Localization of fodrin during fertilization and early development of sea urchins and mice
10
DEVELOPMENTAL BIOLOGY 118,457-466 (1986) Localization of Fodrin during Fertilization and Early Development of Sea Urchins and Mice HEIDE SCHATTEN,’ RICHARD CHENEY,* RON BALCZON~ MARK WILLARD,* CHRISTI CLINE, CALVIN SIMERLY,’ AND GERALD SCHATTEN’ Department of Biological Science, Florida State University, Tallahassee, Florida 52306 and “Deportment of Anatomy and Neurobiology, School of Medicine, Washington University, St. Louis, Missouri 63110 Received March 17, 1986; accepted in revised fwm June 10, 1986 Fodrin, a spectrin-like protein, is localized in gametes, zygotes, and embryos from sea urchins and mice. Mammalian fodrin comprises two polypeptides with molecular weights of approximately 240 kDa (alpha) and 235 kDa (beta). An antibody specific for mammalian a-fodrin cross-reacted with a 240-kDa polypeptide from sea urchin egg extracts. This indicates that sea urchins contain a protein of similar electrophoretic mobility and immunological properties to mam- malian a-fodrin. When this antibody was used to stain the sea urchin gametes with indirect immunofluorescence, fodrin- specific fluorescence was localized to the acrosome of the sperm and was distributed over the entire egg near the surface in a punctate pattern similar to the distribution of polymeric actin. During sperm incorporation, the fodrin-specific fluorescence is found at the site of sperm incorporation, in the fertilization cone. After fertilization, the intensity of fodrin fluorescence increases. During mitosis and eytokinesis in sea urchins, the entire surface of the egg remains stained, the cleavage furrow also was stained but no more intensely than was the rest of the egg surface. Antibody labeling with colloidal gold followed by electron microscopy showed that fodrin was loated in the cytoplasm immediately beneath the plasma membrane. In unfertilized mouse oocytes, both actin and fodrin were stained most intensely beneath the membrane adjacent to the meiotic spindle. After insemination, the cell surfaces of the pronucleate egg and the second polar body were stained, however, the actin matrix surrounding the apposed pronuclei did not bind the fodrin antibody. During cytokinesis in the mouse, the cleavage furrow stained more intensely than did the rest of the egg cortex, and in embryos the cell borders were delineated. These results indicate that organisms as unrelated to mammals as sea urchins have fodrin-like proteins; the rearrangements of such proteins suggest that they participate in the actin- mediated events at the cell surface during fertilization and early development in both mice and sea urchins. o 1986 Academic Press, Inc. INTRODUCTION Dramatic examples of actin reorganization on the cy- toplasmic face of the plasma membrane occur during fertilization and early development (reviewed by Vac- quier, 1981). In sea urchins, these events include the as- sembly of microfilament bundles in sperm during the acrosome reaction (Tilney, 1976a,b), the engulfment of the sperm by egg microvilli during sperm incorporation (Schatten and Schatten, 1980), the elongation of micro- villi after fertilization (Eddy and Shapiro, 1976; Schroe- der, 1981), and the formation of the cleavage furrow at cytokinesis (reviewed by Schroeder, 1981). During mouse fertilization, actin also undergoes dynamic rearrange- ments: In the unfertilized oocyte, actin is concentrated at the cortex adjacent to the meiotic spindle as well as in the microvilli covering the remainder of the egg sur- face (Maro et al., 1984; Longo and Chen, 1985). In pronu- 1 Present address: Integrated Microscopy Facility for Biomedical Research, Zoology Research Building, University of Wisconsin, 1117 West Johnson St., Madison, Wise. 53706. ’ Present address: Department of Cell Biology and Anatomy, Uni- versity of Alabama, School of Medicine, Birmingham, Ala., 35294. cleate eggs, cytoplasmic actin is found surrounding the apposed pronuclei (Maro et al., 1984) in addition to the cortical concentrations. Because spectrin-like proteins, such as fodrin, are widely distributed and are actin-binding proteins with central roles in the cortical cytoskeleton, and because these proteins redistribute in other major reorganiza- tions of the cell surface such as capping and exocytosis (Levine and Willard, 1983), it seemed likely that fertil- ization and early development may be accompanied by changes in the distribution and organization of spectrin- like proteins. In order to investigate the relationship of fodrin to these surface rearrangements, we have com- pared the distribution of fodrin and actin during fertil- ization, cytokinesis, and early development in sea ur- chins and mice. MATERIALS AND METHODS Gamete collection and fertilization. Gametes from the Gulf Coast sea urchin Lytechinus variegatus were col- lected by intracoelomic stimulation and fertilization was 457 0012-1606186 $3.00 Copyright 0 1986 by Academic Press. Inc. All rights of reproduction in any form reserved.
