysed on ice in NTEN buffer containing Complete protease inhibitor
cocktail (Roche) for 30 min. Lysates were subjected to a 30 min centrifuga-
tion at 14 000x g, 48C. An aliquot of the supernatant was kept for Western
blot analyses for hnRNP A1, hnRNP A2, HIV-1 Gag and GAPDH. Before RNA
extraction, cell lysates were treated with 40 U of RNaseout and 114 U of
DNase I for 5 min at room temperature. Total RNA was extracted from cell
lysates using Trizol LS (Invitrogen) according to the procedure suggested by
the manufacturer. Five micrograms of glycogen (Roche, Montreal, Quebec,
Canada) was used as carrier for RNA precipitation.
Resulting RNAs were subjected to RT-PCR amplification for the 4-kbp,
singly spliced and 2-kbp, multiply spliced HIV-1 RNA species using specific
primer combinations exactly as described previously (13,17) except that the
RNA PCR Core Kit (Applied Biosystems, Streetsville, ON, USA) was used
for the two-step reverse transcription (RT) and PCR reactions. A negative
control included the exclusion of RT in the RT step. After 20 and 30 cycles,
15% of the total RT-PCR products were visualized on 1% agarose gel using
Gel Doc System and quantitated by the Quantity One Software version
4.4.1 (BioRad). Inverted images of the ethidium-bromide-stained gels are
shown in the article. For a more sensitive detection of each of the 4- and
2-kbp HIV-1 RNA species, 20% of RT-PCR products at 20 cycles were
subjected to two additional PCR cycles in the presence of [32P]dCTP and
labeled PCR products were resolved on polyacrylamide/urea gels as
described (13,17).
Cell cycle and apoptosis determinationsCell cycle analysis was performed using fluorescence activated cell sorting
as described (76,77).
Acknowledgments
We thank Jean-Francois Clement for contributions in preliminary experi-
ments; Markus Thali for sharing data prior to publication; William Rigby,
Benoit Chabot, Minora Fukuda and Paul Luzio for antibodies; Fred Maxfield
and Markus Thali for helpful discussions and expression vectors; Kimberly
Hu for technical assistance; and Alan Cochrane for reagents and for critical
reading of the manuscript. A. J. M. is supported by Canadian Institutes of
Health Research (CIHR) New Investigator Award and this work was
supported by grants from the CIHR, the Canadian Foundation for AIDS
Research and the Canadian Foundation for Innovation to A. J. M.
Supplementary Materials
Figure S1: The localization of genomic RNA in siA2-treated cells in
relation to Golgi and late endosomal membranes. HeLa cells were
transfected with HxBRU and siA2. IF/FISH coanalyses at 30–36 h post-
transfection. In all images, the green fluorescence signal and blue are the
genomic RNA and DAPI staining of the nucleus, respectively. The Golgi
apparatus, TGN and late endosomal membranes were identified as red
fluorescence signals by fluorescence microscopy using Bodipy-ceramide
(panels A–C), an anti-M6PR antibody (panels D–F) and an anti-Lamp-1
antibody (panels G–I). Panels A, D and G show the distribution of RNA
(green fluorescence) in relation to the nuclei; panels B, E and H show the
distribution of the genomic RNA in relation to the staining pattern of the
organellar markers and panels C, F and I show the merges of all three
components. Arrows in panels G–I indicate genomic RNA in a radial
distribution pattern in the perinuclear space and the asterisk is placed for
orientation. White bar ¼ 10 mm in panels A, D and G. (See supplementary
materials for results.)
Figure S2: Perinuclear distribution of Lamp-1 late endosomal com-
partments. HeLa cells were mock transfected or transfected with HxBRU
proviral DNA and siNS (panel A), siA1 (not shown) or siA2 (panels C and D).
