
Timing of maternal exposure to a high fat diet and development of obesity and hyperinsulinemia in...
11
Hindawi Publishing Corporation Journal of Nutrition and Metabolism Volume 2013, Article ID 517384, 11 pages http://dx.doi.org/10.1155/2013/517384 Research Article Timing of Maternal Exposure to a High Fat Diet and Development of Obesity and Hyperinsulinemia in Male Rat Offspring: Same Metabolic Phenotype, Different Developmental Pathways? Graham J. Howie, 1 Deborah M. Sloboda, 1,2 Clare M. Reynolds, 1 and Mark H. Vickers 1 1 Liggins Institute and Gravida: National Centre for Growth and Development, University of Auckland, 2-6 Park Avenue, Graſton, Auckland 1010, New Zealand 2 Department of Biochemistry and Biomedical Sciences, McMaster University, Hamilton, ON, Canada L8S 4L8 Correspondence should be addressed to Mark H. Vickers; [email protected] Received 11 February 2013; Revised 8 April 2013; Accepted 20 April 2013 Academic Editor: Peter M. Cliſton Copyright © 2013 Graham J. Howie et al. is is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Objective. Offspring born to mothers either fed an obesogenic diet throughout their life or restricted to pregnancy and lactation demonstrate obesity, hyperinsulinemia, and hyperleptinemia, irrespective of their postweaning diet. We examined whether timing of a maternal obesogenic diet results in differential regulation of pancreatic adipoinsular and inflammatory signaling pathways in offspring. Methods. Female Wistar rats were randomized into 3 groups: (1) control (CONT): fed a control diet preconceptionally and during pregnancy and lactation; (2) maternal high fat (MHF): fed an HF diet throughout their life and during pregnancy and lactation; (3) pregnancy and lactation HF (PLHF): fed a control diet throughout life until mating, then HF diet during pregnancy and lactation. Male offspring were fed the control diet postweaning. Plasma and pancreatic tissue were collected, and mRNA concentrations of key factors regulating adipoinsular axis signaling were determined. Results. MHF and PLHF offspring exhibited increased adiposity and were hyperinsulinemic and hyperleptinemic compared to CONT. Despite a similar anthropometric phenotype, MHF and PLHF offspring exhibited distinctly different expression for key pancreatic genes, dependent upon maternal preconceptional nutritional background. Conclusions. ese data suggest that despite using differential signaling pathways, obesity in offspring may be an adaptive outcome of early life exposure to HF during critical developmental windows. 1. Introduction Early life events contribute substantially to the likelihood of an individual becoming obese, although underlying mech- anisms are not well understood. Obesity in women of reproductive age (15 to 44 years) is increasing rapidly, and up to 50% of women in this age range in the USA are now either overweight or obese [1]. is has translated to an exponential increase in the prevalence of obesity during pregnancy with up to 20% of women entering pregnancy with a BMI which would define them as obese [2]. Obesity in pregnancy increases the risks for complications of pregnancy including miscarriage, hypertension, and gestational diabetes [3–5]. Furthermore, it is now well established that maternal obesity leads to an increased risk of obesity and metabolic and cardiovascular disorders in offspring [6–9]. In view of the rising prevalence of obesity in pregnancy and its association with adverse maternal and offspring outcomes, there is a great deal of interest in understanding the mechanistic pathways that link maternal obesity and excess maternal nutrition to increased risk of disease in childhood and beyond [10–13]. Clinical and experimental studies have consistently shown that maternal obesity predicts obesity, associated metabolic comorbidities, and reproductive disorders in adult offspring [2, 14–16], oſten underpinned by a proinflammatory status. However, there is a need to understand the relative contributions and interactions between maternal prepreg- nancy obesity and a gestational obesogenic environment
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Timing of maternal exposure to a high fat diet and development of obesity and hyperinsulinemia in...

Hindawi Publishing CorporationJournal of Nutrition and MetabolismVolume 2013 Article ID 517384 11 pageshttpdxdoiorg1011552013517384
Research ArticleTiming of Maternal Exposure to a High Fat Dietand Development of Obesity and Hyperinsulinemiain Male Rat Offspring Same Metabolic PhenotypeDifferent Developmental Pathways
Graham J Howie1 Deborah M Sloboda12 Clare M Reynolds1 and Mark H Vickers1
1 Liggins Institute and Gravida National Centre for Growth and Development University of Auckland2-6 Park Avenue Grafton Auckland 1010 New Zealand
2Department of Biochemistry and Biomedical Sciences McMaster University Hamilton ON Canada L8S 4L8
Correspondence should be addressed to Mark H Vickers mvickersaucklandacnz
Received 11 February 2013 Revised 8 April 2013 Accepted 20 April 2013
Academic Editor Peter M Clifton
Copyright copy 2013 Graham J Howie et al This is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in any medium provided the original work is properlycited
Objective Offspring born to mothers either fed an obesogenic diet throughout their life or restricted to pregnancy and lactationdemonstrate obesity hyperinsulinemia and hyperleptinemia irrespective of their postweaning diet We examined whether timingof a maternal obesogenic diet results in differential regulation of pancreatic adipoinsular and inflammatory signaling pathways inoffspring Methods Female Wistar rats were randomized into 3 groups (1) control (CONT) fed a control diet preconceptionallyand during pregnancy and lactation (2) maternal high fat (MHF) fed an HF diet throughout their life and during pregnancy andlactation (3) pregnancy and lactation HF (PLHF) fed a control diet throughout life until mating then HF diet during pregnancyand lactation Male offspring were fed the control diet postweaning Plasma and pancreatic tissue were collected and mRNAconcentrations of key factors regulating adipoinsular axis signaling were determined ResultsMHF and PLHF offspring exhibitedincreased adiposity and were hyperinsulinemic and hyperleptinemic compared to CONT Despite a similar anthropometricphenotype MHF and PLHF offspring exhibited distinctly different expression for key pancreatic genes dependent upon maternalpreconceptional nutritional background ConclusionsThese data suggest that despite using differential signaling pathways obesityin offspring may be an adaptive outcome of early life exposure to HF during critical developmental windows
1 Introduction
Early life events contribute substantially to the likelihood ofan individual becoming obese although underlying mech-anisms are not well understood Obesity in women ofreproductive age (15 to 44 years) is increasing rapidly andup to 50 of women in this age range in the USA arenow either overweight or obese [1] This has translated toan exponential increase in the prevalence of obesity duringpregnancywith up to 20ofwomen entering pregnancywitha BMI which would define them as obese [2] Obesity inpregnancy increases the risks for complications of pregnancyincludingmiscarriage hypertension and gestational diabetes[3ndash5] Furthermore it is now well established that maternal
obesity leads to an increased risk of obesity and metabolicand cardiovascular disorders in offspring [6ndash9] In view of therising prevalence of obesity in pregnancy and its associationwith adversematernal and offspring outcomes there is a greatdeal of interest in understanding the mechanistic pathwaysthat link maternal obesity and excess maternal nutrition toincreased risk of disease in childhood and beyond [10ndash13]
Clinical and experimental studies have consistentlyshown that maternal obesity predicts obesity associatedmetabolic comorbidities and reproductive disorders in adultoffspring [2 14ndash16] often underpinned by a proinflammatorystatus However there is a need to understand the relativecontributions and interactions between maternal prepreg-nancy obesity and a gestational obesogenic environment
2 Journal of Nutrition and Metabolism
on subsequent health and well-being of offspring We havepreviously shown that obesity in adult rat offspring arising asa consequence of maternal high fat (HF) nutrition was inde-pendent of the pre-conceptional maternal diet [14] We haveshown that offspring of mothers fed an HF diet restricted topregnancy and lactation compared to those born to mothersfed a HF diet prior to conception and throughout pregnancyand lactation both exhibit a remarkably similar obesogenicphenotype characterised by increased fat mass hyperin-sulinemia and hyperleptinemia These data suggested thatdespite a common obesity phenotype differential mecha-nisms based on the timing of maternal exposure to HF nutri-tion may underpin pancreatic adipoinsular axis dysfunctionin these offspring [17ndash19]We therefore investigated key com-ponents of the leptin-insulin (adipoinsular) and inflamma-tory signaling pathways in pancreatic tissue of offspring bornto mothers fed a high fat diet during pregnancy in the pres-ence or absence of pre-conceptional HF diet-induced obesity
2 Methods
The rat model of maternal high fat nutrition has beendescribed by our group previously [14 15 20ndash24] In briefmale and female Wistar rats were acquired at a weaning age(22 days Vernon Jansen Unit University of Auckland) andhoused two per cage under standard conditions with a 1212light dark cycle and free access to water Females were weight-matched and assigned to receive either standard rat chow(119899 = 16 5 fat Diet 2018 Harlan Teklad Oxon) or a highfat (HF) diet (119899 = 8 24 fat D12451 Research Diets NJUSA) to be fed ad libitum for the duration of the trial Maleswere fed standard chow ad libitum for the duration of thepremating period Body weights were recorded every 3 daysuntil postnatal day 120
At postnatal day 110 body composition in females wasquantified using dual energy X-ray absorptiometry (DEXALunar Prodigy GE Medical Systems Madison WI USA) Atpostnatal day 120 females were time-mated using an estruscycle monitor (Model EC40 Fine Science Tools CA USA)Upon confirmation of mating three maternal dietary groupswere established (1) controls (CONT) females fed a standardchow diet throughout their life andmaintained on a standardchow diet throughout pregnancy and lactation (2) maternalhigh fat diet (MHF) females fed an HF diet throughout theirlife and maintained on the HF diet throughout pregnancyand lactation and (3) pregnancy + lactation HF diet (PLHF)females fed a standard chow diet until conception and thenan HF diet throughout pregnancy and lactation All pregnantdams were weighed and had food intakes measured dailythroughout pregnancy Following birth pup weights andbody lengths were recorded and litter size was randomlyadjusted to 8 pups per litter to ensure standardized nutritionuntil weaning Body weights and food intakes of dams weremeasured throughout the lactation period andoffspringwereweighed every three days until weaning
At weaning (d22) offspring were housed 2 per cage (2per littermaternal background) and fed a chow diet until theend of the trial At postnatal day 150 body composition wasmeasured in male offspring (119899 = 10ndash18 per group) by DEXA
At postnatal day 160 animals were fasted overnight and killedby decapitation following anaesthesia with sodium pentobar-bitone (60mgkg IP) Blood was collected into heparinizedtubes centrifuged and plasma supernatant stored for futureanalysis Whole pancreata were removed and immediatelysnap-frozen in liquid nitrogen for later molecular analysisAll animal experiments were approved under guidelines ofthe Animal Ethics Committee at the University of Auckland(Approval R652)
21 Plasma Analysis Fasting plasma leptin and insulin con-centrations were analysed using commercial rat-specificELISAs (CrystalChem 90040 and 90060 resp DownersGrove IL USA) Fasting whole blood glucose concentrationswere measured using a glucose meter at the time of tissuecollection (d160) (Roche AccuChek) Plasma tumor necrosisfactor (TNF-) alpha interleukin (IL-) 1120573 and IL-6 weremeasured using commercially available ELISAs (Quantikinekits RTA00 RLB00 and R6000B resp RampD SystemsEurope Abingdon UK) The homeostatic model assessment(HOMA)method was used as a measure of insulin resistance(IR) and beta-cell function (HOMA-IR = plasma glucose timesinsulin225) [25 26]
22 Gene Expression Quantitative PCR methods have beendescribed previously in detail [23 27] Briefly whole pancreastissue was disrupted using a homogenizer and total RNAwasextracted using commercially available kits (Qiagen Mini-Prep catalogue number 80204) according to the manufac-turerrsquos instructions RNA integrity and quality were assessedusing a NanoDrop spectrophotometer (ND-1000 BioLabLtd) using NanoDrop software (version 312) where sampleratio A260280 and A260230 were sim20 and intact bandswere visualised by gel electrophoresis Complimentary (c)DNA was synthesised from RNA (5 120583g) by reverse transcrip-tion PCR using MMLV-RT (Invitrogen CA USA) Quanti-tative PCR assays were performed in 15 120583L reaction volumesusing EXPRESS SYBR GreenER (Invitrogen) and fluores-cence was measured and quantified using an ABI-7900HTVer23 Sequence Detection System (Applied Bio SystemsCA USA) The qPCR thermal profile for amplification of allgenes was as follows melting at 95∘C for 15 secs followed byannealingextending at 60∘C for 1min for 40 cycles
Primers were obtained either commercially or designedde novo (Table 1) using NCBI ldquoPrimer-BLASTrdquo A standardcurve was generated using 6-fold serial 1 5 dilutions ofpooled stock cDNA All samples were assayed in triplicateand only those assays displaying a single PCR peak inthe melt-curve analysis were used All qPCR results werenormalized against the geometricmean of housekeeper genescyclophilin and HPRT according to the geNorm method ofVandesompele et al [28] TNF IL-1R1 andCD68 (and respec-tive controlsHPRTand cyclophilin)were analysed usingTaq-man gene expression assays To control for between-samplevariability mRNA levels were normalized to the geometricmean of the controls The ΔCt for each treatment sample wascompared to the mean ΔCt for control samples using therelative quantification 2-(ΔΔCt) method to determine fold-change [29]
Journal of Nutrition and Metabolism 3
Table 1 Sequences of forward and reverse primers
Entrez geneID Forward primer Reverse primer
K+ channel (Kir62) 83535 51015840-GAA GGA GGC AAA TGA TTG GA-31015840 51015840-AGT GTC CCC CAG ACA AAG TG-31015840
PDE3B 29516 51015840-GAC CGT CGT TGC CTT GTA TT-31015840 51015840-CGA TCG CCT TTC TCT ACT GG-31015840
SOCS3 89829 51015840-TTC TTT ACC ACC GAC GGA AC-31015840 51015840-CGT TGA CAG TCT TCC GAC AA-31015840
Insulin1 24505 Commercially prepared primer QuantiTect Primer Assay Cat no QT00373303 (Qiagen)Insulin2 24506 Commercially prepared primer QuantiTect Primer Assay Cat no QT00177380 (Qiagen)Insulin receptor 24954 51015840-ATC CGT CGC TCC TAT GCT CT-31015840 51015840-TCG TGA GGT TGT GCT TGT TC-31015840
Leptin 25608 Commercially prepared primer QuantiTect Primer Assay Cat no QT00190960 (Qiagen)ObRa 24536 51015840-TGA TAT CGC CAA ACA GCA AA-31015840 51015840-AGT GTC CGC TCT CTT TTG GA-31015840
ObRb 24536 51015840-AAA GCC TGA AAC ATT TGA GCA TC-31015840 51015840-CCA GAA GAA GAG GAC CAA ATA TCA C-31015840
Pdx1 29535 Commercially prepared primer QuantiTect Primer Assay Cat no QT00405328 (Qiagen)STAT3 25125 Commercially prepared primer QuantiTect Primer Assay Cat no QT00183512 (Qiagen)STAT5B 25126 Commercially prepared primer QuantiTect Primer Assay Cat no QT00192024 (Qiagen)CD68 287435 Commercially prepared primer TaqMan expression assay Cat no Rn01495634 (Applied Biosystems)TNF-120572 24835 Commercially prepared primer TaqMan expression assay Cat no Rn01525859 (Applied Biosystems)Cyclophilin 25518 51015840-TTG GGT CGC GTC TGC TTC GA-31015840 51015840-GCC AGG ACC TGT ATG CTT CA-31015840
HPRT 24465 51015840-AGT CCC AGC GTC GTG ATT AG-31015840 51015840-CCC CCT TCA GCA CAC AGA-31015840
23 Statistics All data were analysed by analysis of variance(ANOVA) with maternal diet as factor with post hoc testingto compare groups of interest as required (Holm-Sidak)Data with nonnormal distributions were log transformed ifrequired Analysis was performed using StatView statisticalsoftware (SAS USA) Litter was used as a covariate witheach litter considered a single biological replicate Data arepresented as means plusmn SEM unless otherwise stated
3 Results
31MaternalWeights and Body FatMass Apre-conceptionalHF diet (MHF) resulted in a significant increase in femalebody weight (chow 270 plusmn 5 g MHF 310 plusmn 8 119875 lt 005)and total fat mass compared to chow fed animals (chow178 plusmn 07 MHF 277 plusmn 17 119875 lt 00001) At the startof pregnancy there were no differences in body weightsbetween females randomly assigned to CONT and PLHFgroups (CONT 270 plusmn 8 g PLHF 269 plusmn 7) Total caloricintake was similar between CONT and MHF groups butwas significantly increased in PLHF dams during the first10 days of pregnancy (average caloric intake CONT 023 plusmn0004 kcalgBWday MHF 023 plusmn 0004 PLHF 032 plusmn 001119875 lt 005 for PLHF versus CONT and MHF) thereaftercaloric intake was similar to that of CONT and MHF dams
32 BodyWeight and Body Composition inMale Offspring Asreported previously [14] MHF and PLHF pups had slightlybut statistically significantly reduced body weights and werehypoinsulinemic and hypoleptinemic compared to CONToffspring at birth MHF and PLHF neonates displayed rapidcatch-up growth and were heavier than CONT neonates atthe time of weaning (day 22 CONT 616 plusmn 07 g MHF 705 plusmn09 PLHF 642 plusmn 12)
Final adult body weights were increased in MHF andPLHF male offspring compared to CONT (CONT 562 plusmn 9 gMHF 635 plusmn 18 PLHF 628 plusmn 17) [14] MHF and PLHF adultmale offspring had significantly increased total body fat massat day 150 compared to CONT (Figure 1) There were nodifferences in total fat mass between MHF and PLHF adultoffspring
33 Plasma Analyses MHF and PLHF adult male offspringwere hyperleptinemic and hyperinsulinemic compared toCONT (Figures 2(a) and 2(b)) Fasting blood glucose levelswere similar between groups (CONT 56 plusmn 02mmolLMHF57 plusmn 03 PLHF 57 plusmn 02) Glucose insulin ratios weresignificantly (119875 lt 0001) reduced in MHF and PLHF maleoffspring compared to control (CONT 25 plusmn 02 MHF 13 plusmn01 PLHF 12 plusmn 01) and offspring exhibited increased insulinresistance using HOMA analysis (Figure 2(c)) Plasma TNF-120572 levels were not significantly different between the groups(CONT 36 plusmn 09 pgmL MHF 67 plusmn 19 PLHF 51 plusmn 19)Plasma IL-1120573 levels were significantly increased inMHF (119875 lt005) offspring compared to control (CONT 319plusmn 07 pgmLMHF 383 plusmn 28 PLHF 361 plusmn 22) but were not differentbetween CONT and PLHF offspring
34 Gene Expression MHF offspring showed significantlyhigher levels of pancreatic mRNA levels of SOCS3 ObRband IRS1 compared to CONT and PLHF groups (Figure 3)mRNA levels of these genes were similar between CONT andPLHF offspring Despite these similarities in expression levelsbetween CONT and PLHF offspring significant positivecorrelations that existed between these and other measuredgenes were primarily found in CONT and MHF offspringonly and were not present in PLHF offspring (Table 2)
4 Journal of Nutrition and Metabolism
CONT MHF PLHF
50
40
30
20
10
0
lowast
lowast
Tota
l bod
y fa
t (
)
Figure 1 Total body fat mass (percent) as quantified by DEXA scan in adult CONT MHF and PLHF offspring Data are means plusmn SEM 119899 =10ndash12 per group lowast119875 lt 005
CONT MHF PLHF0
5
10
15
20
25lowast
lowast
Lept
in (n
gm
L)
(a)
CONT MHF PLHF0
1
2
3
4
5
6lowast
lowast
Insu
lin (n
gm
L)
(b)
CONT MHF PLHF0
05
1
15 lowast
lowast
HO
MA
IR
(c)
Figure 2 Fasting plasma leptin (a) insulin (b) and homeostatic model assessment (HOMA) to quantify insulin resistance and beta-cellfunction (c) in adult CONT MHF and PLHF offspring Data are means plusmn SEM 119899 = 10ndash18 per group lowast119875 lt 005
Levels of mRNA of IRS2 PDX-1 and Kir62 were sig-nificantly reduced in PLHF offspring compared to MHF off-spring but were not different between any other groups Pan-creatic INS1 and INS2 expression was significantly increasedin MHF and PLHF offspring compared to CONT (Figure 3)
STAT3 expression was significantly increased inPLHF offspring compared to CONT and MHF offspring(Figure 4(a)) PI3K expression was significantly decreased inPLHF compared to CONT and MHF (Figure 4(b)) Of notePI3K was positively correlated with IRS1 and IRS2 expressionin PLHF offspring (1198772 = 044 119875 lt 005 and 1198772 = 072119875 lt 00005 resp) but not in CONT or MHF offspring
Insulin receptor leptin ObRa and phosphodiesterase 3B(PDE3B) were similar between groups (data not shown) Ahighly significant relationship was observed in all offspringgroups between PDX-1 and ATP-sensitive potassium (Kir62)channel (Figure 5) Given the potential link between SOCS3and inflammation mRNA levels of key proinflammatorycytokines were also evaluated There was an increase inTNF-120572 in PLHF offspring compared to CONT and MHF
offspring (Figure 6(a)) The macrophage marker CD68 wasincreased in MHF and PLHF offspring compared to CONT(Figure 6(b))Therewas an increase in IL-1R1mRNA levels inMHF offspring compared to CONT offspring (Figure 6(c))There were no differences in IL-6R cardiotropin IL-1BGLUT2 GP130 CTNF LIF or TNF-R1 mRNA expressionbetween any of the offspring groups (data not shown)
4 Discussion
There is clear clinical and experimental evidence that mater-nal obesity predisposes to obesity and metabolic disorders inoffspring [14 30] However despite the common phenotypeof offspring obesity and hyperinsulinemia the present studyshows that at least in an animal model that maternal nutri-tional history determines the pattern of mRNA expressionof key regulatory genes in the pancreas of adult offspringThese data suggest that different regulatory pathways maylead to a similar metabolic phenotype and are in agreementwith previous experimental studies suggesting that there
Journal of Nutrition and Metabolism 5
SOCS3
0
100
200
300
400
500
600
700
800
( o
f con
trol)
lowast lowast
CONT MHF PLHF
(a)
ObRb
0
500
1000
1500
2000
2500 lowast lowast
( o
f con
trol)
CONT MHF PLHF
(b)
IRS1
0
50
100
150
200
250
300 lowastlowast
( o
f con
trol)
CONT MHF PLHF
(c)
IRS2
0
50
100
150
200lowast
( o
f con
trol)
CONT MHF PLHF
(d)
PDX-1
0
50
100
150
200
250 lowast
( o
f con
trol)
CONT MHF PLHF
(e)
0
50
100
150
200
250
300 lowast
K+ channel
CONT MHF PLHF
( o
f con
trol)
(f)
Ins1
0
50
100
150
200
250
300 lowast
lowast
CONT MHF PLHF
( o
f con
trol)
(g)
Ins2
CONT MHF PLHF0
50100150200250300350400 lowast
lowast
( o
f con
trol)
(h)
Figure 3 Pancreatic mRNA expression in adult CONT MHF and PLHF offspring SOCS3 suppressor of cytokine signaling 3 IRS insulinreceptor substrate PDX1 pancreatic and duodenal homeobox 1 ObRb full length leptin receptor Ins insulin Kir62 potassium channelsubunit Data are means plusmn SEM 119899 = 10ndash18 per group lowast119875 lt 005
6 Journal of Nutrition and Metabolism
Table 2 Correlations between SOCS3 and other key genes N = 8ndash13 per group
CONT MHF PLHFSOCS3 versus ObRb 119877
2 = 069 P = 008 R2 = 087 P = 00008 1198772 = 003 P = 064
SOCS3 versus IR R2 = 067 P = 002 1198772 = 002 P = 071 119877
2 = 0002 P = 090SOCS3 versus IRS1 R2 = 084 P = 001 R2 = 058 P = 002 119877
2 = 004 P = 051SOCS3 versus IRS2 R2 = 098 P = 0002 R2 = 058 P = 002 119877
2 = 015 P = 021SOCS3 versus Kir62 channel R2 = 082 P = 003 R2 = 093 P lt 00001 119877
2 = 0002 P = 090SOCS3 versus Pdx1 R2 = 078 P = 0009 R2 = 091 P = 00003 119877
2 = 007 P = 040
CONT MHF PLHF0
100
200
300
400
500
STAT
3 (
of c
ontro
l)
lowast
lowast
(a)
PI3K
( o
f con
trol)
CONT MHF PLHF0
50
100
150lowast
lowast
(b)
Figure 4 Pancreatic STAT3 (a) and PI3K mRNA expression (b) Data are means plusmn SEM 119899 = 10ndash18 per group STAT3 signal transducer andactivator of transcription 3 PI3K phosphatidylinositol 3-kinase lowast119875 lt 005
may be separate influences of maternal obesity during thepericonceptional period and late gestation on adiposity inoffspring [31] We have previously shown that ldquoprogrammedrdquoobesity and simple diet-induced obesity can develop viadifferent underlying mechanisms [32] The present studyfurther reinforces the importance of the timing of the dietaryinsult on the metabolic derangements that ensue in offspringduring adulthood
SOCS3 inhibition of ObRb signaling has been proposedas a mechanism for leptin resistance in a range of tissues [33ndash35] including pancreatic 120573 cells [36] It is clear that leptinresistance in PLHF pancreata was mediated by mechanismsthat did not involve SOCS3 and this has been reported previ-ously in rat models of HF feeding [37 38] SOCS3 is normallyincreased at sites of both acute and chronic inflammation [39]and has been shown to specifically suppress STAT3 activation[40] This agrees with the present study a marked increasein SOCS3 mRNA levels in MHF offspring is paralleled bySTAT3 levels similar to control offspring Leptin is knownto increase SOCS3 expression in isolated human pancreaticislets and in islets from obob mice treated in vivo [36] ourdata imply that leptin-induced SOCS3 stimulation appearsintact in MHF offspring Conversely in PLHF offspring
SOCS3 expression levels were similar to controls in thepresence of markedly elevated STAT3 expression Of notedespite similar expression levels of SOCS3 between CONTand PLHF offspring the positive correlations that wereobserved between SOCS3 and other key signaling genes wererestricted mainly to CONT and MHF offspring only Thismay point to a maladaptation in PLHF offspring due to theacute HF dietary exposure and a change in homeostatic set-points in MHF offspring due to mothers entering pregnancyin an already obese state Despite the link between SOCS3and inflammation there were no observed changes in TNF-120572 mRNA levels in the MHF group whilst there was asignificant increase in TNF-120572 expression in PLHF offspringIn addition mRNA levels of the macrophage marker CD68were increased in both HF groups and taken togethermay indicate differing macrophage phenotypespolarizationstates resulting in divergent offspring inflammatory profilesdependent upon prior maternal nutritional background Ofnote despite marked changes in inflammatorymarkers at thetissue level we observed no changes in circulating plasmaTNF-120572 concentrationHowever despite enhanced plasma IL-1120573 concentrations pancreatic IL-1120573 expression was unaltered
Journal of Nutrition and Metabolism 7
CONT
0 250 500 7500
250
500
750
PDX-1
1199032= 097
K+ch
anne
l
(a)
MHF
0 250 500 750 10000
500
1000
15001199032= 094
PDX-1
K+ch
anne
l
(b)
PLHF
0 50 100 150 200 250 3000
50
100
150
200
250
3001199032= 078
PDX-1
K+ch
anne
l
(c)
Figure 5 Relationship between PDX-1 and potassium channel subunit Kir62 in adult CONT MHF and PLHF male offspring PDX-1pancreatic and duodenal homeobox 1119873 = 8ndash10 per group
between groups implicating other tissues as possible systemicsources of this proinflammatory cytokine
PLHF offspring exhibited decreased PI3K mRNA levelscompared to CONT animals Induction of the ObRbPI3Ksignaling pathway prolongs potassium channel openingleading to decreased insulin exocytosis a direct action ofleptin which inhibits insulin secretion [41] Moreover thelowered PI3K mRNA levels in PLHF offspring were con-comitant with a reduction in IRS1 and IRS2 mRNA whosephosphorylation is coupled to PI3K activation Of notedownstreamof PI3K PDE3B gene expression levels remainedsimilar to controls suggesting that altered leptin signalingwas achieved in earlier stages of this pathway Thus attenua-tion of the ObRbPI3K signaling pathwaymaymediate leptinresistance in these hyperleptinemic PLHF offspring Sucha mechanism has been observed before in hypothalamicneurons of rats with diet-induced obesity (DIO) [42]
IRS proteins and PI3K have leptin independent rolesin the 120573 cell Both are found downstream of the insulinreceptor in the IRIRSPI3K pathway which mediates theautocrine actions of the insulin-IR complex [43 44] Ourobserved attenuation of PI3K and IRS mRNA did nothowever affect insulin synthesismdashPLHF animals were hyper-insulinemic and preproinsulin mRNA concentrations wereelevated Furthermore mRNA concentrations of IR wereunaffected potentially due to lack of SOCS3 induction inPLHF offspring similar to Controls Thus these mRNAchanges of PI3K coupled to IRS likely reflect a mechanismof leptin resistance in this group
Mean concentrations of ObRb mRNA in PLHF offspringwere similar to controls but a significant inverse relationshipbetween plasma leptin and ObRb mRNA levels in the PLHFgroup was suggestive of ObRb downregulation A downregu-lation of receptor populations in the face of elevated ligand
8 Journal of Nutrition and Metabolism
CONT MHF PLHF0
50
100
150
200
TNF-120572
mRN
A (
of c
ontro
l)
lowast
lowast
(a)
CD68
mRN
A (
of c
ontro
l)
CONT MHF PLHF0
50
100
150
200
lowast
lowast
(b)
IL1-
R1 m
RNA
( o
f con
trol)
CONT MHF PLHF0
50
100
150
200lowast
(c)
Figure 6 Pancreatic mRNA expression of TNF-120572 (a) CD68 (b) and IL1-R1 (c) in adult CONT MHF and PLHF offspring TNF-120572 tumornecrosis factor-alpha CD68 cluster of differentiation 68 IL1-R1 interleukin 1 receptor type 1 Data are means plusmn SEM 119899 = 10ndash18 per grouplowast
119875 lt 005
concentrations is not uncommon in resistant states It isseen for example in the hypothalamus in the presence ofhyperleptinemia [45ndash47] and during pregnancy which is anatural state of leptin resistance [48]
A rise in IRS1 and IRS2 mRNA associated with elevatedObRb and SOCS3 mRNA in MHF offspring may seemunexpected since SOCS3 is known to target insulin receptorsignaling by interference with IRS action [49] However sev-eral studies have demonstrated that SOCS3 mediates insulinresistance in both adipocytes and hepatocytes through post-translational modifications such as inhibition of IRS tyro-sine phosphorylation [50] and targeted ubiquitin-mediateddegradation of IRS1 and IRS2 without effecting transcription[49 51] IRS proteins are involved in both leptin signalingand insulin signaling so altered gene expression levels mayrepresent changes in either (or both) pathways although IRSresponse to leptin is reportedly much lower than that seenwith insulin [52] We observed an increase in IRS1 expressionin MHF offspring compared to Controls This upregulationof IRS1 expression may be related to the increased insulin
production of the MHF offspring given the known role ofIRS1 in the regulation of insulin secretion [53]
SOCS3 is also reported to repress preproinsulin genetranscription [41] although other mechanisms clearly coun-tered this action inMHFoffspring since Ins1 and Ins2mRNAexpression levels remained elevated This observation wasconfirmed by the significantly raised levels of circulatingplasma insulin and the lack of any correlation in levels ofmRNA between SOCS3 and Ins1 or Ins2
Some of the observed differences may relate to theinitial hyperphagia observed in the PLHF maternal groupas reported previously [14] Since rodents primarily eat ona calorific basis they will passively over-consume calorieswhen initially placed on an HF diet and self-regulate backto a normal total caloric intake over a period of a few daysIt has been shown in other rodent models that alterations innutrition around the time of implantation can lead to lastingmetabolic effects in the offspring [54] Maternal obesity hasbeen shown to exert adverse effects as early as the oocyteand preimplantation embryo stage and that these effects may
Journal of Nutrition and Metabolism 9
contribute to the lasting morbidity observed in offspring[55] Similarly even a very short period of altered nutrition(restricted to one ovulatory cycle) prior to natural mating canelicit lasting effects on the offspring [56] and this representsa further point of difference between the MHF and PLHFgroups
The present study was not designed to incorporate theeffect of a reversal paradigm whereby mothers fed thepre-conceptional HF diet were placed on a standard dietthroughout pregnancy and lactation However while there isevidence that a period of dietary restriction in obese mothersmay ablate the programming of obesity in offspring [57]it may be associated with an activation of the stress axisin the offspring [31] Further the data are inconsistent ithas also been shown that dams with diet-induced obesityfed a standard diet throughout pregnancy and lactation stilltransmit the propensity for obesity and metabolic disease totheir offspring [58]
We recognize that gene expression analysis on wholepancreatic tissue is a limitation of the present study Dueto the size of the cohort and logistical considerations onlywhole pancreatic tissue was a possibility whereas ideallyonly pure islet populations would have been examined [59]However for the key genes presented such as ObRb SOCS3and IRS expression has been shown to be primarily confinedto the endocrine islets and not been detected in the exocrinepancreas [39 41 60] therefore we are confident that theobserved changes in the genes studied represent changes inthe islets Further although we have shown that differencesin pancreatic gene expression in offspring are dependentupon the timing of maternal high fat diet exposure this doesnot itself imply causation in the development of the obesephenotypemdashother relevant pathways including liver skeletalmuscle and fat metabolism will also be integrated into theprogrammed phenotype development
This study has demonstrated enduring deleterious effectsof a maternal HF diet during early windows of develop-ment Maternal HF feeding both before conception andorthrough pregnancy and lactation induced obesity hyper-insulinemia and hyperleptinemia in adult offspring eventhough offspring were fed a standard diet postnatally Timingof exposure to the maternal HF diet differentially alteredgene expression in signaling pathways downstream of thepancreatic leptin receptor that mediated leptin resistanceThis also raises the possibility that despite common offspringobesity treatment modalities may only be efficacious in asubpopulation of treated offspring as the current data suggestthat the metabolic pathways disrupted will be dependentupon the timing of the early life dietary insult
Abbreviations
CD68 Cluster of differentiation 68CONT ControlDEXA Dual energy X-ray absorptiometryELISA Enzyme-linked immunosorbent assayGLUT2 Glucose transporter 2GP130 Glycoprotein 130HF High fat
HOMA Homeostatic model assessmentHPRT HypoxanthinephophoribosyltransferaseIL InterleukinIns InsulinIR Insulin resistanceIRS Insulin receptor substrateKir62 Potassium channel subunitLIF Leukemia inhibitory factorMHF Maternal high fatOBR Leptin receptorPDE3B Phosphodiesterase 3BPDX-1 Pancreatic and duodenal homeobox-1PI3K Phosphatidylinositide 3-kinasePLHF Pregnancy and lactation high fatSOCS3 Suppressor of cytokine signaling 3STAT3 Signal transducer and activator of
transcription 3TNF-120572 Tumor necrosis factor-alpha
Conflict of Interests
The authors have no conflict of interests with any financialorganization regarding the material discussed in this paper
Author Contributions
Mark H Vickers developed the animal model designed thestudy completed the data analysis and wrote the first draftof the paper Graham J Howie maintained the experimentalcohort performed the gene expression analysis and con-tributed to the original draft paper Clare M Reynolds per-formed the inflammatory related assays and gene expressionwork Deborah M Sloboda provided input into data analysisand interpretation