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Localization of fodrin during fertilization and early development of sea urchins and mice

DEVELOPMENTAL BIOLOGY 118,457-466 (1986)
Localization of Fodrin during Fertilization and Early Development of Sea Urchins and Mice
HEIDE SCHATTEN,’ RICHARD CHENEY,* RON BALCZON~ MARK WILLARD,* CHRISTI CLINE, CALVIN SIMERLY,’ AND GERALD SCHATTEN’
Department of Biological Science, Florida State University, Tallahassee, Florida 52306 and “Deportment of Anatomy and Neurobiology, School of Medicine, Washington University, St. Louis, Missouri 63110
Received March 17, 1986; accepted in revised fwm June 10, 1986
Fodrin, a spectrin-like protein, is localized in gametes, zygotes, and embryos from sea urchins and mice. Mammalian fodrin comprises two polypeptides with molecular weights of approximately 240 kDa (alpha) and 235 kDa (beta). An antibody specific for mammalian a-fodrin cross-reacted with a 240-kDa polypeptide from sea urchin egg extracts. This indicates that sea urchins contain a protein of similar electrophoretic mobility and immunological properties to mam- malian a-fodrin. When this antibody was used to stain the sea urchin gametes with indirect immunofluorescence, fodrin- specific fluorescence was localized to the acrosome of the sperm and was distributed over the entire egg near the surface in a punctate pattern similar to the distribution of polymeric actin. During sperm incorporation, the fodrin-specific fluorescence is found at the site of sperm incorporation, in the fertilization cone. After fertilization, the intensity of fodrin fluorescence increases. During mitosis and eytokinesis in sea urchins, the entire surface of the egg remains stained, the cleavage furrow also was stained but no more intensely than was the rest of the egg surface. Antibody labeling with colloidal gold followed by electron microscopy showed that fodrin was loated in the cytoplasm immediately beneath the plasma membrane. In unfertilized mouse oocytes, both actin and fodrin were stained most intensely beneath the membrane adjacent to the meiotic spindle. After insemination, the cell surfaces of the pronucleate egg and the second polar body were stained, however, the actin matrix surrounding the apposed pronuclei did not bind the fodrin antibody. During cytokinesis in the mouse, the cleavage furrow stained more intensely than did the rest of the egg cortex, and in embryos the cell borders were delineated. These results indicate that organisms as unrelated to mammals as sea urchins have fodrin-like proteins; the rearrangements of such proteins suggest that they participate in the actin- mediated events at the cell surface during fertilization and early development in both mice and sea urchins. o 1986
Academic Press, Inc.
INTRODUCTION
Dramatic examples of actin reorganization on the cy- toplasmic face of the plasma membrane occur during fertilization and early development (reviewed by Vac- quier, 1981). In sea urchins, these events include the as- sembly of microfilament bundles in sperm during the acrosome reaction (Tilney, 1976a,b), the engulfment of the sperm by egg microvilli during sperm incorporation (Schatten and Schatten, 1980), the elongation of micro- villi after fertilization (Eddy and Shapiro, 1976; Schroe- der, 1981), and the formation of the cleavage furrow at cytokinesis (reviewed by Schroeder, 1981). During mouse fertilization, actin also undergoes dynamic rearrange- ments: In the unfertilized oocyte, actin is concentrated at the cortex adjacent to the meiotic spindle as well as in the microvilli covering the remainder of the egg sur- face (Maro et al., 1984; Longo and Chen, 1985). In pronu-
1 Present address: Integrated Microscopy Facility for Biomedical Research, Zoology Research Building, University of Wisconsin, 1117 West Johnson St., Madison, Wise. 53706.
’ Present address: Department of Cell Biology and Anatomy, Uni- versity of Alabama, School of Medicine, Birmingham, Ala., 35294.
cleate eggs, cytoplasmic actin is found surrounding the apposed pronuclei (Maro et al., 1984) in addition to the cortical concentrations.
Because spectrin-like proteins, such as fodrin, are widely distributed and are actin-binding proteins with central roles in the cortical cytoskeleton, and because these proteins redistribute in other major reorganiza- tions of the cell surface such as capping and exocytosis (Levine and Willard, 1983), it seemed likely that fertil- ization and early development may be accompanied by changes in the distribution and organization of spectrin- like proteins. In order to investigate the relationship of fodrin to these surface rearrangements, we have com- pared the distribution of fodrin and actin during fertil- ization, cytokinesis, and early development in sea ur- chins and mice.
