
Topo thérapeutique Angelman ms Juin2012
24
médecine/sciences m/s n° 6-7, vol. 28, juin-juillet 2012 DOI : 10.1051/medsci/2012286002 médecine/sciences 2012 ; 28 : 000-00 NOUVELLES MAGAZINE 1 1 UR892 Inra (Institut national de la recherche agronomique), virologie et immunologie moléculaires, 78350 Jouy-en Josas, France ; 2 UMR1313 INRA, génétique animale et biologie intégrative, 78350 Jouy-en-Josas, France. [email protected] Un agent infectieux de nature protéique Le prion est le pathogène responsable, chez les mammifères, de désordres neu- rodégénératifs d’issue fatale dénommés encéphalopathies spongiformes trans- missibles (EST) ou maladies à prion. Les EST affectent à la fois l’homme (maladie de Creutzfeldt-Jakob [MCJ]) et les ani- maux de rente ou sauvages (tremblante du mouton, encéphalopathie spongi- forme bovine [ESB], maladie du dépéris- sement chronique des cervidés [MDC]). Les prions sont essentiellement compo- sés d’agrégats d’une forme anormale- ment repliée (PrP Sc ) d’une protéine de l’hôte, la protéine prion cellulaire (PrP C ). La PrP C est une protéine membranaire glycosylée, exprimée de façon quasi ubi- quiste dans l’organisme. Sa séquence en acides aminés est très conservée entre les espèces. Bien que son rôle physiolo- gique ne soit pas formellement établi, sa présence apparaît importante au cours du développement embryonnaire [1, 2]. L’invalidation expérimentale de PrP C rend les animaux résistants aux prions. Lors des étapes initiales de l’infection, l’inte- raction physique entre les oligomères de PrP Sc du prion infectant et la PrP C de l’hôte induirait son changement de conformation, notamment un enrichis- sement en feuillets b, enclenchant ainsi un cycle auto-entretenu de multiplica- tion des prions au cours duquel la PrP C produite par les cellules serait convertie par les molécules de PrP Sc précédem- ment néosynthétisées [3]. Dans le cer- veau, cela conduit à l’accumulation de PrP Sc sous forme de dépôts de type amy- loïde, contribuant ainsi aux désordres neurodégénératifs caractéristiques des EST. Bien que ce mode de propagation s’apparente à celui décrit dans d’autres protéinopathies telles que la maladie d’Alzheimer ou de Parkinson, il produit dans le cas des EST un agent authenti- quement infectieux pour la même espèce ou d’autres espèces de mammifères. Compatibilité conformationnelle et barrière d’espèce Comme les pathogènes convention- nels, les prions arborent une diver- sité de souches au sein d’une même espèce-hôte. Ces souches se différen- cient par leurs propriétés biologiques (temps d’incubation chez l’hôte infecté, neuropathologie) et physicochimiques (profil électrophorétique, résistance de la PrP Sc à des agents dénaturants). En l’absence d’acides nucléiques spéci- fiques, la diversité des souches tien- drait à la capacité de la PrP Sc d’exister sous plusieurs conformations stables, capables de transmettre fidèlement une information biologique spécifique [4]. Le support structural de cette information biologique reste à déterminer. Il pourrait impliquer la structure tertiaire et/ou quaternaire de la protéine [5]. Les prions peuvent également se trans- mettre d’une espèce à une autre et pré- senter un risque zoonotique : le variant de la MCJ apparu en 1996 chez l’homme lors de la crise dite de la « vache folle » provient de l’ingestion d’aliments conta- minés par les prions ESB. La capacité des prions à se propager entre espèces différentes est limitée par une barrière, communément appelée barrière d’es- pèce. La recherche de signes neurololo- giques typiques de la présence des EST et de PrP Sc dans le cerveau est le moyen classiquement utilisé pour apprécier la porosité de cette barrière, l’absence concomitante de ces marqueurs tradui- sant la faible réceptivité apparente des animaux inoculés. Au plan moléculaire, la transmission interespèces reposerait sur le degré de compatibilité spatiale entre les conformations de la PrP C de l’hôte et de la PrP Sc du prion infec- tant (la souche). Intrinsèquement, la PrP C de l’hôte pourrait n’adopter qu’un nombre fini de conformations dans sa forme PrP Sc pathologique. La barrière d’espèce sera faible si la PrP Sc infectante appartient à ce panel de conformations possibles. Sinon, la barrière sera forte et ne sera éventuellement franchie que via l’émergence d’un « mutant » compatible susceptible de présenter de nouvelles propriétés biologiques [4, 6]. À l’inverse, l’expression chez une souris transgénique du gène Prnp (codant pour PrP C ) de l’espèce hétérologue donneuse de prion permet d’estomper la barrière de trans- mission entre cette espèce et la souris (Figure 1). Tissu extraneural et transmission interespèces À partir de ce modèle de compatibilité conformationnelle s’est posée la question de l’influence du tissu d’un même hôte sur l’efficacité de transmission interspé- Transmission interespèces des prions Le tissu lymphoïde s’en mêle Vincent Béringue 1 , Jean-Luc Vilotte 2 , Hubert Laude 1 NOUVELLE
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Topo thérapeutique Angelman ms Juin2012

médecine/sciences
m/s n° 6-7, vol. 28, juin-juillet 2012DOI : 10.1051/medsci/2012286002
médecine/sciences 2012 ; 28 : 000-00
NOUV
ELLE
SM
AGAZ
INE
1
1 UR892 Inra (Institut national de la recherche agronomique), virologie et immunologie moléculaires, 78350 Jouy-en Josas, France ; 2 UMR1313 INRA, génétique animale et biologie intégrative, 78350 Jouy-en-Josas, [email protected]
Un agent infectieux de nature protéiqueLe prion est le pathogène responsable, chez les mammifères, de désordres neu-rodégénératifs d’issue fatale dénommés encéphalopathies spongiformes trans-missibles (EST) ou maladies à prion. Les EST affectent à la fois l’homme (maladie de Creutzfeldt-Jakob [MCJ]) et les ani-maux de rente ou sauvages (tremblante du mouton, encéphalopathie spongi-forme bovine [ESB], maladie du dépéris-sement chronique des cervidés [MDC]).Les prions sont essentiellement compo-sés d’agrégats d’une forme anormale-ment repliée (PrPSc) d’une protéine de l’hôte, la protéine prion cellulaire (PrPC). La PrPC est une protéine membranaire glycosylée, exprimée de façon quasi ubi-quiste dans l’organisme. Sa séquence en acides aminés est très conservée entre les espèces. Bien que son rôle physiolo-gique ne soit pas formellement établi, sa présence apparaît importante au cours du développement embryonnaire [1, 2]. L’invalidation expérimentale de PrPC rend les animaux résistants aux prions. Lors des étapes initiales de l’infection, l’inte-raction physique entre les oligomères de PrPSc du prion infectant et la PrPC de l’hôte induirait son changement de conformation, notamment un enrichis-sement en feuillets b, enclenchant ainsi un cycle auto-entretenu de multiplica-tion des prions au cours duquel la PrPC produite par les cellules serait convertie par les molécules de PrPSc précédem-ment néosynthétisées [3]. Dans le cer-veau, cela conduit à l’accumulation de
PrPSc sous forme de dépôts de type amy-loïde, contribuant ainsi aux désordres neurodégénératifs caractéristiques des EST. Bien que ce mode de propagation s’apparente à celui décrit dans d’autres protéinopathies telles que la maladie d’Alzheimer ou de Parkinson, il produit dans le cas des EST un agent authenti-quement infectieux pour la même espèce ou d’autres espèces de mammifères.
Compatibilité conformationnelle et barrière d’espèceComme les pathogènes convention-nels, les prions arborent une diver-sité de souches au sein d’une même espèce-hôte. Ces souches se différen-cient par leurs propriétés biologiques (temps d’incubation chez l’hôte infecté, neuropathologie) et physicochimiques (profil électrophorétique, résistance de la PrPSc à des agents dénaturants). En l’absence d’acides nucléiques spéci-fiques, la diversité des souches tien-drait à la capacité de la PrPSc d’exister sous plusieurs conformations stables, capables de transmettre fidèlement une information biologique spécifique [4]. Le support structural de cette information biologique reste à déterminer. Il pourrait impliquer la structure tertiaire et/ou quaternaire de la protéine [5].Les prions peuvent également se trans-mettre d’une espèce à une autre et pré-senter un risque zoonotique : le variant de la MCJ apparu en 1996 chez l’homme lors de la crise dite de la « vache folle » provient de l’ingestion d’aliments conta-minés par les prions ESB. La capacité
des prions à se propager entre espèces différentes est limitée par une barrière, communément appelée barrière d’es-pèce. La recherche de signes neurololo-giques typiques de la présence des EST et de PrPSc dans le cerveau est le moyen classiquement utilisé pour apprécier la porosité de cette barrière, l’absence concomitante de ces marqueurs tradui-sant la faible réceptivité apparente des animaux inoculés. Au plan moléculaire, la transmission interespèces reposerait sur le degré de compatibilité spatiale entre les conformations de la PrPC de l’hôte et de la PrPSc du prion infec-tant (la souche). Intrinsèquement, la PrPC de l’hôte pourrait n’adopter qu’un nombre fini de conformations dans sa forme PrPSc pathologique. La barrière d’espèce sera faible si la PrPSc infectante appartient à ce panel de conformations possibles. Sinon, la barrière sera forte et ne sera éventuellement franchie que via l’émergence d’un « mutant » compatible susceptible de présenter de nouvelles propriétés biologiques [4, 6]. À l’inverse, l’expression chez une souris transgénique du gène Prnp (codant pour PrPC) de l’espèce hétérologue donneuse de prion permet d’estomper la barrière de trans-mission entre cette espèce et la souris (Figure 1).
Tissu extraneural et transmission interespècesÀ partir de ce modèle de compatibilité conformationnelle s’est posée la question de l’influence du tissu d’un même hôte sur l’efficacité de transmission interspé-
Transmission interespèces des prionsLe tissu lymphoïde s’en mêleVincent Béringue1, Jean-Luc Vilotte2, Hubert Laude1
NOUVELLE

m/s n° 6-7, vol. 28, juin-juillet 2012 2
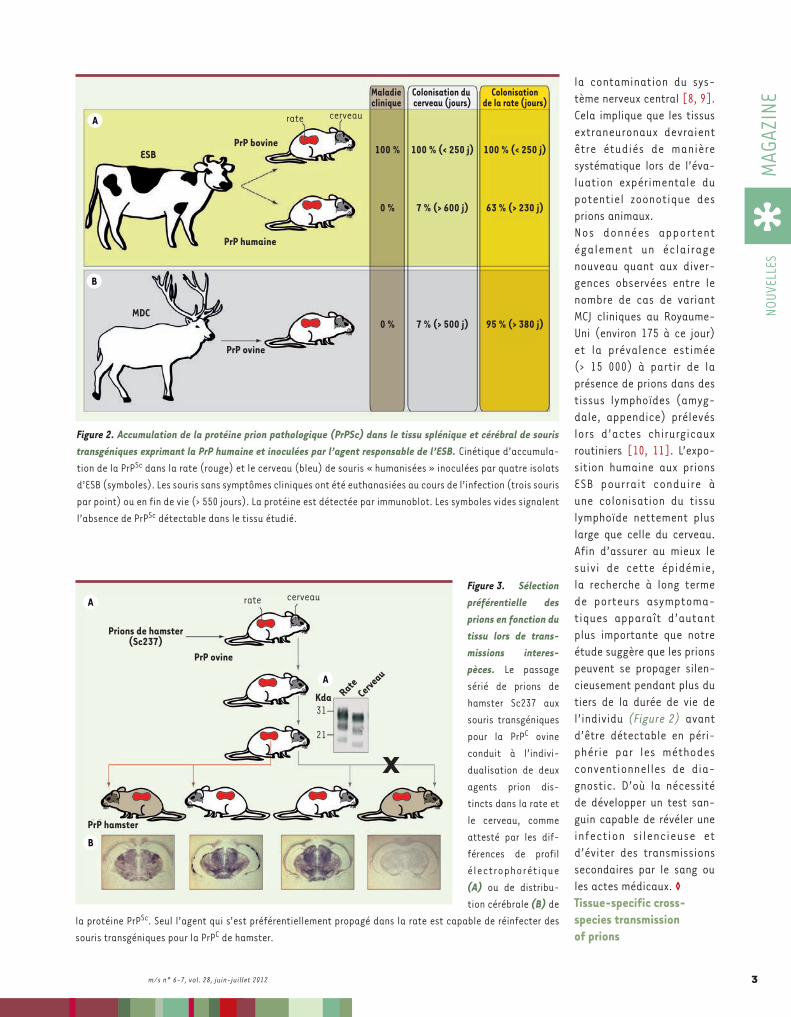
cet agent Sc237 sur la séquence ovine a aussi permis de montrer que, lorsque la barrière d’es-pèce a finalement été franchie par l’émergence d’un prion « mutant » dans le cerveau des animaux inoculés, un agent dif-férent se propageait dans la rate, se distinguant notamment par sa capacité unique à réin-fecter l’hôte de départ (souris transgéniques pour la PrPC du hamster) (Figure 3).Pourquoi la rate apparaît-elle plus permissive que le cerveau à un prion étranger ? Par ana-logie avec le concept de quasi-espèce en virologie1, les prions pourraient héberger un conti-nuum de conformations de PrPSc [4]. Certaines pourraient-elles être plus ou moins compatibles avec les différents tissus ? Des facteurs spécifiques des tis-sus de l’hôte pourraient aussi réguler la dynamique de répli-cation du prion. Ainsi, la PrPC, du fait de variations de confor-mations, ou d’autres créant un environnement favorable, permettraient par exemple une persistance accrue et propice à l’initiation de la conversion hétérospécifique. Quel que soit le mécanisme mis en jeu, notre
étude révèle une capacité d’adaptation insoupçonnée de ce pathogène neu-rotrope lors d’une pression sélective, lui permettant au final d’étendre son spectre d’hôte.
Perspectives pour la santé publiqueLa barrière d’espèce prions ESB-homme apparaît ainsi moins robuste qu’ini-tialement anticipé avec des modèles expérimentaux humanisés focalisés sur
1 Les virus à ARN expriment un taux élevé de mutations - dues aux erreurs de réplication - ce qui entraîne une grande hétérogénétié dans les populations, mais ces variants conti-nuent d’évoluer comme si ils ne formaient « quasiment une seule et même espèce » (espèce au sens d’espèce chimique ou moléculaire et non au sens d’espèce biologique).
dans le cerveau de souris transgéniques pour la PrPC ovine et humaine et non plus murine, les prions se sont quasiment exclusivement multipliés dans le tissu lymphoïde, en l’occurrence la rate, et ce relativement précocement au regard de la durée de vie de l’animal (Figures 1 et 2). Quantitativement, la barrière d’espèce s’est avérée être environ dix fois moins étanche dans la rate que dans le cerveau (Figures 1 et 2). De plus, l’ino-culation à la souris transgénique pour la PrPC ovine de prions de hamster (souche Sc237) a dévoilé que cette facilitation de la réplication splénique était indépen-dante du lymphotropisme intrinsèque de l’agent inoculé. L’adaptation sériée de
cifique des prions, du fait par exemple de variations moléculaires de la protéine PrPC ou d’un environnement cellulaire différent. Le tissu lymphoïde était déjà connu pour être un lieu de transit, quelle que soit la voie d’entrée du prion dans l’organisme [12], et être permissif à un certain nombre de souches lors de transmissions intraespèces. Nous nous sommes donc focalisés sur ce tissu et avons étudié sa permissivité relative par rapport au tissu nerveux lors de transmissions interespèces en apparence infructueuses, destinées à modéliser une transmission bovin-homme ou cervidé-ovin [7]. Lorsque les prions responsables de la MDC ou de l’ESB ont été injectés
Maladieclinique
ESB
MDC
rate cerveau
100 %
0 %
0 %
100 % (< 250 j)
7 % (> 600 j)
0 % 7 % (> 500 j)
0 % 95 % (> 380 j)
63 % (> 230 j)
PrP bovine
PrP ovine
PrP humaine
Colonisationdu tissu (jours)
A
B
Figure 1. Efficacité de la transmission interespèces des prions en fonction du tissu. Les souris transgéniques exprimant la PrPC bovine sont parfaitement susceptibles à l’agent responsable de l’encéphalopathie spon-giforme bovine (ESB) (A). En revanche, les souris transgéniques exprimant la PrPC humaine ne développent pas de symptômes cliniques après inoculation du même agent (A). Il en va de même de souris transgéniques exprimant la PrPC ovine inoculées par l’agent responsable de la maladie du dépérissement chronique des cervidés (MDC) (B). Signe supplémentaire d’une forte barrière d’espèce apparente, un nombre très limité de souris transgéniques pour pour la PrPC ovine et humaine, toutes en fin de vie, accumule de la protéine prion pathologique dans le cerveau (AB). Néanmoins, une grande majorité de souris analysées présente des taux détectables de protéine prion pathologique dans la rate, et ce dès une année postinfection (AB).
m/s n° 6-7, vol. 28, juin-juillet 2012
NOUV
ELLE
SM
AGAZ
INE
3
la contamination du sys-tème nerveux central [8, 9]. Cela implique que les tissus extraneuronaux devraient être étudiés de manière systématique lors de l’éva-luation expérimentale du potentiel zoonotique des prions animaux.Nos données apportent également un éclairage nouveau quant aux diver-gences observées entre le nombre de cas de variant MCJ cliniques au Royaume-Uni (environ 175 à ce jour) et la prévalence estimée (> 15 000) à partir de la présence de prions dans des tissus lymphoïdes (amyg-dale, appendice) prélevés lors d’actes chirurgicaux routiniers [10, 11]. L’expo-sition humaine aux prions ESB pourrait conduire à une colonisation du tissu lymphoïde nettement plus large que celle du cerveau. Afin d’assurer au mieux le suivi de cette épidémie, la recherche à long terme de porteurs asymptoma-tiques apparaît d’autant plus importante que notre étude suggère que les prions peuvent se propager silen-cieusement pendant plus du tiers de la durée de vie de l’individu (Figure 2) avant d’être détectable en péri-phérie par les méthodes conventionnelles de dia-gnostic. D’où la nécessité de développer un test san-guin capable de révéler une infection silencieuse et d’éviter des transmissions secondaires par le sang ou les actes médicaux. ◊Tissue-specific cross-species transmission of prions
Figure 3. Sélection préférentielle des prions en fonction du tissu lors de trans-missions interes-pèces. Le passage sérié de prions de hamster Sc237 aux souris transgéniques pour la PrPC ovine conduit à l’indivi-dualisation de deux agents prion dis-tincts dans la rate et le cerveau, comme attesté par les dif-férences de profil électrophorétique (A) ou de distribu-tion cérébrale (B) de
la protéine PrPSc. Seul l’agent qui s’est préférentiellement propagé dans la rate est capable de réinfecter des souris transgéniques pour la PrPC de hamster.
Maladieclinique
ESB
MDC
rate cerveau
100 %
0 %
100 % (< 250 j)
7 % (> 600 j)
0 % 7 % (> 500 j) 95 % (> 380 j)
63 % (> 230 j)
100 % (< 250 j)PrP bovine
PrP ovine
PrP humaine
Colonisation du cerveau (jours)
Colonisationde la rate (jours)
A
B
Prions de hamster(Sc237)
rate cerveau
21
31Kda Rate
Cerveau
X
PrP ovine
PrP hamster
A
A
B
Figure 2. Accumulation de la protéine prion pathologique (PrPSc) dans le tissu splénique et cérébral de souris transgéniques exprimant la PrP humaine et inoculées par l’agent responsable de l’ESB. Cinétique d’accumula-tion de la PrPSc dans la rate (rouge) et le cerveau (bleu) de souris « humanisées » inoculées par quatre isolats d’ESB (symboles). Les souris sans symptômes cliniques ont été euthanasiées au cours de l’infection (trois souris par point) ou en fin de vie (> 550 jours). La protéine est détectée par immunoblot. Les symboles vides signalent l’absence de PrPSc détectable dans le tissu étudié.