Acknowledgments
The authors acknowledge the technical support of TaniaKamal with endocrine assays The authors acknowledgefunding support from the Health Research Council of NewZealand and Gravida National Centre for Growth andDevelopment
References
[1] M M Hillemeier C S Weisman C Chuang D S Downs JMcCall-Hosenfeld and F Camacho ldquoTransition to overweightor obesity among women of reproductive agerdquo Journal ofWomenrsquos Health vol 20 no 5 pp 703ndash710 2011
[2] J E Norman and RM Reynolds ldquoThe consequences of obesityand excess weight gain in pregnancyrdquo Proceedings of the Nutri-tion Society vol 70 no 4 pp 450ndash456 2011
[3] H A Abenhaim R A Kinch L Morin A Benjamin and RUsher ldquoEffect of prepregnancy body mass index categories onobstetrical and neonatal outcomesrdquo Archives of Gynecology andObstetrics vol 275 no 1 pp 39ndash43 2007
[4] S DHMalnick andH Knobler ldquoThemedical complications ofobesityrdquo Oxford Journals Medicine vol 99 no 9 pp 565ndash5792006
10 Journal of Nutrition and Metabolism
[5] P M Catalano ldquoManagement of obesity in pregnancyrdquo Obstet-rics and Gynecology vol 109 no 2 pp 419ndash433 2007
[6] L Poston ldquoGestational weight gain influences on the long-termhealth of the childrdquo Current Opinion in Clinical Nutrition andMetabolic Care vol 15 pp 252ndash257 2012
[7] H Hochner Y Friedlander R Calderon-Margalit et al ldquoAsso-ciations of maternal prepregnancy body mass index and gesta-tional weight gain with adult offspring cardiometabolic riskfactors the Jerusalem perinatal family follow-up studyrdquo Circu-lation vol 125 no 11 pp 1381ndash1389 2012
[8] S J Spencer ldquoEarly life programming of obesity the impactof the perinatal environment on the development of obesityand metabolic dysfunction in the offspringrdquo Current DiabetesReviews vol 8 no 1 pp 55ndash68 2012
[9] H Chen D Simar K Lambert J Mercier and M J MorrisldquoMaternal and postnatal overnutrition differentially impactappetite regulators and fuel metabolismrdquo Endocrinology vol149 no 11 pp 5348ndash5356 2008
[10] P M Catalano ldquoObesity and pregnancymdashthe propagation of aviscous cyclerdquo Journal of Clinical Endocrinology and Metabo-lism vol 88 no 8 pp 3505ndash3506 2003
[11] M W Gillman S L Rifas-Shiman K Kleinman E Oken JW Rich-Edwards and EM Taveras ldquoDevelopmental origins ofchildhood overweight potential public health impactrdquo Obesityvol 16 no 7 pp 1651ndash1656 2008
[12] E Oken EM Taveras K P Kleinman JW Rich-Edwards andM W Gillman ldquoGestational weight gain and child adiposity atage 3 yearsrdquoTheAmerican Journal of Obstetrics and Gynecologyvol 196 no 4 pp 322e1ndash322e8 2007
[13] H M Ehrenberg B M Mercer and P M Catalano ldquoThe influ-ence of obesity and diabetes on the prevalence of macrosomiardquoThe American Journal of Obstetrics and Gynecology vol 191 no3 pp 964ndash968 2004
[14] G J Howie D M Sloboda T Kamal and M H VickersldquoMaternal nutritional history predicts obesity in adult offspringindependent of postnatal dietrdquo Journal of Physiology vol 587no 4 pp 905ndash915 2009
[15] D M Sloboda G J Howie A Pleasants P D Gluckman andM H Vickers ldquoPre- and postnatal nutritional histories influ-ence reproductive maturation and ovarian function in the ratrdquoPLoS ONE vol 4 no 8 Article ID e6744 2009
[16] M C Battista M F Hivert K Duval and J P BaillargeonldquoIntergenerational cycle of obesity and diabetes how can wereduce the burdens of these conditions on the health of futuregenerationsrdquo Experimental Diabetes Research vol 2011 ArticleID 596060 19 pages 2011
[17] T J Kieffer and J F Habener ldquoThe adipoinsular axis effects ofleptin on pancreatic 120573-cellsrdquo The American Journal of Physiol-ogy vol 278 no 1 pp E1ndashE14 2000
[18] S L Gray C Donald A Jetha S D Covey and T J KiefferldquoHyperinsulinemia precedes insulin resistance in mice lackingpancreatic 120573-cell leptin signalingrdquo Endocrinology vol 151 no 9pp 4178ndash4186 2010
[19] S D Covey R DWideman CMcDonald et al ldquoThe pancreat-ic 120573 cell is a key site formediating the effects of leptin on glucosehomeostasisrdquo Cell Metabolism vol 4 no 4 pp 291ndash302 2006
[20] K J Dudley DM Sloboda K L Connor J Beltrand andMHVickers ldquoOffspring ofmothers fed a high fat diet display hepaticcell cycle inhibition and associated changes in gene expressionand DNA methylationrdquo PLoS ONE vol 6 no 7 Article IDe21662 2011
[21] K L Connor M H Vickers J Beltrand M J Meaney and DM Sloboda ldquoNature nurture or nutrition Impact of maternalnutrition on maternal care offspring development and repro-ductive functionrdquo Journal of Physiology vol 590 part 9 pp2167ndash2180 2012
[22] J Beltrand D M Sloboda K L Connor M Truong and MH Vickers ldquoThe effect of neonatal leptin antagonism in malerat offspring is dependent upon the interaction between priormaternal nutritional status and post-weaning dietrdquo Journal ofNutrition andMetabolism vol 2012 Article ID 296935 10 pages2012
[23] K L Connor M H Vickers C Cupido E Sirimanne and DM Sloboda ldquoMaternal high fat diet during critical windowsof development alters adrenal cortical and medullary enzymeexpression in adult male rat offspringrdquo Journal of the Develop-mental Origins of Health and Disease vol 1 no 4 pp 245ndash2542010
[24] P J Mark C Sisala K Connor et al ldquoA maternal high-fat dietin rat pregnancy reduces growth of the fetus and the placen-tal junctional zone but not placental labyrinth zone growthrdquoJournal of the Developmental Origins of Health and Disease vol2 no 1 pp 63ndash70 2011
[25] D R Matthews J P Hosker A S Rudenski B A Naylor DF Treacher and R C Turner ldquoHomeostasis model assessmentinsulin resistance and 120573-cell function from fasting plasmaglucose and insulin concentrations in manrdquo Diabetologia vol28 no 7 pp 412ndash419 1985
[26] M P Hermans J C Levy R J Morris and R C TurnerldquoComparison of tests of120573-cell function across a range of glucosetolerance from normal to diabetesrdquo Diabetes vol 48 no 9 pp1779ndash1786 1999
[27] A B Bernal M H Vickers M B Hampton R A Poynton andD M Sloboda ldquoMaternal undernutrition significantly impactsovarian follicle number and increases ovarian oxidative stress inadult rat offspringrdquo PLoS ONE vol 5 no 12 Article ID e155582010
[28] J Vandesompele K de Preter F Pattyn et al ldquoAccurate nor-malization of real-time quantitative RT-PCR data by geometricaveraging of multiple internal control genesrdquo Genome biologyvol 3 no 7 Article ID RESEARCH0034 2002
[29] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the 2-ΔΔCT methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
[30] J Armitage L Poston and P Taylor ldquoDevelopmental originsof obesity and the metabolic syndrome the role of maternalobesityrdquo Frontiers of Hormone Research vol 36 pp 73ndash84 2007
[31] S Zhang L Rattanatray J L Morrison L M Nicholas S Lieand I C McMillen ldquoMaternal obesity and the early originsof childhood obesity weighing up the benefits and costs ofmaternal weight loss in the periconceptional period for theoffspringrdquo Experimental Diabetes Research vol 2011 Article ID585749 10 pages 2011
[32] N M Thompson A M Norman S S Donkin et al ldquoPrenataland postnatal pathways to obesity different underlying mech-anisms different metabolic outcomesrdquo Endocrinology vol 148no 5 pp 2345ndash2354 2007
[33] T Fuentes I Ara A Guadalupe-Grau et al ldquoLeptin receptor170 kDa (OB-R170) protein expression is reduced in obesehuman skeletal muscle a potential mechanism of leptin resis-tancerdquo Experimental Physiology vol 95 no 1 pp 160ndash171 2010
[34] Z Wang Y T Zhou T Kakuma et al ldquoLeptin resistance ofadipocytes in obesity role of suppressors of cytokine signalingrdquo
Journal of Nutrition and Metabolism 11
Biochemical and Biophysical Research Communications vol 277no 1 pp 20ndash26 2000
[35] C Bjoslashrbaeligk K El-Haschimi J D Frantz and J S Flier ldquoThe roleof SOCS-3 in leptin signaling and leptin resistancerdquoThe Journalof Biological Chemistry vol 274 no 42 pp 30059ndash30065 1999
[36] K Laubner T J Kieffer N T Lam X Niu F Jakob and JSeufert ldquoInhibition of preproinsulin gene expression by leptininduction of suppressor of cytokine signaling 3 in pancreatic 120573-cellsrdquo Diabetes vol 54 no 12 pp 3410ndash3417 2005
[37] C Peiser G P McGregor and R E Lang ldquoLeptin receptorexpression and suppressor of cytokine signaling transcriptlevels in high-fat-fed ratsrdquo Life Sciences vol 67 no 24 pp 2971ndash2981 2000
[38] G R Steinberg A C Smith S Wormald P Malenfant CCollier and D J Dyck ldquoEndurance training partially reversesdietary-induced leptin resistance in rodent skeletal musclerdquoTheAmerican Journal of Physiology vol 286 no 1 pp E57ndashE632004
[39] G E White A Cotterill M R Addley E J Soilleux and DR Greaves ldquoSuppressor of cytokine signalling protein SOCS3expression is increased at sites of acute and chronic inflamma-tionrdquo Journal of Molecular Histology vol 42 no 2 pp 137ndash1512011
[40] H YasukawaMOhishiHMori et al ldquoIL-6 induces an anti-in-flammatory response in the absence of SOCS3 inmacrophagesrdquoNature Immunology vol 4 no 6 pp 551ndash556 2003
[41] J Seufert ldquoLeptin effects on pancreatic 120573-cell gene expressionand functionrdquo Diabetes vol 53 no 1 pp S152ndashS158 2004
[42] A S Metlakunta M Sahu and A Sahu ldquoHypothalamicphosphatidylinositol 3-kinase pathway of leptin signaling isimpaired during the development of diet-induced obesity inFVBNmicerdquo Endocrinology vol 149 no 3 pp 1121ndash1128 2008
[43] C A Aspinwall W J Qian M G Roper R N Kulkarni C RKahn and R T Kennedy ldquoRoles of insulin receptor substrate-1phosphatidylinositol 3-kinase and release of intracellular Ca2+stores in insulin-stimulated insulin secretion in 120573-cellsrdquo TheJournal of Biological Chemistry vol 275 no 29 pp 22331ndash223382000
[44] I B Leibiger B Leibiger T Moede and P O BerggrenldquoExocytosis of insulin promotes insulin gene transcription viathe insulin receptorPI-3 kinasep70 s6 kinase and CaM kinasepathwaysrdquoMolecular Cell vol 1 no 6 pp 933ndash938 1998
[45] R L Martin E Perez Y J He R Dawson Jr andW J MillardldquoLeptin resistance is associated with hypothalamic leptin recep-tor mRNA and protein downregulationrdquo Metabolism vol 49no 11 pp 1479ndash1484 2000
[46] J Wilsey and P J Scarpace ldquoCaloric restriction reverses thedeficits in leptin receptor protein and leptin signaling capacityassociated with diet-induced obesity role of leptin in the reg-ulation of hypothalamic long-form leptin receptor expressionrdquoJournal of Endocrinology vol 181 no 2 pp 297ndash306 2004
[47] Y Zhang and P J Scarpace ldquoThe role of leptin in leptin resist-ance and obesityrdquo Physiology and Behavior vol 88 no 3 pp249ndash256 2006
[48] D R Grattan S R Ladyman and R A Augustine ldquoHormonalinduction of leptin resistance during pregnancyrdquoPhysiology andBehavior vol 91 no 4 pp 366ndash374 2007
[49] L RuiM YuanD Frantz S Shoelson andM FWhite ldquoSOCS-1 and SOCS-3 block insulin signaling by ubiquitin-mediateddegradation of IRS1 and IRS2rdquoThe Journal of Biological Chem-istry vol 277 no 44 pp 42394ndash42398 2002
[50] K Ueki T Kondo and C R Kahn ldquoSuppressor of cytokine sig-naling 1 (SOCS-1) and SOCS-3 cause insulin resistance throughinhibition of tyrosine phosphorylation of insulin receptor sub-strate proteins by discrete mechanismsrdquoMolecular and CellularBiology vol 24 no 12 pp 5434ndash5446 2004
[51] S B Jorgensen O rsquoNeill HM L Sylow et al ldquoDeletion of skel-etal muscle SOCS3 prevents insulin resistance in obesityrdquo Dia-betes vol 62 no 1 pp 56ndash64 2013
[52] W Becker ldquoLeptin signal transductionrdquo in Leptin and LeptinAntagonists A Gertler Ed pp 1ndash9 Landes Bioscience AustinTex USA 2009
[53] R N Kulkarni J N Winnay M Daniels et al ldquoAltered func-tion of insulin receptor substrate 1-deficient mouse islets andcultured 120573-cell linesrdquo Journal of Clinical Investigation vol 104no 12 pp R69ndashR75 1999
[54] W Y Kwong A E Wild P Roberts A C Willis and T PFleming ldquoMaternal undernutrition during the preimplantationperiod of rat development causes blastocyst abnormalities andprogramming of postnatal hypertensionrdquoDevelopment vol 127no 19 pp 4195ndash4202 2000
[55] E S Jungheim E L Schoeller K L Marquard E D LoudenJ E Schaffer and K H Moley ldquoDiet-induced obesity modelabnormal oocytes and persistent growth abnormalities in theoffspringrdquo Endocrinology vol 151 no 8 pp 4039ndash4046 2010
[56] A J Watkins A Wilkins C Cunningham et al ldquoLow proteindiet fed exclusively during mouse oocyte maturation leadsto behavioural and cardiovascular abnormalities in offspringrdquoJournal of Physiology vol 586 no 8 pp 2231ndash2244 2008
[57] E Zambrano P M Martınez-Samayoa G L Rodrıguez-Gonzalez and P W Nathanielsz ldquoDietary intervention prior topregnancy reverses metabolic programming in male offspringof obese ratsrdquo Journal of Physiology vol 588 no 10 pp 1791ndash1799 2010
[58] M S Patel andM Srinivasan ldquoMetabolic programming causesand consequencesrdquoThe Journal of Biological Chemistry vol 277no 3 pp 1629ndash1632 2002
[59] R A Simmons ldquoRole of metabolic programming in the patho-genesis of 120573-cell failure in postnatal liferdquo Reviews in Endocrineand Metabolic Disorders vol 8 no 2 pp 95ndash104 2007
[60] I B Leibiger B Leibiger and P O Berggren ldquoInsulin signalingin the pancreatic 120573-cellrdquoAnnual Review of Nutrition vol 28 pp233ndash251 2008

2 Journal of Nutrition and Metabolism
on subsequent health and well-being of offspring We havepreviously shown that obesity in adult rat offspring arising asa consequence of maternal high fat (HF) nutrition was inde-pendent of the pre-conceptional maternal diet [14] We haveshown that offspring of mothers fed an HF diet restricted topregnancy and lactation compared to those born to mothersfed a HF diet prior to conception and throughout pregnancyand lactation both exhibit a remarkably similar obesogenicphenotype characterised by increased fat mass hyperin-sulinemia and hyperleptinemia These data suggested thatdespite a common obesity phenotype differential mecha-nisms based on the timing of maternal exposure to HF nutri-tion may underpin pancreatic adipoinsular axis dysfunctionin these offspring [17ndash19]We therefore investigated key com-ponents of the leptin-insulin (adipoinsular) and inflamma-tory signaling pathways in pancreatic tissue of offspring bornto mothers fed a high fat diet during pregnancy in the pres-ence or absence of pre-conceptional HF diet-induced obesity
2 Methods
The rat model of maternal high fat nutrition has beendescribed by our group previously [14 15 20ndash24] In briefmale and female Wistar rats were acquired at a weaning age(22 days Vernon Jansen Unit University of Auckland) andhoused two per cage under standard conditions with a 1212light dark cycle and free access to water Females were weight-matched and assigned to receive either standard rat chow(119899 = 16 5 fat Diet 2018 Harlan Teklad Oxon) or a highfat (HF) diet (119899 = 8 24 fat D12451 Research Diets NJUSA) to be fed ad libitum for the duration of the trial Maleswere fed standard chow ad libitum for the duration of thepremating period Body weights were recorded every 3 daysuntil postnatal day 120
At postnatal day 110 body composition in females wasquantified using dual energy X-ray absorptiometry (DEXALunar Prodigy GE Medical Systems Madison WI USA) Atpostnatal day 120 females were time-mated using an estruscycle monitor (Model EC40 Fine Science Tools CA USA)Upon confirmation of mating three maternal dietary groupswere established (1) controls (CONT) females fed a standardchow diet throughout their life andmaintained on a standardchow diet throughout pregnancy and lactation (2) maternalhigh fat diet (MHF) females fed an HF diet throughout theirlife and maintained on the HF diet throughout pregnancyand lactation and (3) pregnancy + lactation HF diet (PLHF)females fed a standard chow diet until conception and thenan HF diet throughout pregnancy and lactation All pregnantdams were weighed and had food intakes measured dailythroughout pregnancy Following birth pup weights andbody lengths were recorded and litter size was randomlyadjusted to 8 pups per litter to ensure standardized nutritionuntil weaning Body weights and food intakes of dams weremeasured throughout the lactation period andoffspringwereweighed every three days until weaning
At weaning (d22) offspring were housed 2 per cage (2per littermaternal background) and fed a chow diet until theend of the trial At postnatal day 150 body composition wasmeasured in male offspring (119899 = 10ndash18 per group) by DEXA
At postnatal day 160 animals were fasted overnight and killedby decapitation following anaesthesia with sodium pentobar-bitone (60mgkg IP) Blood was collected into heparinizedtubes centrifuged and plasma supernatant stored for futureanalysis Whole pancreata were removed and immediatelysnap-frozen in liquid nitrogen for later molecular analysisAll animal experiments were approved under guidelines ofthe Animal Ethics Committee at the University of Auckland(Approval R652)
21 Plasma Analysis Fasting plasma leptin and insulin con-centrations were analysed using commercial rat-specificELISAs (CrystalChem 90040 and 90060 resp DownersGrove IL USA) Fasting whole blood glucose concentrationswere measured using a glucose meter at the time of tissuecollection (d160) (Roche AccuChek) Plasma tumor necrosisfactor (TNF-) alpha interleukin (IL-) 1120573 and IL-6 weremeasured using commercially available ELISAs (Quantikinekits RTA00 RLB00 and R6000B resp RampD SystemsEurope Abingdon UK) The homeostatic model assessment(HOMA)method was used as a measure of insulin resistance(IR) and beta-cell function (HOMA-IR = plasma glucose timesinsulin225) [25 26]
22 Gene Expression Quantitative PCR methods have beendescribed previously in detail [23 27] Briefly whole pancreastissue was disrupted using a homogenizer and total RNAwasextracted using commercially available kits (Qiagen Mini-Prep catalogue number 80204) according to the manufac-turerrsquos instructions RNA integrity and quality were assessedusing a NanoDrop spectrophotometer (ND-1000 BioLabLtd) using NanoDrop software (version 312) where sampleratio A260280 and A260230 were sim20 and intact bandswere visualised by gel electrophoresis Complimentary (c)DNA was synthesised from RNA (5 120583g) by reverse transcrip-tion PCR using MMLV-RT (Invitrogen CA USA) Quanti-tative PCR assays were performed in 15 120583L reaction volumesusing EXPRESS SYBR GreenER (Invitrogen) and fluores-cence was measured and quantified using an ABI-7900HTVer23 Sequence Detection System (Applied Bio SystemsCA USA) The qPCR thermal profile for amplification of allgenes was as follows melting at 95∘C for 15 secs followed byannealingextending at 60∘C for 1min for 40 cycles
Primers were obtained either commercially or designedde novo (Table 1) using NCBI ldquoPrimer-BLASTrdquo A standardcurve was generated using 6-fold serial 1 5 dilutions ofpooled stock cDNA All samples were assayed in triplicateand only those assays displaying a single PCR peak inthe melt-curve analysis were used All qPCR results werenormalized against the geometricmean of housekeeper genescyclophilin and HPRT according to the geNorm method ofVandesompele et al [28] TNF IL-1R1 andCD68 (and respec-tive controlsHPRTand cyclophilin)were analysed usingTaq-man gene expression assays To control for between-samplevariability mRNA levels were normalized to the geometricmean of the controls The ΔCt for each treatment sample wascompared to the mean ΔCt for control samples using therelative quantification 2-(ΔΔCt) method to determine fold-change [29]
Journal of Nutrition and Metabolism 3
Table 1 Sequences of forward and reverse primers
Entrez geneID Forward primer Reverse primer
K+ channel (Kir62) 83535 51015840-GAA GGA GGC AAA TGA TTG GA-31015840 51015840-AGT GTC CCC CAG ACA AAG TG-31015840
PDE3B 29516 51015840-GAC CGT CGT TGC CTT GTA TT-31015840 51015840-CGA TCG CCT TTC TCT ACT GG-31015840
SOCS3 89829 51015840-TTC TTT ACC ACC GAC GGA AC-31015840 51015840-CGT TGA CAG TCT TCC GAC AA-31015840
Insulin1 24505 Commercially prepared primer QuantiTect Primer Assay Cat no QT00373303 (Qiagen)Insulin2 24506 Commercially prepared primer QuantiTect Primer Assay Cat no QT00177380 (Qiagen)Insulin receptor 24954 51015840-ATC CGT CGC TCC TAT GCT CT-31015840 51015840-TCG TGA GGT TGT GCT TGT TC-31015840
Leptin 25608 Commercially prepared primer QuantiTect Primer Assay Cat no QT00190960 (Qiagen)ObRa 24536 51015840-TGA TAT CGC CAA ACA GCA AA-31015840 51015840-AGT GTC CGC TCT CTT TTG GA-31015840
ObRb 24536 51015840-AAA GCC TGA AAC ATT TGA GCA TC-31015840 51015840-CCA GAA GAA GAG GAC CAA ATA TCA C-31015840
Pdx1 29535 Commercially prepared primer QuantiTect Primer Assay Cat no QT00405328 (Qiagen)STAT3 25125 Commercially prepared primer QuantiTect Primer Assay Cat no QT00183512 (Qiagen)STAT5B 25126 Commercially prepared primer QuantiTect Primer Assay Cat no QT00192024 (Qiagen)CD68 287435 Commercially prepared primer TaqMan expression assay Cat no Rn01495634 (Applied Biosystems)TNF-120572 24835 Commercially prepared primer TaqMan expression assay Cat no Rn01525859 (Applied Biosystems)Cyclophilin 25518 51015840-TTG GGT CGC GTC TGC TTC GA-31015840 51015840-GCC AGG ACC TGT ATG CTT CA-31015840
HPRT 24465 51015840-AGT CCC AGC GTC GTG ATT AG-31015840 51015840-CCC CCT TCA GCA CAC AGA-31015840
23 Statistics All data were analysed by analysis of variance(ANOVA) with maternal diet as factor with post hoc testingto compare groups of interest as required (Holm-Sidak)Data with nonnormal distributions were log transformed ifrequired Analysis was performed using StatView statisticalsoftware (SAS USA) Litter was used as a covariate witheach litter considered a single biological replicate Data arepresented as means plusmn SEM unless otherwise stated
3 Results
31MaternalWeights and Body FatMass Apre-conceptionalHF diet (MHF) resulted in a significant increase in femalebody weight (chow 270 plusmn 5 g MHF 310 plusmn 8 119875 lt 005)and total fat mass compared to chow fed animals (chow178 plusmn 07 MHF 277 plusmn 17 119875 lt 00001) At the startof pregnancy there were no differences in body weightsbetween females randomly assigned to CONT and PLHFgroups (CONT 270 plusmn 8 g PLHF 269 plusmn 7) Total caloricintake was similar between CONT and MHF groups butwas significantly increased in PLHF dams during the first10 days of pregnancy (average caloric intake CONT 023 plusmn0004 kcalgBWday MHF 023 plusmn 0004 PLHF 032 plusmn 001119875 lt 005 for PLHF versus CONT and MHF) thereaftercaloric intake was similar to that of CONT and MHF dams
32 BodyWeight and Body Composition inMale Offspring Asreported previously [14] MHF and PLHF pups had slightlybut statistically significantly reduced body weights and werehypoinsulinemic and hypoleptinemic compared to CONToffspring at birth MHF and PLHF neonates displayed rapidcatch-up growth and were heavier than CONT neonates atthe time of weaning (day 22 CONT 616 plusmn 07 g MHF 705 plusmn09 PLHF 642 plusmn 12)
Final adult body weights were increased in MHF andPLHF male offspring compared to CONT (CONT 562 plusmn 9 gMHF 635 plusmn 18 PLHF 628 plusmn 17) [14] MHF and PLHF adultmale offspring had significantly increased total body fat massat day 150 compared to CONT (Figure 1) There were nodifferences in total fat mass between MHF and PLHF adultoffspring
33 Plasma Analyses MHF and PLHF adult male offspringwere hyperleptinemic and hyperinsulinemic compared toCONT (Figures 2(a) and 2(b)) Fasting blood glucose levelswere similar between groups (CONT 56 plusmn 02mmolLMHF57 plusmn 03 PLHF 57 plusmn 02) Glucose insulin ratios weresignificantly (119875 lt 0001) reduced in MHF and PLHF maleoffspring compared to control (CONT 25 plusmn 02 MHF 13 plusmn01 PLHF 12 plusmn 01) and offspring exhibited increased insulinresistance using HOMA analysis (Figure 2(c)) Plasma TNF-120572 levels were not significantly different between the groups(CONT 36 plusmn 09 pgmL MHF 67 plusmn 19 PLHF 51 plusmn 19)Plasma IL-1120573 levels were significantly increased inMHF (119875 lt005) offspring compared to control (CONT 319plusmn 07 pgmLMHF 383 plusmn 28 PLHF 361 plusmn 22) but were not differentbetween CONT and PLHF offspring
34 Gene Expression MHF offspring showed significantlyhigher levels of pancreatic mRNA levels of SOCS3 ObRband IRS1 compared to CONT and PLHF groups (Figure 3)mRNA levels of these genes were similar between CONT andPLHF offspring Despite these similarities in expression levelsbetween CONT and PLHF offspring significant positivecorrelations that existed between these and other measuredgenes were primarily found in CONT and MHF offspringonly and were not present in PLHF offspring (Table 2)
4 Journal of Nutrition and Metabolism
CONT MHF PLHF
50
40
30
20
10
0
lowast
lowast
Tota
l bod
y fa
t (
)
Figure 1 Total body fat mass (percent) as quantified by DEXA scan in adult CONT MHF and PLHF offspring Data are means plusmn SEM 119899 =10ndash12 per group lowast119875 lt 005
CONT MHF PLHF0
5
10
15
20
25lowast
lowast
Lept
in (n
gm
L)
(a)
CONT MHF PLHF0
1
2
3
4
5
6lowast
lowast
Insu
lin (n
gm
L)
(b)
CONT MHF PLHF0
05
1
15 lowast
lowast
HO
MA
IR
(c)
Figure 2 Fasting plasma leptin (a) insulin (b) and homeostatic model assessment (HOMA) to quantify insulin resistance and beta-cellfunction (c) in adult CONT MHF and PLHF offspring Data are means plusmn SEM 119899 = 10ndash18 per group lowast119875 lt 005
Levels of mRNA of IRS2 PDX-1 and Kir62 were sig-nificantly reduced in PLHF offspring compared to MHF off-spring but were not different between any other groups Pan-creatic INS1 and INS2 expression was significantly increasedin MHF and PLHF offspring compared to CONT (Figure 3)
STAT3 expression was significantly increased inPLHF offspring compared to CONT and MHF offspring(Figure 4(a)) PI3K expression was significantly decreased inPLHF compared to CONT and MHF (Figure 4(b)) Of notePI3K was positively correlated with IRS1 and IRS2 expressionin PLHF offspring (1198772 = 044 119875 lt 005 and 1198772 = 072119875 lt 00005 resp) but not in CONT or MHF offspring
Insulin receptor leptin ObRa and phosphodiesterase 3B(PDE3B) were similar between groups (data not shown) Ahighly significant relationship was observed in all offspringgroups between PDX-1 and ATP-sensitive potassium (Kir62)channel (Figure 5) Given the potential link between SOCS3and inflammation mRNA levels of key proinflammatorycytokines were also evaluated There was an increase inTNF-120572 in PLHF offspring compared to CONT and MHF
offspring (Figure 6(a)) The macrophage marker CD68 wasincreased in MHF and PLHF offspring compared to CONT(Figure 6(b))Therewas an increase in IL-1R1mRNA levels inMHF offspring compared to CONT offspring (Figure 6(c))There were no differences in IL-6R cardiotropin IL-1BGLUT2 GP130 CTNF LIF or TNF-R1 mRNA expressionbetween any of the offspring groups (data not shown)
4 Discussion
There is clear clinical and experimental evidence that mater-nal obesity predisposes to obesity and metabolic disorders inoffspring [14 30] However despite the common phenotypeof offspring obesity and hyperinsulinemia the present studyshows that at least in an animal model that maternal nutri-tional history determines the pattern of mRNA expressionof key regulatory genes in the pancreas of adult offspringThese data suggest that different regulatory pathways maylead to a similar metabolic phenotype and are in agreementwith previous experimental studies suggesting that there
Journal of Nutrition and Metabolism 5
SOCS3
0
100
200
300
400
500
600
700
800
( o
f con
trol)
lowast lowast
CONT MHF PLHF
(a)
ObRb
0
500
1000
1500
2000
2500 lowast lowast
( o
f con
trol)
CONT MHF PLHF
(b)
IRS1
0
50
100
150
200
250
300 lowastlowast
( o
f con
trol)
CONT MHF PLHF
(c)
IRS2
0
50
100
150
200lowast
( o
f con
trol)
CONT MHF PLHF
(d)
PDX-1
0
50
100
150
200
250 lowast
( o
f con
trol)
CONT MHF PLHF
(e)
0
50
100
150
200
250
300 lowast
K+ channel
CONT MHF PLHF
( o
f con
trol)
(f)
Ins1
0
50
100
150
200
250
300 lowast
lowast
CONT MHF PLHF
( o
f con
trol)
(g)
Ins2
CONT MHF PLHF0
50100150200250300350400 lowast
lowast
( o
f con
trol)
(h)
Figure 3 Pancreatic mRNA expression in adult CONT MHF and PLHF offspring SOCS3 suppressor of cytokine signaling 3 IRS insulinreceptor substrate PDX1 pancreatic and duodenal homeobox 1 ObRb full length leptin receptor Ins insulin Kir62 potassium channelsubunit Data are means plusmn SEM 119899 = 10ndash18 per group lowast119875 lt 005
6 Journal of Nutrition and Metabolism
Table 2 Correlations between SOCS3 and other key genes N = 8ndash13 per group
CONT MHF PLHFSOCS3 versus ObRb 119877
2 = 069 P = 008 R2 = 087 P = 00008 1198772 = 003 P = 064
SOCS3 versus IR R2 = 067 P = 002 1198772 = 002 P = 071 119877
2 = 0002 P = 090SOCS3 versus IRS1 R2 = 084 P = 001 R2 = 058 P = 002 119877
2 = 004 P = 051SOCS3 versus IRS2 R2 = 098 P = 0002 R2 = 058 P = 002 119877
2 = 015 P = 021SOCS3 versus Kir62 channel R2 = 082 P = 003 R2 = 093 P lt 00001 119877
2 = 0002 P = 090SOCS3 versus Pdx1 R2 = 078 P = 0009 R2 = 091 P = 00003 119877
2 = 007 P = 040
CONT MHF PLHF0
100
200
300
400
500
STAT
3 (
of c
ontro
l)
lowast
lowast
(a)
PI3K
( o
f con
trol)
CONT MHF PLHF0
50
100
150lowast
lowast
(b)
Figure 4 Pancreatic STAT3 (a) and PI3K mRNA expression (b) Data are means plusmn SEM 119899 = 10ndash18 per group STAT3 signal transducer andactivator of transcription 3 PI3K phosphatidylinositol 3-kinase lowast119875 lt 005
may be separate influences of maternal obesity during thepericonceptional period and late gestation on adiposity inoffspring [31] We have previously shown that ldquoprogrammedrdquoobesity and simple diet-induced obesity can develop viadifferent underlying mechanisms [32] The present studyfurther reinforces the importance of the timing of the dietaryinsult on the metabolic derangements that ensue in offspringduring adulthood
SOCS3 inhibition of ObRb signaling has been proposedas a mechanism for leptin resistance in a range of tissues [33ndash35] including pancreatic 120573 cells [36] It is clear that leptinresistance in PLHF pancreata was mediated by mechanismsthat did not involve SOCS3 and this has been reported previ-ously in rat models of HF feeding [37 38] SOCS3 is normallyincreased at sites of both acute and chronic inflammation [39]and has been shown to specifically suppress STAT3 activation[40] This agrees with the present study a marked increasein SOCS3 mRNA levels in MHF offspring is paralleled bySTAT3 levels similar to control offspring Leptin is knownto increase SOCS3 expression in isolated human pancreaticislets and in islets from obob mice treated in vivo [36] ourdata imply that leptin-induced SOCS3 stimulation appearsintact in MHF offspring Conversely in PLHF offspring
SOCS3 expression levels were similar to controls in thepresence of markedly elevated STAT3 expression Of notedespite similar expression levels of SOCS3 between CONTand PLHF offspring the positive correlations that wereobserved between SOCS3 and other key signaling genes wererestricted mainly to CONT and MHF offspring only Thismay point to a maladaptation in PLHF offspring due to theacute HF dietary exposure and a change in homeostatic set-points in MHF offspring due to mothers entering pregnancyin an already obese state Despite the link between SOCS3and inflammation there were no observed changes in TNF-120572 mRNA levels in the MHF group whilst there was asignificant increase in TNF-120572 expression in PLHF offspringIn addition mRNA levels of the macrophage marker CD68were increased in both HF groups and taken togethermay indicate differing macrophage phenotypespolarizationstates resulting in divergent offspring inflammatory profilesdependent upon prior maternal nutritional background Ofnote despite marked changes in inflammatorymarkers at thetissue level we observed no changes in circulating plasmaTNF-120572 concentrationHowever despite enhanced plasma IL-1120573 concentrations pancreatic IL-1120573 expression was unaltered
Journal of Nutrition and Metabolism 7
CONT
0 250 500 7500
250
500
750
PDX-1
1199032= 097
K+ch
anne
l
(a)
MHF
0 250 500 750 10000
500
1000
15001199032= 094
PDX-1
K+ch
anne
l
(b)
PLHF
0 50 100 150 200 250 3000
50
100
150
200
250
3001199032= 078
PDX-1
K+ch
anne
l
(c)
Figure 5 Relationship between PDX-1 and potassium channel subunit Kir62 in adult CONT MHF and PLHF male offspring PDX-1pancreatic and duodenal homeobox 1119873 = 8ndash10 per group
between groups implicating other tissues as possible systemicsources of this proinflammatory cytokine
PLHF offspring exhibited decreased PI3K mRNA levelscompared to CONT animals Induction of the ObRbPI3Ksignaling pathway prolongs potassium channel openingleading to decreased insulin exocytosis a direct action ofleptin which inhibits insulin secretion [41] Moreover thelowered PI3K mRNA levels in PLHF offspring were con-comitant with a reduction in IRS1 and IRS2 mRNA whosephosphorylation is coupled to PI3K activation Of notedownstreamof PI3K PDE3B gene expression levels remainedsimilar to controls suggesting that altered leptin signalingwas achieved in earlier stages of this pathway Thus attenua-tion of the ObRbPI3K signaling pathwaymaymediate leptinresistance in these hyperleptinemic PLHF offspring Sucha mechanism has been observed before in hypothalamicneurons of rats with diet-induced obesity (DIO) [42]
IRS proteins and PI3K have leptin independent rolesin the 120573 cell Both are found downstream of the insulinreceptor in the IRIRSPI3K pathway which mediates theautocrine actions of the insulin-IR complex [43 44] Ourobserved attenuation of PI3K and IRS mRNA did nothowever affect insulin synthesismdashPLHF animals were hyper-insulinemic and preproinsulin mRNA concentrations wereelevated Furthermore mRNA concentrations of IR wereunaffected potentially due to lack of SOCS3 induction inPLHF offspring similar to Controls Thus these mRNAchanges of PI3K coupled to IRS likely reflect a mechanismof leptin resistance in this group
Mean concentrations of ObRb mRNA in PLHF offspringwere similar to controls but a significant inverse relationshipbetween plasma leptin and ObRb mRNA levels in the PLHFgroup was suggestive of ObRb downregulation A downregu-lation of receptor populations in the face of elevated ligand
8 Journal of Nutrition and Metabolism
CONT MHF PLHF0
50
100
150
200
TNF-120572
mRN
A (
of c
ontro
l)
lowast
lowast
(a)
CD68
mRN
A (
of c
ontro
l)
CONT MHF PLHF0
50
100
150
200
lowast
lowast
(b)
IL1-
R1 m
RNA
( o
f con
trol)
CONT MHF PLHF0
50
100
150
200lowast
(c)
Figure 6 Pancreatic mRNA expression of TNF-120572 (a) CD68 (b) and IL1-R1 (c) in adult CONT MHF and PLHF offspring TNF-120572 tumornecrosis factor-alpha CD68 cluster of differentiation 68 IL1-R1 interleukin 1 receptor type 1 Data are means plusmn SEM 119899 = 10ndash18 per grouplowast
119875 lt 005
concentrations is not uncommon in resistant states It isseen for example in the hypothalamus in the presence ofhyperleptinemia [45ndash47] and during pregnancy which is anatural state of leptin resistance [48]
A rise in IRS1 and IRS2 mRNA associated with elevatedObRb and SOCS3 mRNA in MHF offspring may seemunexpected since SOCS3 is known to target insulin receptorsignaling by interference with IRS action [49] However sev-eral studies have demonstrated that SOCS3 mediates insulinresistance in both adipocytes and hepatocytes through post-translational modifications such as inhibition of IRS tyro-sine phosphorylation [50] and targeted ubiquitin-mediateddegradation of IRS1 and IRS2 without effecting transcription[49 51] IRS proteins are involved in both leptin signalingand insulin signaling so altered gene expression levels mayrepresent changes in either (or both) pathways although IRSresponse to leptin is reportedly much lower than that seenwith insulin [52] We observed an increase in IRS1 expressionin MHF offspring compared to Controls This upregulationof IRS1 expression may be related to the increased insulin
