
Temperate grassland Herbicide Conservation Species composition Forage quality

1 3
Mar BiolDOI 10.1007/s00227-013-2344-z
OrIgInal PaPer
Thermal tolerance of early development in tropical and temperate sea urchins: inferences for the tropicalization of eastern Australia
Natasha A. Hardy · Miles Lamare · Sven Uthicke · Kennedy Wolfe · Steve Doo · Symon Dworjanyn · Maria Byrne
received: 28 april 2013 / accepted: 4 October 2013 © Springer-Verlag Berlin Heidelberg 2013
tolerance of development was determined in embryos and larvae reared at twelve temperatures. For A. placenta, the ambient water temperature near Townsville and experimen-tal control were 24 °C and treatments ranged from 14 to 37 °C. For C. rodgersii, ambient Sydney water temperature and experimental control were 17 °C, and the treatment range was 9–31 °C. A. placenta had a broader developmen-tal thermal envelope (14 °C range 17–31 °C) than C. rodg-ersii (9 °C range 13–22 °C). Both species developed suc-cessfully at temperatures well below ambient, suggesting that cooler water is not a barrier to poleward migration for either species. Both species presently live near the upper thermal limits for larval development, and future ocean warming could lead to contractions of their northern range limits. This study provides insights into the factors influ-encing the realized and potential distribution of planktonic life stages and changes to adult distribution in response to global change.
Introduction
The latitudinal distribution of many marine ectotherms is closely related to thermal tolerance limits of the adult and planktonic stages (andronikov 1975; Jones et al. 2009; Sunday et al. 2012). Climate-driven warming of the ocean surface is already occurring (0.6 °C in the last 10 years and 2–4 °C by 2100, IPCC 2007), and associated redistribu-tion marine biota has emerged as a significant biological response to this change (Walther et al. 2002; Poloczanska et al. 2007; Sunday et al. 2012). These redistributions are predicted to be range boundary changes with contractions equatorially and invasions poleward (Poloczanska et al. 2007; Jones et al. 2009; Sunday et al. 2012; Wernberg et al. 2011). Indeed, poleward range shifts have already occurred
Abstract The thermal envelope of development to the larval stage of two echinoids from eastern australia was characterized to determine whether they fill their potential latitudinal ranges as indicated by tolerance limits. The trop-ical sand dollar, Arachnoides placenta, a species that is not known to have shifted its range, was investigated in Towns-ville, northern australia (19°20′S, 146°77′e), during its autumn spawning season (May 2012). The subtropical/tem-perate sea urchin, Centrostephanus rodgersii, a species that has undergone poleward range expansion, was investigated in Sydney, southern australia (33°58′S, 151°14′e), during its winter spawning season (august 2012). The thermal
Communicated by U. Sommer.
Electronic supplementary material The online version of this article (doi:10.1007/s00227-013-2344-z) contains supplementary material, which is available to authorized users.
n. a. Hardy (*) · K. Wolfe · S. Doo · M. Byrne School of Medical Sciences, University of Sydney, Sydney, nSW 2006, australiae-mail: [email protected]
M. lamare Department of Marine Science, University of Otago, Dunedin, new Zealand
S. Uthicke australian Institute of Marine Science, Townsville, QlD, australia
K. Wolfe · S. Doo · M. Byrne School of Biological Sciences, University of Sydney, Sydney, nSW 2006, australia
S. Dworjanyn national Marine Science Centre, Southern Cross University, Coffs Harbour, nSW, australia

Mar Biol
1 3
for many marine species (Barry et al. 1995; Figueira and Booth 2010; ling et al. 2008). Thus, determining the “ther-mal envelope”, the cool and warm tolerance, of a species is crucial to understanding and predicting changes in abun-dance and distribution of ectotherms in a changing ocean (Pörtner et al. 2007).
Temperature is one of the most important environmen-tal factors controlling development in marine invertebrates, with a general response of faster progression through developmental stages and shorter planktonic larval duration (PlD) as temperature increases, up to a limit (Pechenik 1987; Chen and Chen 1992; Hoegh-guldberg and Pearse 1995). Thus, temperature plays a key role in the recruit-ment dynamics of many species (Pechenik 1987; Hart and Scheibling 1988; O’Connor et al. 2007). The hypothesis that thermal tolerance of planktonic propagules limits the geographical distribution of marine invertebrates is sup-ported by the correlation between spawning temperature and successful development of many species (andronikov 1975; Fujisawa 1995; reitzel et al. 2005). However, devel-opment in some species is tolerant to temperatures well above and below ambient spawning temperatures (rupp 1973; Byrne 2010). It is also suggested that the lower temperature range for normal development does not limit the distribution of many marine invertebrates (Sewell and Young 1999). In the context of these observations and in a warming ocean, it is important to better understand the thermal tolerance of development in marine invertebrates, as local habitats warm and propagules disperse to cooler climes.
The inherent flexibility of the life history of many marine invertebrates subject to environmental stress-ors, particularly temperature, is suggested to increase the probability of species persistence through geological time (Palmer 1994; Hadfield and Strathmann 1996). Most stud-ies on the impacts of temperature on echinoid development have focused on upper thermal thresholds of embryos and larvae (Mcedward 1985; Chen and Chen 1992; Hoegh-guldberg and Pearse 1995), but it is also important to understand lower thermal thresholds as we attempt to pre-dict how marine species may respond to changing ocean conditions (Pörtner et al. 2007). a transition to thermotol-erant species is predicted as a coastal ecosystem response to ocean warming, in parallel with local extinctions of less-tolerant species (Poloczanska et al. 2007).
The tropicalization of temperate australia is under way as sea surface temperatures (SSTs) have warmed by 1 °C over the last 60 years (Hobday and lough 2011). This warming is mainly due to climate-driven increased pole-ward flow of the east australian Current (eaC), a major boundary current, by ~350 km during this time (ridg-way 2007; Hobday and lough 2011). Increased poleward flow of tropical waters of the eaC is driving southerly
migration of tropical and subtropical fish, echinoderms and corals (ling et al. 2009; Figueira and Booth 2010; Baird et al. 2012; Johnson et al. 2012). These migrations have the potential to cause widespread ecological change, as in the case of Centrostephanus rodgersii in Tasmania (ling et al. 2009), and thus, it is important to understand species’ potential for migration with respect to thermal tolerance of their planktonic stages.
We investigated the thermal tolerance of embryos and larvae of two sea urchin species in eastern australia to determine whether they fill their potential latitudinal ranges as indicated by developmental tolerance limits, and to deter-mine the effects of projected near-future ocean warming. We focused on two ecologically important and abundant species, Arachnoides placenta from tropical Queensland and C. rodgersii from temperate nSW. A. placenta is an intertidal sand dollar with a broad distribution from Taiwan to western australia. In australia, A. placenta occurs from Moreton Bay, southern Queensland (27°12′S, 153°14′e) to Broome, western australia (17°57′S, 122°14′e) (Miskelly 2002; Haycock 2004). This species is not known to have shifted its range. In Queensland, A. placenta spawns in autumn from april to June (SST: 22–28 °C in Townsville) and larvae are estimated to spend 7–14 days in the plankton prior to settlement (Haycock 2004). Centrostephanus rodg-ersii is a temperate sea urchin that occurs from subtropi-cal northern new South Wales (nSW) (28°37′S, 153°38′e) to north-eastern Tasmania (41°52′S, 148°18′e) (Byrne and andrew 2013). This species has undergone marked poleward range expansion in eastern australia (ling et al. 2008), and its presence in northern new Zealand may also be recent (Pecorino et al. 2013). This species spawns in winter, July to august (SST: 15–20 °C in Sydney), and has a long planktonic phase of 3 months (Swanson et al. 2012; Byrne and andrew 2013).