m/s n° 6-7, vol. 28, juin-juillet 2012 4
NOUVELLE
Un transporteur du magnésiumNouvel intermédiaire dans les voies de signalisation du TCRGilles Besin1,2
1 In-Cell-Art ; 2Institut du thorax, UMR 1087 Inserm/UMR 6291 CNRS/IRT-UN, 8, quai Moncousu, 44007 Nantes Cedex 1, [email protected]
> Les ions magnésium (Mg2+) sont les cations les plus abondants dans les cellules mammifères et constituent un cofacteur essentiel pour l’ATP, les acides nucléiques, et de nombreuses enzymes chez les animaux et les plantes [1, 2]. Toutefois, contrairement aux ions calcium (Ca2+) pour lesquels les concentrations intracellulaires d’ions libres ([Ca2+]Int environ 0,1 mM) et les concentrations extracellulaires ([Ca2+]
ex environ 1 mm) permettent des mou-vements et flux ioniques, aucune étude n’a démontré clairement le rôle des ions Mg2+ en tant que messager secondaire dans la signalisation intracellulaire. Les ions magnésium intracellulaires ([Mg2+]Int environ 20 mm) sont surtout com-plexés avec l’ATP, l’ADN, des enzymes ou d’autres molécules, et seulement 1 à 5 % de ces ions magnésium cellulaires (0,2-1 mm) sont sous une forme libre dans le cytosol. Fait intéressant, dans diverses études, l’utilisation de sondes sensibles aux flux d’ions magnésium a permis de démontrer des changements
dans la concentration intracellulaire de ces ions dans certaines cellules du système immunitaire comme les lym-phocytes (après stimulation par des lectines) [3-5].Le système immunitaire représente la principale défense contre les infections virales, bactériennes ou les cancers et tout défaut du système immunitaire entraîne une sensibilité à ces agressions pouvant avoir de graves conséquences. Le décryptage moléculaire de multiples déficits immunitaires primaires révèle souvent de nouveaux acteurs interve-nant dans l’activation lymphocytaire. Ainsi, en étudiant trois patients immu-nodéficients, l’équipe de M. Lenardo (NIH, Bethesda, États-Unis) a identi-fié récemment Magnesium Tranporter Protein 1 (MAGT1) comme un nouveau maillon de la voie d’activation des lym-phocytes T [6].L’activation des lymphocytes T est déclenchée par l’interaction entre le récepteur des lymphocytes T (TCR) et une molécule du complexe majeur
d’histocompatibilité (CMH) chargée d’un peptide antigénique, exprimée à la surface d’une cellule présentatrice d’antigènes (APC). Cet engagement du TCR entraîne l’activation des enzymes kinases Lck, ZAP-70 (zeta-chain-asso-ciated protein kinase of 70 kDa) et Itk (leukocyte tyrosine kinase) qui phos-phorylent la protéine PLCg1 (phospho-lipase Cg) ainsi que d’autres protéines. La phosphorylation de la PLCg entraîne l’augmentation des taux d’ions calcium intracellulaires ([Ca2+]Int) et l’activa-tion de plusieurs facteurs de transcrip-tion : NFAT [nuclear factor of activated T-cells], NF-kB, etc. Des mutations de ces molécules clés de l’activation de la voie du TCR (Lck, ZAP-70, Itk, LAT [lin-ker of activation of T cell], PLCg1) ont été identifiées chez plusieurs patients immunodéficients [7].
Un nouveau déficit immunitaire lié à l’XLe groupe du Pr Lenardo a étudié trois patients issus de deux familles
8. Asante EA, Linehan JM, Desbruslais M, et al. BSE prions propagate as either variant CJD-like or sporadic CJD-like prion strains in transgenic mice expressing human prion protein. EMBO J 2002 ; 21 : 6358-66.
9. Bishop MT, Hart P, Aitchison L, et al. Predicting susceptibility and incubation time of human-to-human transmission of vCJD. Lancet Neurol 2006 ; 5 : 393-8.
10. Health Protection Agency, UK. http://www.hpa.org.uk/hpr/archives/2011/news3611.htm.
11. Hilton DA, Ghani AC, Conyers L, et al. Prevalence of lymphoreticular prion protein accumulation in UK tissue samples. J Pathol 2004 ; 203 : 733-9.
12. Bachy V, Aucouturier P. Maladies à prions. Quel rôle pour les cellules dendritiques dans la pathogenèse des formes transmises ? Med Sci (Paris) 2010 ; 26 : 615-20.
2. Malaga-Trillo E, Solis GP, Schrock Y, et al. Regulation of embryonic cell adhesion by the prion protein. PLoS Biol 2009 ; 7 : e55.
3. Caughey B, Baron GS, Chesebro B, Jeffrey M. Getting a grip on prions: oligomers, amyloids, and pathological membrane interactions. Annu Rev Biochem 2009 ; 78 : 177-204.
4. Collinge J, Clarke AR. A general model of prion strains and their pathogenicity. Science 2007 ; 318 : 930-6.
5. Tixador P, Herzog L, Reine F, et al. The physical relationship between infectivity and prion protein aggregates is strain-dependent. PLoS Pathog 2010 ; 6 : e1000859.
6. Beringue V, Vilotte JL, Laude H. Prion agent diversity and species barrier. Vet Res 2008 ; 39 : 47.
7. Beringue V, Herzog L, Jaumain E, et al. Facilitated cross-species transmission of prions in extraneural tissue. Science 2012 ; 335 : 472-5.
LIENS D’INTÉRÊTLes auteurs déclarent n’avoir aucun lien d’intérêt concernant les données publiées dans cet article.
REMERCIEMENTSLe travail résumé dans cet article a bénéficié en partie du soutien d’INRA-transfert et de la Fondation Alliance Biosecure. Merci à Claire Laude pour l’iconographie.
RÉFÉRENCES
1. Khalife M, Young R, Passet B, et al. Transcriptomic analysis brings new insight into the biological role of the prion protein during mouse embryogenesis. PLoS One 2011 ; 6 : e23253.

m/s n° 6-7, vol. 28, juin-juillet 2012
NOUV
ELLE
SM
AGAZ
INE
5
confirmée par séquençage haut débit et capture d’exon, qui ont mis en évidence une délétion de 10 paires de bases au sein du gène MAGT1 des deux frères, absente chez la mère, mais présente chez la grand-mère et l’arrière grand-mère des 2 patients. Cette délétion sup-prime un site donneur d’épissage situé à la jonction exon-intron de l’exon 7. Les transcrits sont effondrés et la protéine indétectable. Chez le troisième patient, il s’agissait d’une mutation non-sens de l’exon 3 de MAGT1.
Implication fonctionnelle du gène MAGT1Le gène MAGT1 code pour un trans-porteur membranaire du magnésium, MAGT1, dont les fonctions physiolo-giques ne sont pas encore totalement élucidées. Alors que la stimulation du TCR induit une entrée massive des ions Mg2+ dans les cellules T normales [3-5], cet influx est indétectable dans les lym-phocytes T des patients. Une analyse précise a montré que ce défaut était très spécifique : il n’affectait pas les processus métaboliques cellulaires pour lesquels le Mg2+ est un cofacteur, mais intervenait uniquement sur les événe-ments d’activation en aval de la phos-phorylation de la protéine PLC-g1 qui est diminuée dans les lymphocytes T des patients. Les fonctions des lympho-
différentes. Le déficit immunitaire de ces patients était classé comme lym-phopénie CD4 idiopathique (ICL)1 et caractérisé par des taux bas de lym-phocytes T CD4+ alors que les autres populations de lymphocytes étaient présentes en nombre normal ou élevé et que les taux d’immunoglobulines et les réponses vaccinales n’étaient anormales que de façon intermittente. Le rapport des lymphocytes CD4/CD8 était inversé et une réduction des cellules CD31+ au sein de la population des lymphocytes T CD4+ naïfs suggérait une diminution de la production thymique. L’activation des lymphocytes T en réponse au signal du TCR ou à un anti-CD3 agoniste était très anormale et le défaut a été localisé précisément à une étape précoce de la cascade d’activation. La réponse immu-nitaire humorale était normale. Clini-quement, ces patients étaient atteints d’infections récurrentes mal contrô-lées, mais essentiellement virales, en particulier par le virus d’Epstein Barr.Orientés par l’atteinte de deux gar-çons dans une des deux familles et un biais d’inactivation du chromosome X chez la mère, les auteurs ont suspecté une atteinte liée à l’X. Celle-ci a été
1 Défini par un taux de CD4+ < 300/mm3 ou < 20 % du total de lymphocytes en l’absence d’infection par le VIH ou d’autres causes évidentes de lymphopénie.
cytes B, qui expriment le gène MAGT1, n’étaient pas touchées chez ces patients par l’absence de fonction du transpor-teur MAGT1 (mais c’est la PLCg2 qui est impliquée dans l’activation du BCR). Des expériences complémentaires in vitro de délétion transitoire du gène MAGT1 dans des lymphocytes normaux, par ARN interférence et de restauration de l’expression du gène MAGT1 nor-mal via des vecteurs lentiviraux dans les lymphocytes T de ces patients ont confirmé le rôle du transporteur MAGT1 dans ces défauts d’activation lympho-cytaire. Curieusement, cette perte de flux ionique d’ions Mg2+ était associée à une baisse de l’influx de Ca2+ qui, lui, est tout à fait connu lors de l’activa-tion T. Cette baisse concomitante de l’influx calcique n’est pas complètement expliquée.Des études antérieures avaient déjà démontré le rôle important des ions Mg2+ pour l’activation des lymphocytes T [8, 9]. Des influx de magnésium ont aussi été observés dans deux lignées de cellules épithéliales en réponse à l’EGF (epidermal growth factor) qui agit aussi via la PLCg1. dans ce modèle aussi, des flux de calcium accompagnent l’influx de Mg2+, comme lors de l’activation des lymphocytes T via le TCR. Cette corré-lation entre les deux influx ioniques n’existe pas lorsque d’autres PLC sont impliquées, PLCg2 ou PLCb.
ConclusionLe travail de ce groupe révèle l’impor-tance physiologique du transporteur MAGT1, qui relaie un influx transitoire de Mg2+ déclenché par l’activation d’un récepteur. Cet influx joue un rôle de second messager dans les lymphocytes T activés (via l’activation de la PLCg1 et l’influx de Ca2+) mais aussi dans les cellules épithéliales en réponse à l’EGF. Comme souvent en immunologie, c’est l’exploration d’un déficit immunitaire qui a permis cette découverte. Il faut toutefois démontrer que la diminution de la réponse des lymphocytes T asso-ciée à une diminution de la fonction de
Zap70Lck
Antigène
TCRCD4MAGT1
Influx de Mg2+
Mg2+
PLC g1
LATltk
SLP76
Ca2+
PiP2
Figure 1. Localisation de l’action de l’influx de Mg2+ relayée par le transporteur MAGT1 dans l’activation lymphocytaire T.