production of the MHF offspring given the known role ofIRS1 in the regulation of insulin secretion [53]
SOCS3 is also reported to repress preproinsulin genetranscription [41] although other mechanisms clearly coun-tered this action inMHFoffspring since Ins1 and Ins2mRNAexpression levels remained elevated This observation wasconfirmed by the significantly raised levels of circulatingplasma insulin and the lack of any correlation in levels ofmRNA between SOCS3 and Ins1 or Ins2
Some of the observed differences may relate to theinitial hyperphagia observed in the PLHF maternal groupas reported previously [14] Since rodents primarily eat ona calorific basis they will passively over-consume calorieswhen initially placed on an HF diet and self-regulate backto a normal total caloric intake over a period of a few daysIt has been shown in other rodent models that alterations innutrition around the time of implantation can lead to lastingmetabolic effects in the offspring [54] Maternal obesity hasbeen shown to exert adverse effects as early as the oocyteand preimplantation embryo stage and that these effects may
Journal of Nutrition and Metabolism 9
contribute to the lasting morbidity observed in offspring[55] Similarly even a very short period of altered nutrition(restricted to one ovulatory cycle) prior to natural mating canelicit lasting effects on the offspring [56] and this representsa further point of difference between the MHF and PLHFgroups
The present study was not designed to incorporate theeffect of a reversal paradigm whereby mothers fed thepre-conceptional HF diet were placed on a standard dietthroughout pregnancy and lactation However while there isevidence that a period of dietary restriction in obese mothersmay ablate the programming of obesity in offspring [57]it may be associated with an activation of the stress axisin the offspring [31] Further the data are inconsistent ithas also been shown that dams with diet-induced obesityfed a standard diet throughout pregnancy and lactation stilltransmit the propensity for obesity and metabolic disease totheir offspring [58]
We recognize that gene expression analysis on wholepancreatic tissue is a limitation of the present study Dueto the size of the cohort and logistical considerations onlywhole pancreatic tissue was a possibility whereas ideallyonly pure islet populations would have been examined [59]However for the key genes presented such as ObRb SOCS3and IRS expression has been shown to be primarily confinedto the endocrine islets and not been detected in the exocrinepancreas [39 41 60] therefore we are confident that theobserved changes in the genes studied represent changes inthe islets Further although we have shown that differencesin pancreatic gene expression in offspring are dependentupon the timing of maternal high fat diet exposure this doesnot itself imply causation in the development of the obesephenotypemdashother relevant pathways including liver skeletalmuscle and fat metabolism will also be integrated into theprogrammed phenotype development
This study has demonstrated enduring deleterious effectsof a maternal HF diet during early windows of develop-ment Maternal HF feeding both before conception andorthrough pregnancy and lactation induced obesity hyper-insulinemia and hyperleptinemia in adult offspring eventhough offspring were fed a standard diet postnatally Timingof exposure to the maternal HF diet differentially alteredgene expression in signaling pathways downstream of thepancreatic leptin receptor that mediated leptin resistanceThis also raises the possibility that despite common offspringobesity treatment modalities may only be efficacious in asubpopulation of treated offspring as the current data suggestthat the metabolic pathways disrupted will be dependentupon the timing of the early life dietary insult
Abbreviations
CD68 Cluster of differentiation 68CONT ControlDEXA Dual energy X-ray absorptiometryELISA Enzyme-linked immunosorbent assayGLUT2 Glucose transporter 2GP130 Glycoprotein 130HF High fat
HOMA Homeostatic model assessmentHPRT HypoxanthinephophoribosyltransferaseIL InterleukinIns InsulinIR Insulin resistanceIRS Insulin receptor substrateKir62 Potassium channel subunitLIF Leukemia inhibitory factorMHF Maternal high fatOBR Leptin receptorPDE3B Phosphodiesterase 3BPDX-1 Pancreatic and duodenal homeobox-1PI3K Phosphatidylinositide 3-kinasePLHF Pregnancy and lactation high fatSOCS3 Suppressor of cytokine signaling 3STAT3 Signal transducer and activator of
transcription 3TNF-120572 Tumor necrosis factor-alpha
Conflict of Interests
The authors have no conflict of interests with any financialorganization regarding the material discussed in this paper
Author Contributions
Mark H Vickers developed the animal model designed thestudy completed the data analysis and wrote the first draftof the paper Graham J Howie maintained the experimentalcohort performed the gene expression analysis and con-tributed to the original draft paper Clare M Reynolds per-formed the inflammatory related assays and gene expressionwork Deborah M Sloboda provided input into data analysisand interpretation
Acknowledgments
The authors acknowledge the technical support of TaniaKamal with endocrine assays The authors acknowledgefunding support from the Health Research Council of NewZealand and Gravida National Centre for Growth andDevelopment
References
[1] M M Hillemeier C S Weisman C Chuang D S Downs JMcCall-Hosenfeld and F Camacho ldquoTransition to overweightor obesity among women of reproductive agerdquo Journal ofWomenrsquos Health vol 20 no 5 pp 703ndash710 2011
[2] J E Norman and RM Reynolds ldquoThe consequences of obesityand excess weight gain in pregnancyrdquo Proceedings of the Nutri-tion Society vol 70 no 4 pp 450ndash456 2011
[3] H A Abenhaim R A Kinch L Morin A Benjamin and RUsher ldquoEffect of prepregnancy body mass index categories onobstetrical and neonatal outcomesrdquo Archives of Gynecology andObstetrics vol 275 no 1 pp 39ndash43 2007
[4] S DHMalnick andH Knobler ldquoThemedical complications ofobesityrdquo Oxford Journals Medicine vol 99 no 9 pp 565ndash5792006
10 Journal of Nutrition and Metabolism
[5] P M Catalano ldquoManagement of obesity in pregnancyrdquo Obstet-rics and Gynecology vol 109 no 2 pp 419ndash433 2007
[6] L Poston ldquoGestational weight gain influences on the long-termhealth of the childrdquo Current Opinion in Clinical Nutrition andMetabolic Care vol 15 pp 252ndash257 2012
[7] H Hochner Y Friedlander R Calderon-Margalit et al ldquoAsso-ciations of maternal prepregnancy body mass index and gesta-tional weight gain with adult offspring cardiometabolic riskfactors the Jerusalem perinatal family follow-up studyrdquo Circu-lation vol 125 no 11 pp 1381ndash1389 2012
[8] S J Spencer ldquoEarly life programming of obesity the impactof the perinatal environment on the development of obesityand metabolic dysfunction in the offspringrdquo Current DiabetesReviews vol 8 no 1 pp 55ndash68 2012
[9] H Chen D Simar K Lambert J Mercier and M J MorrisldquoMaternal and postnatal overnutrition differentially impactappetite regulators and fuel metabolismrdquo Endocrinology vol149 no 11 pp 5348ndash5356 2008
[10] P M Catalano ldquoObesity and pregnancymdashthe propagation of aviscous cyclerdquo Journal of Clinical Endocrinology and Metabo-lism vol 88 no 8 pp 3505ndash3506 2003
[11] M W Gillman S L Rifas-Shiman K Kleinman E Oken JW Rich-Edwards and EM Taveras ldquoDevelopmental origins ofchildhood overweight potential public health impactrdquo Obesityvol 16 no 7 pp 1651ndash1656 2008
[12] E Oken EM Taveras K P Kleinman JW Rich-Edwards andM W Gillman ldquoGestational weight gain and child adiposity atage 3 yearsrdquoTheAmerican Journal of Obstetrics and Gynecologyvol 196 no 4 pp 322e1ndash322e8 2007
[13] H M Ehrenberg B M Mercer and P M Catalano ldquoThe influ-ence of obesity and diabetes on the prevalence of macrosomiardquoThe American Journal of Obstetrics and Gynecology vol 191 no3 pp 964ndash968 2004
[14] G J Howie D M Sloboda T Kamal and M H VickersldquoMaternal nutritional history predicts obesity in adult offspringindependent of postnatal dietrdquo Journal of Physiology vol 587no 4 pp 905ndash915 2009
[15] D M Sloboda G J Howie A Pleasants P D Gluckman andM H Vickers ldquoPre- and postnatal nutritional histories influ-ence reproductive maturation and ovarian function in the ratrdquoPLoS ONE vol 4 no 8 Article ID e6744 2009
[16] M C Battista M F Hivert K Duval and J P BaillargeonldquoIntergenerational cycle of obesity and diabetes how can wereduce the burdens of these conditions on the health of futuregenerationsrdquo Experimental Diabetes Research vol 2011 ArticleID 596060 19 pages 2011
[17] T J Kieffer and J F Habener ldquoThe adipoinsular axis effects ofleptin on pancreatic 120573-cellsrdquo The American Journal of Physiol-ogy vol 278 no 1 pp E1ndashE14 2000
[18] S L Gray C Donald A Jetha S D Covey and T J KiefferldquoHyperinsulinemia precedes insulin resistance in mice lackingpancreatic 120573-cell leptin signalingrdquo Endocrinology vol 151 no 9pp 4178ndash4186 2010
[19] S D Covey R DWideman CMcDonald et al ldquoThe pancreat-ic 120573 cell is a key site formediating the effects of leptin on glucosehomeostasisrdquo Cell Metabolism vol 4 no 4 pp 291ndash302 2006
[20] K J Dudley DM Sloboda K L Connor J Beltrand andMHVickers ldquoOffspring ofmothers fed a high fat diet display hepaticcell cycle inhibition and associated changes in gene expressionand DNA methylationrdquo PLoS ONE vol 6 no 7 Article IDe21662 2011
[21] K L Connor M H Vickers J Beltrand M J Meaney and DM Sloboda ldquoNature nurture or nutrition Impact of maternalnutrition on maternal care offspring development and repro-ductive functionrdquo Journal of Physiology vol 590 part 9 pp2167ndash2180 2012
[22] J Beltrand D M Sloboda K L Connor M Truong and MH Vickers ldquoThe effect of neonatal leptin antagonism in malerat offspring is dependent upon the interaction between priormaternal nutritional status and post-weaning dietrdquo Journal ofNutrition andMetabolism vol 2012 Article ID 296935 10 pages2012
[23] K L Connor M H Vickers C Cupido E Sirimanne and DM Sloboda ldquoMaternal high fat diet during critical windowsof development alters adrenal cortical and medullary enzymeexpression in adult male rat offspringrdquo Journal of the Develop-mental Origins of Health and Disease vol 1 no 4 pp 245ndash2542010
[24] P J Mark C Sisala K Connor et al ldquoA maternal high-fat dietin rat pregnancy reduces growth of the fetus and the placen-tal junctional zone but not placental labyrinth zone growthrdquoJournal of the Developmental Origins of Health and Disease vol2 no 1 pp 63ndash70 2011
[25] D R Matthews J P Hosker A S Rudenski B A Naylor DF Treacher and R C Turner ldquoHomeostasis model assessmentinsulin resistance and 120573-cell function from fasting plasmaglucose and insulin concentrations in manrdquo Diabetologia vol28 no 7 pp 412ndash419 1985
[26] M P Hermans J C Levy R J Morris and R C TurnerldquoComparison of tests of120573-cell function across a range of glucosetolerance from normal to diabetesrdquo Diabetes vol 48 no 9 pp1779ndash1786 1999
[27] A B Bernal M H Vickers M B Hampton R A Poynton andD M Sloboda ldquoMaternal undernutrition significantly impactsovarian follicle number and increases ovarian oxidative stress inadult rat offspringrdquo PLoS ONE vol 5 no 12 Article ID e155582010
[28] J Vandesompele K de Preter F Pattyn et al ldquoAccurate nor-malization of real-time quantitative RT-PCR data by geometricaveraging of multiple internal control genesrdquo Genome biologyvol 3 no 7 Article ID RESEARCH0034 2002
[29] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the 2-ΔΔCT methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
[30] J Armitage L Poston and P Taylor ldquoDevelopmental originsof obesity and the metabolic syndrome the role of maternalobesityrdquo Frontiers of Hormone Research vol 36 pp 73ndash84 2007
[31] S Zhang L Rattanatray J L Morrison L M Nicholas S Lieand I C McMillen ldquoMaternal obesity and the early originsof childhood obesity weighing up the benefits and costs ofmaternal weight loss in the periconceptional period for theoffspringrdquo Experimental Diabetes Research vol 2011 Article ID585749 10 pages 2011
[32] N M Thompson A M Norman S S Donkin et al ldquoPrenataland postnatal pathways to obesity different underlying mech-anisms different metabolic outcomesrdquo Endocrinology vol 148no 5 pp 2345ndash2354 2007
[33] T Fuentes I Ara A Guadalupe-Grau et al ldquoLeptin receptor170 kDa (OB-R170) protein expression is reduced in obesehuman skeletal muscle a potential mechanism of leptin resis-tancerdquo Experimental Physiology vol 95 no 1 pp 160ndash171 2010
[34] Z Wang Y T Zhou T Kakuma et al ldquoLeptin resistance ofadipocytes in obesity role of suppressors of cytokine signalingrdquo
Journal of Nutrition and Metabolism 11
Biochemical and Biophysical Research Communications vol 277no 1 pp 20ndash26 2000
[35] C Bjoslashrbaeligk K El-Haschimi J D Frantz and J S Flier ldquoThe roleof SOCS-3 in leptin signaling and leptin resistancerdquoThe Journalof Biological Chemistry vol 274 no 42 pp 30059ndash30065 1999
[36] K Laubner T J Kieffer N T Lam X Niu F Jakob and JSeufert ldquoInhibition of preproinsulin gene expression by leptininduction of suppressor of cytokine signaling 3 in pancreatic 120573-cellsrdquo Diabetes vol 54 no 12 pp 3410ndash3417 2005
[37] C Peiser G P McGregor and R E Lang ldquoLeptin receptorexpression and suppressor of cytokine signaling transcriptlevels in high-fat-fed ratsrdquo Life Sciences vol 67 no 24 pp 2971ndash2981 2000
[38] G R Steinberg A C Smith S Wormald P Malenfant CCollier and D J Dyck ldquoEndurance training partially reversesdietary-induced leptin resistance in rodent skeletal musclerdquoTheAmerican Journal of Physiology vol 286 no 1 pp E57ndashE632004
[39] G E White A Cotterill M R Addley E J Soilleux and DR Greaves ldquoSuppressor of cytokine signalling protein SOCS3expression is increased at sites of acute and chronic inflamma-tionrdquo Journal of Molecular Histology vol 42 no 2 pp 137ndash1512011
[40] H YasukawaMOhishiHMori et al ldquoIL-6 induces an anti-in-flammatory response in the absence of SOCS3 inmacrophagesrdquoNature Immunology vol 4 no 6 pp 551ndash556 2003
[41] J Seufert ldquoLeptin effects on pancreatic 120573-cell gene expressionand functionrdquo Diabetes vol 53 no 1 pp S152ndashS158 2004
[42] A S Metlakunta M Sahu and A Sahu ldquoHypothalamicphosphatidylinositol 3-kinase pathway of leptin signaling isimpaired during the development of diet-induced obesity inFVBNmicerdquo Endocrinology vol 149 no 3 pp 1121ndash1128 2008
[43] C A Aspinwall W J Qian M G Roper R N Kulkarni C RKahn and R T Kennedy ldquoRoles of insulin receptor substrate-1phosphatidylinositol 3-kinase and release of intracellular Ca2+stores in insulin-stimulated insulin secretion in 120573-cellsrdquo TheJournal of Biological Chemistry vol 275 no 29 pp 22331ndash223382000
[44] I B Leibiger B Leibiger T Moede and P O BerggrenldquoExocytosis of insulin promotes insulin gene transcription viathe insulin receptorPI-3 kinasep70 s6 kinase and CaM kinasepathwaysrdquoMolecular Cell vol 1 no 6 pp 933ndash938 1998
[45] R L Martin E Perez Y J He R Dawson Jr andW J MillardldquoLeptin resistance is associated with hypothalamic leptin recep-tor mRNA and protein downregulationrdquo Metabolism vol 49no 11 pp 1479ndash1484 2000
[46] J Wilsey and P J Scarpace ldquoCaloric restriction reverses thedeficits in leptin receptor protein and leptin signaling capacityassociated with diet-induced obesity role of leptin in the reg-ulation of hypothalamic long-form leptin receptor expressionrdquoJournal of Endocrinology vol 181 no 2 pp 297ndash306 2004
[47] Y Zhang and P J Scarpace ldquoThe role of leptin in leptin resist-ance and obesityrdquo Physiology and Behavior vol 88 no 3 pp249ndash256 2006
[48] D R Grattan S R Ladyman and R A Augustine ldquoHormonalinduction of leptin resistance during pregnancyrdquoPhysiology andBehavior vol 91 no 4 pp 366ndash374 2007
[49] L RuiM YuanD Frantz S Shoelson andM FWhite ldquoSOCS-1 and SOCS-3 block insulin signaling by ubiquitin-mediateddegradation of IRS1 and IRS2rdquoThe Journal of Biological Chem-istry vol 277 no 44 pp 42394ndash42398 2002
[50] K Ueki T Kondo and C R Kahn ldquoSuppressor of cytokine sig-naling 1 (SOCS-1) and SOCS-3 cause insulin resistance throughinhibition of tyrosine phosphorylation of insulin receptor sub-strate proteins by discrete mechanismsrdquoMolecular and CellularBiology vol 24 no 12 pp 5434ndash5446 2004
[51] S B Jorgensen O rsquoNeill HM L Sylow et al ldquoDeletion of skel-etal muscle SOCS3 prevents insulin resistance in obesityrdquo Dia-betes vol 62 no 1 pp 56ndash64 2013
[52] W Becker ldquoLeptin signal transductionrdquo in Leptin and LeptinAntagonists A Gertler Ed pp 1ndash9 Landes Bioscience AustinTex USA 2009
[53] R N Kulkarni J N Winnay M Daniels et al ldquoAltered func-tion of insulin receptor substrate 1-deficient mouse islets andcultured 120573-cell linesrdquo Journal of Clinical Investigation vol 104no 12 pp R69ndashR75 1999
[54] W Y Kwong A E Wild P Roberts A C Willis and T PFleming ldquoMaternal undernutrition during the preimplantationperiod of rat development causes blastocyst abnormalities andprogramming of postnatal hypertensionrdquoDevelopment vol 127no 19 pp 4195ndash4202 2000
[55] E S Jungheim E L Schoeller K L Marquard E D LoudenJ E Schaffer and K H Moley ldquoDiet-induced obesity modelabnormal oocytes and persistent growth abnormalities in theoffspringrdquo Endocrinology vol 151 no 8 pp 4039ndash4046 2010
[56] A J Watkins A Wilkins C Cunningham et al ldquoLow proteindiet fed exclusively during mouse oocyte maturation leadsto behavioural and cardiovascular abnormalities in offspringrdquoJournal of Physiology vol 586 no 8 pp 2231ndash2244 2008
[57] E Zambrano P M Martınez-Samayoa G L Rodrıguez-Gonzalez and P W Nathanielsz ldquoDietary intervention prior topregnancy reverses metabolic programming in male offspringof obese ratsrdquo Journal of Physiology vol 588 no 10 pp 1791ndash1799 2010
[58] M S Patel andM Srinivasan ldquoMetabolic programming causesand consequencesrdquoThe Journal of Biological Chemistry vol 277no 3 pp 1629ndash1632 2002
[59] R A Simmons ldquoRole of metabolic programming in the patho-genesis of 120573-cell failure in postnatal liferdquo Reviews in Endocrineand Metabolic Disorders vol 8 no 2 pp 95ndash104 2007
[60] I B Leibiger B Leibiger and P O Berggren ldquoInsulin signalingin the pancreatic 120573-cellrdquoAnnual Review of Nutrition vol 28 pp233ndash251 2008

Journal of Nutrition and Metabolism 3
Table 1 Sequences of forward and reverse primers
Entrez geneID Forward primer Reverse primer
K+ channel (Kir62) 83535 51015840-GAA GGA GGC AAA TGA TTG GA-31015840 51015840-AGT GTC CCC CAG ACA AAG TG-31015840
PDE3B 29516 51015840-GAC CGT CGT TGC CTT GTA TT-31015840 51015840-CGA TCG CCT TTC TCT ACT GG-31015840
SOCS3 89829 51015840-TTC TTT ACC ACC GAC GGA AC-31015840 51015840-CGT TGA CAG TCT TCC GAC AA-31015840
Insulin1 24505 Commercially prepared primer QuantiTect Primer Assay Cat no QT00373303 (Qiagen)Insulin2 24506 Commercially prepared primer QuantiTect Primer Assay Cat no QT00177380 (Qiagen)Insulin receptor 24954 51015840-ATC CGT CGC TCC TAT GCT CT-31015840 51015840-TCG TGA GGT TGT GCT TGT TC-31015840
Leptin 25608 Commercially prepared primer QuantiTect Primer Assay Cat no QT00190960 (Qiagen)ObRa 24536 51015840-TGA TAT CGC CAA ACA GCA AA-31015840 51015840-AGT GTC CGC TCT CTT TTG GA-31015840
ObRb 24536 51015840-AAA GCC TGA AAC ATT TGA GCA TC-31015840 51015840-CCA GAA GAA GAG GAC CAA ATA TCA C-31015840
Pdx1 29535 Commercially prepared primer QuantiTect Primer Assay Cat no QT00405328 (Qiagen)STAT3 25125 Commercially prepared primer QuantiTect Primer Assay Cat no QT00183512 (Qiagen)STAT5B 25126 Commercially prepared primer QuantiTect Primer Assay Cat no QT00192024 (Qiagen)CD68 287435 Commercially prepared primer TaqMan expression assay Cat no Rn01495634 (Applied Biosystems)TNF-120572 24835 Commercially prepared primer TaqMan expression assay Cat no Rn01525859 (Applied Biosystems)Cyclophilin 25518 51015840-TTG GGT CGC GTC TGC TTC GA-31015840 51015840-GCC AGG ACC TGT ATG CTT CA-31015840
HPRT 24465 51015840-AGT CCC AGC GTC GTG ATT AG-31015840 51015840-CCC CCT TCA GCA CAC AGA-31015840
23 Statistics All data were analysed by analysis of variance(ANOVA) with maternal diet as factor with post hoc testingto compare groups of interest as required (Holm-Sidak)Data with nonnormal distributions were log transformed ifrequired Analysis was performed using StatView statisticalsoftware (SAS USA) Litter was used as a covariate witheach litter considered a single biological replicate Data arepresented as means plusmn SEM unless otherwise stated
3 Results
31MaternalWeights and Body FatMass Apre-conceptionalHF diet (MHF) resulted in a significant increase in femalebody weight (chow 270 plusmn 5 g MHF 310 plusmn 8 119875 lt 005)and total fat mass compared to chow fed animals (chow178 plusmn 07 MHF 277 plusmn 17 119875 lt 00001) At the startof pregnancy there were no differences in body weightsbetween females randomly assigned to CONT and PLHFgroups (CONT 270 plusmn 8 g PLHF 269 plusmn 7) Total caloricintake was similar between CONT and MHF groups butwas significantly increased in PLHF dams during the first10 days of pregnancy (average caloric intake CONT 023 plusmn0004 kcalgBWday MHF 023 plusmn 0004 PLHF 032 plusmn 001119875 lt 005 for PLHF versus CONT and MHF) thereaftercaloric intake was similar to that of CONT and MHF dams
32 BodyWeight and Body Composition inMale Offspring Asreported previously [14] MHF and PLHF pups had slightlybut statistically significantly reduced body weights and werehypoinsulinemic and hypoleptinemic compared to CONToffspring at birth MHF and PLHF neonates displayed rapidcatch-up growth and were heavier than CONT neonates atthe time of weaning (day 22 CONT 616 plusmn 07 g MHF 705 plusmn09 PLHF 642 plusmn 12)
Final adult body weights were increased in MHF andPLHF male offspring compared to CONT (CONT 562 plusmn 9 gMHF 635 plusmn 18 PLHF 628 plusmn 17) [14] MHF and PLHF adultmale offspring had significantly increased total body fat massat day 150 compared to CONT (Figure 1) There were nodifferences in total fat mass between MHF and PLHF adultoffspring
33 Plasma Analyses MHF and PLHF adult male offspringwere hyperleptinemic and hyperinsulinemic compared toCONT (Figures 2(a) and 2(b)) Fasting blood glucose levelswere similar between groups (CONT 56 plusmn 02mmolLMHF57 plusmn 03 PLHF 57 plusmn 02) Glucose insulin ratios weresignificantly (119875 lt 0001) reduced in MHF and PLHF maleoffspring compared to control (CONT 25 plusmn 02 MHF 13 plusmn01 PLHF 12 plusmn 01) and offspring exhibited increased insulinresistance using HOMA analysis (Figure 2(c)) Plasma TNF-120572 levels were not significantly different between the groups(CONT 36 plusmn 09 pgmL MHF 67 plusmn 19 PLHF 51 plusmn 19)Plasma IL-1120573 levels were significantly increased inMHF (119875 lt005) offspring compared to control (CONT 319plusmn 07 pgmLMHF 383 plusmn 28 PLHF 361 plusmn 22) but were not differentbetween CONT and PLHF offspring
34 Gene Expression MHF offspring showed significantlyhigher levels of pancreatic mRNA levels of SOCS3 ObRband IRS1 compared to CONT and PLHF groups (Figure 3)mRNA levels of these genes were similar between CONT andPLHF offspring Despite these similarities in expression levelsbetween CONT and PLHF offspring significant positivecorrelations that existed between these and other measuredgenes were primarily found in CONT and MHF offspringonly and were not present in PLHF offspring (Table 2)
4 Journal of Nutrition and Metabolism
CONT MHF PLHF
50
40
30
20
10
0
lowast
lowast
Tota
l bod
y fa
t (
)
Figure 1 Total body fat mass (percent) as quantified by DEXA scan in adult CONT MHF and PLHF offspring Data are means plusmn SEM 119899 =10ndash12 per group lowast119875 lt 005
CONT MHF PLHF0
5
10
15
20
25lowast
lowast
Lept
in (n
gm
L)
(a)
CONT MHF PLHF0
1
2
3
4
5
6lowast
lowast
Insu
lin (n
gm
L)
(b)
CONT MHF PLHF0
05
1
15 lowast
lowast
HO
MA
IR
(c)
Figure 2 Fasting plasma leptin (a) insulin (b) and homeostatic model assessment (HOMA) to quantify insulin resistance and beta-cellfunction (c) in adult CONT MHF and PLHF offspring Data are means plusmn SEM 119899 = 10ndash18 per group lowast119875 lt 005
Levels of mRNA of IRS2 PDX-1 and Kir62 were sig-nificantly reduced in PLHF offspring compared to MHF off-spring but were not different between any other groups Pan-creatic INS1 and INS2 expression was significantly increasedin MHF and PLHF offspring compared to CONT (Figure 3)
STAT3 expression was significantly increased inPLHF offspring compared to CONT and MHF offspring(Figure 4(a)) PI3K expression was significantly decreased inPLHF compared to CONT and MHF (Figure 4(b)) Of notePI3K was positively correlated with IRS1 and IRS2 expressionin PLHF offspring (1198772 = 044 119875 lt 005 and 1198772 = 072119875 lt 00005 resp) but not in CONT or MHF offspring
Insulin receptor leptin ObRa and phosphodiesterase 3B(PDE3B) were similar between groups (data not shown) Ahighly significant relationship was observed in all offspringgroups between PDX-1 and ATP-sensitive potassium (Kir62)channel (Figure 5) Given the potential link between SOCS3and inflammation mRNA levels of key proinflammatorycytokines were also evaluated There was an increase inTNF-120572 in PLHF offspring compared to CONT and MHF
offspring (Figure 6(a)) The macrophage marker CD68 wasincreased in MHF and PLHF offspring compared to CONT(Figure 6(b))Therewas an increase in IL-1R1mRNA levels inMHF offspring compared to CONT offspring (Figure 6(c))There were no differences in IL-6R cardiotropin IL-1BGLUT2 GP130 CTNF LIF or TNF-R1 mRNA expressionbetween any of the offspring groups (data not shown)
4 Discussion
There is clear clinical and experimental evidence that mater-nal obesity predisposes to obesity and metabolic disorders inoffspring [14 30] However despite the common phenotypeof offspring obesity and hyperinsulinemia the present studyshows that at least in an animal model that maternal nutri-tional history determines the pattern of mRNA expressionof key regulatory genes in the pancreas of adult offspringThese data suggest that different regulatory pathways maylead to a similar metabolic phenotype and are in agreementwith previous experimental studies suggesting that there
Journal of Nutrition and Metabolism 5
SOCS3
0
100
200
300
400
500
600
700
800
( o
f con
trol)
lowast lowast
CONT MHF PLHF
(a)
ObRb
0
500
1000
1500
2000
2500 lowast lowast
( o
f con
trol)
CONT MHF PLHF
(b)
IRS1
0
50
100
150
200
250
300 lowastlowast
( o
f con
trol)
CONT MHF PLHF
(c)
IRS2
0
50
100
150
200lowast
( o
f con
trol)
CONT MHF PLHF
(d)
PDX-1
0
50
100
150
200
250 lowast
( o
f con
trol)
CONT MHF PLHF
(e)
0
50
100
150
200
250
300 lowast
K+ channel
CONT MHF PLHF
( o
f con
trol)
(f)
Ins1
0
50
100
150
200
250
300 lowast
lowast
CONT MHF PLHF
( o
f con
trol)
(g)
Ins2
CONT MHF PLHF0
50100150200250300350400 lowast
lowast
( o
f con
trol)
(h)
Figure 3 Pancreatic mRNA expression in adult CONT MHF and PLHF offspring SOCS3 suppressor of cytokine signaling 3 IRS insulinreceptor substrate PDX1 pancreatic and duodenal homeobox 1 ObRb full length leptin receptor Ins insulin Kir62 potassium channelsubunit Data are means plusmn SEM 119899 = 10ndash18 per group lowast119875 lt 005
6 Journal of Nutrition and Metabolism
Table 2 Correlations between SOCS3 and other key genes N = 8ndash13 per group
CONT MHF PLHFSOCS3 versus ObRb 119877
2 = 069 P = 008 R2 = 087 P = 00008 1198772 = 003 P = 064
SOCS3 versus IR R2 = 067 P = 002 1198772 = 002 P = 071 119877
2 = 0002 P = 090SOCS3 versus IRS1 R2 = 084 P = 001 R2 = 058 P = 002 119877
2 = 004 P = 051SOCS3 versus IRS2 R2 = 098 P = 0002 R2 = 058 P = 002 119877
2 = 015 P = 021SOCS3 versus Kir62 channel R2 = 082 P = 003 R2 = 093 P lt 00001 119877
2 = 0002 P = 090SOCS3 versus Pdx1 R2 = 078 P = 0009 R2 = 091 P = 00003 119877
2 = 007 P = 040
CONT MHF PLHF0
100
200
300
400
500
STAT
3 (
of c
ontro
l)
lowast
lowast
(a)
PI3K
( o
f con
trol)
CONT MHF PLHF0
50
100
150lowast
lowast
(b)
Figure 4 Pancreatic STAT3 (a) and PI3K mRNA expression (b) Data are means plusmn SEM 119899 = 10ndash18 per group STAT3 signal transducer andactivator of transcription 3 PI3K phosphatidylinositol 3-kinase lowast119875 lt 005
may be separate influences of maternal obesity during thepericonceptional period and late gestation on adiposity inoffspring [31] We have previously shown that ldquoprogrammedrdquoobesity and simple diet-induced obesity can develop viadifferent underlying mechanisms [32] The present studyfurther reinforces the importance of the timing of the dietaryinsult on the metabolic derangements that ensue in offspringduring adulthood
SOCS3 inhibition of ObRb signaling has been proposedas a mechanism for leptin resistance in a range of tissues [33ndash35] including pancreatic 120573 cells [36] It is clear that leptinresistance in PLHF pancreata was mediated by mechanismsthat did not involve SOCS3 and this has been reported previ-ously in rat models of HF feeding [37 38] SOCS3 is normallyincreased at sites of both acute and chronic inflammation [39]and has been shown to specifically suppress STAT3 activation[40] This agrees with the present study a marked increasein SOCS3 mRNA levels in MHF offspring is paralleled bySTAT3 levels similar to control offspring Leptin is knownto increase SOCS3 expression in isolated human pancreaticislets and in islets from obob mice treated in vivo [36] ourdata imply that leptin-induced SOCS3 stimulation appearsintact in MHF offspring Conversely in PLHF offspring
SOCS3 expression levels were similar to controls in thepresence of markedly elevated STAT3 expression Of notedespite similar expression levels of SOCS3 between CONTand PLHF offspring the positive correlations that wereobserved between SOCS3 and other key signaling genes wererestricted mainly to CONT and MHF offspring only Thismay point to a maladaptation in PLHF offspring due to theacute HF dietary exposure and a change in homeostatic set-points in MHF offspring due to mothers entering pregnancyin an already obese state Despite the link between SOCS3and inflammation there were no observed changes in TNF-120572 mRNA levels in the MHF group whilst there was asignificant increase in TNF-120572 expression in PLHF offspringIn addition mRNA levels of the macrophage marker CD68were increased in both HF groups and taken togethermay indicate differing macrophage phenotypespolarizationstates resulting in divergent offspring inflammatory profilesdependent upon prior maternal nutritional background Ofnote despite marked changes in inflammatorymarkers at thetissue level we observed no changes in circulating plasmaTNF-120572 concentrationHowever despite enhanced plasma IL-1120573 concentrations pancreatic IL-1120573 expression was unaltered
Journal of Nutrition and Metabolism 7
CONT
0 250 500 7500
250
500
750
PDX-1
1199032= 097
K+ch
anne
l
(a)
MHF
0 250 500 750 10000
500
1000
15001199032= 094
PDX-1
K+ch
anne
l
(b)
PLHF
0 50 100 150 200 250 3000
50
100
150
200
250
3001199032= 078
PDX-1
K+ch
anne
l
(c)
Figure 5 Relationship between PDX-1 and potassium channel subunit Kir62 in adult CONT MHF and PLHF male offspring PDX-1pancreatic and duodenal homeobox 1119873 = 8ndash10 per group
between groups implicating other tissues as possible systemicsources of this proinflammatory cytokine
PLHF offspring exhibited decreased PI3K mRNA levelscompared to CONT animals Induction of the ObRbPI3Ksignaling pathway prolongs potassium channel openingleading to decreased insulin exocytosis a direct action ofleptin which inhibits insulin secretion [41] Moreover thelowered PI3K mRNA levels in PLHF offspring were con-comitant with a reduction in IRS1 and IRS2 mRNA whosephosphorylation is coupled to PI3K activation Of notedownstreamof PI3K PDE3B gene expression levels remainedsimilar to controls suggesting that altered leptin signalingwas achieved in earlier stages of this pathway Thus attenua-tion of the ObRbPI3K signaling pathwaymaymediate leptinresistance in these hyperleptinemic PLHF offspring Sucha mechanism has been observed before in hypothalamicneurons of rats with diet-induced obesity (DIO) [42]
IRS proteins and PI3K have leptin independent rolesin the 120573 cell Both are found downstream of the insulinreceptor in the IRIRSPI3K pathway which mediates theautocrine actions of the insulin-IR complex [43 44] Ourobserved attenuation of PI3K and IRS mRNA did nothowever affect insulin synthesismdashPLHF animals were hyper-insulinemic and preproinsulin mRNA concentrations wereelevated Furthermore mRNA concentrations of IR wereunaffected potentially due to lack of SOCS3 induction inPLHF offspring similar to Controls Thus these mRNAchanges of PI3K coupled to IRS likely reflect a mechanismof leptin resistance in this group
Mean concentrations of ObRb mRNA in PLHF offspringwere similar to controls but a significant inverse relationshipbetween plasma leptin and ObRb mRNA levels in the PLHFgroup was suggestive of ObRb downregulation A downregu-lation of receptor populations in the face of elevated ligand
8 Journal of Nutrition and Metabolism
CONT MHF PLHF0
50
100
150
200
TNF-120572
mRN
A (
of c
ontro
l)
lowast
lowast
(a)
CD68
mRN
A (
of c
ontro
l)
CONT MHF PLHF0
50
100
150
200
lowast
lowast
(b)
IL1-
R1 m
RNA
( o
f con
trol)
CONT MHF PLHF0
50
100
150
200lowast
(c)
Figure 6 Pancreatic mRNA expression of TNF-120572 (a) CD68 (b) and IL1-R1 (c) in adult CONT MHF and PLHF offspring TNF-120572 tumornecrosis factor-alpha CD68 cluster of differentiation 68 IL1-R1 interleukin 1 receptor type 1 Data are means plusmn SEM 119899 = 10ndash18 per grouplowast
119875 lt 005
concentrations is not uncommon in resistant states It isseen for example in the hypothalamus in the presence ofhyperleptinemia [45ndash47] and during pregnancy which is anatural state of leptin resistance [48]
A rise in IRS1 and IRS2 mRNA associated with elevatedObRb and SOCS3 mRNA in MHF offspring may seemunexpected since SOCS3 is known to target insulin receptorsignaling by interference with IRS action [49] However sev-eral studies have demonstrated that SOCS3 mediates insulinresistance in both adipocytes and hepatocytes through post-translational modifications such as inhibition of IRS tyro-sine phosphorylation [50] and targeted ubiquitin-mediateddegradation of IRS1 and IRS2 without effecting transcription[49 51] IRS proteins are involved in both leptin signalingand insulin signaling so altered gene expression levels mayrepresent changes in either (or both) pathways although IRSresponse to leptin is reportedly much lower than that seenwith insulin [52] We observed an increase in IRS1 expressionin MHF offspring compared to Controls This upregulationof IRS1 expression may be related to the increased insulin
production of the MHF offspring given the known role ofIRS1 in the regulation of insulin secretion [53]
SOCS3 is also reported to repress preproinsulin genetranscription [41] although other mechanisms clearly coun-tered this action inMHFoffspring since Ins1 and Ins2mRNAexpression levels remained elevated This observation wasconfirmed by the significantly raised levels of circulatingplasma insulin and the lack of any correlation in levels ofmRNA between SOCS3 and Ins1 or Ins2
Some of the observed differences may relate to theinitial hyperphagia observed in the PLHF maternal groupas reported previously [14] Since rodents primarily eat ona calorific basis they will passively over-consume calorieswhen initially placed on an HF diet and self-regulate backto a normal total caloric intake over a period of a few daysIt has been shown in other rodent models that alterations innutrition around the time of implantation can lead to lastingmetabolic effects in the offspring [54] Maternal obesity hasbeen shown to exert adverse effects as early as the oocyteand preimplantation embryo stage and that these effects may
