
Toxicity of Functionalized Single-Walled Carbon Nanotubes on Soil Microbial Communities:...
9
Toxicity of Functionalized Single-Walled Carbon Nanotubes on Soil Microbial Communities: Implications for Nutrient Cycling in Soil Debora F. Rodrigues,* ,†,§ Deb P. Jaisi, ‡,§ and Menachem Elimelech § † Department of Civil and Environmental Engineering, University of Houston, Houston, Texas 77204-5003, United States ‡ Department of Plant and Soil Sciences, University of Delaware, Newark, Delaware 19716, United States § Department of Chemical and Environmental Engineering, Yale University, New Haven, Connecticut 06520-8286, United States * S Supporting Information ABSTRACT: Culture-dependent and -independent methods were employed to determine the impact of carboxyl-functionalized single-walled carbon nanotubes (SWNTs) on fungal and bacterial soil microbial communities. Soil samples were exposed to 0 (control), 250, and 500 μg of SWNTs per gram of soil. Aliquots of soil were sampled for up to 14 days for culture-dependent analyses, namely, plate count agar and bacterial community level physiological profiles, and culture- independent analyses, namely, quantitative real-time polymerase chain reaction (qPCR), mutliplex-terminal restriction fragment length polymorphism (M- TRFLP), and clone libraries. Results from culture-independent and -dependent methods show that the bacterial soil community is transiently affected by the presence of SWNTs. The major impact of SWNTs on bacterial community was observed after 3 days of exposure, but the bacterial community completely recovered after 14 days. However, no recovery of the fungal community was observed for the duration of the experiment. Physiological and DNA microbial community analyses suggest that fungi and bacteria involved in carbon and phosphorus biogeochemical cycles can be adversely affected by the presence of SWNTs. This study suggests that high concentrations of SWNTs can have widely varying effects on microbial communities and biogeochemical cycling of nutrients in soils. ■ INTRODUCTION The advancement in nanotechnology is leading to the production of novel materials that are used for diverse industrial products. Current estimates suggest that more than 1317 consumable products, including cell phone batteries, computer screens, sporting equipment, and cosmetics, already contain nanomaterials. 1 In fact, the global market that uses nanomaterials has grown steadily from U.S. $7.5 billion in 2003 to U.S. $12.7 billion in 2008 and is expect to exceed U.S. $27 billion over the next 5 years. 1 Among nanomaterials, carbon nanotubes (CNTs) are the most promising because of their numerous unique properties compared to bulk materials. 2 CNTs are used in many applications ranging from consumer electronics to drug delivery. 3 CNTs have great potential for new applications because of the endless possibilities that exist to functionalize CNT surfaces with exotic functional chemistries. Because of such potential for new applications and the likelihood for large- scale production, it is likely that CNTs will find their way into aquatic and terrestrial environments, potentially resulting in ecotoxicological effects. Knowledge on the impact of nanomaterials, including CNTs, on the environment is currently limited; hence, there is a critical need for safety regulations due to the potential use of nanomaterials at large industrial scales. 4,5 So far, the majority of studies on the toxicological effects of CNTs have been investigated mostly in pure cultures. 6,7 For instance, toxicity studies with Escherichia coli as a model microorganism showed that over 80% of the cells attached to nonfunctionalized single- walled carbon nanotube (SWNT) aggregates were dead after an incubation time of 60 min. 8 Gene expression analyses of the E. coli exposed to different types of CNTs showed a high expression of stress-related genes, corroborating the cytotoxic effects of CNTs on E. coli. In the environment, there are a handful of studies that investigated the toxic effects of CNTs on microbial communities. The majority of the studies were done in aquatic environments, 7,9,10 and only one study has investigated the toxicity of multiwalled carbon nanotubes on complex microbial communities in soil environments. 11 The soil ecosystem is likely to be the ultimate recipient for nanomaterials; 4 therefore, soil microorganisms could be significantly affected when exposed to nanomaterials. 12 Soil microbial communities play an important role in nutrient cycling and have been a sensitive indicator of soils’ response to environmental stressors, such as heavy metals and antimicrobial agents. 13−15 It is possible that once CNTs come in contact with soil, they can be retained within the soil profile and prolong the exposure to soil microorganisms. 16 Hence, the impact of CNTs on soil microbial ecology must be considered in light of recent results indicating their adverse effects on microorganisms. 11 Additionally, analysis of temporal impact of CNTs on soil Received: October 2, 2012 Revised: November 28, 2012 Accepted: December 3, 2012 Published: December 3, 2012 Article pubs.acs.org/est © 2012 American Chemical Society 625 dx.doi.org/10.1021/es304002q | Environ. Sci. Technol. 2013, 47, 625−633
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Toxicity of Functionalized Single-Walled Carbon Nanotubes on Soil Microbial Communities:...

Toxicity of Functionalized Single-Walled Carbon Nanotubes on SoilMicrobial Communities: Implications for Nutrient Cycling in SoilDebora F. Rodrigues,*,†,§ Deb P. Jaisi,‡,§ and Menachem Elimelech§
†Department of Civil and Environmental Engineering, University of Houston, Houston, Texas 77204-5003, United States‡Department of Plant and Soil Sciences, University of Delaware, Newark, Delaware 19716, United States§Department of Chemical and Environmental Engineering, Yale University, New Haven, Connecticut 06520-8286, United States
*S Supporting Information
ABSTRACT: Culture-dependent and -independent methods were employed todetermine the impact of carboxyl-functionalized single-walled carbon nanotubes(SWNTs) on fungal and bacterial soil microbial communities. Soil samples wereexposed to 0 (control), 250, and 500 μg of SWNTs per gram of soil. Aliquots ofsoil were sampled for up to 14 days for culture-dependent analyses, namely, platecount agar and bacterial community level physiological profiles, and culture-independent analyses, namely, quantitative real-time polymerase chain reaction(qPCR), mutliplex-terminal restriction fragment length polymorphism (M-TRFLP), and clone libraries. Results from culture-independent and -dependentmethods show that the bacterial soil community is transiently affected by the presence of SWNTs. The major impact of SWNTson bacterial community was observed after 3 days of exposure, but the bacterial community completely recovered after 14 days.However, no recovery of the fungal community was observed for the duration of the experiment. Physiological and DNAmicrobial community analyses suggest that fungi and bacteria involved in carbon and phosphorus biogeochemical cycles can beadversely affected by the presence of SWNTs. This study suggests that high concentrations of SWNTs can have widely varyingeffects on microbial communities and biogeochemical cycling of nutrients in soils.
