
Traditional Management and Morphological Patterns of Myrtillocactus schenckii (Cactaceae) in the...
13
Traditional Management and Morphological Patterns of Myrtillocactus schenckii (Cactaceae) in the Tehuacán Valley, Central Mexico 1 JOSÉ BLANCAS 2 ,ALEJANDRO CASAS * ,2 ,RAFAEL LIRA 3 , AND JAVIER CABALLERO 4 2 Centro de Investigaciones en Ecosistemas, Universidad Nacional Autónoma de México, (Campus Morelia), Apartado postal 27-3 (Santa María de Guido), Morelia, Michoacán 58190, México 3 Laboratorio de Recursos Naturales, UBIPRO, F. E. S.-Iztacala, Universidad Nacional Autónoma de México, Av. de los Barrios No.1, Los Reyes Iztacala, Tlalnepantla, C. P. 54090, Estado de México, Mexico 4 Jardín Botánico, Instituto de Biología, Universidad Nacional Autónoma de México, Apartado Postal 04710, México D.F., Mexico *Corresponding author; e-mail: [email protected] Traditional Management and Morphological Patterns of Myrtillocactus schenckii (Cactaceae) in the Tehuacán Valley, Central Mexico Myrtillocactus schenckii is a columnar cactus end- emic to central México and appreciated for its edible fruit. In the Tehuacán Valley it occurs wild in thorn-scrub forests, but it is also under silviculture management—tolerate in agrofo- restry systems and cultivated in home gardens. Ethnobotanical and morphometric studies were conducted to document its use and management forms, as well as consequences of artificial selection on morphological patterns in managed populations. In silviculture popula- tions artificial selection occurs through selective tolerance of the phenotypes producing more, larger, and sweeter fruits, whereas in home gardens people plant branches of the preferred phenotypes. Morphological differences were found especially in fruit size and production, the main targets of artificial selection. The highest average values of fruit size and production were found in cultivated populations (0.993 cm 3 and 204.45 fruits per branch, respectively), intermediate in silviculture populations (0.819 cm 3 and 70.63 fruits per branch), and the lowest in the wild (0.68 cm 3 and 59.75 fruits per branch). Multivariate statistical analyses differentiated populations according to their management type. Morphological diversity was higher in managed populations (0.703±0.029 and 0.679±0.019 in silviculture and cultivated populations, respectively) than in the wild (0.652±0.016). Managed plants of M. schenckii do not show signs of depending on humans for survival and reproduction and, therefore, domestication should be considered incipient. Key Words: Columnar cacti, domestication, genetic resources, Mesoamerica, non-timber forest products, Tehuacán Valley. Introduction The Tehuacán Valley is one of the areas of Mesoamerica where the history of plant manage- ment has been better documented. Archaeological studies by MacNeish (1967) reported informa- tion on plants used and managed from prehistory, including some of the oldest remains of agricul- ture in the New World. In addition, ethno- botanical studies (Casas et al. 2001; Lira et al. 2009) have documented a broad spectrum of forms of interactions between people and plants currently occurring in the region, among them a variety of agricultural and silviculture practices such as tolerance, promotion, special care, trans- planting, and sowing of nearly 120 native plant species (Casas et al. 2008). Management of all these species is conducted with different inten- sities according to the human energy invested, tools, productivity, and practice of artificial selection, but how intensive the plant manage- 1 Received 13 April 2009; accepted 5 October 2009; published online 7 November 2009. Economic Botany, 63(4), 2009, pp. 375–387. © 2009, by The New York Botanical Garden Press, Bronx, NY 10458-5126 U.S.A.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Traditional Management and Morphological Patterns of Myrtillocactus schenckii (Cactaceae) in the...

Traditional Management and Morphological Patternsof Myrtillocactus schenckii (Cactaceae) in the Tehuacán Valley,Central Mexico1
JOSÉ BLANCAS2, ALEJANDRO CASAS*,2, RAFAEL LIRA3, AND JAVIER CABALLERO
4
2Centro de Investigaciones en Ecosistemas, Universidad Nacional Autónoma de México, (CampusMorelia), Apartado postal 27-3 (Santa María de Guido), Morelia, Michoacán 58190, México
3Laboratorio de Recursos Naturales, UBIPRO, F. E. S.-Iztacala, Universidad Nacional Autónoma deMéxico, Av. de los Barrios No.1, Los Reyes Iztacala, Tlalnepantla, C. P. 54090, Estado de México,Mexico
4Jardín Botánico, Instituto de Biología, Universidad Nacional Autónoma de México, Apartado Postal04710, México D.F., Mexico*Corresponding author; e-mail: [email protected]
Traditional Management and Morphological Patterns of Myrtillocactus schenckii (Cactaceae)in the Tehuacán Valley, Central Mexico Myrtillocactus schenckii is a columnar cactus end-emic to central México and appreciated for its edible fruit. In the Tehuacán Valley it occurswild in thorn-scrub forests, but it is also under silviculture management—tolerate in agrofo-restry systems and cultivated in home gardens. Ethnobotanical and morphometric studieswere conducted to document its use and management forms, as well as consequences ofartificial selection on morphological patterns in managed populations. In silviculture popula-tions artificial selection occurs through selective tolerance of the phenotypes producing more,larger, and sweeter fruits, whereas in home gardens people plant branches of the preferredphenotypes. Morphological differences were found especially in fruit size and production, themain targets of artificial selection. The highest average values of fruit size and productionwere found in cultivated populations (0.993 cm3 and 204.45 fruits per branch, respectively),intermediate in silviculture populations (0.819 cm3 and 70.63 fruits per branch), and thelowest in the wild (0.68 cm3 and 59.75 fruits per branch). Multivariate statistical analysesdifferentiated populations according to their management type. Morphological diversity washigher in managed populations (0.703±0.029 and 0.679±0.019 in silviculture and cultivatedpopulations, respectively) than in the wild (0.652±0.016). Managed plants of M. schenckiido not show signs of depending on humans for survival and reproduction and, therefore,domestication should be considered incipient.
Key Words: Columnar cacti, domestication, genetic resources, Mesoamerica,non-timber forest products, Tehuacán Valley.