Cells were processed for IF/FISH coanalyses. A rabbit anti-Lamp-1 was
used to identify late endosomal compartments in all series of cells (red
fluorescence in panels A–D). Green fluorescence represents the genomic
RNA and DAPI staining identifies the nuclei (in blue). Merged images are
shown above. Lamp-1 is expressed in all cells and is found in a discrete
patch or slightly diffused region immediately adjacent to the nucleus in
most cells. Genomic RNA was found widely distributed in a cell in
a punctate staining pattern (panel B) and siA2 treatment to knockdown
1190 Traffic 2006; 7: 1177–1193
Levesque et al.

hnRNP A2 expression resulted in the perinuclear distribution of genomic
RNA that was found in close proximity to the late endosomal compartments
(panels C and D). White bar ¼ 10 mm.
Figure S3: The RILP induces a perinuclear distribution of genomic
RNA in all siRNA conditions. HeLa cells were mock transfected (not
shown; see Figure 7) or transfected with HxBRU proviral DNA and siNS
(panels A-C), siA1 (panel D) or siA2 (panel E). RILP was coexpressed to
recruit the dynein/dynactin motor complexes to the MTOC. Cells were
processed for IF/FISH coanalyses. A rabbit anti-hnRNP A2 was used to
identify hnRNP A2 expression in all series of cells (red fluorescence in
panels A–E). Green fluorescence represents the genomic RNA and DAPI
staining identifies the nuclei (in blue). Merged images are shown above. A
merged image including phase contrast (PC; panel A) of a cell that was
transfected with HxBRU, siNS and vector control only in the same
experiment is shown to demonstrate wild-type distribution of genomic
RNA before RILP expression (panel A). In siNS- and siA1-treated cells,
hnRNP A2 levels are not modulated and is expressed in all cells (panels B–
D), whereas siA2 treatment resulted in a near-complete reduction in the
hnRNP A2 expression in four of the five cells shown (panel E). The HIV-1
genomic RNA was recruited to the MTOC in all conditions when RILP was
expressed. White bar ¼ 10 mm.
Figure S4: The siA2 treatment of HIV-1-/A2RE-expressing cells pro-
motes nucleocytoplamic transport of genomic RNA. Cells were trans-
fected as described in legend Figure 1, except that the a provirus harboring
amutation in the A2RE-2/A8Gwas transfected alone or with the siRNAs as
indicated. A) Western blot analyses reveal efficient knockdown of
hnRNPs. B–E) HIV-1 genomic RNA is identified by FISH (as green
fluorescence) in cells expressing the HIV-1/A2RE provirus alone B) or with
siNS C), siA2 D) or siA1 E). White bar ¼ 10 mm. (See supplementary
material for results.)
Figure S5: The siA2 treatment modulates genomic RNA content in
virions. Because the siA2 RNA localization phenotype is quite dramatic, we
investigated how siA2 treatment impacted on the genomic RNA content in
virions. A) HeLa cells were transfected with HxBRU alone or HxBRU and
siNS, siA1 or siA2. Virus was isolated from cells at 36 h posttransfection,
concentrated by ultracentrifugation, and quantitated by p24 enzyme-linked
immunosorbent assay (ELISA) (17,75). A) p24 virus equivalents per milliliter
are presented in the histogram. Virus was treated with DNase I (13) and
then solubilized in 1% sodium dodecyl sulphate and deproteinized with
proteinase K. The purified RNA was separated in denaturing formaldehyde/
agarose gels and blotted onto a nylon membrane. Northern blot analysis
was performed using a probe that corresponds to a Apa1/Sph1 HIV-1 Gag
DNA fragment that will recognize genomic RNA (75). Genomic RNA signals
were quantitated as described previously (13). This analysis revealed that
siA2 treatment enhanced genomic RNA content by 30% and the genomic
RNA content corresponded directly to p24 levels for HxBRU, siNS and siA1
conditions, as shown by the p24/RNA ratios that are near 1. B) To
independently confirm this result, virus was prepared in an identical manner
in two additional experiments. Virus was normalized for p24 content by
ELISA, and RNA was isolated from pelleted virus [2750 ng equivalents (17)]
and was then solubilized in an equal volume of guanidium thiocyanate
buffer. Aliquots were denatured and slot blotted onto membranes. The
membrane was probed as described above. The O.D. of the resultant
signals were quantitated by densitometry, and the mean genomic RNA
content between the two experiments is shown below blots. These
analyses show that siA2 mediated a 30–60% increase in virion-associated
genomic RNA, suggesting that siA2 impacted on virion RNA content as
shown in A, and this affected negatively on infectivity.