■ INTRODUCTION
The advancement in nanotechnology is leading to theproduction of novel materials that are used for diverseindustrial products. Current estimates suggest that more than1317 consumable products, including cell phone batteries,computer screens, sporting equipment, and cosmetics, alreadycontain nanomaterials.1 In fact, the global market that usesnanomaterials has grown steadily from U.S. $7.5 billion in 2003to U.S. $12.7 billion in 2008 and is expect to exceed U.S. $27billion over the next 5 years.1
Among nanomaterials, carbon nanotubes (CNTs) are themost promising because of their numerous unique propertiescompared to bulk materials.2 CNTs are used in manyapplications ranging from consumer electronics to drugdelivery.3 CNTs have great potential for new applicationsbecause of the endless possibilities that exist to functionalizeCNT surfaces with exotic functional chemistries. Because ofsuch potential for new applications and the likelihood for large-scale production, it is likely that CNTs will find their way intoaquatic and terrestrial environments, potentially resulting inecotoxicological effects.Knowledge on the impact of nanomaterials, including CNTs,
on the environment is currently limited; hence, there is acritical need for safety regulations due to the potential use ofnanomaterials at large industrial scales.4,5 So far, the majority ofstudies on the toxicological effects of CNTs have beeninvestigated mostly in pure cultures.6,7 For instance, toxicitystudies with Escherichia coli as a model microorganism showed
that over 80% of the cells attached to nonfunctionalized single-walled carbon nanotube (SWNT) aggregates were dead after anincubation time of 60 min.8 Gene expression analyses of the E.coli exposed to different types of CNTs showed a highexpression of stress-related genes, corroborating the cytotoxiceffects of CNTs on E. coli. In the environment, there are ahandful of studies that investigated the toxic effects of CNTs onmicrobial communities. The majority of the studies were donein aquatic environments,7,9,10 and only one study hasinvestigated the toxicity of multiwalled carbon nanotubes oncomplex microbial communities in soil environments.11
The soil ecosystem is likely to be the ultimate recipient fornanomaterials;4 therefore, soil microorganisms could besignificantly affected when exposed to nanomaterials.12 Soilmicrobial communities play an important role in nutrientcycling and have been a sensitive indicator of soils’ response toenvironmental stressors, such as heavy metals and antimicrobialagents.13−15 It is possible that once CNTs come in contact withsoil, they can be retained within the soil profile and prolong theexposure to soil microorganisms.16 Hence, the impact of CNTson soil microbial ecology must be considered in light of recentresults indicating their adverse effects on microorganisms.11
Additionally, analysis of temporal impact of CNTs on soil
Received: October 2, 2012Revised: November 28, 2012Accepted: December 3, 2012Published: December 3, 2012
Article
pubs.acs.org/est
© 2012 American Chemical Society 625 dx.doi.org/10.1021/es304002q | Environ. Sci. Technol. 2013, 47, 625−633

microbial activity will provide new insights on how CNTs mayalter key ecosystem functions in soils.Most of the current studies on the toxicity of CNTs to
microbial communities employ conventional culture methods.Molecular biology methods have been developed rapidly overthe past decade, overcoming many of the problems associatedwith conventional culture methods.17 Nucleic acid-basedcommunity analysis has lately gained prominence because ofthe greater resolving power compared to other methods, suchas phospholipid fatty acid analysis (PLFA).18 Most of thesemolecular methods are polymerase chain reaction (PCR) basedand target the rRNA gene cluster. The combination of PCRamplification from soil DNA samples with fingerprintingtechniques, such as denaturing gradient gel electrophoresis(DGGE) and terminal restriction fragment length poly-morphism (TRFLP), provide detailed information about themicrobial composition of whole communities.The present study investigates the impact of SWNTs with
carboxyl functional groups on fungal and bacterial microbialcommunities in soil by culture-dependent and -independentmethods. Soil samples were exposed to high concentrations ofSWNTs (250 and 500 μg/g), which may represent worst-casescenarios of SWNT discharge (e.g., industrial spill).19 Ourresults show that the bacterial microbial community istransiently affected by the presence of SWNTs. Fungalcommunities, on the other hand, were found to be muchmore sensitive to the presence of SWNTs and did not recoverto their original cell concentration during the 14 daysexperiment course. The results demonstrate that different soilmicrobial communities have different sensitivities to thepresence of SWNTs, which may have direct implications forimportant biogeochemical processes in soils.
■ MATERIALS AND METHODSSoil Sample Collection and Characterization. Soil
samples were collected from a turf grass field in New Haven,CT (latitude 41.339 16°; longitude 72.904 44°) by use of sterilespatulas. The soil samples were stored in sterile whirl-pak bagsat 4 °C until being transferred to the laboratory as previouslydescribed.20 The sampling site has isolated large trees and doesnot present any known soil contamination or excessivemodification of the natural land environment. Soil texture,organic matter content, and particle size distribution weredetermined in the Soil Testing Laboratory at South DakotaState University.Preparation and Characterization of SWNTs. Commer-
cial carboxyl-functionalized SWNTs were purchased fromCheap Tubes, Inc. (Brattleboro, VT). SWNTs were fullycharacterized before use as previously described.16 Briefly, theSWNT diameter was calculated from Raman spectra (Jasco)collected at an excitation wavelength of 785 nm. The degree ofpurity was calculated from thermogravimetric analysis (TGA)(Setsys 16/18) by heating the SWNTs from 200 to 1000 °C ata rate of 10 °C/min. The metal content was also analyzed byusing scanning electron microscopy with energy-dispersive X-ray spectroscopy (SEM-EDX).Preparation of SWNT Stock Solution. The SWNT
suspension (2.0 g/L) was ultrasonicated (Misonix 3000,Misonix Inc., Farmingdale, NY) in deionized water for three30-min cycles. Between each sonication cycle, the larger SWNTaggregates were allowed to settle for 30 min and the well-suspended SWNTs were transferred to a new vial andultrasonicated again for 30 min. This cycle was repeated 3
times to yield a very well dispersed solution of SWNTs. Thedispersed SWNTs from the third cycle, which were stable insolution for several weeks, were used as a stock solution for thesubsequent toxicity experiments.16 The concentration of theSWNT stock solution was determined gravimetrically on anultramicrobalance (Mettler Toledo MX5) with a repeatabilityof 2.5 μg.
Soil Exposure to SWNTs. The collected soil washomogenized in a sterile container by use of sterile spatulas,after removal of pebble grains and roots, and then was saturatedwith sterile deionized (DI) water. The soil was divided into 9flasks with 40 g each to obtain triplicate samples to be exposedto three different concentrations of SWNTs [0 (control), 250,and 500 μg/g]. A specific amount of SWNTs was addeddirectly to all flasks except the control flasks, where only DIwater was added. The soils were then mixed for 20 min beforeincubation at 25 °C. Throughout the experiments, the soilsamples were kept saturated by addition of sterile DI water.Aliquots of soil were collected at different time points (0, 3, and24 h and 3, 7, and 14 days) from each flask to be used for DNAextraction, total heterotrophic plate counts, and community-level physiological profile (CLPP) analyses.
DNA Extraction and Purification. For the DNAextraction procedure, one aliquot of 0.5 g of soil was collectedat each time point from each flask. The total communitygenomic DNA was extracted with the PowerSoil DNA isolationkit from MoBio Laboratory, Inc. (Carlsbad, CA) as describedby the manufacturer. The quality of the extracted DNA wasanalyzed by electrophoresis on a 1% agarose gel. DNAconcentrations were measured by absorbance at 260 nm.17
Enumeration of Total Soil Bacterial Community byPlate. For the enumeration of microorganisms in the soil, anagar medium containing soil extract was prepared to maintainsimilar nutrient conditions for the microbial community, aspreviously described.21 Briefly, the soil (500 g) was ground anddispersed with a mortar and pestle. The soil was mixed with 1 Lof sterile water for 15 min and then autoclaved (121 °C for 15min). After autoclaving, an additional 500 mL of sterile waterwas added and the mixture was mixed for 15 min, followed bycentrifugation at 8000 rpm for 30 min. Since the solution wasstill cloudy after the centrifugation, 1 g of calcium carbonate(CaCO3) was added to the suspension and allowed to react for5 min, and then the suspension was centrifuged again. The agarmedium was prepared from 500 mL of soil extract, 500 mL ofwater, 1 g of glucose, 0.5 g of yeast extract, 0.5 g of K2HPO4,and 15 g of agar. The mixed medium was autoclaved for 15 minand plated in sterile Petri dishes.After the medium was prepared for the experiment, the plates
were used to determine the total microbial community from thesoil exposed to different concentrations of SWNTs. Twoaliquots of 1 g from the soil samples exposed to the differentSWNT concentrations were diluted in 9 mL of 1× PBS(phosphate-buffered saline; 1 mM KH2PO4, 1 mM Na2HPO4,2.7 mM KCl, and 13.7 mM NaCl at pH 7.4). The mixture wasvortexed for 20 min, then diluted through a serial dilution andplated by the spread plate method. In total, there were 6replicates for each SWNT concentration. All plates wereincubated for 1 week at 25 °C, which was close to thetemperature where the soils were collected. Plates werequantified for all samples exposed for 0, 3, and 24 h and 3, 7,and 14 days to the different concentrations of SWNTs (0, 250,and 500 μg/g).
