
Toxicity and bioaccumulation of tributyltin and triphenyltin on oysters and rock shells collected...
10
Toxicity and bioaccumulation of tributyltin and triphenyltin on oysters and rock shells collected from Taiwan maricuture area Pei-Jie Meng a, T , Jih-Terng Wang b , Li-Lian Liu c , Ming-Hui Chen a,c , Tsu-Chang Hung d a National Museum of Marine Biology and Aquarium, Biology Department, 2 Houwan Road, Checheng, Pentung, Pingtung 944, Taiwan b Tajen Institute of Technology, Ping-Tong 907, Taiwan c Institute of Marine Biology, National Sun Yat-sen University, Kaohsiung 804, Taiwan d Institute of Chemistry, Academia Sinica, Taipei 115, Taiwan Received 30 August 2004; accepted 12 January 2005 Available online 11 March 2005 Abstract The present study was undertaken to evaluate the toxicity of tributyltin (TBT) on oysters (Crassostrea gigas ) and bioaccumulation of TBT and triphenyltin (TPhT) on oysters and rock shells (Thais clavigera ) from mariculture areas of Taiwan. When treated with concentrations of 0.08, 0.40, 2.00, 10.00 and 50.00 Ag TBT/L, the 48-, 72-, 96- and 120-h LC50s of oysters were 44.6, 18.4, 17.9 and 14.3 Ag TBT/L, respectively. In the bioaccumulation experiments, oysters and rock shells were exposed to various concentrations of organotins, i.e. A: control, B: 0.40 Ag TBT/L, C: 0.40 Ag TPhT/L, and D: 0.20 Ag TBT/L + 0.20 Ag TPhT/L. In general, TPhT was faster accumulated than TBT in both oysters and rock shells and oysters had a higher elimination capability than rock shells. Additionally, greater bioaccumulation and elimination rates had been observed in female oysters than males. To rock shells, the bioaccumulation rate of organotins in imposex females was greater than males and females. D 2005 Elsevier B.V. All rights reserved. Keywords: Bioaccumulation; Organotin compounds; Rock shells and oysters; Sexual phenomena; Hermaproditic/imposex 1. Introduction Since tributyltin (TBT) was first applied as a moth- proofing agent in 1925 (Thompson et al., 1985), many organotin compounds had been produced and used as biocidal agents, catalysts and stabilizers for polyvinyl chloride polymers (Evans and Karpel, 1985). These compounds such as TBT and triphenyltin (TPhT) have caused serious pollution problems in coastal areas of many countries (Meinema et al., 1986; Alzieu, 1989, 1991, 2000; Salazer and Salazer, 1991; Iwata et al., 1995). Compounds of TBT and TPhT can be degraded to dibutyltin (DBT), monobutyltin (MBT), diphenyl- tin (DPhT) and monophenyltin (MPhT), respectively, by solar radiation, bacterial biodegradation, and/or biological decomposition (Maguire et al., 1983). 0048-9697/$ - see front matter D 2005 Elsevier B.V. All rights reserved. doi:10.1016/j.scitotenv.2005.01.016 T Corresponding author. Fax: +8868 8825034. E-mail address: [email protected] (P.-J. Meng). Science of the Total Environment 349 (2005) 140– 149 www.elsevier.com/locate/scitotenv
Transcript of Toxicity and bioaccumulation of tributyltin and triphenyltin on oysters and rock shells collected...

www.elsevier.com/locate/scitotenv
Science of the Total Environm
Toxicity and bioaccumulation of tributyltin and triphenyltin on
oysters and rock shells collected from Taiwan maricuture area
Pei-Jie Menga,T, Jih-Terng Wangb, Li-Lian Liuc, Ming-Hui Chena,c, Tsu-Chang Hungd
aNational Museum of Marine Biology and Aquarium, Biology Department, 2 Houwan Road, Checheng, Pentung, Pingtung 944, TaiwanbTajen Institute of Technology, Ping-Tong 907, Taiwan
cInstitute of Marine Biology, National Sun Yat-sen University, Kaohsiung 804, TaiwandInstitute of Chemistry, Academia Sinica, Taipei 115, Taiwan
Received 30 August 2004; accepted 12 January 2005
Available online 11 March 2005
Abstract
The present study was undertaken to evaluate the toxicity of tributyltin (TBT) on oysters (Crassostrea gigas) and
bioaccumulation of TBT and triphenyltin (TPhT) on oysters and rock shells (Thais clavigera) from mariculture areas of Taiwan.
When treated with concentrations of 0.08, 0.40, 2.00, 10.00 and 50.00 Ag TBT/L, the 48-, 72-, 96- and 120-h LC50s of oysters
were 44.6, 18.4, 17.9 and 14.3 Ag TBT/L, respectively. In the bioaccumulation experiments, oysters and rock shells were exposed
to various concentrations of organotins, i.e. A: control, B: 0.40 Ag TBT/L, C: 0.40 Ag TPhT/L, and D: 0.20 Ag TBT/L + 0.20 AgTPhT/L. In general, TPhT was faster accumulated than TBT in both oysters and rock shells and oysters had a higher
elimination capability than rock shells. Additionally, greater bioaccumulation and elimination rates had been observed in
female oysters than males. To rock shells, the bioaccumulation rate of organotins in imposex females was greater than males
and females.
D 2005 Elsevier B.V. All rights reserved.
Keywords: Bioaccumulation; Organotin compounds; Rock shells and oysters; Sexual phenomena; Hermaproditic/imposex
1. Introduction
Since tributyltin (TBT) was first applied as a moth-
proofing agent in 1925 (Thompson et al., 1985), many
organotin compounds had been produced and used as
biocidal agents, catalysts and stabilizers for polyvinyl
0048-9697/$ - see front matter D 2005 Elsevier B.V. All rights reserved.
doi:10.1016/j.scitotenv.2005.01.016
T Corresponding author. Fax: +8868 8825034.
E-mail address: [email protected] (P.-J. Meng).
chloride polymers (Evans and Karpel, 1985). These
compounds such as TBT and triphenyltin (TPhT) have
caused serious pollution problems in coastal areas of
many countries (Meinema et al., 1986; Alzieu, 1989,
1991, 2000; Salazer and Salazer, 1991; Iwata et al.,
1995). Compounds of TBT and TPhT can be degraded
to dibutyltin (DBT), monobutyltin (MBT), diphenyl-
tin (DPhT) and monophenyltin (MPhT), respectively,
by solar radiation, bacterial biodegradation, and/or
biological decomposition (Maguire et al., 1983).
ent 349 (2005) 140–149
P.-J. Meng et al. / Science of the Total Environment 349 (2005) 140–149 141
These compounds are toxic to various marine
organisms especially TBT, for instance, as low as
ppb level in seawater, which can induce oyster shell
abnormalities and spat development failure (Alzieu,
1986, 1991, 2000), imposex on snails (e.g. Wilken et
al., 1994; Fent and Meier, 1994; Horiguchi et al.,
1994; Bryan et al., 1986) and etc.