Studies on the thermal biology of echinoid development have largely focused on upper thermal limits, and it appears that moderate warming (≤4 °C) is tolerated by the develop-mental stages of many species, but greater warming is del-eterious (Byrne 2010). Upper thermal limits for develop-ment provide insights into potential for local extinction and range contraction in a warming world, whilst lower limits provide insights into potential for poleward range extension through dispersal of planktonic larvae. The thermal enve-lope for development in A. placenta is not known, although an upper thermal tolerance limit of 28–31 °C is reported for populations from Taiwan (Chen and Chen 1992) where the annual temperature range is 20–30 °C (Physical Oceanog-raphy DaaC Ocean eSIP Tool; http://poet.jpl.nasa.gov/). The thermal envelope of development in C. rodgersii has been determined for southern (12–20 °C) and northern (17.3–24.5 °C) range edge populations in Tasmania and northern nSW, respectively (ling et al. 2008; Pecorino

Mar Biol
1 3
et al. 2013). a recent study indicated that the northern new Zealand population has a developmental thermal envelope of 17.2–23.5 °C (Pecorino et al. 2013). Here, we investi-gated the thermal biology of development for C. rodgersii from populations near the middle of its range in new South Wales, which based on genetic evidence, were the source for the invasive populations in Tasmania (Banks et al. 2007). Data on upper thermal limits for development in A. placenta and C. rodgersii were used to assess the potential for persistence in their current habitat and potential north-ern range expansion in the face of warming. The lower thermal limits were determined to assess the potential for southern range expansion to cooler latitudes.
The aims of this study were to characterize the thermal envelope of development to the larval stage in A. placenta and C. rodgersii. as A. placenta is a tropical species with a broad distribution, which in eastern australia is largely restricted to northern Queensland, we expected that pole-ward migration of this species would be limited by the cool tolerance limits of larval development. In contrast, the inva-sive populations of C. rodgersii in Tasmania are living at the low thermal limit for successful development (12 °C) (ling et al. 2008). For C. rodgersii, we expected that pole-ward migration was facilitated by the cool tolerance of larval development in mid-range populations that are the likely source of propagules for the invasive populations. The following hypotheses were addressed: (1) cool tem-perature is a barrier to southern migration in A. placenta; (2) the cool tolerance of development in nSW C. rodgersii facilitated invasion to Tasmania; (3) development in both species will be vulnerable to projected near-future ocean warming (~4 °C).
Materials and methods
Specimen collection
Arachnoides placenta (at least 20-mm diameter) were collected at low tide from Pallarenda Beach, Queensland (19°20′S, 146°77′e), in May 2012 and transferred to flow-through aquaria at ambient SST (~24 °C), at the austral-ian Institute of Marine Science (aIMS). Centrostephanus rodgersii (at least 60-mm diameter) were collected (1–2 m depth) by snorkelling at low tide from little Bay, Sydney nSW (33°58′S, 151°14′e), in august 2012 and maintained in aquaria at ambient SST (~17 °C) for several hours before use.
Sea surface temperatures
Continuous measurements of SSTs were available for both study sites. These data were used to determine the
control temperature treatment and to place experiments in an environmentally relevant context, for present and near-future ocean-warming projections (IPCC 2007; Hobday and lough 2011). Daily average SST data for the coastal habitat of A. placenta have been collected near Townsville (aIMS mooring, 19°06′S, 146°51′e, Mag-netic Island, 6.8 m depth) over 16 years (1996–2011). Multiple daily measurements of SST for the coastal habi-tat of C. rodgersii were available for 8 years (2002–2010) from the Integrated Marine Observing System moor-ing near Sydney (Waverider Buoy, 33°78′S, 151°42′e, Bluenet: http://www.metoc.gov.au/geonetwork/srv/en/search.external?any=%22Sydney+offshore+wave+data%22).
Sea surface temperatures for eastern australia were determined from the CSIrO atlas of regional Seas (CarS2009) (see Condie and Dunn 2006) and used to cre-ate contour plots of marine isotherms in eastern australia for present autumn and winter conditions (Fig. 1a, b), when the larvae of the two species are in the plankton. Marine isotherm data were overlain on coastal maps in arcgIS (eSrI) to create contour plots. These data were used to illustrate broad thermal trends for the east coast of aus-tralia and to assess the present distribution of the study spe-cies with respect to their developmental thermotolerances. Future changes in the planktonic habitats of A. placenta and C. rodgersii were assessed based on present latitudinal isotherms and +4 °C overall projections for 2100 (Hobday and lough 2011).
Spawning and fertilization
Mature A. placenta were induced to spawn by intra-coe-lomic injection of 1–2 ml of 0.5 M KCl. Females were inverted over a beaker of filtered seawater (1.0 μm) and left to spawn for 15 min. Sperm was collected as it was released. Sperm from 5 males was pooled in a plastic tube and stored at 4 °C until use (<30 min). Mature C. rodgersii were induced to spawn using 2–4 ml of 0.5 M KCl. For C. rodgersii, eggs and sperm were collected dry from the surface of the gonopores as they were released using glass pipettes. The eggs were transferred to beakers and sperm were placed dry in small dishes, pooled, and stored at 4 °C until use (<30 min). The gametes of both species were checked microscopically for quality (e.g. egg shape, sperm motility).
For each species, eggs from 3 to 5 females were pooled in 250-ml beakers and rinsed several times. The number of eggs in the suspension was determined from averag-ing counts of 100 μl aliquots (n = 5). The total number of eggs required for each experiment was placed in a 500-ml beaker of FSW. Sperm counts were determined from an average of 5 haemocytometer counts. a concentration

Mar Biol
1 3
of 104 sperm ml−1 was used for A. placenta to ensure high fertilization, low polyspermy, and acceptable rates of normal development to the larval stage (>75 %) in con-trols (see gonzalez-Bernat et al. 2013). For C. rodgersii, a concentration of 102 sperm ml−1 was used (see Byrne et al. 2010). The percentage of fertilization was checked in counts of 50 randomly selected eggs to ensure fertilization rates were acceptable (>90 %). eggs were rinsed 15 min post-fertilization (2–3 times) to remove excess sperm, and seawater was renewed. The embryos were then placed in experimental treatments.
experimental treatments and rearing
Twelve temperature levels were used to determine the lower and upper thermal thresholds for normal develop-ment in A. placenta and C. rodgersii based on recent aver-age SST. Temperature treatments included controls (ambi-ent SST), the extremes currently experienced, near-future projections for increasing SSTs up to 4 °C by 2100 (Hob-day and lough 2011), and the range of temperatures larvae might encounter at cooler latitudes (Table 1). For A. pla-centa, these temperature treatments were ~14, 17, 19, 22, 24, 26, 27, 29, 31, 32, 35, 37 °C with an ambient control of 24 °C. For C. rodgersii, temperature treatments were ~9, 11, 13, 15, 17, 20, 22, 24, 26, 27, 29, 31 °C with an ambient control of 17 °C. experimental treatments were achieved
using an aluminium block with a cold water input at one end and a warm water input at the other end creating a ther-mal gradient. although 12 temperature levels were used, due to high mortality in extreme treatments (see “results” section), the analyses involved fewer treatments due high mortality in extreme treatments (see “Statistics” section).