m/s n° 6-7, vol. 28, juin-juillet 2012 6
4. Modiano JF, Kelepouris E, Kern JA, Nowell PC. Requirement for extracellular calcium or magnesium in mitogen-induced activation of human peripheral blood lymphocytes. J Cell Physiol 1988 ; 135 : 451-8.
5. Whitney RB, Sutherland RM. The influence of calcium, magnesium and cyclic adenosine 3’, 5’-monophosphate on the mixed lymphocyte reaction. J Immunol 1972 ; 108 : 1179-83.
6. Li FY, Chaigne-Delalande B, Kanellopoulou C, et al. Second messenger role for Mg2+ revealed by human T-cell immunodeficiency. Nature 2011 ; 475 : 471-6.
7. Casanova JL, Abel L. Primary immunodeficiencies: a field in its infancy. Science 2007 ; 317 : 617-9.
8. Rijkers GT, Griffioen AW. Changes in free cytoplasmic magnesium following activation of human lymphocytes. Biochem J 1993 ; 289 : 373-7.
9. Rijkers GT, Henriquez N, Griffioen AW. Intracellular magnesium movements and lymphocyte activation. Magnes Res 1993 ; 6 : 205-13.
puissent être dus à l’absence de fonction de MAGT1. ◊Magnesium transporter protein 1, a new intermediate in the TCR signaling
LIENS D’INTÉRÊTL’auteur déclare n’avoir aucun lien d’intérêt concer-nant les données publiées dans cet article.
RÉFÉRENCES
1. Cowan JA. Structural and catalytic chemistry of magnesium-dependent enzymes. Biometals 2002 ; 15 : 225-35.
2. Yang W, Lee JY, Nowotny M. Making and breaking nucleic acids: two-Mg2+-ion catalysis and substrate specificity. Mol Cell 2006 ; 22 : 5-13.
3. Abboud CN, Scully SP, Lichtman AH, et al. The requirements for ionized calcium and magnesium in lymphocyte proliferation. J Cell Physiol 1985 ; 122 : 64-72.
MAGT1 est uniquement due à la baisse des flux de magnésium. Il se pourrait que le transporteur MAGT1 possède d’autres fonctions importantes, mais non encore identifiées, dans l’activation des lymphocytes T.Enfin, il est curieux que les fonctions des lymphocytes B, qui expriment le gène MAGT1, ne semblent pas touchées chez ces patients par l’absence de fonction du transporteur MAGT1. Il serait intéres-sant de compléter l’analyse de l’expres-sion de MAGT1 dans les autres cellules du système immunitaire et de détermi-ner les éventuels défauts de fonction de ces dernières. Enfin, on ne peut exclure que d’autres défauts des lymphocytes T
Inserm UMR1037, centre de recherche en cancérologie de Toulouse, Toulouse, France ; université de Toulouse, 118 Route de Narbonne, 31062 Toulouse Cedex 09, France. CNRS équipe de recherche labellisée (ERL) 5294, Toulouse, [email protected]
p27KIP1 : un inhibiteur des complexes cycline/CDKLa progression dans le cycle de divi-sion cellulaire est régie par l’activa-tion séquentielle des complexes cycline/CDK (cyclin-dependent kinases). Ces complexes sont finement régulés à de multiples niveaux, notamment par des inhibiteurs de CDK dont p27KIP1 (cyclin-dependent kinase inhibitor 1B ou p27) [1, 2]. L’importance de p27 dans le contrôle des transitions de G0 (quiescence) à G1, et de G1 à S a été abondamment décrite depuis le clonage du gène codant pour p27 en 1994 [1]. Le niveau de p27 est élevé dans les cellules quiescentes, et diminue lors de la transition G1/S pour rester faible dans les phases S, G2 et M du cycle cellulaire. La dégradation de p27 est déclenchée par sa phosphorylation par le complexe cycline E/CDK2, ce qui crée un site de reconnaissance pour l’E3
ubiquitine ligase Skp2 (S-phase kinase-associated protein 2) [1]. L’invalidation chez la souris du gène cdkn1b, qui code pour p27, souligne l’importance de p27
dans le contrôle de la prolifération cel-lulaire puisque la taille de ces animaux augmente d’environ 30 %, et que cette croissance s’accompagne d’une hyper-plasie de divers organes et d’une pré-disposition à la tumorigenèse spontanée ou induite par des carcinogènes [1]. De plus, la perte de l’expression nucléaire de p27 est un facteur de mauvais pronostic dans de nombreux types de cancers chez l’humain [1]. En outre, la diminution nucléaire de p27 n’est pas associée à des mutations génétiques, comme pour les suppresseurs de tumeurs classiques, mais est causée par l’augmentation de sa dégradation protéolytique ou par son exclusion du noyau où il inhibe les complexes cycline/CDK.
p27KIP1 : protéine multifonctionnelleDe nombreuses études indiquent que le rôle de p27 ne se limite pas à l’inhibition du cycle cellulaire, et que cette protéine participe en fait à la régulation d’autres processus cellulaires via son interaction avec divers partenaires protéiques [1, 2]. En effet, p27 est impliquée dans le contrôle de la migration cellulaire, de l’apoptose, de la transcription et du devenir des cellules souches pro-génitrices [1, 3, 4]. Afin d’étudier les fonctions indépendantes des complexes cycline/CDK de p27, nous avons généré des souris knock-in exprimant un allèle de p27 incapable de lier les cyclines et les CDK (p27CK-) [2, 4, 5]. À l’in-verse des souris p27-/- qui présentent des tumeurs spontanées uniquement au niveau de l’hypophyse, les souris p27CK-
développent des tumeurs dans divers organes dont les poumons. Ce modèle
Un nouveau rôle de p27KIP1 en mitose ?Bérénice Leclercq, Arnaud Besson
NOUVELLE

m/s n° 6-7, vol. 28, juin-juillet 2012
NOUV
ELLE
SM
AGAZ
INE
7
d’actomyosine et du midbody. De plus, la surexpression du domaine minimal de citron-K qui lie p27 est suffisante pour empêcher le phénotype de multinu-cléation causé par p27CK-. L’expression d’un mutant de p27CK- incapable de lier citron-K n’entraîne plus non plus de multinucléation [10]. Ces observations confirment donc au niveau fonctionnel l’importance de l’interaction citron-K/p27 dans le phénotype des cellules p27CK-. Enfin, il apparaît que l’interac-tion de p27 avec citron-K empêche cette dernière d’interagir avec son activateur Rho [10]. Par conséquent, p27 semble impliquée dans les phases finales de la cytocinèse via la régulation de l’activa-tion de citron-K par les protéines Rho.Ces résultats fournissent un mécanisme potentiel qui pourrait expliquer la sus-ceptibilité accrue des souris p27CK- à la tumorigenèse [4]. Chez l’homme, des études cliniques associent la locali-sation cytoplasmique de p27 avec des tumeurs agressives, de haut grade, et la présence de métastases [1]. Par consé-quent, dans les cellules tumorales où p27 est principalement cytoplasmique, l’entrée en mitose avec un niveau élevé de p27 pourrait alors causer des défauts de cytocinèse, entraînant la multi-nucléation et la polyploïdie, contri-buant ainsi à promouvoir l’instabilité génétique.Plusieurs questions restent en suspens : quelle est l’importance de ce nouveau rôle de p27 au niveau physiologique dans une cellule normale ? Comment est régulée l’interaction de p27 avec citron-K ? Une hypothèse particulière-ment intéressante serait que p27 soit capable de cibler des complexes cycline/CDK vers des protéines spécifiques du midbody (comme citron-K) ou puisse inhiber l’activité de ces complexes au midbody, à un moment donné, pour réguler le processus d’abscission. De manière intéressante, lors d’un criblage protéomique réalisé afin d’identifier les protéines présentes au midbody, CDK1 et CDK4 ont effectivement été trouvées [11]. Reste à comprendre le
tissus, dont le foie et les reins [10]. De plus, l’expression de p27CK- est capable d’induire la multinucléation de cel-lules en culture [10]. Ces résultats indiquent donc que p27 semble jouer un rôle indépendant des cycline/CDK en fin de mitose, lors de l’étape finale de la division cellulaire qui conduit à la sépa-ration des cellules filles : la cytocinèse.La cytocinèse est divisée en plusieurs étapes incluant l’assemblage de l’an-neau contractile d’actomyosine, sa contraction au niveau du sillon de cli-vage jusqu’à la formation d’une zone dense (midbody) au niveau de la zone d’interdigitation des microtubules du fuseau mitotique. La phase finale de la cytocinèse est l’abscission où les cellules se séparent physiquement. Des analyses de vidéomicroscopie ont révélé qu’une fraction de fibroblastes primaires issus de souris p27CK- effec-tuent une mitose normale, mais ont un défaut de fin de cytocinèse, lors de l’abscission, avec une réouverture du pont intercellulaire [10].Par ailleurs, nous avons identifié la pro-téine citron kinase (citron-K) lors d’un criblage protéomique visant à identifier des partenaires de p27 [10]. Citron-K est une Ser/Thr kinase effectrice des GTPases Rho qui possède plusieurs domaines impliqués dans de nombreuses interactions protéine/protéine [10]. Citron-K joue un rôle essentiel dans la cytocinèse et est localisée sur l’anneau d’actomyosine du sillon de clivage, et au midbody. Chez la drosophile et dans les cellules HeLa, la perte de fonction de citron-K entraîne un défaut d’abscission et une réouverture du pont intercel-lulaire, conduisant à la formation de cellules multinucléées.L’interaction de p27 avec citron-K a été confirmée in vitro et in vivo, et le domaine d’interaction de p27 sur citron-K a été cartographié au niveau d’une région qui participe à l’interaction de citron-K avec son activateur Rho [10]. Des expériences d’immunofluorescence ont également permis de colocaliser p27 et citron-K au niveau de l’anneau
murin a donc révélé un rôle oncogénique pour p27, indépendant de ses fonctions d’inhibiteur de cycline/CDK.Une autre caractéristique de la protéine p27CK- est qu’en absence de liaison aux complexes cycline/CDK, Skp2 est inca-pable de s’y lier, ce qui empêche son ubiquitinylation et sa dégradation. Par conséquent, le niveau de p27CK- est anormalement élevé dans les phases S/G2/M du cycle cellulaire [5].
Rôles de p27 dans les phases G2/MAlors que le rôle de p27 à la transi-tion G1/S est abondamment décrit, son implication dans les phases tardives du cycle cellulaire l’est beaucoup moins. Néanmoins, plusieurs études indiquent que p27 joue un rôle important dans le contrôle de l’entrée en mitose. Les souris Skp2-/- présentent un phénotype de polyploïdie, une amplification des centrosomes et un défaut de proliféra-tion qui sont absents dans les animaux Skp2-/-/p27-/-, indiquant que l’accumu-lation de p27 en phases G2/M causée par la perte de Skp2 est responsable d’un défaut de progression en mitose [6, 7]. p27 est aussi impliquée dans l’arrêt des cellules en phases G2/M en réponse aux dommages à l’ADN [8, 9]. p27 poten-tialise également l’accumulation de la protéine Rad51 lors de la réparation des cassures double brin de l’ADN. En effet, elle inhibe la phosphorylation de BRCA2 (breast cancer 2) par le com-plexe cycline-B/CDK1, ce qui empêche la formation des complexes BRCA2/Rad51 et la réparation des dommages à l’ADN dépendante de Rad51 [9].Nous avons donc utilisé le fait que le niveau de p27CK- est comparable à celui de p27 dans les souris Skp2-/- pour déterminer si p27 intervenait au cours de la mitose indépendamment des cycline/CDK.
Un nouveau rôle de p27 dans le contrôle de la cytocinèse via la régulation de citron kinaseLes souris p27CK- présentent un phéno-type de multinucléation dans plusieurs

m/s n° 6-7, vol. 28, juin-juillet 2012 8
7. Kossatz U, Dietrich N, Zender L, et al. Skp2-dependent degradation of p27kip1 is essential for cell cycle progression. Genes Dev 2004 ; 18 : 2602-7.
8. Cuadrado M, Gutierrez-Martinez P, Swat A, et al. p27Kip1 stabilization is essential for the maintenance of cell cycle arrest in response to DNA damage. Cancer Res 2009 ; 69 : 8726-32.
9. See WL, Miller JP, Squatrito M, et al. Defective DNA double-strand break repair underlies enhanced tumorigenesis and chromosomal instability in p27-deficient mice with growth factor-induced oligodendrogliomas. Oncogene 2010 ; 29 : 1720-31.
10. Serres MP, Kossatz U, Chi Y, et al. p27(Kip1) controls cytokinesis via the regulation of citron kinase activation. J Clin Invest 2012 ; 122 : 844-58.
11. Skop AR, Liu H, Yates J 3rd, et al. Dissection of the mammalian midbody proteome reveals conserved cytokinesis mechanisms. Science 2004 ; 305 : 61-6.
2. Besson A. p27Kip1, suppresseur de tumeur… et oncogène ? Med Sci (Paris) 2007 ; 23 : 1089-91.
3. Pippa R, Espinosa L, Gundem G, et al. p27(Kip1) represses transcription by direct interaction with p130/E2F4 at the promoters of target genes. Oncogene 2011 ; 19 décembre. doi: 10.1038/onc.2011.582.
4. Besson A, Hwang HC, Donovan SL, et al. Discovery of an oncogenic activity in p27Kip1 that causes stem cell expansion and a multiple tumor phenotype. Genes Dev 2007 ; 21 : 1731-46.
5. Besson A, Gurian-West M, Chen X, et al. A pathway in quiescent cells that controls p27Kip1 stability, subcellular localization and tumor suppression. Genes Dev 2006 ; 20 : 47-64.
6. Nakayama K, Nagahama H, Minamishima YA, et al. Skp2-mediated degradation of p27 regulates progression into mitosis. Dev Cell 2004 ; 6 : 661-72.
rôle de chacune de ces protéines dans la cytocinèse et, plus particulièrement, dans l’interaction entre p27 et citron-K ou RhoA. ◊A new function for p27KIP1 into mitosis
LIENS D’INTÉRÊTLes auteurs déclarent n’avoir aucun lien d’intérêt concernant les données publiées dans cet article.
RÉFÉRENCES
1. Besson A, Dowdy SF, Roberts JM. CDK inhibitors: cell cycle regulators and beyond. Dev Cell 2008 ; 14 : 159-69.
NOUVELLE
Topo thérapeutique pour le syndrome d’Angelman ?Bernard Dan1,2, Karine Pelc1, Guy Chéron1,2
1 Hôpital universitaire des Enfants Reine Fabiola, 15, avenue J.J. Crocq, 1020 Bruxelles, Belgique ;2 Université libre de Bruxelles, faculté des sciences de la motricité, CP640, 808, route de Lennik, 1070 Bruxelles, [email protected]
Le syndrome d’Angelman : défaut d’expression d’un gène soumis à empreinteLe syndrome d’Angelman [1] est une affection neurogénétique caractérisée par une déficience intellectuelle sévère, un comportement exubérant et jovial (Figure 1), l’absence de langage, des troubles moteurs, de l’épilepsie et un électroencéphalogramme typique. Il est relativement rare (prévalence estimée à environ un individu sur 12 000), mais les implications épigénétiques en cause dans cette pathologie pourraient éga-lement concerner de nombreuses autres affections. Il est dû au déficit d’expres-sion du gène UBE3A [2, 3] dans les cellules cérébrales. Ce gène code pour l’ubiquitine-protéine ligase E3 qui agit sur l’étape d’ubiquitinylation du système protéolytique ubiquitine-protéasome et, mais les liens physiologiques entre le déficit moléculaire et les manifestations cliniques restent à clarifier. L’absence d’expression de l’UBE3A pourrait entra-ver des processus neuronaux en rapport avec la dégradation et le remplacement
de protéines spécifiques, interférer avec la transduction des signaux, le cycle cellulaire, la réparation de l’ADN et/ou la régulation de la transcription. Dans le cerveau, le déficit d’expression du gène pourrait entraîner des anoma-lies de la formation des épines dendri-tique et de la plasticité synaptique [4], ainsi que de la transmission synaptique (acide g-aminobutyrique [GABAA] et acide N-méthyl-D-aspartique [NMDA]) et non synaptique (gap junction) dans plusieurs régions, notamment au niveau de l’hippocampe [5] et du cervelet [6].Dans les neurones, le gène UBE3A est soumis à empreinte : il n’est norma-lement exprimé qu’à partir de l’allèle maternel sur le chromosome 15, tandis que l’allèle d’origine paternelle reste virtuellement silencieux. Il semble en effet qu’un centre de contrôle situé à côté du gène UBE3A sur le chromosome 15 paternel force la transcription de cette portion de l’ADN à l’envers, et que le transcrit antisens qui en résulte inhibe l’expression du gène UBE3A [7], agissant comme un signal « Ne me lisez
pas ! ». Sur le chromosome maternel, ce centre de contrôle est méthylé, ce qui inhibe la transcription antisens de l’ADN, comme si elle occultait le signal « Ne me lisez pas ! », et rend ainsi pos-sible l’expression de l’UBE3A à partir de l’allèle maternel.Plusieurs mécanismes moléculaires peuvent empêcher l’expression normale d’UBE3A à partir du chromosome mater-nel et ainsi causer le syndrome d’Angel-man (Figure 2). Il peut s’agir d’une délé-tion ou d’une mutation du gène UBE3A d’origine maternelle. Plus rarement, c’est l’empreinte marquant le chromo-some comme hérité de la mère, c’est-à-dire la méthylation de l’antisens, qui est absente. Enfin, il arrive que les deux chromosomes 15 soient hérités du père et aucun de la mère : on parle alors de disomie uniparentale paternelle. Tous ces mécanismes empêchent l’expression du gène UBE3A, soit par anomalie du gène lui-même (délétion ou mutation), soit parce qu’il ne peut pas être transcrit (absence de méthylation de l’antisens). Tous ces mécanismes provoquent le