Although ship related industries contribute
unknown quantities of organotins into the coastal
waters of Taiwan, the Taiwan Agriculture Industry
Association (TAIA, 1997) reported that more than
150 tons of 45% and 20 tons of 2% TPhT acetate
were used for agriculture in 1996 and banned in
2001. Therefore, it is not surprising that as high as
2500 ng/g dry wt of TBT were found in sediments
dredged from the dumpsites in Keelung Harbor
(Hung and Liu, 1998), and high TBT and TPhT (i.e.
1510 and 590 ng/g dry wt, respectively) were
observed in oysters (Crassostrea gigas) from mar-
iculture areas of Shiangsan and Lukang (Hung et al.,
1998). Rock shells (Thais clavigera) in the oyster
0
10
20
30
40
50
60
70
80
90
100
0 20 40 60Hou
0 20 40 60Hour
MO
RT
AL
ITY
(%
)M
OR
TA
LIT
Y (
%)
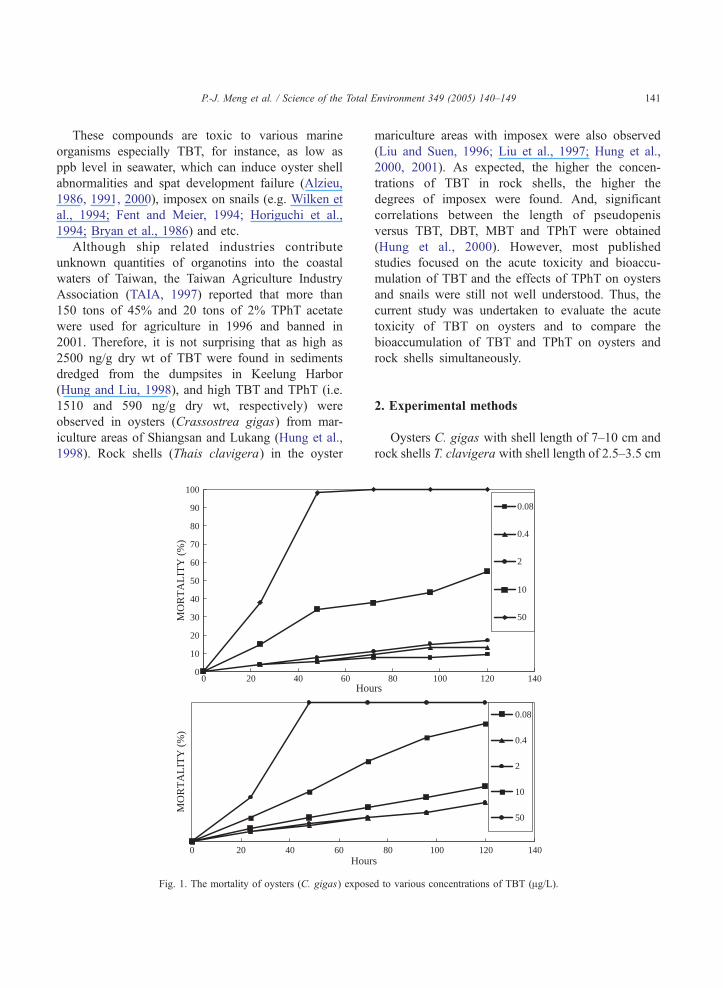
Fig. 1. The mortality of oysters (C. gigas) expose
mariculture areas with imposex were also observed
(Liu and Suen, 1996; Liu et al., 1997; Hung et al.,
2000, 2001). As expected, the higher the concen-
trations of TBT in rock shells, the higher the
degrees of imposex were found. And, significant
correlations between the length of pseudopenis
versus TBT, DBT, MBT and TPhT were obtained
(Hung et al., 2000). However, most published
studies focused on the acute toxicity and bioaccu-
mulation of TBT and the effects of TPhT on oysters
and snails were still not well understood. Thus, the
current study was undertaken to evaluate the acute
toxicity of TBT on oysters and to compare the
bioaccumulation of TBT and TPhT on oysters and
rock shells simultaneously.
2. Experimental methods
Oysters C. gigas with shell length of 7–10 cm and
rock shells T. clavigera with shell length of 2.5–3.5 cm
80 100 120 140rs
80 100 120 140s
0.08
0.4
2
10
50
0.08
0.4
2
10
50
d to various concentrations of TBT (Ag/L).
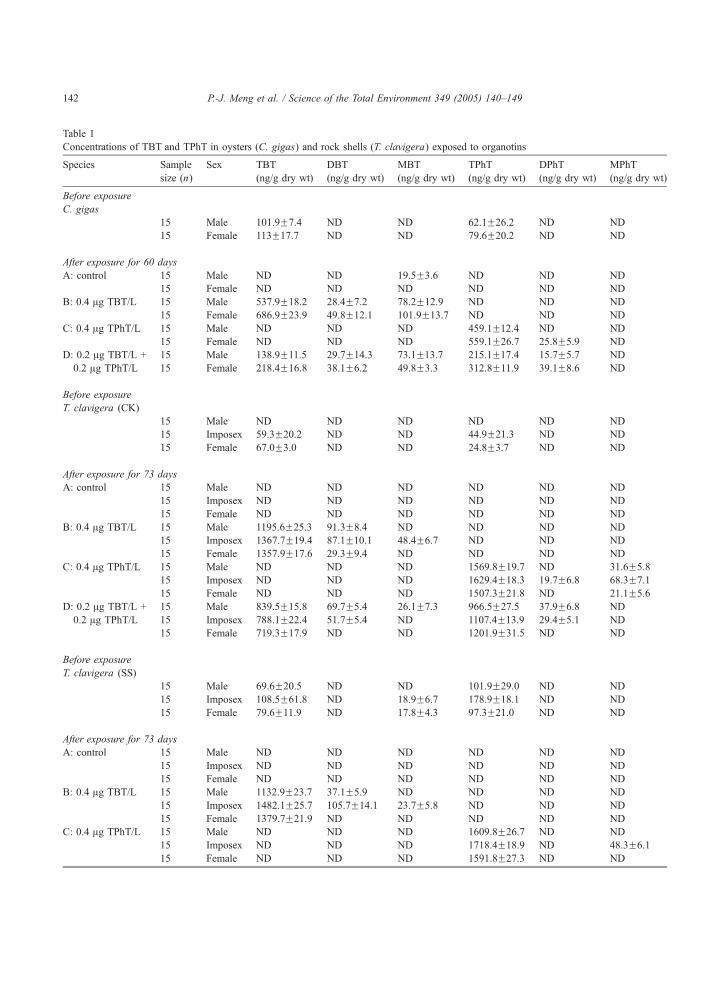
Table 1
Concentrations of TBT and TPhT in oysters (C. gigas) and rock shells (T. clavigera) exposed to organotins
Species Sample
size (n)
Sex TBT
(ng/g dry wt)
DBT
(ng/g dry wt)
MBT
(ng/g dry wt)
TPhT
(ng/g dry wt)
DPhT
(ng/g dry wt)
MPhT
(ng/g dry wt)
Before exposure
C. gigas
15 Male 101.9F7.4 ND ND 62.1F26.2 ND ND
15 Female 113F17.7 ND ND 79.6F20.2 ND ND
After exposure for 60 days
A: control 15 Male ND ND 19.5F3.6 ND ND ND
15 Female ND ND ND ND ND ND