Filtered seawater (0.5 μm, Millipore) from the flow-through system at aIMS was used for experiments with A. placenta. For C. rodgersii experiments, seawater was col-lected from a nearby open ocean site, long Bay, Sydney nSW (33°57′S, 151°15′e), at high tide and filtered (1 μm, Millipore). The salinity, dissolved oxygen (required >90 %
Fig. 1 Marine isotherms (in °C) for present-day eastern aus-tralia a in autumn, b in winter (see Hobday and lough 2011, CarS2009)
Table 1 Temperature range used in experiments with Arachnoides placenta and Centrostephanus rodgersii in context with present and future conditions
Data from environmental monitoring (see “Methods” section)
Treatments A. placenta (May–June, Townsville, QlD) (°C)
C. rodgersii (July–august, Sydney, nSW) (°C)
Control temperature 24 17
Spawning season range 22–28 15–20
Spawning season warming projections (ca. +4 °C)
26–32 19–24
low-temperature treatments 14–19 9–13
High-temperature treatments 35, 37 26–31

Mar Biol
1 3
DO), temperature and pH were recorded for all experimen-tal water, using a WTW multimeter (Wissenschaftlich-Technische-Werkstätten, model Multiline 3400i). For A. placenta, temperature in the vials was checked with a ther-mometer twice daily (Table S.1). For C. rodgersii, temper-ature loggers (Onset Thermodata) were placed in vials in the fourth row of the temperature blocks. Temperature was recorded every 5 min for the 72-h duration of the experi-ments (Table S.2).
newly fertilized A. placenta (Fig. 2a) and C. rodgersii (Fig. 3a) embryos (20 egg ml−1) were placed into vials at the appropriate temperatures. Two temperature blocks, each with four rows, were used for experiments. For A. placenta, two experiments were conducted in May/June 2012: the first, lasting 48 h, used both temperature blocks simultaneously with all 8 replicate rows used and bringing water to temperature separately for a water change at 24 h, whilst the second experiment, lasting 72 h, used 7 rows as
replicates with the 8th used to keep additional vials at test temperature as multiple water changes were needed for the additional day. For C. rodgersii, two experiments were conducted in august 2012, each using a single temperature block with three replicate rows per treatment and a fourth row used for keeping isothermal water for water changes, thus achieving 6 replicates across both experiments. Arach-noides placenta and C. rodgersii larvae were cultured for up to 72 h to the 8-armed echinopluteus stage and the 2-armed echinopluteus transversus larva, respectively (Soars et al. 2009; gonzalez-Bernat et al. 2013), with par-tial replacement of half the volume of the treatment water (ca. 15 ml) at 24 and 48 h.
early embryonic and larval development
embryos and larvae were randomly sampled, by gently inverting the vials to homogenize their distribution and
Fig. 2 normal development in Arachnoides placenta: a fertilized egg, b cleavage, c late blastula, d 4-armed pluteus. e–h abnormal develop-ment: e arrested fertilized egg, f asymmetrical cleavage, g abnormal, degenerating blastula, h abnormal pluteus. Scale bar 100 μm
Fig. 3 normal development in Centrostephanus rodgersii: a fertilized egg, b cleavage, c blastula, d prism/early larva. Scale bar 100 μm

Mar Biol
1 3
3 ml was removed at each time point. For A. placenta, four time points were sampled by repeatedly sampling the vials at 6, 10, 24 and 48 h with an additional sample time point at 60 h in the second experiment, as the number of larvae left in the treatments after repeated sampling to 48 h in the first experiment was too low for analysis at 60 h. In the sec-ond experiment, we reared embryos to 72 h to determine the time to the 8-armed echinopluteus stage. For C. rodg-ersii, three time points were sampled repeatedly from the vials: 18, 42 and 72 h.
For both species, samples were scored according to devel-opmental stage reached as well as for abnormal/arrested development or mortality in random counts of the first 50 specimens encountered in each sample. normal and abnor-mal embryos of A. placenta are shown in Fig. 2a–h, and for C. rodgersii, normal development is shown in Fig. 3a–d. examples of abnormal development in C. rodgersii are avail-able in Doo et al. (2012). normal development included symmetrical cleavage embryos (Figs. 2b, 3b), considerations of shape in blastulae (Figs. 2c, 3c), and symmetrical pluteal arms (Fig. 2d) and shape (Fig. 3d) in larvae. This contrasted quite clearly with abnormal development in both species which included asymmetrical cleavage (Fig. 2f), deformed embryos and larvae at any stage (Fig. 2g), and arm asym-metries in plutei (Fig. 2h) (Doo et al. 2012). For 48 h post-fertilization, the 32 °C treatment for A. placenta had too few embryos (<10) and was excluded from the analysis.
For A. placenta, changes in the time of appearance of 4-, 6- and 8-armed stages across temperature treatments at 60 h were assessed. For C. rodgersii, changes in the devel-opmental chronology in cold treatments to the 2-armed pluteus characteristic of this species were estimated after 72 h. There were no morphological characteristics (i.e. additional arms) to assess the impact of increased tempera-ture on developmental timing after the appearance of this pair of arms. For each time point, the developmental stage achieved by the majority of embryos or larvae (>50 % stage achieved) was recorded for each species to assess changes to the timing of developmental stages.
Statistical analyses
Data from repeated sampling of the vials over time were analysed by one-way repeated measures analysis of vari-ance (rManOVa). To assess the potential effect of using two temperature blocks, a two-way rManOVa with tem-perature and block as fixed factors was used to analyse per-centage normal data in A. placenta for 6–48 h and for C. rodgersii for 18–72 h. The block effect was not significant (p > 0.25) for both experiments (A. placenta: F1,59 = 1.32, p = 0.254; C. rodgersii: F1,24 = 0.03, p = 0.402), and was therefore removed from the statistical model (Underwood 1997). Data in each treatment were thus pooled across
blocks. These data were subsequently analysed using a one-way rManOVa for each species with temperature as the fixed factor. The separate experiment to 60 h for A. placenta was analysed by two-way anOVa with tem-perature and block as fixed factors. although block was not removed (p < 0.25), the effect of block was not signifi-cant (F1,1 = 3.84, p = 0.057), and there was no interactive term of block and temperature (F7,7 = 1.41, p = 0.228) and therefore block was ignored.
To meet the assumptions of anOVa (homogeneity of variance and normality), percentage normal data underwent angular transformation prior to analyses. Homogeneity of variance was met when extreme treatments (i.e. 0 % survi-vorship) were removed from further analysis (A. placenta: 14, 35, 37 °C; C. rodgersii: 9, 11, 26–31 °C). Thus, for A. placenta, the number of temperature treatments included in the rManOVa was 10 for the initial 6-h time point (removal of 14, 37 °C), 9 for 10, 24 and 60 h (removal of 14, 35, 37 °C), and 8 for 48 h due to low recovery of larvae in the 32 °C treatment. Temperature treatments that were not included in the statistical analysis can be graphically analysed. For C. rodgersii, the number of temperature treat-ments included in the rManOVa was 6 at all time points (18–72 h). normality of data at 60 h for A. placenta was assessed by plotting residuals against normal distribution, but was not met due to trends in the data set being driven by extreme treatments. But as large data sets using anOVa analyses are robust to deviations of normality, the anOVa proceeded (Quinn and Keough 2002). The Mauchley’s test criterion was used to assess sphericity in the rManOVa analyses and, where it was met, univariate unadjusted p val-ues were used for the effects tests (A. placenta: Mauchley criterion = 0.9497, df = 5, p = 0.6951). Where it was not met, univariate greenhouse-geisser adjusted epsilon p val-ues were used (C. rodgersii: Mauchley criterion = 0.7022, df = 2, p = 0.006).
Tukey’s HSD post hoc tests were performed on one-way anOVas to identify significant differences between treatments. The thermal tolerances for A. placenta and C. rodgersii are illustrated for the 24- and 72-h time points, respectively. results for the other stages are presented in supplementary material (Table S.3, S.4, Figs. S.1–6). all statistical analyses were performed in JMP 8.0 (SaS Insti-tute Inc.).
Results
Sea surface temperatures
The latitudinal coastal marine isotherms for the autumn and winter spawning seasons in eastern australia are illus-trated (Fig. 1a, b). During the autumn/winter spawning and

Mar Biol
1 3
planktonic development period of A. placenta (april–June) near Townsville, the SST ranged from 22 to 28 °C with an average in May of 24.3 °C (n = 592, Se < 0.1) (Fig. 1a, b). During the winter spawning and planktonic develop-ment period of C. rodgersii (July–august), the SST of Syd-ney ranged from 15 to 20 °C, with an average in august of 17.6 °C (n = 6562, Se < 0.1) (Fig. 1b).