MATERIALS AND METHODS
Gamete collection and fertilization. Gametes from the Gulf Coast sea urchin Lytechinus variegatus were col- lected by intracoelomic stimulation and fertilization was
457 0012-1606186 $3.00 Copyright 0 1986 by Academic Press. Inc. All rights of reproduction in any form reserved.

458 DEVELOPMENTAL BIOLOGY VOLUME 118, 1986
performed by conventional methods. Superovulated mouse oocytes, denuded of the cumulus and zona, were processed directly for studies on unfertilized oocytes or fertilized in vitro with epididymal sperm (Whittingham, 1968) and cultured through first division. Blastocysts were collected from naturally mated CD-l females at 96 hr after the estimated time of ovulation.
Antibody purification and western blotting. Fodrin from guinea pig brain and affinity-purified antifodrin antibody were prepared as described by Levine and Wil- lard (1981). The antifodrin used in this study reacts with the 240-kDa a-subunit of mammalian fodrin, but not with the 235-kDa P-subunit. Crude gels from unfertilized sea urchin eggs were prepared following the methods of Kane (1980), and mitotic sea urchin eggs were extracted by the protocol of Balczon and Schatten (1983). These preparations were dissolved in sample buffer (Laemmli, 1970) and analyzed by western blotting after electro- phoresis on SDS polyacrylamide gradient gels (4-12% w/v linear gradient of acrylamide with a constant 33.3/ 1 w/w ratio of acrylamide to N,N’-methylenebisacryl- amide), using the buffer system of Laemmli (1970). Electrophoretic transfer to nitrocellulose, amido black staining of the transferred proteins, and immunochem- ical staining were carried out as described by Hirokawa et a2. (1983), except that the transfer buffer contained 0.1% SDS. Nitrocellulose strips were incubated in 5 ml of affinity-purified antifodrin or of nonimmune IgG at a concentration of 1 pg/ml, and were stained using a biotinylated secondary antibody and an avidin-biotin- ylated horseradish peroxidase complex (Hirokawa et al, 1983).
Immunojhorescence, jluorescence, and immunoelec- tron microscopy. Immunofluorescence microscopy was performed with sea urchin eggs following the extraction protocol of Balczon and Schatten (1983). Mouse oocytes and zygotes were extracted in 25% glycerol, 50 mMKC1, 0.5 mM MgCl,, 0.1 mM EDTA, 1 mM EGTA, 1 mM 2- mercaptoethanol, 50 mM imidazole, pH 6.7, with 1% Tri- ton X-100 (Schatten et al, 1985). Sperm were fixed with- out extraction. The cells were affixed to polylysine-coated coverslips (Mazia et al, 1975), fixed in methanol at -lO”C, and incubated in affinity-purified fodrin antibody for 45 min at 37°C in a humidified environment. Following rinses with phosphate-buffered saline, staining with flu- orescein-labeled goat anti-rabbit second antibody (Cap- pel Laboratories), and a final rinse in phosphate-buffered saline, Zeiss epifluorescence microscopy with appropriate filter sets was used. The Hoechst DNA binding dye No. 33258 (2.5 pg/ml) was included in the penultimate rinsing medium.
Actin and fodrin were colocalized by permeabilization with 50 pg/ml of lysolecithin and fixation with 2% para- formaldehyde in calcium-free seawater, pH 8.1 (Cline and Schatten, 1986), or phosphate-buffered saline. Rho- daminyl-phalloidin (Wieland and Faulstich, 1977; Barak
et ah, 1981), courtesy of Professor Th. Wieland (Max- Planck-Institut, Heidelberg), was applied at a concen- tration of 40 rig/ml in a humidified chamber for 1 hr at 37°C. Following three washes with phosphate-buffered saline, the protocols previously described for antifodrin immunofluorescence and DNA fluorescence microscopy were performed.
Immunoelectron microscopy with protein A gold (DeMey, 1983; Erdos and Whitaker, 1983), courtesy of Dr. G. Erdos and Dr. H. Aldrich (University of Florida), was performed on detergent-extracted sea urchin eggs, incubated with fodrin antibody and then protein A-gold, and embedded in Spurr’s resin. Electron microscopy was performed with a JEOL 100 CX-II scanning transmission electron microscope. As controls for the immunogold la- beling, parallel samples were labeled with only protein A-gold in which case few particles remained after pro- cessing, or with affinity-purified antitubulin antibody as the primary antibody in which case the cell surface was free of gold particles and the cytoplasmic microtubules were heavily laden with gold.