Journal of Nutrition and Metabolism 9
contribute to the lasting morbidity observed in offspring[55] Similarly even a very short period of altered nutrition(restricted to one ovulatory cycle) prior to natural mating canelicit lasting effects on the offspring [56] and this representsa further point of difference between the MHF and PLHFgroups
The present study was not designed to incorporate theeffect of a reversal paradigm whereby mothers fed thepre-conceptional HF diet were placed on a standard dietthroughout pregnancy and lactation However while there isevidence that a period of dietary restriction in obese mothersmay ablate the programming of obesity in offspring [57]it may be associated with an activation of the stress axisin the offspring [31] Further the data are inconsistent ithas also been shown that dams with diet-induced obesityfed a standard diet throughout pregnancy and lactation stilltransmit the propensity for obesity and metabolic disease totheir offspring [58]
We recognize that gene expression analysis on wholepancreatic tissue is a limitation of the present study Dueto the size of the cohort and logistical considerations onlywhole pancreatic tissue was a possibility whereas ideallyonly pure islet populations would have been examined [59]However for the key genes presented such as ObRb SOCS3and IRS expression has been shown to be primarily confinedto the endocrine islets and not been detected in the exocrinepancreas [39 41 60] therefore we are confident that theobserved changes in the genes studied represent changes inthe islets Further although we have shown that differencesin pancreatic gene expression in offspring are dependentupon the timing of maternal high fat diet exposure this doesnot itself imply causation in the development of the obesephenotypemdashother relevant pathways including liver skeletalmuscle and fat metabolism will also be integrated into theprogrammed phenotype development
This study has demonstrated enduring deleterious effectsof a maternal HF diet during early windows of develop-ment Maternal HF feeding both before conception andorthrough pregnancy and lactation induced obesity hyper-insulinemia and hyperleptinemia in adult offspring eventhough offspring were fed a standard diet postnatally Timingof exposure to the maternal HF diet differentially alteredgene expression in signaling pathways downstream of thepancreatic leptin receptor that mediated leptin resistanceThis also raises the possibility that despite common offspringobesity treatment modalities may only be efficacious in asubpopulation of treated offspring as the current data suggestthat the metabolic pathways disrupted will be dependentupon the timing of the early life dietary insult
Abbreviations
CD68 Cluster of differentiation 68CONT ControlDEXA Dual energy X-ray absorptiometryELISA Enzyme-linked immunosorbent assayGLUT2 Glucose transporter 2GP130 Glycoprotein 130HF High fat
HOMA Homeostatic model assessmentHPRT HypoxanthinephophoribosyltransferaseIL InterleukinIns InsulinIR Insulin resistanceIRS Insulin receptor substrateKir62 Potassium channel subunitLIF Leukemia inhibitory factorMHF Maternal high fatOBR Leptin receptorPDE3B Phosphodiesterase 3BPDX-1 Pancreatic and duodenal homeobox-1PI3K Phosphatidylinositide 3-kinasePLHF Pregnancy and lactation high fatSOCS3 Suppressor of cytokine signaling 3STAT3 Signal transducer and activator of
transcription 3TNF-120572 Tumor necrosis factor-alpha
Conflict of Interests
The authors have no conflict of interests with any financialorganization regarding the material discussed in this paper
Author Contributions
Mark H Vickers developed the animal model designed thestudy completed the data analysis and wrote the first draftof the paper Graham J Howie maintained the experimentalcohort performed the gene expression analysis and con-tributed to the original draft paper Clare M Reynolds per-formed the inflammatory related assays and gene expressionwork Deborah M Sloboda provided input into data analysisand interpretation
Acknowledgments
The authors acknowledge the technical support of TaniaKamal with endocrine assays The authors acknowledgefunding support from the Health Research Council of NewZealand and Gravida National Centre for Growth andDevelopment
References
[1] M M Hillemeier C S Weisman C Chuang D S Downs JMcCall-Hosenfeld and F Camacho ldquoTransition to overweightor obesity among women of reproductive agerdquo Journal ofWomenrsquos Health vol 20 no 5 pp 703ndash710 2011
[2] J E Norman and RM Reynolds ldquoThe consequences of obesityand excess weight gain in pregnancyrdquo Proceedings of the Nutri-tion Society vol 70 no 4 pp 450ndash456 2011
[3] H A Abenhaim R A Kinch L Morin A Benjamin and RUsher ldquoEffect of prepregnancy body mass index categories onobstetrical and neonatal outcomesrdquo Archives of Gynecology andObstetrics vol 275 no 1 pp 39ndash43 2007
[4] S DHMalnick andH Knobler ldquoThemedical complications ofobesityrdquo Oxford Journals Medicine vol 99 no 9 pp 565ndash5792006
10 Journal of Nutrition and Metabolism
[5] P M Catalano ldquoManagement of obesity in pregnancyrdquo Obstet-rics and Gynecology vol 109 no 2 pp 419ndash433 2007
[6] L Poston ldquoGestational weight gain influences on the long-termhealth of the childrdquo Current Opinion in Clinical Nutrition andMetabolic Care vol 15 pp 252ndash257 2012
[7] H Hochner Y Friedlander R Calderon-Margalit et al ldquoAsso-ciations of maternal prepregnancy body mass index and gesta-tional weight gain with adult offspring cardiometabolic riskfactors the Jerusalem perinatal family follow-up studyrdquo Circu-lation vol 125 no 11 pp 1381ndash1389 2012
[8] S J Spencer ldquoEarly life programming of obesity the impactof the perinatal environment on the development of obesityand metabolic dysfunction in the offspringrdquo Current DiabetesReviews vol 8 no 1 pp 55ndash68 2012
[9] H Chen D Simar K Lambert J Mercier and M J MorrisldquoMaternal and postnatal overnutrition differentially impactappetite regulators and fuel metabolismrdquo Endocrinology vol149 no 11 pp 5348ndash5356 2008
[10] P M Catalano ldquoObesity and pregnancymdashthe propagation of aviscous cyclerdquo Journal of Clinical Endocrinology and Metabo-lism vol 88 no 8 pp 3505ndash3506 2003
[11] M W Gillman S L Rifas-Shiman K Kleinman E Oken JW Rich-Edwards and EM Taveras ldquoDevelopmental origins ofchildhood overweight potential public health impactrdquo Obesityvol 16 no 7 pp 1651ndash1656 2008
[12] E Oken EM Taveras K P Kleinman JW Rich-Edwards andM W Gillman ldquoGestational weight gain and child adiposity atage 3 yearsrdquoTheAmerican Journal of Obstetrics and Gynecologyvol 196 no 4 pp 322e1ndash322e8 2007
[13] H M Ehrenberg B M Mercer and P M Catalano ldquoThe influ-ence of obesity and diabetes on the prevalence of macrosomiardquoThe American Journal of Obstetrics and Gynecology vol 191 no3 pp 964ndash968 2004
[14] G J Howie D M Sloboda T Kamal and M H VickersldquoMaternal nutritional history predicts obesity in adult offspringindependent of postnatal dietrdquo Journal of Physiology vol 587no 4 pp 905ndash915 2009
[15] D M Sloboda G J Howie A Pleasants P D Gluckman andM H Vickers ldquoPre- and postnatal nutritional histories influ-ence reproductive maturation and ovarian function in the ratrdquoPLoS ONE vol 4 no 8 Article ID e6744 2009
[16] M C Battista M F Hivert K Duval and J P BaillargeonldquoIntergenerational cycle of obesity and diabetes how can wereduce the burdens of these conditions on the health of futuregenerationsrdquo Experimental Diabetes Research vol 2011 ArticleID 596060 19 pages 2011
[17] T J Kieffer and J F Habener ldquoThe adipoinsular axis effects ofleptin on pancreatic 120573-cellsrdquo The American Journal of Physiol-ogy vol 278 no 1 pp E1ndashE14 2000
[18] S L Gray C Donald A Jetha S D Covey and T J KiefferldquoHyperinsulinemia precedes insulin resistance in mice lackingpancreatic 120573-cell leptin signalingrdquo Endocrinology vol 151 no 9pp 4178ndash4186 2010
[19] S D Covey R DWideman CMcDonald et al ldquoThe pancreat-ic 120573 cell is a key site formediating the effects of leptin on glucosehomeostasisrdquo Cell Metabolism vol 4 no 4 pp 291ndash302 2006
[20] K J Dudley DM Sloboda K L Connor J Beltrand andMHVickers ldquoOffspring ofmothers fed a high fat diet display hepaticcell cycle inhibition and associated changes in gene expressionand DNA methylationrdquo PLoS ONE vol 6 no 7 Article IDe21662 2011
[21] K L Connor M H Vickers J Beltrand M J Meaney and DM Sloboda ldquoNature nurture or nutrition Impact of maternalnutrition on maternal care offspring development and repro-ductive functionrdquo Journal of Physiology vol 590 part 9 pp2167ndash2180 2012
[22] J Beltrand D M Sloboda K L Connor M Truong and MH Vickers ldquoThe effect of neonatal leptin antagonism in malerat offspring is dependent upon the interaction between priormaternal nutritional status and post-weaning dietrdquo Journal ofNutrition andMetabolism vol 2012 Article ID 296935 10 pages2012
[23] K L Connor M H Vickers C Cupido E Sirimanne and DM Sloboda ldquoMaternal high fat diet during critical windowsof development alters adrenal cortical and medullary enzymeexpression in adult male rat offspringrdquo Journal of the Develop-mental Origins of Health and Disease vol 1 no 4 pp 245ndash2542010
[24] P J Mark C Sisala K Connor et al ldquoA maternal high-fat dietin rat pregnancy reduces growth of the fetus and the placen-tal junctional zone but not placental labyrinth zone growthrdquoJournal of the Developmental Origins of Health and Disease vol2 no 1 pp 63ndash70 2011
[25] D R Matthews J P Hosker A S Rudenski B A Naylor DF Treacher and R C Turner ldquoHomeostasis model assessmentinsulin resistance and 120573-cell function from fasting plasmaglucose and insulin concentrations in manrdquo Diabetologia vol28 no 7 pp 412ndash419 1985
[26] M P Hermans J C Levy R J Morris and R C TurnerldquoComparison of tests of120573-cell function across a range of glucosetolerance from normal to diabetesrdquo Diabetes vol 48 no 9 pp1779ndash1786 1999
[27] A B Bernal M H Vickers M B Hampton R A Poynton andD M Sloboda ldquoMaternal undernutrition significantly impactsovarian follicle number and increases ovarian oxidative stress inadult rat offspringrdquo PLoS ONE vol 5 no 12 Article ID e155582010
[28] J Vandesompele K de Preter F Pattyn et al ldquoAccurate nor-malization of real-time quantitative RT-PCR data by geometricaveraging of multiple internal control genesrdquo Genome biologyvol 3 no 7 Article ID RESEARCH0034 2002
[29] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the 2-ΔΔCT methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
[30] J Armitage L Poston and P Taylor ldquoDevelopmental originsof obesity and the metabolic syndrome the role of maternalobesityrdquo Frontiers of Hormone Research vol 36 pp 73ndash84 2007
[31] S Zhang L Rattanatray J L Morrison L M Nicholas S Lieand I C McMillen ldquoMaternal obesity and the early originsof childhood obesity weighing up the benefits and costs ofmaternal weight loss in the periconceptional period for theoffspringrdquo Experimental Diabetes Research vol 2011 Article ID585749 10 pages 2011
[32] N M Thompson A M Norman S S Donkin et al ldquoPrenataland postnatal pathways to obesity different underlying mech-anisms different metabolic outcomesrdquo Endocrinology vol 148no 5 pp 2345ndash2354 2007
[33] T Fuentes I Ara A Guadalupe-Grau et al ldquoLeptin receptor170 kDa (OB-R170) protein expression is reduced in obesehuman skeletal muscle a potential mechanism of leptin resis-tancerdquo Experimental Physiology vol 95 no 1 pp 160ndash171 2010
[34] Z Wang Y T Zhou T Kakuma et al ldquoLeptin resistance ofadipocytes in obesity role of suppressors of cytokine signalingrdquo
Journal of Nutrition and Metabolism 11
Biochemical and Biophysical Research Communications vol 277no 1 pp 20ndash26 2000
[35] C Bjoslashrbaeligk K El-Haschimi J D Frantz and J S Flier ldquoThe roleof SOCS-3 in leptin signaling and leptin resistancerdquoThe Journalof Biological Chemistry vol 274 no 42 pp 30059ndash30065 1999
[36] K Laubner T J Kieffer N T Lam X Niu F Jakob and JSeufert ldquoInhibition of preproinsulin gene expression by leptininduction of suppressor of cytokine signaling 3 in pancreatic 120573-cellsrdquo Diabetes vol 54 no 12 pp 3410ndash3417 2005
[37] C Peiser G P McGregor and R E Lang ldquoLeptin receptorexpression and suppressor of cytokine signaling transcriptlevels in high-fat-fed ratsrdquo Life Sciences vol 67 no 24 pp 2971ndash2981 2000
[38] G R Steinberg A C Smith S Wormald P Malenfant CCollier and D J Dyck ldquoEndurance training partially reversesdietary-induced leptin resistance in rodent skeletal musclerdquoTheAmerican Journal of Physiology vol 286 no 1 pp E57ndashE632004
[39] G E White A Cotterill M R Addley E J Soilleux and DR Greaves ldquoSuppressor of cytokine signalling protein SOCS3expression is increased at sites of acute and chronic inflamma-tionrdquo Journal of Molecular Histology vol 42 no 2 pp 137ndash1512011
[40] H YasukawaMOhishiHMori et al ldquoIL-6 induces an anti-in-flammatory response in the absence of SOCS3 inmacrophagesrdquoNature Immunology vol 4 no 6 pp 551ndash556 2003
[41] J Seufert ldquoLeptin effects on pancreatic 120573-cell gene expressionand functionrdquo Diabetes vol 53 no 1 pp S152ndashS158 2004
[42] A S Metlakunta M Sahu and A Sahu ldquoHypothalamicphosphatidylinositol 3-kinase pathway of leptin signaling isimpaired during the development of diet-induced obesity inFVBNmicerdquo Endocrinology vol 149 no 3 pp 1121ndash1128 2008
[43] C A Aspinwall W J Qian M G Roper R N Kulkarni C RKahn and R T Kennedy ldquoRoles of insulin receptor substrate-1phosphatidylinositol 3-kinase and release of intracellular Ca2+stores in insulin-stimulated insulin secretion in 120573-cellsrdquo TheJournal of Biological Chemistry vol 275 no 29 pp 22331ndash223382000
[44] I B Leibiger B Leibiger T Moede and P O BerggrenldquoExocytosis of insulin promotes insulin gene transcription viathe insulin receptorPI-3 kinasep70 s6 kinase and CaM kinasepathwaysrdquoMolecular Cell vol 1 no 6 pp 933ndash938 1998
[45] R L Martin E Perez Y J He R Dawson Jr andW J MillardldquoLeptin resistance is associated with hypothalamic leptin recep-tor mRNA and protein downregulationrdquo Metabolism vol 49no 11 pp 1479ndash1484 2000
[46] J Wilsey and P J Scarpace ldquoCaloric restriction reverses thedeficits in leptin receptor protein and leptin signaling capacityassociated with diet-induced obesity role of leptin in the reg-ulation of hypothalamic long-form leptin receptor expressionrdquoJournal of Endocrinology vol 181 no 2 pp 297ndash306 2004
[47] Y Zhang and P J Scarpace ldquoThe role of leptin in leptin resist-ance and obesityrdquo Physiology and Behavior vol 88 no 3 pp249ndash256 2006
[48] D R Grattan S R Ladyman and R A Augustine ldquoHormonalinduction of leptin resistance during pregnancyrdquoPhysiology andBehavior vol 91 no 4 pp 366ndash374 2007
[49] L RuiM YuanD Frantz S Shoelson andM FWhite ldquoSOCS-1 and SOCS-3 block insulin signaling by ubiquitin-mediateddegradation of IRS1 and IRS2rdquoThe Journal of Biological Chem-istry vol 277 no 44 pp 42394ndash42398 2002
[50] K Ueki T Kondo and C R Kahn ldquoSuppressor of cytokine sig-naling 1 (SOCS-1) and SOCS-3 cause insulin resistance throughinhibition of tyrosine phosphorylation of insulin receptor sub-strate proteins by discrete mechanismsrdquoMolecular and CellularBiology vol 24 no 12 pp 5434ndash5446 2004
[51] S B Jorgensen O rsquoNeill HM L Sylow et al ldquoDeletion of skel-etal muscle SOCS3 prevents insulin resistance in obesityrdquo Dia-betes vol 62 no 1 pp 56ndash64 2013
[52] W Becker ldquoLeptin signal transductionrdquo in Leptin and LeptinAntagonists A Gertler Ed pp 1ndash9 Landes Bioscience AustinTex USA 2009
[53] R N Kulkarni J N Winnay M Daniels et al ldquoAltered func-tion of insulin receptor substrate 1-deficient mouse islets andcultured 120573-cell linesrdquo Journal of Clinical Investigation vol 104no 12 pp R69ndashR75 1999
[54] W Y Kwong A E Wild P Roberts A C Willis and T PFleming ldquoMaternal undernutrition during the preimplantationperiod of rat development causes blastocyst abnormalities andprogramming of postnatal hypertensionrdquoDevelopment vol 127no 19 pp 4195ndash4202 2000
[55] E S Jungheim E L Schoeller K L Marquard E D LoudenJ E Schaffer and K H Moley ldquoDiet-induced obesity modelabnormal oocytes and persistent growth abnormalities in theoffspringrdquo Endocrinology vol 151 no 8 pp 4039ndash4046 2010
[56] A J Watkins A Wilkins C Cunningham et al ldquoLow proteindiet fed exclusively during mouse oocyte maturation leadsto behavioural and cardiovascular abnormalities in offspringrdquoJournal of Physiology vol 586 no 8 pp 2231ndash2244 2008
[57] E Zambrano P M Martınez-Samayoa G L Rodrıguez-Gonzalez and P W Nathanielsz ldquoDietary intervention prior topregnancy reverses metabolic programming in male offspringof obese ratsrdquo Journal of Physiology vol 588 no 10 pp 1791ndash1799 2010
[58] M S Patel andM Srinivasan ldquoMetabolic programming causesand consequencesrdquoThe Journal of Biological Chemistry vol 277no 3 pp 1629ndash1632 2002
[59] R A Simmons ldquoRole of metabolic programming in the patho-genesis of 120573-cell failure in postnatal liferdquo Reviews in Endocrineand Metabolic Disorders vol 8 no 2 pp 95ndash104 2007
[60] I B Leibiger B Leibiger and P O Berggren ldquoInsulin signalingin the pancreatic 120573-cellrdquoAnnual Review of Nutrition vol 28 pp233ndash251 2008
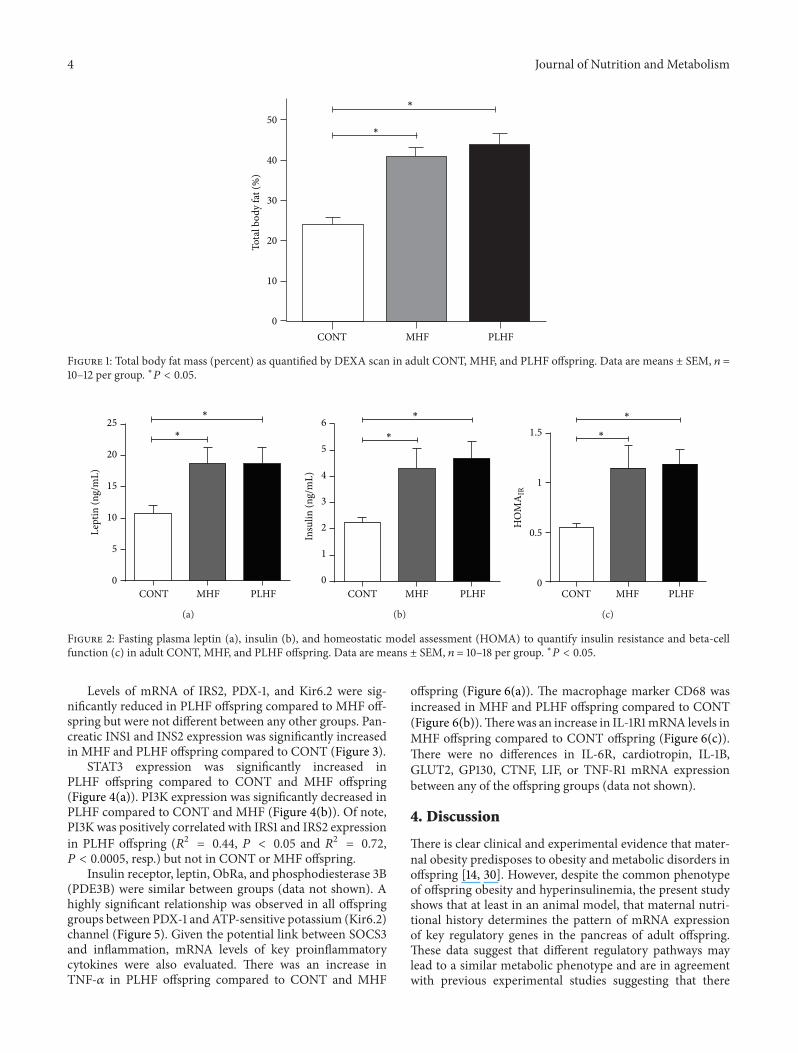
4 Journal of Nutrition and Metabolism
CONT MHF PLHF
50
40
30
20
10
0
lowast
lowast
Tota
l bod
y fa
t (
)
Figure 1 Total body fat mass (percent) as quantified by DEXA scan in adult CONT MHF and PLHF offspring Data are means plusmn SEM 119899 =10ndash12 per group lowast119875 lt 005
CONT MHF PLHF0
5
10
15
20
25lowast
lowast
Lept
in (n
gm
L)
(a)
CONT MHF PLHF0
1
2
3
4
5
6lowast
lowast
Insu
lin (n
gm
L)
(b)
CONT MHF PLHF0
05
1
15 lowast
lowast
HO
MA
IR
(c)
Figure 2 Fasting plasma leptin (a) insulin (b) and homeostatic model assessment (HOMA) to quantify insulin resistance and beta-cellfunction (c) in adult CONT MHF and PLHF offspring Data are means plusmn SEM 119899 = 10ndash18 per group lowast119875 lt 005
Levels of mRNA of IRS2 PDX-1 and Kir62 were sig-nificantly reduced in PLHF offspring compared to MHF off-spring but were not different between any other groups Pan-creatic INS1 and INS2 expression was significantly increasedin MHF and PLHF offspring compared to CONT (Figure 3)
STAT3 expression was significantly increased inPLHF offspring compared to CONT and MHF offspring(Figure 4(a)) PI3K expression was significantly decreased inPLHF compared to CONT and MHF (Figure 4(b)) Of notePI3K was positively correlated with IRS1 and IRS2 expressionin PLHF offspring (1198772 = 044 119875 lt 005 and 1198772 = 072119875 lt 00005 resp) but not in CONT or MHF offspring
Insulin receptor leptin ObRa and phosphodiesterase 3B(PDE3B) were similar between groups (data not shown) Ahighly significant relationship was observed in all offspringgroups between PDX-1 and ATP-sensitive potassium (Kir62)channel (Figure 5) Given the potential link between SOCS3and inflammation mRNA levels of key proinflammatorycytokines were also evaluated There was an increase inTNF-120572 in PLHF offspring compared to CONT and MHF
offspring (Figure 6(a)) The macrophage marker CD68 wasincreased in MHF and PLHF offspring compared to CONT(Figure 6(b))Therewas an increase in IL-1R1mRNA levels inMHF offspring compared to CONT offspring (Figure 6(c))There were no differences in IL-6R cardiotropin IL-1BGLUT2 GP130 CTNF LIF or TNF-R1 mRNA expressionbetween any of the offspring groups (data not shown)
4 Discussion
There is clear clinical and experimental evidence that mater-nal obesity predisposes to obesity and metabolic disorders inoffspring [14 30] However despite the common phenotypeof offspring obesity and hyperinsulinemia the present studyshows that at least in an animal model that maternal nutri-tional history determines the pattern of mRNA expressionof key regulatory genes in the pancreas of adult offspringThese data suggest that different regulatory pathways maylead to a similar metabolic phenotype and are in agreementwith previous experimental studies suggesting that there
Journal of Nutrition and Metabolism 5
SOCS3
0
100
200
300
400
500
600
700
800
( o
f con
trol)
lowast lowast
CONT MHF PLHF
(a)
ObRb
0
500
1000
1500
2000
2500 lowast lowast
( o
f con
trol)
CONT MHF PLHF
(b)
IRS1
0
50
100
150
200
250
300 lowastlowast
( o
f con
trol)
CONT MHF PLHF
(c)
IRS2
0
50
100
150
200lowast
( o
f con
trol)
CONT MHF PLHF
(d)
PDX-1
0
50
100
150
200
250 lowast
( o
f con
trol)
CONT MHF PLHF
(e)
0
50
100
150
200
250
300 lowast
K+ channel
CONT MHF PLHF
( o
f con
trol)
(f)
Ins1
0
50
100
150
200
250
300 lowast
lowast
CONT MHF PLHF
( o
f con
trol)
(g)
Ins2
CONT MHF PLHF0
50100150200250300350400 lowast
lowast
( o
f con
trol)
(h)
Figure 3 Pancreatic mRNA expression in adult CONT MHF and PLHF offspring SOCS3 suppressor of cytokine signaling 3 IRS insulinreceptor substrate PDX1 pancreatic and duodenal homeobox 1 ObRb full length leptin receptor Ins insulin Kir62 potassium channelsubunit Data are means plusmn SEM 119899 = 10ndash18 per group lowast119875 lt 005
6 Journal of Nutrition and Metabolism
Table 2 Correlations between SOCS3 and other key genes N = 8ndash13 per group
CONT MHF PLHFSOCS3 versus ObRb 119877
2 = 069 P = 008 R2 = 087 P = 00008 1198772 = 003 P = 064
SOCS3 versus IR R2 = 067 P = 002 1198772 = 002 P = 071 119877
2 = 0002 P = 090SOCS3 versus IRS1 R2 = 084 P = 001 R2 = 058 P = 002 119877
2 = 004 P = 051SOCS3 versus IRS2 R2 = 098 P = 0002 R2 = 058 P = 002 119877
2 = 015 P = 021SOCS3 versus Kir62 channel R2 = 082 P = 003 R2 = 093 P lt 00001 119877
2 = 0002 P = 090SOCS3 versus Pdx1 R2 = 078 P = 0009 R2 = 091 P = 00003 119877
2 = 007 P = 040
CONT MHF PLHF0
100
200
300
400
500
STAT
3 (
of c
ontro
l)
lowast
lowast
(a)
PI3K
( o
f con
trol)
CONT MHF PLHF0
50
100
150lowast
lowast
(b)
Figure 4 Pancreatic STAT3 (a) and PI3K mRNA expression (b) Data are means plusmn SEM 119899 = 10ndash18 per group STAT3 signal transducer andactivator of transcription 3 PI3K phosphatidylinositol 3-kinase lowast119875 lt 005
may be separate influences of maternal obesity during thepericonceptional period and late gestation on adiposity inoffspring [31] We have previously shown that ldquoprogrammedrdquoobesity and simple diet-induced obesity can develop viadifferent underlying mechanisms [32] The present studyfurther reinforces the importance of the timing of the dietaryinsult on the metabolic derangements that ensue in offspringduring adulthood
SOCS3 inhibition of ObRb signaling has been proposedas a mechanism for leptin resistance in a range of tissues [33ndash35] including pancreatic 120573 cells [36] It is clear that leptinresistance in PLHF pancreata was mediated by mechanismsthat did not involve SOCS3 and this has been reported previ-ously in rat models of HF feeding [37 38] SOCS3 is normallyincreased at sites of both acute and chronic inflammation [39]and has been shown to specifically suppress STAT3 activation[40] This agrees with the present study a marked increasein SOCS3 mRNA levels in MHF offspring is paralleled bySTAT3 levels similar to control offspring Leptin is knownto increase SOCS3 expression in isolated human pancreaticislets and in islets from obob mice treated in vivo [36] ourdata imply that leptin-induced SOCS3 stimulation appearsintact in MHF offspring Conversely in PLHF offspring
SOCS3 expression levels were similar to controls in thepresence of markedly elevated STAT3 expression Of notedespite similar expression levels of SOCS3 between CONTand PLHF offspring the positive correlations that wereobserved between SOCS3 and other key signaling genes wererestricted mainly to CONT and MHF offspring only Thismay point to a maladaptation in PLHF offspring due to theacute HF dietary exposure and a change in homeostatic set-points in MHF offspring due to mothers entering pregnancyin an already obese state Despite the link between SOCS3and inflammation there were no observed changes in TNF-120572 mRNA levels in the MHF group whilst there was asignificant increase in TNF-120572 expression in PLHF offspringIn addition mRNA levels of the macrophage marker CD68were increased in both HF groups and taken togethermay indicate differing macrophage phenotypespolarizationstates resulting in divergent offspring inflammatory profilesdependent upon prior maternal nutritional background Ofnote despite marked changes in inflammatorymarkers at thetissue level we observed no changes in circulating plasmaTNF-120572 concentrationHowever despite enhanced plasma IL-1120573 concentrations pancreatic IL-1120573 expression was unaltered
Journal of Nutrition and Metabolism 7
CONT
0 250 500 7500
250
500
750
PDX-1
1199032= 097
K+ch
anne
l
(a)
MHF
0 250 500 750 10000
500
1000
15001199032= 094
PDX-1
K+ch
anne
l
(b)
PLHF
0 50 100 150 200 250 3000
50
100
150
200
250
3001199032= 078
PDX-1
K+ch
anne
l
(c)
Figure 5 Relationship between PDX-1 and potassium channel subunit Kir62 in adult CONT MHF and PLHF male offspring PDX-1pancreatic and duodenal homeobox 1119873 = 8ndash10 per group
between groups implicating other tissues as possible systemicsources of this proinflammatory cytokine
PLHF offspring exhibited decreased PI3K mRNA levelscompared to CONT animals Induction of the ObRbPI3Ksignaling pathway prolongs potassium channel openingleading to decreased insulin exocytosis a direct action ofleptin which inhibits insulin secretion [41] Moreover thelowered PI3K mRNA levels in PLHF offspring were con-comitant with a reduction in IRS1 and IRS2 mRNA whosephosphorylation is coupled to PI3K activation Of notedownstreamof PI3K PDE3B gene expression levels remainedsimilar to controls suggesting that altered leptin signalingwas achieved in earlier stages of this pathway Thus attenua-tion of the ObRbPI3K signaling pathwaymaymediate leptinresistance in these hyperleptinemic PLHF offspring Sucha mechanism has been observed before in hypothalamicneurons of rats with diet-induced obesity (DIO) [42]
IRS proteins and PI3K have leptin independent rolesin the 120573 cell Both are found downstream of the insulinreceptor in the IRIRSPI3K pathway which mediates theautocrine actions of the insulin-IR complex [43 44] Ourobserved attenuation of PI3K and IRS mRNA did nothowever affect insulin synthesismdashPLHF animals were hyper-insulinemic and preproinsulin mRNA concentrations wereelevated Furthermore mRNA concentrations of IR wereunaffected potentially due to lack of SOCS3 induction inPLHF offspring similar to Controls Thus these mRNAchanges of PI3K coupled to IRS likely reflect a mechanismof leptin resistance in this group
Mean concentrations of ObRb mRNA in PLHF offspringwere similar to controls but a significant inverse relationshipbetween plasma leptin and ObRb mRNA levels in the PLHFgroup was suggestive of ObRb downregulation A downregu-lation of receptor populations in the face of elevated ligand
8 Journal of Nutrition and Metabolism
CONT MHF PLHF0
50
100
150
200
TNF-120572
mRN
A (
of c
ontro
l)
lowast
lowast
(a)
CD68
mRN
A (
of c
ontro
l)
CONT MHF PLHF0
50
100
150
200
lowast
lowast
(b)
IL1-
R1 m
RNA
( o
f con
trol)
CONT MHF PLHF0
50
100
150
200lowast
(c)
Figure 6 Pancreatic mRNA expression of TNF-120572 (a) CD68 (b) and IL1-R1 (c) in adult CONT MHF and PLHF offspring TNF-120572 tumornecrosis factor-alpha CD68 cluster of differentiation 68 IL1-R1 interleukin 1 receptor type 1 Data are means plusmn SEM 119899 = 10ndash18 per grouplowast
119875 lt 005
concentrations is not uncommon in resistant states It isseen for example in the hypothalamus in the presence ofhyperleptinemia [45ndash47] and during pregnancy which is anatural state of leptin resistance [48]
A rise in IRS1 and IRS2 mRNA associated with elevatedObRb and SOCS3 mRNA in MHF offspring may seemunexpected since SOCS3 is known to target insulin receptorsignaling by interference with IRS action [49] However sev-eral studies have demonstrated that SOCS3 mediates insulinresistance in both adipocytes and hepatocytes through post-translational modifications such as inhibition of IRS tyro-sine phosphorylation [50] and targeted ubiquitin-mediateddegradation of IRS1 and IRS2 without effecting transcription[49 51] IRS proteins are involved in both leptin signalingand insulin signaling so altered gene expression levels mayrepresent changes in either (or both) pathways although IRSresponse to leptin is reportedly much lower than that seenwith insulin [52] We observed an increase in IRS1 expressionin MHF offspring compared to Controls This upregulationof IRS1 expression may be related to the increased insulin
production of the MHF offspring given the known role ofIRS1 in the regulation of insulin secretion [53]
SOCS3 is also reported to repress preproinsulin genetranscription [41] although other mechanisms clearly coun-tered this action inMHFoffspring since Ins1 and Ins2mRNAexpression levels remained elevated This observation wasconfirmed by the significantly raised levels of circulatingplasma insulin and the lack of any correlation in levels ofmRNA between SOCS3 and Ins1 or Ins2
Some of the observed differences may relate to theinitial hyperphagia observed in the PLHF maternal groupas reported previously [14] Since rodents primarily eat ona calorific basis they will passively over-consume calorieswhen initially placed on an HF diet and self-regulate backto a normal total caloric intake over a period of a few daysIt has been shown in other rodent models that alterations innutrition around the time of implantation can lead to lastingmetabolic effects in the offspring [54] Maternal obesity hasbeen shown to exert adverse effects as early as the oocyteand preimplantation embryo stage and that these effects may
Journal of Nutrition and Metabolism 9
contribute to the lasting morbidity observed in offspring[55] Similarly even a very short period of altered nutrition(restricted to one ovulatory cycle) prior to natural mating canelicit lasting effects on the offspring [56] and this representsa further point of difference between the MHF and PLHFgroups
The present study was not designed to incorporate theeffect of a reversal paradigm whereby mothers fed thepre-conceptional HF diet were placed on a standard dietthroughout pregnancy and lactation However while there isevidence that a period of dietary restriction in obese mothersmay ablate the programming of obesity in offspring [57]it may be associated with an activation of the stress axisin the offspring [31] Further the data are inconsistent ithas also been shown that dams with diet-induced obesityfed a standard diet throughout pregnancy and lactation stilltransmit the propensity for obesity and metabolic disease totheir offspring [58]
We recognize that gene expression analysis on wholepancreatic tissue is a limitation of the present study Dueto the size of the cohort and logistical considerations onlywhole pancreatic tissue was a possibility whereas ideallyonly pure islet populations would have been examined [59]However for the key genes presented such as ObRb SOCS3and IRS expression has been shown to be primarily confinedto the endocrine islets and not been detected in the exocrinepancreas [39 41 60] therefore we are confident that theobserved changes in the genes studied represent changes inthe islets Further although we have shown that differencesin pancreatic gene expression in offspring are dependentupon the timing of maternal high fat diet exposure this doesnot itself imply causation in the development of the obesephenotypemdashother relevant pathways including liver skeletalmuscle and fat metabolism will also be integrated into theprogrammed phenotype development
This study has demonstrated enduring deleterious effectsof a maternal HF diet during early windows of develop-ment Maternal HF feeding both before conception andorthrough pregnancy and lactation induced obesity hyper-insulinemia and hyperleptinemia in adult offspring eventhough offspring were fed a standard diet postnatally Timingof exposure to the maternal HF diet differentially alteredgene expression in signaling pathways downstream of thepancreatic leptin receptor that mediated leptin resistanceThis also raises the possibility that despite common offspringobesity treatment modalities may only be efficacious in asubpopulation of treated offspring as the current data suggestthat the metabolic pathways disrupted will be dependentupon the timing of the early life dietary insult
Abbreviations
CD68 Cluster of differentiation 68CONT ControlDEXA Dual energy X-ray absorptiometryELISA Enzyme-linked immunosorbent assayGLUT2 Glucose transporter 2GP130 Glycoprotein 130HF High fat
HOMA Homeostatic model assessmentHPRT HypoxanthinephophoribosyltransferaseIL InterleukinIns InsulinIR Insulin resistanceIRS Insulin receptor substrateKir62 Potassium channel subunitLIF Leukemia inhibitory factorMHF Maternal high fatOBR Leptin receptorPDE3B Phosphodiesterase 3BPDX-1 Pancreatic and duodenal homeobox-1PI3K Phosphatidylinositide 3-kinasePLHF Pregnancy and lactation high fatSOCS3 Suppressor of cytokine signaling 3STAT3 Signal transducer and activator of
transcription 3TNF-120572 Tumor necrosis factor-alpha
Conflict of Interests
The authors have no conflict of interests with any financialorganization regarding the material discussed in this paper
Author Contributions
Mark H Vickers developed the animal model designed thestudy completed the data analysis and wrote the first draftof the paper Graham J Howie maintained the experimentalcohort performed the gene expression analysis and con-tributed to the original draft paper Clare M Reynolds per-formed the inflammatory related assays and gene expressionwork Deborah M Sloboda provided input into data analysisand interpretation
Acknowledgments
The authors acknowledge the technical support of TaniaKamal with endocrine assays The authors acknowledgefunding support from the Health Research Council of NewZealand and Gravida National Centre for Growth andDevelopment
References
[1] M M Hillemeier C S Weisman C Chuang D S Downs JMcCall-Hosenfeld and F Camacho ldquoTransition to overweightor obesity among women of reproductive agerdquo Journal ofWomenrsquos Health vol 20 no 5 pp 703ndash710 2011
[2] J E Norman and RM Reynolds ldquoThe consequences of obesityand excess weight gain in pregnancyrdquo Proceedings of the Nutri-tion Society vol 70 no 4 pp 450ndash456 2011
[3] H A Abenhaim R A Kinch L Morin A Benjamin and RUsher ldquoEffect of prepregnancy body mass index categories onobstetrical and neonatal outcomesrdquo Archives of Gynecology andObstetrics vol 275 no 1 pp 39ndash43 2007
[4] S DHMalnick andH Knobler ldquoThemedical complications ofobesityrdquo Oxford Journals Medicine vol 99 no 9 pp 565ndash5792006
10 Journal of Nutrition and Metabolism
[5] P M Catalano ldquoManagement of obesity in pregnancyrdquo Obstet-rics and Gynecology vol 109 no 2 pp 419ndash433 2007
[6] L Poston ldquoGestational weight gain influences on the long-termhealth of the childrdquo Current Opinion in Clinical Nutrition andMetabolic Care vol 15 pp 252ndash257 2012
[7] H Hochner Y Friedlander R Calderon-Margalit et al ldquoAsso-ciations of maternal prepregnancy body mass index and gesta-tional weight gain with adult offspring cardiometabolic riskfactors the Jerusalem perinatal family follow-up studyrdquo Circu-lation vol 125 no 11 pp 1381ndash1389 2012
[8] S J Spencer ldquoEarly life programming of obesity the impactof the perinatal environment on the development of obesityand metabolic dysfunction in the offspringrdquo Current DiabetesReviews vol 8 no 1 pp 55ndash68 2012