Supplemental materials are available as part of the online article at http://
www.blackwell-synergy.com
References
1. Coffin JM. HIV population dynamics in vivo: implications for genetic
variation, pathogenesis, and therapy [Comments]. Science 1995;267:
483–489.
2. HoDD,Neumann AU, Perelson AS, ChenW, Leonard JM,MarkowitzM.
Rapid turnover of plasma virions and CD4 lymphocytes in HIV-1
infection [Comments]. Nature 1995;373:123–126.
3. Pomerantz RJ, Seshamma T, Trono D. Efficient replication of human
immunodeficiency virus type 1 requires a threshold level of Rev:
potential implications for latency. J Virol 1992;66:1809–1813.
4. Malim MH, Hauber J, Le SY, Maizel JV, Cullen BR. The HIV-1 rev
trans-activator acts through a structured target sequence to activate
nuclear export of unspliced viral mRNA. Nature 1989;338:254–257.
5. Yedavalli VS, Neuveut C, Chi YH, Kleiman L, Jeang KT. Requirement of
DDX3 DEAD box RNA helicase for HIV-1 Rev-RRE export function. Cell
2004;119:381–392.
6. Kjems J, Askjaer P. Rev protein and its cellular partners. Adv Pharmacol
2000;48:251–298.
7. Hofmann W, Reichart B, Ewald A, Muller E, Schmitt I, Stauber RH,
Lottspeich F, Jockusch BM, Scheer U, Hauber J, Dabauvalle MC.
Cofactor requirements for nuclear export of Rev response element
(RRE)- and constitutive transport element (CTE)-containing retroviral
RNAs. An unexpected role for actin. J Cell Biol 2001;152:895–910.
8. Yu Z, Sanchez-Velar N, Catrina IE, Kittler EL, Udofia EB, Zapp ML. The
cellular HIV-1 Rev cofactor hRIP is required for viral replication. Proc
Natl Acad Sci U S A 2005;102:4027–4032.
9. Sanchez-Velar N, Udofia EB, Yu Z., Zapp ML. hRIP, a cellular cofactor
for Rev function, promotes release of HIV-1 RNAs from the perinuclear
region. Genes Dev 2004;18: 23–24.
10. Tang Y, Winkler U, Freed EO, Torrey TA, Kim W, Li H, Goff SP., Morse
HC III. Cellular motor protein KIF-4 associates with retroviral Gag.
J Virol 1999;73:10508–10513.
11. Kim W, Tang Y, Okada Y, Torrey TA, Chattopadhyay SK, Pfleiderer M,
Falkner FG, Dorner F, Choi W, Hirokawa N., Morse HC III. Binding of
murine leukemia virus Gag polyproteins to KIF4, a microtubule-based
motor protein. J Virol 1998;72:6898–6901.
12. Mouland AJ, Xu H, Cui H, Krueger W, Munro TP, Prasol M, Mercier J,
Rekosh D, Smith R, Barbarese E, Cohen EA, Carson JH. RNA
trafficking signals in human immunodeficiency virus type 1. Mol Cell
Biol 2001;21:2133–2143.
13. Beriault V, Clement JF, Levesque K, Lebel C, Yong X, Chabot B, Cohen
EA, Cochrane AW, Rigby WF, Mouland AJ. A late role for the
association of hnRNP A2 with the HIV-1 hnRNP A2 response elements
in genomic RNA, Gag, and Vpr localization. J Biol Chem 2004;
279:44141–44153.