RESULTS
Western blots were performed to determine whether sea urchin eggs contain polypeptides related to mam- malian fodrin. An antibody which is specific for the a- subunit of guinea pig fodrin (Fig. la) reacted with a single polypeptide of similar molecular weight (approx- imately 240 kDa) from cytoplasmic gels produced from homogenates of unfertilized sea urchin eggs by the pro- cedure of Kane (1980) (Figs. lc-e) and from cytomatrix extracts of mitotic sea urchin eggs (Figs. If-h). The ex- traction protocol used to prepare the cytomatrix fraction (Figs. If-h) removes most all of the cellular actin; in the gelated fraction (Figs. lc-e) a band at 43 kDa probably corresponds to total egg actin. This result indicates that certain immunological and physical properties of fodrin- like proteins are shared in organisms as distantly related as guinea pigs and sea urchins.
This antibody was used to determine the location of fodrin-like proteins in sea urchin gametes and zygotes by means of indirect immunofluorescence microscopy. In sea urchin sperm, it reacted specifically with the ac- rosomal region (Fig. 2a). Corresponding sperm heads, imaged with the DNA-specific fluorescence Hoechst dye No. 33258, are marked with arrows in Fig. 2b.
In unfertilized eggs, fodrin-specific fluorescence formed a pun&ate pattern over the entire cortex of the egg (Fig. 3a). This arrangement is very similar to that produced by the staining of cortical actin with rhoda- minyl phalloidin (Fig. 3b).
During sperm incorporation, antifodrin immunoflu- orescence detects an increased fluorescence at the site of sperm entry, in the fertilization cone, particularly apparent after detergent extraction (Fig. 5). In Fig. 4

SCHATTEN ET AL. FoclrCn during Fertilization 459
a b c d e f g h FIG. 1. Western blotting with antifodrin. Samples were electropho-
resed on SDS polyacrylamide gradient gels, transferred to nitrocel- lulose, and then the nitrocellulose strips were incubated with antibodies and staining reagents as described under Materials and Methods. (a, b) The purified 240-kDa subunit of mammalian fodrin stained with antifodrin or nonimmune IgG, respectively. (c, d) Cytoplasmic gels from unfertilized sea urchin eggs stained with anti-+&-in or nonim- mune IgG, respectively. An immunoreactive polypeptide with an ap- parent molecular weight of 240 kDa is specifically stained by the anti- fitiin (e) Proteins of the cytoplasmic gels from unfertilized sea urchin eggs visualized by amido black staining. (f, g) Mitotic sea urchin egg extracts stained with anti-f&in or nonimmune IgG, respectively. This sample also contains an immunoreactive polypeptide with an apparent molecular weight of 240 kDa that is selectively stained by the anti- fodrin (h) Proteins of the mitotic sea urchin eggs visualized by amido black staining. Actin is removed during the detergent extraction. Ap- proximately 40 pg of protein was loaded per lane c, d, and e, while 80 wg was loaded per lane in f, g, and h. The molecular weights in kilo- daltons of standard proteins are indicated at the left.
on a permeabilized egg, triple staining of an egg for actin (Fig. 4a), fodrin (Fig. 4b), and DNA of the male and female pronuclei (Fig. 4~) demonstrates that fodrin and actin are found together with a punctate distribution at the egg cortex. The recruitment of actin in the fertiliza- tion cone is apparent (Fig. 4a, arrow) and fodrin may be similarly concentrated at this site (Fig. 4b, arrow). In Figs. 5a-c on detergent-extracted eggs, the accumu- lation of fodrin in fertilization cones (arrows) is more apparent. Though both actin and fodrin are preserved during lysolecithin permeabilization (Fig. 4), the actin is removed during Triton extraction in Fig. 5. The loss of this protein during extraction is shown in Fig. lh. This suggests that the fodrin may be more firmly as- sociated with the cytomatrix than is the actin.
At division, fodrin retains its pun&ate distribution at the cell surface and does not appear to concentrate at the cleavage furrow. In Fig. 6a at telophase, fodrin is found at pun&ate sites uniformly distributed on the en- tire cell surface. During cytokinesis (Fig. 6~) there does not appear to be a marked accumulation at the cleavage furrow.