[9] H Chen D Simar K Lambert J Mercier and M J MorrisldquoMaternal and postnatal overnutrition differentially impactappetite regulators and fuel metabolismrdquo Endocrinology vol149 no 11 pp 5348ndash5356 2008
[10] P M Catalano ldquoObesity and pregnancymdashthe propagation of aviscous cyclerdquo Journal of Clinical Endocrinology and Metabo-lism vol 88 no 8 pp 3505ndash3506 2003
[11] M W Gillman S L Rifas-Shiman K Kleinman E Oken JW Rich-Edwards and EM Taveras ldquoDevelopmental origins ofchildhood overweight potential public health impactrdquo Obesityvol 16 no 7 pp 1651ndash1656 2008
[12] E Oken EM Taveras K P Kleinman JW Rich-Edwards andM W Gillman ldquoGestational weight gain and child adiposity atage 3 yearsrdquoTheAmerican Journal of Obstetrics and Gynecologyvol 196 no 4 pp 322e1ndash322e8 2007
[13] H M Ehrenberg B M Mercer and P M Catalano ldquoThe influ-ence of obesity and diabetes on the prevalence of macrosomiardquoThe American Journal of Obstetrics and Gynecology vol 191 no3 pp 964ndash968 2004
[14] G J Howie D M Sloboda T Kamal and M H VickersldquoMaternal nutritional history predicts obesity in adult offspringindependent of postnatal dietrdquo Journal of Physiology vol 587no 4 pp 905ndash915 2009
[15] D M Sloboda G J Howie A Pleasants P D Gluckman andM H Vickers ldquoPre- and postnatal nutritional histories influ-ence reproductive maturation and ovarian function in the ratrdquoPLoS ONE vol 4 no 8 Article ID e6744 2009
[16] M C Battista M F Hivert K Duval and J P BaillargeonldquoIntergenerational cycle of obesity and diabetes how can wereduce the burdens of these conditions on the health of futuregenerationsrdquo Experimental Diabetes Research vol 2011 ArticleID 596060 19 pages 2011
[17] T J Kieffer and J F Habener ldquoThe adipoinsular axis effects ofleptin on pancreatic 120573-cellsrdquo The American Journal of Physiol-ogy vol 278 no 1 pp E1ndashE14 2000
[18] S L Gray C Donald A Jetha S D Covey and T J KiefferldquoHyperinsulinemia precedes insulin resistance in mice lackingpancreatic 120573-cell leptin signalingrdquo Endocrinology vol 151 no 9pp 4178ndash4186 2010
[19] S D Covey R DWideman CMcDonald et al ldquoThe pancreat-ic 120573 cell is a key site formediating the effects of leptin on glucosehomeostasisrdquo Cell Metabolism vol 4 no 4 pp 291ndash302 2006
[20] K J Dudley DM Sloboda K L Connor J Beltrand andMHVickers ldquoOffspring ofmothers fed a high fat diet display hepaticcell cycle inhibition and associated changes in gene expressionand DNA methylationrdquo PLoS ONE vol 6 no 7 Article IDe21662 2011
[21] K L Connor M H Vickers J Beltrand M J Meaney and DM Sloboda ldquoNature nurture or nutrition Impact of maternalnutrition on maternal care offspring development and repro-ductive functionrdquo Journal of Physiology vol 590 part 9 pp2167ndash2180 2012
[22] J Beltrand D M Sloboda K L Connor M Truong and MH Vickers ldquoThe effect of neonatal leptin antagonism in malerat offspring is dependent upon the interaction between priormaternal nutritional status and post-weaning dietrdquo Journal ofNutrition andMetabolism vol 2012 Article ID 296935 10 pages2012
[23] K L Connor M H Vickers C Cupido E Sirimanne and DM Sloboda ldquoMaternal high fat diet during critical windowsof development alters adrenal cortical and medullary enzymeexpression in adult male rat offspringrdquo Journal of the Develop-mental Origins of Health and Disease vol 1 no 4 pp 245ndash2542010
[24] P J Mark C Sisala K Connor et al ldquoA maternal high-fat dietin rat pregnancy reduces growth of the fetus and the placen-tal junctional zone but not placental labyrinth zone growthrdquoJournal of the Developmental Origins of Health and Disease vol2 no 1 pp 63ndash70 2011
[25] D R Matthews J P Hosker A S Rudenski B A Naylor DF Treacher and R C Turner ldquoHomeostasis model assessmentinsulin resistance and 120573-cell function from fasting plasmaglucose and insulin concentrations in manrdquo Diabetologia vol28 no 7 pp 412ndash419 1985
[26] M P Hermans J C Levy R J Morris and R C TurnerldquoComparison of tests of120573-cell function across a range of glucosetolerance from normal to diabetesrdquo Diabetes vol 48 no 9 pp1779ndash1786 1999
[27] A B Bernal M H Vickers M B Hampton R A Poynton andD M Sloboda ldquoMaternal undernutrition significantly impactsovarian follicle number and increases ovarian oxidative stress inadult rat offspringrdquo PLoS ONE vol 5 no 12 Article ID e155582010
[28] J Vandesompele K de Preter F Pattyn et al ldquoAccurate nor-malization of real-time quantitative RT-PCR data by geometricaveraging of multiple internal control genesrdquo Genome biologyvol 3 no 7 Article ID RESEARCH0034 2002
[29] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the 2-ΔΔCT methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
[30] J Armitage L Poston and P Taylor ldquoDevelopmental originsof obesity and the metabolic syndrome the role of maternalobesityrdquo Frontiers of Hormone Research vol 36 pp 73ndash84 2007
[31] S Zhang L Rattanatray J L Morrison L M Nicholas S Lieand I C McMillen ldquoMaternal obesity and the early originsof childhood obesity weighing up the benefits and costs ofmaternal weight loss in the periconceptional period for theoffspringrdquo Experimental Diabetes Research vol 2011 Article ID585749 10 pages 2011
[32] N M Thompson A M Norman S S Donkin et al ldquoPrenataland postnatal pathways to obesity different underlying mech-anisms different metabolic outcomesrdquo Endocrinology vol 148no 5 pp 2345ndash2354 2007
[33] T Fuentes I Ara A Guadalupe-Grau et al ldquoLeptin receptor170 kDa (OB-R170) protein expression is reduced in obesehuman skeletal muscle a potential mechanism of leptin resis-tancerdquo Experimental Physiology vol 95 no 1 pp 160ndash171 2010
[34] Z Wang Y T Zhou T Kakuma et al ldquoLeptin resistance ofadipocytes in obesity role of suppressors of cytokine signalingrdquo
Journal of Nutrition and Metabolism 11
Biochemical and Biophysical Research Communications vol 277no 1 pp 20ndash26 2000
[35] C Bjoslashrbaeligk K El-Haschimi J D Frantz and J S Flier ldquoThe roleof SOCS-3 in leptin signaling and leptin resistancerdquoThe Journalof Biological Chemistry vol 274 no 42 pp 30059ndash30065 1999
[36] K Laubner T J Kieffer N T Lam X Niu F Jakob and JSeufert ldquoInhibition of preproinsulin gene expression by leptininduction of suppressor of cytokine signaling 3 in pancreatic 120573-cellsrdquo Diabetes vol 54 no 12 pp 3410ndash3417 2005
[37] C Peiser G P McGregor and R E Lang ldquoLeptin receptorexpression and suppressor of cytokine signaling transcriptlevels in high-fat-fed ratsrdquo Life Sciences vol 67 no 24 pp 2971ndash2981 2000
[38] G R Steinberg A C Smith S Wormald P Malenfant CCollier and D J Dyck ldquoEndurance training partially reversesdietary-induced leptin resistance in rodent skeletal musclerdquoTheAmerican Journal of Physiology vol 286 no 1 pp E57ndashE632004
[39] G E White A Cotterill M R Addley E J Soilleux and DR Greaves ldquoSuppressor of cytokine signalling protein SOCS3expression is increased at sites of acute and chronic inflamma-tionrdquo Journal of Molecular Histology vol 42 no 2 pp 137ndash1512011
[40] H YasukawaMOhishiHMori et al ldquoIL-6 induces an anti-in-flammatory response in the absence of SOCS3 inmacrophagesrdquoNature Immunology vol 4 no 6 pp 551ndash556 2003
[41] J Seufert ldquoLeptin effects on pancreatic 120573-cell gene expressionand functionrdquo Diabetes vol 53 no 1 pp S152ndashS158 2004
[42] A S Metlakunta M Sahu and A Sahu ldquoHypothalamicphosphatidylinositol 3-kinase pathway of leptin signaling isimpaired during the development of diet-induced obesity inFVBNmicerdquo Endocrinology vol 149 no 3 pp 1121ndash1128 2008
[43] C A Aspinwall W J Qian M G Roper R N Kulkarni C RKahn and R T Kennedy ldquoRoles of insulin receptor substrate-1phosphatidylinositol 3-kinase and release of intracellular Ca2+stores in insulin-stimulated insulin secretion in 120573-cellsrdquo TheJournal of Biological Chemistry vol 275 no 29 pp 22331ndash223382000
[44] I B Leibiger B Leibiger T Moede and P O BerggrenldquoExocytosis of insulin promotes insulin gene transcription viathe insulin receptorPI-3 kinasep70 s6 kinase and CaM kinasepathwaysrdquoMolecular Cell vol 1 no 6 pp 933ndash938 1998
[45] R L Martin E Perez Y J He R Dawson Jr andW J MillardldquoLeptin resistance is associated with hypothalamic leptin recep-tor mRNA and protein downregulationrdquo Metabolism vol 49no 11 pp 1479ndash1484 2000
[46] J Wilsey and P J Scarpace ldquoCaloric restriction reverses thedeficits in leptin receptor protein and leptin signaling capacityassociated with diet-induced obesity role of leptin in the reg-ulation of hypothalamic long-form leptin receptor expressionrdquoJournal of Endocrinology vol 181 no 2 pp 297ndash306 2004
[47] Y Zhang and P J Scarpace ldquoThe role of leptin in leptin resist-ance and obesityrdquo Physiology and Behavior vol 88 no 3 pp249ndash256 2006
[48] D R Grattan S R Ladyman and R A Augustine ldquoHormonalinduction of leptin resistance during pregnancyrdquoPhysiology andBehavior vol 91 no 4 pp 366ndash374 2007
[49] L RuiM YuanD Frantz S Shoelson andM FWhite ldquoSOCS-1 and SOCS-3 block insulin signaling by ubiquitin-mediateddegradation of IRS1 and IRS2rdquoThe Journal of Biological Chem-istry vol 277 no 44 pp 42394ndash42398 2002
[50] K Ueki T Kondo and C R Kahn ldquoSuppressor of cytokine sig-naling 1 (SOCS-1) and SOCS-3 cause insulin resistance throughinhibition of tyrosine phosphorylation of insulin receptor sub-strate proteins by discrete mechanismsrdquoMolecular and CellularBiology vol 24 no 12 pp 5434ndash5446 2004
[51] S B Jorgensen O rsquoNeill HM L Sylow et al ldquoDeletion of skel-etal muscle SOCS3 prevents insulin resistance in obesityrdquo Dia-betes vol 62 no 1 pp 56ndash64 2013
[52] W Becker ldquoLeptin signal transductionrdquo in Leptin and LeptinAntagonists A Gertler Ed pp 1ndash9 Landes Bioscience AustinTex USA 2009
[53] R N Kulkarni J N Winnay M Daniels et al ldquoAltered func-tion of insulin receptor substrate 1-deficient mouse islets andcultured 120573-cell linesrdquo Journal of Clinical Investigation vol 104no 12 pp R69ndashR75 1999
[54] W Y Kwong A E Wild P Roberts A C Willis and T PFleming ldquoMaternal undernutrition during the preimplantationperiod of rat development causes blastocyst abnormalities andprogramming of postnatal hypertensionrdquoDevelopment vol 127no 19 pp 4195ndash4202 2000
[55] E S Jungheim E L Schoeller K L Marquard E D LoudenJ E Schaffer and K H Moley ldquoDiet-induced obesity modelabnormal oocytes and persistent growth abnormalities in theoffspringrdquo Endocrinology vol 151 no 8 pp 4039ndash4046 2010
[56] A J Watkins A Wilkins C Cunningham et al ldquoLow proteindiet fed exclusively during mouse oocyte maturation leadsto behavioural and cardiovascular abnormalities in offspringrdquoJournal of Physiology vol 586 no 8 pp 2231ndash2244 2008
[57] E Zambrano P M Martınez-Samayoa G L Rodrıguez-Gonzalez and P W Nathanielsz ldquoDietary intervention prior topregnancy reverses metabolic programming in male offspringof obese ratsrdquo Journal of Physiology vol 588 no 10 pp 1791ndash1799 2010
[58] M S Patel andM Srinivasan ldquoMetabolic programming causesand consequencesrdquoThe Journal of Biological Chemistry vol 277no 3 pp 1629ndash1632 2002
[59] R A Simmons ldquoRole of metabolic programming in the patho-genesis of 120573-cell failure in postnatal liferdquo Reviews in Endocrineand Metabolic Disorders vol 8 no 2 pp 95ndash104 2007
[60] I B Leibiger B Leibiger and P O Berggren ldquoInsulin signalingin the pancreatic 120573-cellrdquoAnnual Review of Nutrition vol 28 pp233ndash251 2008
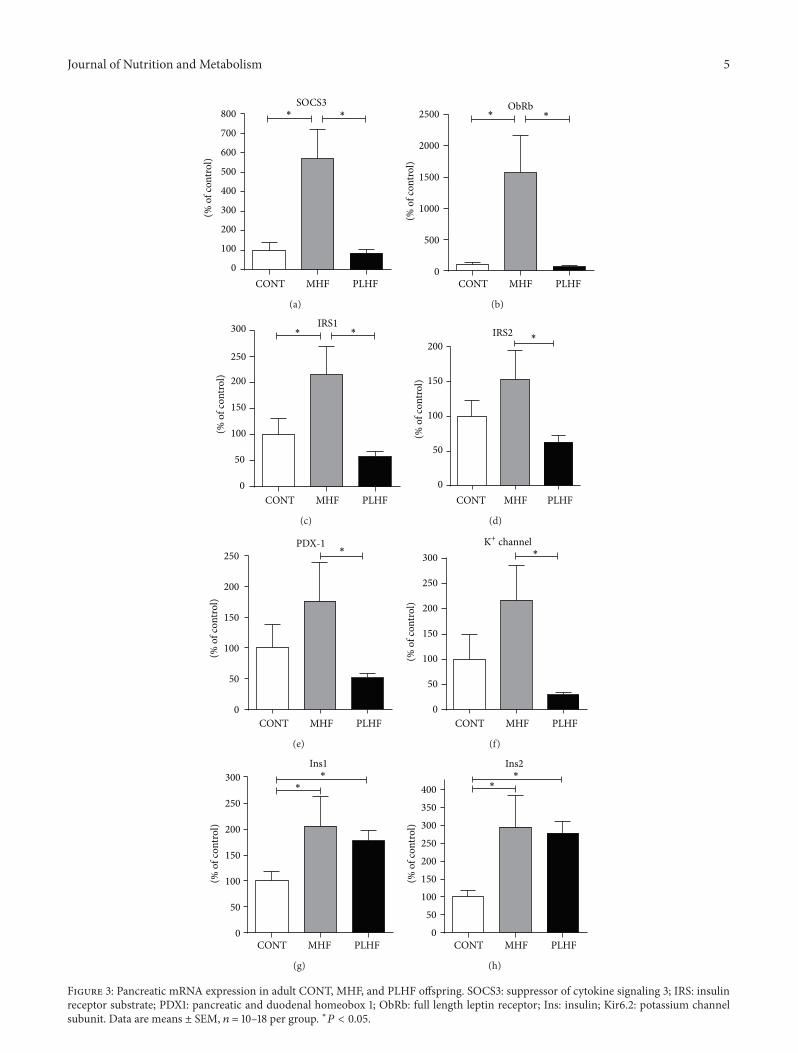
Journal of Nutrition and Metabolism 5
SOCS3
0
100
200
300
400
500
600
700
800
( o
f con
trol)
lowast lowast
CONT MHF PLHF
(a)
ObRb
0
500
1000
1500
2000
2500 lowast lowast
( o
f con
trol)
CONT MHF PLHF
(b)
IRS1
0
50
100
150
200
250
300 lowastlowast
( o
f con
trol)
CONT MHF PLHF
(c)
IRS2
0
50
100
150
200lowast
( o
f con
trol)
CONT MHF PLHF
(d)
PDX-1
0
50
100
150
200
250 lowast
( o
f con
trol)
CONT MHF PLHF
(e)
0
50
100
150
200
250
300 lowast
K+ channel
CONT MHF PLHF
( o
f con
trol)
(f)
Ins1
0
50
100
150
200
250
300 lowast
lowast
CONT MHF PLHF
( o
f con
trol)
(g)
Ins2
CONT MHF PLHF0
50100150200250300350400 lowast
lowast
( o
f con
trol)
(h)
Figure 3 Pancreatic mRNA expression in adult CONT MHF and PLHF offspring SOCS3 suppressor of cytokine signaling 3 IRS insulinreceptor substrate PDX1 pancreatic and duodenal homeobox 1 ObRb full length leptin receptor Ins insulin Kir62 potassium channelsubunit Data are means plusmn SEM 119899 = 10ndash18 per group lowast119875 lt 005
6 Journal of Nutrition and Metabolism
Table 2 Correlations between SOCS3 and other key genes N = 8ndash13 per group
CONT MHF PLHFSOCS3 versus ObRb 119877
2 = 069 P = 008 R2 = 087 P = 00008 1198772 = 003 P = 064
SOCS3 versus IR R2 = 067 P = 002 1198772 = 002 P = 071 119877
2 = 0002 P = 090SOCS3 versus IRS1 R2 = 084 P = 001 R2 = 058 P = 002 119877
2 = 004 P = 051SOCS3 versus IRS2 R2 = 098 P = 0002 R2 = 058 P = 002 119877
2 = 015 P = 021SOCS3 versus Kir62 channel R2 = 082 P = 003 R2 = 093 P lt 00001 119877
2 = 0002 P = 090SOCS3 versus Pdx1 R2 = 078 P = 0009 R2 = 091 P = 00003 119877
2 = 007 P = 040
CONT MHF PLHF0
100
200
300
400
500
STAT
3 (
of c
ontro
l)
lowast
lowast
(a)
PI3K
( o
f con
trol)
CONT MHF PLHF0
50
100
150lowast
lowast
(b)
Figure 4 Pancreatic STAT3 (a) and PI3K mRNA expression (b) Data are means plusmn SEM 119899 = 10ndash18 per group STAT3 signal transducer andactivator of transcription 3 PI3K phosphatidylinositol 3-kinase lowast119875 lt 005
may be separate influences of maternal obesity during thepericonceptional period and late gestation on adiposity inoffspring [31] We have previously shown that ldquoprogrammedrdquoobesity and simple diet-induced obesity can develop viadifferent underlying mechanisms [32] The present studyfurther reinforces the importance of the timing of the dietaryinsult on the metabolic derangements that ensue in offspringduring adulthood
SOCS3 inhibition of ObRb signaling has been proposedas a mechanism for leptin resistance in a range of tissues [33ndash35] including pancreatic 120573 cells [36] It is clear that leptinresistance in PLHF pancreata was mediated by mechanismsthat did not involve SOCS3 and this has been reported previ-ously in rat models of HF feeding [37 38] SOCS3 is normallyincreased at sites of both acute and chronic inflammation [39]and has been shown to specifically suppress STAT3 activation[40] This agrees with the present study a marked increasein SOCS3 mRNA levels in MHF offspring is paralleled bySTAT3 levels similar to control offspring Leptin is knownto increase SOCS3 expression in isolated human pancreaticislets and in islets from obob mice treated in vivo [36] ourdata imply that leptin-induced SOCS3 stimulation appearsintact in MHF offspring Conversely in PLHF offspring
SOCS3 expression levels were similar to controls in thepresence of markedly elevated STAT3 expression Of notedespite similar expression levels of SOCS3 between CONTand PLHF offspring the positive correlations that wereobserved between SOCS3 and other key signaling genes wererestricted mainly to CONT and MHF offspring only Thismay point to a maladaptation in PLHF offspring due to theacute HF dietary exposure and a change in homeostatic set-points in MHF offspring due to mothers entering pregnancyin an already obese state Despite the link between SOCS3and inflammation there were no observed changes in TNF-120572 mRNA levels in the MHF group whilst there was asignificant increase in TNF-120572 expression in PLHF offspringIn addition mRNA levels of the macrophage marker CD68were increased in both HF groups and taken togethermay indicate differing macrophage phenotypespolarizationstates resulting in divergent offspring inflammatory profilesdependent upon prior maternal nutritional background Ofnote despite marked changes in inflammatorymarkers at thetissue level we observed no changes in circulating plasmaTNF-120572 concentrationHowever despite enhanced plasma IL-1120573 concentrations pancreatic IL-1120573 expression was unaltered
Journal of Nutrition and Metabolism 7
CONT
0 250 500 7500
250
500
750
PDX-1
1199032= 097
K+ch
anne
l
(a)
MHF
0 250 500 750 10000
500
1000
15001199032= 094
PDX-1
K+ch
anne
l
(b)
PLHF
0 50 100 150 200 250 3000
50
100
150
200
250
3001199032= 078
PDX-1
K+ch
anne
l
(c)
Figure 5 Relationship between PDX-1 and potassium channel subunit Kir62 in adult CONT MHF and PLHF male offspring PDX-1pancreatic and duodenal homeobox 1119873 = 8ndash10 per group
between groups implicating other tissues as possible systemicsources of this proinflammatory cytokine
PLHF offspring exhibited decreased PI3K mRNA levelscompared to CONT animals Induction of the ObRbPI3Ksignaling pathway prolongs potassium channel openingleading to decreased insulin exocytosis a direct action ofleptin which inhibits insulin secretion [41] Moreover thelowered PI3K mRNA levels in PLHF offspring were con-comitant with a reduction in IRS1 and IRS2 mRNA whosephosphorylation is coupled to PI3K activation Of notedownstreamof PI3K PDE3B gene expression levels remainedsimilar to controls suggesting that altered leptin signalingwas achieved in earlier stages of this pathway Thus attenua-tion of the ObRbPI3K signaling pathwaymaymediate leptinresistance in these hyperleptinemic PLHF offspring Sucha mechanism has been observed before in hypothalamicneurons of rats with diet-induced obesity (DIO) [42]
IRS proteins and PI3K have leptin independent rolesin the 120573 cell Both are found downstream of the insulinreceptor in the IRIRSPI3K pathway which mediates theautocrine actions of the insulin-IR complex [43 44] Ourobserved attenuation of PI3K and IRS mRNA did nothowever affect insulin synthesismdashPLHF animals were hyper-insulinemic and preproinsulin mRNA concentrations wereelevated Furthermore mRNA concentrations of IR wereunaffected potentially due to lack of SOCS3 induction inPLHF offspring similar to Controls Thus these mRNAchanges of PI3K coupled to IRS likely reflect a mechanismof leptin resistance in this group
Mean concentrations of ObRb mRNA in PLHF offspringwere similar to controls but a significant inverse relationshipbetween plasma leptin and ObRb mRNA levels in the PLHFgroup was suggestive of ObRb downregulation A downregu-lation of receptor populations in the face of elevated ligand
8 Journal of Nutrition and Metabolism
CONT MHF PLHF0
50
100
150
200
TNF-120572
mRN
A (
of c
ontro
l)
lowast
lowast
(a)
CD68
mRN
A (
of c
ontro
l)
CONT MHF PLHF0
50
100
150
200
lowast
lowast
(b)
IL1-
R1 m
RNA
( o
f con
trol)
CONT MHF PLHF0
50
100
150
200lowast
(c)
Figure 6 Pancreatic mRNA expression of TNF-120572 (a) CD68 (b) and IL1-R1 (c) in adult CONT MHF and PLHF offspring TNF-120572 tumornecrosis factor-alpha CD68 cluster of differentiation 68 IL1-R1 interleukin 1 receptor type 1 Data are means plusmn SEM 119899 = 10ndash18 per grouplowast
119875 lt 005
concentrations is not uncommon in resistant states It isseen for example in the hypothalamus in the presence ofhyperleptinemia [45ndash47] and during pregnancy which is anatural state of leptin resistance [48]
A rise in IRS1 and IRS2 mRNA associated with elevatedObRb and SOCS3 mRNA in MHF offspring may seemunexpected since SOCS3 is known to target insulin receptorsignaling by interference with IRS action [49] However sev-eral studies have demonstrated that SOCS3 mediates insulinresistance in both adipocytes and hepatocytes through post-translational modifications such as inhibition of IRS tyro-sine phosphorylation [50] and targeted ubiquitin-mediateddegradation of IRS1 and IRS2 without effecting transcription[49 51] IRS proteins are involved in both leptin signalingand insulin signaling so altered gene expression levels mayrepresent changes in either (or both) pathways although IRSresponse to leptin is reportedly much lower than that seenwith insulin [52] We observed an increase in IRS1 expressionin MHF offspring compared to Controls This upregulationof IRS1 expression may be related to the increased insulin
production of the MHF offspring given the known role ofIRS1 in the regulation of insulin secretion [53]
SOCS3 is also reported to repress preproinsulin genetranscription [41] although other mechanisms clearly coun-tered this action inMHFoffspring since Ins1 and Ins2mRNAexpression levels remained elevated This observation wasconfirmed by the significantly raised levels of circulatingplasma insulin and the lack of any correlation in levels ofmRNA between SOCS3 and Ins1 or Ins2
Some of the observed differences may relate to theinitial hyperphagia observed in the PLHF maternal groupas reported previously [14] Since rodents primarily eat ona calorific basis they will passively over-consume calorieswhen initially placed on an HF diet and self-regulate backto a normal total caloric intake over a period of a few daysIt has been shown in other rodent models that alterations innutrition around the time of implantation can lead to lastingmetabolic effects in the offspring [54] Maternal obesity hasbeen shown to exert adverse effects as early as the oocyteand preimplantation embryo stage and that these effects may
Journal of Nutrition and Metabolism 9
contribute to the lasting morbidity observed in offspring[55] Similarly even a very short period of altered nutrition(restricted to one ovulatory cycle) prior to natural mating canelicit lasting effects on the offspring [56] and this representsa further point of difference between the MHF and PLHFgroups
The present study was not designed to incorporate theeffect of a reversal paradigm whereby mothers fed thepre-conceptional HF diet were placed on a standard dietthroughout pregnancy and lactation However while there isevidence that a period of dietary restriction in obese mothersmay ablate the programming of obesity in offspring [57]it may be associated with an activation of the stress axisin the offspring [31] Further the data are inconsistent ithas also been shown that dams with diet-induced obesityfed a standard diet throughout pregnancy and lactation stilltransmit the propensity for obesity and metabolic disease totheir offspring [58]
We recognize that gene expression analysis on wholepancreatic tissue is a limitation of the present study Dueto the size of the cohort and logistical considerations onlywhole pancreatic tissue was a possibility whereas ideallyonly pure islet populations would have been examined [59]However for the key genes presented such as ObRb SOCS3and IRS expression has been shown to be primarily confinedto the endocrine islets and not been detected in the exocrinepancreas [39 41 60] therefore we are confident that theobserved changes in the genes studied represent changes inthe islets Further although we have shown that differencesin pancreatic gene expression in offspring are dependentupon the timing of maternal high fat diet exposure this doesnot itself imply causation in the development of the obesephenotypemdashother relevant pathways including liver skeletalmuscle and fat metabolism will also be integrated into theprogrammed phenotype development
This study has demonstrated enduring deleterious effectsof a maternal HF diet during early windows of develop-ment Maternal HF feeding both before conception andorthrough pregnancy and lactation induced obesity hyper-insulinemia and hyperleptinemia in adult offspring eventhough offspring were fed a standard diet postnatally Timingof exposure to the maternal HF diet differentially alteredgene expression in signaling pathways downstream of thepancreatic leptin receptor that mediated leptin resistanceThis also raises the possibility that despite common offspringobesity treatment modalities may only be efficacious in asubpopulation of treated offspring as the current data suggestthat the metabolic pathways disrupted will be dependentupon the timing of the early life dietary insult
Abbreviations
CD68 Cluster of differentiation 68CONT ControlDEXA Dual energy X-ray absorptiometryELISA Enzyme-linked immunosorbent assayGLUT2 Glucose transporter 2GP130 Glycoprotein 130HF High fat
HOMA Homeostatic model assessmentHPRT HypoxanthinephophoribosyltransferaseIL InterleukinIns InsulinIR Insulin resistanceIRS Insulin receptor substrateKir62 Potassium channel subunitLIF Leukemia inhibitory factorMHF Maternal high fatOBR Leptin receptorPDE3B Phosphodiesterase 3BPDX-1 Pancreatic and duodenal homeobox-1PI3K Phosphatidylinositide 3-kinasePLHF Pregnancy and lactation high fatSOCS3 Suppressor of cytokine signaling 3STAT3 Signal transducer and activator of
transcription 3TNF-120572 Tumor necrosis factor-alpha
Conflict of Interests
The authors have no conflict of interests with any financialorganization regarding the material discussed in this paper
Author Contributions
Mark H Vickers developed the animal model designed thestudy completed the data analysis and wrote the first draftof the paper Graham J Howie maintained the experimentalcohort performed the gene expression analysis and con-tributed to the original draft paper Clare M Reynolds per-formed the inflammatory related assays and gene expressionwork Deborah M Sloboda provided input into data analysisand interpretation
Acknowledgments
The authors acknowledge the technical support of TaniaKamal with endocrine assays The authors acknowledgefunding support from the Health Research Council of NewZealand and Gravida National Centre for Growth andDevelopment
References
[1] M M Hillemeier C S Weisman C Chuang D S Downs JMcCall-Hosenfeld and F Camacho ldquoTransition to overweightor obesity among women of reproductive agerdquo Journal ofWomenrsquos Health vol 20 no 5 pp 703ndash710 2011
[2] J E Norman and RM Reynolds ldquoThe consequences of obesityand excess weight gain in pregnancyrdquo Proceedings of the Nutri-tion Society vol 70 no 4 pp 450ndash456 2011
[3] H A Abenhaim R A Kinch L Morin A Benjamin and RUsher ldquoEffect of prepregnancy body mass index categories onobstetrical and neonatal outcomesrdquo Archives of Gynecology andObstetrics vol 275 no 1 pp 39ndash43 2007
[4] S DHMalnick andH Knobler ldquoThemedical complications ofobesityrdquo Oxford Journals Medicine vol 99 no 9 pp 565ndash5792006
10 Journal of Nutrition and Metabolism
[5] P M Catalano ldquoManagement of obesity in pregnancyrdquo Obstet-rics and Gynecology vol 109 no 2 pp 419ndash433 2007
[6] L Poston ldquoGestational weight gain influences on the long-termhealth of the childrdquo Current Opinion in Clinical Nutrition andMetabolic Care vol 15 pp 252ndash257 2012
[7] H Hochner Y Friedlander R Calderon-Margalit et al ldquoAsso-ciations of maternal prepregnancy body mass index and gesta-tional weight gain with adult offspring cardiometabolic riskfactors the Jerusalem perinatal family follow-up studyrdquo Circu-lation vol 125 no 11 pp 1381ndash1389 2012
[8] S J Spencer ldquoEarly life programming of obesity the impactof the perinatal environment on the development of obesityand metabolic dysfunction in the offspringrdquo Current DiabetesReviews vol 8 no 1 pp 55ndash68 2012
[9] H Chen D Simar K Lambert J Mercier and M J MorrisldquoMaternal and postnatal overnutrition differentially impactappetite regulators and fuel metabolismrdquo Endocrinology vol149 no 11 pp 5348ndash5356 2008
[10] P M Catalano ldquoObesity and pregnancymdashthe propagation of aviscous cyclerdquo Journal of Clinical Endocrinology and Metabo-lism vol 88 no 8 pp 3505ndash3506 2003
[11] M W Gillman S L Rifas-Shiman K Kleinman E Oken JW Rich-Edwards and EM Taveras ldquoDevelopmental origins ofchildhood overweight potential public health impactrdquo Obesityvol 16 no 7 pp 1651ndash1656 2008
[12] E Oken EM Taveras K P Kleinman JW Rich-Edwards andM W Gillman ldquoGestational weight gain and child adiposity atage 3 yearsrdquoTheAmerican Journal of Obstetrics and Gynecologyvol 196 no 4 pp 322e1ndash322e8 2007
[13] H M Ehrenberg B M Mercer and P M Catalano ldquoThe influ-ence of obesity and diabetes on the prevalence of macrosomiardquoThe American Journal of Obstetrics and Gynecology vol 191 no3 pp 964ndash968 2004
[14] G J Howie D M Sloboda T Kamal and M H VickersldquoMaternal nutritional history predicts obesity in adult offspringindependent of postnatal dietrdquo Journal of Physiology vol 587no 4 pp 905ndash915 2009
[15] D M Sloboda G J Howie A Pleasants P D Gluckman andM H Vickers ldquoPre- and postnatal nutritional histories influ-ence reproductive maturation and ovarian function in the ratrdquoPLoS ONE vol 4 no 8 Article ID e6744 2009
[16] M C Battista M F Hivert K Duval and J P BaillargeonldquoIntergenerational cycle of obesity and diabetes how can wereduce the burdens of these conditions on the health of futuregenerationsrdquo Experimental Diabetes Research vol 2011 ArticleID 596060 19 pages 2011
[17] T J Kieffer and J F Habener ldquoThe adipoinsular axis effects ofleptin on pancreatic 120573-cellsrdquo The American Journal of Physiol-ogy vol 278 no 1 pp E1ndashE14 2000
[18] S L Gray C Donald A Jetha S D Covey and T J KiefferldquoHyperinsulinemia precedes insulin resistance in mice lackingpancreatic 120573-cell leptin signalingrdquo Endocrinology vol 151 no 9pp 4178ndash4186 2010
[19] S D Covey R DWideman CMcDonald et al ldquoThe pancreat-ic 120573 cell is a key site formediating the effects of leptin on glucosehomeostasisrdquo Cell Metabolism vol 4 no 4 pp 291ndash302 2006
[20] K J Dudley DM Sloboda K L Connor J Beltrand andMHVickers ldquoOffspring ofmothers fed a high fat diet display hepaticcell cycle inhibition and associated changes in gene expressionand DNA methylationrdquo PLoS ONE vol 6 no 7 Article IDe21662 2011
[21] K L Connor M H Vickers J Beltrand M J Meaney and DM Sloboda ldquoNature nurture or nutrition Impact of maternalnutrition on maternal care offspring development and repro-ductive functionrdquo Journal of Physiology vol 590 part 9 pp2167ndash2180 2012
[22] J Beltrand D M Sloboda K L Connor M Truong and MH Vickers ldquoThe effect of neonatal leptin antagonism in malerat offspring is dependent upon the interaction between priormaternal nutritional status and post-weaning dietrdquo Journal ofNutrition andMetabolism vol 2012 Article ID 296935 10 pages2012
[23] K L Connor M H Vickers C Cupido E Sirimanne and DM Sloboda ldquoMaternal high fat diet during critical windowsof development alters adrenal cortical and medullary enzymeexpression in adult male rat offspringrdquo Journal of the Develop-mental Origins of Health and Disease vol 1 no 4 pp 245ndash2542010
[24] P J Mark C Sisala K Connor et al ldquoA maternal high-fat dietin rat pregnancy reduces growth of the fetus and the placen-tal junctional zone but not placental labyrinth zone growthrdquoJournal of the Developmental Origins of Health and Disease vol2 no 1 pp 63ndash70 2011
[25] D R Matthews J P Hosker A S Rudenski B A Naylor DF Treacher and R C Turner ldquoHomeostasis model assessmentinsulin resistance and 120573-cell function from fasting plasmaglucose and insulin concentrations in manrdquo Diabetologia vol28 no 7 pp 412ndash419 1985
[26] M P Hermans J C Levy R J Morris and R C TurnerldquoComparison of tests of120573-cell function across a range of glucosetolerance from normal to diabetesrdquo Diabetes vol 48 no 9 pp1779ndash1786 1999
[27] A B Bernal M H Vickers M B Hampton R A Poynton andD M Sloboda ldquoMaternal undernutrition significantly impactsovarian follicle number and increases ovarian oxidative stress inadult rat offspringrdquo PLoS ONE vol 5 no 12 Article ID e155582010
[28] J Vandesompele K de Preter F Pattyn et al ldquoAccurate nor-malization of real-time quantitative RT-PCR data by geometricaveraging of multiple internal control genesrdquo Genome biologyvol 3 no 7 Article ID RESEARCH0034 2002
[29] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the 2-ΔΔCT methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
[30] J Armitage L Poston and P Taylor ldquoDevelopmental originsof obesity and the metabolic syndrome the role of maternalobesityrdquo Frontiers of Hormone Research vol 36 pp 73ndash84 2007
[31] S Zhang L Rattanatray J L Morrison L M Nicholas S Lieand I C McMillen ldquoMaternal obesity and the early originsof childhood obesity weighing up the benefits and costs ofmaternal weight loss in the periconceptional period for theoffspringrdquo Experimental Diabetes Research vol 2011 Article ID585749 10 pages 2011
[32] N M Thompson A M Norman S S Donkin et al ldquoPrenataland postnatal pathways to obesity different underlying mech-anisms different metabolic outcomesrdquo Endocrinology vol 148no 5 pp 2345ndash2354 2007
[33] T Fuentes I Ara A Guadalupe-Grau et al ldquoLeptin receptor170 kDa (OB-R170) protein expression is reduced in obesehuman skeletal muscle a potential mechanism of leptin resis-tancerdquo Experimental Physiology vol 95 no 1 pp 160ndash171 2010
[34] Z Wang Y T Zhou T Kakuma et al ldquoLeptin resistance ofadipocytes in obesity role of suppressors of cytokine signalingrdquo
Journal of Nutrition and Metabolism 11
Biochemical and Biophysical Research Communications vol 277no 1 pp 20ndash26 2000
[35] C Bjoslashrbaeligk K El-Haschimi J D Frantz and J S Flier ldquoThe roleof SOCS-3 in leptin signaling and leptin resistancerdquoThe Journalof Biological Chemistry vol 274 no 42 pp 30059ndash30065 1999
[36] K Laubner T J Kieffer N T Lam X Niu F Jakob and JSeufert ldquoInhibition of preproinsulin gene expression by leptininduction of suppressor of cytokine signaling 3 in pancreatic 120573-cellsrdquo Diabetes vol 54 no 12 pp 3410ndash3417 2005
[37] C Peiser G P McGregor and R E Lang ldquoLeptin receptorexpression and suppressor of cytokine signaling transcriptlevels in high-fat-fed ratsrdquo Life Sciences vol 67 no 24 pp 2971ndash2981 2000
[38] G R Steinberg A C Smith S Wormald P Malenfant CCollier and D J Dyck ldquoEndurance training partially reversesdietary-induced leptin resistance in rodent skeletal musclerdquoTheAmerican Journal of Physiology vol 286 no 1 pp E57ndashE632004
[39] G E White A Cotterill M R Addley E J Soilleux and DR Greaves ldquoSuppressor of cytokine signalling protein SOCS3expression is increased at sites of acute and chronic inflamma-tionrdquo Journal of Molecular Histology vol 42 no 2 pp 137ndash1512011
[40] H YasukawaMOhishiHMori et al ldquoIL-6 induces an anti-in-flammatory response in the absence of SOCS3 inmacrophagesrdquoNature Immunology vol 4 no 6 pp 551ndash556 2003
[41] J Seufert ldquoLeptin effects on pancreatic 120573-cell gene expressionand functionrdquo Diabetes vol 53 no 1 pp S152ndashS158 2004
[42] A S Metlakunta M Sahu and A Sahu ldquoHypothalamicphosphatidylinositol 3-kinase pathway of leptin signaling isimpaired during the development of diet-induced obesity inFVBNmicerdquo Endocrinology vol 149 no 3 pp 1121ndash1128 2008
[43] C A Aspinwall W J Qian M G Roper R N Kulkarni C RKahn and R T Kennedy ldquoRoles of insulin receptor substrate-1phosphatidylinositol 3-kinase and release of intracellular Ca2+stores in insulin-stimulated insulin secretion in 120573-cellsrdquo TheJournal of Biological Chemistry vol 275 no 29 pp 22331ndash223382000
[44] I B Leibiger B Leibiger T Moede and P O BerggrenldquoExocytosis of insulin promotes insulin gene transcription viathe insulin receptorPI-3 kinasep70 s6 kinase and CaM kinasepathwaysrdquoMolecular Cell vol 1 no 6 pp 933ndash938 1998
[45] R L Martin E Perez Y J He R Dawson Jr andW J MillardldquoLeptin resistance is associated with hypothalamic leptin recep-tor mRNA and protein downregulationrdquo Metabolism vol 49no 11 pp 1479ndash1484 2000
[46] J Wilsey and P J Scarpace ldquoCaloric restriction reverses thedeficits in leptin receptor protein and leptin signaling capacityassociated with diet-induced obesity role of leptin in the reg-ulation of hypothalamic long-form leptin receptor expressionrdquoJournal of Endocrinology vol 181 no 2 pp 297ndash306 2004