Environmental Science & Technology Article
dx.doi.org/10.1021/es304002q | Environ. Sci. Technol. 2013, 47, 625−633626

Quantification of Bacteria and Fungi MicrobialCommunities by Real-Time PCR. For relative quantificationof microbial communities in the soil, real-time polymerasechain reaction (qPCR) was used as previously described byRodrigues and Tiedje.17 Briefly, the increase in fluorescenceemission was monitored during PCR amplification by the 7700sequence detector (PE Applied Biosystems). The Ct valuesobtained for each sample were compared with the standardcurve to determine the initial copy number of the target gene.For eubacteria, Pseudomonas stutzeri was used to build thestandard curve, since this microorganism is commonly found insoil and has five 16SrRNA gene copies (which is estimated tobe the average 16S copy number of soil microorganisms insoil).22,23 For fungi, a clone containing both the amplifiedinternal transcribed spacer (ITS) and 5.8S obtained from thesoil was used to build the standard curve. The primers used inthe qPCR were E8F (5′-AGAGTTTGATCCTGGCTCAG-3′)and E533R (5′-TIACCGIICTICTGGCAC-3′) for the 16SrRNA gene to quantify eubacterial communities24 and ITS3F(5′-GCATCGGATGAAGAACGCAGC-3′) and ITS4R (5′-TCCTCCGCTTATTGATATGC-3′) for the 5.8S and ITS2regions to quantify the fungal community.25 The qPCRreactions and primer optimizations were done as describedby Rodrigues and Tiedje.17 The reagents used for the reactionwere the 2× PCR master mix (Applied Biosystems) withaddition of 0.4 mg·mL−1 final concentration bovine serumalbumin (Roche). A primer concentration of 300 nM was usedfor all primers. The annealing and extension temperatures forthe bacterial and fungal primers were 55 and 55.3 °C,respectively. The sizes of the amplicons were 525 and 937 bpfor bacteria and fungi, respectively. The cycle number wherethe fluorescence signal crossed the threshold, threshold (Ctvalue) was used to calculate relative amounts of originaltemplate using the formula 1/eCt as previously described.24
Assessment of Microbial Community Structures byMultiplex-TRFLP Analysis. The multiplex- (M-) TRFLP wasdone as previously described.25 Briefly, the eubacterial DNAsamples were amplified with the primers 63F (5′-CAGGCC-TAACACATGCAAGTC-3′) and 1087R labeled with VICfluorophore, VIC-(5′-CTCGTTGCGGGACTTACCCC-3′).Similarly, ITS1F primer labeled with 6-carboxyfluorescein,FAM-(5′-CTTGGTCATTTAGAGGAAGTAA-3′), and ITS4R(3′-TCCTCCGCTTATTGATATGC-5′) were used forfungi.25 Fifty microliter reactions were performed in triplicatefor each sample. The reactions contained 0.02 unit·μL−1 TaqDNA polymerase, 1× reaction buffer, 1.5 mM MgCl2, 0.2 mMeach dNTP, 0.2 μM each primer, 400 ng·μL−1 bovine serumalbumin (BSA) and 2 ng·μL−1 template. The followingconditions were chosen for amplification after optimization:initial denaturation for 5 min at 95 °C; 30 cycles with 30 s at 95°C for denaturation, 30 s at 55 °C for annealing, and 1 min at72 °C for extension; and final extension for 10 min at 72 °C.Triplicates were combined and purified with the Qiagen PCRpurification kit after gel examination. Prior to digestion, purifiedPCR product concentrations were determined with aspectrophotometer at 260 nm. All PCR products obtainedfrom PCRs were digested separately with the HhaI and MspIrestriction enzymes in 20 μL reaction mixtures containing 500ng of PCR products, 1× buffer, 0.1 μg·μL−1 acetylated BSA, and20 units of restriction enzyme (all reagents were obtained fromPromega, Southampton, U.K.). Samples were incubated at 37°C for 3 h, followed by deactivation at 95 °C for 15 min. Afterthat, the DNA was precipitated overnight in a solution
containing 1.5 μL of 3 M sodium acetate, 0.3 μL of glycogen(20 mg·μL−1), and 42 μL of absolute ethanol. The sampleswere then centrifuged for 35 min at 14 000 rpm and thesupernatant was discarded. The precipitated DNA was washedwith 70% ethanol for 5−10 min, centrifuged for 20 min, andthen vacuum-dried. The DNA pellets were resuspended in 9.85μL of deionized formamide. ROX size standard (0.15 μL) wasadded 15 min before analysis in the ABI system analyzer.24 AllTRFLP analyses were performed at the RTSF facility atMichigan State University. For the identification and sizing ofthe restriction fragments generated, the Peak Scanner program(Applied Biosystems) was used. After identification of therestriction fragments and their intensity with the Peak Scanner,the triplicate samples for each condition were analyzed in Excel,and only peaks that were present in at least 2 replicates wereused for the subsequent analyses. The statistical analysis wasperformed with the web-based tool T-REX (http://trex.biohpc.org/).26 In this program, the terminal restriction fragments(TRFs) were aligned and a matrix was generated to determinethe variance between main and interaction effects as well assample heterogeneity, then additive main effects and multi-plicative interaction model (AMMI) were employed to analyzethe data.