14. Nydegger S, Foti M, Derdowski A, Spearman P, Thali M. HIV-1 egress
is gated through late endosomal membranes. Traffic 2003;4:902–910.
15. Ono A, Freed EO. Cell-type dependent targeting of HIV-1 assembly to
the plasma membrane and the multivesicular body. J Virol 2004;
78:1552–1565.
16. Patry C, Bouchard L, Labrecque P, Gendron D, Lemieux B, Toutant J,
Lapointe E, Wellinger R, Chabot B. Small interfering RNA-mediated
reduction in heterogeneous nuclear ribonucleoparticule A1/A2 proteins
induces apoptosis in human cancer cells but not in normal mortal cell
lines. Cancer Res 2003;63:7679–7688.
17. Chatel-Chaix L, Clement J-F, Martel C, Beriault V, Gatignol A, Des-
Groseillers L, Mouland AJ. Staufen is part of the HIV-1 Gag ribo-
nucleoprotein complex and is involved in the generation of infectious
viral particles. Mol Cell Biol 2004;24:2637–2648.
18. Honda A, Al-Awar OS, Hay JC, Donaldson JG. Targeting of Arf-1 to the
early Golgi by membrin, an ER-Golgi SNARE. J Cell Biol 2005;
168:1039–51.
Traffic 2006; 7: 1177–1193 1191
hnRNP A2-Mediated HIV-1 RNA Trafficking

19. Jordens I, Fernandez-Borja M, Marsman M, Dusseljee S, Janssen L,
Calafat J, Janssen H, Wubbolts R, Neefjes J. The Rab7 effector protein
RILP controls lysosomal transport by inducing the recruitment of
dynein-dynactin motors. Curr Biol 2001;11:1680–1685.
20. Lichtenstein M, Guo W, Tartakoff AM. Control of nuclear export of
hnRNP A1. Traffic 2001;2:261–267.
21. Cochrane AW, McNally MT, Mouland AJ. The retrovirus RNA traffick-
ing granule: from birth to maturity. Retrovirology 2006;3:18.
22. Wu M, Wang T, Loh E, Hong W, Song H. Structural basis for
recruitment of RILP by small GTPase Rab7. EMBO J 2005;24:
1491–1501.
23. Kosturko LD, Maggipinto MJ, D’Sa C, Carson JH, Barbarese E. The
microtubule-associated protein, TOG, binds to the RNA trafficking
protein, hnRNP A2. Mol Biol Cell 2005.
24. Yano T, Lopez de Quinto S, Matsui Y, Shevchenko A, Shevchenko A,
Ephrussi A. Hrp48, a Drosophila hnRNPA/B homolog, binds and
regulates translation of oskar mRNA. Dev Cell 2004;6:637–648.
25. Huynh JR, Munro TP, Smith-Litiere K, Lepesant JA, St Johnston D. The
Drosophila hnRNPA/B homolog, Hrp48, is specifically required for
a distinct step in osk mRNA localization. Dev Cell 2004;6:625–635.
26. Kim VN, Dreyfus G. Nuclear mRNA binding proteins couple pre-mRNA
splicing and post-splicing events. Mol Cell 2001;12:1–10.
27. Kim VN, Yong J, Kataoka N, Abel L, Diem MD, Dreyfuss G. The Y14
protein communicates to the cytoplasm the position of exon-exon
junctions. EMBO J 2001;20:2062–2068.
28. Kress TL, Yoon YJ, Mowry KL. Nuclear RNP complex assembly
initiates cytoplasmic RNA localization. J Cell Biol 2004;165:203–211.
29. Kamma H, Portman DS, Dreyfuss G. Cell type-specific expression of
hnRNP proteins. Exp Cell Res 1995;221:187–196.
30. Farina KL, Singer RH. The nuclear connection in RNA transport and
localization. Trends Cell Biol 2002;12:466–472.