Thermal tolerance of development in A. placenta
In the first experiment, where A. placenta embryos were reared to 48 h, percentage normal development was affected by temperature (rManOVa, F8,60 = 47.8, p < 0.001) (Table 2; Fig. 4). at 6-, 10-, 24- and 48-h normal develop-ment was >80 % between 17 and 31 °C (Fig. 3). Outside of this range, percentage normal development was signifi-cantly reduced (Fig. 4) and certain treatments were excluded from the analysis due to little or no recovery of larvae. no normal development occurred at the extreme cold and warm treatments (14, 35, 37 °C), from the beginning of the experi-ment (i.e. 100 % mortality). In the second experiment where larvae were reared to 60 h, a similar pattern was observed (anOVa, F8,60 = 3.5, p < 0.005; Table 3).
Developmental success at 32 °C varied among stages (Fig. 4). This temperature corresponds to 4 °C above the maximum temperature in May, 28 °C (Fig. 1). By 6 h, 67 % (Se = 7.6 %) of embryos were normal at 32 °C (Table 2). By 24 h, percentage normal development decreased to 55 % (Se = 10.7 %), and by 60 h to 35 % (Se = 12.9 %) (Fig. 4).
Tukey’s HSD post hoc tests showed that there was no difference in percentage normal embryos and larvae between each pair of temperature treatments from 19 to 31 °C for the 6- and 10-h time points, and between the 17 and 32 °C treatments for the 6- and 10-h time points (Table 2; Fig. 4). However, percentage normal develop-ment was <70 % at 32 °C and >80 % at 17 °C at 6 and 10 h (Fig. 4). at 24, 48 and 60 h, percentage normal larvae did not differ among the 17–31 °C treatments (Tables 2, 3; Fig. 4). The success of larval development was lower in the low- and high-temperature treatments (14 and >32 °C) (Tables 2, 3). Thus, the thermal envelope for development to the advanced echinopluteus stage in A. placenta was determined to be 17–31 °C (Fig. 4).
effect of temperature on larval development times of A. placenta
The faster progression of A. placenta through developmen-tal stages in warm treatments was evident from the first time point scored (6 h) (Table 4, Fig. S.1). Time to first swimming and gastrulation was reduced at warmer tem-peratures and increased at cooler temperatures (Table 4, Fig. S.1, S.2). The relationship between higher temperature and developmental stage achieved is illustrated for the plu-teus stage (24 h) (Fig. 5) and for other stages (Fig. S.1–4), showing more advanced development in warmer treat-ments. This relationship changed at the upper threshold temperature of 32 °C, where delayed embryos were evident (Fig. 4, S.1–4). at 72 h, larvae in control treatments were all 8-armed plutei, the terminal larval stage of A. placenta, whilst larvae in cool treatments were still 4-armed. The lar-val stages present in the temperature range were compared at 60 h (Table 4, Fig. S.3, S.4), with that at 72 h. at 17 °C, the 4-armed larvae were 36 h behind controls, which were 6-armed plutei (ca. 15 % increase in development time). In contrast, the larvae were forming their 8th pair of arms at 31 °C, 12 h ahead of controls (ca. 37.5 % decrease in devel-opment time) (Table 4).
Thermal tolerance of development in C. rodgersii
The percentage of normal C. rodgersii embryos and larvae reared to 72 h was affected by temperature at all time points (rManOVa, F5,30 = 1147, p < 0.001) (Table 5; Fig. 6). at 18 h, % blastulae was >95 % between 13 and 24 °C (Table 5; Fig. 6, S.5), although Tukey’s HSD post hoc tests indicated that the 13 and 24 °C treatments differed from the 15–22 °C treatments which had ~97 % normal blastulae. at temperatures above and below this range (13–24 °C), percentage normal blastulae was greatly reduced (Table 5; Fig. 6) and 6 treatments were excluded from the analysis due to little or no recovery of larvae, leaving 6 treatments
Table 2 repeated measures anOVa of percentage normal develop-ment data for Arachnoides placenta across time (6–48 h post-ferti-lization) in all temperature treatments with >0 % recoverable larvae (6 h: 10 treatments; 10, 24 and 60 h: 9 treatments; 48 h: 8 treatments) (n = 8)
The 32 °C treatment was removed from the analysis for 48 h due to low recovery of larvae. Tukey’s HSD test underlined treatments do not differ. F exact F, df degrees of freedom
Source of variation F(df) p
Between subjects
Temperature 47.78(8,60) <0.0001
Within subjects
Time 3.09(3,180) =0.0284
Time × temperature 3.77(24,180) <0.0001
Mauchley criterion = 0.9497, df = 5, p = 0.6951
Time point (h) Tukey’s HSD post hoc test on univariate anOVas
6 35° < 17°, 32° < 19°, 21°, 24°, 26°, 27°, 29°, 31°
10 17°, 32° < 19°, 21°, 24°, 26°, 27°, 29°, 31°
24 32° < 17°, 19°, 21°, 24°, 26°, 27°, 29°, 31°
48 17°, 19°, 21°, 24°, 26°, 27°, 29°, 31°

Mar Biol
1 3
analysed by rManOVa. The thermal tolerance differed with respect to time and developmental stage (Fig. 6). at 42 h, percentage normal gastrulae was >90 % between 15 and 22 °C, with percentage normal gastrulae greatly reduced outside of this range (13 °C: 40 ± 2.6 %; 24 °C: 45 ± 2 %) (Table 5; Fig. 6). Tukey’s HSD post hoc tests indicated that the 13 and 24 °C treatments differed from 15 to 22 °C, but not from each other. However, the reduction
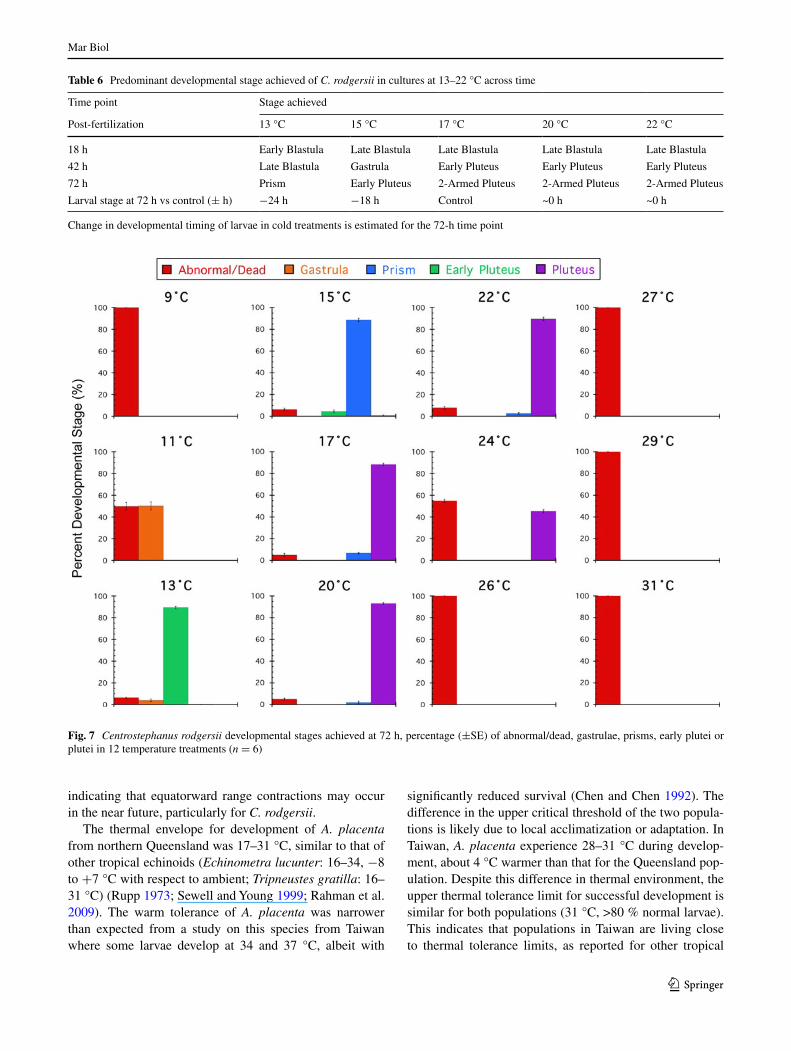
in percentage normal embryos at 13 °C was due not to abnormal development or mortality, as at 24 °C, but due to slower development, with >92 % of the embryos in the normal late blastula stage. at 72 h, the temperature range for normal prism/early plutei was 13–22 °C, with >90 % normal prisms/early plutei (Fig. 6). Outside of this range, there were <50 % prisms/plutei at 24 °C. There was no nor-mal development in extreme cold and warm treatments (9, 26–31 °C) (Fig. 6). Thus, the thermal envelope for normal development to the 2-armed echinopluteus of C. rodgersii was determined to be 13–22 °C.