Bacterial Community-Level Physiological Profiles. ForCLPP analysis, Eco-plates were used. The Eco-plates aredesigned for intraplate replication, as each 96-well plate forEco-plate provides 31 different carbon substrates in a triplicatearray. The Biolog Eco-plate technique relies on measurementsof different carbon substrates utilization by bacteria. Fungi donot metabolize the tetrazolium in the Eco-plates wells, andtherefore they do not contribute to the microbial “metabolicfingerprint” on these plates.27 In this present analysis, to assessthe changes of bacterial communities exposed to differentSWNT concentrations, we used an aliquot (1 g) of soilcollected at different time periods (0, 3, and 24 h and 3, 7, and14 days), as described earlier. The sample was thoroughlymixed with 9 mL of 1× PBS by vortexing for 10 min28 and thenallowed to settle for 15 min. Supernatant was diluted to 1:50 in1× PBS and inoculated in Eco-Plates for bacterial communitydetermination. The soil sample solution prepared above wasvortexed, the surpenatant was poured into a sterile Petri dish,and 100 μL of the solution was added to each well. Theprepared plate was then immediately incubated at 25 °C andthe absorbance was read on a Spectramax 340 plate reader(Molecular Devices Corp., San Jose,́ CA) at 590 nm at time 0and thereafter once a day for 14 days. All wells with change incolor or turbidity were considered positive for growth. We onlyconsidered positive if at least two wells containing the samesubstrate presented growth. The statistical analysis of Biologdata included differences in color development among samplesand the average well color development (AWCD) method.29
The AWCD was calculated according to Garland and Mills,29
that is, AWCD = ∑(C − R)/n, where C is the optical densitymeasured within each well, R is the absorbance value of thecontrol well, and n is the number of substrates (Eco-plates, n =31). For statistical analyses of the data, principal componentanalyses (PCA) were performed. Prior to the statistical analysis,the data were transformed by dividing, for each substrate, thedifference in optical density at 120 h, relative to the controlwell, by the AWCD of the plate at 120 h, that is, (C − R)/AWCD.29 Principal component analysis (PCA) was performedon the transformed AWCD after normalization of the data withBOX-COX transformation statistical tool in the XLSTAT
Environmental Science & Technology Article
dx.doi.org/10.1021/es304002q | Environ. Sci. Technol. 2013, 47, 625−633627

program (Addinsoft Inc.).20 PCA analysis projects original dataonto new, statistically independent axes (principal compo-nents). Each principal component extracts a portion of thevariance from the original data. Relationships among sampleswere obtained by plotting scores of their first two principalcomponents in two dimensions.Assessment of Effects of SWNTs on Different Groups
of Soil Microorganisms. The same primers utilized for theqPCR experiments were used to generate the clone libraries forboth bacteria and fungi. DNA samples from three soil replicatestaken after 3 day exposure to SWNTs were combined togenerate the clone libraries. This time period was selected onthe basis of the highest impact of SWNTs on the microbialcommunity. The clone libraries were generated as previouslydescribed by Rodrigues et al.30 Briefly, the specific geneamplicons for bacteria and fungi were cloned using the TOPOTA cloning kit for sequencing (Invitrogen Life Technologies)and sent to Macrogen Inc. (Seoul, Korea; www.Macrogen.com)for sequencing with primers for the cloning vector (M13F).The 16S rRNA gene sequences from the clone libraries wereprocessed using the pipeline quality filter tools on theRibosomal Database Project (RDP-II) Web site (http://rdp.cme.msu.edu).31 The Classifier tool provided by RDP-II wasused to assign the 16S rRNA gene sequences to thetaxonomical hierarchy. The aligned sequences were derepli-cated by uploading into the FastGroupII program32 to clustersequences with 99% similarity including gaps. For the fungi, wefollowed the cloning procedures and data analyses as previouslydescribed.33,34 Briefly, the sequences obtained were alignedwith BioEdit V7.1.7 program, and the aligned sequences werealso dereplicated by the FastGroupII program. Operationaltaxonomic units (OTUs) were defined as a group of sequencessharing at least 97% pairwise similarity, which is within therange of intraspecific ITS sequence divergence. Representativesequences for each OTU were queried against GenBank withthe BLAST tool. Each sequence was identified to the lowesttaxonomic rank common to all of the top BLAST hitsaccording to the taxonomy of Kirk et al.35 The uniquesequences for fungi and bacteria were deposited in the NCBIdatabase (Accession Numbers JX545155−JX545204).
■ RESULTS AND DISCUSSIONSoil and SWNT Properties. The soil was composed of
sand, slit, and clay at 54%, 30%, and 16%, respectively, andclassified as sandy loam based on the USDA soil triangleclassification. The soil appeared gray-brown in color and wasrelatively poor in organic matter (0.5 wt %). The functionalizedSWNTs had diameters ranging from 0.9 to 1.44 nm asdetermined from the Raman spectra breathing mode.16 TGAand EDX analyses indicated a metal content of 5.04%, withcopper and iron as the major metals. Similarly, the ratio of D toG bands in the Raman spectra was about 0.45, suggesting highlevels of defects in the nanotubes, which is consistent withother functionalized SWNTs.16
Impact of SWNTs on Soil Microbial CommunityAbundance. Past studies have investigated the impact ofSWNTs on microorganisms under pure culture conditions,indicating that SWNTs are toxic to bacteria.6−8 There are onlytwo published studies on the effects of CNTs on nativemicrobial communities in the environmental. One studyinvestigated the effects of multiwalled carbon nanotubes onsoil microbial communities,11 and the other study wasperformed with single-walled carbon nanotubes in contact
with activated sludge microbial communities.9 Both studiespresented some impact of CNTs on the microbial commun-ities, depending on the concentration of CNTs. However, nostudies so far have addressed the effects of SWNTs on soilmicrobial communities by use of both conventional andculture-independent methods.Both culture-dependent and -independent methods are very
useful tools in determining the effects of certain contaminantson microbial communities; however, either approach has itsown limitation. Hence, to overcome their individual deficien-cies, we combined culture-dependent and -independentapproaches to determine the overall changes in microbialabundance and function after their exposure to differentconcentrations of functionalized SWNTs. In this investigation,three replicates for each SWNT concentration were tested tocover various microenvironments in the soil system aspreviously described in various studies with whole cell countingexperiments36,37 or biomass measurements,38 as well as specificbacterial enumerations.39,40 In these studies, it was demon-strated that the soil system, at microscale, has microenviron-ments differing in size, stability, microbial populations, andphysical and chemical properties.41−44 Despite our efforts tomix well the soil before division of the bulk soil into aliquots tobe tested with different concentrations of SWNTs, the numberof microbial populations in each aliquot still presented somevariation, as observed in Figures 1 and 2. However, the initial
control of each aliquot without the nanomaterials served as abaseline for comparisons on the changes of microbialcommunities over time for each SWNT concentration.The results for the culture-dependent method (plate count)
also showed that the impact of SWNTs varied with the SWNTconcentration (Figure 1). A similar dose-dependency effect onsoil microbial activity was also observed with multiwalledcarbon nanotubes (MWNTs).11 The highest concentration ofSWNTs (500 μg) used in this study presented the highestimpact on the microbial community (Figures 1 and 2). We notethat the concentration of SWNTs used in the present study was10 times less than that of the study with MWNTs11 but still
Figure 1. Plate counts in agar plates to quantify the total bacterialcommunity in soils exposed to 250 or 500 μg of SWNTs per gram ofsoil for different time periods (3 or 24 h or 3, 7, or 14 days). Thecontrol (soil with no SWNTs) is also presented. The results representthe averages and standard deviations of 6 replicates incubated at 25 °Cfor 10 days. Only soils exposed to 250 and 500 μg of SWNTs for 24 hand 3 days presented statistically significant different results in the t-test (p-value <0.05) than their respective control samples.