31. Gu W, Pan F, Zhang H, Bassell GJ, Singer RH. A predominantly nuclear
protein affecting cytoplasmic localization of beta-actin mRNA in fibro-
blasts and neurons. J Cell Biol 2002;156:41–51.
32. Sfakianos JN, LaCasse RA, Hunter E. The M-PMV cytoplasmic
targeting-retention signal directs nascent Gag polypeptides to a peri-
centriolar region of the cell. Traffic 2003;4:660–670.
33. Sfakianos JN, Hunter E. M-PMV capsid transport is mediated by Env/
Gag interactions at the pericentriolar recycling endosome. Traffic
2003;4:671–680.
34. Petit C, Giron ML, Tobaly-Tapiero J, Bittoun P, Real E, Jacob Y, Tordo
N, De The H, Saib A. Targeting of incoming retroviral Gag to the
centrosome involves a direct interaction with the dynein light chain 8.
J Cell Sci 2003;116:3433–3442.
35. McDonald D, Vodicka M, Lucero G, Svitkina Y, Borisy G, Emerman M,
Hope TJ. Visualization of the intracellular behavior of HIV in living cells.
J Cell Biol 2002;159:441–452.
36. Kashima T, Manley JL. A negative element in SMN2 exon 7 inhibits
splicing in spinal muscular atrophy. Nat Genet 2003;34:460–463.
37. Bilodeau PS, Domsic JK, Mayeda A, Krainer AR, Stoltzfus CM. RNA
splicing at human immunodeficiency virus type 1 3’ splice site A2 is
regulated by binding of hnRNP A/B proteins to an exonic splicing
silencer element. J Virol 2001;75:8487–8497.
38. Caputi M, Mayeda A, Krainer AR, Zahler AM. hnRNP A/B proteins are
required for inhibition of HIV-1 pre-mRNA splicing. EMBO J 1999;
18:4060–4067.
39. Buck CB, Shen X, Egan MA, Pierson TC, Walker CM, Siliciano RF. The
human immunodeficiency virus type 1 gag gene encodes an internal
ribosome entry site. J Virol 2001;75:181–191.
40. Brasey A, Lopez-Lastra M, Ohlmann T, Beerens N, Berkhout B, Darlix
JL, Sonenberg N. The leader of human immunodeficiency virus type 1
genomic RNA harbors an internal ribosome entry segment that is active
during the G2/M phase of the cell cycle. J Virol 2003;77:3939–3949.
41. Bonnal S, Pileur F, Orsini C, Parker F, Pujol F, Prats AC, Vagner S.
Heterogeneous nuclear ribonucleoprotein A1 is a novel internal ribo-
some entry site trans-acting factor that modulates alternative initiation
of translation of the fibroblast growth factor 2 mRNA. J Biol Chem
2005;280:4144–4153.
42. Kress E, Baydoun HH, Bex F, Gazzolo L, Duc Dodon M. Critical role of
hnRNP A1 in HTLV-1 replication in human transformed T lymphocytes.
Retrovirology 2005;2:8.
43. Najera I, Krieg M, Karn J. Synergistic stimulation of HIV-1 rev-depend-
ent export of unspliced mRNA to the cytoplasm by hnRNP A1. J Mol
Biol 1999;285:1951–1964.
44. Pomerantz RJ, Trono D, Feinberg MB, Baltimore D. Cells nonproduc-
tively infected with HIV-1 exhibit an aberrant pattern of viral RNA
expression: a molecular model for latency. Cell 1990;61:1271–1276.
45. Neville M, Stutz F, Lee L, Davis LI, Rosbash M. The importin-beta
family member Crm1p bridges the interaction between Rev and the
nuclear pore complex during nuclear export. Curr Biol 1997;7:
767–775.
46. Xu Y, Reddy TR, FischerWH,Wong-Staal F. A Novel hnRNP specifically
interacts with HIV-1 RRE RNA. J Biomed Sci 1996;3:82–91.