effects of temperature on larval development times of C. rodgersii
The faster progression of C. rodgersii through develop-mental stages in warm treatments was evident from the first time point scored (18 h) (Table 6, Fig. S.5). The per-centage of individuals swimming and completing gastru-lation at each time point indicated that the time to reach both were reduced at warmer temperatures and increased at cooler temperatures (Table 6, Fig. S.5, S.6). The rela-tionship between increased temperature and development
Fig. 4 Percentage (±Se) of normally developing Arachnoides placenta embryos and larvae, in 12 temperature treatments from 6 to 48 h (n = 8) and to 60 h post-fertilization (n = 7)
Table 3 anOVa for percentage normal development data for Arach-noides placenta larvae at 60 h post-fertilization, in 9 temperature treatments with >0 % recoverable larvae (n = 7)
Tukey’s HSD test underlined treatments do not differ. F exact F, df degrees of freedom
Source of variation df F p
Temperature 8 11.56 <0.0001
Block 1 3.84 0.0571
Temperature × block 7 1.41 0.2275
error 52
C. Total 60
Tukey’s HSD 32° < 17°, 19°, 22°, 24°, 26°, 27°, 29°, 31°

Mar Biol
1 3
time is illustrated for the pluteus stage (72 h) (Fig. 7). Once C. rodgersii larvae reach the 2-armed stage, no additional arms are added, and therefore, developmental delay in cool temperatures was scored using this landmark (Table 6). By 72 h, larvae reared at 13 °C were at the prism stage, a delay of over 32 h (ca. 44 % increase in development time)
compared to the controls, which were 2-armed plutei at 17 °C (Table 6; Fig. 7). although development occurred at 11 °C, this was very slow with embryos still gastrulae at 72 h, a delay of 2 days compared to the controls, and most embryos reared at this temperature were dead or arrested (Fig. 7).
Table 4 Predominant developmental stage achieved of Arachnoides placenta in cultures at 17–31 °C across time for the 6–48-h experiment and for the 72-h experiment carried out separately
Change in developmental timing of larvae in cold and warm treatments is estimated for the 60-h time point
Time point Stage achieved
Post-fertilization 17 °C 22 °C 24 °C 27 °C 31 °C
6 h early Blastula early Blastula Hatching Hatching Mesenchyme Blastula
10 h early Blastula Hatching gastrula Prism Prism
24 h gastrula 2-armed Pluteus 4-armed Pluteus 4-armed Pluteus 4-armed Pluteus
48 h 2-armed Pluteus 6-armed Pluteus 6-armed Pluteus 6-armed Pluteus 6-armed Pluteus
60 h 4-armed Pluteus 6-armed Pluteus 6-armed Pluteus 6-armed Pluteus 6-/8-armed Pluteus
72 h 4-armed Pluteus 6-armed 6-/8-armed 6-/8-armed 8-armed
larval stage at 60 h vs control (± h) −36 h ~0 h Control ~0 h +12 h
Fig. 5 Arachnoides placenta developmental stages achieved at 24 h, percentage (±Se) of abnormal/dead, gastrulae, prisms, 4-armed plutei or 6-armed plutei in 12 temperature treatments (n = 8)

Mar Biol
1 3
Discussion
as the ocean warms, marine species are likely to exhibit changes in their developmental dynamics as seen here for Arachnoides placenta and Centrostephanus rodgersii, with faster development and shorter PlDs, up to tolerance lim-its. The thermal envelope for larval development in both species exceeded the temperature ranges experienced in the water column in their present habitats. early development proceeded at temperatures 7 °C below ambient in A. pla-centa (i.e. 17 °C) and 4 °C below ambient in C. rodgersii (i.e. 13 °C). Thus, thermal tolerance of the planktonic stage is unlikely to limit southern migration of these species in
the study populations. Our first hypothesis, that low tem-perature tolerance in A. placenta larvae may limit the southern distribution of this species, is rejected. Our second hypothesis, that low temperature tolerance in C. rodgersii larvae spawned in south-eastern australia facilitated inva-sion to Tasmania is supported, and propagules from these populations are likely to continue to be dispersed to Tas-mania. although development in A. placenta was observed at +7 °C and in C. rodgersii at +5 °C, most embryos were sensitive to a 4 °C increase above the maximal spawning temperatures. Thus, ocean warming may be more delete-rious to the embryos of C. rodgersii than those of A. pla-centa, providing partial support for hypothesis three, and
Table 5 repeated measures anOVa of percentage normal development data for Centrostephanus rodgersii embryos and larvae across time (18–72 h post-fertilization) in 6 temperature treatments with >0 % recoverable larvae (n = 6)
Tukey’s HSD test underlined treatments do not differ. F exact F, df degrees of freedom
Source of variation F(df) p
Between subjects
Temperature 366.03(5,30) <0.0001
Within subjects
Time 314.27(2,60) <0.0001
Time × temperature 148.61(10,60) <0.0001
Mauchley criterion = 0.7022, df = 2, p = 0.0059
Time point (h) Tukey’s HSD
18 24°, 13° < 15°, 17°, 20°, 22°
42 24°, 13° < 15°, 17°, 20°, 22°
72 24° < 13°, 15°, 17°, 20°, 22°
Fig. 6 Percentage (±Se) of normally developing Cen-trostephanus rodgersii embryos and larvae in 12 temperature treatments from 18 to 72 h post-fertilization (n = 6)
Mar Biol
1 3
indicating that equatorward range contractions may occur in the near future, particularly for C. rodgersii.
The thermal envelope for development of A. placenta from northern Queensland was 17–31 °C, similar to that of other tropical echinoids (Echinometra lucunter: 16–34, −8 to +7 °C with respect to ambient; Tripneustes gratilla: 16–31 °C) (rupp 1973; Sewell and Young 1999; rahman et al. 2009). The warm tolerance of A. placenta was narrower than expected from a study on this species from Taiwan where some larvae develop at 34 and 37 °C, albeit with
significantly reduced survival (Chen and Chen 1992). The difference in the upper critical threshold of the two popula-tions is likely due to local acclimatization or adaptation. In Taiwan, A. placenta experience 28–31 °C during develop-ment, about 4 °C warmer than that for the Queensland pop-ulation. Despite this difference in thermal environment, the upper thermal tolerance limit for successful development is similar for both populations (31 °C, >80 % normal larvae). This indicates that populations in Taiwan are living close to thermal tolerance limits, as reported for other tropical
Table 6 Predominant developmental stage achieved of C. rodgersii in cultures at 13–22 °C across time
Change in developmental timing of larvae in cold treatments is estimated for the 72-h time point
Time point Stage achieved
Post-fertilization 13 °C 15 °C 17 °C 20 °C 22 °C
18 h early Blastula late Blastula late Blastula late Blastula late Blastula
42 h late Blastula gastrula early Pluteus early Pluteus early Pluteus
72 h Prism early Pluteus 2-armed Pluteus 2-armed Pluteus 2-armed Pluteus
larval stage at 72 h vs control (± h) −24 h −18 h Control ~0 h ~0 h
Fig. 7 Centrostephanus rodgersii developmental stages achieved at 72 h, percentage (±Se) of abnormal/dead, gastrulae, prisms, early plutei or plutei in 12 temperature treatments (n = 6)

Mar Biol
1 3
invertebrates (Somero 2010; Morley et al. 2012). The tol-erance of A. placenta development to temperatures above ambient in Taiwan is also much narrower (+3 °C) than that of the australian population (+7 °C). Thus, in its current habitat, A. placenta will become increasingly vulnerable to warming, but its thermal tolerance indicates potential resil-ience to 4 °C warming.