m/s n° 6-7, vol. 28, juin-juillet 2012
NOUV
ELLE
SM
AGAZ
INE
9
En clinique oncologique, le mécanisme thérapeutique du Topo repose sur une inhibition de la réplication de l’ADN conduisant à la mort des cellules can-céreuses ce qui confère à cette molécule un effet cytotoxique puissant. L’équipe de B. Philpot et M. Zylka a démontré, chez la souris, qu’il favorise l’expres-sion de l’allèle paternel en réduisant la transcription de l’antisens par un mécanisme distinct de la méthylation du centre de contrôle et qui reste à découvrir. L’identification du mécanisme d’action du Topo via cette activation de l’expression de l’allèle paternel du gène UBE3A pourrait permettre le dévelop-pement de nouvelles approches phar-macologiques pour traiter les patients atteints du syndrome d’Angelman.De fait, de nombreuses interrogations persistent avant que l’on puisse affirmer que ces résultats obtenus chez la sou-ris peuvent s’appliquer à des humains. Par exemple, les effets de cette molé-cule sur l’expression du gène Ube3a sont-ils stables à long terme chez la souris ? Quels seraient les effets du Topo sur l’expression clinique du syn-drome d’Angelman chez l’animal ? Quelles seraient les doses nécessaires ? Y aurait-il des effets secondaires ? Le traitement serait-il sûr et efficace chez des humains ? À quel type de patient l’administrer ? Ce n’est que lorsqu’une démarche scientifique aura permis de clarifier ces aspects que l’on pourra envisager dans quelle mesure des molé-cules comme le Topo seraient à même de soulager les symptômes des individus atteints du syndrome d’Angelman.Ce sont de nouvelles questions qui sou-lèvent l’enthousiasme des équipes qui
intact mais non fonctionnel, notamment par des approches diététiques visant à modifier la méthylation de l’ADN afin d’entraver le signal « Ne me lisez pas ! » [8]. Ces tentatives sont restées infruc-tueuses. Tout récemment, une équipe américaine dirigée par B. Philpot et M. Zylka a testé plus de 2 000 médicaments sur un modèle murin afin d’évaluer si certains d’entre eux pouvaient activer l’allèle paternel du gène [9]. Parmi ces molécules, les inhibiteurs de la topoi-somérase I1, qui appartiennent à une famille de médicaments anticancéreux interagissant avec l’ADN, se sont révé-lés capables d’activer l’allèle paternel silencieux. La molécule la plus promet-teuse est le topotécan ou Topo. C’est un dérivé semi-synthétique de la camp-tothécine, un alcaloïde naturellement présent dans Camptotheca acuminata, un arbre ornemental.
1 La topoisomérase I (protéine nucléaire monomérique de 91 kDa) coupe un seul brin du duplex d’ADN en formant un complexe entre l’ADN et un résidu de tyrosine de l’enzyme. La coupure d’un seul brin diminue la tension de l’hélice et assurer sa relaxation. Les inhibiteurs stabilisent ce complexe covalent, et empêchent ainsi la religation des brins d’ADN, ce qui entraîne l’action cytotoxique de la molécule.
syndrome d’Angelman, mais la gravité des symptômes varie (statistiquement) en fonction du mécanisme sous-jacent. La déficience intellectuelle, les troubles du langage et l’épilepsie tendent à être moins marqués chez les individus pré-sentant un défaut d’empreinte ou une disomie uniparentale, que chez ceux chez lesquels le gène UBE3A est absent ou muté.Cette variabilité suggère la persistance d’une expression résiduelle - quoique très faible - de l’allèle intact du gène malgré la présence du signal « Ne me lisez pas ! ». Les patients ayant une délétion ou une mutation possèdent un seul allèle intact, mais virtuelle-ment non opérationnel, du gène UBE3A (celui qui est hérité du père) ; ceux qui ont une disomie uniparentale ou un trouble de l’empreinte possèdent deux allèles (deux d’origine paternelle ou un paternel et un maternel).
Le topotécan : levée de l’inhibition de l’allèle UBE3A paternelDepuis 2000, plusieurs équipes ont tenté de provoquer l’expression de l’allèle
Figure 1. Aspects caractéristiques du syn-drome d’Angelman. On peut observer plusieurs aspects très caractéristiques du syndrome d’Angelman : le contact visuel, l’hypoplasie de l’étage moyen de la face, la bouche large, souriante, la position basse de la langue, la sialorrhée et la posture reflétant un tonus musculaire bas.

m/s n° 6-7, vol. 28, juin-juillet 2012 10
4. Greer PL, Hanayama R, Bloodgood BL, et al. The Angelman syndrome protein Ube3A regulates synapse development by ubiquitinating arc. Cell 2010 ; 140 : 704-16.
5. Weeber EJ, Jiang YH, Elgersma Y, et al. Derangements of hippocampal calcium/calmodulin-dependent protein kinase II in a mouse model for Angelman mental retardation syndrome. J Neurosci 2003 ; 23 : 2634-44.
6. Cheron G, Servais L, Wagstaff J, Dan B. Fast cerebellar oscillation associated with ataxia in a mouse model of Angelman syndrome. Neuroscience 2005 ; 130 : 631-7.
7. Rougeulle C, Cardoso C, Fontes M, et al. An imprinted antisense RNA overlaps UBE3A and a second maternally expressed transcript. Nat Genet 1998 ; 19 : 15-6.
8. Bird LM, Tan WH, Bacino CA, et al. A therapeutic trial of pro-methylation dietary supplements in Angelman syndrome. Am J Med Genet 2011 ; 155A : 2956-63.
9. Huang HS, Allen JA, Mabb AM, et al. Topoisomerase inhibitors unsilence the dormant allele of Ube3a in neurons. Nature 2011 ; 481 : 185-9.
y travaillent tout en n’oubliant pas que l’éthique de la science médicale impose de suivre des chemins balisés et pro-gressifs qui ne mènent aux essais cli-niques qu’après obtention des réponses à toutes les questions préliminaires. ◊A new topo to targeted management of Angelman syndrome?
LIENS D’INTÉRÊTSLes auteurs déclarent n’avoir aucun lien d’intérêt concernant les données publiées dans cet article.
RÉFÉRENCES
1. Dan B. Angelman syndrome. London : Mac Keith Press, Wiley-Blackwell, 2008 : 256 p.
2. Kishino T, Lalande M, Wagstaff J. UBE3A/E6-AP mutations cause Angelman syndrome. Nat Genet 1997 ; 15 : 70-3.
3. Matsuura T, Sutcliffe JS, Fang P, et al. De novo truncating mutations in E6-AP ubiquitin-protein ligase gene (UBE3A) in Angelman syndrome. Nat Genet 1997 ; 15 : 74-7.
Figure 2. Schéma explicatif des différentes classes moléculaires du syndrome d’Angelman. Le chromosome 15 hérité de la mère est représenté par une femme, celui hérité du père par un homme. Le panneau de circulation à sens unique indique l’expression de l’allèle du gène UBE3A, le panneau d’impasse l’inactivation de l’allèle, le panneau de chantier une délétion ou une mutation du gène et le panneau du virage la réactivation de l’allèle inactif. (1) Dans la situation physiologique, seul le chromosome 15 hérité de la mère porte un signal épigénétique permettant l’expression du gène UBE3A, alors que l’allèle paternel de ce gène est inactif. (2) En cas de délé-tion ou de mutation de l’allèle maternel du gène UBE3A, celui-ci ne peut pas être exprimé et l’al-lèle paternel est inactif comme dans la situation physiologique. (3) Si l’empreinte génomique fait défaut sur le chromosome 15 hérité de la mère, le gène UBE3A n’est pas exprimé, puisque l’allèle maternel ainsi que l’allèle paternel sont inactifs. (4) Si les deux chromosomes 15 sont hérités du père, les deux allèles sont inactifs. (5) Une voie thérapeutique serait de favoriser l’expression de l’allèle paternel (© Christyan Fox).

m/s n° 6-7, vol. 28, juin-juillet 2012
NOUV
ELLE
SM
AGAZ
INE
11
Institut de biologie de l’École normale supérieure (ENS), CNRS-Inserm-ENS, 46, rue d’Ulm, 75230 Paris Cedex 05, [email protected]
> Chez de nombreux organismes, l’intro-duction artificielle d’un fragment d’ARN double brin (ARNdb) correspondant à la séquence d’un gène inactive spé-cifiquement l’expression de ce gène. Ce phénomène d’interférence à l’ARN (ARNi) a été mis en évidence chez le nématode C. elegans [1], un petit ver d’un millimètre qui est devenu un orga-nisme modèle de laboratoire. L’ARNi est devenu un outil pratique d’inactivation des gènes, et cette découverte a valu le prix Nobel de médecine à Andy Fire et Craig Mello en 2006. Des résultats récents montrent que la réponse ARNi peut être transmise de génération en génération en affectant l’état des his-tones et la régulation transcriptionnelle du gène. Le rôle physiologique de cette transmission héréditaire pourrait-il être la défense antivirale ?
Inactivation post-transcriptionnelle… mais aussi transcriptionnelleLe mécanisme de l’ARNi a été amplement étudié depuis 15 ans [2]. Chez C. elegans, l’injection ou l’ingestion d’un ARNdb peut déclencher la réponse d’interfé-rence. La ribonucléase Dicer/DCR-1 clive dans le cytoplasme l’ARNdb en fragments de 21 nucléotides, nommés siARN (si pour small interfering) (Figure 1). Ces siARN-1 recrutent et guident la pro-téine Argonaute RDE-1 vers une autre copie de l’ARN messager complémentaire (ARNm) qui sert de matrice à la syn-thèse de siARN-2 antisens par une ARN polymérase dépendante de l’ARN. Ces
siARN-2 amplifiés lient alors d’autres protéines Argonautes, et ces complexes induisent la dégradation des ARNm cor-respondants. Ceci conduit à l’extinction post-transcriptionnelle (PTGS pour post-transcriptional gene silencing) du gène.Un second mécanisme d’action des siARN 2° a été mis en évidence chez C. elegans par le laboratoire de Scott Kennedy. Les siARN-2 se lient à la protéine Argonaute NRDE-3, et le complexe entre dans le noyau. L’extinction du gène correspon-dant se fait alors au niveau de la trans-cription (TGS pour transcriptional gene silencing) (Figure 1). Les gènes impliqués dans ce phénomène ont été identifiés par un crible génétique et appelés nrde (nuclear-RNAi deficient).
Extinction transgénérationnelle de certains gènes à partir d’un ARNdb spécifiqueChez C. elegans, l’ARNi peut, dans cer-tains cas, persister pendant plusieurs générations, mimant les effets trans-générationnels de l’extinction de gènes connus chez certains champignons et plantes. Ainsi, chez C. elegans, l’extinc-tion d’un transgène exprimant la pro-téine fluorescente GFP (green fluores-cent protein) après l’ingestion d’ARNdb spécifique de la GFP peut être transmise jusqu’à 20 générations [3]. Cette trans-mission fonctionne pour certains gènes seulement (13/171 gènes testés sont ainsi éteints sur plusieurs générations) [3-5], sans que l’on comprenne pour l’instant ce qui les caractérise.
Une diminution de la quantité d’ARNdb injectée réduit la durée de l’extinction du gène, suggèrant que la transmission de l’effet ARNi est diluée au fil des géné-rations [5]. La même étude du labora-toire d’Andrew Fire révèle que les deux types de gamètes, mâle et femelle, sont capables de transmettre ces facteurs, mais que les spermatozoïdes le font plus efficacement.
Deux mécanismes de transmission transgénérationnelleLa transmission de l’extinction ARNi ne requiert pas que la descendance puisse elle même produire des siRNA-1. En effet, cette transmission se fait norma-lement chez un descendant incapable de produire des siRNA-1 (mutant rde-1 ou rde-4) [4]. En revanche, les gènes nécessaires à l’amplification des siARN 2°, comme mut-7 ou rde-2, doivent, eux, être fonctionnels dans la descendance [4], comme doivent l’être aussi les gènes nrde-2, nrde-3 et nrde-4 nécessaires à la voie de régulation transcription-nelle en aval des siARN-2 [6]. La trans-mission transgénérationnelle requiert donc, à chaque génération, la forma-tion de siARN-2 et, en aval, une voie de répression transcriptionnelle intacte (Figure 1).La répression transcriptionnelle par les siARN-2 semble passer par des modifi-cations d’histones. Les histones sont des protéines de la chromatine qui régulent le degré de condensation et l’accessibilité de cette dernière pour que s’enclenche la
Transmission multigénérationnelle de l’interférence à l’ARN chez le nématode Caenorhabditis elegansTony Bélicard, Marie-Anne Félix
NOUVELLE

m/s n° 6-7, vol. 28, juin-juillet 2012 12
Gu et al. [7], du laboratoire d’Andrew Fire, ont récemment réussi à suivre la quantité de siARN-2 spécifiques et le taux de méthylation de H3K9 au locus correspondant chez un ver adulte ayant absorbé des ARNdb spécifiques et dans sa descendance pendant quatre géné-rations. Dans la génération parentale (P0), la synthèse des siARN spécifiques est d’abord détectée, puis une marque H3K9me au niveau du gène ciblé est observée. Les siARN-2 et la méthylation sont ensuite détectés chez les descen-dants et ce jusqu’à trois générations dans cette étude, mais avec un mar-quage d’intensité décroissante au fil des générations [7].Quel peut être le facteur transmis par les gamètes qui permet l’héritabilité de l’extinction transcriptionnelle ? Les modifications d’histones sont trans-missibles lors des divisions cellulaires, mais elles sont généralement « réini-tialisées » dans la lignée germinale chez C. elegans. De plus, l’extinction transcriptionnelle peut aussi s’appliquer à une copie du gène nouvellement intro-duite par croisement, et l’action des siARN-2 sur la méthylation des histones semble devoir être répétée à chaque génération. La transmission de siARN via les gamètes pourrait permettre une persistance et une réamplification de l’extinction transcriptionnelle de cer-tains gènes au fil des générations. Pour l’instant, il n’est pas clair s’il s’agit de siARN-1 ou 2 (voir plus loin).
L’enjeu évolutif de la transmission de l’ARNi serait-il l’immunisation de la descendance contre des pathogènes naturels ?En principe, la transmission transgéné-rationnelle du statut de l’expression des gènes permet à un parent de transmettre à sa descendance les conséquences qu’ont imprimées à son organisme les influences de l’environnement (effet « lamarckien »). Ceci peut concerner par exemple la défense antivirale. En effet, chez C. elegans comme chez les plantes et les insectes, la voie d’ARNi
la transcription. De plus, la transmission de l’extinction d’expression génique est abolie chez des mutants affectant la structure chromatinienne [3].
transcription. En effet, la voie nucléaire des siARN active la méthylation de la lysine 9 de l’histone 3 (H3K9me) [6], une modification qui inhibe localement
ARNdb
RDE-1
AAA
Dégradation ARNm(PTGS)
siARN 1°
siARN 2°
RdRp
Noyau
NRDE-3
Parent P0
P0 Gamètes F
1
F3 Gamètes F
4
Cytoplasme
DCR-1
siARN ?
H3K9me au locus
Répression transcriptionnelledu gène (TGS)
H3K9me au locus
H3K9me au locus
TGS
PTGS ?
PTGS ?
TGS
siARN ?
A
B
Figure 1. Mécanismes transcriptionnels et post-transcriptionnels de l’extinction de gènes par l’ARN double-brin et transmission multigénérationnelle. A. Dans le parent P0 exposé à des ARNdb, deux mécanismes de répression de l’expression du gène correspondant peuvent être déclenchés. L’ARNdb (en vert) est fragmenté par l’enzyme DCR-1. Les petits siARN-1 produits (extrémité 5’ monophosphate) sont pris en charge par l’Argonaute RDE-1. Ce complexe s’hybride à son ARNm complémentaire et produit des siARN antisens dits 2 (extrémité 5’ triphosphate). Ces derniers vont, soit participer directement au clivage des ARNm correspondants (voie PTGS ou post-transcriptional gene silencing, en orange), soit être pris en charge par l’Argonaute NRDE-3. Le complexe siARN 2°/NRDE-3 entre dans le noyau et réprime la transcription du gène correspondant (voie TGS ou transcriptional gene silencing, en bleu). B. Transmission de l’extinction du gène dans les générations suivantes (F1, etc.). Les siARN-1 semblent être transmis dans les gamètes et, même en absence d’ARNdb exogène dans la descendance, permettent l’extinction des gènes ciblés via la méthylation de l’ADN. La quantité de siARN transmis décroît au fil des générations.