IntroductionThe Tehuacán Valley is one of the areas of
Mesoamerica where the history of plant manage-ment has been better documented. Archaeologicalstudies by MacNeish (1967) reported informa-tion on plants used and managed from prehistory,including some of the oldest remains of agricul-ture in the New World. In addition, ethno-
botanical studies (Casas et al. 2001; Lira et al.2009) have documented a broad spectrum offorms of interactions between people and plantscurrently occurring in the region, among them avariety of agricultural and silviculture practicessuch as tolerance, promotion, special care, trans-planting, and sowing of nearly 120 native plantspecies (Casas et al. 2008). Management of allthese species is conducted with different inten-sities according to the human energy invested,tools, productivity, and practice of artificialselection, but how intensive the plant manage-
1 Received 13 April 2009; accepted 5 October2009; published online 7 November 2009.
Economic Botany, 63(4), 2009, pp. 375–387.© 2009, by The New York Botanical Garden Press, Bronx, NY 10458-5126 U.S.A.

ment can be is a question still analyzed by severalauthors (Casas et al. 1999a, 2007; González-Insuasti and Caballero 2007). Columnar cactihave been interesting study systems to analyzethis research question, and studies available onthese plants offer the possibility of documentinghow the intensity of artificial selection may varywithin and among related species (Casas et al.1999a). Also, it is possible to analyze how theintensity of artificial selection, life history aspects,and interactions among the coexisting wild andmanaged plant populations may influence thedegree of domestication (Casas et al. 2007).Columnar cacti produce edible fruits that are
generally gathered from wild populations, butsome are also in situ or silviculture managed inassociation with agroforestry systems (Casas et al.1999a). Silviculture is directed to maintain orincrease availability of useful plant species orparticularly good phenotypes of some species(with larger and sweeter fruits, and with thinnerand fewer spines in the fruit peel), and thismanagement form has been interpreted as artifi-cial selection (Casas et al. 1999b). Some species ofcolumnar cacti are also cultivated in homegardens, where artificial selection is conductedmore intensely by planting both sexual andasexual propagules of the desirable phenotypes,and continually replacing plant materials in thecultivated areas (Casas et al. 2006, 2007).Several species of columnar cacti have been
studied from ethnobotanical, morphometric,reproductive, and population genetics approachesin order to document how artificial selection isconducted and its impacts on plant populations(Casas et al. 2007). The morphometric studies ofStenocereus stellatus (Casas et al. 1999b), Polaskiachende (Cruz and Casas 2002), P. chichipe, andEscontria chiotilla (Casas et al. 2007) suggest thatmorphological divergence between wild andmanaged populations within and between speciesis influenced by the intensity of artificial selectionas well as characteristics of the plant speciesdetermining the degree of reproductive isolationbetween wild and managed populations (Casas etal. 2007). Within species, wild and cultivatedpopulations are generally the most divergent,whereas the silviculture populations are inter-mediate. Among species, morphological diver-gence between wild and managed populations ishigher in species under higher artificial selectionintensity (Casas et al. 2007). But the degree ofreproductive isolation between wild and managed
populations may also influence the degree ofdifferentiation (Otero-Arnaiz et al. 2005; Parra etal. 2008; Tinoco et al. 2005). Gene flow amongpopulations occurs through movement of pollen,seeds, and asexual propagules, but it stronglydepends on the establishing success of propagules.Pollen movement is determined by bats in speciessuch as Stenocereus spp. (Casas et al. 2007),whereas bees are the most important pollinatorsof Escontria chiotilla (Oaxaca-Villa et al. 2006),Polaskia spp. (Cruz and Casas 2002; Otero-Arnaiz et al. 2003), and Myrtillocactus spp. (Ortízet al. n.d.). Seeds of bat-pollinated species havehigher water requirements to germinate andsurvive (Guillén et al. 2009), but vegetativepropagation is more successful with faster growththan branches of bee-pollinated species. Bee-pollinated species have more limited pollenmobility but their seeds have lower water require-ments (Guillén et al. 2009), thus establishingtheir seedlings more successfully; however, thesespecies show limited capacity for vegetativepropagation and grow slower than bat-pollinatedspecies. But these capacities are variable amongspecies; for instance, Polaskia chichipe has goodcapacity of seedling establishment and intermedi-ate capacity of vegetative propagation and growthvelocity. Differentiation between wild and man-aged populations has been found to be higher inS. stellatus compared with Escontria chiotilla,species representing the highest and lowestmanagement intensity levels, respectively (Casaset al. 2007).Management and domestication of Myrtillocac-
tus schenckii has not been analyzed, but informa-tion already available indicates that this species isunder relatively intermediate management inten-sity since, although it can be found undercultivation, it is scarcer than other species inhome gardens (Casas et al. 1999a), and repro-ductive biology studies (Ortíz et al. in press)indicate that vegetative propagation is possiblebut only partially successful. M. schenckii isendemic to semi-arid regions of Puebla andOaxaca, central Mexico, growing wild in tropicaldeciduous and thorn-scrub forests in areas of soilsderived from volcanic rocks at elevations of 1,300to 1,900 m. It is 3 to 5 m in height, with darkgreen branches. Flowers are small (an open corollabeing nearly 2.5 cm diam), white, with diurnalanthesis. Blooming occurs from November toApril, flower production being much higher incultivated populations, where self-pollination also
376 ECONOMIC BOTANY [VOL 63

is more frequent (Ortíz et al. in press). Plabeiamexicana (stingless bee), Apis mellifera, Tabanussp., and Xylocopa mexicanorum (carpenter bee) arethe main flower visitors (Ortíz et al. in press).Fruits are spherical or ellipsoid, 0.8 to 1.5 cm inlength and 0.6 to 1.5 cm in width. Fruiting occursfrom May to July (Bravo-Hollis 1978). Accordingto Ortíz et al. (in press), human management ofM. schenckii influences differences in phenology,frequency of flower visitors, and breeding systemsin populations, which may limit pollen flowbetween wild and managed populations.
In the Tehuacán Valley M. schenckii is called“garambullo” and is appreciated for its ediblefruit. It occurs wild in the cactus forest called“garambullal,” where it is a dominant species andwhere people gather its fruit. But it also occurs inagroforestry systems, where people tolerate juve-nile and adult plants, and in home gardens, whereit is a cultivated plant. We expected to find higherdifferentiation in fruit features between wild andcultivated populations and intermediate differ-entiation between silviculture populations inagroforestry systems and the other populations.The information available also allows us tohypothesize that M. schenckii, similarly to P.chichipe, exhibits an intermediate degree ofdomestication. The purpose of this study wastherefore to document how M. schenckii is usedand managed, how artificial selection is con-ducted, and what are its effects on phenotypefrequencies in populations.