Environmental Science & Technology Article
dx.doi.org/10.1021/es304002q | Environ. Sci. Technol. 2013, 47, 625−633628
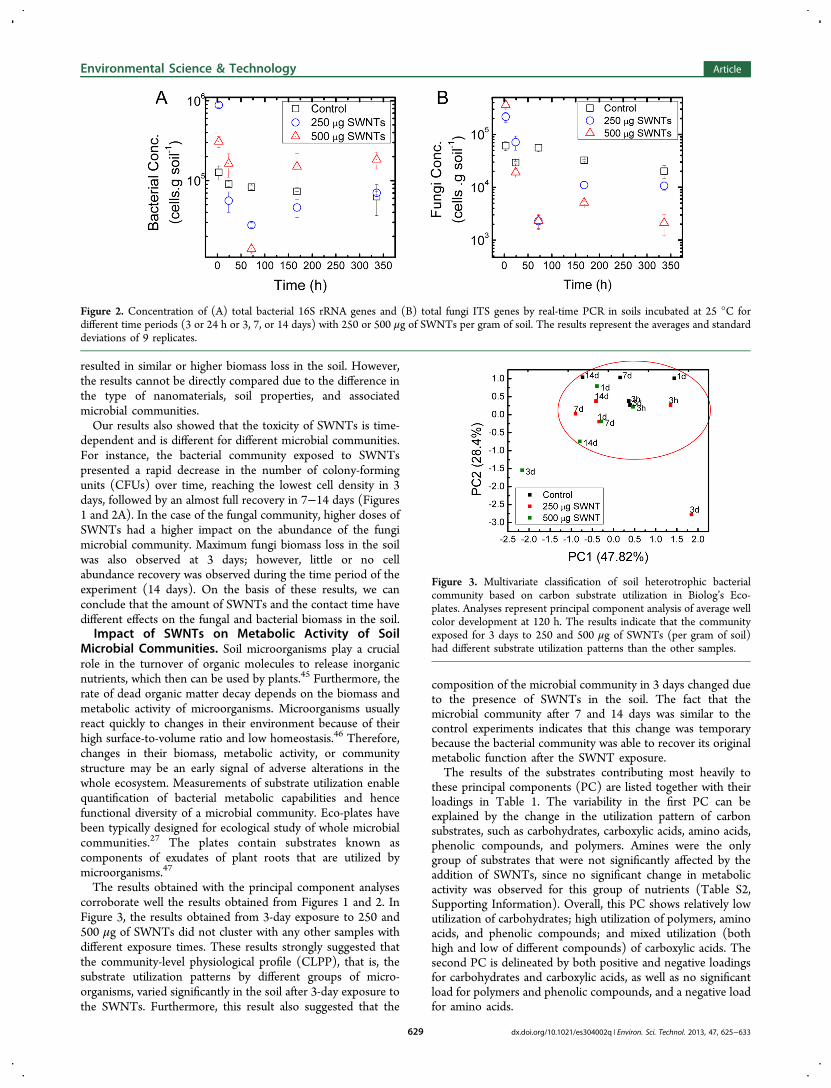
resulted in similar or higher biomass loss in the soil. However,the results cannot be directly compared due to the difference inthe type of nanomaterials, soil properties, and associatedmicrobial communities.Our results also showed that the toxicity of SWNTs is time-
dependent and is different for different microbial communities.For instance, the bacterial community exposed to SWNTspresented a rapid decrease in the number of colony-formingunits (CFUs) over time, reaching the lowest cell density in 3days, followed by an almost full recovery in 7−14 days (Figures1 and 2A). In the case of the fungal community, higher doses ofSWNTs had a higher impact on the abundance of the fungimicrobial community. Maximum fungi biomass loss in the soilwas also observed at 3 days; however, little or no cellabundance recovery was observed during the time period of theexperiment (14 days). On the basis of these results, we canconclude that the amount of SWNTs and the contact time havedifferent effects on the fungal and bacterial biomass in the soil.Impact of SWNTs on Metabolic Activity of Soil
Microbial Communities. Soil microorganisms play a crucialrole in the turnover of organic molecules to release inorganicnutrients, which then can be used by plants.45 Furthermore, therate of dead organic matter decay depends on the biomass andmetabolic activity of microorganisms. Microorganisms usuallyreact quickly to changes in their environment because of theirhigh surface-to-volume ratio and low homeostasis.46 Therefore,changes in their biomass, metabolic activity, or communitystructure may be an early signal of adverse alterations in thewhole ecosystem. Measurements of substrate utilization enablequantification of bacterial metabolic capabilities and hencefunctional diversity of a microbial community. Eco-plates havebeen typically designed for ecological study of whole microbialcommunities.27 The plates contain substrates known ascomponents of exudates of plant roots that are utilized bymicroorganisms.47
The results obtained with the principal component analysescorroborate well the results obtained from Figures 1 and 2. InFigure 3, the results obtained from 3-day exposure to 250 and500 μg of SWNTs did not cluster with any other samples withdifferent exposure times. These results strongly suggested thatthe community-level physiological profile (CLPP), that is, thesubstrate utilization patterns by different groups of micro-organisms, varied significantly in the soil after 3-day exposure tothe SWNTs. Furthermore, this result also suggested that the
composition of the microbial community in 3 days changed dueto the presence of SWNTs in the soil. The fact that themicrobial community after 7 and 14 days was similar to thecontrol experiments indicates that this change was temporarybecause the bacterial community was able to recover its originalmetabolic function after the SWNT exposure.The results of the substrates contributing most heavily to
these principal components (PC) are listed together with theirloadings in Table 1. The variability in the first PC can beexplained by the change in the utilization pattern of carbonsubstrates, such as carbohydrates, carboxylic acids, amino acids,phenolic compounds, and polymers. Amines were the onlygroup of substrates that were not significantly affected by theaddition of SWNTs, since no significant change in metabolicactivity was observed for this group of nutrients (Table S2,Supporting Information). Overall, this PC shows relatively lowutilization of carbohydrates; high utilization of polymers, aminoacids, and phenolic compounds; and mixed utilization (bothhigh and low of different compounds) of carboxylic acids. Thesecond PC is delineated by both positive and negative loadingsfor carbohydrates and carboxylic acids, as well as no significantload for polymers and phenolic compounds, and a negative loadfor amino acids.
Figure 2. Concentration of (A) total bacterial 16S rRNA genes and (B) total fungi ITS genes by real-time PCR in soils incubated at 25 °C fordifferent time periods (3 or 24 h or 3, 7, or 14 days) with 250 or 500 μg of SWNTs per gram of soil. The results represent the averages and standarddeviations of 9 replicates.
Figure 3. Multivariate classification of soil heterotrophic bacterialcommunity based on carbon substrate utilization in Biolog’s Eco-plates. Analyses represent principal component analysis of average wellcolor development at 120 h. The results indicate that the communityexposed for 3 days to 250 and 500 μg of SWNTs (per gram of soil)had different substrate utilization patterns than the other samples.
Environmental Science & Technology Article
dx.doi.org/10.1021/es304002q | Environ. Sci. Technol. 2013, 47, 625−633629

The apparent resistance and greater survival of communitiesthat can degrade phenolic compounds and complex polymersagrees with microbes that can tolerate, and in some cases candegrade, high loads of pollutants, such as polyaromatichydrocarbons and polychlorinated biphenyls.30,48,49 It ispossible that the recovery of the microbial community after14 days could be triggered by biological degradation of SWNTsto a less toxic material, as previously observed with anothercarbon-based nanomaterial (C60-fullerols).