B: 0.4 Ag TBT/L 15 Male 537.9F18.2 28.4F7.2 78.2F12.9 ND ND ND
15 Female 686.9F23.9 49.8F12.1 101.9F13.7 ND ND ND
C: 0.4 Ag TPhT/L 15 Male ND ND ND 459.1F12.4 ND ND
15 Female ND ND ND 559.1F26.7 25.8F5.9 ND
D: 0.2 Ag TBT/L +
0.2 Ag TPhT/L
15 Male 138.9F11.5 29.7F14.3 73.1F13.7 215.1F17.4 15.7F5.7 ND
15 Female 218.4F16.8 38.1F6.2 49.8F3.3 312.8F11.9 39.1F8.6 ND
Before exposure
T. clavigera (CK)
15 Male ND ND ND ND ND ND
15 Imposex 59.3F20.2 ND ND 44.9F21.3 ND ND
15 Female 67.0F3.0 ND ND 24.8F3.7 ND ND
After exposure for 73 days
A: control 15 Male ND ND ND ND ND ND
15 Imposex ND ND ND ND ND ND
15 Female ND ND ND ND ND ND
B: 0.4 Ag TBT/L 15 Male 1195.6F25.3 91.3F8.4 ND ND ND ND
15 Imposex 1367.7F19.4 87.1F10.1 48.4F6.7 ND ND ND
15 Female 1357.9F17.6 29.3F9.4 ND ND ND ND
C: 0.4 Ag TPhT/L 15 Male ND ND ND 1569.8F19.7 ND 31.6F5.8
15 Imposex ND ND ND 1629.4F18.3 19.7F6.8 68.3F7.1
15 Female ND ND ND 1507.3F21.8 ND 21.1F5.6
D: 0.2 Ag TBT/L +
0.2 Ag TPhT/L
15 Male 839.5F15.8 69.7F5.4 26.1F7.3 966.5F27.5 37.9F6.8 ND
15 Imposex 788.1F22.4 51.7F5.4 ND 1107.4F13.9 29.4F5.1 ND
15 Female 719.3F17.9 ND ND 1201.9F31.5 ND ND
Before exposure
T. clavigera (SS)
15 Male 69.6F20.5 ND ND 101.9F29.0 ND ND
15 Imposex 108.5F61.8 ND 18.9F6.7 178.9F18.1 ND ND
15 Female 79.6F11.9 ND 17.8F4.3 97.3F21.0 ND ND
After exposure for 73 days
A: control 15 Male ND ND ND ND ND ND
15 Imposex ND ND ND ND ND ND
15 Female ND ND ND ND ND ND
B: 0.4 Ag TBT/L 15 Male 1132.9F23.7 37.1F5.9 ND ND ND ND
15 Imposex 1482.1F25.7 105.7F14.1 23.7F5.8 ND ND ND
15 Female 1379.7F21.9 ND ND ND ND ND
C: 0.4 Ag TPhT/L 15 Male ND ND ND 1609.8F26.7 ND ND
15 Imposex ND ND ND 1718.4F18.9 ND 48.3F6.1
15 Female ND ND ND 1591.8F27.3 ND ND
P.-J. Meng et al. / Science of the Total Environment 349 (2005) 140–149142
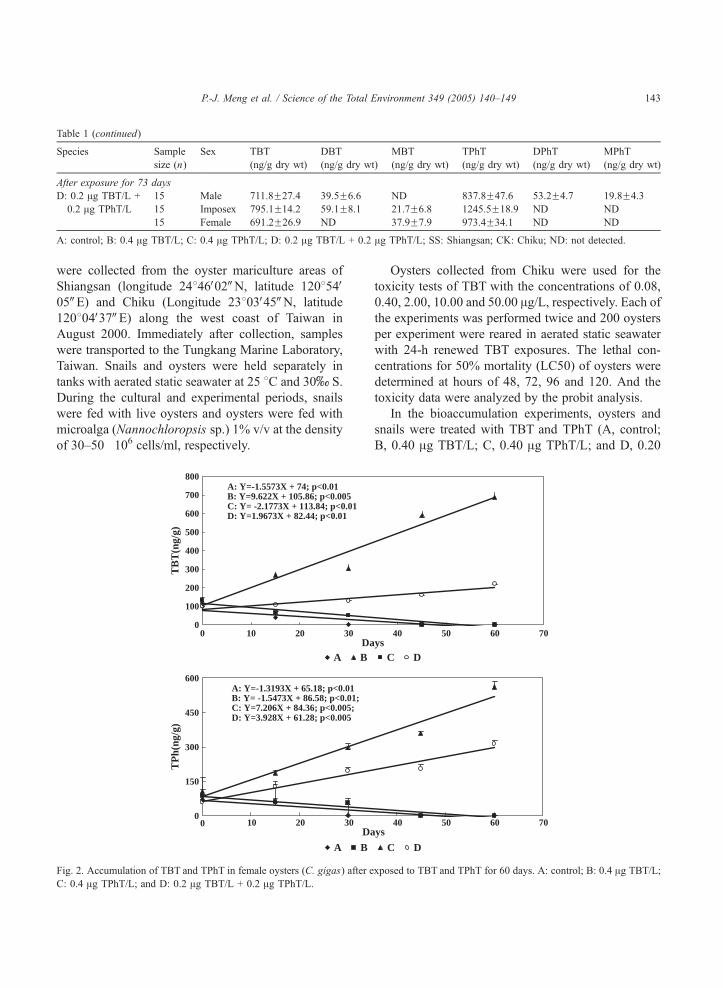
Table 1 (continued)
Species Sample
size (n)
Sex TBT
(ng/g dry wt)
DBT
(ng/g dry wt)
MBT
(ng/g dry wt)
TPhT
(ng/g dry wt)
DPhT
(ng/g dry wt)
MPhT
(ng/g dry wt)
After exposure for 73 days
D: 0.2 Ag TBT/L +
0.2 Ag TPhT/L
15 Male 711.8F27.4 39.5F6.6 ND 837.8F47.6 53.2F4.7 19.8F4.3
15 Imposex 795.1F14.2 59.1F8.1 21.7F6.8 1245.5F18.9 ND ND
15 Female 691.2F26.9 ND 37.9F7.9 973.4F34.1 ND ND
A: control; B: 0.4 Ag TBT/L; C: 0.4 Ag TPhT/L; D: 0.2 Ag TBT/L + 0.2 Ag TPhT/L; SS: Shiangsan; CK: Chiku; ND: not detected.