MethodsSTUDY AREA
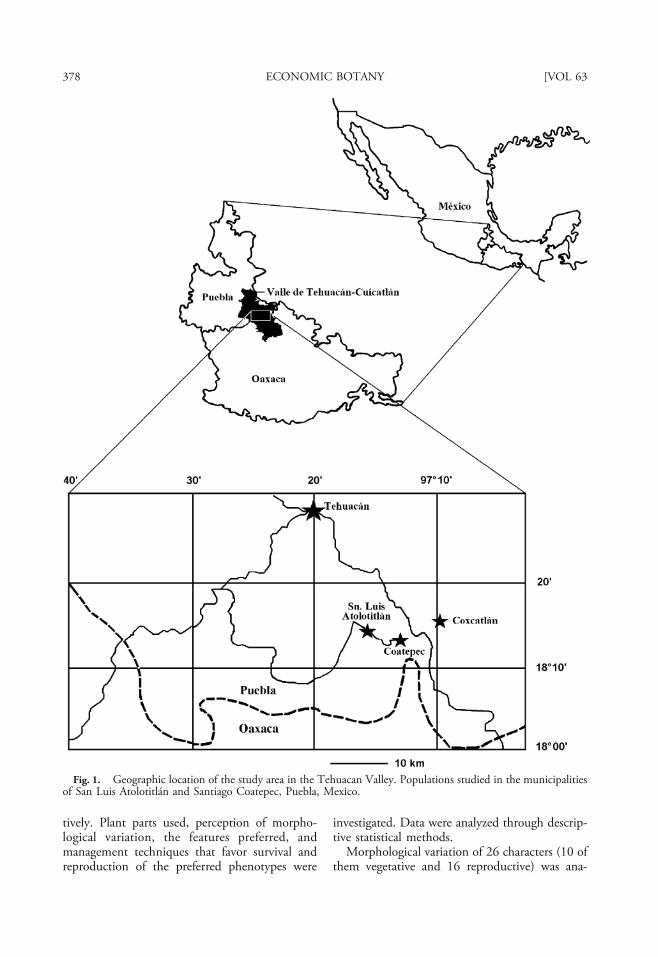
The study was conducted in the territory of thevillages of San Luis Atolotitlán and SantiagoCoatepec, in the state of Puebla, Mexico(Fig. 1), within the Tehuacán-Cuicatlán bio-sphere reserve. Climate of the area is semi-arid,with annual mean temperature of 18.5°C, andannual mean precipitation of 400 mm. Popula-tions of M. schenckii were sampled in elevationsfrom 1,800 to 2,000 m, and included three wild,three silviculture managed, and two cultivatedpopulations. People of the villages are of Náhuatlorigin, whose subsistence depends on rain-fedmaize agriculture, raising goats, and extraction offorest products. Commercialization of productssuch as agave mescal, palm handcrafts, and fruitsfrom their home gardens complement theireconomy.
PLANT POPULATIONS STUDIED
Wild populations occur in natural vegetationcalled garambullal where M. schenckii is adominant species along with trees and shrubssuch as Polaskia chichipe, P. chende, Pittocaulonpraecox, Agave salmiana, Mimosa luisana, Acaciacochlyacantha, Ipomoea arborescens, and Opuntiapillifera. Silviculture populations represent M.schenckii established in agroforestry systemsderived from natural garambullal that have beenunder recurrent cycles of use and fallow forcenturies. Polaskia spp., O. pillifera, I. arbores-cens, and A. salmiana were tolerated when thefields were cleared for cultivation. Cultivatedpopulations are those within home gardens inthe villages of San Luis Atolotitlán and SantiagoCoatepec. Home gardens in these villages areapproximately 200 m2 and frequently contain asmany as 60 species of edible, medicinal, andornamental plant species. Other columnar cactiare cultivated in home gardens, the mostcommon being S. pruinosus (11 individualplants per plot on average) and S. stellatus (7individual plants per plot on average), followedby P. chichipe (5 individual plants per plot onaverage) and M. schenckii (3 individual plantsper plot on average). Samples of 30 adultindividual plants selected randomly werestudied per population (240 individual plantsin total). In wild populations we established1,000 m2 squares where reproductive individualswere sampled at random. We completed thesample of 30 individual plants in a total of 7 and6 plots of agroforestry systems in San LuisAtolotitlán and Coatepec, respectively. Forsampling home gardens, we assigned a numberto each house in a village and selected the samplethrough random numbers. In the TehuacánValley, M. schenckii has a distribution asso-ciated with localized and scarce patches of soilsderived from volcanic rocks, and the villagesstudied are representative of the environments(and populations) where M. schenckii is found inthe region.
ETHNOBOTANICAL AND MORPHOMETRIC
STUDIES
Information on use and management ofgarambullo was documented through ethnobo-tanical semistructured interviews and a survey to30 and 35 households (nearly 20% of the total)randomly selected in the villages studied, respec-
377BLANCAS ET. AL: TRADITIONAL MANAGEMENT AND MORPHOLOGICAL PATTERNS2009]
tively. Plant parts used, perception of morpho-logical variation, the features preferred, andmanagement techniques that favor survival andreproduction of the preferred phenotypes were
investigated. Data were analyzed through descrip-tive statistical methods.Morphological variation of 26 characters (10 of
them vegetative and 16 reproductive) was ana-
Fig. 1. Geographic location of the study area in the Tehuacan Valley. Populations studied in the municipalitiesof San Luis Atolotitlán and Santiago Coatepec, Puebla, Mexico.
378 ECONOMIC BOTANY [VOL 63

lyzed (Table 1). Three to five structures weremeasured and averaged per individual plant.Flowers were collected and preserved in 70%ethanol, whereas fruits were collected and main-tained in plastic bags, refrigerated, and thenmeasured in the laboratory. Fruit size wascalculated based on measures of two perpendicu-lar diameters and using the formula of sphericalbodies volume (v=4/3 π a2b, where a and b arehalf of the diameters measured). Fruit sweetnesswas determined by measuring percentage ofsugars in pulp (ºBrix) through a refractometerAtago (0–32 ºBrixs), and acidity was measured inpH units through a pH meter Denver Instrument215. Measurements of vegetative parts wereconducted in the field. General biomass ofindividual plants was measured using the formulaof an inverted truncated cone v=πh/3 (R2+Rr+r2), in which h is the plant height, R is the canopydiameter, which was estimated as the average oftwo perpendicular measures (north-south andeast-west oriented) of canopy diameter, and r isthe diameter of the main stem measured justbelow the first ramification. Branch diameter wasmeasured in the mid-part of branches at the thirdlevel of ramification, where also number, width,and depth of ribs and distance between areoleswere measured.