50 Another possibil-ity for this recovery could be that the soil microorganisms haveother methods of mitigating toxicity, for example, production ofintracellular or extracellular materials for overcoming reactiveoxygen species produced by the nanomaterials, cell repairmechanisms, or even microbial community shifts toward moreresistant organisms.8 However, further experiments are neededto test these hypotheses.Impact of SWNTs on Soil Microbial Community
Structure. In the present study, TRFLP analysis was used todetermine unique operational taxonomic units (OTU) in eachsample.51,52 In our analysis, after identification of each OTU, itwas observed that the diversity of both bacterial and fungal soilcommunities changed with the addition of SWNTs. Interest-ingly though, for all SWNT concentrations tested, the bacterialcommunity after 14 days presented very similar microbialcommunities since they were clustered together. This resultindicates that the bacterial community recovered after 14 days(Figure 4A). In the case of the fungal soil community, thepopulations of fungi changed drastically with the addition ofSWNTs and remained different even after 14 days. This resultindicates that the fungal soil community is more sensitive to the
presence of SWNTs and does not recover easily from theexposure to SWNTs.The specific bacterium and fungus populations that were
most heavily affected by the presence of SWNTs wereinvestigated through clone libraries after 3 days of exposureto SWNTs. The results (Figure 5) show that some populationsof bacteria and even fungi remained unaffected by the presenceof SWNTs; that is, they did not present a population change ofmore than 5% when compared to the control sample. In thecase of bacteria, the majority of the groups were unaffected,except for genera of the phyla Cyanobacteria, Chloroflexi, andActinobacteria that showed higher than 5% change comparedto the control sample (Figure 5A). In the case of fungi, thegenera Pseudeurotium, Penicillium, and Eupenicillium were themost affected by the presence of SWNTs (Figure 5B).The bacteria of Chloroflexi and Cyanobacteria phyla are
mainly photosynthetic microorganisms and are commonlyfound in soil environments.53,54 These microorganisms havebeen described to influence the CO2 dynamics in soil as theyplay an important role in the carbon biogeochemical cycling insoils.54,55 In the case of Actinobacteria and Penicillium, they arecommonly described as microorganisms responsible for thedegradation of cellulolytic and hemicellulolytic compounds insoils.56,57 These groups of microorganisms are also consideredimportant in the carbon biogeochemical cycling in soils.44,45
These clone library results seem to be consistent with the
Table 1. Results of the Graphical Analysis Presented inFigure 3 Obtained from the Eco-plate Dataa
substrate PC1 PC2
Carbohydratemethyl β-D-glucoside −0.84D-galactonic acid γ-lactone −0.61D-mannitol −0.81N-acetyl-D-glucosamine −0.77D-cellobiose −0.62glucose 1-phosphate −0.71D-xylose −0.96D-galacturonic acid −0.81i-erythritol 0.71
Carboxylic Aciditaconic acid −0.70α-ketobutyric acid 0.60 0.55pyruvic acid methyl ester −0.56D-glucosaminic acid −0.59 0.57
Amino Acidsglycyl-L-glutamic acid 0.75L-phenylalanine 0.52L-serine −0.65L-asparagine −0.70
Phenolic Compounds2-hydroxybenzoic acid 0.57
PolymersTween 80 0.59α-cyclodextrin 0.92glycogen 0.59
aThe carbon substrates significantly utilized on the first two principalcomponents are described as PC1 and PC2.
Figure 4. Results of the additive main effects and multiplicativeinteraction model (AMMI) analysis of T-RFLP data sets of (A)bacteria and (B) fungi in soil samples exposed to different SWNTconcentrations for different time periods. The model uses analysis ofvariance (ANOVA) to first partition the variation into main effects andinteractions, and then it applies principal component analysis (PCA)to the interactions to create interaction principal components axes(IPCAs).
Environmental Science & Technology Article
dx.doi.org/10.1021/es304002q | Environ. Sci. Technol. 2013, 47, 625−633630

CLPP results that showed that the bacterial communityinvolved in the metabolism of cellulose and hemicellulosecarbohydrates such as D-cellobiose and D-xylose were affected inthe presence of SWNTs (Table 1). The results show that boththe culture-independent and -dependent methods indicate thatthe presence of SWNTs can affect the degradation of plantbiomass in soil. A similar effect on carbon biogeochemicalcycling was observed for soil microbial communities exposed toMWNTs.11
The genera Penicillium, Pseudeurotium, and Eupenicillium, onthe other hand, are also commonly described as being capableof solubilizing inorganic or hydrolyzing organic phospho-rus.58,59 Phosphorus is an essential nutrient for plant andmicroorganisms, and its availability largely depends on the rateof its solubilization from phosphate minerals or hydrolysis oforganic phosphorus compounds by microorganisms. Similarly,in a MWNT study with soil communities, it was demonstratedthat, in the presence of MWNTs, the phosphorus biogeochem-ical cycling was reduced.11 Our present study complements theMWNT results, in the sense that it shows that both SWNTsand MWNTs have negative impacts on soil nutrient cycling.Additionally, the present study shows that SWNTs seem to bemore toxic than MWNTs in the soil, since the concentration ofSWNTs, which was 10 times less, induced similar soil stressesas the MWNTs. These findings agree with previous pureculture studies with SWNTs and MWNTs, which show thatSWNTs are more toxic than MWNTs.8,10,11
Implications. Physicochemical and biological processes actin concert in soils across time and space to maintain a balancedecosystem. Certain groups of biota can have a profound effecton the cycling of biogeochemically reactive elements (macro-nutrients such as carbon, nitrogen, sulfur, and phosphorus aswell as trace elements vital for life). The presence of foreignmaterials in the ecosystem can have a detrimental effect if theyimpact microorganisms that are involved in nutrient or other
major element cycling in soil. Our results indicate that thepresence of SWNTs in soils impact microorganisms that arelargely related to carbon and phosphorus cycling. The temporalchange of this impact, however, was different among differentgroups of microorganisms. For example, it was transient for thebacterial community, as they recovered over time (within 14days) from the impact of SWNTs. However, the impact on thefungal soil community appeared to be long-lasting. Our resultsfrom both culture-dependent and -independent methodscorroborated well the SWNT impact observed in the soilcommunities; however, the impact appeared to be morepronounced in the latter method. Therefore, selection ofspecific research methods and analysis of targeted communitiesof microorganisms such as those related to nutrient cycling areimportant to better understand the impact of nanomaterials inthe environment.
■ ASSOCIATED CONTENT
*S Supporting InformationTwo tables, listing unpaired t-test statistical analyses of Figures1 and 2A and results of all substrate loading values calculatedfor both PCA 1 and PCA 2 presented in Figure 3; and onefigure, showing rarefaction curves for the fungi and eubacterialclone libraries. This material is available free of charge via theInternet at http://pubs.acs.org/.
■ AUTHOR INFORMATION
Corresponding Author*E-mail: [email protected].
NotesThe authors declare no competing financial interest.
■ ACKNOWLEDGMENTS
We acknowledge the National Science Foundation underResearch Grant CBET-0828795.
■ REFERENCES(1) Rejeski, D. Project on Emerging Nanotechnologies; Bethesda, MD,2009; p 12.(2) Odom, T. W.; Huang, J.-L.; Kim, P.; Lieber, C. M. Atomicstructure and electronic properties of single-walled carbon nanotubes.Nature 1998, 391, 62−64.(3) Grobert., N. Carbon nanotubes: Becoming clean. Mater. Today2007, 10, 28−35.(4) Nowack, B.; Bucheli, T. D. Occurrence, behavior and effects ofnanoparticles in the environment. Environ. Pollut. 2007, 150 (1), 5−22.(5) Klaine, S. J.; Alvarez, P. J.; Batley, G. E.; Fernandes, T. F.; Handy,R. D.; Lyon, D. Y.; Mahendra, S.; McLaughlin, M. J.; Lead, J. R.Nanomaterials in the environment: behavior, fate, bioavailability, andeffects. Environ. Toxicol. Chem./SETAC 2008, 27 (9), 1825−1851.(6) Ahmed, F.; Santos, C. M.; Vergara, R. A.; Tria, M. C.; Advincula,R.; Rodrigues, D. F. Antimicrobial applications of electroactive PVK-SWNT nanocomposites. Environ. Sci. Technol. 2012, 46 (3), 1804−1810.(7) Rodrigues, D. F.; Elimelech, M. Toxic effects of single-walledcarbon nanotubes in the development of E. coli biofilm. Environ. Sci.Technol. 2010, 44 (12), 4583−4589.(8) Kang, S.; Herzberg, M.; Rodrigues, D. F.; Elimelech, M.Antibacterial effects of carbon nanotubes: size does matter! Langmuir2008, 24 (13), 6409−6413.(9) Goyal, D.; Zhang, X. J.; Rooney-Varga, J. N. Impacts of single-walled carbon nanotubes on microbial community structure inactivated sludge. Lett. Appl. Microbiol. 2010, 51 (4), 428−435.