P.-J. Meng et al. / Science of the Total Environment 349 (2005) 140–149 143
were collected from the oyster mariculture areas of
Shiangsan (longitude 24846V02WN, latitude 120854V05WE) and Chiku (Longitude 23803V45WN, latitude
120804V37WE) along the west coast of Taiwan in
August 2000. Immediately after collection, samples
were transported to the Tungkang Marine Laboratory,
Taiwan. Snails and oysters were held separately in
tanks with aerated static seawater at 25 8C and 30x S.
During the cultural and experimental periods, snails
were fed with live oysters and oysters were fed with
microalga (Nannochloropsis sp.) 1% v/v at the density
of 30–50�106 cells/ml, respectively.
A: Y=-1.5573X + 74; p<0.01B: Y=9.622X + 105.86; p<0.005C: Y= -2.1773X + 113.84; p<0.01D: Y=1.9673X + 82.44; p<0.01
0
100
200
300
400
500
600
700
800
0 10 20 30
10 20 30
Da
TB
T(n
g/g)
A B
A B
A: Y=-1.3193X + 65.18; p<0.01B: Y= -1.5473X + 86.58; p<0.01;C: Y=7.206X + 84.36; p<0.005;D: Y=3.928X + 61.28; p<0.005
0
150
300
450
600
0Da
TP
h(ng
/g)
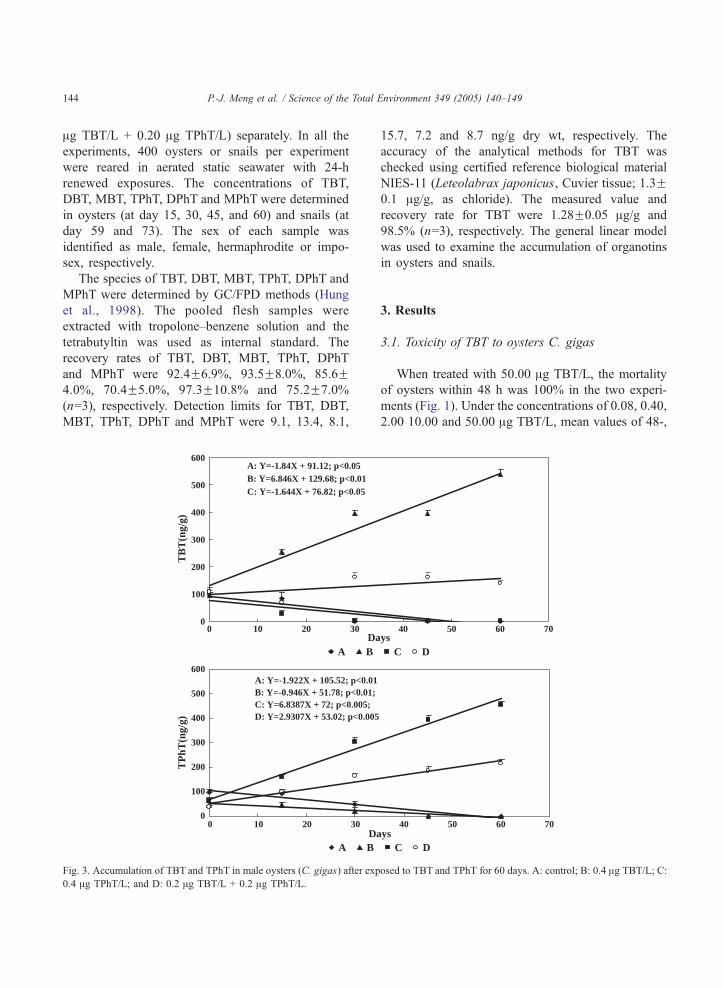
Fig. 2. Accumulation of TBT and TPhT in female oysters (C. gigas) after e
C: 0.4 Ag TPhT/L; and D: 0.2 Ag TBT/L + 0.2 Ag TPhT/L.
Oysters collected from Chiku were used for the
toxicity tests of TBT with the concentrations of 0.08,
0.40, 2.00, 10.00 and 50.00 Ag/L, respectively. Each ofthe experiments was performed twice and 200 oysters
per experiment were reared in aerated static seawater
with 24-h renewed TBT exposures. The lethal con-
centrations for 50% mortality (LC50) of oysters were
determined at hours of 48, 72, 96 and 120. And the
toxicity data were analyzed by the probit analysis.
In the bioaccumulation experiments, oysters and
snails were treated with TBT and TPhT (A, control;
B, 0.40 Ag TBT/L; C, 0.40 Ag TPhT/L; and D, 0.20
40 50 60 70
40 50 60 70
ysC D
C D
ys
xposed to TBT and TPhT for 60 days. A: control; B: 0.4 Ag TBT/L;
P.-J. Meng et al. / Science of the Total Environment 349 (2005) 140–149144
Ag TBT/L + 0.20 Ag TPhT/L) separately. In all the
experiments, 400 oysters or snails per experiment
were reared in aerated static seawater with 24-h
renewed exposures. The concentrations of TBT,
DBT, MBT, TPhT, DPhT and MPhT were determined
in oysters (at day 15, 30, 45, and 60) and snails (at
day 59 and 73). The sex of each sample was
identified as male, female, hermaphrodite or impo-
sex, respectively.
The species of TBT, DBT, MBT, TPhT, DPhT and
MPhT were determined by GC/FPD methods (Hung
et al., 1998). The pooled flesh samples were
extracted with tropolone–benzene solution and the
tetrabutyltin was used as internal standard. The
recovery rates of TBT, DBT, MBT, TPhT, DPhT
and MPhT were 92.4F6.9%, 93.5F8.0%, 85.6F4.0%, 70.4F5.0%, 97.3F10.8% and 75.2F7.0%
(n=3), respectively. Detection limits for TBT, DBT,
MBT, TPhT, DPhT and MPhT were 9.1, 13.4, 8.1,
A: Y=-1.84X + 91.12; p<0.05B: Y=6.846X + 129.68; p<0.01C: Y=-1.644X + 76.82; p<0.05
0
100
200
300
400
500
600
0 10 20 30
0 10 20 30
Da
TB
T(n
g/g)
BA
BA
A: Y=-1.922X + 105.52; p<0.01B: Y=-0.946X + 51.78; p<0.01;C: Y=6.8387X + 72; p<0.005;D: Y=2.9307X + 53.02; p<0.005
0
100
200
300
400
500
600
Da
TP
hT(n
g/g)
Fig. 3. Accumulation of TBT and TPhT in male oysters (C. gigas) after exp
0.4 Ag TPhT/L; and D: 0.2 Ag TBT/L + 0.2 Ag TPhT/L.
15.7, 7.2 and 8.7 ng/g dry wt, respectively. The
accuracy of the analytical methods for TBT was
checked using certified reference biological material
NIES-11 (Leteolabrax japonicus, Cuvier tissue; 1.3F0.1 Ag/g, as chloride). The measured value and
recovery rate for TBT were 1.28F0.05 Ag/g and
98.5% (n=3), respectively. The general linear model
was used to examine the accumulation of organotins
in oysters and snails.