DATA ANALYSES
Principal Component (PCA) and DiscriminantFunction Analyses (DFA, Sneath and Sokal 1973)were conducted in order to classify individualplants analyzed according to their general mor-phological similarity, and to explore if theirsimilarity is related to the management regime.PCA was also used to identify those characterswith higher contribution (higher eigenvectorsscores) to the classification, and DFA was directedto test whether the wild, silviculture, and culti-vated groups are significantly different amongthemselves and to identify the proportion ofindividual plants of each group resembling plantsof different groups. A basic data matrix wasconstructed with individual plants as operativetaxonomic units (OTUs) and morphologicalcharacters as variables. Considering the differenttypes of variables and measurement units, thebasic data matrix was standardized through thealgorithm Y’=(Y-a)/b, in which Y’ is the stand-ardized value, Y is the real value of that character,a is its average value, and b is its standard
deviation. PCA was performed based on a Pearsoncorrelation between characters. These analyseswere conducted through NYSYS (Rohlf 1993).
One-way ANOVA and Tukey (95% confi-dence) multiple-range tests were performedthrough JMP (JMP-SAS Institute 1996) toidentify how morphological features differedamong all populations studied and among pop-ulations pooled according to their managementtype. The purpose of these analyses was tovisualize trends of variation among populations,but general differences among groups of popula-tions were tested through DFA.
An index of morphological diversity (MD) wascalculated based on the Simpson diversity index, inorder to summarize information on the amount ofvariation of all variables in a single value. This indexwas defined as MD=1-Σ1-s(pi)
2 in which pi is theproportion of the total number of individual plantssampled in a population showing the ith state ofmorphological character and s is the number ofstates of that character (Casas et al. 2006).Frequencies of character states were first calculated;since all characters studied are quantitative, aprevious conversion to qualitative states wasnecessary. Such transformation was conductedbased on the significance intervals calculatedthrough the ANOVA and Tukey multiple rangetests, similar to the method used by Casas et al.(2006) and Blancas et al. (2006). Phenotypicdifferentiation between pools of wild, silviculturemanaged, and cultivated populations was analyzedusing the algorithm of Nei’s genetic distance (Nei1972), which, in our study, considered the typesand frequencies of morphological character states inpopulations. The degree of differentiation of M.schenckii was compared with those calculated forStenocereus stellatus, Polaskia chichipe, and P. chende,based on data by Casas et al. (2007) and Cruz andCasas (2002).
ResultsUSE, MANAGEMENT, AND ARTIFICIAL
SELECTION
Garambullo is widely used by all peoplesurveyed. Fruits are the main useful part, butdry branches and roots are occasionally used asfuelwood and the entire plant for living fences.Most people (nearly 67%) considered that con-sumption of garambullo fruits has been similarthroughout time, but the remaining peopleinterviewed considered that it was higher in the
379BLANCAS ET. AL: TRADITIONAL MANAGEMENT AND MORPHOLOGICAL PATTERNS2009]
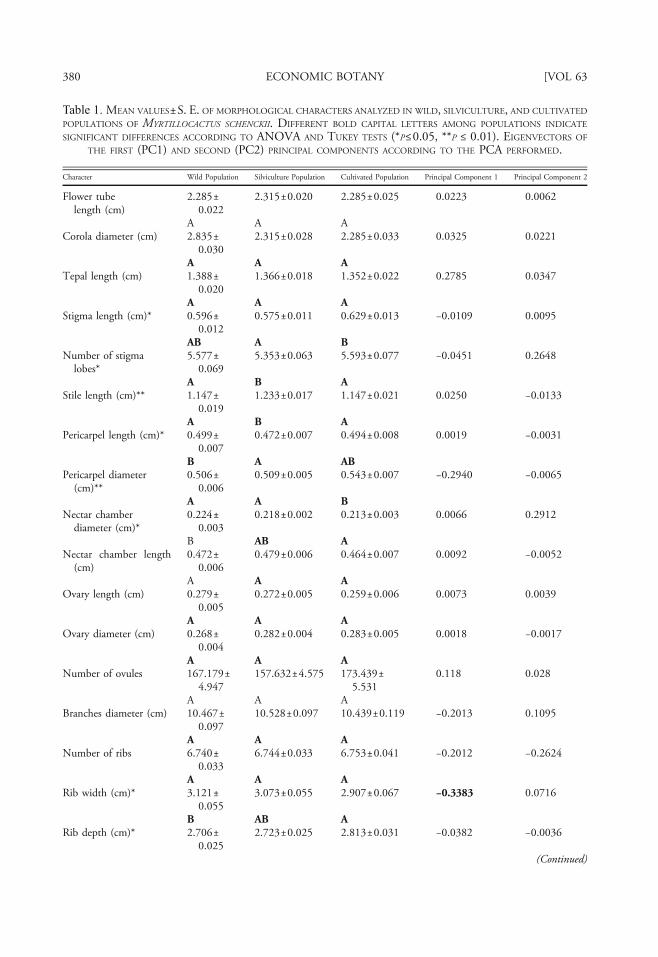
Table 1. MEAN VALUES±S. E. OF MORPHOLOGICAL CHARACTERS ANALYZED IN WILD, SILVICULTURE, AND CULTIVATED
POPULATIONS OF MYRTILLOCACTUS SCHENCKII. DIFFERENT BOLD CAPITAL LETTERS AMONG POPULATIONS INDICATE
SIGNIFICANT DIFFERENCES ACCORDING TO ANOVA AND TUKEY TESTS (*P≤0.05, **P ≤ 0.01). EIGENVECTORS OF
THE FIRST (PC1) AND SECOND (PC2) PRINCIPAL COMPONENTS ACCORDING TO THE PCA PERFORMED.