[47] Y Zhang and P J Scarpace ldquoThe role of leptin in leptin resist-ance and obesityrdquo Physiology and Behavior vol 88 no 3 pp249ndash256 2006
[48] D R Grattan S R Ladyman and R A Augustine ldquoHormonalinduction of leptin resistance during pregnancyrdquoPhysiology andBehavior vol 91 no 4 pp 366ndash374 2007
[49] L RuiM YuanD Frantz S Shoelson andM FWhite ldquoSOCS-1 and SOCS-3 block insulin signaling by ubiquitin-mediateddegradation of IRS1 and IRS2rdquoThe Journal of Biological Chem-istry vol 277 no 44 pp 42394ndash42398 2002
[50] K Ueki T Kondo and C R Kahn ldquoSuppressor of cytokine sig-naling 1 (SOCS-1) and SOCS-3 cause insulin resistance throughinhibition of tyrosine phosphorylation of insulin receptor sub-strate proteins by discrete mechanismsrdquoMolecular and CellularBiology vol 24 no 12 pp 5434ndash5446 2004
[51] S B Jorgensen O rsquoNeill HM L Sylow et al ldquoDeletion of skel-etal muscle SOCS3 prevents insulin resistance in obesityrdquo Dia-betes vol 62 no 1 pp 56ndash64 2013
[52] W Becker ldquoLeptin signal transductionrdquo in Leptin and LeptinAntagonists A Gertler Ed pp 1ndash9 Landes Bioscience AustinTex USA 2009
[53] R N Kulkarni J N Winnay M Daniels et al ldquoAltered func-tion of insulin receptor substrate 1-deficient mouse islets andcultured 120573-cell linesrdquo Journal of Clinical Investigation vol 104no 12 pp R69ndashR75 1999
[54] W Y Kwong A E Wild P Roberts A C Willis and T PFleming ldquoMaternal undernutrition during the preimplantationperiod of rat development causes blastocyst abnormalities andprogramming of postnatal hypertensionrdquoDevelopment vol 127no 19 pp 4195ndash4202 2000
[55] E S Jungheim E L Schoeller K L Marquard E D LoudenJ E Schaffer and K H Moley ldquoDiet-induced obesity modelabnormal oocytes and persistent growth abnormalities in theoffspringrdquo Endocrinology vol 151 no 8 pp 4039ndash4046 2010
[56] A J Watkins A Wilkins C Cunningham et al ldquoLow proteindiet fed exclusively during mouse oocyte maturation leadsto behavioural and cardiovascular abnormalities in offspringrdquoJournal of Physiology vol 586 no 8 pp 2231ndash2244 2008
[57] E Zambrano P M Martınez-Samayoa G L Rodrıguez-Gonzalez and P W Nathanielsz ldquoDietary intervention prior topregnancy reverses metabolic programming in male offspringof obese ratsrdquo Journal of Physiology vol 588 no 10 pp 1791ndash1799 2010
[58] M S Patel andM Srinivasan ldquoMetabolic programming causesand consequencesrdquoThe Journal of Biological Chemistry vol 277no 3 pp 1629ndash1632 2002
[59] R A Simmons ldquoRole of metabolic programming in the patho-genesis of 120573-cell failure in postnatal liferdquo Reviews in Endocrineand Metabolic Disorders vol 8 no 2 pp 95ndash104 2007
[60] I B Leibiger B Leibiger and P O Berggren ldquoInsulin signalingin the pancreatic 120573-cellrdquoAnnual Review of Nutrition vol 28 pp233ndash251 2008
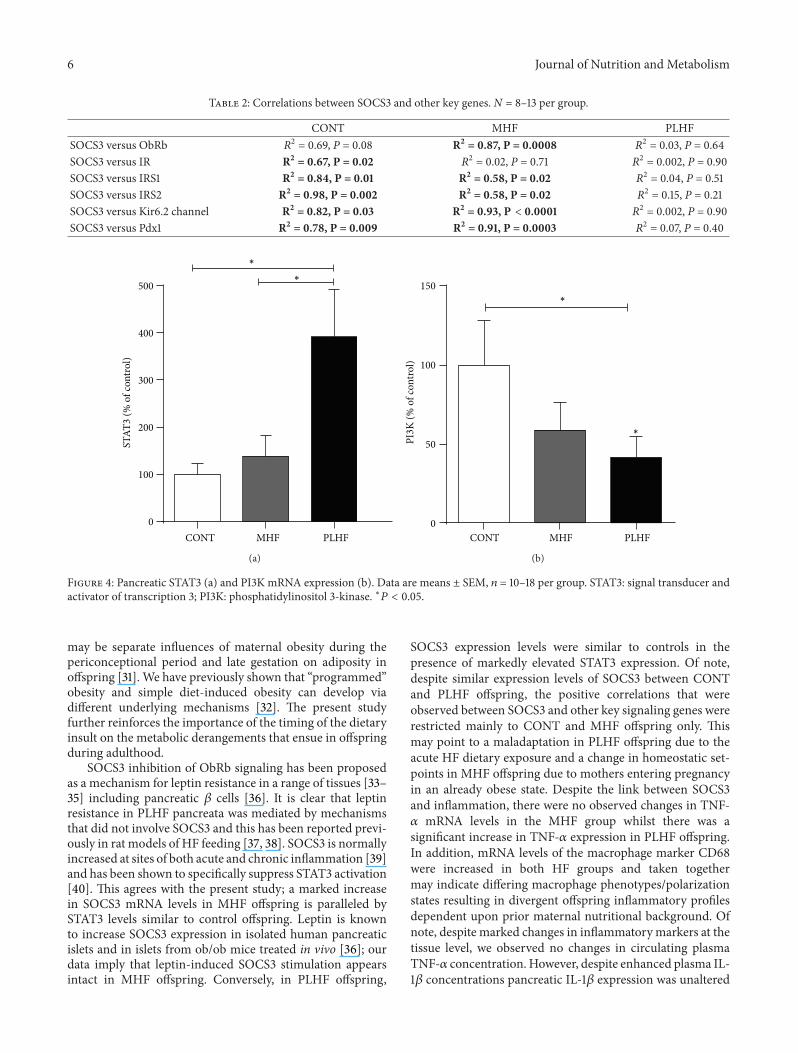
6 Journal of Nutrition and Metabolism
Table 2 Correlations between SOCS3 and other key genes N = 8ndash13 per group
CONT MHF PLHFSOCS3 versus ObRb 119877
2 = 069 P = 008 R2 = 087 P = 00008 1198772 = 003 P = 064
SOCS3 versus IR R2 = 067 P = 002 1198772 = 002 P = 071 119877
2 = 0002 P = 090SOCS3 versus IRS1 R2 = 084 P = 001 R2 = 058 P = 002 119877
2 = 004 P = 051SOCS3 versus IRS2 R2 = 098 P = 0002 R2 = 058 P = 002 119877
2 = 015 P = 021SOCS3 versus Kir62 channel R2 = 082 P = 003 R2 = 093 P lt 00001 119877
2 = 0002 P = 090SOCS3 versus Pdx1 R2 = 078 P = 0009 R2 = 091 P = 00003 119877
2 = 007 P = 040
CONT MHF PLHF0
100
200
300
400
500
STAT
3 (
of c
ontro
l)
lowast
lowast
(a)
PI3K
( o
f con
trol)
CONT MHF PLHF0
50
100
150lowast
lowast
(b)
Figure 4 Pancreatic STAT3 (a) and PI3K mRNA expression (b) Data are means plusmn SEM 119899 = 10ndash18 per group STAT3 signal transducer andactivator of transcription 3 PI3K phosphatidylinositol 3-kinase lowast119875 lt 005
may be separate influences of maternal obesity during thepericonceptional period and late gestation on adiposity inoffspring [31] We have previously shown that ldquoprogrammedrdquoobesity and simple diet-induced obesity can develop viadifferent underlying mechanisms [32] The present studyfurther reinforces the importance of the timing of the dietaryinsult on the metabolic derangements that ensue in offspringduring adulthood
SOCS3 inhibition of ObRb signaling has been proposedas a mechanism for leptin resistance in a range of tissues [33ndash35] including pancreatic 120573 cells [36] It is clear that leptinresistance in PLHF pancreata was mediated by mechanismsthat did not involve SOCS3 and this has been reported previ-ously in rat models of HF feeding [37 38] SOCS3 is normallyincreased at sites of both acute and chronic inflammation [39]and has been shown to specifically suppress STAT3 activation[40] This agrees with the present study a marked increasein SOCS3 mRNA levels in MHF offspring is paralleled bySTAT3 levels similar to control offspring Leptin is knownto increase SOCS3 expression in isolated human pancreaticislets and in islets from obob mice treated in vivo [36] ourdata imply that leptin-induced SOCS3 stimulation appearsintact in MHF offspring Conversely in PLHF offspring
SOCS3 expression levels were similar to controls in thepresence of markedly elevated STAT3 expression Of notedespite similar expression levels of SOCS3 between CONTand PLHF offspring the positive correlations that wereobserved between SOCS3 and other key signaling genes wererestricted mainly to CONT and MHF offspring only Thismay point to a maladaptation in PLHF offspring due to theacute HF dietary exposure and a change in homeostatic set-points in MHF offspring due to mothers entering pregnancyin an already obese state Despite the link between SOCS3and inflammation there were no observed changes in TNF-120572 mRNA levels in the MHF group whilst there was asignificant increase in TNF-120572 expression in PLHF offspringIn addition mRNA levels of the macrophage marker CD68were increased in both HF groups and taken togethermay indicate differing macrophage phenotypespolarizationstates resulting in divergent offspring inflammatory profilesdependent upon prior maternal nutritional background Ofnote despite marked changes in inflammatorymarkers at thetissue level we observed no changes in circulating plasmaTNF-120572 concentrationHowever despite enhanced plasma IL-1120573 concentrations pancreatic IL-1120573 expression was unaltered
Journal of Nutrition and Metabolism 7
CONT
0 250 500 7500
250
500
750
PDX-1
1199032= 097
K+ch
anne
l
(a)
MHF
0 250 500 750 10000
500
1000
15001199032= 094
PDX-1
K+ch
anne
l
(b)
PLHF
0 50 100 150 200 250 3000
50
100
150
200
250
3001199032= 078
PDX-1
K+ch
anne
l
(c)
Figure 5 Relationship between PDX-1 and potassium channel subunit Kir62 in adult CONT MHF and PLHF male offspring PDX-1pancreatic and duodenal homeobox 1119873 = 8ndash10 per group
between groups implicating other tissues as possible systemicsources of this proinflammatory cytokine
PLHF offspring exhibited decreased PI3K mRNA levelscompared to CONT animals Induction of the ObRbPI3Ksignaling pathway prolongs potassium channel openingleading to decreased insulin exocytosis a direct action ofleptin which inhibits insulin secretion [41] Moreover thelowered PI3K mRNA levels in PLHF offspring were con-comitant with a reduction in IRS1 and IRS2 mRNA whosephosphorylation is coupled to PI3K activation Of notedownstreamof PI3K PDE3B gene expression levels remainedsimilar to controls suggesting that altered leptin signalingwas achieved in earlier stages of this pathway Thus attenua-tion of the ObRbPI3K signaling pathwaymaymediate leptinresistance in these hyperleptinemic PLHF offspring Sucha mechanism has been observed before in hypothalamicneurons of rats with diet-induced obesity (DIO) [42]
IRS proteins and PI3K have leptin independent rolesin the 120573 cell Both are found downstream of the insulinreceptor in the IRIRSPI3K pathway which mediates theautocrine actions of the insulin-IR complex [43 44] Ourobserved attenuation of PI3K and IRS mRNA did nothowever affect insulin synthesismdashPLHF animals were hyper-insulinemic and preproinsulin mRNA concentrations wereelevated Furthermore mRNA concentrations of IR wereunaffected potentially due to lack of SOCS3 induction inPLHF offspring similar to Controls Thus these mRNAchanges of PI3K coupled to IRS likely reflect a mechanismof leptin resistance in this group
Mean concentrations of ObRb mRNA in PLHF offspringwere similar to controls but a significant inverse relationshipbetween plasma leptin and ObRb mRNA levels in the PLHFgroup was suggestive of ObRb downregulation A downregu-lation of receptor populations in the face of elevated ligand
8 Journal of Nutrition and Metabolism
CONT MHF PLHF0
50
100
150
200
TNF-120572
mRN
A (
of c
ontro
l)
lowast
lowast
(a)
CD68
mRN
A (
of c
ontro
l)
CONT MHF PLHF0
50
100
150
200
lowast
lowast
(b)
IL1-
R1 m
RNA
( o
f con
trol)
CONT MHF PLHF0
50
100
150
200lowast
(c)
Figure 6 Pancreatic mRNA expression of TNF-120572 (a) CD68 (b) and IL1-R1 (c) in adult CONT MHF and PLHF offspring TNF-120572 tumornecrosis factor-alpha CD68 cluster of differentiation 68 IL1-R1 interleukin 1 receptor type 1 Data are means plusmn SEM 119899 = 10ndash18 per grouplowast
119875 lt 005
concentrations is not uncommon in resistant states It isseen for example in the hypothalamus in the presence ofhyperleptinemia [45ndash47] and during pregnancy which is anatural state of leptin resistance [48]
A rise in IRS1 and IRS2 mRNA associated with elevatedObRb and SOCS3 mRNA in MHF offspring may seemunexpected since SOCS3 is known to target insulin receptorsignaling by interference with IRS action [49] However sev-eral studies have demonstrated that SOCS3 mediates insulinresistance in both adipocytes and hepatocytes through post-translational modifications such as inhibition of IRS tyro-sine phosphorylation [50] and targeted ubiquitin-mediateddegradation of IRS1 and IRS2 without effecting transcription[49 51] IRS proteins are involved in both leptin signalingand insulin signaling so altered gene expression levels mayrepresent changes in either (or both) pathways although IRSresponse to leptin is reportedly much lower than that seenwith insulin [52] We observed an increase in IRS1 expressionin MHF offspring compared to Controls This upregulationof IRS1 expression may be related to the increased insulin
production of the MHF offspring given the known role ofIRS1 in the regulation of insulin secretion [53]
SOCS3 is also reported to repress preproinsulin genetranscription [41] although other mechanisms clearly coun-tered this action inMHFoffspring since Ins1 and Ins2mRNAexpression levels remained elevated This observation wasconfirmed by the significantly raised levels of circulatingplasma insulin and the lack of any correlation in levels ofmRNA between SOCS3 and Ins1 or Ins2
Some of the observed differences may relate to theinitial hyperphagia observed in the PLHF maternal groupas reported previously [14] Since rodents primarily eat ona calorific basis they will passively over-consume calorieswhen initially placed on an HF diet and self-regulate backto a normal total caloric intake over a period of a few daysIt has been shown in other rodent models that alterations innutrition around the time of implantation can lead to lastingmetabolic effects in the offspring [54] Maternal obesity hasbeen shown to exert adverse effects as early as the oocyteand preimplantation embryo stage and that these effects may
Journal of Nutrition and Metabolism 9
contribute to the lasting morbidity observed in offspring[55] Similarly even a very short period of altered nutrition(restricted to one ovulatory cycle) prior to natural mating canelicit lasting effects on the offspring [56] and this representsa further point of difference between the MHF and PLHFgroups
The present study was not designed to incorporate theeffect of a reversal paradigm whereby mothers fed thepre-conceptional HF diet were placed on a standard dietthroughout pregnancy and lactation However while there isevidence that a period of dietary restriction in obese mothersmay ablate the programming of obesity in offspring [57]it may be associated with an activation of the stress axisin the offspring [31] Further the data are inconsistent ithas also been shown that dams with diet-induced obesityfed a standard diet throughout pregnancy and lactation stilltransmit the propensity for obesity and metabolic disease totheir offspring [58]
We recognize that gene expression analysis on wholepancreatic tissue is a limitation of the present study Dueto the size of the cohort and logistical considerations onlywhole pancreatic tissue was a possibility whereas ideallyonly pure islet populations would have been examined [59]However for the key genes presented such as ObRb SOCS3and IRS expression has been shown to be primarily confinedto the endocrine islets and not been detected in the exocrinepancreas [39 41 60] therefore we are confident that theobserved changes in the genes studied represent changes inthe islets Further although we have shown that differencesin pancreatic gene expression in offspring are dependentupon the timing of maternal high fat diet exposure this doesnot itself imply causation in the development of the obesephenotypemdashother relevant pathways including liver skeletalmuscle and fat metabolism will also be integrated into theprogrammed phenotype development
This study has demonstrated enduring deleterious effectsof a maternal HF diet during early windows of develop-ment Maternal HF feeding both before conception andorthrough pregnancy and lactation induced obesity hyper-insulinemia and hyperleptinemia in adult offspring eventhough offspring were fed a standard diet postnatally Timingof exposure to the maternal HF diet differentially alteredgene expression in signaling pathways downstream of thepancreatic leptin receptor that mediated leptin resistanceThis also raises the possibility that despite common offspringobesity treatment modalities may only be efficacious in asubpopulation of treated offspring as the current data suggestthat the metabolic pathways disrupted will be dependentupon the timing of the early life dietary insult
Abbreviations
CD68 Cluster of differentiation 68CONT ControlDEXA Dual energy X-ray absorptiometryELISA Enzyme-linked immunosorbent assayGLUT2 Glucose transporter 2GP130 Glycoprotein 130HF High fat
HOMA Homeostatic model assessmentHPRT HypoxanthinephophoribosyltransferaseIL InterleukinIns InsulinIR Insulin resistanceIRS Insulin receptor substrateKir62 Potassium channel subunitLIF Leukemia inhibitory factorMHF Maternal high fatOBR Leptin receptorPDE3B Phosphodiesterase 3BPDX-1 Pancreatic and duodenal homeobox-1PI3K Phosphatidylinositide 3-kinasePLHF Pregnancy and lactation high fatSOCS3 Suppressor of cytokine signaling 3STAT3 Signal transducer and activator of
transcription 3TNF-120572 Tumor necrosis factor-alpha
Conflict of Interests
The authors have no conflict of interests with any financialorganization regarding the material discussed in this paper
Author Contributions
Mark H Vickers developed the animal model designed thestudy completed the data analysis and wrote the first draftof the paper Graham J Howie maintained the experimentalcohort performed the gene expression analysis and con-tributed to the original draft paper Clare M Reynolds per-formed the inflammatory related assays and gene expressionwork Deborah M Sloboda provided input into data analysisand interpretation
Acknowledgments
The authors acknowledge the technical support of TaniaKamal with endocrine assays The authors acknowledgefunding support from the Health Research Council of NewZealand and Gravida National Centre for Growth andDevelopment
References
[1] M M Hillemeier C S Weisman C Chuang D S Downs JMcCall-Hosenfeld and F Camacho ldquoTransition to overweightor obesity among women of reproductive agerdquo Journal ofWomenrsquos Health vol 20 no 5 pp 703ndash710 2011
[2] J E Norman and RM Reynolds ldquoThe consequences of obesityand excess weight gain in pregnancyrdquo Proceedings of the Nutri-tion Society vol 70 no 4 pp 450ndash456 2011
[3] H A Abenhaim R A Kinch L Morin A Benjamin and RUsher ldquoEffect of prepregnancy body mass index categories onobstetrical and neonatal outcomesrdquo Archives of Gynecology andObstetrics vol 275 no 1 pp 39ndash43 2007
[4] S DHMalnick andH Knobler ldquoThemedical complications ofobesityrdquo Oxford Journals Medicine vol 99 no 9 pp 565ndash5792006
10 Journal of Nutrition and Metabolism
[5] P M Catalano ldquoManagement of obesity in pregnancyrdquo Obstet-rics and Gynecology vol 109 no 2 pp 419ndash433 2007
[6] L Poston ldquoGestational weight gain influences on the long-termhealth of the childrdquo Current Opinion in Clinical Nutrition andMetabolic Care vol 15 pp 252ndash257 2012
[7] H Hochner Y Friedlander R Calderon-Margalit et al ldquoAsso-ciations of maternal prepregnancy body mass index and gesta-tional weight gain with adult offspring cardiometabolic riskfactors the Jerusalem perinatal family follow-up studyrdquo Circu-lation vol 125 no 11 pp 1381ndash1389 2012
[8] S J Spencer ldquoEarly life programming of obesity the impactof the perinatal environment on the development of obesityand metabolic dysfunction in the offspringrdquo Current DiabetesReviews vol 8 no 1 pp 55ndash68 2012
[9] H Chen D Simar K Lambert J Mercier and M J MorrisldquoMaternal and postnatal overnutrition differentially impactappetite regulators and fuel metabolismrdquo Endocrinology vol149 no 11 pp 5348ndash5356 2008
[10] P M Catalano ldquoObesity and pregnancymdashthe propagation of aviscous cyclerdquo Journal of Clinical Endocrinology and Metabo-lism vol 88 no 8 pp 3505ndash3506 2003
[11] M W Gillman S L Rifas-Shiman K Kleinman E Oken JW Rich-Edwards and EM Taveras ldquoDevelopmental origins ofchildhood overweight potential public health impactrdquo Obesityvol 16 no 7 pp 1651ndash1656 2008
[12] E Oken EM Taveras K P Kleinman JW Rich-Edwards andM W Gillman ldquoGestational weight gain and child adiposity atage 3 yearsrdquoTheAmerican Journal of Obstetrics and Gynecologyvol 196 no 4 pp 322e1ndash322e8 2007
[13] H M Ehrenberg B M Mercer and P M Catalano ldquoThe influ-ence of obesity and diabetes on the prevalence of macrosomiardquoThe American Journal of Obstetrics and Gynecology vol 191 no3 pp 964ndash968 2004
[14] G J Howie D M Sloboda T Kamal and M H VickersldquoMaternal nutritional history predicts obesity in adult offspringindependent of postnatal dietrdquo Journal of Physiology vol 587no 4 pp 905ndash915 2009
[15] D M Sloboda G J Howie A Pleasants P D Gluckman andM H Vickers ldquoPre- and postnatal nutritional histories influ-ence reproductive maturation and ovarian function in the ratrdquoPLoS ONE vol 4 no 8 Article ID e6744 2009
[16] M C Battista M F Hivert K Duval and J P BaillargeonldquoIntergenerational cycle of obesity and diabetes how can wereduce the burdens of these conditions on the health of futuregenerationsrdquo Experimental Diabetes Research vol 2011 ArticleID 596060 19 pages 2011
[17] T J Kieffer and J F Habener ldquoThe adipoinsular axis effects ofleptin on pancreatic 120573-cellsrdquo The American Journal of Physiol-ogy vol 278 no 1 pp E1ndashE14 2000
[18] S L Gray C Donald A Jetha S D Covey and T J KiefferldquoHyperinsulinemia precedes insulin resistance in mice lackingpancreatic 120573-cell leptin signalingrdquo Endocrinology vol 151 no 9pp 4178ndash4186 2010
[19] S D Covey R DWideman CMcDonald et al ldquoThe pancreat-ic 120573 cell is a key site formediating the effects of leptin on glucosehomeostasisrdquo Cell Metabolism vol 4 no 4 pp 291ndash302 2006
[20] K J Dudley DM Sloboda K L Connor J Beltrand andMHVickers ldquoOffspring ofmothers fed a high fat diet display hepaticcell cycle inhibition and associated changes in gene expressionand DNA methylationrdquo PLoS ONE vol 6 no 7 Article IDe21662 2011
[21] K L Connor M H Vickers J Beltrand M J Meaney and DM Sloboda ldquoNature nurture or nutrition Impact of maternalnutrition on maternal care offspring development and repro-ductive functionrdquo Journal of Physiology vol 590 part 9 pp2167ndash2180 2012
[22] J Beltrand D M Sloboda K L Connor M Truong and MH Vickers ldquoThe effect of neonatal leptin antagonism in malerat offspring is dependent upon the interaction between priormaternal nutritional status and post-weaning dietrdquo Journal ofNutrition andMetabolism vol 2012 Article ID 296935 10 pages2012
[23] K L Connor M H Vickers C Cupido E Sirimanne and DM Sloboda ldquoMaternal high fat diet during critical windowsof development alters adrenal cortical and medullary enzymeexpression in adult male rat offspringrdquo Journal of the Develop-mental Origins of Health and Disease vol 1 no 4 pp 245ndash2542010
[24] P J Mark C Sisala K Connor et al ldquoA maternal high-fat dietin rat pregnancy reduces growth of the fetus and the placen-tal junctional zone but not placental labyrinth zone growthrdquoJournal of the Developmental Origins of Health and Disease vol2 no 1 pp 63ndash70 2011
[25] D R Matthews J P Hosker A S Rudenski B A Naylor DF Treacher and R C Turner ldquoHomeostasis model assessmentinsulin resistance and 120573-cell function from fasting plasmaglucose and insulin concentrations in manrdquo Diabetologia vol28 no 7 pp 412ndash419 1985
[26] M P Hermans J C Levy R J Morris and R C TurnerldquoComparison of tests of120573-cell function across a range of glucosetolerance from normal to diabetesrdquo Diabetes vol 48 no 9 pp1779ndash1786 1999
[27] A B Bernal M H Vickers M B Hampton R A Poynton andD M Sloboda ldquoMaternal undernutrition significantly impactsovarian follicle number and increases ovarian oxidative stress inadult rat offspringrdquo PLoS ONE vol 5 no 12 Article ID e155582010
[28] J Vandesompele K de Preter F Pattyn et al ldquoAccurate nor-malization of real-time quantitative RT-PCR data by geometricaveraging of multiple internal control genesrdquo Genome biologyvol 3 no 7 Article ID RESEARCH0034 2002
[29] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the 2-ΔΔCT methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
[30] J Armitage L Poston and P Taylor ldquoDevelopmental originsof obesity and the metabolic syndrome the role of maternalobesityrdquo Frontiers of Hormone Research vol 36 pp 73ndash84 2007
[31] S Zhang L Rattanatray J L Morrison L M Nicholas S Lieand I C McMillen ldquoMaternal obesity and the early originsof childhood obesity weighing up the benefits and costs ofmaternal weight loss in the periconceptional period for theoffspringrdquo Experimental Diabetes Research vol 2011 Article ID585749 10 pages 2011
[32] N M Thompson A M Norman S S Donkin et al ldquoPrenataland postnatal pathways to obesity different underlying mech-anisms different metabolic outcomesrdquo Endocrinology vol 148no 5 pp 2345ndash2354 2007
[33] T Fuentes I Ara A Guadalupe-Grau et al ldquoLeptin receptor170 kDa (OB-R170) protein expression is reduced in obesehuman skeletal muscle a potential mechanism of leptin resis-tancerdquo Experimental Physiology vol 95 no 1 pp 160ndash171 2010
[34] Z Wang Y T Zhou T Kakuma et al ldquoLeptin resistance ofadipocytes in obesity role of suppressors of cytokine signalingrdquo
Journal of Nutrition and Metabolism 11
Biochemical and Biophysical Research Communications vol 277no 1 pp 20ndash26 2000
[35] C Bjoslashrbaeligk K El-Haschimi J D Frantz and J S Flier ldquoThe roleof SOCS-3 in leptin signaling and leptin resistancerdquoThe Journalof Biological Chemistry vol 274 no 42 pp 30059ndash30065 1999
[36] K Laubner T J Kieffer N T Lam X Niu F Jakob and JSeufert ldquoInhibition of preproinsulin gene expression by leptininduction of suppressor of cytokine signaling 3 in pancreatic 120573-cellsrdquo Diabetes vol 54 no 12 pp 3410ndash3417 2005
[37] C Peiser G P McGregor and R E Lang ldquoLeptin receptorexpression and suppressor of cytokine signaling transcriptlevels in high-fat-fed ratsrdquo Life Sciences vol 67 no 24 pp 2971ndash2981 2000
[38] G R Steinberg A C Smith S Wormald P Malenfant CCollier and D J Dyck ldquoEndurance training partially reversesdietary-induced leptin resistance in rodent skeletal musclerdquoTheAmerican Journal of Physiology vol 286 no 1 pp E57ndashE632004
[39] G E White A Cotterill M R Addley E J Soilleux and DR Greaves ldquoSuppressor of cytokine signalling protein SOCS3expression is increased at sites of acute and chronic inflamma-tionrdquo Journal of Molecular Histology vol 42 no 2 pp 137ndash1512011
[40] H YasukawaMOhishiHMori et al ldquoIL-6 induces an anti-in-flammatory response in the absence of SOCS3 inmacrophagesrdquoNature Immunology vol 4 no 6 pp 551ndash556 2003
[41] J Seufert ldquoLeptin effects on pancreatic 120573-cell gene expressionand functionrdquo Diabetes vol 53 no 1 pp S152ndashS158 2004
[42] A S Metlakunta M Sahu and A Sahu ldquoHypothalamicphosphatidylinositol 3-kinase pathway of leptin signaling isimpaired during the development of diet-induced obesity inFVBNmicerdquo Endocrinology vol 149 no 3 pp 1121ndash1128 2008
[43] C A Aspinwall W J Qian M G Roper R N Kulkarni C RKahn and R T Kennedy ldquoRoles of insulin receptor substrate-1phosphatidylinositol 3-kinase and release of intracellular Ca2+stores in insulin-stimulated insulin secretion in 120573-cellsrdquo TheJournal of Biological Chemistry vol 275 no 29 pp 22331ndash223382000
[44] I B Leibiger B Leibiger T Moede and P O BerggrenldquoExocytosis of insulin promotes insulin gene transcription viathe insulin receptorPI-3 kinasep70 s6 kinase and CaM kinasepathwaysrdquoMolecular Cell vol 1 no 6 pp 933ndash938 1998
[45] R L Martin E Perez Y J He R Dawson Jr andW J MillardldquoLeptin resistance is associated with hypothalamic leptin recep-tor mRNA and protein downregulationrdquo Metabolism vol 49no 11 pp 1479ndash1484 2000
[46] J Wilsey and P J Scarpace ldquoCaloric restriction reverses thedeficits in leptin receptor protein and leptin signaling capacityassociated with diet-induced obesity role of leptin in the reg-ulation of hypothalamic long-form leptin receptor expressionrdquoJournal of Endocrinology vol 181 no 2 pp 297ndash306 2004
[47] Y Zhang and P J Scarpace ldquoThe role of leptin in leptin resist-ance and obesityrdquo Physiology and Behavior vol 88 no 3 pp249ndash256 2006
[48] D R Grattan S R Ladyman and R A Augustine ldquoHormonalinduction of leptin resistance during pregnancyrdquoPhysiology andBehavior vol 91 no 4 pp 366ndash374 2007
[49] L RuiM YuanD Frantz S Shoelson andM FWhite ldquoSOCS-1 and SOCS-3 block insulin signaling by ubiquitin-mediateddegradation of IRS1 and IRS2rdquoThe Journal of Biological Chem-istry vol 277 no 44 pp 42394ndash42398 2002
[50] K Ueki T Kondo and C R Kahn ldquoSuppressor of cytokine sig-naling 1 (SOCS-1) and SOCS-3 cause insulin resistance throughinhibition of tyrosine phosphorylation of insulin receptor sub-strate proteins by discrete mechanismsrdquoMolecular and CellularBiology vol 24 no 12 pp 5434ndash5446 2004
[51] S B Jorgensen O rsquoNeill HM L Sylow et al ldquoDeletion of skel-etal muscle SOCS3 prevents insulin resistance in obesityrdquo Dia-betes vol 62 no 1 pp 56ndash64 2013
[52] W Becker ldquoLeptin signal transductionrdquo in Leptin and LeptinAntagonists A Gertler Ed pp 1ndash9 Landes Bioscience AustinTex USA 2009
[53] R N Kulkarni J N Winnay M Daniels et al ldquoAltered func-tion of insulin receptor substrate 1-deficient mouse islets andcultured 120573-cell linesrdquo Journal of Clinical Investigation vol 104no 12 pp R69ndashR75 1999
[54] W Y Kwong A E Wild P Roberts A C Willis and T PFleming ldquoMaternal undernutrition during the preimplantationperiod of rat development causes blastocyst abnormalities andprogramming of postnatal hypertensionrdquoDevelopment vol 127no 19 pp 4195ndash4202 2000
[55] E S Jungheim E L Schoeller K L Marquard E D LoudenJ E Schaffer and K H Moley ldquoDiet-induced obesity modelabnormal oocytes and persistent growth abnormalities in theoffspringrdquo Endocrinology vol 151 no 8 pp 4039ndash4046 2010
[56] A J Watkins A Wilkins C Cunningham et al ldquoLow proteindiet fed exclusively during mouse oocyte maturation leadsto behavioural and cardiovascular abnormalities in offspringrdquoJournal of Physiology vol 586 no 8 pp 2231ndash2244 2008
[57] E Zambrano P M Martınez-Samayoa G L Rodrıguez-Gonzalez and P W Nathanielsz ldquoDietary intervention prior topregnancy reverses metabolic programming in male offspringof obese ratsrdquo Journal of Physiology vol 588 no 10 pp 1791ndash1799 2010
[58] M S Patel andM Srinivasan ldquoMetabolic programming causesand consequencesrdquoThe Journal of Biological Chemistry vol 277no 3 pp 1629ndash1632 2002
[59] R A Simmons ldquoRole of metabolic programming in the patho-genesis of 120573-cell failure in postnatal liferdquo Reviews in Endocrineand Metabolic Disorders vol 8 no 2 pp 95ndash104 2007
[60] I B Leibiger B Leibiger and P O Berggren ldquoInsulin signalingin the pancreatic 120573-cellrdquoAnnual Review of Nutrition vol 28 pp233ndash251 2008
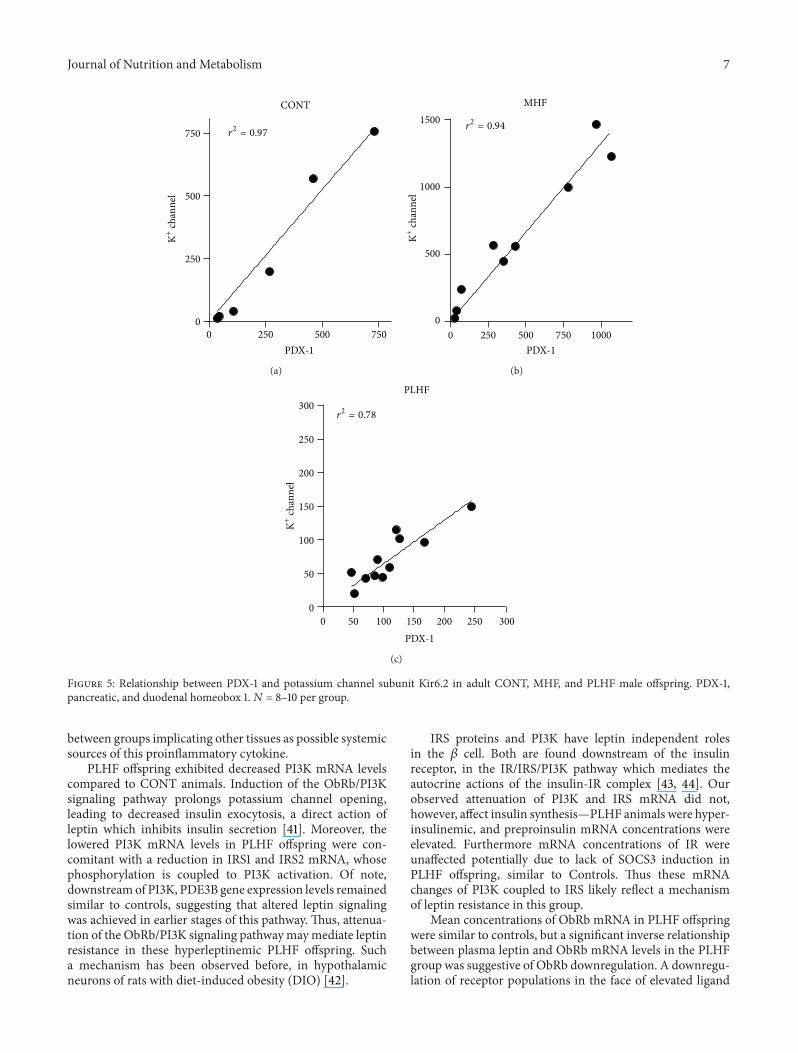
Journal of Nutrition and Metabolism 7
CONT
0 250 500 7500
250
500
750
PDX-1
1199032= 097
K+ch
anne
l
(a)
MHF
0 250 500 750 10000
500
1000
15001199032= 094
PDX-1
K+ch
anne
l
(b)
PLHF
0 50 100 150 200 250 3000
50
100
150
200
250
3001199032= 078
PDX-1
K+ch
anne
l
(c)
Figure 5 Relationship between PDX-1 and potassium channel subunit Kir62 in adult CONT MHF and PLHF male offspring PDX-1pancreatic and duodenal homeobox 1119873 = 8ndash10 per group
between groups implicating other tissues as possible systemicsources of this proinflammatory cytokine
PLHF offspring exhibited decreased PI3K mRNA levelscompared to CONT animals Induction of the ObRbPI3Ksignaling pathway prolongs potassium channel openingleading to decreased insulin exocytosis a direct action ofleptin which inhibits insulin secretion [41] Moreover thelowered PI3K mRNA levels in PLHF offspring were con-comitant with a reduction in IRS1 and IRS2 mRNA whosephosphorylation is coupled to PI3K activation Of notedownstreamof PI3K PDE3B gene expression levels remainedsimilar to controls suggesting that altered leptin signalingwas achieved in earlier stages of this pathway Thus attenua-tion of the ObRbPI3K signaling pathwaymaymediate leptinresistance in these hyperleptinemic PLHF offspring Sucha mechanism has been observed before in hypothalamicneurons of rats with diet-induced obesity (DIO) [42]
IRS proteins and PI3K have leptin independent rolesin the 120573 cell Both are found downstream of the insulinreceptor in the IRIRSPI3K pathway which mediates theautocrine actions of the insulin-IR complex [43 44] Ourobserved attenuation of PI3K and IRS mRNA did nothowever affect insulin synthesismdashPLHF animals were hyper-insulinemic and preproinsulin mRNA concentrations wereelevated Furthermore mRNA concentrations of IR wereunaffected potentially due to lack of SOCS3 induction inPLHF offspring similar to Controls Thus these mRNAchanges of PI3K coupled to IRS likely reflect a mechanismof leptin resistance in this group
Mean concentrations of ObRb mRNA in PLHF offspringwere similar to controls but a significant inverse relationshipbetween plasma leptin and ObRb mRNA levels in the PLHFgroup was suggestive of ObRb downregulation A downregu-lation of receptor populations in the face of elevated ligand
8 Journal of Nutrition and Metabolism
CONT MHF PLHF0
50
100
150
200
TNF-120572
mRN
A (
of c
ontro
l)
lowast
lowast
(a)
CD68
mRN
A (
of c
ontro
l)
CONT MHF PLHF0
50
100
150
200
lowast
lowast
(b)
IL1-
R1 m
RNA
( o
f con
trol)
CONT MHF PLHF0
50
100
150
200lowast
(c)
Figure 6 Pancreatic mRNA expression of TNF-120572 (a) CD68 (b) and IL1-R1 (c) in adult CONT MHF and PLHF offspring TNF-120572 tumornecrosis factor-alpha CD68 cluster of differentiation 68 IL1-R1 interleukin 1 receptor type 1 Data are means plusmn SEM 119899 = 10ndash18 per grouplowast
119875 lt 005
concentrations is not uncommon in resistant states It isseen for example in the hypothalamus in the presence ofhyperleptinemia [45ndash47] and during pregnancy which is anatural state of leptin resistance [48]
A rise in IRS1 and IRS2 mRNA associated with elevatedObRb and SOCS3 mRNA in MHF offspring may seemunexpected since SOCS3 is known to target insulin receptorsignaling by interference with IRS action [49] However sev-eral studies have demonstrated that SOCS3 mediates insulinresistance in both adipocytes and hepatocytes through post-translational modifications such as inhibition of IRS tyro-sine phosphorylation [50] and targeted ubiquitin-mediateddegradation of IRS1 and IRS2 without effecting transcription[49 51] IRS proteins are involved in both leptin signalingand insulin signaling so altered gene expression levels mayrepresent changes in either (or both) pathways although IRSresponse to leptin is reportedly much lower than that seenwith insulin [52] We observed an increase in IRS1 expressionin MHF offspring compared to Controls This upregulationof IRS1 expression may be related to the increased insulin
production of the MHF offspring given the known role ofIRS1 in the regulation of insulin secretion [53]
SOCS3 is also reported to repress preproinsulin genetranscription [41] although other mechanisms clearly coun-tered this action inMHFoffspring since Ins1 and Ins2mRNAexpression levels remained elevated This observation wasconfirmed by the significantly raised levels of circulatingplasma insulin and the lack of any correlation in levels ofmRNA between SOCS3 and Ins1 or Ins2
Some of the observed differences may relate to theinitial hyperphagia observed in the PLHF maternal groupas reported previously [14] Since rodents primarily eat ona calorific basis they will passively over-consume calorieswhen initially placed on an HF diet and self-regulate backto a normal total caloric intake over a period of a few daysIt has been shown in other rodent models that alterations innutrition around the time of implantation can lead to lastingmetabolic effects in the offspring [54] Maternal obesity hasbeen shown to exert adverse effects as early as the oocyteand preimplantation embryo stage and that these effects may
Journal of Nutrition and Metabolism 9
contribute to the lasting morbidity observed in offspring[55] Similarly even a very short period of altered nutrition(restricted to one ovulatory cycle) prior to natural mating canelicit lasting effects on the offspring [56] and this representsa further point of difference between the MHF and PLHFgroups
The present study was not designed to incorporate theeffect of a reversal paradigm whereby mothers fed thepre-conceptional HF diet were placed on a standard dietthroughout pregnancy and lactation However while there isevidence that a period of dietary restriction in obese mothersmay ablate the programming of obesity in offspring [57]it may be associated with an activation of the stress axisin the offspring [31] Further the data are inconsistent ithas also been shown that dams with diet-induced obesityfed a standard diet throughout pregnancy and lactation stilltransmit the propensity for obesity and metabolic disease totheir offspring [58]
We recognize that gene expression analysis on wholepancreatic tissue is a limitation of the present study Dueto the size of the cohort and logistical considerations onlywhole pancreatic tissue was a possibility whereas ideallyonly pure islet populations would have been examined [59]However for the key genes presented such as ObRb SOCS3and IRS expression has been shown to be primarily confinedto the endocrine islets and not been detected in the exocrinepancreas [39 41 60] therefore we are confident that theobserved changes in the genes studied represent changes inthe islets Further although we have shown that differencesin pancreatic gene expression in offspring are dependentupon the timing of maternal high fat diet exposure this doesnot itself imply causation in the development of the obesephenotypemdashother relevant pathways including liver skeletalmuscle and fat metabolism will also be integrated into theprogrammed phenotype development