m/s n° 6-7, vol. 28, juin-juillet 2012
NOUV
ELLE
SM
AGAZ
INE
13
utiliser l’amplification de petits ARN spécifiques et la répression transcrip-tionnelle à chaque génération. L’impact de ce processus de transmission sur l’adaptation phénotypique des orga-nismes en milieu naturel reste cepen-dant à démontrer. Il est possible que la cible principale de cette extinction mul-tigénérationnelle soit les transposons insérés dans le génome. ◊????????? ???????
LIENS D’INTÉRÊTSLes auteurs déclarent n’avoir aucun lien d’intérêt concernant les données publiées dans cet article.
REMERCIEMENTSL’équipe est financée par le CNRS, l’ENS, l’Inserm, et pour la recherche sur les virus, la subvention de l’ANR11 BSV3 01301. Nous remercions Valérie Robert (ENS Lyon) pour ses commentaires.
RÉFÉRENCES
1. Fire A, Xu S, Montgomery MK, et al. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998 ; 391 : 806-11.
2. Fischer SEJ. Small RNA-mediated gene silencing pathways in C. elegans. Int J Biochem Cell Biol 2010 ; 42 : 1306-15.
3. Vastenhouw NL, Brunschwig K, Okihara KL, et al. Gene expression: long-term gene silencing by RNAi. Nature 2006 ; 442 : 882.
4. Grishok A, Tabara H, Mello CC. Genetic requirements for inheritance of RNAi in C. elegans. Science 2000 ; 287 : 2494-7.
5. Alcazar RM, Lin R, Fire AZ. Transmission dynamics of heritable silencing induced by double-stranded RNA in Caenorhabditis elegans. Genetics 2008 ; 180 : 1275-88.
6. Burton NO, Burkhart KB, Kennedy S. Nuclear RNAi maintains heritable gene silencing in Caenorhabditis elegans. Proc Natl Acad Sci USA 2011 ; 108 : 19683-8.
7. Gu SG, Pak J, Guang S, et al. Amplification of siRNA in Caenorhabditis elegans generates a transgenerational sequence-targeted histone H3 lysine 9 methylation footprint. Nat Genet 2012 ; 44 : 157-64.
8. Félix M-A, Ashe A, Piffaretti J, et al. Natural and experimental infection of Caenorhabditis nematodes by novel viruses related to nodaviruses. PLoS Biol 2011 ; 9 : e1000586.
9. Lu R, Maduro M, Li F, et al. Animal virus replication and RNAi-mediated antiviral silencing in Caenorhabditis elegans. Nature 2005 ; 436 : 1040-3.
10. Rechavi O, Minevich G, Hobert O. Transgenerational inheritance of an acquired small RNA-based antiviral response in C. elegans. Cell 2011 ; 147 : 1248-56.
peuvent être reconstituées et suivies en mesurant l’expression ou l’extinc-tion de la GFP [9]. Rechavi et al. [10] ont récemment étudié la transmissibi-lité intergénérationnelle de petits ARN antiviraux dérivés de l’expression d’un tel transgène. Comme c’est le cas pour un gène endogène (voir plus haut) ou la GFP, l’extinction des ARN viraux peut se transmettre en absence de la machine-rie de formation des siARN 1° chez les descendants (mutant rde-1), et peut se transmettre à une nouvelle copie du gène testé introduite par croisement. Cependant, cette réponse requiert à chaque génération l’amplification de siARN-2 - elle est abolie dans le mutant rrf-1 affectant une RdRP du ver. Des siARN-1 correspondant aux séquences virales peuvent être détectés sur plu-sieurs générations, quoique en très petit nombre dès la troisième génération. Ceci suggère donc que la transmission peut s’effectuer via les siARN-1, qui pourraient enclencher l’amplification de novo de siARN-2 à chaque génération.Pour ce qui est de la portée de cette étude sur la transmission d’une défense antivirale, on peut regretter que les séquences virales y soient portées sur un transgène artificiel. En effet, l’inter-férence ne peut s’appliquer ici que au niveau transcriptionnel. Ce ne peut pas être le cas avec un virus relativement proche, tel que celui d’Orsay, dont le génome n’existe pas sous une forme ADN. Aucun rétrovirus n’est pour l’ins-tant connu chez les nématodes, mais des rétrotransposons y sont insérés dans le génome.
En conclusionL’extinction d’un gène par l’ARNdb cor-respondant peut se transmettre au fil des générations chez des animaux, comme chez les plantes et les cham-pignons. Chez C. elegans, elle semble
joue un rôle dans la défense antivirale - de nombreux virus ayant un génome de type ARNdb.Le virus d’Orsay - découvert récem-ment - est le premier virus naturel de C. elegans, [8]. Son génome s’apparente à celui des nodavirus : il est composé de deux segments d’ARN simple brin positif (ARN1 et ARN2). L’ARN1 code pour une ARN polymérase dépendante de l’ARN (RdRP) qui synthétise le brin complémentaire des ARN viraux et, par conséquent, crée des ARNdb. Ceux-ci déclenchent alors une réponse siARN contre le génome viral. Les mutants C. elegans des gènes impliqués dans la réponse siARN sont plus sensibles au virus que le génotype sauvage de réfé-rence [8]. La réponse siARN est donc requise pour la défense antivirale chez C. elegans.La réponse à un virus peut-elle se trans-mettre d’une génération à l’autre, et ainsi « immuniser » plusieurs généra-tions ? Chez C. elegans, nous n’avons pas encore la réponse avec un virus infectieux comme le virus d’Orsay, mais un système artificiel a testé cette hypothèse.Un système transgénique artifi-ciel reconstituant la réplication du Flock house virus (FHV), un nodavirus d’arthropode qui n’infecte pas naturel-lement C. elegans, a été mis au point il y a quelques années [9]. Le génome du virus FHV est composé de deux ARN dont l’un code pour une RdRP et pour la protéine B2 qui inhibe la réponse ARNi de l’hôte. En remplaçant cette protéine B2 par la GFP et en construisant un ver transgénique dans lequel les séquences ADN correspondant à RdRP et B2 sont sous le contrôle d’un promoteur induc-tible par un choc thermique, la répli-cation de l’ARN1 viral dans le ver (qui est assurée ensuite de façon autonome par la RdRP) et la réponse d’extinction
m/s n° 6-7, vol. 28, juin-juillet 2012 14
NOUVELLE
Positionnement du noyau dans les musclesNouvel acteur de la fonction musculaireVincent Gache1, Bruno Cadot1, Edgar R. Gomes1,2
1UMR S 787 Inserm, université Paris 6 Pierre et Marie Curie, 75634 Paris, France ;2Groupe hospitalier Pitié-Salpêtrière, institut de myologie, 105, boulevard de l’Hôpital, 75634 Paris Cedex 13, [email protected]. Gache et B. Cadot ont contribué équitablement à ce travail.
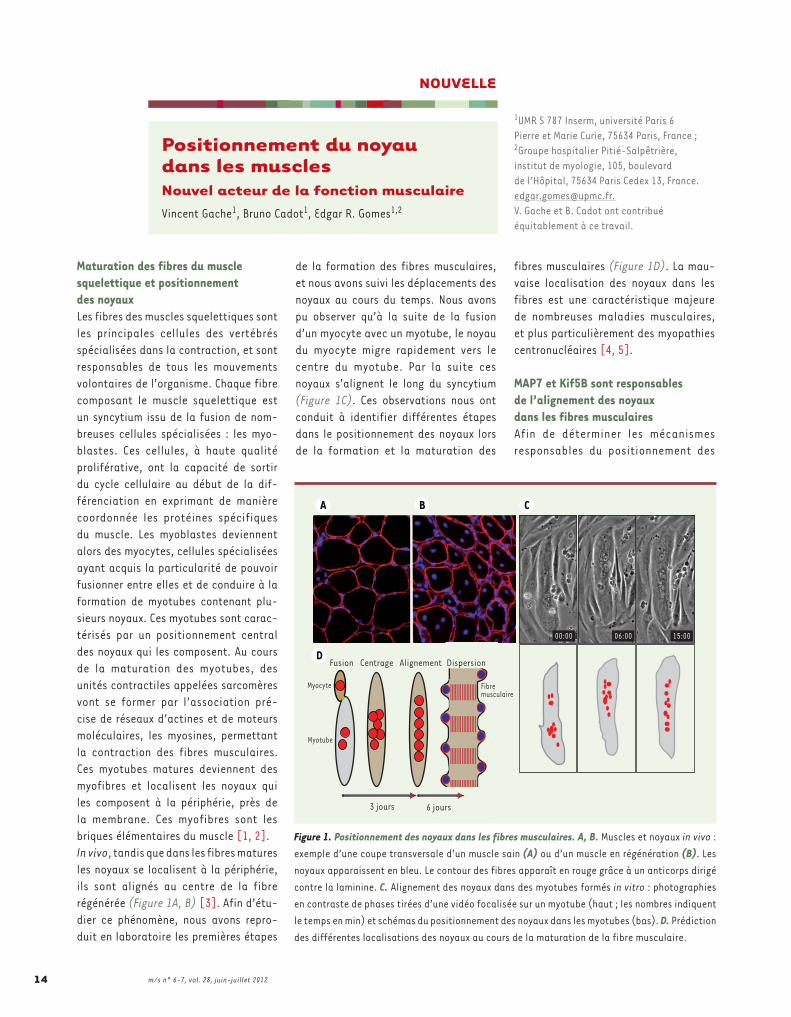
Maturation des fibres du muscle squelettique et positionnement des noyauxLes fibres des muscles squelettiques sont les principales cellules des vertébrés spécialisées dans la contraction, et sont responsables de tous les mouvements volontaires de l’organisme. Chaque fibre composant le muscle squelettique est un syncytium issu de la fusion de nom-breuses cellules spécialisées : les myo-blastes. Ces cellules, à haute qualité proliférative, ont la capacité de sortir du cycle cellulaire au début de la dif-férenciation en exprimant de manière coordonnée les protéines spécifiques du muscle. Les myoblastes deviennent alors des myocytes, cellules spécialisées ayant acquis la particularité de pouvoir fusionner entre elles et de conduire à la formation de myotubes contenant plu-sieurs noyaux. Ces myotubes sont carac-térisés par un positionnement central des noyaux qui les composent. Au cours de la maturation des myotubes, des unités contractiles appelées sarcomères vont se former par l’association pré-cise de réseaux d’actines et de moteurs moléculaires, les myosines, permettant la contraction des fibres musculaires. Ces myotubes matures deviennent des myofibres et localisent les noyaux qui les composent à la périphérie, près de la membrane. Ces myofibres sont les briques élémentaires du muscle [1, 2].In vivo, tandis que dans les fibres matures les noyaux se localisent à la périphérie, ils sont alignés au centre de la fibre régénérée (Figure 1A, B) [3]. Afin d’étu-dier ce phénomène, nous avons repro-duit en laboratoire les premières étapes
de la formation des fibres musculaires, et nous avons suivi les déplacements des noyaux au cours du temps. Nous avons pu observer qu’à la suite de la fusion d’un myocyte avec un myotube, le noyau du myocyte migre rapidement vers le centre du myotube. Par la suite ces noyaux s’alignent le long du syncytium (Figure 1C). Ces observations nous ont conduit à identifier différentes étapes dans le positionnement des noyaux lors de la formation et la maturation des
fibres musculaires (Figure 1D). La mau-vaise localisation des noyaux dans les fibres est une caractéristique majeure de nombreuses maladies musculaires, et plus particulièrement des myopathies centronucléaires [4, 5].
MAP7 et Kif5B sont responsables de l’alignement des noyaux dans les fibres musculairesAfin de déterminer les mécanismes responsables du positionnement des
A
D
B C
Fusion Centrage Alignement Dispersion
Myocyte
Myotube
3 jours 6 jours
Fibre musculaire
00:00 06:00 15:00
Figure 1. Positionnement des noyaux dans les fibres musculaires. A, B. Muscles et noyaux in vivo : exemple d’une coupe transversale d’un muscle sain (A) ou d’un muscle en régénération (B). Les noyaux apparaissent en bleu. Le contour des fibres apparaît en rouge grâce à un anticorps dirigé contre la laminine. C. Alignement des noyaux dans des myotubes formés in vitro : photographies en contraste de phases tirées d’une vidéo focalisée sur un myotube (haut ; les nombres indiquent le temps en min) et schémas du positionnement des noyaux dans les myotubes (bas). D. Prédiction des différentes localisations des noyaux au cours de la maturation de la fibre musculaire.
m/s n° 6-7, vol. 28, juin-juillet 2012
NOUV
ELLE
SM
AGAZ
INE
15
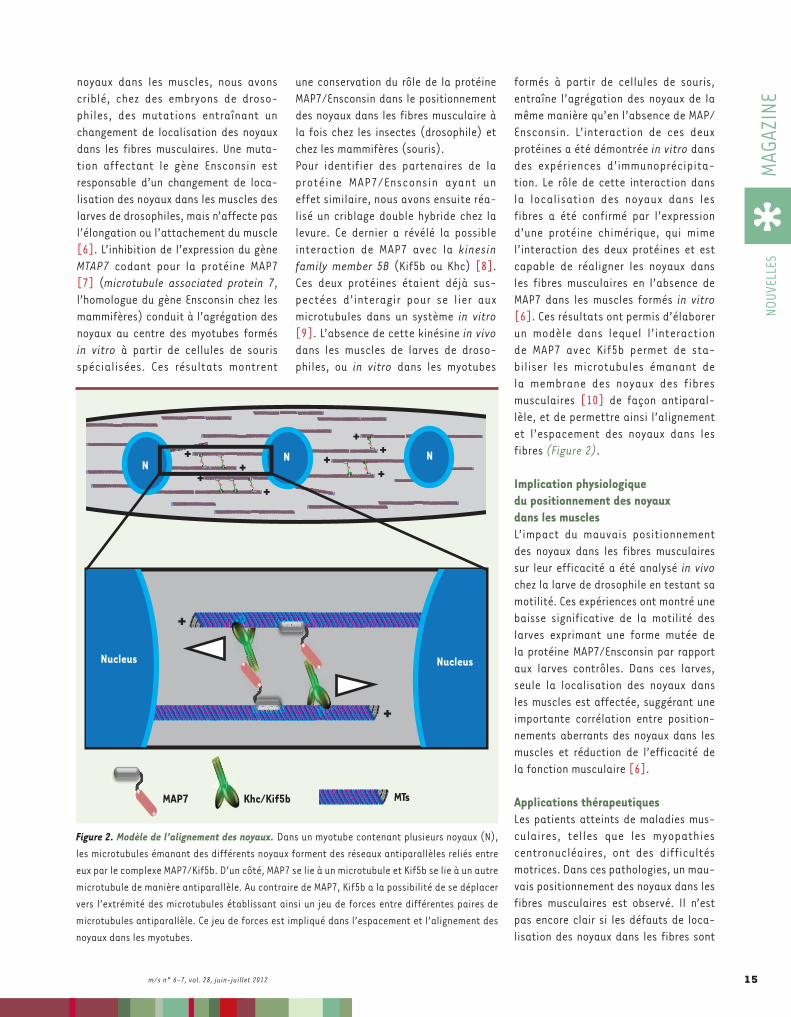
formés à partir de cellules de souris, entraîne l’agrégation des noyaux de la même manière qu’en l’absence de MAP/Ensconsin. L’interaction de ces deux protéines a été démontrée in vitro dans des expériences d’immunoprécipita-tion. Le rôle de cette interaction dans la localisation des noyaux dans les fibres a été confirmé par l’expression d’une protéine chimérique, qui mime l’interaction des deux protéines et est capable de réaligner les noyaux dans les fibres musculaires en l’absence de MAP7 dans les muscles formés in vitro [6]. Ces résultats ont permis d’élaborer un modèle dans lequel l’interaction de MAP7 avec Kif5b permet de sta-biliser les microtubules émanant de la membrane des noyaux des fibres musculaires [10] de façon antiparal-lèle, et de permettre ainsi l’alignement et l’espacement des noyaux dans les fibres (Figure 2).
Implication physiologique du positionnement des noyaux dans les musclesL’impact du mauvais positionnement des noyaux dans les fibres musculaires sur leur efficacité a été analysé in vivo chez la larve de drosophile en testant sa motilité. Ces expériences ont montré une baisse significative de la motilité des larves exprimant une forme mutée de la protéine MAP7/Ensconsin par rapport aux larves contrôles. Dans ces larves, seule la localisation des noyaux dans les muscles est affectée, suggérant une importante corrélation entre position-nements aberrants des noyaux dans les muscles et réduction de l’efficacité de la fonction musculaire [6].
Applications thérapeutiquesLes patients atteints de maladies mus-culaires, telles que les myopathies centronucléaires, ont des difficultés motrices. Dans ces pathologies, un mau-vais positionnement des noyaux dans les fibres musculaires est observé. Il n’est pas encore clair si les défauts de loca-lisation des noyaux dans les fibres sont
une conservation du rôle de la protéine MAP7/Ensconsin dans le positionnement des noyaux dans les fibres musculaire à la fois chez les insectes (drosophile) et chez les mammifères (souris).Pour identifier des partenaires de la protéine MAP7/Ensconsin ayant un effet similaire, nous avons ensuite réa-lisé un criblage double hybride chez la levure. Ce dernier a révélé la possible interaction de MAP7 avec la kinesin family member 5B (Kif5b ou Khc) [8]. Ces deux protéines étaient déjà sus-pectées d’inter agir pour se lier aux microtubules dans un système in vitro [9]. L’absence de cette kinésine in vivo dans les muscles de larves de droso-philes, ou in vitro dans les myotubes
noyaux dans les muscles, nous avons criblé, chez des embryons de droso-philes, des mutations entraînant un changement de localisation des noyaux dans les fibres musculaires. Une muta-tion affectant le gène Ensconsin est responsable d’un changement de loca-lisation des noyaux dans les muscles des larves de drosophiles, mais n’affecte pas l’élongation ou l’attachement du muscle [6]. L’inhibition de l’expression du gène MTAP7 codant pour la protéine MAP7 [7] (microtubule associated protein 7, l’homologue du gène Ensconsin chez les mammifères) conduit à l’agrégation des noyaux au centre des myotubes formés in vitro à partir de cellules de souris spécialisées. Ces résultats montrent
MAP7 MTsKhc/Kif5b
+
++++
++
+
+
+
Nucleus Nucleus
NN N
Figure 2. Modèle de l’alignement des noyaux. Dans un myotube contenant plusieurs noyaux (N), les microtubules émanant des différents noyaux forment des réseaux antiparallèles reliés entre eux par le complexe MAP7/Kif5b. D’un côté, MAP7 se lie à un microtubule et Kif5b se lie à un autre microtubule de manière antiparallèle. Au contraire de MAP7, Kif5b a la possibilité de se déplacer vers l’extrémité des microtubules établissant ainsi un jeu de forces entre différentes paires de microtubules antiparallèle. Ce jeu de forces est impliqué dans l’espacement et l’alignement des noyaux dans les myotubes.