Character Wild Population Silviculture Population Cultivated Population Principal Component 1 Principal Component 2
Flower tubelength (cm)
2.285±0.022
2.315±0.020 2.285±0.025 0.0223 0.0062
A A ACorola diameter (cm) 2.835±
0.0302.315±0.028 2.285±0.033 0.0325 0.0221
A A ATepal length (cm) 1.388±
0.0201.366±0.018 1.352±0.022 0.2785 0.0347
A A AStigma length (cm)* 0.596±
0.0120.575±0.011 0.629±0.013 −0.0109 0.0095
AB A BNumber of stigmalobes*
5.577±0.069
5.353±0.063 5.593±0.077 −0.0451 0.2648
A B AStile length (cm)** 1.147±
0.0191.233±0.017 1.147±0.021 0.0250 −0.0133
A B APericarpel length (cm)* 0.499±
0.0070.472±0.007 0.494±0.008 0.0019 −0.0031
B A ABPericarpel diameter(cm)**
0.506±0.006
0.509±0.005 0.543±0.007 −0.2940 −0.0065
A A BNectar chamberdiameter (cm)*
0.224±0.003
0.218±0.002 0.213±0.003 0.0066 0.2912
B AB ANectar chamber length(cm)
0.472±0.006
0.479±0.006 0.464±0.007 0.0092 −0.0052
A A AOvary length (cm) 0.279±
0.0050.272±0.005 0.259±0.006 0.0073 0.0039
A A AOvary diameter (cm) 0.268±
0.0040.282±0.004 0.283±0.005 0.0018 −0.0017
A A ANumber of ovules 167.179±
4.947157.632±4.575 173.439±
5.5310.118 0.028
A A ABranches diameter (cm) 10.467±
0.09710.528±0.097 10.439±0.119 −0.2013 0.1095
A A ANumber of ribs 6.740±
0.0336.744±0.033 6.753±0.041 −0.2012 −0.2624
A A ARib width (cm)* 3.121±
0.0553.073±0.055 2.907±0.067 −0.3383 0.0716
B AB ARib depth (cm)* 2.706±
0.0252.723±0.025 2.813±0.031 −0.0382 −0.0036
(Continued)
380 ECONOMIC BOTANY [VOL 63

past, when “people lived using more wildresources and these were more abundant.” Thislast group said that at present some nativeresources (including garambullo fruits) have beenreplaced by other fruits obtained in the market.In the past, they said, the lack of roads limitedcommerce and people had to be more self-sufficient than at present. People depended moreupon agriculture and gathering resources in thewild, whereas at present more people work insmall industries in the neighboring villages andmigrate to other regions of Mexico or the U.S.A.Nearly 67% of people interviewed consideredthat garambullo has decreased its abundancebecause clearing of land for agriculture hasincreased. Some people (15%) also perceived thatrain has decreased and that it is more irregularthan in the past.
Most people interviewed (83%) said thatgarambullo plants are tolerated when land iscleared to establish agricultural fields, whereasthe rest said they remove them. The main reasons
mentioned to tolerate them were that these plants(1) produce edible fruits (43%), (2) should bemaintained according to customary norms (25%),(3) are part of an agricultural practice inheritedfrom their parents (11%), (4) allow shade areasfor people and domestic animals (8%), (5) help toprevent soil erosion (6%), (6) are used for stubblestoring (5%), and (7) may be living fences tolimit the parcels (2%). In contrast, people thatremove garambullo plants argued that theirpresence makes agricultural labors more difficultand occupies space that could be used for maize;they said that because garamabullo is abundant inthe area, it is not affected by its removal fromagricultural fields.
Approximately 20% of people said they trans-plant garambullo plants from wild populationsinto their home gardens. Nearly 50% said thatvegetative propagation is difficult or of uncertainsuccess, whereas the others said that vegetativepropagation is possible but more difficult than forother columnar cacti such as Stenocereus spp.
Table 1. (CONTINUED).
Character Wild Population Silviculture Population Cultivated Population Principal Component 1 Principal Component 2
A AB BDistance betweenareoles (cm)**
1.167±0.015
1.112±0.015 1.225±0.018 −0.0258 0.0043
B A CIndividual plantbiomass (m3)
72.640±6.313
55.554±6.313 66.993±7.732 −0.1413 0.1588
A A AFruit weight (g)** 0.837±
0.0330.960±0.033 1.089±0.040 0.3075 0.1757
A B CFruit volume (cm3)** 0.681±
0.0300.820±0.029 0.994±0.035 0.2753 0.2459
A B CSeed weight (mg)** 0.467±
0.0060.479±0.007 0.435±0.008 −0.310 −0.106
B B ANumber of seeds 118.317±
4.472118.064±4.553 112.797±
5.414−9.9262 −10.4303
A A AFruits per branch** 59.748±
9.5570.625±9.55 204.452±
11.696113.4985 15.5239
A A BSugar content in pulp(ºBrix)
15.956±0.345
16.101±0.341 15.363±0.408 −0.2013 0.1095
A A APulp acidity (pH)** 4.113±
0.0414.042±0.041 3.865±0.049 −0.0248 −0.0043
B B A
381BLANCAS ET. AL: TRADITIONAL MANAGEMENT AND MORPHOLOGICAL PATTERNS2009]

However, only 10% recognized that individualplants producing more and larger fruits arepreferred when making decisions on which plantto tolerate or to transplant.All people interviewed agreed that garambullo
plants occurring in home gardens and agrofor-estry systems do not receive special care ormaintenance, but they indirectly benefit fromsome practices. For instance, in home gardenswhere M. schenckii grows in association with thedragon fruit locally called pitahaya (Hylocereusundatus), people observed that garambullo plantsare more vigorous because of the occasionalirrigation given to pitahayas during the dryseason. According to respondents, garambullosoccurring in agroforestry systems benefit fromagricultural practices such as weeding and tillage.