47. Dodon MD, Hamaia S, Martin J, Gazzolo L. Heterogeneous nuclear
ribonucleoprotein A1 interferes with the binding of the human T cell
leukemia virus type 1 rex regulatory protein to its response element.
J Biol Chem 2002;277:18744–18752.
48. Yang JP, Reddy TR, Truong KT, Suhasini M, Wong-Staal F. Functional
interaction of Sam68 and heterogeneous nuclear ribonucleoprotein K.
Oncogene 2002;21:7187–7194.
49. Murray JL, Mavrakis M, McDonald NJ, Yilla M, Sheng J, Bellini WJ,
Zhao L, Le Doux JM, Shaw MW, Luo CC, Lippincott-Schwartz J,
Sanchez A, Rubin DH, Hodge TW. Rab9 GTPase is required for
replication of human immunodeficiency virus type 1, filoviruses, and
measles virus. J Virol 2005;79:11742–11751.
50. Alroy I, Tuvia S, Greener T, Gordon D, Barr HM, Taglicht D, Mandil-
Levin R, Ben-Avraham D, Konforty D, Nir A, Levius O, Bicoviski V, Dori
M, Cohen S, Yaar L et al. The trans-Golgi network-associated human
ubiquitin-protein ligase POSH is essential for HIV type 1 production.
Proc Natl Acad Sci U S A 2005;102:1478–1483.
51. Maxfield FR, McGraw TE. Endocytic recycling. Nat Rev Mol Cell Biol
2004;5:121–132.
52. Poole E, Strappe P, Mok HP, Hicks R, Lever AM. HIV-1 Gag-RNA
interaction occurs at a perinuclear/centrosomal site; analysis by confo-
cal microscopy and FRET. Traffic 2005;6:741–755.
53. Butsch M, Boris-Lawrie K. Translation is not required to generate virion
precursor RNA in human immunodeficiency virus type 1-infected T
cells. J Virol 2000;74:11531–11537.
54. LeBlanc JJ, Beemon KL. Unspliced Rous sarcoma virus genomic RNAs
are translated and subjected to nonsense-mediated mRNA decay
before packaging. J Virol 2004;78:5139–5146.
55. Poon DT, Chertova EN, Ott DE. Human immunodeficiency virus type 1
preferentially encapsidates genomic RNAs that encode Pr55(Gag):
functional linkage between translation and RNA packaging. Virology
2002;293:368–378.
56. Rudner L, Nydegger S, Coren LV, Nagashima K, Thali M, Ott DE.
Dynamic fluorescent imaging of human immunodeficiency virus
type 1 gag in live cells by biarsenical labeling. J Virol 2005;79:
4055–4065.
57. Itin C, Ulitzur N, Muhlbauer B, Pfeffer SR. Mapmodulin, cytoplasmic
dynein, and microtubules enhance the transport of mannose 6-phos-
phate receptors from endosomes to the trans-golgi network. Mol Biol
Cell 1999;10:2191–2197.
58. Alfadhli A, Dhenub TC, Still A, Barklis E. Analysis of human immuno-
deficiency virus type 1 Gag dimerization-induced assembly. J Virol
2005;79:14498–14506.
1192 Traffic 2006; 7: 1177–1193
Levesque et al.

59. Ott DE, Coren LV, Gagliardi TD. Redundant roles for nucleocapsid and
matrix RNA-binding sequences in human immunodeficiency virus type
1 assembly. J Virol 2005;79:13839–13847.
60. Huseby D, Barklis RL, Alfadhli A, Barklis E. Assembly of human
immunodeficiency virus precursor gag proteins. J Biol Chem
2005;280:17664–17670.
61. Derdowski A, Ding L, Spearman P. A novel fluorescence resonance
energy transfer assay demonstrates that the human immunodeficiency
virus type 1 Pr55Gag I domain mediates Gag-Gag interactions. J Virol
2004;78:1230–1242.