This study has demonstrated enduring deleterious effectsof a maternal HF diet during early windows of develop-ment Maternal HF feeding both before conception andorthrough pregnancy and lactation induced obesity hyper-insulinemia and hyperleptinemia in adult offspring eventhough offspring were fed a standard diet postnatally Timingof exposure to the maternal HF diet differentially alteredgene expression in signaling pathways downstream of thepancreatic leptin receptor that mediated leptin resistanceThis also raises the possibility that despite common offspringobesity treatment modalities may only be efficacious in asubpopulation of treated offspring as the current data suggestthat the metabolic pathways disrupted will be dependentupon the timing of the early life dietary insult
Abbreviations
CD68 Cluster of differentiation 68CONT ControlDEXA Dual energy X-ray absorptiometryELISA Enzyme-linked immunosorbent assayGLUT2 Glucose transporter 2GP130 Glycoprotein 130HF High fat
HOMA Homeostatic model assessmentHPRT HypoxanthinephophoribosyltransferaseIL InterleukinIns InsulinIR Insulin resistanceIRS Insulin receptor substrateKir62 Potassium channel subunitLIF Leukemia inhibitory factorMHF Maternal high fatOBR Leptin receptorPDE3B Phosphodiesterase 3BPDX-1 Pancreatic and duodenal homeobox-1PI3K Phosphatidylinositide 3-kinasePLHF Pregnancy and lactation high fatSOCS3 Suppressor of cytokine signaling 3STAT3 Signal transducer and activator of
transcription 3TNF-120572 Tumor necrosis factor-alpha
Conflict of Interests
The authors have no conflict of interests with any financialorganization regarding the material discussed in this paper
Author Contributions
Mark H Vickers developed the animal model designed thestudy completed the data analysis and wrote the first draftof the paper Graham J Howie maintained the experimentalcohort performed the gene expression analysis and con-tributed to the original draft paper Clare M Reynolds per-formed the inflammatory related assays and gene expressionwork Deborah M Sloboda provided input into data analysisand interpretation
Acknowledgments
The authors acknowledge the technical support of TaniaKamal with endocrine assays The authors acknowledgefunding support from the Health Research Council of NewZealand and Gravida National Centre for Growth andDevelopment
References
[1] M M Hillemeier C S Weisman C Chuang D S Downs JMcCall-Hosenfeld and F Camacho ldquoTransition to overweightor obesity among women of reproductive agerdquo Journal ofWomenrsquos Health vol 20 no 5 pp 703ndash710 2011
[2] J E Norman and RM Reynolds ldquoThe consequences of obesityand excess weight gain in pregnancyrdquo Proceedings of the Nutri-tion Society vol 70 no 4 pp 450ndash456 2011
[3] H A Abenhaim R A Kinch L Morin A Benjamin and RUsher ldquoEffect of prepregnancy body mass index categories onobstetrical and neonatal outcomesrdquo Archives of Gynecology andObstetrics vol 275 no 1 pp 39ndash43 2007
[4] S DHMalnick andH Knobler ldquoThemedical complications ofobesityrdquo Oxford Journals Medicine vol 99 no 9 pp 565ndash5792006
10 Journal of Nutrition and Metabolism
[5] P M Catalano ldquoManagement of obesity in pregnancyrdquo Obstet-rics and Gynecology vol 109 no 2 pp 419ndash433 2007
[6] L Poston ldquoGestational weight gain influences on the long-termhealth of the childrdquo Current Opinion in Clinical Nutrition andMetabolic Care vol 15 pp 252ndash257 2012
[7] H Hochner Y Friedlander R Calderon-Margalit et al ldquoAsso-ciations of maternal prepregnancy body mass index and gesta-tional weight gain with adult offspring cardiometabolic riskfactors the Jerusalem perinatal family follow-up studyrdquo Circu-lation vol 125 no 11 pp 1381ndash1389 2012
[8] S J Spencer ldquoEarly life programming of obesity the impactof the perinatal environment on the development of obesityand metabolic dysfunction in the offspringrdquo Current DiabetesReviews vol 8 no 1 pp 55ndash68 2012
[9] H Chen D Simar K Lambert J Mercier and M J MorrisldquoMaternal and postnatal overnutrition differentially impactappetite regulators and fuel metabolismrdquo Endocrinology vol149 no 11 pp 5348ndash5356 2008
[10] P M Catalano ldquoObesity and pregnancymdashthe propagation of aviscous cyclerdquo Journal of Clinical Endocrinology and Metabo-lism vol 88 no 8 pp 3505ndash3506 2003
[11] M W Gillman S L Rifas-Shiman K Kleinman E Oken JW Rich-Edwards and EM Taveras ldquoDevelopmental origins ofchildhood overweight potential public health impactrdquo Obesityvol 16 no 7 pp 1651ndash1656 2008
[12] E Oken EM Taveras K P Kleinman JW Rich-Edwards andM W Gillman ldquoGestational weight gain and child adiposity atage 3 yearsrdquoTheAmerican Journal of Obstetrics and Gynecologyvol 196 no 4 pp 322e1ndash322e8 2007
[13] H M Ehrenberg B M Mercer and P M Catalano ldquoThe influ-ence of obesity and diabetes on the prevalence of macrosomiardquoThe American Journal of Obstetrics and Gynecology vol 191 no3 pp 964ndash968 2004
[14] G J Howie D M Sloboda T Kamal and M H VickersldquoMaternal nutritional history predicts obesity in adult offspringindependent of postnatal dietrdquo Journal of Physiology vol 587no 4 pp 905ndash915 2009
[15] D M Sloboda G J Howie A Pleasants P D Gluckman andM H Vickers ldquoPre- and postnatal nutritional histories influ-ence reproductive maturation and ovarian function in the ratrdquoPLoS ONE vol 4 no 8 Article ID e6744 2009
[16] M C Battista M F Hivert K Duval and J P BaillargeonldquoIntergenerational cycle of obesity and diabetes how can wereduce the burdens of these conditions on the health of futuregenerationsrdquo Experimental Diabetes Research vol 2011 ArticleID 596060 19 pages 2011
[17] T J Kieffer and J F Habener ldquoThe adipoinsular axis effects ofleptin on pancreatic 120573-cellsrdquo The American Journal of Physiol-ogy vol 278 no 1 pp E1ndashE14 2000
[18] S L Gray C Donald A Jetha S D Covey and T J KiefferldquoHyperinsulinemia precedes insulin resistance in mice lackingpancreatic 120573-cell leptin signalingrdquo Endocrinology vol 151 no 9pp 4178ndash4186 2010
[19] S D Covey R DWideman CMcDonald et al ldquoThe pancreat-ic 120573 cell is a key site formediating the effects of leptin on glucosehomeostasisrdquo Cell Metabolism vol 4 no 4 pp 291ndash302 2006
[20] K J Dudley DM Sloboda K L Connor J Beltrand andMHVickers ldquoOffspring ofmothers fed a high fat diet display hepaticcell cycle inhibition and associated changes in gene expressionand DNA methylationrdquo PLoS ONE vol 6 no 7 Article IDe21662 2011
[21] K L Connor M H Vickers J Beltrand M J Meaney and DM Sloboda ldquoNature nurture or nutrition Impact of maternalnutrition on maternal care offspring development and repro-ductive functionrdquo Journal of Physiology vol 590 part 9 pp2167ndash2180 2012
[22] J Beltrand D M Sloboda K L Connor M Truong and MH Vickers ldquoThe effect of neonatal leptin antagonism in malerat offspring is dependent upon the interaction between priormaternal nutritional status and post-weaning dietrdquo Journal ofNutrition andMetabolism vol 2012 Article ID 296935 10 pages2012
[23] K L Connor M H Vickers C Cupido E Sirimanne and DM Sloboda ldquoMaternal high fat diet during critical windowsof development alters adrenal cortical and medullary enzymeexpression in adult male rat offspringrdquo Journal of the Develop-mental Origins of Health and Disease vol 1 no 4 pp 245ndash2542010
[24] P J Mark C Sisala K Connor et al ldquoA maternal high-fat dietin rat pregnancy reduces growth of the fetus and the placen-tal junctional zone but not placental labyrinth zone growthrdquoJournal of the Developmental Origins of Health and Disease vol2 no 1 pp 63ndash70 2011
[25] D R Matthews J P Hosker A S Rudenski B A Naylor DF Treacher and R C Turner ldquoHomeostasis model assessmentinsulin resistance and 120573-cell function from fasting plasmaglucose and insulin concentrations in manrdquo Diabetologia vol28 no 7 pp 412ndash419 1985
[26] M P Hermans J C Levy R J Morris and R C TurnerldquoComparison of tests of120573-cell function across a range of glucosetolerance from normal to diabetesrdquo Diabetes vol 48 no 9 pp1779ndash1786 1999
[27] A B Bernal M H Vickers M B Hampton R A Poynton andD M Sloboda ldquoMaternal undernutrition significantly impactsovarian follicle number and increases ovarian oxidative stress inadult rat offspringrdquo PLoS ONE vol 5 no 12 Article ID e155582010
[28] J Vandesompele K de Preter F Pattyn et al ldquoAccurate nor-malization of real-time quantitative RT-PCR data by geometricaveraging of multiple internal control genesrdquo Genome biologyvol 3 no 7 Article ID RESEARCH0034 2002
[29] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the 2-ΔΔCT methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
[30] J Armitage L Poston and P Taylor ldquoDevelopmental originsof obesity and the metabolic syndrome the role of maternalobesityrdquo Frontiers of Hormone Research vol 36 pp 73ndash84 2007
[31] S Zhang L Rattanatray J L Morrison L M Nicholas S Lieand I C McMillen ldquoMaternal obesity and the early originsof childhood obesity weighing up the benefits and costs ofmaternal weight loss in the periconceptional period for theoffspringrdquo Experimental Diabetes Research vol 2011 Article ID585749 10 pages 2011
[32] N M Thompson A M Norman S S Donkin et al ldquoPrenataland postnatal pathways to obesity different underlying mech-anisms different metabolic outcomesrdquo Endocrinology vol 148no 5 pp 2345ndash2354 2007
[33] T Fuentes I Ara A Guadalupe-Grau et al ldquoLeptin receptor170 kDa (OB-R170) protein expression is reduced in obesehuman skeletal muscle a potential mechanism of leptin resis-tancerdquo Experimental Physiology vol 95 no 1 pp 160ndash171 2010
[34] Z Wang Y T Zhou T Kakuma et al ldquoLeptin resistance ofadipocytes in obesity role of suppressors of cytokine signalingrdquo
Journal of Nutrition and Metabolism 11
Biochemical and Biophysical Research Communications vol 277no 1 pp 20ndash26 2000
[35] C Bjoslashrbaeligk K El-Haschimi J D Frantz and J S Flier ldquoThe roleof SOCS-3 in leptin signaling and leptin resistancerdquoThe Journalof Biological Chemistry vol 274 no 42 pp 30059ndash30065 1999
[36] K Laubner T J Kieffer N T Lam X Niu F Jakob and JSeufert ldquoInhibition of preproinsulin gene expression by leptininduction of suppressor of cytokine signaling 3 in pancreatic 120573-cellsrdquo Diabetes vol 54 no 12 pp 3410ndash3417 2005
[37] C Peiser G P McGregor and R E Lang ldquoLeptin receptorexpression and suppressor of cytokine signaling transcriptlevels in high-fat-fed ratsrdquo Life Sciences vol 67 no 24 pp 2971ndash2981 2000
[38] G R Steinberg A C Smith S Wormald P Malenfant CCollier and D J Dyck ldquoEndurance training partially reversesdietary-induced leptin resistance in rodent skeletal musclerdquoTheAmerican Journal of Physiology vol 286 no 1 pp E57ndashE632004
[39] G E White A Cotterill M R Addley E J Soilleux and DR Greaves ldquoSuppressor of cytokine signalling protein SOCS3expression is increased at sites of acute and chronic inflamma-tionrdquo Journal of Molecular Histology vol 42 no 2 pp 137ndash1512011
[40] H YasukawaMOhishiHMori et al ldquoIL-6 induces an anti-in-flammatory response in the absence of SOCS3 inmacrophagesrdquoNature Immunology vol 4 no 6 pp 551ndash556 2003
[41] J Seufert ldquoLeptin effects on pancreatic 120573-cell gene expressionand functionrdquo Diabetes vol 53 no 1 pp S152ndashS158 2004
[42] A S Metlakunta M Sahu and A Sahu ldquoHypothalamicphosphatidylinositol 3-kinase pathway of leptin signaling isimpaired during the development of diet-induced obesity inFVBNmicerdquo Endocrinology vol 149 no 3 pp 1121ndash1128 2008
[43] C A Aspinwall W J Qian M G Roper R N Kulkarni C RKahn and R T Kennedy ldquoRoles of insulin receptor substrate-1phosphatidylinositol 3-kinase and release of intracellular Ca2+stores in insulin-stimulated insulin secretion in 120573-cellsrdquo TheJournal of Biological Chemistry vol 275 no 29 pp 22331ndash223382000
[44] I B Leibiger B Leibiger T Moede and P O BerggrenldquoExocytosis of insulin promotes insulin gene transcription viathe insulin receptorPI-3 kinasep70 s6 kinase and CaM kinasepathwaysrdquoMolecular Cell vol 1 no 6 pp 933ndash938 1998
[45] R L Martin E Perez Y J He R Dawson Jr andW J MillardldquoLeptin resistance is associated with hypothalamic leptin recep-tor mRNA and protein downregulationrdquo Metabolism vol 49no 11 pp 1479ndash1484 2000
[46] J Wilsey and P J Scarpace ldquoCaloric restriction reverses thedeficits in leptin receptor protein and leptin signaling capacityassociated with diet-induced obesity role of leptin in the reg-ulation of hypothalamic long-form leptin receptor expressionrdquoJournal of Endocrinology vol 181 no 2 pp 297ndash306 2004
[47] Y Zhang and P J Scarpace ldquoThe role of leptin in leptin resist-ance and obesityrdquo Physiology and Behavior vol 88 no 3 pp249ndash256 2006
[48] D R Grattan S R Ladyman and R A Augustine ldquoHormonalinduction of leptin resistance during pregnancyrdquoPhysiology andBehavior vol 91 no 4 pp 366ndash374 2007
[49] L RuiM YuanD Frantz S Shoelson andM FWhite ldquoSOCS-1 and SOCS-3 block insulin signaling by ubiquitin-mediateddegradation of IRS1 and IRS2rdquoThe Journal of Biological Chem-istry vol 277 no 44 pp 42394ndash42398 2002
[50] K Ueki T Kondo and C R Kahn ldquoSuppressor of cytokine sig-naling 1 (SOCS-1) and SOCS-3 cause insulin resistance throughinhibition of tyrosine phosphorylation of insulin receptor sub-strate proteins by discrete mechanismsrdquoMolecular and CellularBiology vol 24 no 12 pp 5434ndash5446 2004
[51] S B Jorgensen O rsquoNeill HM L Sylow et al ldquoDeletion of skel-etal muscle SOCS3 prevents insulin resistance in obesityrdquo Dia-betes vol 62 no 1 pp 56ndash64 2013
[52] W Becker ldquoLeptin signal transductionrdquo in Leptin and LeptinAntagonists A Gertler Ed pp 1ndash9 Landes Bioscience AustinTex USA 2009
[53] R N Kulkarni J N Winnay M Daniels et al ldquoAltered func-tion of insulin receptor substrate 1-deficient mouse islets andcultured 120573-cell linesrdquo Journal of Clinical Investigation vol 104no 12 pp R69ndashR75 1999
[54] W Y Kwong A E Wild P Roberts A C Willis and T PFleming ldquoMaternal undernutrition during the preimplantationperiod of rat development causes blastocyst abnormalities andprogramming of postnatal hypertensionrdquoDevelopment vol 127no 19 pp 4195ndash4202 2000
[55] E S Jungheim E L Schoeller K L Marquard E D LoudenJ E Schaffer and K H Moley ldquoDiet-induced obesity modelabnormal oocytes and persistent growth abnormalities in theoffspringrdquo Endocrinology vol 151 no 8 pp 4039ndash4046 2010
[56] A J Watkins A Wilkins C Cunningham et al ldquoLow proteindiet fed exclusively during mouse oocyte maturation leadsto behavioural and cardiovascular abnormalities in offspringrdquoJournal of Physiology vol 586 no 8 pp 2231ndash2244 2008
[57] E Zambrano P M Martınez-Samayoa G L Rodrıguez-Gonzalez and P W Nathanielsz ldquoDietary intervention prior topregnancy reverses metabolic programming in male offspringof obese ratsrdquo Journal of Physiology vol 588 no 10 pp 1791ndash1799 2010
[58] M S Patel andM Srinivasan ldquoMetabolic programming causesand consequencesrdquoThe Journal of Biological Chemistry vol 277no 3 pp 1629ndash1632 2002
[59] R A Simmons ldquoRole of metabolic programming in the patho-genesis of 120573-cell failure in postnatal liferdquo Reviews in Endocrineand Metabolic Disorders vol 8 no 2 pp 95ndash104 2007
[60] I B Leibiger B Leibiger and P O Berggren ldquoInsulin signalingin the pancreatic 120573-cellrdquoAnnual Review of Nutrition vol 28 pp233ndash251 2008
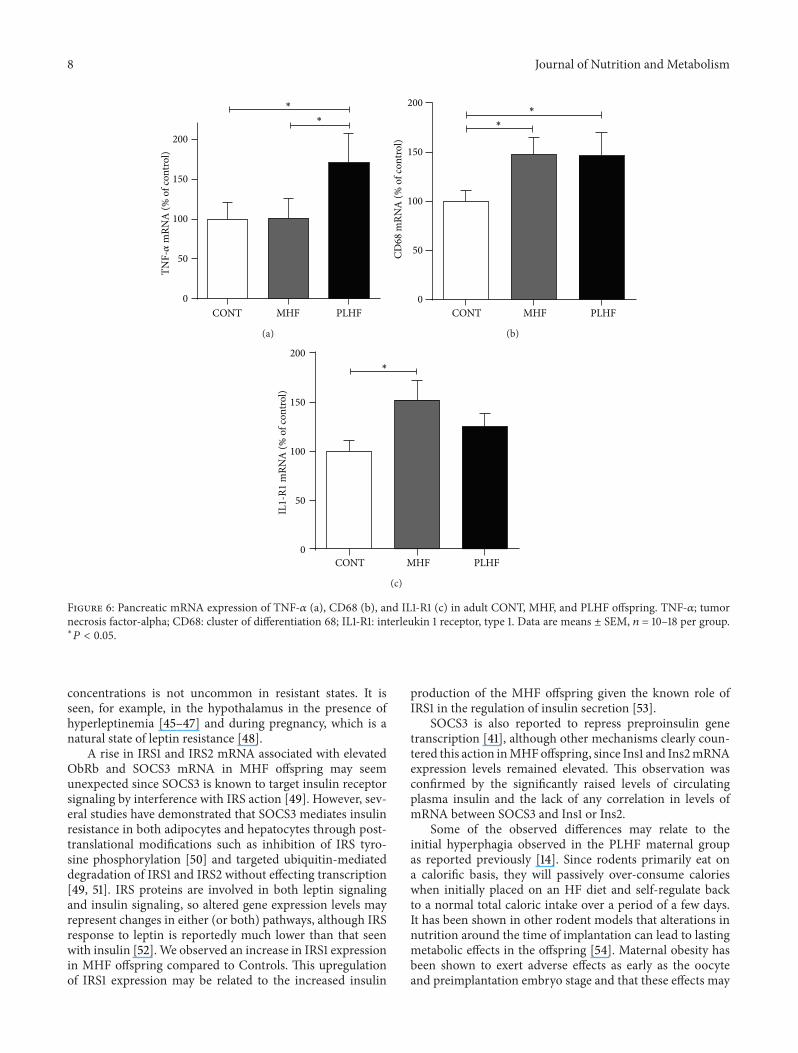
8 Journal of Nutrition and Metabolism
CONT MHF PLHF0
50
100
150
200
TNF-120572
mRN
A (
of c
ontro
l)
lowast
lowast
(a)
CD68
mRN
A (
of c
ontro
l)
CONT MHF PLHF0
50
100
150
200
lowast
lowast
(b)
IL1-
R1 m
RNA
( o
f con
trol)
CONT MHF PLHF0
50
100
150
200lowast
(c)
Figure 6 Pancreatic mRNA expression of TNF-120572 (a) CD68 (b) and IL1-R1 (c) in adult CONT MHF and PLHF offspring TNF-120572 tumornecrosis factor-alpha CD68 cluster of differentiation 68 IL1-R1 interleukin 1 receptor type 1 Data are means plusmn SEM 119899 = 10ndash18 per grouplowast
119875 lt 005
concentrations is not uncommon in resistant states It isseen for example in the hypothalamus in the presence ofhyperleptinemia [45ndash47] and during pregnancy which is anatural state of leptin resistance [48]
A rise in IRS1 and IRS2 mRNA associated with elevatedObRb and SOCS3 mRNA in MHF offspring may seemunexpected since SOCS3 is known to target insulin receptorsignaling by interference with IRS action [49] However sev-eral studies have demonstrated that SOCS3 mediates insulinresistance in both adipocytes and hepatocytes through post-translational modifications such as inhibition of IRS tyro-sine phosphorylation [50] and targeted ubiquitin-mediateddegradation of IRS1 and IRS2 without effecting transcription[49 51] IRS proteins are involved in both leptin signalingand insulin signaling so altered gene expression levels mayrepresent changes in either (or both) pathways although IRSresponse to leptin is reportedly much lower than that seenwith insulin [52] We observed an increase in IRS1 expressionin MHF offspring compared to Controls This upregulationof IRS1 expression may be related to the increased insulin
production of the MHF offspring given the known role ofIRS1 in the regulation of insulin secretion [53]
SOCS3 is also reported to repress preproinsulin genetranscription [41] although other mechanisms clearly coun-tered this action inMHFoffspring since Ins1 and Ins2mRNAexpression levels remained elevated This observation wasconfirmed by the significantly raised levels of circulatingplasma insulin and the lack of any correlation in levels ofmRNA between SOCS3 and Ins1 or Ins2
Some of the observed differences may relate to theinitial hyperphagia observed in the PLHF maternal groupas reported previously [14] Since rodents primarily eat ona calorific basis they will passively over-consume calorieswhen initially placed on an HF diet and self-regulate backto a normal total caloric intake over a period of a few daysIt has been shown in other rodent models that alterations innutrition around the time of implantation can lead to lastingmetabolic effects in the offspring [54] Maternal obesity hasbeen shown to exert adverse effects as early as the oocyteand preimplantation embryo stage and that these effects may
Journal of Nutrition and Metabolism 9
contribute to the lasting morbidity observed in offspring[55] Similarly even a very short period of altered nutrition(restricted to one ovulatory cycle) prior to natural mating canelicit lasting effects on the offspring [56] and this representsa further point of difference between the MHF and PLHFgroups
The present study was not designed to incorporate theeffect of a reversal paradigm whereby mothers fed thepre-conceptional HF diet were placed on a standard dietthroughout pregnancy and lactation However while there isevidence that a period of dietary restriction in obese mothersmay ablate the programming of obesity in offspring [57]it may be associated with an activation of the stress axisin the offspring [31] Further the data are inconsistent ithas also been shown that dams with diet-induced obesityfed a standard diet throughout pregnancy and lactation stilltransmit the propensity for obesity and metabolic disease totheir offspring [58]
We recognize that gene expression analysis on wholepancreatic tissue is a limitation of the present study Dueto the size of the cohort and logistical considerations onlywhole pancreatic tissue was a possibility whereas ideallyonly pure islet populations would have been examined [59]However for the key genes presented such as ObRb SOCS3and IRS expression has been shown to be primarily confinedto the endocrine islets and not been detected in the exocrinepancreas [39 41 60] therefore we are confident that theobserved changes in the genes studied represent changes inthe islets Further although we have shown that differencesin pancreatic gene expression in offspring are dependentupon the timing of maternal high fat diet exposure this doesnot itself imply causation in the development of the obesephenotypemdashother relevant pathways including liver skeletalmuscle and fat metabolism will also be integrated into theprogrammed phenotype development
This study has demonstrated enduring deleterious effectsof a maternal HF diet during early windows of develop-ment Maternal HF feeding both before conception andorthrough pregnancy and lactation induced obesity hyper-insulinemia and hyperleptinemia in adult offspring eventhough offspring were fed a standard diet postnatally Timingof exposure to the maternal HF diet differentially alteredgene expression in signaling pathways downstream of thepancreatic leptin receptor that mediated leptin resistanceThis also raises the possibility that despite common offspringobesity treatment modalities may only be efficacious in asubpopulation of treated offspring as the current data suggestthat the metabolic pathways disrupted will be dependentupon the timing of the early life dietary insult
Abbreviations
CD68 Cluster of differentiation 68CONT ControlDEXA Dual energy X-ray absorptiometryELISA Enzyme-linked immunosorbent assayGLUT2 Glucose transporter 2GP130 Glycoprotein 130HF High fat
HOMA Homeostatic model assessmentHPRT HypoxanthinephophoribosyltransferaseIL InterleukinIns InsulinIR Insulin resistanceIRS Insulin receptor substrateKir62 Potassium channel subunitLIF Leukemia inhibitory factorMHF Maternal high fatOBR Leptin receptorPDE3B Phosphodiesterase 3BPDX-1 Pancreatic and duodenal homeobox-1PI3K Phosphatidylinositide 3-kinasePLHF Pregnancy and lactation high fatSOCS3 Suppressor of cytokine signaling 3STAT3 Signal transducer and activator of
transcription 3TNF-120572 Tumor necrosis factor-alpha
Conflict of Interests
The authors have no conflict of interests with any financialorganization regarding the material discussed in this paper
Author Contributions
Mark H Vickers developed the animal model designed thestudy completed the data analysis and wrote the first draftof the paper Graham J Howie maintained the experimentalcohort performed the gene expression analysis and con-tributed to the original draft paper Clare M Reynolds per-formed the inflammatory related assays and gene expressionwork Deborah M Sloboda provided input into data analysisand interpretation
Acknowledgments
The authors acknowledge the technical support of TaniaKamal with endocrine assays The authors acknowledgefunding support from the Health Research Council of NewZealand and Gravida National Centre for Growth andDevelopment
References
[1] M M Hillemeier C S Weisman C Chuang D S Downs JMcCall-Hosenfeld and F Camacho ldquoTransition to overweightor obesity among women of reproductive agerdquo Journal ofWomenrsquos Health vol 20 no 5 pp 703ndash710 2011
[2] J E Norman and RM Reynolds ldquoThe consequences of obesityand excess weight gain in pregnancyrdquo Proceedings of the Nutri-tion Society vol 70 no 4 pp 450ndash456 2011
[3] H A Abenhaim R A Kinch L Morin A Benjamin and RUsher ldquoEffect of prepregnancy body mass index categories onobstetrical and neonatal outcomesrdquo Archives of Gynecology andObstetrics vol 275 no 1 pp 39ndash43 2007
[4] S DHMalnick andH Knobler ldquoThemedical complications ofobesityrdquo Oxford Journals Medicine vol 99 no 9 pp 565ndash5792006
10 Journal of Nutrition and Metabolism
[5] P M Catalano ldquoManagement of obesity in pregnancyrdquo Obstet-rics and Gynecology vol 109 no 2 pp 419ndash433 2007
[6] L Poston ldquoGestational weight gain influences on the long-termhealth of the childrdquo Current Opinion in Clinical Nutrition andMetabolic Care vol 15 pp 252ndash257 2012
[7] H Hochner Y Friedlander R Calderon-Margalit et al ldquoAsso-ciations of maternal prepregnancy body mass index and gesta-tional weight gain with adult offspring cardiometabolic riskfactors the Jerusalem perinatal family follow-up studyrdquo Circu-lation vol 125 no 11 pp 1381ndash1389 2012
[8] S J Spencer ldquoEarly life programming of obesity the impactof the perinatal environment on the development of obesityand metabolic dysfunction in the offspringrdquo Current DiabetesReviews vol 8 no 1 pp 55ndash68 2012
[9] H Chen D Simar K Lambert J Mercier and M J MorrisldquoMaternal and postnatal overnutrition differentially impactappetite regulators and fuel metabolismrdquo Endocrinology vol149 no 11 pp 5348ndash5356 2008
[10] P M Catalano ldquoObesity and pregnancymdashthe propagation of aviscous cyclerdquo Journal of Clinical Endocrinology and Metabo-lism vol 88 no 8 pp 3505ndash3506 2003
[11] M W Gillman S L Rifas-Shiman K Kleinman E Oken JW Rich-Edwards and EM Taveras ldquoDevelopmental origins ofchildhood overweight potential public health impactrdquo Obesityvol 16 no 7 pp 1651ndash1656 2008
[12] E Oken EM Taveras K P Kleinman JW Rich-Edwards andM W Gillman ldquoGestational weight gain and child adiposity atage 3 yearsrdquoTheAmerican Journal of Obstetrics and Gynecologyvol 196 no 4 pp 322e1ndash322e8 2007
[13] H M Ehrenberg B M Mercer and P M Catalano ldquoThe influ-ence of obesity and diabetes on the prevalence of macrosomiardquoThe American Journal of Obstetrics and Gynecology vol 191 no3 pp 964ndash968 2004
[14] G J Howie D M Sloboda T Kamal and M H VickersldquoMaternal nutritional history predicts obesity in adult offspringindependent of postnatal dietrdquo Journal of Physiology vol 587no 4 pp 905ndash915 2009
[15] D M Sloboda G J Howie A Pleasants P D Gluckman andM H Vickers ldquoPre- and postnatal nutritional histories influ-ence reproductive maturation and ovarian function in the ratrdquoPLoS ONE vol 4 no 8 Article ID e6744 2009
[16] M C Battista M F Hivert K Duval and J P BaillargeonldquoIntergenerational cycle of obesity and diabetes how can wereduce the burdens of these conditions on the health of futuregenerationsrdquo Experimental Diabetes Research vol 2011 ArticleID 596060 19 pages 2011
[17] T J Kieffer and J F Habener ldquoThe adipoinsular axis effects ofleptin on pancreatic 120573-cellsrdquo The American Journal of Physiol-ogy vol 278 no 1 pp E1ndashE14 2000
[18] S L Gray C Donald A Jetha S D Covey and T J KiefferldquoHyperinsulinemia precedes insulin resistance in mice lackingpancreatic 120573-cell leptin signalingrdquo Endocrinology vol 151 no 9pp 4178ndash4186 2010
[19] S D Covey R DWideman CMcDonald et al ldquoThe pancreat-ic 120573 cell is a key site formediating the effects of leptin on glucosehomeostasisrdquo Cell Metabolism vol 4 no 4 pp 291ndash302 2006
[20] K J Dudley DM Sloboda K L Connor J Beltrand andMHVickers ldquoOffspring ofmothers fed a high fat diet display hepaticcell cycle inhibition and associated changes in gene expressionand DNA methylationrdquo PLoS ONE vol 6 no 7 Article IDe21662 2011
[21] K L Connor M H Vickers J Beltrand M J Meaney and DM Sloboda ldquoNature nurture or nutrition Impact of maternalnutrition on maternal care offspring development and repro-ductive functionrdquo Journal of Physiology vol 590 part 9 pp2167ndash2180 2012
[22] J Beltrand D M Sloboda K L Connor M Truong and MH Vickers ldquoThe effect of neonatal leptin antagonism in malerat offspring is dependent upon the interaction between priormaternal nutritional status and post-weaning dietrdquo Journal ofNutrition andMetabolism vol 2012 Article ID 296935 10 pages2012
[23] K L Connor M H Vickers C Cupido E Sirimanne and DM Sloboda ldquoMaternal high fat diet during critical windowsof development alters adrenal cortical and medullary enzymeexpression in adult male rat offspringrdquo Journal of the Develop-mental Origins of Health and Disease vol 1 no 4 pp 245ndash2542010
[24] P J Mark C Sisala K Connor et al ldquoA maternal high-fat dietin rat pregnancy reduces growth of the fetus and the placen-tal junctional zone but not placental labyrinth zone growthrdquoJournal of the Developmental Origins of Health and Disease vol2 no 1 pp 63ndash70 2011
[25] D R Matthews J P Hosker A S Rudenski B A Naylor DF Treacher and R C Turner ldquoHomeostasis model assessmentinsulin resistance and 120573-cell function from fasting plasmaglucose and insulin concentrations in manrdquo Diabetologia vol28 no 7 pp 412ndash419 1985
[26] M P Hermans J C Levy R J Morris and R C TurnerldquoComparison of tests of120573-cell function across a range of glucosetolerance from normal to diabetesrdquo Diabetes vol 48 no 9 pp1779ndash1786 1999
[27] A B Bernal M H Vickers M B Hampton R A Poynton andD M Sloboda ldquoMaternal undernutrition significantly impactsovarian follicle number and increases ovarian oxidative stress inadult rat offspringrdquo PLoS ONE vol 5 no 12 Article ID e155582010
[28] J Vandesompele K de Preter F Pattyn et al ldquoAccurate nor-malization of real-time quantitative RT-PCR data by geometricaveraging of multiple internal control genesrdquo Genome biologyvol 3 no 7 Article ID RESEARCH0034 2002
[29] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the 2-ΔΔCT methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
[30] J Armitage L Poston and P Taylor ldquoDevelopmental originsof obesity and the metabolic syndrome the role of maternalobesityrdquo Frontiers of Hormone Research vol 36 pp 73ndash84 2007
[31] S Zhang L Rattanatray J L Morrison L M Nicholas S Lieand I C McMillen ldquoMaternal obesity and the early originsof childhood obesity weighing up the benefits and costs ofmaternal weight loss in the periconceptional period for theoffspringrdquo Experimental Diabetes Research vol 2011 Article ID585749 10 pages 2011
[32] N M Thompson A M Norman S S Donkin et al ldquoPrenataland postnatal pathways to obesity different underlying mech-anisms different metabolic outcomesrdquo Endocrinology vol 148no 5 pp 2345ndash2354 2007
[33] T Fuentes I Ara A Guadalupe-Grau et al ldquoLeptin receptor170 kDa (OB-R170) protein expression is reduced in obesehuman skeletal muscle a potential mechanism of leptin resis-tancerdquo Experimental Physiology vol 95 no 1 pp 160ndash171 2010
[34] Z Wang Y T Zhou T Kakuma et al ldquoLeptin resistance ofadipocytes in obesity role of suppressors of cytokine signalingrdquo
Journal of Nutrition and Metabolism 11
Biochemical and Biophysical Research Communications vol 277no 1 pp 20ndash26 2000
[35] C Bjoslashrbaeligk K El-Haschimi J D Frantz and J S Flier ldquoThe roleof SOCS-3 in leptin signaling and leptin resistancerdquoThe Journalof Biological Chemistry vol 274 no 42 pp 30059ndash30065 1999
[36] K Laubner T J Kieffer N T Lam X Niu F Jakob and JSeufert ldquoInhibition of preproinsulin gene expression by leptininduction of suppressor of cytokine signaling 3 in pancreatic 120573-cellsrdquo Diabetes vol 54 no 12 pp 3410ndash3417 2005
[37] C Peiser G P McGregor and R E Lang ldquoLeptin receptorexpression and suppressor of cytokine signaling transcriptlevels in high-fat-fed ratsrdquo Life Sciences vol 67 no 24 pp 2971ndash2981 2000
[38] G R Steinberg A C Smith S Wormald P Malenfant CCollier and D J Dyck ldquoEndurance training partially reversesdietary-induced leptin resistance in rodent skeletal musclerdquoTheAmerican Journal of Physiology vol 286 no 1 pp E57ndashE632004
[39] G E White A Cotterill M R Addley E J Soilleux and DR Greaves ldquoSuppressor of cytokine signalling protein SOCS3expression is increased at sites of acute and chronic inflamma-tionrdquo Journal of Molecular Histology vol 42 no 2 pp 137ndash1512011
[40] H YasukawaMOhishiHMori et al ldquoIL-6 induces an anti-in-flammatory response in the absence of SOCS3 inmacrophagesrdquoNature Immunology vol 4 no 6 pp 551ndash556 2003
[41] J Seufert ldquoLeptin effects on pancreatic 120573-cell gene expressionand functionrdquo Diabetes vol 53 no 1 pp S152ndashS158 2004
[42] A S Metlakunta M Sahu and A Sahu ldquoHypothalamicphosphatidylinositol 3-kinase pathway of leptin signaling isimpaired during the development of diet-induced obesity inFVBNmicerdquo Endocrinology vol 149 no 3 pp 1121ndash1128 2008
[43] C A Aspinwall W J Qian M G Roper R N Kulkarni C RKahn and R T Kennedy ldquoRoles of insulin receptor substrate-1phosphatidylinositol 3-kinase and release of intracellular Ca2+stores in insulin-stimulated insulin secretion in 120573-cellsrdquo TheJournal of Biological Chemistry vol 275 no 29 pp 22331ndash223382000
[44] I B Leibiger B Leibiger T Moede and P O BerggrenldquoExocytosis of insulin promotes insulin gene transcription viathe insulin receptorPI-3 kinasep70 s6 kinase and CaM kinasepathwaysrdquoMolecular Cell vol 1 no 6 pp 933ndash938 1998
[45] R L Martin E Perez Y J He R Dawson Jr andW J MillardldquoLeptin resistance is associated with hypothalamic leptin recep-tor mRNA and protein downregulationrdquo Metabolism vol 49no 11 pp 1479ndash1484 2000
[46] J Wilsey and P J Scarpace ldquoCaloric restriction reverses thedeficits in leptin receptor protein and leptin signaling capacityassociated with diet-induced obesity role of leptin in the reg-ulation of hypothalamic long-form leptin receptor expressionrdquoJournal of Endocrinology vol 181 no 2 pp 297ndash306 2004
[47] Y Zhang and P J Scarpace ldquoThe role of leptin in leptin resist-ance and obesityrdquo Physiology and Behavior vol 88 no 3 pp249ndash256 2006
[48] D R Grattan S R Ladyman and R A Augustine ldquoHormonalinduction of leptin resistance during pregnancyrdquoPhysiology andBehavior vol 91 no 4 pp 366ndash374 2007
[49] L RuiM YuanD Frantz S Shoelson andM FWhite ldquoSOCS-1 and SOCS-3 block insulin signaling by ubiquitin-mediateddegradation of IRS1 and IRS2rdquoThe Journal of Biological Chem-istry vol 277 no 44 pp 42394ndash42398 2002
[50] K Ueki T Kondo and C R Kahn ldquoSuppressor of cytokine sig-naling 1 (SOCS-1) and SOCS-3 cause insulin resistance throughinhibition of tyrosine phosphorylation of insulin receptor sub-strate proteins by discrete mechanismsrdquoMolecular and CellularBiology vol 24 no 12 pp 5434ndash5446 2004
[51] S B Jorgensen O rsquoNeill HM L Sylow et al ldquoDeletion of skel-etal muscle SOCS3 prevents insulin resistance in obesityrdquo Dia-betes vol 62 no 1 pp 56ndash64 2013
[52] W Becker ldquoLeptin signal transductionrdquo in Leptin and LeptinAntagonists A Gertler Ed pp 1ndash9 Landes Bioscience AustinTex USA 2009
[53] R N Kulkarni J N Winnay M Daniels et al ldquoAltered func-tion of insulin receptor substrate 1-deficient mouse islets andcultured 120573-cell linesrdquo Journal of Clinical Investigation vol 104no 12 pp R69ndashR75 1999
[54] W Y Kwong A E Wild P Roberts A C Willis and T PFleming ldquoMaternal undernutrition during the preimplantationperiod of rat development causes blastocyst abnormalities andprogramming of postnatal hypertensionrdquoDevelopment vol 127no 19 pp 4195ndash4202 2000
[55] E S Jungheim E L Schoeller K L Marquard E D LoudenJ E Schaffer and K H Moley ldquoDiet-induced obesity modelabnormal oocytes and persistent growth abnormalities in theoffspringrdquo Endocrinology vol 151 no 8 pp 4039ndash4046 2010
[56] A J Watkins A Wilkins C Cunningham et al ldquoLow proteindiet fed exclusively during mouse oocyte maturation leadsto behavioural and cardiovascular abnormalities in offspringrdquoJournal of Physiology vol 586 no 8 pp 2231ndash2244 2008
[57] E Zambrano P M Martınez-Samayoa G L Rodrıguez-Gonzalez and P W Nathanielsz ldquoDietary intervention prior topregnancy reverses metabolic programming in male offspringof obese ratsrdquo Journal of Physiology vol 588 no 10 pp 1791ndash1799 2010
[58] M S Patel andM Srinivasan ldquoMetabolic programming causesand consequencesrdquoThe Journal of Biological Chemistry vol 277no 3 pp 1629ndash1632 2002
[59] R A Simmons ldquoRole of metabolic programming in the patho-genesis of 120573-cell failure in postnatal liferdquo Reviews in Endocrineand Metabolic Disorders vol 8 no 2 pp 95ndash104 2007