PATTERNS OF MORPHOLOGICAL VARIATION
According to PCA, the first three principalcomponents explain 93.32% of the variation.Individual plants under different managementregimes were found forming a continuous gra-dient of similarity rather than discrete identifiablegroups (Fig. 2). The first principal componentsclassified most of the cultivated individual plants
in the upper-right side of the plot and most of thewild and silviculture in situ managed individualsgrouped at the left lower area of the plot.Eigenvectors showed that characters with highercontribution to the first principal components aredimensions of fruit (larger in cultivated popula-tion) and seed weight (smaller in cultivatedindividual plants), whereas in the second principalcomponent the number of fruits per branch is themost relevant character (Table 1).According to DFA, the multivariate morpho-
logical differences among the three populationtypes were significant (Table 2), most individualplants being classified according to their manage-ment type. However, it is noteworthy that sometrees from a population are similar to most treesin a population under different management type(Table 3). For example, nearly 27% of wild treesare similar to the silviculture group and only 2wild trees (nearly 3%) are similar to the cultivatedgroup; approximately 21% trees from silviculturepopulations are similar to the wild group, butonly 6 of them (nearly 7%) are similar to thecultivated group; almost 85% cultivated treeswere classified within the cultivated group and thefew remaining trees were similar to the wild andsilviculture groups. This information indicates
Fig. 2. Projection of individuals of Myrtillocactus schenckii in the space of the first and second principal compo-nents. Black symbols are wild populations (▲=San Luis Atolotitlán wild 1; ■=San Luis Atolotitlán wild 2; ●=Coatepec wild); gray symbols are silviculture managed populations ( =San Luis Atolotitlán silviculture 1; =SanLuis Atolotitlán silviculture 2; =Coatepec silviculture); white symbols are cultivated populations (△=San LuisAtolotitlán cultivated; □=Coatepec cultivated).
382 ECONOMIC BOTANY [VOL 63

that wild and silviculture in situ managedpopulations are similar among themselves andclearly different from the cultivated stands, as alsoillustrated in Fig. 3.
One-way ANOVAs identified significant dif-ferences in 14 of the 26 characters analyzed, all ofthem showing significant differences betweencultivated populations and the others. In partic-ular 8 characters (stigma length, pericarpel diam-eter, rib depth, distance between areoles, fruitvolume and weight, seed weight, and number offruits per branch) showed marked differencesaccording to the management type (Table 1).
The highest values of morphological diversity(MD) were found in the silviculture populations(Table 5), which averaged 0.768±0.016, followedby the cultivated populations (MD=0.679±0.019) and the wild populations (MD=0.652±0.016). The highest diversity was found partic-ularly in the population in situ managed 2 (MD=0.842±0.017) (Table 5) and the cultivatedpopulation 1 (MD=0.764±0.017), whereas thelowest values were found in the wild population 1(MD=0.547±0.026).
Discussion
Compared with other columnar cacti species ofthe Tehuacán Valley, Myrtillocactus schenckii canbe considered as a plant resource of relativelyintermediate importance since quantities of fruitconsumed are higher than those reported forPachycereus hollianus (Rodríguez-Arévalo et al.2006) and Myrtillocactus geometrizans, (Casas etal. 1999a, b), but lower than those reported forStenocereus stellatus, Escontria chiotilla, and Polas-kia spp. (Casas et al. 1999a, 2007). In addition, itshould be mentioned that M. schenckii is culti-vated (higher management intensity than E.chiotilla and P. chende, which are not cultivated),but trees of this species are scarcer than othercolumnar cacti, as mentioned above. Fruits arethe main useful part of garambullo, and bothsilviculture management and cultivation involveartificial selection mainly directed to favor phe-notypes producing larger fruits and more fruitsper branch. Most people tolerate garambulloplants in agroforestry systems, some transplantindividual plants or branches in both agroforestry
Table 2. SIGNIFICANCE TESTS OF THE DISCRIMINANT FUNCTION ANALYSIS (DFA).
Discriminant Function Eigen Value Relative Percentage
Canonical Correlation
1 1.426 66.08 0.767
2 0.381 33.92 0.525
Derived Function Wilks’ lambda X2 F L. D. Significance
0 0.23 256.96 6.955 48 0.000000
1 0.723 68.649 23 0.000002
Table 3. CLASSIFICATION OF THE REAL WILD, IN SITU MANAGED, AND CULTIVATED INDIVIDUAL PLANTS INTO
PREDICTED GROUPS ACCORDING TO THE DISCRIMINANT FUNCTION ANALYSIS (DFA).
Actual Group
Predicted Group
Wild In situ Managed Cultivated Total
No. % No. % No. % No. %
Wild 54 70.12 21 27.27 2 2.59 77 100In situ Managed 19 21.11 65 72.22 6 6.66 90 100Cultivated 4 6.66 4 6.66 52 86.66 60 100
383BLANCAS ET. AL: TRADITIONAL MANAGEMENT AND MORPHOLOGICAL PATTERNS2009]

systems and home gardens, but only 10% peoplementioned practicing selection favoring the mostproductive plants. The cultural importance andmanagement intensity appear to have decreased inthe last decades as access to commercialized exoticfruits and out migration increased.The multivariate statistical analyses performed
showed that individual plants are morphologicallysimilar among themselves according to themanagement type, which suggests that manage-
ment influences the morphological patternsreported. The populations studied divergedaccording to the management intensity, thedifferences between wild and cultivated popula-tions being higher than those between wild andsilviculture populations. The one-way ANOVAsperformed consistently found that fruit charactersdiffer among populations according to theirmanagement regime (Table 1), suggesting thatmorphological patterns among populations differ
Fig. 3. Discriminant function analysis. Classification of individuals of Myrtillocactus schenckii according toforms of management. Black symbols are wild populations (▲=San Luis Atolotitlán wild 1; ■=San Luis Atol-otitlán wild 2; ●=Coatepec wild); gray symbols are silviculture managed populations ( =San Luis Atolotitlánsilviculture 1; =San Luis Atolotitlán silviculture 2; =Coatepec silviculture); white symbols are cultivatedpopulations (△=San Luis Atolotitlán cultivated; □=Coatepec cultivated).
Table 4. MORPHOLOGICAL DIVERSITY INDEXES (MD) PER POPULATION STUDIED AND AVERAGED PER MANAGEMENT
TYPE.
Population (MD)
Wild 1 (0.547±0.026)Wild 2 (0.703±0.029)Wild 3 (0.706±0.036)Average (0.652±0.016)Silviculture managed 1 (0.735±0.018)Silviculture managed 2 (0.842±0.017)Silviculture managed 3 (0.729±0.022)Average (0.703±0.029)Cultivated 1 (0.764±0.017)Cultivated 2 (0.595±0.025)Average (0.679±0.019)TOTAL AVERAGE 0.703±0.029
384 ECONOMIC BOTANY [VOL 63

because human management changes the fre-quencies of phenotypes in populations and,consequently, the average values of the charactersanalyzed. Fruit size and number per branch arethe highest in cultivated populations and thelowest in the wild, a pattern similar to thosedocumented in other species of columnar cacti(Casas et al. 2007). Long-term experiments(because of the long life cycle of M. schenckii) ofquantitative genetics are still necessary to testwhether the morphological characters analyzedare heritable, but the occurrence of morphologicalvariation within a single site as reported in thisstudy suggests that genes contribute significantlyto the phenotypic variation.