m/s n° 6-7, vol. 28, juin-juillet 2012 16
5. Jungbluth H, Wallgren-Pettersson C, Laporte J. Centronuclear (myotubular) myopathy. Orphanet J Rare Dis 2008 ; 3 : 26.
6. Metzger T, Gache V, Xu M, et al. MAP and kinesin-dependent nuclear positioning is required for skeletal muscle function. Nature 2012 ; 484 : 120-4.
7. Masson D, Kreis TE. Identification and molecular characterization of E-MAP-115, a novel microtubule-associated protein predominantly expressed in epithelial cells. J Cell Biol 1993 ; 123 : 357-71.
8. Vale RD, Reese TS, Sheetz MP. Identification of a novel force-generating protein, kinesin, involved in microtubule-based motility. Cell 1985 ; 42 : 39-50.
9. Sung HH, Telley IA, Papadaki P, et al. Drosophila ensconsin promotes productive recruitment of Kinesin-1 to microtubules. Dev Cell 2008 ; 15 : 866-76.
10. Tassin AM, Maro B, Bornens M. Fate of microtubule-organizing centers during myogenesis in vitro. J Cell Biol 1985 ; 100 : 35-46.
LIENS D’INTÉRÊTLes auteurs déclarent n’avoir aucun lien d’intérêt concernant les données publiées dans cet article.
RÉFÉRENCES 1. Carlson BM. Muscle regeneration in amphibians and
mammals: passing the torch. Dev Dyn 2003 ; 226 : 167-81.
2. Dhawan J, Rando TA. Stem cells in postnatal myogenesis: molecular mechanisms of satellite cell quiescence, activation and replenishment. Trends Cell Biol 2005 ; 15 : 666-73.
3. Bruusgaard JC, Liestøl K, Ekmark M, et al. Number and spatial distribution of nuclei in the muscle fibres of normal mice studied in vivo. J Physiol (Lond) 2003 ; 551 : 467-78.
4. Cohn RD, Campbell KP. Molecular basis of muscular dystrophies. Muscle Nerve 2000 ; 23 : 1456-71.
une cause de la faiblesse musculaire ou le résultat d’un dysfonctionnement des fibres. Toutefois, nos résultats montrent une corrélation entre le bon position-nement des noyaux dans les fibres et la fonction des muscles. Nous suggérons qu’une correction des défauts de posi-tionnement des noyaux musculaires chez des patients atteints de myopathies pourrait entraîner une amélioration de leur fonction musculaire. ◊Spatial distribution of nuclei in the muscle fibres: a novel actor of muscular fonction
UMR 698 Inserm, université Paris-Diderot, Sorbonne Paris-Cité, 75018, Paris, [email protected]
> En dépit d’importants investissements et des stratégies thérapeutiques dispo-nibles - prévention des facteurs de risque et traitements interventionnels -, les maladies athérothrombotiques restent une des premières causes de morbidité et mortalité dans les pays développés. Ce constat nous force à approfondir ce que sont réellement les lésions d’athé-rothrombose chez l’homme, à détermi-ner comment elles se développent et comment elles évoluent vers leur expres-sion clinique.
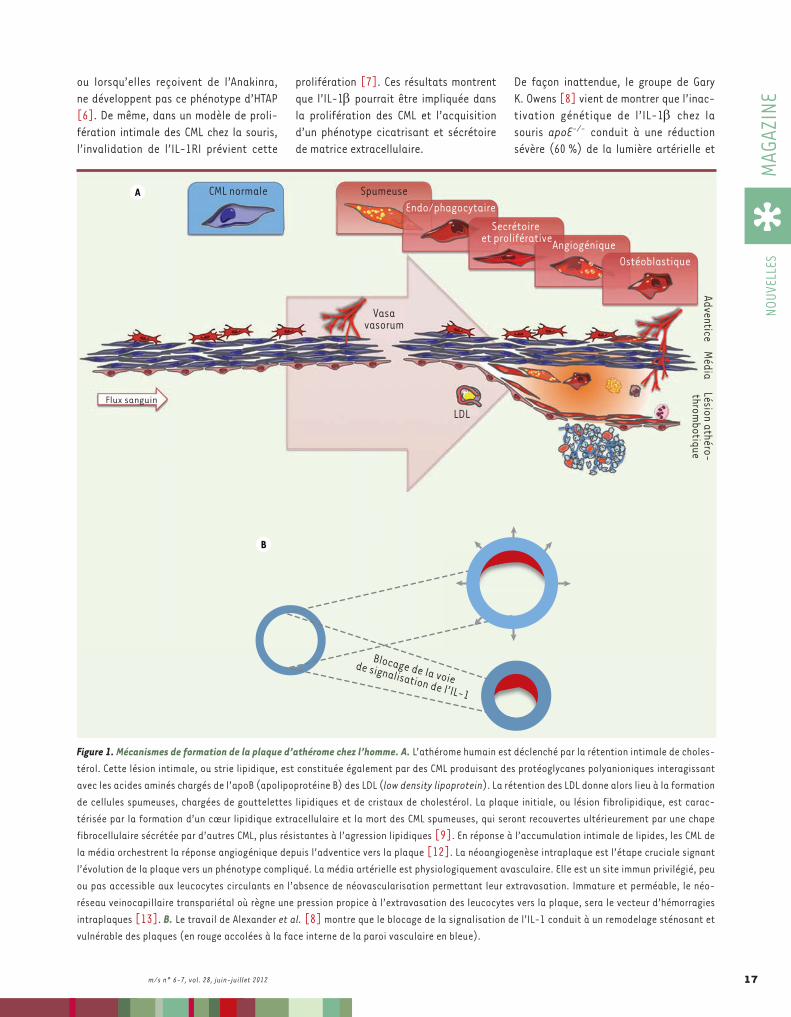
Interleukine-1 et lésions d’athérothromboseDepuis plus de dix ans, le paradigme dominant est que l’inflammation est le moteur de la progression des lésions d’athérothrombose [1], alors même que les traitements anti-inflammatoires classiques augmentent la fréquence de l’expression clinique de la maladie. Dans ce contexte du « tout inflammatoire », l’intérêt s’est récemment porté sur l’in-terleukine 1 bêta (IL-1b), une cytokine pro-inflammatoire produite par les
plaques à tous les stades d’athérome, en rapport avec la surcharge lipidique [2] et la cristallisation du cholestérol [3]. Contrairement à l’IL-1a dont elle par-tage les activités biologiques, l’IL-1b est une cytokine secrétée après son clivage par la caspase 1, cette protéase étant elle-même activée par l’« inflamma-some ». Si les sources principales d’IL-1b sont les leucocytes, les plaquettes et de très nombreuses cellules stromales sont aussi capables de la produire. Les récep-teurs de l’IL-1 sont également présents sur de nombreuses cellules de l’orga-nisme, et pas seulement sur les leuco-cytes. L’IL-1b et son récepteur principal, l’IL-1RI, représentent donc un système extrêmement ubiquitaire.
Essai thérapeutique de blocage de l’interleukine-1 dans la maladie athéromateuseUn essai clinique de blocage de l’IL-1b par un anticorps monoclonal humanisé, le Canakinumab (ILARIS®, étude CANTOS [Canakinumab anti-inflammatory throm-bosis outcomes study]), est en cours
dans l’athérothrombose [4], et ce malgré l’arrêt - pour une raison qui n’a pas été rendue publique - d’un essai du Canaki-numab dans la polyarthrite rhumatoïde chez l’homme (clinicaltrials.gov, Iden-tifier NCT00784628). Le Canakinumab a cependant été approuvé par la FDA (Food and drug administration) (États-Unis) et par l’EMA (European medicines agency) (Europe) dans l’indication des cryopyrino-pathies de l’enfant, une pathologie s’ac-compagnant de mutations dans le gène NLRP3 (NLR family, pyrin domain contai-ning 3) codant pour une protéine impli-quée dans l’activation de la caspase 1. L’Anakinra, un antagoniste synthétique des récepteurs de l’IL-1b, est peu effi-cace dans la polyarthrite rhumatoïde [5] et n’a pas encore été testé dans l’athé-rothrombose humaine. La souris apoE-/-, lorsqu’elle est soumise à un régime hyper-cholestérolémique, développe une hyper-tension artérielle pulmonaire (HTAP) caractérisée par une prolifération diffuse des cellules musculaires lisses (CML). Ces souris, lorsqu’elles sont croisées avec d’autres souris invalidées pour l’IL-1RI,
Faut-il bloquer l’interleukine-1 dans l’athérothrombose ?Antonino Nicoletti, Giuseppina Caligiuri, Benoit Ho-Tin-Noé, Jean-Baptiste Michel
NOUVELLE
m/s n° 6-7, vol. 28, juin-juillet 2012
NOUV
ELLE
SM
AGAZ
INE
17
De façon inattendue, le groupe de Gary K. Owens [8] vient de montrer que l’inac-tivation génétique de l’IL-1b chez la souris apoE-/- conduit à une réduction sévère (60 %) de la lumière artérielle et
prolifération [7]. Ces résultats montrent que l’IL-1b pourrait être impliquée dans la prolifération des CML et l’acquisition d’un phénotype cicatrisant et sécrétoire de matrice extracellulaire.
ou lorsqu’elles reçoivent de l’Anakinra, ne développent pas ce phénotype d’HTAP [6]. De même, dans un modèle de proli-fération intimale des CML chez la souris, l’invalidation de l’IL-1RI prévient cette
Flux sanguin
Vasavasorum
Blocage de la voie de signalisation de l’IL-1
LDL
CML normale SpumeuseEndo/phagocytaire
Secrétoire et proliférativeAngiogénique
Ostéoblastique
AdventiceM
édiaLésion athéro-throm
botique
A
B
Figure 1. Mécanismes de formation de la plaque d’athérome chez l’homme. A. L’athérome humain est déclenché par la rétention intimale de choles-térol. Cette lésion intimale, ou strie lipidique, est constituée également par des CML produisant des protéoglycanes polyanioniques interagissant avec les acides aminés chargés de l’apoB (apolipoprotéine B) des LDL (low density lipoprotein). La rétention des LDL donne alors lieu à la formation de cellules spumeuses, chargées de gouttelettes lipidiques et de cristaux de cholestérol. La plaque initiale, ou lésion fibrolipidique, est carac-térisée par la formation d’un cœur lipidique extracellulaire et la mort des CML spumeuses, qui seront recouvertes ultérieurement par une chape fibrocellulaire sécrétée par d’autres CML, plus résistantes à l’agression lipidiques [9]. En réponse à l’accumulation intimale de lipides, les CML de la média orchestrent la réponse angiogénique depuis l’adventice vers la plaque [12]. La néoangiogenèse intraplaque est l’étape cruciale signant l’évolution de la plaque vers un phénotype compliqué. La média artérielle est physiologiquement avasculaire. Elle est un site immun privilégié, peu ou pas accessible aux leucocytes circulants en l’absence de néovascularisation permettant leur extravasation. Immature et perméable, le néo-réseau veinocapillaire transpariétal où règne une pression propice à l’extravasation des leucocytes vers la plaque, sera le vecteur d’hémorragies intraplaques [13]. B. Le travail de Alexander et al. [8] montre que le blocage de la signalisation de l’IL-1 conduit à un remodelage sténosant et vulnérable des plaques (en rouge accolées à la face interne de la paroi vasculaire en bleue).