Figure 5. (A) Bacterial and (B) fungal community distributions in soilsamples exposed to different SWNT concentrations (per gram of soil)after 3 days. Each subdivision of the bars corresponds to thepercentage of microorganisms present at different SNWT exposure.
Environmental Science & Technology Article
dx.doi.org/10.1021/es304002q | Environ. Sci. Technol. 2013, 47, 625−633631

(10) Kang, S.; Mauter, M. S.; Elimelech, M. Microbial cytotoxicity ofcarbon-based nanomaterials: implications for river water and waste-water effluent. Environ. Sci. Technol. 2009, 43 (7), 2648−2453.(11) Chung, H.; Son, Y.; Yoon, T. K.; Kim, S.; Kim, W. The effect ofmulti-walled carbon nanotubes on soil microbial activity. Ecotoxicol.Environ. Saf. 2011, 74 (4), 569−575.(12) Navarro, E.; Baun, A.; Behra, R.; Hartmann, N. B.; Filser, J.;Miao, A. J.; Quigg, A.; Santschi, P. H.; Sigg, L. Environmental behaviorand ecotoxicity of engineered nanoparticles to algae, plants, and fungi.Ecotoxicology 2008, 17 (5), 372−386.(13) Yang, Y.; Campbell, C. D.; Clark, L.; Cameron, C. M.; Paterson,E. Microbial indicators of heavy metal contamination in urban andrural soils. Chemosphere 2006, 63 (11), 1942−1952.(14) Liu, F.; Ying, G. G.; Yang, L. H.; Zhou, Q. X. Terrestrialecotoxicological effects of the antimicrobial agent triclosan. Ecotoxicol.Environ. Saf. 2009, 72 (1), 86−92.(15) Madsen, E. L. Microorganisms and their roles in fundamentalbiogeochemical cycles. Curr. Opin. Biotechnol. 2011, 22 (3), 456−464.(16) Jaisi, D. P.; Saleh, N. B.; Blake, R. E.; Elimelech, M. Transport ofsingle-walled carbon nanotubes in porous media: filtration mecha-nisms and reversibility. Environ. Sci. Technol. 2008, 42 (22), 8317−8323.(17) Rodrigues, D. F.; Tiedje, J. M. Multi-locus real-time PCR forquantitation of bacteria in the environment reveals Exiguobacterium tobe prevalent in permafrost. FEMS Microbiol. Ecol. 2007, 59 (2), 489−499.(18) Singh, B. K.; Millard, P.; Whiteley, A. S.; Murrell, J. C.Unravelling rhizosphere−microbial interactions: opportunities andlimitations. Trends Microbiol. 2004, 12 (8), 386−393.(19) Petersen, E. J.; Pinto, R. A.; Landrum, P. F.; Weber, W. J., Jr.Influence of carbon nanotubes on pyrene bioaccumulation fromcontaminated soils by earthworms. Environ. Sci. Technol. 2009, 43(11), 4181−4187.(20) Rodrigues, D. F.; da C Jesus, E.; Ayala-del-Río, H. L.; Pellizari,V. H.; Gilichinsky, D.; Sepulveda-Torres, L.; Tiedje, J. M.Biogeography of two cold-adapted genera: Psychrobacter andExiguobacterium. ISME J. 2009, 3 (6), 658−665.(21) Hamaki, T.; Suzuki, M.; Fudou, R.; Jojima, Y.; Kajiura, T.;Tabuchi, A.; Sen, K.; Shibai, H. Isolation of novel bacteria andactinomycetes using soil-extract agar medium. J. Biosci. Bioeng. 2005,99 (5), 485−492.(22) Klappenbach, J. A.; Dunbar, J. M.; Schmidt, T. M. rRNA operoncopy number reflects ecological strategies of bacteria. Appl. Environ.Microbiol. 2000, 66 (4), 1328−1333.(23) Gruntzig, V.; Nold, S. C.; Zhou, J.; Tiedje, J. M. Pseudomonasstutzeri nitrite reductase gene abundance in environmental samplesmeasured by real-time PCR. Appl. Environ. Microbiol. 2001, 67 (2),760−768.(24) Manerkar, M. A.; Seena, S.; Barlocher, F. Q-RT-PCR forassessing archaea, bacteria, and fungi during leaf decomposition in astream. Microb. Ecol. 2008, 56 (3), 467−473.(25) Singh, B. K.; Nazaries, L.; Munro, S.; Anderson, I. C.; Campbell,C. D. Use of multiplex terminal restriction fragment lengthpolymorphism for rapid and simultaneous analysis of differentcomponents of the soil microbial community. Appl. Environ. Microbiol.2006, 72 (11), 7278−7285.(26) Culman, S. W.; Bukowski, R.; Gauch, H. G.; Cadillo-Quiroz, H.;Buckley, D. H. T-REX: software for the processing and analysis of T-RFLP data. BMC Bioinf. 2009, 10, 171.(27) Preston-Mafham, J.; Boddy, L.; Randerson, P. F. Analysis ofmicrobial community functional diversity using sole-carbon-sourceutilisation profiles: a critique. FEMS Microbiol. Ecol. 2002, 42 (1), 1−14.(28) Hadwin, A. M.; Del Rio, L. F.; Pinto, L. J.; Painter, M.;Routledge, R.; Moore, M. M. Microbial communities in wetlands ofthe Athabasca oil sands: genetic and metabolic characterization. FEMSMicrobiol. Ecol. 2006, 55 (1), 68−78.(29) Garland, J. L.; Mills, A. L. Classification and characterization ofheterotrophic microbial communities on the basis of patterns of
community-level sole-carbon-source utilization. Appl. Environ. Micro-biol. 1991, 57 (8), 2351−2359.(30) Rodrigues, D.; Sakata, S.; Comasseto, J.; Bícego, M.; Pellizari, V.Diversity of hydrocarbon-degrading Klebsiella strains isolated fromhydrocarbon-contaminated estuaries. J. Appl. Microbiol. 2009, 106 (4),1304−1314.(31) Cole, J. R.; Chai, B.; Farris, R. J.; Wang, Q.; Kulam, S. A.;McGarrell, D. M.; Garrity, G. M.; Tiedje, J. M. The RibosomalDatabase Project (RDP-II): sequences and tools for high-throughputrRNA analysis. Nucleic Acids Res. 2005, 33 (Database issue), D294−D296.(32) Yu, Y.; Breitbart, M.; McNairnie, P.; Rohwer, F. FastGroupII: aweb-based bioinformatics platform for analyses of large 16S rDNAlibraries. BMC Bioinf. 2006, 7, 57.(33) O’Brien, H. E.; Parrent, J. L.; Jackson, J. A.; Moncalvo, J. M.;Vilgalys, R. Fungal community analysis by large-scale sequencing ofenvironmental samples. Appl. Environ. Microbiol. 2005, 71 (9), 5544−5550.(34) Taylor, D. L.; Herriott, I. C.; Long, J.; O’Neill, K. TOPO TA isA-OK: a test of phylogenetic bias in fungal environmental clone libraryconstruction. Environ. Microbiol. 2007, 9 (5), 1329−1334.(35) Kirk, P. M.; Cannon, P. F.; David, J. C.; Stalpers, J. A., Ainsworth& Bisby’s Dictionary of the Fungi, 9th ed.; CABI Bioscience: Cambridge,U.K., 2001.