3. Results
3.1. Toxicity of TBT to oysters C. gigas
When treated with 50.00 Ag TBT/L, the mortality
of oysters within 48 h was 100% in the two experi-
ments (Fig. 1). Under the concentrations of 0.08, 0.40,
2.00 10.00 and 50.00 Ag TBT/L, mean values of 48-,
40 50 60 70
40 50 60 70
ysC D
C Dys
osed to TBT and TPhT for 60 days. A: control; B: 0.4 Ag TBT/L; C:
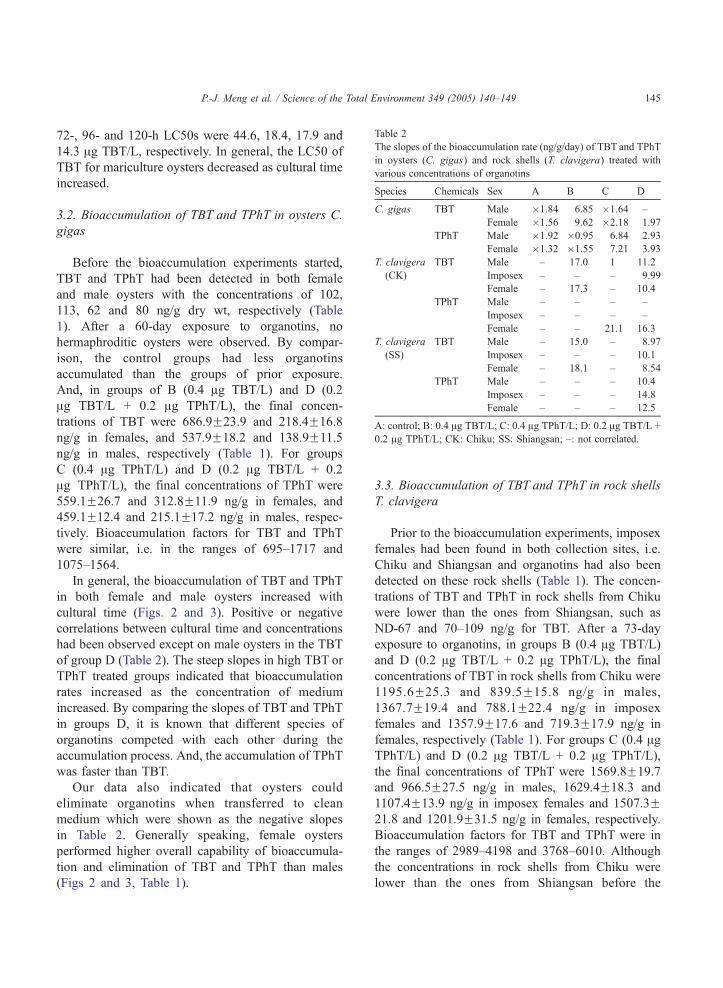
Table 2
The slopes of the bioaccumulation rate (ng/g/day) of TBT and TPhT
in oysters (C. gigas) and rock shells (T. clavigera) treated with
various concentrations of organotins
Species Chemicals Sex A B C D
C. gigas TBT Male �1.84 6.85 �1.64 –
Female �1.56 9.62 �2.18 1.97
TPhT Male �1.92 �0.95 6.84 2.93
Female �1.32 �1.55 7.21 3.93
T. clavigera
(CK)
TBT Male – 17.0 1 11.2
Imposex – – – 9.99
Female – 17.3 – 10.4
TPhT Male – – – –
Imposex – – – –
Female – – 21.1 16.3
T. clavigera
(SS)
TBT Male – 15.0 – 8.97
Imposex – – – 10.1
Female – 18.1 – 8.54
TPhT Male – – – 10.4
Imposex – – – 14.8
Female – – – 12.5
A: control; B: 0.4 Ag TBT/L; C: 0.4 Ag TPhT/L; D: 0.2 Ag TBT/L +
0.2 Ag TPhT/L; CK: Chiku; SS: Shiangsan; –: not correlated.
P.-J. Meng et al. / Science of the Total Environment 349 (2005) 140–149 145
72-, 96- and 120-h LC50s were 44.6, 18.4, 17.9 and
14.3 Ag TBT/L, respectively. In general, the LC50 of
TBT for mariculture oysters decreased as cultural time
increased.
3.2. Bioaccumulation of TBT and TPhT in oysters C.
gigas
Before the bioaccumulation experiments started,
TBT and TPhT had been detected in both female
and male oysters with the concentrations of 102,
113, 62 and 80 ng/g dry wt, respectively (Table
1). After a 60-day exposure to organotins, no
hermaphroditic oysters were observed. By compar-
ison, the control groups had less organotins
accumulated than the groups of prior exposure.
And, in groups of B (0.4 Ag TBT/L) and D (0.2
Ag TBT/L + 0.2 Ag TPhT/L), the final concen-
trations of TBT were 686.9F23.9 and 218.4F16.8
ng/g in females, and 537.9F18.2 and 138.9F11.5
ng/g in males, respectively (Table 1). For groups
C (0.4 Ag TPhT/L) and D (0.2 Ag TBT/L + 0.2
Ag TPhT/L), the final concentrations of TPhT were
559.1F26.7 and 312.8F11.9 ng/g in females, and
459.1F12.4 and 215.1F17.2 ng/g in males, respec-
tively. Bioaccumulation factors for TBT and TPhT
were similar, i.e. in the ranges of 695–1717 and
1075–1564.
In general, the bioaccumulation of TBT and TPhT
in both female and male oysters increased with
cultural time (Figs. 2 and 3). Positive or negative
correlations between cultural time and concentrations
had been observed except on male oysters in the TBT
of group D (Table 2). The steep slopes in high TBT or
TPhT treated groups indicated that bioaccumulation
rates increased as the concentration of medium
increased. By comparing the slopes of TBT and TPhT
in groups D, it is known that different species of
organotins competed with each other during the
accumulation process. And, the accumulation of TPhT
was faster than TBT.