m/s n° 6-7, vol. 28, juin-juillet 2012 18
De phénomènes cellulaires tels que l’extravasation de leucocytes ? Si oui, quels phénomènes parmi ceux-ci et de quelles fonctions sont-ils dotés ? Le progrès médical ne peut se suffire de telles approximations, surtout, lorsque des essais thérapeutiques incluant la participation humaine sont en jeu. ◊Should we block interleukin-1 in atherothrombosis ?
LIENS D’INTÉRÊTSLes auteurs déclarent n’avoir aucun lien d’intérêt concernant les données publiées dans cet article.
RÉFÉRENCES
1. Ross R. Atherosclerosis-an inflammatory disease. N Engl J Med 1999 ; 340 : 115-26.
2. Clarke MC, Talib S, Figg NL, Bennett MR. Vascular smooth muscle cell apoptosis induces interleukin-1-directed inflammation: effects of hyperlipidemia-mediated inhibition of phagocytosis. Circ Res 2010 ; 106 : 363-72.
3. Duewell P, Kono H, Rayner KJ, et al. NLRP3 inflammasomes are required for atherogenesis and activated by cholesterol crystals. Nature 2010 ; 464 : 1357-61.
4. Ridker PM, Thuren T, Zalewski A, Libby P. Interleukin-1 beta inhibition and the prevention of recurrent cardiovascular events: rationale and design of the Canakinumab anti-inflammatory thrombosis outcomes study (CANTOS). Am Heart J 2011 ; 162 : 597-605.
5. Mertens M, Singh JA. Anakinra for rheumatoid arthritis: a systematic review. J Rheumatol 2009 ; 36 : 1118-25.
6. Lawrie A, Hameed AG, Chamberlain J, et al. Paigen diet-fed apolipoprotein E knockout mice develop severe pulmonary hypertension in an interleukin-1-dependent manner. Am J Pathol 2011 ; 179 : 1693-705.
7. Saxena A, Rauch U, Berg KE, et al. The vascular repair process after injury of the carotid artery is regulated by IL-1RI and MyD88 signalling. Cardiovasc Res 2011 ; 91 : 350-7.
8. Alexander MR, Moehle CW, Johnson JL, et al. Genetic inactivation of IL-1 signaling enhances atherosclerotic plaque instability and reduces outward vessel remodeling in advanced atherosclerosis in mice. J Clin Invest 2012 ; 122 : 70-9.
9. Lacolley P, Regnault V, Nicoletti A, et al. The vascular smooth muscle cell in arterial pathology: a cell that can take on multiple roles. Cardiovasc Res 2012 (sous-presse).
10. Ait-Oufella H, Salomon BL, Potteaux S, et al. Natural regulatory T cells control the development of atherosclerosis in mice. Nat Med 2006 ; 12 : 178-80.
11. Caligiuri G, Nicoletti A, Poirier B, Hansson GK. Protective immunity against atherosclerosis carried by B cells of hypercholesterolemic mice. J Clin Invest 2002 ; 109 : 745-53.
12. Ho-Tin-Noé B, Le Dall J, Gomez D et al. Early atheroma-derived agonists of peroxisome proliferator-activated receptor-g trigger intramedial angiogenesis in a smooth muscle cell-dependent manner. Circ Res 2011 ; 109 : 1003-14.
13. Le Dall J, Ho-Tin-Noé B, Louedec L et al. Immaturity of microvessels in haemorrhagic plaques is associated with proteolytic degradation of angiogenic factors. Cardiovasc Res 2010 ; 85 : 184-93.
La cristallisation du cholestérol dans le microenvironnement des CML est un évè-nement macromoléculaire probablement capital dans ces changements fonction-nels, comme cela a été montré pour le macrophage [3]. Les capacités cicatri-santes des CML limitent le risque de rup-ture des plaques, et leur mort participe de la pathogenèse de plaques de plus en plus complexes. Les CML mériteraient donc d’occuper une place plus impor-tante dans l’athérosclérose humaine. Or, l’implication préférentielle des leu-cocytes mononucléés, suggérée par les travaux chez la souris, nous incline à utiliser des stratégies anti-inflamma-toires possiblement inappropriées car ne pointant pas les bonnes cibles et/ou sans doute trop radicales.À cet égard, l’extravasation de monocytes et de lymphocytes vers la plaque est sou-vent présentée comme étant un proces-sus délétère, alors que la réponse immune innée et certaines réponses adaptatives peuvent être athéroprotectrices [10, 11]. Dans un tel contexte de mixité et de plas-ticité des cellules stromales et des cel-lules d’origine hématopoïétique, l’impact du blocage d’une cytokine telle que l’IL-1b, produite par et agissant sur les deux familles de cellules, est difficile à pré-dire. C’est précisément ce que montre le travail d’Alexander et al. [8] dans lequel l’inhibition de la signalisation de l’IL-1b n’est pas univoque et intègre à la fois des effets bénéfiques sur les leucocytes et la taille des lésions et des effets délétères sur le remodelage sténosant et vulnérable des plaques via les CML.L’ensemble de la communication scien-tifique autour de l’athérothrombose pose finalement des questions d’ordre sémantique, épistémologique et éthique autour de l’emploi abusif du mot inflammation. L’emploi de ce terme, ini-tialement issu de la séméiologie médi-cale, est, dans les sciences biologiques, dans la plupart des cas approximatif, ne traduisant pas de façon analytique le phénomène étudié. De quoi s’agit-il ? De signalisation intracellulaire ? De sécrétion de cytokines ou chémokines ?
ce malgré une réduction de la surface des plaques lipidiques (d’environ 30 %). Ceci s’explique par l’abolition chez ces souris du remodelage excentrique de la média pariétale en regard de la plaque, un pro-cessus de compensation qui permet le maintien du diamètre de la lumière arté-rielle malgré la croissance de la lésion athéromateuse. L’invalidation de l’IL-1RI s’accompagne également d’une diminu-tion de la taille de la chape fibreuse et cellulaire et de l’augmentation de la den-sité de globules rouges dans les plaques. Ces données suggèrent qu’en l’absence de signalisation de l’IL-1b, la maladie athéromateuse évolue défavorablement avec des sténoses plus sévères et des plaques plus vulnérables (Figure 1). Cette étude est donc un signal d’alarme sur les risques d’un blocage pharmacologique de la voie de l’IL-1.
Athérogenèse et inflammation : un paradigme abusif ? Ce travail offre l’opportunité de discuter plusieurs points essentiels concernant l’utilisation du terme inflammation dans la pathologie athéromateuse : existe-t-il une véritable relation de causalité entre l’extravasation leucocytaire dans les plaques et leur évolution morbide ? Quelles sont les limites des modèles murins dans la compréhension de la maladie humaine ? Le processus d’athérogenèse chez l’homme, succinctement décrit dans la Figure 1, met en lumière les limites des modèles murins dans lesquels l’angioge-nèse transpariétale et les hémorragies intraplaques sont extrêmement rares. Elle place aussi les CML au centre des évolu-tions successives de la pathologie chez l’homme : prolifération intimale initiale, cellules spumeuses, chape fibrocellulaire, organisation des réponses adventitielles [9]. Si les CML peuvent adopter des phé-notypes aussi divers que contractile, pro-lifératif, sécrétoire, phagocytaire, voire ostéoblastique, c’est qu’elles préservent une forte plasticité phénotypique chez l’adulte [9] (�).
(�) Voir la Nouvelle de K. Treguer et al., page ??? de ce numéro

m/s n° 6-7, vol. 28, juin-juillet 2012
NOUV
ELLE
SM
AGAZ
INE
19
NOUVELLE
L’astrocyte, un partenaire clé des neurones au cours de la transmission synaptique de baseAude Panatier1,2,3, Richard Robitaille1,2
1 Département de physiologie, Université de Montréal ; 2 Groupe de recherche sur le système nerveux central, université de Montréal, PO box 6128 Station centre-ville, Montréal, QC, H3C 3J7 Canada ;3 Inserm U862, neurocentre Magendie ; UMR 5297, institut interdisciplinaire des neurosciences, 146, rue Léo Saignat, 33077 Bordeaux Cedex, [email protected]
Le concept de la synapse tripartiteLe fonctionnement du cerveau repose sur la transmission d’informations de neurone en neurone, à travers une structure spécialisée, la synapse. Très simplement, l’information se propage sous la forme d’un signal électrique, le potentiel d’action, le long de l’axone neuronal. Lorsque le potentiel d’ac-tion arrive au niveau de la terminaison synaptique (élément présynaptique) ce signal électrique est converti en un signal chimique. Les neurotransmet-teurs libérés dans la fente synaptique activent par la suite des récepteurs présents sur l’élément postsynaptique et l’information est transmise.Il a longtemps été considéré que la trans-mission synaptique était uniquement une histoire de neurone. Cependant, en 1999, la synapse s’est complexifiée avec l’émergence du concept de la synapse tripartite qui considère les astrocytes comme des éléments fonctionnels des synapses [1]. Au cours de ces vingt der-nières années, de nombreux travaux réa-lisés dans plusieurs régions du cerveau ont révélé que les astrocytes étaient capables de détecter les transmetteurs libérés dans la fente synaptique, et d’y répondre [2-4].Les astrocytes appartiennent au groupe des cellules gliales qui, contrairement aux neurones, n’émettent pas de poten-tiel d’action. Parmi leurs nombreux rôles, les astrocytes régulent l’efficacité du transfert d’information en libérant par exemple des transmetteurs, appelés gliotransmetteurs (comme le glutamate, la D-sérine ou encore les purines) [5, 6].
Pour ce faire, au cours d’une activité synaptique intense qui recrute un réseau de neurones, les astrocytes détectent les neurotransmetteurs libérés dans la fente synaptique via des récepteurs exprimés à leur surface. L’activation de ces récepteurs provoque une augmenta-tion de calcium dans les astrocytes. En retour, ces cellules gliales libèrent des gliotransmetteurs qui se lieront à des récepteurs soit neuronaux présynap-tiques, soit neuronaux postsynaptiques afin de réguler l’efficacité du transfert d’informations [2, 3].En accord avec la vision classique, les astrocytes détectent les transmetteurs libérés dans la fente synaptique en réponse à l’arrivée de plusieurs poten-tiels d’actions dans l’élément présynap-tique. Cependant, en plus des activités synaptiques intenses faisant intervenir des réseaux synaptiques, la communica-tion neuronale a aussi lieu au niveau de synapses individuelles, à des fréquences beaucoup plus faibles. Cette forme de communication détermine le tonus synaptique des neurones et façonne les mécanismes de plasticité synaptique qui sous-tendent diverses formes d’appren-tissage. Afin de mieux comprendre com-ment l’information excitatrice glutama-tergique est transmise dans le cerveau, nous avons voulu savoir si les astrocytes étaient également capables de détec-ter et de réguler la libération d’une seule vésicule de neurotransmetteurs (glutamate, neurotransmetteur exci-tateur) formée en réponse à l’arrivée présumée d’un seul potentiel d’action dans l’élément présynaptique [7]. Cette
transmission, appelée transmission synaptique de base, est fondamentale pour le fonctionnement du cerveau.
Les astrocytes régulent la transmission synaptique de baseCette étude a été réalisée dans des tranches « aiguës1 » d’hippocampe de rat (région CA1), une région du cer-veau impliquée dans les processus de mémoire et d’apprentissage. Une combinaison d’approches de micros-copie confocale et multiphotonique et d’enregistrements électrophysiologiques a été utilisée.Les astrocytes sont caractérisés par la présence d’un prolongement prin-cipal et de nombreux prolongements qui leur confèrent une morphologie en forme « d’éponge ». Ces prolongements astrocytaires ne sont pas rectilignes. En effet, leur diamètre s’élargit le long de l’axe principal, puis s’affine à nou-veau, délimitant des compartiments morphologiques (taille d’un comparti-ment mesurée le long de l’axe principal du prolongement : environ 1 µm). Tout d’abord, nous avons mis en évidence que ces compartiments sont fonctionnels : ils interagissent avec les synapses et expriment à leur surface des récepteurs glutamatergiques.Au cours de la transmission synaptique de base, l’astrocyte détecte les molé-cules de glutamate libérées dans la fente synaptique via des récepteurs
1 Ce terme désigne des tranches de cerveau fraîchement prélevées et conservées dans un milieu artificiel pendant 6 heures afin de réaliser les expériences.

m/s n° 6-7, vol. 28, juin-juillet 2012 20
l’échelle des synapses individuelles - jusqu’à l’activité synaptique intense, à l’échelle d’un réseau. En fonction du niveau et du patron d’activité synap-tique, il est possible que le signal cal-cique se propage à l’échelle d’un pro-longement, voir de la cellule dans son ensemble. Une étape supplémentaire, serait de déterminer si ce signal cal-cique va avoir des conséquences sur l’efficacité de la transmission au niveau des synapses voisines, avec lesquelles l’astrocyte est en interaction. Désor-mais, si nous voulons comprendre les fonctionnements et les dysfonctionne-ments du cerveau, nous devons consi-dérer l’astrocyte comme un partenaire intime des neurones. ◊Astrocyte, a key partner of neurones during basal synaptic transmission
LIENS D’INTÉRÊTSLes auteurs déclarent n’avoir aucun lien d’intérêt concernant les données publiées dans cet article.
RÉFÉRENCES
1. Araque A, Parpura V, Sanzgiri RP, Haydon PG. Tripartite synapses: glia, the unacknowledged partner. Trends Neurosci 1999 ; 22 : 208-15.
2. Perea G, Navarrete M, Araque A. Tripartite synapses: astrocytes process and control synaptic information. Trends Neurosci 2009 ; 32 : 421-31.
3. Halassa MM, Haydon PG. Integrated brain circuits: astrocytic networks modulate neuronal activity and behavior. Annu Rev Physiol 2010 ; 72 : 335-55.
4. Tasker JG, Oliet SH, Bains JS, et al. Glial regulation of neuronal function: from synapse to systems physiology. J Neuroendocrinol 2012 ; 24 : 566-76.
5. Volterra A, Meldolesi J. Astrocytes, from brain glue to communication elements: the revolution continues. Nat Rev Neurosci 2005 ; 6 : 626-40.
6. Parpura V, Heneka MT, Montana V, et al. Glial cells in (patho)physiology. J Neurochem 2012 ; 121 : 4-27.
7. Panatier A, Vallée J, Haber M, Murai KK, Lacaille JC, Robitaille R. Astrocytes are endogenous regulators of basal transmission at central synapses. Cell 2011 ; 146 : 785-98.
8. Di Castro MA, Chuquet J, Liaudet N, et al. Local Ca2+ detection and modulation of synaptic release by astrocytes. Nat Neurosci 2011 ; 14 : 1276-84.
En résumé, nos travaux indiquent que l’as-trocyte est un partenaire clé des neurones au cours de la transmission synaptique de base. Tout comme les neurones, les astro-cytes sont capables de traiter l’informa-tion localement, au niveau de compar-timents fonctionnels. Il est important de noter que cette sensibilité astrocytaire a également été rapportée récemment dans une autre région de l’hippocampe [8]. Désormais, la communication élémentaire ne peut plus être considérée comme un phénomène uniquement neuronal.
PerspectivesIl semble que l’astrocyte agisse comme un intégrateur d’informations, depuis la transmission synaptique de base - à
métabotropiques du glutamate de type 5 (mGluR5, Figure 1). L’activation de ces récepteurs conduit à une augmentation de calcium dans le compartiment astro-cytaire. En retour, l’astrocyte libère des purines via un mécanisme mettant en jeu les protéines de type SNARE (soluble N-ethylmaleimide-sensitive factor attachment protein receptor), sensibles à la toxine tétanique. Une fois libérées dans la fente synaptique, les purines activent des récepteurs à l’adénosine de type A2A (A2AR, Figure 1) présents sur la surface de l’élément neuronal présy-naptique. L’activation de ces récepteurs conduit alors à une facilitation de la transmission synaptique de base au niveau de cette synapse (Figure 1).
Neuroneprésynaptique
Neuronepostsynaptique
mGluR5
A2AR
Ca2+
+
++
PurinesGlutamate
AMPANMDA
Astrocyte Potentield’action
Figure 1. Les astrocytes facilitent la transmission synaptique de base. Représentation schématique résumant l’implication des astrocytes au cours de la transmission synaptique de base. AMPA et NMDA représentent les récepteurs glutamatergiques de type AMPA et NMDA. mGluR5, récepteur métabotropique du glutamate de type 5 ; A2AR, récepteur à l’adénosine de type A2A (adapté de [7]).

m/s n° 6-7, vol. 28, juin-juillet 2012
NOUV
ELLE
SM
AGAZ
INE
21
1 Centre for Molecular Medicine, Institute of Cardiovascular Regeneration, University of Frankfurt Medical School, 60590 Frankfurt am Main, Allemagne ; 2 adresse actuelle : UMR CNRS 5164, Université Bordeaux 2, 146, rue Léo Saignat, 33076 Bordeaux, [email protected]
> L’athérosclérose, maladie fréquente correspondant à la modification patho-logique des artères, est la cause de nom-breuses maladies telles que l’infarctus du myocarde et les accidents vasculaires cérébraux. Dans les pays développés, ces pathologies associées représen-tent la première cause de mortalité et constituent un enjeu important de santé publique. L’athérosclérose est un phé-nomène progressif normal au cours de
la vie, mais qui peut devenir patholo-gique s’il s’accélère ou s’amplifie. En effet, les artères sont le siège de dépôts évoluant avec l’âge au niveau de leur paroi : c’est le vieillissement artériel. Ce processus peut être aggravé par les perturbations du flux sanguin perçues par les cellules endothéliales (CE). Dans les zones de turbulences, en général proche des bifurcations artérielles ou d’une plaque d’athérome, le flux sanguin
n’est plus laminaire, il est turbulent, ce qui induit une diminution des forces de cisaillement1. Cette information va être transmise aux cellules musculaires lisses (CML) qui entourent les vaisseaux. Les CML vont alors s’activer, se dédiffé-rencier et proliférer, ce qui accélère et aggrave le phénomène d’athérosclérose naturel [1] (Figure 1). Ces dernières années, le facteur de transcription Krüp-pel-like factor 2 (KLF2) a été identifié
comme un senseur jouant un rôle clé dans les effets béné-fiques athéroprotecteurs du flux laminaire dans les vaisseaux [2]. En l’absence de KLF2 dans les CE, les souris meurent au stade embryonnaire E10,5 ; leur phénotype associe une absence de tonus vasculaire, des hémor-ragies et des dysfonctions car-diaques secondaires à des com-plications vasculaires [3, 4]. De façon intéressante, l’endothé-lium de ces souris est normal, ce qui n’est pas le cas des CML, dysfonctionnelles et désorgani-sées. Cette observation indique que l’expression endothéliale de KLF2 exerce un rôle physio-logique important sur la couche sous jacente de CML [5]. Nous nous sommes donc intéressés au mécanisme de cette communi-
1 Les forces de cisaillement ou contraintes de cisaillement sont les forces de friction exer-cées par le sang sur la paroi du vaisseau. Elles dépendent du débit sanguin, de la viscosité du sang et du diamètre du vaisseau. En l’absence d’obstacle ou de bifurcation, le flux est laminaire et les forces de cisaillement sont maximales.
Protection des vaisseaux sanguins contre l’athéroscléroseLe rôle des miARN sécrétés par l’endothéliumKarine Tréguer1,2, Susanne Heydt1, Eduard Hergenreider1
NOUVELLE
KLF2
Cellule musculaire lisse (CML)Cellule endothéliale (CE)Lymphocyte TMacrophage
Goutelettes lipidiquesCholésterolFlux sanguin laminaireFlux sanguin turbulent
Intima
Média
Adventice ?
Phénotypeprolifératif
Athérosclérose
KLF2
?
Phénotypecontractile
Athéroprotection
Figure 1. Effet du flux dans la mise en place de la plaque d’athérome. Cadre supérieur : le flux laminaire induit l’expression de KLF2 dans les cellules endothéliales (CE). KLF2 est responsable du maintien du phéno-type contractile et athéroprotecteur des cellules musculaires lisses (CML) sous-jacentes. Cadre inférieur : lorsque le flux est turbulent, aux abords d’une bifurcation ou d’une plaque d’athérome, l’expression de KLF2 dans les CE diminue et les CML sous-jacentes adoptent un phénotype prolifératif proathérosclérotique. La communication entre les CE et les CML via KLF2 fait l’objet de cette étude.