(36) Hattori, T. Soil aggregates as microhabitats of microorganisms.Biol. Fertil. Soil 1988, 6, 189−203.(37) Ranjard, L.; Richaume, A.; Monrozier, J. L.; Nazaret, S.Response of soil bacteria to Hg(II) in relation to soil characteristicsand cell location. FEMS Microbiol. Ecol. 1997, 24, 321−331.(38) Monrozier, J. L.; Ladd, J. N.; Fitzpatrick, R. W.; Foster, R. C.;Raupach, M. Components and microbial biomass content of sizefractions in soils of contrasting aggregation. Geoderma 1991, 49, 37−62.(39) Kabir, M.; Chotte, J. L.; Rahman, M.; Bally, R.; Monrozier, J. L.Distribution of soil fractions and location of soil bacteria in a vertisolunder cultivation and perennial grass. Plant Soil 1994, 163, 243−255.(40) Mendes, I. C.; Bottomley, P. J. Distribution of a population ofRhizobium leguminosarum bv. trifolii among different size classes of soilaggregates. Appl. Environ. Microbiol. 1998, 64 (3), 970−975.(41) Franklin, R. B.; Mills, A. L. Multi-scale variation in spatialheterogeneity for microbial community structure in an eastern Virginiaagricultural field. FEMS Microbiol. Ecol. 2003, 44 (3), 335−346.(42) Murray, R. E.; Feig, Y. S.; Tiedje, J. M. Spatial heterogeneity inthe distribution of denitrifying bacteria associated with denitrificationactivity zones. Appl. Environ. Microbiol. 1995, 61 (7), 2791−2793.(43) Ranjard, L.; Poly, F.; Combrisson, J.; Richaume, A.; Gourbiere,F.; Thioulouse, J.; Nazaret, S. Heterogeneous cell density and geneticstructure of bacterial pools associated with various soil microenviron-ments as determined by enumeration and DNA fingerprintingapproach (RISA). Microb. Ecol. 2000, 39 (4), 263−272.(44) Zhou, J.; Xia, B.; Huang, H.; Palumbo, A. V.; Tiedje, J. M.Microbial diversity and heterogeneity in sandy subsurface soils. Appl.Environ. Microbiol. 2004, 70 (3), 1723−1734.(45) Bradford, M. A.; Jones, T. H.; Bardgett, R. D.; Black, H. I.; Boag,B.; Bonkowski, M.; Cook, R.; Eggers, T.; Gange, A. C.; Grayston, S. J.;Kandeler, E.; McCaig, A. E.; Newington, J. E.; Prosser, J. I.; Setala, H.;Staddon, P. L.; Tordoff, G. M.; Tscherko, D.; Lawton, J. H. Impacts ofsoil faunal community composition on model grassland ecosystems.Science 2002, 298 (5593), 615−618.(46) Boivin, M. E.; Breure, A. M.; Posthuma, L.; Rutgers, M.Determination of field effects of contaminants-significance ofpollution-induced community tolerance. Hum. Ecol. Risk Assess.2002, 8 (5), 1035.(47) Campbell, C. D.; Grayston, S. J.; Hirst, D. J. Use of rhizospherecarbon sources in sole carbon source test to discriminate soil microbialcommunities. J. Microbiol. Methods 1997, 30, 33−41.(48) Leigh, M. B.; Pellizari, V. H.; Uhlik, O.; Sutka, R.; Rodrigues, J.;Ostrom, N. E.; Zhou, J.; Tiedje, J. M. Biphenyl-utilizing bacteria and
Environmental Science & Technology Article
dx.doi.org/10.1021/es304002q | Environ. Sci. Technol. 2013, 47, 625−633632

their functional genes in a pine root zone contaminated withpolychlorinated biphenyls (PCBs). ISME J. 2007, 1 (2), 134−148.(49) Pellizari, V. H.; Bezborodnikov, S.; Quensen, J. F., 3rd; Tiedje, J.M. Evaluation of strains isolated by growth on naphthalene andbiphenyl for hybridization of genes to dioxygenase probes andpolychlorinated biphenyl-degrading ability. Appl. Environ. Microbiol.1996, 62 (6), 2053−2058.(50) Schreiner, K. M.; Filley, T. R.; Blanchette, R. A.; Bowen, B. B.;Bolskar, R. D.; Hockaday, W. C.; Masiello, C. A.; Raebiger, J. W.White-rot basidiomycete-mediated decomposition of C60 fullerol.Environ. Sci. Technol. 2009, 43 (9), 3162−3168.(51) Blackwood, C. B.; Marsh, T.; Kim, S. H.; Paul, E. A. Terminalrestriction fragment length polymorphism data analysis for quantitativecomparison of microbial communities. Appl. Environ. Microbiol. 2003,69 (2), 926−932.(52) Marsh, T. L. Terminal restriction fragment length poly-morphism (T-RFLP): an emerging method for characterizing diversityamong homologous populations of amplification products. Curr. Opin.Microbiol. 1999, 2 (3), 323−327.(53) Yousuf, B.; Sanadhya, P.; Keshri, J.; Jha, B. Comparativemolecular analysis of chemolithoautotrophic bacterial diversity andcommunity structure from coastal saline soils, Gujarat, India. BMCMicrobiol. 2012, 12 (1), 150.(54) Yousuf, B.; Keshri, J.; Mishra, A.; Jha, B. Application of targetedmetagenomics to explore abundance and diversity of CO(2)-fixingbacterial community using cbbL gene from the rhizosphere of Arachishypogaea. Gene 2012, 506 (1), 18−24.(55) Freeman, K. R.; Pescador, M. Y.; Reed, S. C.; Costello, E. K.;Robeson, M. S.; Schmidt, S. K. Soil CO2 flux and photoautotrophiccommunity composition in high-elevation, ’barren’ soil. Environ.Microbiol. 2009, 11 (3), 674−686.(56) Eida, M. F.; Nagaoka, T.; Wasaki, J.; Kouno, K. Evaluation ofcellulolytic and hemicellulolytic abilities of fungi isolated from coffeeresidue and sawdust composts. Microbes Environ./JSME 2011, 26 (3),220−227.(57) Soares, F. L., Jr.; Melo, I. S.; Dias, A. C.; Andreote, F. D.Cellulolytic bacteria from soils in harsh environments. World J.Microbiol. Biotechnol. 2012, 28 (5), 2195−203.(58) Vyas, P.; Rahi, P.; Chauhan, A.; Gulati, A. Phosphatesolubilization potential and stress tolerance of Eupenicillium parvumfrom tea soil. Mycol. Res. 2007, 111 (Pt. 8), 931−938.(59) Aseri, G. K.; Jain, N.; Tarafdar, J. C. Hydrolysis of organicphosphate forms by phosphatases and phytase producing fungi of aridand semi arid soils in India. Am.−Eurasian J. Agric. Environ. Sci. 2009, 5(4), 564−670.
Environmental Science & Technology Article
dx.doi.org/10.1021/es304002q | Environ. Sci. Technol. 2013, 47, 625−633633




