Our data also indicated that oysters could
eliminate organotins when transferred to clean
medium which were shown as the negative slopes
in Table 2. Generally speaking, female oysters
performed higher overall capability of bioaccumula-
tion and elimination of TBT and TPhT than males
(Figs 2 and 3, Table 1).
3.3. Bioaccumulation of TBT and TPhT in rock shells
T. clavigera
Prior to the bioaccumulation experiments, imposex
females had been found in both collection sites, i.e.
Chiku and Shiangsan and organotins had also been
detected on these rock shells (Table 1). The concen-
trations of TBT and TPhT in rock shells from Chiku
were lower than the ones from Shiangsan, such as
ND-67 and 70–109 ng/g for TBT. After a 73-day
exposure to organotins, in groups B (0.4 Ag TBT/L)
and D (0.2 Ag TBT/L + 0.2 Ag TPhT/L), the final
concentrations of TBT in rock shells from Chiku were
1195.6F25.3 and 839.5F15.8 ng/g in males,
1367.7F19.4 and 788.1F22.4 ng/g in imposex
females and 1357.9F17.6 and 719.3F17.9 ng/g in
females, respectively (Table 1). For groups C (0.4 AgTPhT/L) and D (0.2 Ag TBT/L + 0.2 Ag TPhT/L),
the final concentrations of TPhT were 1569.8F19.7
and 966.5F27.5 ng/g in males, 1629.4F18.3 and
1107.4F13.9 ng/g in imposex females and 1507.3F21.8 and 1201.9F31.5 ng/g in females, respectively.
Bioaccumulation factors for TBT and TPhT were in
the ranges of 2989–4198 and 3768–6010. Although
the concentrations in rock shells from Chiku were
lower than the ones from Shiangsan before the
P.-J. Meng et al. / Science of the Total Environment 349 (2005) 140–149146
exposure experiments started, after a 73-day exposure,
the final concentrations of organotins and bio-
accumulation factors from both sites were similar
(Table 1).
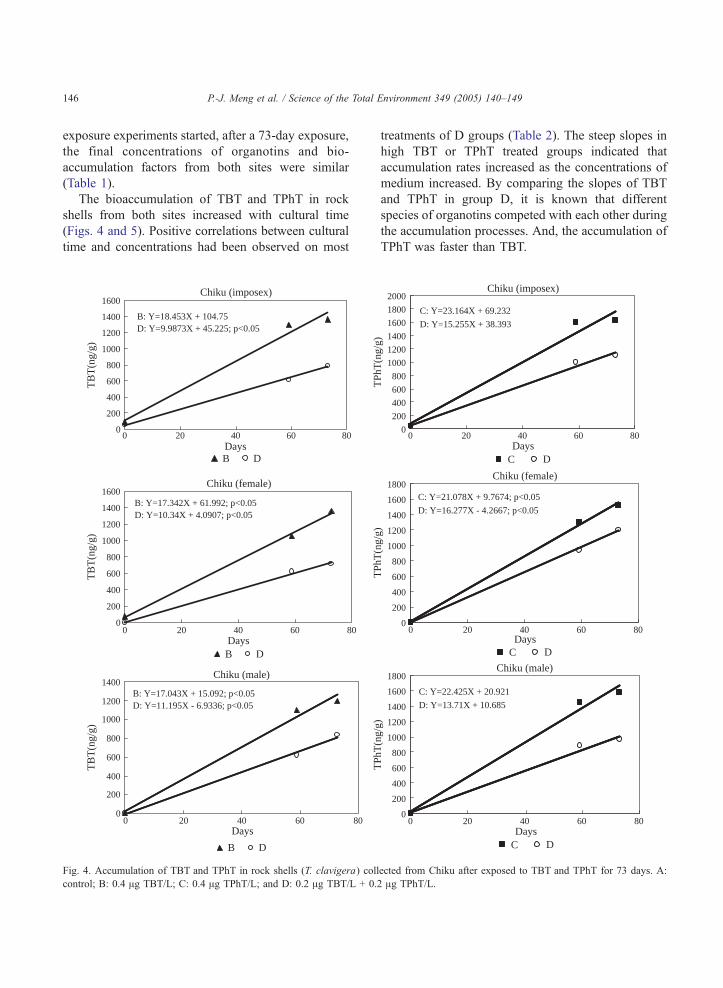
The bioaccumulation of TBT and TPhT in rock
shells from both sites increased with cultural time
(Figs. 4 and 5). Positive correlations between cultural
time and concentrations had been observed on most
Chiku (imposex)
0
200
400
600
800
1000
1200
1400
1600
0 20 40 60 80Days
TB
T(n
g/g)
B D
B D
B D
B: Y=18.453X + 104.75D: Y=9.9873X + 45.225; p<0.05
Chiku (female)
0
200
400
600
800
1000
1200
1400
1600
0 20 40 60 80
20 40 60 80
Days
TB
T(n
g/g)
B: Y=17.342X + 61.992; p<0.05D: Y=10.34X + 4.0907; p<0.05
Chiku (male)
0
200
400
600
800
1000
1200
1400
0Days
TB
T(n
g/g)
B: Y=17.043X + 15.092; p<0.05D: Y=11.195X - 6.9336; p<0.05
Fig. 4. Accumulation of TBT and TPhT in rock shells (T. clavigera) coll
control; B: 0.4 Ag TBT/L; C: 0.4 Ag TPhT/L; and D: 0.2 Ag TBT/L + 0.
treatments of D groups (Table 2). The steep slopes in
high TBT or TPhT treated groups indicated that
accumulation rates increased as the concentrations of
medium increased. By comparing the slopes of TBT
and TPhT in group D, it is known that different
species of organotins competed with each other during
the accumulation processes. And, the accumulation of
TPhT was faster than TBT.
Chiku (imposex)
0
200
400
600
800
1000
1200
1400
1600
1800
2000
0 20 40 60 80
20 40 60 80
20 40 60 80
Days
TPh
T(n
g/g)
C D
C D
C D
C: Y=23.164X + 69.232
D: Y=15.255X + 38.393
Chiku (female)
0
200
400
600
800
1000
1200
1400
1600
1800
0Days
TPh
T(n
g/g)
C: Y=21.078X + 9.7674; p<0.05
D: Y=16.277X - 4.2667; p<0.05
Chiku (male)
0
200
400
600
800
1000
1200
1400
1600
1800
0Days
TPh
T(n
g/g)
C: Y=22.425X + 20.921
D: Y=13.71X + 10.685
ected from Chiku after exposed to TBT and TPhT for 73 days. A:
2 Ag TPhT/L.
Shiangsan (imposex)
0
200
400
600
800
1000
1200
1400
1600
1800
0 20 40 60 80
20 40 60 80
Days
TB
T(n
g/g)
B D
B D
B: Y=18.302X + 213.36D: Y=10.13X + 73.687; p<0.05
Shiangsan (female)
0
200
400
600
800
1000
1200
1400
1600
0Days
20 40 60 80
B D
0Days
TB
T(n
g/g)
B: Y=18.052X + 89.689; p<0.05D: Y=8.543X + 83.608; p<0.05
Shiangsan (male)
0
200
400
600
800
1000
1200
1400
TB
T(n
g/g)
B: Y=15.029X + 64.652; p<0.05D: Y=8.9733X + 84.742; p<0.05
Shiangsan (imposex)
0
500
1000
1500
2000
0 20 40 60 80
Days
TPh
T(n
g/g)
C D
0 20 40 60 80
20 40 60 80
DaysC D
DaysC D
C: Y=22.051X + 209D: Y=14.768X + 155.58; p<0.05
Shiangsan (female)
0
500
1000
1500
2000
TPh
T(n
g/g)
C: Y=21.319X + 116.39D: Y=12.537X + 71.957; p<0.05
Shiangsan (male)
0
200
400
600
800
1000
1200
1400
1600
1800
0
TPh
T(n
g/g)
C: Y=20.966X + 162.31D: Y=10.359X + 108.88; p<0.05
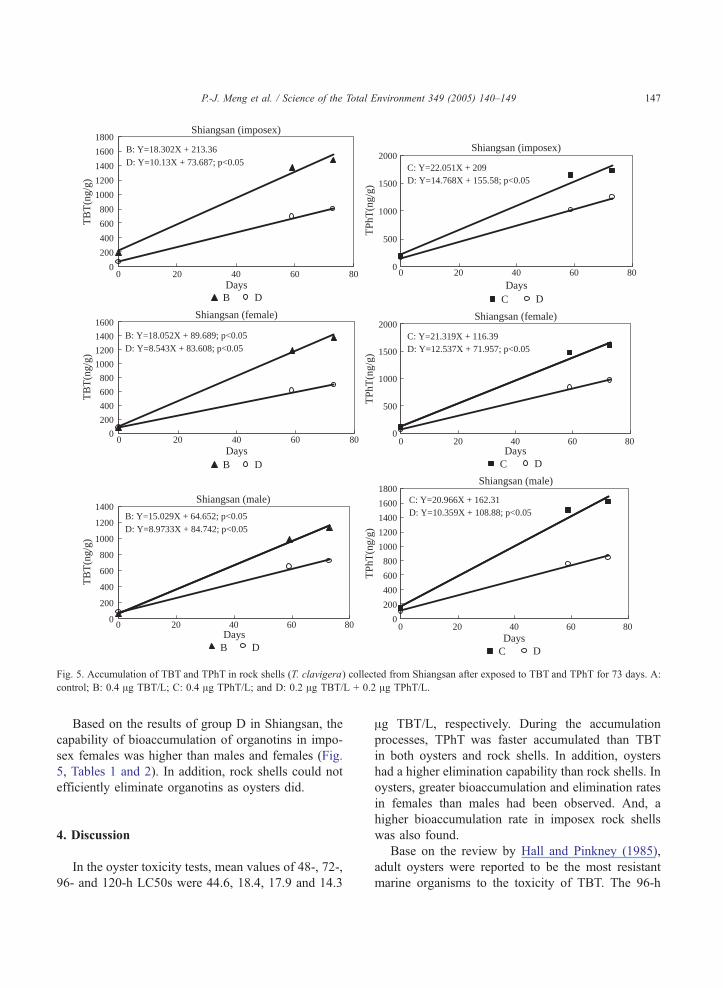
Fig. 5. Accumulation of TBT and TPhT in rock shells (T. clavigera) collected from Shiangsan after exposed to TBT and TPhT for 73 days. A:
control; B: 0.4 Ag TBT/L; C: 0.4 Ag TPhT/L; and D: 0.2 Ag TBT/L + 0.2 Ag TPhT/L.
P.-J. Meng et al. / Science of the Total Environment 349 (2005) 140–149 147
Based on the results of group D in Shiangsan, the
capability of bioaccumulation of organotins in impo-
sex females was higher than males and females (Fig.
5, Tables 1 and 2). In addition, rock shells could not
efficiently eliminate organotins as oysters did.
4. Discussion
In the oyster toxicity tests, mean values of 48-, 72-,
96- and 120-h LC50s were 44.6, 18.4, 17.9 and 14.3
Ag TBT/L, respectively. During the accumulation
processes, TPhT was faster accumulated than TBT
in both oysters and rock shells. In addition, oysters
had a higher elimination capability than rock shells. In
oysters, greater bioaccumulation and elimination rates
in females than males had been observed. And, a
higher bioaccumulation rate in imposex rock shells
was also found.
Base on the review by Hall and Pinkney (1985),
adult oysters were reported to be the most resistant
marine organisms to the toxicity of TBT. The 96-h

P.-J. Meng et al. / Science of the Total Environment 349 (2005) 140–149148
LC50s of 210, 290 and 560–1000 Ag/L TBTO were
reported for adult European oysters (Ostreia edulis),
adult Pacific oysters (C. gigas) and adult eastern
oysters (C. virginica). However, our comparable data,
i.e. 17.9 Ag TBT/L, was much lower than those studies.
It is known that environmental factors such as
temperature and salinity affect the tolerance of
organisms in a great extend which may result in
enlarge, narrow, or shift the ranges of the lower or
upper limits (Kinne, 1963). In some species, increas-
ing salinity has been found to increase resistance to
high temperatures, while in others no effect has been
observed. In the studies of fiddler crab zoeae (Uca
pugilator), the 48-h LC50s at 30x S and 10, 20 and
28 8C were 1.4, 0.37 and 0.08 mg trimethyltin/L,
respectively (Hall and Pinkney, 1985). When treated
at 20x S and 10, 20 and 28 8C, the 48-h LC50s were
0.20, 0.04 and 0.03 mg trimethyltin/L, respectively.
As the temperature increased, the values of LC50
decreased accordingly in the case of fiddler crab
zoeae. In the present study, the mean seawater
temperature was 28.5 8C during the experimental
period and a lower tolerance was expected. However,
it cannot be confirmed based on available literatures.
Although the bioaccumulation of organotins by
aquatic organisms has been extensively studied in the
past three decades, available information was mostly
focused on TBT (Laughlin, 1996). The accumulation
factor of TBT in adult oyster C. virginica was 27,800.