62. Larson DR, Ma YM, Vogt VM, Webb WW. Direct measurement of
Gag-Gag interaction during retrovirus assembly with FRET and
fluorescence correlation spectroscopy. J Cell Biol 2003;162:
1233–1244.
63. Muriaux D, Mirro J, Harvin D, Rein A. RNA is a structural element in
retrovirus particles. Proc Natl Acad Sci U S A 2001;98:5246–5251.
64. Mizuno N, Toba S, Edamatsu M, Watai-Nishii J, Hirokawa N,
Toyoshima YY, Kikkawa M. Dynein and kinesin share an overlapping
microtubule-binding site. EMBO J 2004;23:2459–2467.
65. Duncan JE, Warrior R. The cytoplasmic dynein and kinesin motors have
interdependent roles in patterning the Drosophila oocyte. Curr Biol
2002;12:1982–1991.
66. Carson JH, Barbarese E. Systems analysis of RNA trafficking in neural
cells. Biol Cell 2005;97:51–62.
67. Kanai Y, Dohmae N, Hirokawa N. Kinesin transports RNA: isolation and
characterization of an RNA-transporting granule. Neuron 2004;43:
513–525.
68. Song J, Carson JH, Barbarese E, Li FY, Duncan ID. RNA transport in
oligodendrocytes from the taiep mutant rat. Mol Cell Neurosci
2003;24:926–938.
69. Ling SC, Fahrner PS, Greenough WT, Gelfand VI. Transport of
Drosophila fragile X mental retardation protein-containing ribonucleo-
protein granules by kinesin-1 and cytoplasmic dynein. Proc Natl Acad
Sci U S A 2004;101:17428–17433.
70. Basyuk E, Galli T, Mougel M, Blanchard JM, Sitbon M, Bertrand E.
Retroviral genomic RNAs are transported to the plasma membrane by
endosomal vesicles. Dev Cell 2003;5:161–174.
71. Yoon YJ, Mowry KL. Xenopus Staufen is a component of a ribonucleo-
protein complex containing Vg1 RNA and kinesin. Development 2004;
131:3035–3045.
72. Kiebler MA, DesGroseillers L. Molecular insights into mRNA transport
and local translation in the mammalian nervous system. Neuron
2000;25:19–28.
73. Cohen RS. The role of membranes and membrane trafficking in RNA
localization. Biol Cell 2005;97:1–13.
74. Carlsson SR, Roth J, Piller F, Fukuda M. Isolation and characterization
of human lysosomal membrane glycoproteins, h-lamp-1 and h-lamp-2.
Major sialoglycoproteins carrying polylactosaminoglycan. J Biol Chem
1988;263:18911–18919.
75. Mouland AJ, Mercier J, Luo M, Bernier L, DesGroseillers L, Cohen EA.
The double-stranded RNA-binding protein Staufen is incorporated in
human immunodeficiency virus type 1: evidence for a role in genomic
RNA encapsidation. J Virol 2000;74:5441–5451.
76. Yao XJ, Mouland AJ, Subbramanian RA, Forget J, Rougeau N,
Bergeron D, Cohen EA. Vpr stimulates viral expression and induces
cell killing in human immunodeficiency virus type 1-infected dividing
Jiurkat T cells. J Virol 1998;72:4686–4693.
77. Mouland AJ, Coady M, Yao XJ, Cohen EA. Hypophosphorylation of
poly(A) polymerase and increased polyadenylation activity are associ-
ated with human immunodeficiency virus type 1 Vpr expression.
Virology 2002;292:221–230.
78. Pelchen-Matthews A, Kramer B, Marsh M. Infectious HIV-1 assembles
in late endosomes in primary macrophages. J Cell Biol 2003;162:
443–455.
Traffic 2006; 7: 1177–1193 1193
hnRNP A2-Mediated HIV-1 RNA Trafficking




