The thermal envelope for development of C. rodgersii from mid-nSW was 13–22 °C, similar to that determined for the Tasmanian population, 12–20 °C, in a study where the larvae of C. rodgersii were reared for 21 days in tem-perature block treatments (ling et al. 2008). The similar developmental thermal envelopes in both populations are consistent with the finding that the nSW populations were the source of migrants to Tasmania (Banks et al. 2010). additionally, this is indicative of a lack of adaptation or acclimation by southern range edge populations to cooler climes since their invasion (~60 years ago) (see also ling et al. 2008). The lower thermal tolerance limit of develop-ment for C. rodgersii in Tasmania and nSW coincides with the average winter SST (12 °C) in Tasmania. Thus, the cool tolerance of development enabled southern range expan-sion of the propagules of this species (ling et al. 2008). as winter temperatures for Tasmania are predicted to reach 16–17 °C by 2100 (Hobday and lough 2011), the optimal developmental temperature for C. rodgersii, there may be changes in local population dynamics perhaps result-ing from greater reproductive success in resident popula-tions (ling et al. 2008). Continued propagule supply from mainland australia is also likely. Further range expansion of C. rodgersii will, however, be limited by the scarcity of shallow rocky reefs south of Tasmania on the edge of the Southern Ocean, but deeper-water reefs may provide habi-tat for expansion (Byrne and andrew 2013).
The thermal envelope for successful larval development in A. placenta (14 °C range, 17–31 °C) was broader than in C. rodgersii (9 °C range, 13–22 °C). This follows the trend towards broader developmental thermal tolerances for trop-ical echinoids compared with higher latitude species (Sun-day et al. 2011; Morley et al. 2012). Despite this greater thermotolerance, A. placenta has not undergone poleward range expansion. The latitudinal isotherm of 17 °C, the cool tolerance limit for development in A. placenta, is at 37°S in south-eastern australia ~1,600 km south of the present range edge of this species in Moreton Bay, where late autumn and winter temperatures average 21 °C. Thus, the distribution of this species does not realize the poten-tial indicated by the lower temperature tolerance of devel-opment. Why A. placenta are not present further south in australia is, therefore, interesting. Other factors such as the thermal tolerance of late-stage larvae, juvenile and adult life stages, habitat availability and oceanography may con-strain A. placenta to Queensland. In Queensland, this sand
dollar is most common on low-energy beaches comprised of medium to fine sand (Haycock 2004). The dissipative, high wave energy and coarser particle sandy beaches in nSW (Short 1993) may not be suitable habitat for A. pla-centa. Studies on sediment grain size and settlement suc-cess of juvenile A. placenta are needed to address this sug-gestion. Oceanographic connectivity may also influence the distribution of this species. In northern nSW at 32°S, the eaC gives rise to the eastward-flowing Tasman Front Current (Condie and Dunn 2006), a potential impediment to southerly migration of A. placenta larvae in the austral winter, although this has not prevented migration of some tropical fish larvae to temperate waters in the austral sum-mer (Booth et al. 2007).
With respect to ocean warming, development to the lar-val stage in A. placenta and C. rodgersii was robust to near-future projections for their planktonic habitat, with toler-ance to 31 °C for A. placenta and 22 °C for C. rodgersii. However, projected SSTs of 26–32 °C during the spawning season of A. placenta and 21–24 °C for C. rodgersii will push the development of the study populations to their pre-sent-day thermal limits. This level of temperature increase may be exacerbated by strong aerial warming (+5 °C by 2070) predicted for the coastal region (Hobday and lough 2011). Development in both species was sensitive to 4 °C above the present spawning season maxima (28 °C for A. placenta, 20 °C for C. rodgersii). This is consistent with the general vulnerability of sea urchin development to >4 °C warming (Byrne 2011). Centrostephanus rodgersii in nSW are living closer to the thermal maximum for suc-cessful development (+5 °C) compared with A. placenta (+7 °C). although there is evidence that C. rodgersii may have the capacity to adapt to higher temperatures, with the offspring of some males (but not others) developing suc-cessfully up to the gastrula stage at 25 °C (Foo et al. 2012), it is unknown if similar plasticity is present for later larval stages in this species. The potential for genetic adaptation will also affect species vulnerabilities in a changing ocean.
For many species, there is a tight relationship between sea temperature and spawning time, and this often corre-sponds to the optimal temperature for larval development (Johnson and Babcock 1994; reitzel et al. 2005; Byrne 2011). Spawning phenology in some invertebrates will likely change in response to ocean warming (Byrne 2011). In A. placenta, spawning may be shifted to late autumn or winter in northern australia, but the duration of the spawn-ing season may also change. The lower thermal tolerance for development in A. placenta indicates that low tempera-ture is not a factor limiting the spawning season in this spe-cies. Other factors such as day length may be more impor-tant, as shown for other sea urchins where photoperiod controls reproduction (Pearse et al. 1986). In C. rodger-sii, the spawning season is unlikely to change because the

Mar Biol
1 3
onset of spawning is tied to the winter solstice (Byrne and andrew 2013).
as expected, there was a strong effect of temperature on the duration of development in A. placenta and C. rodger-sii, being greatly reduced with just a 2–4 °C temperature increase, as documented for other echinoids (rupp 1973; Mcedward 1985; Fujisawa 1989, 1995; Chen and Chen 1992; Sewell and Young 1999; ling et al. 2008). Changes in the timing of planktonic development due to ocean warming can lead to trophic mismatch, as previously shown for other echinoids (edwards and richardson 2004; O’Connor et al. 2007). The PlDs of A. placenta (~7–14 days) and C. rodgersii (~3 months) can be expected to shorten in response to ocean warming, but may also stay unchanged or lengthen as larvae are dispersed to cooler climes. Ocean warming and reduced PlD alter larval dis-persal distance with potential unfavourable impacts on population connectivity, community structure and regional patterns of biodiversity (O’Connor et al. 2007). For species with short PlD, such as A. placenta, ocean temperatures for the source and sink populations at spawning and at lar-val settlement are likely to be similar. For C. rodgersii with long PlD, ocean temperatures at spawning will likely dif-fer from those at settlement. For the latter species, the tem-perature range applied in our experiments (9–31 °C) would likely encompass the range of sea temperatures encoun-tered by larvae during their development.