[60] I B Leibiger B Leibiger and P O Berggren ldquoInsulin signalingin the pancreatic 120573-cellrdquoAnnual Review of Nutrition vol 28 pp233ndash251 2008

Journal of Nutrition and Metabolism 9
contribute to the lasting morbidity observed in offspring[55] Similarly even a very short period of altered nutrition(restricted to one ovulatory cycle) prior to natural mating canelicit lasting effects on the offspring [56] and this representsa further point of difference between the MHF and PLHFgroups
The present study was not designed to incorporate theeffect of a reversal paradigm whereby mothers fed thepre-conceptional HF diet were placed on a standard dietthroughout pregnancy and lactation However while there isevidence that a period of dietary restriction in obese mothersmay ablate the programming of obesity in offspring [57]it may be associated with an activation of the stress axisin the offspring [31] Further the data are inconsistent ithas also been shown that dams with diet-induced obesityfed a standard diet throughout pregnancy and lactation stilltransmit the propensity for obesity and metabolic disease totheir offspring [58]
We recognize that gene expression analysis on wholepancreatic tissue is a limitation of the present study Dueto the size of the cohort and logistical considerations onlywhole pancreatic tissue was a possibility whereas ideallyonly pure islet populations would have been examined [59]However for the key genes presented such as ObRb SOCS3and IRS expression has been shown to be primarily confinedto the endocrine islets and not been detected in the exocrinepancreas [39 41 60] therefore we are confident that theobserved changes in the genes studied represent changes inthe islets Further although we have shown that differencesin pancreatic gene expression in offspring are dependentupon the timing of maternal high fat diet exposure this doesnot itself imply causation in the development of the obesephenotypemdashother relevant pathways including liver skeletalmuscle and fat metabolism will also be integrated into theprogrammed phenotype development
This study has demonstrated enduring deleterious effectsof a maternal HF diet during early windows of develop-ment Maternal HF feeding both before conception andorthrough pregnancy and lactation induced obesity hyper-insulinemia and hyperleptinemia in adult offspring eventhough offspring were fed a standard diet postnatally Timingof exposure to the maternal HF diet differentially alteredgene expression in signaling pathways downstream of thepancreatic leptin receptor that mediated leptin resistanceThis also raises the possibility that despite common offspringobesity treatment modalities may only be efficacious in asubpopulation of treated offspring as the current data suggestthat the metabolic pathways disrupted will be dependentupon the timing of the early life dietary insult
Abbreviations
CD68 Cluster of differentiation 68CONT ControlDEXA Dual energy X-ray absorptiometryELISA Enzyme-linked immunosorbent assayGLUT2 Glucose transporter 2GP130 Glycoprotein 130HF High fat
HOMA Homeostatic model assessmentHPRT HypoxanthinephophoribosyltransferaseIL InterleukinIns InsulinIR Insulin resistanceIRS Insulin receptor substrateKir62 Potassium channel subunitLIF Leukemia inhibitory factorMHF Maternal high fatOBR Leptin receptorPDE3B Phosphodiesterase 3BPDX-1 Pancreatic and duodenal homeobox-1PI3K Phosphatidylinositide 3-kinasePLHF Pregnancy and lactation high fatSOCS3 Suppressor of cytokine signaling 3STAT3 Signal transducer and activator of
transcription 3TNF-120572 Tumor necrosis factor-alpha
Conflict of Interests
The authors have no conflict of interests with any financialorganization regarding the material discussed in this paper
Author Contributions
Mark H Vickers developed the animal model designed thestudy completed the data analysis and wrote the first draftof the paper Graham J Howie maintained the experimentalcohort performed the gene expression analysis and con-tributed to the original draft paper Clare M Reynolds per-formed the inflammatory related assays and gene expressionwork Deborah M Sloboda provided input into data analysisand interpretation
Acknowledgments
The authors acknowledge the technical support of TaniaKamal with endocrine assays The authors acknowledgefunding support from the Health Research Council of NewZealand and Gravida National Centre for Growth andDevelopment
References
[1] M M Hillemeier C S Weisman C Chuang D S Downs JMcCall-Hosenfeld and F Camacho ldquoTransition to overweightor obesity among women of reproductive agerdquo Journal ofWomenrsquos Health vol 20 no 5 pp 703ndash710 2011
[2] J E Norman and RM Reynolds ldquoThe consequences of obesityand excess weight gain in pregnancyrdquo Proceedings of the Nutri-tion Society vol 70 no 4 pp 450ndash456 2011
[3] H A Abenhaim R A Kinch L Morin A Benjamin and RUsher ldquoEffect of prepregnancy body mass index categories onobstetrical and neonatal outcomesrdquo Archives of Gynecology andObstetrics vol 275 no 1 pp 39ndash43 2007
[4] S DHMalnick andH Knobler ldquoThemedical complications ofobesityrdquo Oxford Journals Medicine vol 99 no 9 pp 565ndash5792006
10 Journal of Nutrition and Metabolism
[5] P M Catalano ldquoManagement of obesity in pregnancyrdquo Obstet-rics and Gynecology vol 109 no 2 pp 419ndash433 2007
[6] L Poston ldquoGestational weight gain influences on the long-termhealth of the childrdquo Current Opinion in Clinical Nutrition andMetabolic Care vol 15 pp 252ndash257 2012
[7] H Hochner Y Friedlander R Calderon-Margalit et al ldquoAsso-ciations of maternal prepregnancy body mass index and gesta-tional weight gain with adult offspring cardiometabolic riskfactors the Jerusalem perinatal family follow-up studyrdquo Circu-lation vol 125 no 11 pp 1381ndash1389 2012
[8] S J Spencer ldquoEarly life programming of obesity the impactof the perinatal environment on the development of obesityand metabolic dysfunction in the offspringrdquo Current DiabetesReviews vol 8 no 1 pp 55ndash68 2012
[9] H Chen D Simar K Lambert J Mercier and M J MorrisldquoMaternal and postnatal overnutrition differentially impactappetite regulators and fuel metabolismrdquo Endocrinology vol149 no 11 pp 5348ndash5356 2008
[10] P M Catalano ldquoObesity and pregnancymdashthe propagation of aviscous cyclerdquo Journal of Clinical Endocrinology and Metabo-lism vol 88 no 8 pp 3505ndash3506 2003
[11] M W Gillman S L Rifas-Shiman K Kleinman E Oken JW Rich-Edwards and EM Taveras ldquoDevelopmental origins ofchildhood overweight potential public health impactrdquo Obesityvol 16 no 7 pp 1651ndash1656 2008
[12] E Oken EM Taveras K P Kleinman JW Rich-Edwards andM W Gillman ldquoGestational weight gain and child adiposity atage 3 yearsrdquoTheAmerican Journal of Obstetrics and Gynecologyvol 196 no 4 pp 322e1ndash322e8 2007
[13] H M Ehrenberg B M Mercer and P M Catalano ldquoThe influ-ence of obesity and diabetes on the prevalence of macrosomiardquoThe American Journal of Obstetrics and Gynecology vol 191 no3 pp 964ndash968 2004
[14] G J Howie D M Sloboda T Kamal and M H VickersldquoMaternal nutritional history predicts obesity in adult offspringindependent of postnatal dietrdquo Journal of Physiology vol 587no 4 pp 905ndash915 2009
[15] D M Sloboda G J Howie A Pleasants P D Gluckman andM H Vickers ldquoPre- and postnatal nutritional histories influ-ence reproductive maturation and ovarian function in the ratrdquoPLoS ONE vol 4 no 8 Article ID e6744 2009
[16] M C Battista M F Hivert K Duval and J P BaillargeonldquoIntergenerational cycle of obesity and diabetes how can wereduce the burdens of these conditions on the health of futuregenerationsrdquo Experimental Diabetes Research vol 2011 ArticleID 596060 19 pages 2011
[17] T J Kieffer and J F Habener ldquoThe adipoinsular axis effects ofleptin on pancreatic 120573-cellsrdquo The American Journal of Physiol-ogy vol 278 no 1 pp E1ndashE14 2000
[18] S L Gray C Donald A Jetha S D Covey and T J KiefferldquoHyperinsulinemia precedes insulin resistance in mice lackingpancreatic 120573-cell leptin signalingrdquo Endocrinology vol 151 no 9pp 4178ndash4186 2010
[19] S D Covey R DWideman CMcDonald et al ldquoThe pancreat-ic 120573 cell is a key site formediating the effects of leptin on glucosehomeostasisrdquo Cell Metabolism vol 4 no 4 pp 291ndash302 2006
[20] K J Dudley DM Sloboda K L Connor J Beltrand andMHVickers ldquoOffspring ofmothers fed a high fat diet display hepaticcell cycle inhibition and associated changes in gene expressionand DNA methylationrdquo PLoS ONE vol 6 no 7 Article IDe21662 2011
[21] K L Connor M H Vickers J Beltrand M J Meaney and DM Sloboda ldquoNature nurture or nutrition Impact of maternalnutrition on maternal care offspring development and repro-ductive functionrdquo Journal of Physiology vol 590 part 9 pp2167ndash2180 2012
[22] J Beltrand D M Sloboda K L Connor M Truong and MH Vickers ldquoThe effect of neonatal leptin antagonism in malerat offspring is dependent upon the interaction between priormaternal nutritional status and post-weaning dietrdquo Journal ofNutrition andMetabolism vol 2012 Article ID 296935 10 pages2012
[23] K L Connor M H Vickers C Cupido E Sirimanne and DM Sloboda ldquoMaternal high fat diet during critical windowsof development alters adrenal cortical and medullary enzymeexpression in adult male rat offspringrdquo Journal of the Develop-mental Origins of Health and Disease vol 1 no 4 pp 245ndash2542010
[24] P J Mark C Sisala K Connor et al ldquoA maternal high-fat dietin rat pregnancy reduces growth of the fetus and the placen-tal junctional zone but not placental labyrinth zone growthrdquoJournal of the Developmental Origins of Health and Disease vol2 no 1 pp 63ndash70 2011
[25] D R Matthews J P Hosker A S Rudenski B A Naylor DF Treacher and R C Turner ldquoHomeostasis model assessmentinsulin resistance and 120573-cell function from fasting plasmaglucose and insulin concentrations in manrdquo Diabetologia vol28 no 7 pp 412ndash419 1985
[26] M P Hermans J C Levy R J Morris and R C TurnerldquoComparison of tests of120573-cell function across a range of glucosetolerance from normal to diabetesrdquo Diabetes vol 48 no 9 pp1779ndash1786 1999
[27] A B Bernal M H Vickers M B Hampton R A Poynton andD M Sloboda ldquoMaternal undernutrition significantly impactsovarian follicle number and increases ovarian oxidative stress inadult rat offspringrdquo PLoS ONE vol 5 no 12 Article ID e155582010
[28] J Vandesompele K de Preter F Pattyn et al ldquoAccurate nor-malization of real-time quantitative RT-PCR data by geometricaveraging of multiple internal control genesrdquo Genome biologyvol 3 no 7 Article ID RESEARCH0034 2002
[29] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the 2-ΔΔCT methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
[30] J Armitage L Poston and P Taylor ldquoDevelopmental originsof obesity and the metabolic syndrome the role of maternalobesityrdquo Frontiers of Hormone Research vol 36 pp 73ndash84 2007
[31] S Zhang L Rattanatray J L Morrison L M Nicholas S Lieand I C McMillen ldquoMaternal obesity and the early originsof childhood obesity weighing up the benefits and costs ofmaternal weight loss in the periconceptional period for theoffspringrdquo Experimental Diabetes Research vol 2011 Article ID585749 10 pages 2011
[32] N M Thompson A M Norman S S Donkin et al ldquoPrenataland postnatal pathways to obesity different underlying mech-anisms different metabolic outcomesrdquo Endocrinology vol 148no 5 pp 2345ndash2354 2007
[33] T Fuentes I Ara A Guadalupe-Grau et al ldquoLeptin receptor170 kDa (OB-R170) protein expression is reduced in obesehuman skeletal muscle a potential mechanism of leptin resis-tancerdquo Experimental Physiology vol 95 no 1 pp 160ndash171 2010
[34] Z Wang Y T Zhou T Kakuma et al ldquoLeptin resistance ofadipocytes in obesity role of suppressors of cytokine signalingrdquo
Journal of Nutrition and Metabolism 11
Biochemical and Biophysical Research Communications vol 277no 1 pp 20ndash26 2000
[35] C Bjoslashrbaeligk K El-Haschimi J D Frantz and J S Flier ldquoThe roleof SOCS-3 in leptin signaling and leptin resistancerdquoThe Journalof Biological Chemistry vol 274 no 42 pp 30059ndash30065 1999
[36] K Laubner T J Kieffer N T Lam X Niu F Jakob and JSeufert ldquoInhibition of preproinsulin gene expression by leptininduction of suppressor of cytokine signaling 3 in pancreatic 120573-cellsrdquo Diabetes vol 54 no 12 pp 3410ndash3417 2005
[37] C Peiser G P McGregor and R E Lang ldquoLeptin receptorexpression and suppressor of cytokine signaling transcriptlevels in high-fat-fed ratsrdquo Life Sciences vol 67 no 24 pp 2971ndash2981 2000
[38] G R Steinberg A C Smith S Wormald P Malenfant CCollier and D J Dyck ldquoEndurance training partially reversesdietary-induced leptin resistance in rodent skeletal musclerdquoTheAmerican Journal of Physiology vol 286 no 1 pp E57ndashE632004
[39] G E White A Cotterill M R Addley E J Soilleux and DR Greaves ldquoSuppressor of cytokine signalling protein SOCS3expression is increased at sites of acute and chronic inflamma-tionrdquo Journal of Molecular Histology vol 42 no 2 pp 137ndash1512011
[40] H YasukawaMOhishiHMori et al ldquoIL-6 induces an anti-in-flammatory response in the absence of SOCS3 inmacrophagesrdquoNature Immunology vol 4 no 6 pp 551ndash556 2003
[41] J Seufert ldquoLeptin effects on pancreatic 120573-cell gene expressionand functionrdquo Diabetes vol 53 no 1 pp S152ndashS158 2004
[42] A S Metlakunta M Sahu and A Sahu ldquoHypothalamicphosphatidylinositol 3-kinase pathway of leptin signaling isimpaired during the development of diet-induced obesity inFVBNmicerdquo Endocrinology vol 149 no 3 pp 1121ndash1128 2008
[43] C A Aspinwall W J Qian M G Roper R N Kulkarni C RKahn and R T Kennedy ldquoRoles of insulin receptor substrate-1phosphatidylinositol 3-kinase and release of intracellular Ca2+stores in insulin-stimulated insulin secretion in 120573-cellsrdquo TheJournal of Biological Chemistry vol 275 no 29 pp 22331ndash223382000
[44] I B Leibiger B Leibiger T Moede and P O BerggrenldquoExocytosis of insulin promotes insulin gene transcription viathe insulin receptorPI-3 kinasep70 s6 kinase and CaM kinasepathwaysrdquoMolecular Cell vol 1 no 6 pp 933ndash938 1998
[45] R L Martin E Perez Y J He R Dawson Jr andW J MillardldquoLeptin resistance is associated with hypothalamic leptin recep-tor mRNA and protein downregulationrdquo Metabolism vol 49no 11 pp 1479ndash1484 2000
[46] J Wilsey and P J Scarpace ldquoCaloric restriction reverses thedeficits in leptin receptor protein and leptin signaling capacityassociated with diet-induced obesity role of leptin in the reg-ulation of hypothalamic long-form leptin receptor expressionrdquoJournal of Endocrinology vol 181 no 2 pp 297ndash306 2004
[47] Y Zhang and P J Scarpace ldquoThe role of leptin in leptin resist-ance and obesityrdquo Physiology and Behavior vol 88 no 3 pp249ndash256 2006
[48] D R Grattan S R Ladyman and R A Augustine ldquoHormonalinduction of leptin resistance during pregnancyrdquoPhysiology andBehavior vol 91 no 4 pp 366ndash374 2007
[49] L RuiM YuanD Frantz S Shoelson andM FWhite ldquoSOCS-1 and SOCS-3 block insulin signaling by ubiquitin-mediateddegradation of IRS1 and IRS2rdquoThe Journal of Biological Chem-istry vol 277 no 44 pp 42394ndash42398 2002
[50] K Ueki T Kondo and C R Kahn ldquoSuppressor of cytokine sig-naling 1 (SOCS-1) and SOCS-3 cause insulin resistance throughinhibition of tyrosine phosphorylation of insulin receptor sub-strate proteins by discrete mechanismsrdquoMolecular and CellularBiology vol 24 no 12 pp 5434ndash5446 2004
[51] S B Jorgensen O rsquoNeill HM L Sylow et al ldquoDeletion of skel-etal muscle SOCS3 prevents insulin resistance in obesityrdquo Dia-betes vol 62 no 1 pp 56ndash64 2013
[52] W Becker ldquoLeptin signal transductionrdquo in Leptin and LeptinAntagonists A Gertler Ed pp 1ndash9 Landes Bioscience AustinTex USA 2009
[53] R N Kulkarni J N Winnay M Daniels et al ldquoAltered func-tion of insulin receptor substrate 1-deficient mouse islets andcultured 120573-cell linesrdquo Journal of Clinical Investigation vol 104no 12 pp R69ndashR75 1999
[54] W Y Kwong A E Wild P Roberts A C Willis and T PFleming ldquoMaternal undernutrition during the preimplantationperiod of rat development causes blastocyst abnormalities andprogramming of postnatal hypertensionrdquoDevelopment vol 127no 19 pp 4195ndash4202 2000
[55] E S Jungheim E L Schoeller K L Marquard E D LoudenJ E Schaffer and K H Moley ldquoDiet-induced obesity modelabnormal oocytes and persistent growth abnormalities in theoffspringrdquo Endocrinology vol 151 no 8 pp 4039ndash4046 2010
[56] A J Watkins A Wilkins C Cunningham et al ldquoLow proteindiet fed exclusively during mouse oocyte maturation leadsto behavioural and cardiovascular abnormalities in offspringrdquoJournal of Physiology vol 586 no 8 pp 2231ndash2244 2008
[57] E Zambrano P M Martınez-Samayoa G L Rodrıguez-Gonzalez and P W Nathanielsz ldquoDietary intervention prior topregnancy reverses metabolic programming in male offspringof obese ratsrdquo Journal of Physiology vol 588 no 10 pp 1791ndash1799 2010
[58] M S Patel andM Srinivasan ldquoMetabolic programming causesand consequencesrdquoThe Journal of Biological Chemistry vol 277no 3 pp 1629ndash1632 2002
[59] R A Simmons ldquoRole of metabolic programming in the patho-genesis of 120573-cell failure in postnatal liferdquo Reviews in Endocrineand Metabolic Disorders vol 8 no 2 pp 95ndash104 2007
[60] I B Leibiger B Leibiger and P O Berggren ldquoInsulin signalingin the pancreatic 120573-cellrdquoAnnual Review of Nutrition vol 28 pp233ndash251 2008

10 Journal of Nutrition and Metabolism
[5] P M Catalano ldquoManagement of obesity in pregnancyrdquo Obstet-rics and Gynecology vol 109 no 2 pp 419ndash433 2007
[6] L Poston ldquoGestational weight gain influences on the long-termhealth of the childrdquo Current Opinion in Clinical Nutrition andMetabolic Care vol 15 pp 252ndash257 2012
[7] H Hochner Y Friedlander R Calderon-Margalit et al ldquoAsso-ciations of maternal prepregnancy body mass index and gesta-tional weight gain with adult offspring cardiometabolic riskfactors the Jerusalem perinatal family follow-up studyrdquo Circu-lation vol 125 no 11 pp 1381ndash1389 2012
[8] S J Spencer ldquoEarly life programming of obesity the impactof the perinatal environment on the development of obesityand metabolic dysfunction in the offspringrdquo Current DiabetesReviews vol 8 no 1 pp 55ndash68 2012
[9] H Chen D Simar K Lambert J Mercier and M J MorrisldquoMaternal and postnatal overnutrition differentially impactappetite regulators and fuel metabolismrdquo Endocrinology vol149 no 11 pp 5348ndash5356 2008
[10] P M Catalano ldquoObesity and pregnancymdashthe propagation of aviscous cyclerdquo Journal of Clinical Endocrinology and Metabo-lism vol 88 no 8 pp 3505ndash3506 2003
[11] M W Gillman S L Rifas-Shiman K Kleinman E Oken JW Rich-Edwards and EM Taveras ldquoDevelopmental origins ofchildhood overweight potential public health impactrdquo Obesityvol 16 no 7 pp 1651ndash1656 2008
[12] E Oken EM Taveras K P Kleinman JW Rich-Edwards andM W Gillman ldquoGestational weight gain and child adiposity atage 3 yearsrdquoTheAmerican Journal of Obstetrics and Gynecologyvol 196 no 4 pp 322e1ndash322e8 2007
[13] H M Ehrenberg B M Mercer and P M Catalano ldquoThe influ-ence of obesity and diabetes on the prevalence of macrosomiardquoThe American Journal of Obstetrics and Gynecology vol 191 no3 pp 964ndash968 2004
[14] G J Howie D M Sloboda T Kamal and M H VickersldquoMaternal nutritional history predicts obesity in adult offspringindependent of postnatal dietrdquo Journal of Physiology vol 587no 4 pp 905ndash915 2009
[15] D M Sloboda G J Howie A Pleasants P D Gluckman andM H Vickers ldquoPre- and postnatal nutritional histories influ-ence reproductive maturation and ovarian function in the ratrdquoPLoS ONE vol 4 no 8 Article ID e6744 2009
[16] M C Battista M F Hivert K Duval and J P BaillargeonldquoIntergenerational cycle of obesity and diabetes how can wereduce the burdens of these conditions on the health of futuregenerationsrdquo Experimental Diabetes Research vol 2011 ArticleID 596060 19 pages 2011
[17] T J Kieffer and J F Habener ldquoThe adipoinsular axis effects ofleptin on pancreatic 120573-cellsrdquo The American Journal of Physiol-ogy vol 278 no 1 pp E1ndashE14 2000
[18] S L Gray C Donald A Jetha S D Covey and T J KiefferldquoHyperinsulinemia precedes insulin resistance in mice lackingpancreatic 120573-cell leptin signalingrdquo Endocrinology vol 151 no 9pp 4178ndash4186 2010
[19] S D Covey R DWideman CMcDonald et al ldquoThe pancreat-ic 120573 cell is a key site formediating the effects of leptin on glucosehomeostasisrdquo Cell Metabolism vol 4 no 4 pp 291ndash302 2006
[20] K J Dudley DM Sloboda K L Connor J Beltrand andMHVickers ldquoOffspring ofmothers fed a high fat diet display hepaticcell cycle inhibition and associated changes in gene expressionand DNA methylationrdquo PLoS ONE vol 6 no 7 Article IDe21662 2011
[21] K L Connor M H Vickers J Beltrand M J Meaney and DM Sloboda ldquoNature nurture or nutrition Impact of maternalnutrition on maternal care offspring development and repro-ductive functionrdquo Journal of Physiology vol 590 part 9 pp2167ndash2180 2012
[22] J Beltrand D M Sloboda K L Connor M Truong and MH Vickers ldquoThe effect of neonatal leptin antagonism in malerat offspring is dependent upon the interaction between priormaternal nutritional status and post-weaning dietrdquo Journal ofNutrition andMetabolism vol 2012 Article ID 296935 10 pages2012
[23] K L Connor M H Vickers C Cupido E Sirimanne and DM Sloboda ldquoMaternal high fat diet during critical windowsof development alters adrenal cortical and medullary enzymeexpression in adult male rat offspringrdquo Journal of the Develop-mental Origins of Health and Disease vol 1 no 4 pp 245ndash2542010
[24] P J Mark C Sisala K Connor et al ldquoA maternal high-fat dietin rat pregnancy reduces growth of the fetus and the placen-tal junctional zone but not placental labyrinth zone growthrdquoJournal of the Developmental Origins of Health and Disease vol2 no 1 pp 63ndash70 2011
[25] D R Matthews J P Hosker A S Rudenski B A Naylor DF Treacher and R C Turner ldquoHomeostasis model assessmentinsulin resistance and 120573-cell function from fasting plasmaglucose and insulin concentrations in manrdquo Diabetologia vol28 no 7 pp 412ndash419 1985
[26] M P Hermans J C Levy R J Morris and R C TurnerldquoComparison of tests of120573-cell function across a range of glucosetolerance from normal to diabetesrdquo Diabetes vol 48 no 9 pp1779ndash1786 1999
[27] A B Bernal M H Vickers M B Hampton R A Poynton andD M Sloboda ldquoMaternal undernutrition significantly impactsovarian follicle number and increases ovarian oxidative stress inadult rat offspringrdquo PLoS ONE vol 5 no 12 Article ID e155582010
[28] J Vandesompele K de Preter F Pattyn et al ldquoAccurate nor-malization of real-time quantitative RT-PCR data by geometricaveraging of multiple internal control genesrdquo Genome biologyvol 3 no 7 Article ID RESEARCH0034 2002
[29] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the 2-ΔΔCT methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
[30] J Armitage L Poston and P Taylor ldquoDevelopmental originsof obesity and the metabolic syndrome the role of maternalobesityrdquo Frontiers of Hormone Research vol 36 pp 73ndash84 2007
[31] S Zhang L Rattanatray J L Morrison L M Nicholas S Lieand I C McMillen ldquoMaternal obesity and the early originsof childhood obesity weighing up the benefits and costs ofmaternal weight loss in the periconceptional period for theoffspringrdquo Experimental Diabetes Research vol 2011 Article ID585749 10 pages 2011
[32] N M Thompson A M Norman S S Donkin et al ldquoPrenataland postnatal pathways to obesity different underlying mech-anisms different metabolic outcomesrdquo Endocrinology vol 148no 5 pp 2345ndash2354 2007
[33] T Fuentes I Ara A Guadalupe-Grau et al ldquoLeptin receptor170 kDa (OB-R170) protein expression is reduced in obesehuman skeletal muscle a potential mechanism of leptin resis-tancerdquo Experimental Physiology vol 95 no 1 pp 160ndash171 2010
[34] Z Wang Y T Zhou T Kakuma et al ldquoLeptin resistance ofadipocytes in obesity role of suppressors of cytokine signalingrdquo
Journal of Nutrition and Metabolism 11
Biochemical and Biophysical Research Communications vol 277no 1 pp 20ndash26 2000
[35] C Bjoslashrbaeligk K El-Haschimi J D Frantz and J S Flier ldquoThe roleof SOCS-3 in leptin signaling and leptin resistancerdquoThe Journalof Biological Chemistry vol 274 no 42 pp 30059ndash30065 1999
[36] K Laubner T J Kieffer N T Lam X Niu F Jakob and JSeufert ldquoInhibition of preproinsulin gene expression by leptininduction of suppressor of cytokine signaling 3 in pancreatic 120573-cellsrdquo Diabetes vol 54 no 12 pp 3410ndash3417 2005
[37] C Peiser G P McGregor and R E Lang ldquoLeptin receptorexpression and suppressor of cytokine signaling transcriptlevels in high-fat-fed ratsrdquo Life Sciences vol 67 no 24 pp 2971ndash2981 2000
[38] G R Steinberg A C Smith S Wormald P Malenfant CCollier and D J Dyck ldquoEndurance training partially reversesdietary-induced leptin resistance in rodent skeletal musclerdquoTheAmerican Journal of Physiology vol 286 no 1 pp E57ndashE632004
[39] G E White A Cotterill M R Addley E J Soilleux and DR Greaves ldquoSuppressor of cytokine signalling protein SOCS3expression is increased at sites of acute and chronic inflamma-tionrdquo Journal of Molecular Histology vol 42 no 2 pp 137ndash1512011
[40] H YasukawaMOhishiHMori et al ldquoIL-6 induces an anti-in-flammatory response in the absence of SOCS3 inmacrophagesrdquoNature Immunology vol 4 no 6 pp 551ndash556 2003
[41] J Seufert ldquoLeptin effects on pancreatic 120573-cell gene expressionand functionrdquo Diabetes vol 53 no 1 pp S152ndashS158 2004
[42] A S Metlakunta M Sahu and A Sahu ldquoHypothalamicphosphatidylinositol 3-kinase pathway of leptin signaling isimpaired during the development of diet-induced obesity inFVBNmicerdquo Endocrinology vol 149 no 3 pp 1121ndash1128 2008
[43] C A Aspinwall W J Qian M G Roper R N Kulkarni C RKahn and R T Kennedy ldquoRoles of insulin receptor substrate-1phosphatidylinositol 3-kinase and release of intracellular Ca2+stores in insulin-stimulated insulin secretion in 120573-cellsrdquo TheJournal of Biological Chemistry vol 275 no 29 pp 22331ndash223382000
[44] I B Leibiger B Leibiger T Moede and P O BerggrenldquoExocytosis of insulin promotes insulin gene transcription viathe insulin receptorPI-3 kinasep70 s6 kinase and CaM kinasepathwaysrdquoMolecular Cell vol 1 no 6 pp 933ndash938 1998
[45] R L Martin E Perez Y J He R Dawson Jr andW J MillardldquoLeptin resistance is associated with hypothalamic leptin recep-tor mRNA and protein downregulationrdquo Metabolism vol 49no 11 pp 1479ndash1484 2000
[46] J Wilsey and P J Scarpace ldquoCaloric restriction reverses thedeficits in leptin receptor protein and leptin signaling capacityassociated with diet-induced obesity role of leptin in the reg-ulation of hypothalamic long-form leptin receptor expressionrdquoJournal of Endocrinology vol 181 no 2 pp 297ndash306 2004
[47] Y Zhang and P J Scarpace ldquoThe role of leptin in leptin resist-ance and obesityrdquo Physiology and Behavior vol 88 no 3 pp249ndash256 2006
[48] D R Grattan S R Ladyman and R A Augustine ldquoHormonalinduction of leptin resistance during pregnancyrdquoPhysiology andBehavior vol 91 no 4 pp 366ndash374 2007
[49] L RuiM YuanD Frantz S Shoelson andM FWhite ldquoSOCS-1 and SOCS-3 block insulin signaling by ubiquitin-mediateddegradation of IRS1 and IRS2rdquoThe Journal of Biological Chem-istry vol 277 no 44 pp 42394ndash42398 2002
[50] K Ueki T Kondo and C R Kahn ldquoSuppressor of cytokine sig-naling 1 (SOCS-1) and SOCS-3 cause insulin resistance throughinhibition of tyrosine phosphorylation of insulin receptor sub-strate proteins by discrete mechanismsrdquoMolecular and CellularBiology vol 24 no 12 pp 5434ndash5446 2004
[51] S B Jorgensen O rsquoNeill HM L Sylow et al ldquoDeletion of skel-etal muscle SOCS3 prevents insulin resistance in obesityrdquo Dia-betes vol 62 no 1 pp 56ndash64 2013
[52] W Becker ldquoLeptin signal transductionrdquo in Leptin and LeptinAntagonists A Gertler Ed pp 1ndash9 Landes Bioscience AustinTex USA 2009
[53] R N Kulkarni J N Winnay M Daniels et al ldquoAltered func-tion of insulin receptor substrate 1-deficient mouse islets andcultured 120573-cell linesrdquo Journal of Clinical Investigation vol 104no 12 pp R69ndashR75 1999
[54] W Y Kwong A E Wild P Roberts A C Willis and T PFleming ldquoMaternal undernutrition during the preimplantationperiod of rat development causes blastocyst abnormalities andprogramming of postnatal hypertensionrdquoDevelopment vol 127no 19 pp 4195ndash4202 2000
[55] E S Jungheim E L Schoeller K L Marquard E D LoudenJ E Schaffer and K H Moley ldquoDiet-induced obesity modelabnormal oocytes and persistent growth abnormalities in theoffspringrdquo Endocrinology vol 151 no 8 pp 4039ndash4046 2010
[56] A J Watkins A Wilkins C Cunningham et al ldquoLow proteindiet fed exclusively during mouse oocyte maturation leadsto behavioural and cardiovascular abnormalities in offspringrdquoJournal of Physiology vol 586 no 8 pp 2231ndash2244 2008
[57] E Zambrano P M Martınez-Samayoa G L Rodrıguez-Gonzalez and P W Nathanielsz ldquoDietary intervention prior topregnancy reverses metabolic programming in male offspringof obese ratsrdquo Journal of Physiology vol 588 no 10 pp 1791ndash1799 2010
[58] M S Patel andM Srinivasan ldquoMetabolic programming causesand consequencesrdquoThe Journal of Biological Chemistry vol 277no 3 pp 1629ndash1632 2002
[59] R A Simmons ldquoRole of metabolic programming in the patho-genesis of 120573-cell failure in postnatal liferdquo Reviews in Endocrineand Metabolic Disorders vol 8 no 2 pp 95ndash104 2007
[60] I B Leibiger B Leibiger and P O Berggren ldquoInsulin signalingin the pancreatic 120573-cellrdquoAnnual Review of Nutrition vol 28 pp233ndash251 2008

Journal of Nutrition and Metabolism 11
Biochemical and Biophysical Research Communications vol 277no 1 pp 20ndash26 2000
[35] C Bjoslashrbaeligk K El-Haschimi J D Frantz and J S Flier ldquoThe roleof SOCS-3 in leptin signaling and leptin resistancerdquoThe Journalof Biological Chemistry vol 274 no 42 pp 30059ndash30065 1999
[36] K Laubner T J Kieffer N T Lam X Niu F Jakob and JSeufert ldquoInhibition of preproinsulin gene expression by leptininduction of suppressor of cytokine signaling 3 in pancreatic 120573-cellsrdquo Diabetes vol 54 no 12 pp 3410ndash3417 2005
[37] C Peiser G P McGregor and R E Lang ldquoLeptin receptorexpression and suppressor of cytokine signaling transcriptlevels in high-fat-fed ratsrdquo Life Sciences vol 67 no 24 pp 2971ndash2981 2000
[38] G R Steinberg A C Smith S Wormald P Malenfant CCollier and D J Dyck ldquoEndurance training partially reversesdietary-induced leptin resistance in rodent skeletal musclerdquoTheAmerican Journal of Physiology vol 286 no 1 pp E57ndashE632004
[39] G E White A Cotterill M R Addley E J Soilleux and DR Greaves ldquoSuppressor of cytokine signalling protein SOCS3expression is increased at sites of acute and chronic inflamma-tionrdquo Journal of Molecular Histology vol 42 no 2 pp 137ndash1512011
[40] H YasukawaMOhishiHMori et al ldquoIL-6 induces an anti-in-flammatory response in the absence of SOCS3 inmacrophagesrdquoNature Immunology vol 4 no 6 pp 551ndash556 2003
[41] J Seufert ldquoLeptin effects on pancreatic 120573-cell gene expressionand functionrdquo Diabetes vol 53 no 1 pp S152ndashS158 2004
[42] A S Metlakunta M Sahu and A Sahu ldquoHypothalamicphosphatidylinositol 3-kinase pathway of leptin signaling isimpaired during the development of diet-induced obesity inFVBNmicerdquo Endocrinology vol 149 no 3 pp 1121ndash1128 2008
[43] C A Aspinwall W J Qian M G Roper R N Kulkarni C RKahn and R T Kennedy ldquoRoles of insulin receptor substrate-1phosphatidylinositol 3-kinase and release of intracellular Ca2+stores in insulin-stimulated insulin secretion in 120573-cellsrdquo TheJournal of Biological Chemistry vol 275 no 29 pp 22331ndash223382000
[44] I B Leibiger B Leibiger T Moede and P O BerggrenldquoExocytosis of insulin promotes insulin gene transcription viathe insulin receptorPI-3 kinasep70 s6 kinase and CaM kinasepathwaysrdquoMolecular Cell vol 1 no 6 pp 933ndash938 1998
[45] R L Martin E Perez Y J He R Dawson Jr andW J MillardldquoLeptin resistance is associated with hypothalamic leptin recep-tor mRNA and protein downregulationrdquo Metabolism vol 49no 11 pp 1479ndash1484 2000
[46] J Wilsey and P J Scarpace ldquoCaloric restriction reverses thedeficits in leptin receptor protein and leptin signaling capacityassociated with diet-induced obesity role of leptin in the reg-ulation of hypothalamic long-form leptin receptor expressionrdquoJournal of Endocrinology vol 181 no 2 pp 297ndash306 2004
[47] Y Zhang and P J Scarpace ldquoThe role of leptin in leptin resist-ance and obesityrdquo Physiology and Behavior vol 88 no 3 pp249ndash256 2006
[48] D R Grattan S R Ladyman and R A Augustine ldquoHormonalinduction of leptin resistance during pregnancyrdquoPhysiology andBehavior vol 91 no 4 pp 366ndash374 2007
[49] L RuiM YuanD Frantz S Shoelson andM FWhite ldquoSOCS-1 and SOCS-3 block insulin signaling by ubiquitin-mediateddegradation of IRS1 and IRS2rdquoThe Journal of Biological Chem-istry vol 277 no 44 pp 42394ndash42398 2002
[50] K Ueki T Kondo and C R Kahn ldquoSuppressor of cytokine sig-naling 1 (SOCS-1) and SOCS-3 cause insulin resistance throughinhibition of tyrosine phosphorylation of insulin receptor sub-strate proteins by discrete mechanismsrdquoMolecular and CellularBiology vol 24 no 12 pp 5434ndash5446 2004
[51] S B Jorgensen O rsquoNeill HM L Sylow et al ldquoDeletion of skel-etal muscle SOCS3 prevents insulin resistance in obesityrdquo Dia-betes vol 62 no 1 pp 56ndash64 2013
[52] W Becker ldquoLeptin signal transductionrdquo in Leptin and LeptinAntagonists A Gertler Ed pp 1ndash9 Landes Bioscience AustinTex USA 2009
[53] R N Kulkarni J N Winnay M Daniels et al ldquoAltered func-tion of insulin receptor substrate 1-deficient mouse islets andcultured 120573-cell linesrdquo Journal of Clinical Investigation vol 104no 12 pp R69ndashR75 1999
[54] W Y Kwong A E Wild P Roberts A C Willis and T PFleming ldquoMaternal undernutrition during the preimplantationperiod of rat development causes blastocyst abnormalities andprogramming of postnatal hypertensionrdquoDevelopment vol 127no 19 pp 4195ndash4202 2000
[55] E S Jungheim E L Schoeller K L Marquard E D LoudenJ E Schaffer and K H Moley ldquoDiet-induced obesity modelabnormal oocytes and persistent growth abnormalities in theoffspringrdquo Endocrinology vol 151 no 8 pp 4039ndash4046 2010
[56] A J Watkins A Wilkins C Cunningham et al ldquoLow proteindiet fed exclusively during mouse oocyte maturation leadsto behavioural and cardiovascular abnormalities in offspringrdquoJournal of Physiology vol 586 no 8 pp 2231ndash2244 2008
[57] E Zambrano P M Martınez-Samayoa G L Rodrıguez-Gonzalez and P W Nathanielsz ldquoDietary intervention prior topregnancy reverses metabolic programming in male offspringof obese ratsrdquo Journal of Physiology vol 588 no 10 pp 1791ndash1799 2010
[58] M S Patel andM Srinivasan ldquoMetabolic programming causesand consequencesrdquoThe Journal of Biological Chemistry vol 277no 3 pp 1629ndash1632 2002
[59] R A Simmons ldquoRole of metabolic programming in the patho-genesis of 120573-cell failure in postnatal liferdquo Reviews in Endocrineand Metabolic Disorders vol 8 no 2 pp 95ndash104 2007
[60] I B Leibiger B Leibiger and P O Berggren ldquoInsulin signalingin the pancreatic 120573-cellrdquoAnnual Review of Nutrition vol 28 pp233ndash251 2008




