m/s n° 6-7, vol. 28, juin-juillet 2012 22
forces de cisaillement dans les CE. Cette double analyse a permis d’identifier de nouvelles cibles de KLF2 en réponse au flux : le cluster mir-143/145. Nous avons validé ces miARN candidats par invalidation de KLF2 in vitro, et nous avons montré qu’en absence de KLF2, ce cluster n’est plus induit par les forces de cisaillement ; de plus, KLF2 est capable de se lier au promoteur de ce cluster et d’induire son expression. Les forces de cisaillement induisent donc l’expres-sion de KLF2 dans les CE, et celui-ci, à son tour, induit l’expression de miR-143/145. Ce cluster était jusque là connu
ou à la dégradation des ARNm cibles [7, 17]. Ces molécules sont déjà connues pour leur rôle important dans la vas-cularisation ainsi que dans différentes pathologies vasculaires [8]. Notre objectif a été de déterminer si KLF2 pouvait réguler l’expression de miARN dans les CE et contribuer à ces effets vasculoprotecteurs.
Les forces de cisaillement régulent l’expression de miARN dans les CE via KLF2Nous avons tout d’abord déterminé les miARN régulés par KLF2 et par les
cation entre les CE et les CML conduisant à l’effet athéroprotecteur relayé par KLF2 [6] (Figure 1).
Les miARN : un nouveau niveau de régulation de l’expression géniqueUn candidat de choix dans cette com-munication entre CE et CML seraient les microARN (miARN). Les miARN sont de petits ARN non codants simple brin (22 nucléotides) qui sont transcrits dans le noyau, clivés notamment par les enzymes Drosha et Dicer et incorpo-rés dans un complexe de silencing qui conduit à l’inhibition de la traduction
Surnageant
Récupérer lesurnageant
13 500 x g ; 5 min
20 500 x g ; 45 min
Concentrer lesurnageant et isoler les ARN
Laver le culot au PBS20 500 x g ; 45 minCulot de microvésicules(100 - 1 000 nm) contient les MV
miARN Protéine de liaison aux miARN
membrane bicouche-phospholipidique
miARN dégradés
Protéinase K
Triton-X 100PhospholipaseCyclodextrine
RNase
RNase
Analyse biochimique de la protection des miARN extracellulaires
Membrane poreuse (pores 0,4 µm)
Cellules musculaires lisses
Cellules endothéliales transduites par eGFP ou transfectées par cel-miR-39
Cellule musculairelisse (CML)
Cellule endothéliale (CE)
Microvésicule (MV)
miARN
Étude du transfert de matériel intercellulaire
A
B
Figure 2. Analyse de la communication intercellulaire. A. Analyse biochimique de la protection des miARN extracellulaires. L’échantillon est traité à la protéinase K ou au détergent pour dégrader les protéines ou les membranes lipidiques, puis les ARN sont dégradés par traitement à la Rnase. S’ils sont dégradés, cela indique qu’ils étaient accessibles et informe sur le type de protection, protéique ou membranaire. B. Système de coculture permettant d’étudier le transfert de matériel entre les CE et les CML.
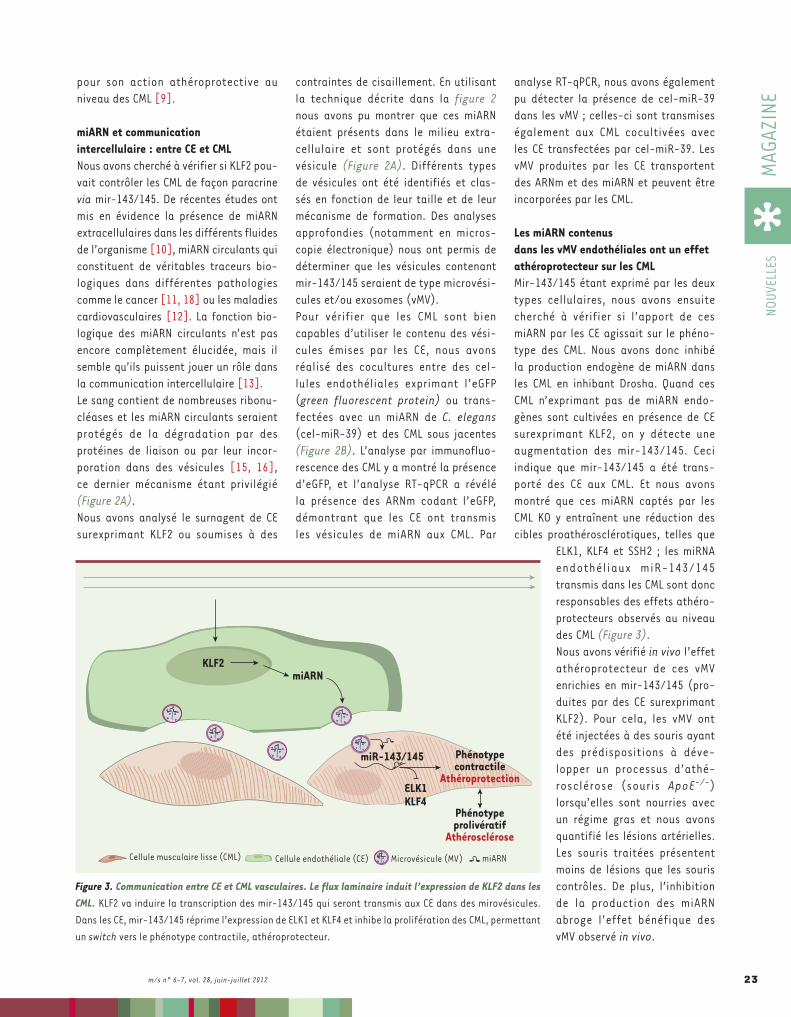
m/s n° 6-7, vol. 28, juin-juillet 2012
NOUV
ELLE
SM
AGAZ
INE
23
analyse RT-qPCR, nous avons également pu détecter la présence de cel-miR-39 dans les vMV ; celles-ci sont transmises également aux CML cocultivées avec les CE transfectées par cel-miR-39. Les vMV produites par les CE transportent des ARNm et des miARN et peuvent être incorporées par les CML.
Les miARN contenus dans les vMV endothéliales ont un effet athéroprotecteur sur les CMLMir-143/145 étant exprimé par les deux types cellulaires, nous avons ensuite cherché à vérifier si l’apport de ces miARN par les CE agissait sur le phéno-type des CML. Nous avons donc inhibé la production endogène de miARN dans les CML en inhibant Drosha. Quand ces CML n’exprimant pas de miARN endo-gènes sont cultivées en présence de CE surexprimant KLF2, on y détecte une augmentation des mir-143/145. Ceci indique que mir-143/145 a été trans-porté des CE aux CML. Et nous avons montré que ces miARN captés par les CML KO y entraînent une réduction des cibles proathérosclérotiques, telles que
ELK1, KLF4 et SSH2 ; les miRNA endothéliaux miR-143/145 transmis dans les CML sont donc responsables des effets athéro-protecteurs observés au niveau des CML (Figure 3).Nous avons vérifié in vivo l’effet athéroprotecteur de ces vMV enrichies en mir-143/145 (pro-duites par des CE surexprimant KLF2). Pour cela, les vMV ont été injectées à des souris ayant des prédispositions à déve-lopper un processus d’athé-rosclérose (souris ApoE-/-) lorsqu’elles sont nourries avec un régime gras et nous avons quantifié les lésions artérielles. Les souris traitées présentent moins de lésions que les souris contrôles. De plus, l’inhibition de la production des miARN abroge l’effet bénéfique des vMV observé in vivo.
contraintes de cisaillement. En utilisant la technique décrite dans la figure 2 nous avons pu montrer que ces miARN étaient présents dans le milieu extra-cellulaire et sont protégés dans une vésicule (Figure 2A). Différents types de vésicules ont été identifiés et clas-sés en fonction de leur taille et de leur mécanisme de formation. Des analyses approfondies (notamment en micros-copie électronique) nous ont permis de déterminer que les vésicules contenant mir-143/145 seraient de type microvési-cules et/ou exosomes (vMV).Pour vérifier que les CML sont bien capables d’utiliser le contenu des vési-cules émises par les CE, nous avons réalisé des cocultures entre des cel-lules endothéliales exprimant l’eGFP (green fluorescent protein) ou trans-fectées avec un miARN de C. elegans (cel-miR-39) et des CML sous jacentes (Figure 2B). L’analyse par immunofluo-rescence des CML y a montré la présence d’eGFP, et l’analyse RT-qPCR a révélé la présence des ARNm codant l’eGFP, démontrant que les CE ont transmis les vésicules de miARN aux CML. Par
pour son action athéroprotective au niveau des CML [9].
miARN et communication intercellulaire : entre CE et CMLNous avons cherché à vérifier si KLF2 pou-vait contrôler les CML de façon paracrine via mir-143/145. De récentes études ont mis en évidence la présence de miARN extracellulaires dans les différents fluides de l’organisme [10], miARN circulants qui constituent de véritables traceurs bio-logiques dans différentes pathologies comme le cancer [11, 18] ou les maladies cardiovasculaires [12]. La fonction bio-logique des miARN circulants n’est pas encore complètement élucidée, mais il semble qu’ils puissent jouer un rôle dans la communication intercellulaire [13].Le sang contient de nombreuses ribonu-cléases et les miARN circulants seraient protégés de la dégradation par des protéines de liaison ou par leur incor-poration dans des vésicules [15, 16], ce dernier mécanisme étant privilégié (Figure 2A).Nous avons analysé le surnagent de CE surexprimant KLF2 ou soumises à des
KLF2miARN
miR-143/145 Phénotypecontractile
AthéroprotectionELK1KLF4
Phénotypeprolivératif
Athérosclérose
Cellule musculaire lisse (CML) Cellule endothéliale (CE) Microvésicule (MV) miARN
Figure 3. Communication entre CE et CML vasculaires. Le flux laminaire indu it l’expression de KLF2 dans les CML. KLF2 va induire la transcription des mir-143/145 qui seront transmis aux CE dan s des mirovésicules. Dans les CE, mir-143/145 réprime l’expression de ELK1 et KLF4 et inhibe la prolifération des CML, permettant un switch vers le phénotype contractile, athéroprotecteur.

m/s n° 6-7, vol. 28, juin-juillet 2012 24
7. Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell 2009 ; 136 : 215-33.
8. Bonauer A, Boon RA, Dimmeler S. Vascular microRNAs. Curr Drug Targets 2010 ; 11 : 943-9.
9. Rangrez AY, Massy ZA, Metzinger-Le Meuth V, Metzinger L. miR-143 and miR-145: molecular keys to switch the phenotype of vascular smooth muscle cells. Circ Cardiovasc Genet 2011 ; 4 : 197-205.
10. Weber JA, Baxter DH, Zhang S, et al. The microRNA spectrum in 12 body fluids. Clin Chem 2010 ; 56 : 1733-41.
11. Schwarzenbach H, Hoon DS, Pantel K. Cell-free nucleic acids as biomarkers in cancer patients. Nat Rev Cancer 2011 ; 11 : 426-37.
12. Fichtlscherer S, Zeiher AM, Dimmeler S. Circulating microRNAs: biomarkers or mediators of cardiovascular diseases? Arterioscler Thromb Vasc Biol 2011 ; 31 : 2383-90.
13. Creemers EE, Tijsen AJ, Pinto YM. Circulating microRNAs: novel biomarkers and extracellular communicators in cardiovascular disease? Circ Res 2012 ; 110 : 483-95.
14. Weickmann JL, Glitz DG. Human ribonucleases. Quantitation of pancreatic-like enzymes in serum, urine, and organ preparations. J Biol Chem 1982 ; 257 : 8705-10.
15. Wang K, Zhang S, Weber J, et al. Export of microRNAs and microRNA-protective protein by mammalian cells. Nucleic Acids Res 2010 ; 38 : 7248-59.
16. Arroyo JD, Chevillet JR, Kroh EM, et al. Argonaute2 complexes carry a population of circulating microRNAs independent of vesicles in human plasma. Proc Natl Acad Sci USA 2011 ; 108 : 5003-8.
17. Patrice Dunoyer. La bataille du silence : mécanisme et inhibition du RNA silencing au cours des interactions plante/virus. Med Sci (Paris) 25 ; 5 : 505-12.
18. Ladeiro Y, Zucman-rossi J. Micro-ARN (miARN) et cancer : le cas des tumeurs hépatocellulaires. Med Sci (Paris) 25 ; 5 : 467-72.
d’adresser spécifiquement les miARN aux cellules cibles à traiter. De plus, les cellules productrices issues du patient peuvent être manipulées afin de modifier le contenu des vMV qui seront délivrées aux cellules ciblées. ◊miRNA secreted in vesicles allow atheroprotective communication in vessel wall
LIENS D’INTÉRÊTLes auteurs déclarent n’avoir aucun lien d’intérêt concernant les données publiées dans cet article.
RÉFÉRENCES
1. Malek AM, Alper SL, Izumo S. Hemodynamic shear stress and its role in atherosclerosis. JAMA 1999 ; 282 : 2035-42.
2. Boon RA, Horrevoets AJ. Key transcriptional regulators of the vasoprotective effects of shear stress. Hamostaseologie 2009 ; 29 : 39-40.
3. Kuo CT, Veselits ML, Barton KP, et al. The LKLF transcription factor is required for normal tunica media formation and blood vessel stabilization during murine embryogenesis. Genes Dev 1997 ; 11 : 2996-3006.
4. Lee JS, Yu Q, Shin JT, et al. Klf2 is an essential regulator of vascular hemodynamic forces in vivo. Dev Cell 2006 ; 11 : 845-57.
5. Wu J, Bohanan CS, Neumann JC, Lingrel JB. KLF2 transcription factor modulates blood vessel maturation through smooth muscle cell migration. J Biol Chem 2008 ; 283 : 3942-50.
6. Hergenreider E, Heydt S, Treguer K, et al. Atheroprotective communication between endothelial cells and smooth muscle cells through miRNAs. Nat Cell Biol 2012 ; 14 : 249-56.
Applications thérapeutiquesCes travaux ont permis de mettre en évidence une communication entre les CE et les CML via les miARN circulants. En réponse au flux sanguin, l’activa-tion de KLF2 permet la production des mir143/145 qui sont transmis aux CML via des vMV. Ces miARN dans les CML vont inhiber l’expression de facteurs proathérogéniques (Figure 3).Ces résultats encourageants pourraient susciter des applications thérapeu-tiques intéressantes. Il pourrait être possible, grâce aux vMV, de réduire le développement des plaques d’athérome chez les personnes à risque, notamment les personnes obèses. Cette stratégie pourrait s’appliquer à d’autres patho-logies où l’expression des miARN est dérégulée (cancer, maladies cardio-vasculaires), car les vMV constituent un moyen de ciblage des traitements par miARN. Comme nous l’avons montré pour l’athérosclérose, l’administration des miARN via les vMV pourrait se faire par voie intraveineuse. Les vMV seraient produites in vitro à partir des cellules du patient lui même, réduisant ainsi le risque d’effets secondaires. Le choix du type cellulaire donneur permettrait




