The thermal tolerance (cool and warm) of the early embryos of A. placenta and C. rodgersii was broader than for larvae, as previously found for E. lucunter (Sewell and Young 1999). For C. rodgersii, the thermal threshold for larvae was 22 °C compared to 24 °C for embryos. The broader thermal tolerance of early development is a fea-ture of echinoid embryos and may be due to protection by maternal protective factors (e.g. chaperone proteins) (Hamdoun and epel 2007). Differences in thermal toler-ance among developmental stages are typical of many marine invertebrates (Sewell and Young 1999; Storch et al. 2011; Higgins et al. 2012). as we only studied the effect of temperature on the early endotrophic stages of larval development, similar information on the planktotrophic lar-val stages and juveniles is needed to better understand the probable impact of changing ocean conditions.
although development in the two species was relatively robust to temperature change, subtler sublethal effects may limit the success of C. rodgersii and A. placenta in chang-ing planktonic conditions. Temperature (warm and cold) has been shown to affect heart rate, growth, survival, metabolism and swimming of crab, octopus and fish larvae (Pörtner and Knust 2007; Storch et al. 2011; Higgins et al. 2012). Chen and Chen (1992) found that the larvae of A. placenta reared at elevated temperatures were significantly smaller with shorter arms and gave rise to smaller juveniles. These thermal effects
on the body form of the echinoplutei have been found in other species and have important consequences for swimming and feeding (Mcedward 1985; Hart and Strathmann 1994; Soars et al. 2009). Thus, larval feeding rates may be considerably affected by temperature (rassoulzadegan and Fenaux 1979). Other parameters, in addition to the morphological criteria used here, affect larval success and the potential for south-ern range expansion. It is also important to understand how responses to stressors during the pelagic phase translate to juvenile settlement and recruitment (gosselin and Qian 1997; Pechenik 2006; Wolfe et al. 2013).
With respect to ocean warming and the developmental thermal maximum, it appears that the study populations of A. placenta and C. rodgersii are currently living relatively near their upper thermal threshold for successful develop-ment. Future climate-driven warming in their current hab-itats will likely lead to contractions at the northern range limits, as suggested for the temperate species Heliocidaris erythrogramma and other taxa (Byrne et al. 2011; Wer-nberg et al. 2011). regarding their potential for poleward range expansion, A. placenta populations are not realizing their potential poleward distribution as indicated by their lower thermal thresholds, whilst C. rodgersii populations are. Southern range expansion has already occurred in C. rodgersii providing southern refugia for this species, and ocean warming will improve conditions for spawning and development for this species at its current southern range edge, Tasmania. For A. placenta, however, other factors apparently limit its southern distribution, so development will be negatively affected by climate-driven warming in the absence of adaptation, acclimatization or range expan-sion. Overall, an understanding of the developmental enve-lope of marine ectotherms, as shown here for A. placenta and C. rodgersii, can provide valuable insights into the fac-tors influencing the realized and potential distribution of planktonic life stages, and changes to adult distribution in response to global change.
Acknowledgments This work was supported by grants from the australian research Council (MB and SaD). We gratefully acknowl-edge the assistance of Michelle liddy (aIMS). Dr alistair Hobday, CSIrO Marine laboratories, Hobart, is thanked for assistance with CarS data extraction and generation of isotherm maps. echino-derms were collected under permit licenses from the new South Wales Department of Industry and the great Barrier reef Marine Park authority.
References
andronikov VB (1975) Heat resistance of gametes of marine inverte-brates in relation to temperature conditions under which the spe-cies exist. Mar Biol 30:1–11
Baird aH, Sommer B, Madin JS (2012) Pole-ward range expansion of Acropora spp. along the east coast of australia. Coral reefs. doi:10.1007/s00338-012-0928-6

Mar Biol
1 3
Banks SC, Piggott MP, Williamson Je, Bové U, Holbrook nJ, Beheregaray lB (2007) Oceanic variability and coastal topogra-phy shape genetic structure in a long-dispersing sea urchin. ecol-ogy 88:3055–3064
Banks SC, ling SD, Johnson Cr, Piggott MP, Williamson Je, Beheregaray lB (2010) genetic structure of a recent climate change-driven range extension. Mol ecol 19:2011–2024
Barry JP, Baxter CH, Sagarin rD, gilman Se (1995) Climate-related, long-term faunal changes in a California rocky intertidal commu-nity. Science 267:672–675
Booth DJ, Figueira WF, gregson Ma, Brown l, Beretta g (2007) Occurrence of tropical fishes in temperate southeastern aus-tralia: role of the east australian Current. estuar Coast Shelf Sci 72:102–114
Byrne M (2010) Impact of climate change stressors on marine inver-tebrate life histories with a focus on the Mollusca and echinoder-mata. In: Yu J, Henderson-Sellers a (eds) Climate alert: climate change monitoring and strategy. University of Sydney Press, Syd-ney, pp 142–185
Byrne M (2011) Impact of ocean warming and ocean acidification on marine invertebrate life history stages: vulnerabilities and poten-tial for persistence in a changing ocean. Ocean Mar Biol annu rev 49:1–42
Byrne M, andrew nl (2013) Centrostephanus rodgersii. In: law-rence JM (ed) edible sea urchins: biology and ecology. elsevier Science, amsterdam
Byrne M, Soars n, Ho M-a, Wong e, Mcelroy D, Selvakumar-aswamy P, Sheppard-Brennand H, Dworjanyn Sa, Davis ar (2010) Fertilisation in a suite of coastal marine invertebrates from Se australia is robust to near-future ocean warming and acidifi-cation. Mar Biol 157:2061–2069
Byrne M, Selvakumaraswamy P, Ho M-a, nguyen HD (2011) Sea urchin development in a global change hotspot, potential for southerly migration of thermotolerant propagules. Deep-Sea res 58:712–719
Chen C-P, Chen B-Y (1992) effects of high temperature on larval development and metamorphosis of Arachnoides placenta (echi-nodermata: echinoidea). Mar Biol 112:445–449
Condie Sa, Dunn Jr (2006) Seasonal characteristics of the surface mixed layer in the australasian region: implications for pri-mary production regimes and biogeography. Mar Freshw res 57:569–590
Doo SS, Dworjanyn Sa, Foo Sa, Soars na, Byrne M (2012) Impacts of ocean acidification on development of the meroplanktonic larval stage of the sea urchin Centrostephanus rodgersii. ICeS J Mar Sci 69:460–464
edwards M, richardson aJ (2004) Impact of climate change on marine pelagic phenology and trophic mismatch. nature 430:881–884
Figueira WF, Booth DJ (2010) Increasing ocean temperatures allow tropical fishes to survive over winter in temperate waters. glob Change Biol 16:506–516
Foo Sa, Dworjanyn Sa, Poore agB, Byrne M (2012) adaptive capacity of the habitat modifying sea urchin Centrostephanus rodgersii to ocean warming and ocean acidification: performance of early embryos. PloS One 7:e42497
Fujisawa H (1989) Differences in temperature dependence of early development of sea urchins with different growing seasons. Biol Bull 176:96–102
Fujisawa H (1995) Variation in embryonic temperature sensitivity among groups of the sea urchin, Hemicentrotus pulcherrimus, which differ in their habitats. Zool Sci 12:583–589
gonzalez-Bernat MJ, lamare M, Uthicke S, Byrne M (2013) Fer-tilisation, embryogenesis and larval development in the tropi-cal intertidal sand dollar Arachnoides placenta in response to reduced seawater pH. Mar Biol 160:1927–1941. doi:10.1007/s00227-012-2034-2
gosselin la, Qian PY (1997) Juvenile mortality in benthic marine invertebrates. Mar ecol Prog Ser 146:265–282