Seed weight was the lowest in cultivatedpopulations and the highest in wild populations,a finding which differs from the common patternreported for other columnar cacti, in which seednumber and size are directly proportional to fruitsize (Casas et al. 2007). Number of ovules wasnot significantly different among populations;therefore, the fewer seeds produced by fruits ofcultivated populations reveal a lower efficiency ofthe pollinators in the cultivated population,probably associated with a relative scarcity ofpollinators in relation to the high number offlowers that cultivated garambullos produce. Butalso, as revealed by Ortíz et al. (in press), self-pollination is significantly higher in cultivatedpopulations than in the wild, and this breedingsystem results in lower production of seeds thanout-crossing. Therefore, differences in seed num-ber and size among populations could be asso-ciated with frequency of self-pollination occurringin populations under different managementregimes. Flower pericarpel was significantly largerin cultivated and silviculture populations than inthe wild, and this difference could be related toartificial selection favoring larger fruits. Therefore,
fruit size of M. schenckii appears to be related tosize of flower parts rather than to seed number.
The populations of M. schenckii studied havethe highest levels of morphological diversitycompared with other species of columnar cacti(Table 4). As in other species, the highest levels ofmorphological diversity were found in managedpopulations, which indicates that human inter-action with those populations favor variablestands, a common pattern in plants underdomestication (Hawkes 1983), and silvicultureand cultivated populations are reservoirs ofvariation that should be taken into account forconservation purposes.
Patterns of morphological diversity and phe-notypic differentiation of M. schenckii comparedwith other species of columnar cacti are appa-rently related to both management intensity andbiological features of the species (Table 5).Polaskia chende is the least morphologicallydiverse and has the lowest phenotypical differ-entiation among wild and managed populations.This species is also the least intensely managed ofthe species compared, and the enrichment ofmanaged stands with selected new variants ismore difficult than in the other species because ofdifficulties controlling vegetative propagation andits slow growth. Differentiation among wild andmanaged populations is low since artificial selec-tion operating in managed stands is slighter thanin the other species compared, and because geneflow among populations is high (Cruz and Casas2002). Polaskia chichipe and M. schenckii, withhigher morphological differentiation and pheno-typic divergence, are comparable with P. chendesince they are pollinated by diurnal insects, theirseeds are dispersed by birds, and they havesuccessful establishment in wild and managedpopulations (Otero-Arnaiz et al. 2003; Ortíz et al.in press). Therefore, both pollen and seed flow
Table 5. COMPARISON BETWEEN THE MORPHOLOGICAL DIVERSITY INDEXES (DM) AND PHENOTYPIC DIFFERENTIA-
TION AMONG FOUR SPECIES OF COLUMNAR CACTI FROM THE TEHUACÁN VALLEY, CENTRAL MEXICO. 1BLANCAS ET
AL. (2006), 2CASAS ET AL. 2006.
Species
Average
morphological
diversity (MD) index
Phenotypic differentiation
between wild and silviculture
managed populations
Phenotypic differentiation
between wild and
cultivated populations
Polaskia chende 0.348±0.0461 0.009 –Myrtillocactus schenckii 0.703±0.029 0.069 0.110Polaskia chichipe 0.590±0.0071 0.193 0.353Stenocereus stellatus 0.450±0.0152 0.251 0.379
385BLANCAS ET. AL: TRADITIONAL MANAGEMENT AND MORPHOLOGICAL PATTERNS2009]

among wild and managed populations are high.In these cases, however, the higher morphologicaldiversity and phenotypic differentiation betweenwild and managed populations could be explainedby the higher management intensity.Stenocereus stellatus is the most intensely
managed of the species compared and shows alsothe highest levels of morphological diversity andphenotypic differentiation among wild and culti-vated populations. The high variation can beexplained because of the high replacement ofvariants in both agroforestry systems and homegardens (Casas et al. 2006). The high differ-entiation between wild and cultivated populationsis associated with the relatively easy separationfrom their wild relatives since sexual reproductionis episodic (Casas et al. 2007).It is possible to expect higher genetic diversity
and gene flow in species with higher success ofsexual reproduction and, consequently, a lowerdifferentiation among populations. But morpho-logical and population genetics data show thecontrary. This pattern can be explained becausehuman intervention through artificial selectionmay change the natural patterns in artificialpopulations of agroforestry systems and homegardens. Seeds and vegetative propagules aredispersed and cared for to different degrees byhumans in artificial environments, and artificialselection changes phenotype frequencies by favor-ing abundance of individual plants preferred bypeople. These actions may alter gene flow and thegenetic structure of populations and are moreeasily conducted in species with easier vegetativepropagation and faster growth, intermediate inthose species with intermediate capacity ofvegetative propagation and growth, and moredifficult in species with no vegetative propagationand slow growth. Although these patterns couldbe explored more deeply in additional cases tocomplete the panorama, this method appears tohave merit as a means of analyzing the relationbetween management and artificial selectionintensity, as well as biological and reproductiveaspects of coexisting wild and managed popula-tions of plant resources.In conclusion, Myrtillocactus schenckii is a
plant resource of relatively intermediate culturalvalue compared with other species of columnarcacti. It is gathered in wild populations but alsomanaged in agroforestry systems and homegardens, artificial selection favoring individualplants producing larger and more fruits, which
have increased their abundance in managedpopulations. This is a sign of an ongoing processof incipient domestication in which plantsforming part of managed stands do not dependon humans to survive and reproduce. Gene flowamong wild and managed populations appears tobe high and, therefore, artificial selection appearsto be the main factor explaining the morpho-logical differentiation found in this study. Bothagroforestry systems and home gardens areimportant scenarios for studying plant manage-ment, artificial selection, and domestication, andfor conservation programs.
AcknowledgmentsThe authors thank the Dirección General de
Asuntos del Personal Académico, UNAM, Mexico(DGAPA, research projects PAPIIT-IN220005and IN219608), and the Royal Botanic Gardensat Kew (Millennium Seed Bank), U.K., forfinancial support. We also thank Edgar PérezNegrón for support in field work, and people ofSan Luis Atolotitlán and Coatepec, Puebla, fortheir hospitality and valuable collaboration in theresearch.