In C. gigas, values varied greatly as 2000, 6000, 7000
and 135,000. In the present study, it was from 695 to
1717 and fell into the lower end of reported data. In
the dogwhelk Nucella lapillus, the reported accumu-
lation factors of TBT were 10,000, 10,500 and
100,000 (Laughlin, 1996). The reports that not only
TBT but also TPT promoted imposex in the rock shell
T. clavigera were published by Horiguchi and co-
workers (Horiguchi et al., 1995, 1997a,b, 1998). The
potency of TPT for promoting the development of
imposex in the rock shell is estimated to be
approximately the same as that of TBT (Horiguchi
et al., 1997a). The accumulation factors of TBT and
TPhT have been estimated at 5000–10,000 and
22,000, respectively. These values are much higher
than our results, i.e. 2989–4198 and 3768–6010. It is
known that the relationship between duration of
exposure and occurrence of a steady state is an
important issue in developing a predictive relationship
between exposure and tissue burdens. Thus, the low
accumulation factors of oysters and rock shells found
in our studies may simply result from the failure of
reaching steady states (shown in Figs. 2–5).
In addition, the notable differences between female
and male oysters or imposex females and other rock
shells in bioaccumulation of organotins might be the
reason why the concentrations of TBT and TPhT in
female oysters and imposex rock shells were generally
higher when surveying organotin contents in mar-
iculture areas (Hung et al., 2000, 2001).
Acknowledgements
We are grateful for the constructive comments of
the anonymous reviewers who have substantially
improved the manuscript. This study was supported
by National Science Council/ROC grant NSC89-
2621-B-002-036.
References
Alzieu C. TBT detrimental effects on oysters culture in France—
evolution since antifouling paint regulation. Proceedings of
the Oceans International Organotin Symposium, vol. 4; 1986.
p. 1130–4.
Alzieu C. Monitoring and assessment of butyltins in Atlantic coastal
water. Mar Pollut Bull 1989;20:22–6.
Alzieu C. Environmental problems caused by TBT in France:
assessment, regulation, prospects. Mar Environ Res 1991;
32:7–17.
Alzieu C. Environmental impact of TBT: the French experience. Sci
Tot Environ 2000;258:99–102.
Bryan GW, Gibbs PE, Hummerstone LG, Burt GR. The decline of
the gastropod Nucella lapillus around southwest England:
evidence for the effect of tributyltin from antifouling paint. J
Mar Biol Assoc UK 1986;66:611–40;
Environ 1986;258:99–102.
Evans CJ, Karpel S. Organotin compounds in modern technology. J
Organomet Chem Libr 1985;16:178–217.
Fent K, Meier W. Effects of tributyltin of fish early life stages. Arch
Environ Contam Toxicol 1994;27:224–31.
Hall Jr LW, Pinkney AE. Acute and sublethal effects of organotin
compounds on aquatic biota: an interpretative literature evalua-
tion. CRC Crit Rev Toxicol 1985;14:159–209.
Horiguchi T, Shiraishi H, Shimizu M, Morita M. Imposex and
organotin compounds in Thais clavigera and T bronni in Japan.
J Mar Biol Assoc UK 1994;74:651–69.
Horiguchi T, Shiraishi H, Shimizu M, Yamazaki S, Morita M.
Imposex in Japanese gastropods (Neogastropoda and Mesogas-

P.-J. Meng et al. / Science of the Total Environment 349 (2005) 140–149 149
tropoda): effects of tributyltin and triphenyltin from antifouling
paints. Mar Pollut Bull 1995;31:402–5.
Horiguchi T, Shiraishi H, Shimizu M, Morita M. Imposex in sea
snails, caused by organotin (tributyltin and triphenyltin)
pollution in Japan: a survey. Appl Organomet Chem 1997a;
11:451–5.
Horiguchi T, Shiraishi H, Shimizu M, Morita M. Effects of
triphenyltin chloride and five other organotin compounds on
the development of imposex in the rock shell, Thais clavigera.
Environ Pollut 1997b;95:85–91.
Horiguchi T, Hyeon-Seo C, Shiraishi H, Shibata Y, Soma M, Morita
M, et al. Field studies on imposex and organotin accumulation
in the rock shell, Thais clavigera, from the Seto Inland Sea and
the Sanriku region Japan. Sci Total Environ 1998;214:65–70.
Hung TC, Liu BP. Determination of tributyltin in sediments from
the Machu and Taiwan coastal areas. Acta Oceanogr Taiwanica
1998;37:105–12.
Hung TC, Lee TY, Liao TF. Determination of butyltins and
phenyltins in oysters and fishes from Taiwan coastal waters.
Environ Pollut 1998;102:197–203.
Hung TC, Hsu WK, Meng PJ, Chuang A. Species of organotins in
imposex of rock shells and hermaphroditic oysters from the
western coast of Taiwan. Bull Inst Chem Academia Sinica
2000;47:1–12.
Hung TC, Hsu WK, Meng PJ, Chuang A. Organotins and imposex
in the rock shells, Thais clavigera, from the Taiwan oyster
mariculture area. Environ Pollut 2001;112:145–52.
Iwata H, Tanabe S, Mizuno T, Tatsukawa R. High accumulation of
toxic butyltins in marine mammals from Japanese coastal
waters. Environ Sci Technol 1995;29:2959–62.
Kinne O. The effects of temperature and salinity on marine and
brackish water animals: I. Temperature. Oceanogr Mar Bio Ann
Rev 1963;1:301–40.
Laughlin Jr RB. Bioaccumulation of TBT by aquatic organisms. In:
Champ MA, Seligman PF, editors. Organotin—environmental
fate and effects. London7 Chapman and Hall; 1996.
Liu LL, Suen IJ. Prosobranch gastropod imposex in the west coast
of Taiwan. Venus 1996;55:207–14.
Liu LL, Chen SJ, Peng WY, Hung JJ. Organotin concentrations in
three intertidal neogastropods from the coastal waters of Taiwan.
Environ Pollut 1997;98:113–8.
Maguire RJ, Carey JH, Hale EJ. Degradation of the tributyltin
species in water. J Agric Food Chem 1983;31:1060–5.
Meinema HA, Van Dam-Meerbeek TG, Vonk WJ. Evaluation of
impact of organotin compounds on the aquatic environment.
Final Rept. of TNO, Utrecht to EEC. Contract No. 85/B/600/
11/007/11/N; 1986.
Salazer MH, Salazer SM. Assessing site-specific effects of TBT
contamination with mussel growth rates. Mar Environ Res 1991;
32:131–50.
TAIA. Domestic manufactures production and sale of pesticides in
1996. Taiwan Agriculture Industry Association; 1997.
Thompson JA, Sheffer MG, Pierce RC, Chau YK, Cooney JJ,
Cullen WP, et al. Organotin compounds in the aquatic
environment: scientific criteria for assessing their effects on
environmental quality. National Research Council Canada;
1985. p. 284.
Wilken RD, Kuballa J, Jantzen E. Organotin: their analysis and
assessment in the Elbe river system, north Germany. Fresenius’
J Anal Chem 1994;25:77–84.




