Hadfield Mg, Strathmann MF (1996) Variability, flexibility and plasticity in life histories of marine invertebrates. Oceanol acta 19:323–324
Hamdoun a, epel D (2007) embryo stability and vulnerability in an always changing world. Proc natl acad Sci 104:1745–1750
Hart MW, Scheibling re (1988) Heat waves, baby booms and the destruction of kelp beds by sea urchins. Mar Biol 99:167–176
Hart MW, Strathmann rr (1994) Functional consequences of pheno-typic plasticity in echinoid larvae. Biol Bull 186:291–299
Haycock lJ (2004) The reproduction and recruitment of the sand dol-lar Arachnoides placenta (l.) (echinoidea: echinodermata) from differing habitats on the north Queensland coast. MS research Thesis, James Cook University, Queensland
Higgins Fa, Bates ae, lamare MD (2012) Heat tolerance, behav-ioural temperature selection and temperature-dependent respira-tion in larval Octopus huttoni. J Therm Biol 37:83–88
Hobday aJ, lough JM (2011) Observed climate change in aus-tralian marine and freshwater environments. Mar Freshw res 62:984–999
Hoegh-guldberg O, Pearse JS (1995) Temperature, food availabil-ity, and the development of marine invertebrate larvae. am Zool 14:415–425
International Panel on Climate Change (IPCC) (2007) Climate change 2007: the fourth assessment report of the International Panel on Climate Change (IPCC). Cambridge University Press, Cambridge
Johnson lg, Babcock rC (1994) Temperature and the larval ecology of the Crown-of-thorns starfish, Acanthaster planci. Biol Bull 187:304–308
Johnson Cr, Banks SC, Barrett nS, Cazassus F, Dunstan PK, edgar gJ, Frusher SD, gardner C, Haddon M, Helidoniotis F, Hill Kl, Holbrook nJ, Hosie gW, last Pr, ling SD, Melbourne-Thomas J, Miller K, Pecl gT, richardson aJ, ridgway Kr, rintoul Sr, ritz Da, ross DJ, Sanderson JC, Shepherd Sa, Slotwinski a, Swadling Ka, Taw n (2012) Climate change cascades: shifts in oceanography, species’ ranges and subtidal marine community dynamics in eastern Tasmania. J exp Mar Biol ecol 400:17–32
Jones SJ, Mieszkowska n, Wethey DS (2009) linking thermal toler-ances and biogeography: Mytilus edulis (l.) at its southern limit on the east coast of the United States. Biol Bull 217:73–85
ling SD, Johnson Cr, Frusher S, King CK (2008) reproductive potential of a marine ecosystem engineer at the edge of a newly expanded range. glob Change Biol 14:907–915
ling SD, Johnson Cr, ridgway K, Hobday aJ, Haddon M (2009) Climate-driven range extension of a sea urchin: inferring future trends by analysis of recent population dynamics. glob Change Biol 15:719–731
Mcedward lr (1985) effects of temperature on the body form, growth, electron transport system activity, and development rate of an echinopluteus. J exp Mar Biol ecol 93:169–181
Miskelly a (2002) Sea urchins of australia and the Indo-Pacific. Cap-ricornia Publications, Sydney
Morley Sa, Martin SM, Day rW, ericson J, lai C-H, lamare M, Tan K-S, Thorne MaS, Peck lS (2012) Thermal reaction norms and the scale of temperature variation: latitudinal vulnerability of intertidal nacellid limpets to climate change. PlosOne 7:e52818
O’Connor MI, Bruno JF, gaines SD, Halpern BS, lester Se, Kinlan BP, Weiss JM (2007) Temperature control of larval dispersal and the implications for marine ecology, evolution and conservation. Proc natl acad Sci 104:1266–1271
Palmer ar (1994) Temperature sensitivity, rate of development, and time to maturity: geographic variation in laboratory reared Nucella and a cross-phyletic overview. In: Wilson WH Jr, Stricker Sa, Shin gl (eds) reproduction and development of marine inverte-brates. Johns Hopkins University Press, Baltimore, pp 177–194

Mar Biol
1 3
Pearse JS, Pearse VB, Davis KK (1986) Photoperiodic regulation of gametogenesis and growth in the sea urchin Strongylocentrotus purpuratus. J exp Zool 237:107–118
Pechenik Ja (1987) environmental influences on larval survival and development. In: giese aC, Pearse JS (eds) reproduction of marine invertebrates. academic Press, new York, pp 551–608
Pechenik Ja (2006) larval experience and latent effects—metamor-phosis is not a new beginning. Int Comp Biol 46:323–333
Pecorino D, lamare MD, Barker MF, Byrne M (2013) Does embry-onic and larval thermal tolerance control the distribution of the sea urchin Centrostephanus rodgersii (Diadematidae) in new Zealand? J exp Mar Biol ecol 445:120–128
Poloczanska eS, Babcock rC, Butler a, Hobday a, Hoegh-guldberg O, Kunz TJ, Matear r, Milton Da, Okey Ta, richardson aJ (2007) Climate change and australian marine life. Oceanogr Mar Biol annu rev 45:407–478
Pörtner H-O, Knust r (2007) Climate change affects marine fishes through the oxygen limitation of thermal tolerance. Science 315:95–97
Pörtner H-O, Peck l, Somero gn (2007) Thermal limits and adapta-tion in marine antarctic ectotherms: an integrative view. Philos Trans r Soc B 362:2233–2258
Quinn gP, Keough MJ (2002) experimental design and data analysis for biologists. Cambridge University Press, Cambridge
rahman S, Tsuchiya M, Uehara T (2009) effects of temperature on hatching rate, embryonic development and early larval survival of the edible sea urchin, Tripneustes gratilla. Biologia 64:768–775
rassoulzadegan F, Fenaux l (1979) grazing of echinoderm larvae (Paracentrotus lividus and Arbacia lixula) on naturally occurring particulate matter. J Plankton res 1:215–223
reitzel aM, Miles CM, Heyland a, Cowart JD, Mcedward lr (2005) The contribution of the facultative feeding period to echi-noid larval development and size at metamorphosis: a compara-tive approach. J exp Mar Biol ecol 317:189–201
ridgway Kr (2007) long-term trend and decadal variability of the southward penetration of the east australian Current. geophys res lett 34:l13613
rupp JH (1973) effects of temperature on fertilization and early cleavage of some tropical echinoderms, with emphasis on Echi-nometra mathaei. Mar Biol 23:183–189
Sewell Ma, Young CM (1999) Temperature limits to fertilization and early development in the tropical sea urchin Echinometra lucunter. J exp Mar Biol ecol 236:291–305
Short aD (1993) Beaches of the new South Wales Coast: a guide to their nature, characteristics, surf and safety. australian Beach Safety and Management Program, nSW
Soars na, Prowse Taa, Byrne M (2009) Overview of phenotypic plasticity in echinoid larvae, ‘Echinopluteus transversus’ type vs. typical echinoplutei. Mar ecol Prog Ser 383:113–125
Somero gn (2010) The physiology of climate change: how potentials for acclimatization and genetic adaptation will determine ‘win-ners’ and ‘losers’. J exp Biol 213:912–920
Storch D, Fernández M, navarrete Sa, Pörtner H-O (2011) Thermal tolerance of larval stages of the Chilean kelp crap Taliepus denta-tus. Mar ecol Prog Ser 429:157–167
Sunday JM, Bates ae, Dulvy nK (2011) global analysis of ther-mal tolerance and latitude in ectotherms. Proc r Soc B 278:1823–1830
Sunday JM, Bates ae, Dulvy nK (2012) Thermal tolerance and the global redistribution of animals. nat Clim Change 2:686–690
Swanson rl, Byrne M, Prowse Taa, Mos B, Dworjanyn Sa, Stein-berg PD (2012) Dissolved histamine: a potential habitat marker promoting settlement and metamorphosis in sea urchins larvae. Mar Biol 159:915–925
Underwood aJ (1997) experiments in ecology: their logical design and interpretation using analysis of variance. Cambridge Univer-sity Press, Cambridge
Walther g-r, Post e, Convey P, Menzel a, Parmesan C, Beebee TJC, Fromentin J-M, Hoegh-guldberg O, Bairlein F (2002) ecological responses to recent climate change. nature 416:389–395
Wernberg T, russel BD, Moore PJ, ling SD, Smale Da, Camp-bell a, Coleman Ma, Steinberg PD, Kendrick ga, Connell SD (2011) Impacts of climate change in a global hotspot for temper-ate marine biodiversity and ocean warming. J exp Mar Biol ecol 400:7–16
Wolfe KDl, Dworjanyn Sa, Byrne M (2013) effects of ocean warm-ing and acidification on survival, growth and skeletal develop-ment in the early benthic juvenile sea urchin, (Heliocidaris eryth-rogramma). glob Change Biol 19:2608–2707
Copyright © 2022 FDOKUMEN




