Literature CitedBlancas, J. J., F. Parra, J. D. Lucio, M. E. Ruíz-
Durán, E. Pérez-Negrón, A. Otero-Arnaiz, N.Pérez-Nasser, and A. Casas. 2006. Manejotradicional y conservación de la diversidadmorfológica y genética de Polaskia spp. (Cac-taceae) en México. Zonas Áridas 10:20–40.
Bravo-Hollis, H. 1978. Las cactáceas de México.Universidad Nacional Autónoma de México,México, Volumen I.
Casas, A., J. Caballero, and A. Valiente-Banuet.1999a. Use, Management and Domesticationof Columnar Cacti in South-central Mexico: AHistorical Perspective. Journal of Ethnobiol-ogy 19:71–95.
———, A. Valiente-Banuet, A. Rojas-Martínez,and P. Dávila. 1999b. Morphological Varia-tion and the Process of Domestication of theColumnar Cactus Stenocereus stellatus in Cen-tral Mexico. American Journal of Botany86:522–533.
———, ———, J. Viveros, J. Caballero, L.Cortés, P. Dávila, R. Lira, and I. Rodríguez.2001. Plant Resources of the Tehuacán-Cuicatlán Valley, Mexico. Economic Botany55(1):129–166.
386 ECONOMIC BOTANY [VOL 63

———, J. Cruse, E. Morales, A. Otero-Arnaiz,and A. Valiente-Banuet. 2006. Maintenanceof Phenotypic and Genotypic Diversity ofStenocereus stellatus (Cactaceae) by IndigenousPeoples in Central Mexico. Biodiversity andConservation 15:879–898.
———, A. Otero-Arnaiz, E. Pérez-Negrón, andA. Valiente-Banuet. 2007. In Situ Manage-ment and Domestication of Plants in Meso-america. Annals of Botany 100:1101–1115.
———, S. Rangel-Landa, I. Torres-García, E.Pérez-Negrón, L. Solís, F. Parra, A. Delgado,J. J. Blancas, B. Farfán, and A. I. Moreno.2008. In Situ Management and Conservationof Plant Resources in the Tehuacán-CuicatlánValley, Mexico: An Ethnobotanical and Eco-logical Perspective. Pages 1–25 in U. P. DeAlbuquerque and M. Alves-Ramos, eds., Cur-rent Topics in Ethnobotany. Research Sign-post, Kerala, India.
Cruz, M. and A. Casas. 2002. MorphologicalVariation and Reproductive Biology of Polas-kia chende (Cactaceae) under Domestication inCentral Mexico. Journal of Arid Environments51:561–576.
González-Insuasti, M. S. and J. Caballero. 2007.Managing Plant Resources: How IntensiveCan It Be? Human Ecology 35:303–314.
Guillén, S., A. Casas, J. Benítez, and M. Martínez.2009. Seed Germination of Wild, Managed inSitu and Cultivated Populations of ColumnarCacti in the Tehuacán-Cuicatlán Valley. Jour-nal of Arid Environments 73:407–413.
Hawkes, J. G. 1983. The Diversity of CropPlants. Harvard University Press, London.
JMP software Version 5.0 of the SAS System forMacintosh. Copyright © 1996. SAS Institute,Inc., Cary, North Carolina
Lira, R., A. Casas, R. Rosas, M. Paredes, E. Pérez-Negrón, S. Rangel-Landa, L. Solís, I. Torres,and P. Dávila. 2009. Traditional Knowledgeand Useful Plant Richness in the Tehuacán-CuicatlánValley, Mexico. Economic Botany63(3):271–287.
MacNeish, R. S. 1967. A Summary of Subsis-tence. Pages 290–309 in D. S. Byers, ed., ThePrehistory of the Tehuacán Valley. Vo1. 1.Environment and Subsistence. University ofTexas Press, Austin.
Nei, M. 1972. Genetic Distance between Popula-tions. The American Naturalist 106:283–292.
Oaxaca-Villa, B., A. Casas, and A. Valiente-Banuet. 2006. Reproductive Biology in Wildand Silvicultural Management Populations ofEscontria chiotilla (Cactaceae) in the Tehua-cánValley, Central México. Genetic Resourcesand Crop Evolution 53:277–287.
Ortíz, F., K. Stoner, E. Pérez-Negrón, and A.Casas. (in press). Pollination Biology ofMyrtillocactus schenckii (Cactaceae) in Wildand Managed Populations of the TehuacanValley, Mexico. Journal of Arid Environments.
Otero-Arnaiz, A., A. Casas, M. C. Bartolo, E.Pérez-Negrón, and A. Valiente-Banuet. 2003.Evolution of Polaskia chichipe (Cactaceae)under Domestication in the Tehuacan Valley,Central México: Reproductive Biology. Amer-ican Journal of Botany 90:593–602.
Otero-Arnaiz, A. C., J. L. Hamrick, and J. Cruse.2005. Genetic Variation and Evolution ofPolaskia chichipe (Cactaceae) under Domesti-cation in the Tehuacán Valley, Central Mex-ico, Analyzed by Microsatellite Polymorphism.Molecular Ecology 14:1603–1611.
Parra, F., N. Pérez-Nasser, D. Pérez-Salicrup, R.Lira, and A. Casas. 2008. Population Geneticsand Process of Domestication of Stenocereuspruinosus in the Tehuacán Valley, Mexico.Journal of Arid Environments 72:1997–2010.
Rodríguez-Arévalo, I., A. Casas, J. Campos, and R.Lira. 2006. Uso, manejo y proceso de domes-tication de Pachycereus hollianus (F.A.C. Weber)Buxb. (Cactaceae) en el Valle de Tehuacán-Cuicatlán, Mexico. Interciencia 31:677–685.
Rohlf, J. 1993. Numerical Taxonomy and Multi-variate Analysis System for the PC Micro-computer (and Compatibles). Version 2.6.Applied Biostatistics, Inc., New York.
Sneath, P. H. A. and R. R. Sokal. 1973.Numerical Taxonomy. The Principles andPractice of Numerical Classification, Freeman,San Francisco, California.
Tinoco, A., A. Casas, R. Luna, and K. Oyama.2005. Population Genetics of Wild andSilvicultural Managed Populations of Escontriachiotilla in the Tehuacán Valley, CentralMexico. Genetic Resources and Crop Evolu-tion 52:525–538.
387BLANCAS ET. AL: TRADITIONAL MANAGEMENT AND MORPHOLOGICAL PATTERNS2009]




