At times, the egg is sheared during processing, leaving the bottom surface of the egg alIixed to the substrate. Inspection of these isolated egg surfaces demonstrates that there is a close, but not precise, correspondence between actin fluorescence (Fig. ‘7a) and fodrin immu- nofluorescence (Fig. 7b). Treatment of unfertilized eggs with 10 PM cytochalasin E results in patchy patterns of fluorescence for both actin (Fig. 7c) and fodrin (Fig. 7d).
Immunoelectron microscopy with protein A gold on detergent-extracted eggs demonstrates that fodrin is confined to the egg surface, near microvilli, as shown in
FIG. 2. Fodrin at the acrosomes in sea urchin sperm. Antifodrin antibodies stain only at the acrosomal region in sperm (a). Corresponding sperm are imaged with the DNA fluorescent Hoechst dye No. 33258 and are marked with arrows in b. Double-labeled sperm for fodrin immunofluorescence microscopy (a) and for DNA fluorescence (b). Bar = 10pm.

460 DEVELOPMENTAL BIOLOGY VOLUME 118, 1986
FIG. 3. Fodrin and actin detection in unfertilized sea urchin eggs. Fodrin is localized in a punctate pattern which is restricted to the egg surface in an unfertilized sea urchin egg (a). Polymeric actin, detected with rhodaminyl-phalloidin fluorescence in a different egg, appears to have a similar distribution and is also restricted to the cortex in b. Bar = 10 pm.
FIG. 4. Colocalization of fodrin and actin in a permeabilized fertilized sea urchin egg. At 10 min postinsemination, actin fluorescence (a) and fodrin immunofluorescence (b) microscopy demonstrates that these proteins are restricted to the egg cortex. There is a recruitment of aetin

SCHATTEN ET AL. Fodrin during Fertilization
FIG. 6. Fodrin during sea urchin division. (a, b) Telophase. Fodrin is uniformally distributed at the cell surface at telophase and does not appear to accumulate at the presumptive cleavage furrow. (c, d) First cleavage. Fodrin remains at the cell surface and does not accumulate markedly at the cleavage furrow. (a, c): fodrin immunofluorescence; (b, d): corresponding DNA fluorescence of telophase chromosomes and blastomere nuclei, respectively. Bar = 10 pm.
a survey image in Fig. 8a. Figure 8b, at higher magni- fication, shows that the gold particles are bound pri- marily at the cell surface. The detergent-extraction pro- tocol and the fixation procedure required for immuno- electron microscopy do not preserve microfilaments or membrane profiles. Fodrin is not found within the cy- toplasm, even though immunogold electron microscopy with antitubulin demonstrates the ability of the gold
particles to penetrate deep into the cytoplasm in these extracted preparations. Immunoelectron microscopy with nonimmune serum does not preferentially associate with any recognizable structure.
Unfertilized mouse oocytes have fodrin concentrated at the cell surface along with cortical actin at the region adjacent to the meiotic spindle (Fig. 9b). The corre- sponding localization of actin with rhodaminyl-
and possibly fodrin in the fertilization cone (arrows). The incorporated sperm has decondensed to form the male pronucleus in c; the female pronucleus, at a different focal plane, is inset at the bottom of c. Triple-labeled eggs stained for actin (a), fodrin (h), and pronuclear DNA (c). Bar = 10 pm.
FIG. 5. Fertilization cones in detergent-extracted sea urchin eggs. Fodrin is retained at the egg cortex following extraction with the detergent Triton X-100. In a-c, it is apparent as bright wispy fibers covering the entire cortex as well as the much larger accumulations found at the fertilization cones (arrows). Bar = 10 Wm.

462 DEVELOPMENTAL BIOLOGY VOLUME 118, 1986
FIG. 7. Fodrin and actin colocalization in sheared surfaces. Actin (a) and fodrin (b) are detected in the same surface isolated during processing. While there is good correspondence between the fluorescence patterns, the coincidence is not precise. 10 @leytochalasin E results in decreased fluorescence of both actin (c) and fodrin (d), and their appearances become patchy. Bar = 10 pm.
phalloidin is depicted in Fig. 9a, and Hoechst DNA flu- orescence of the meiotic chromosomes aligned at the equator of the meiotic spindle is shown in Fig. 9c. Fodrin and actin are present as a cortical band at the site which is destined to constrict forming the second polar body.
During mouse fertilization, fodrin is found along the entire oocyte and second polar body surfaces (Figs. 9d- f). In Fig. 9e, at the pronucleate stage, fodrin is restricted to the cortex of the oocyte and of the second polar body, while actin in the same oocyte (Fig. 9d) is present at these regions and also within the cytoplasm around the apposed pronuclei (Fig. 9d).
At cleavage in mouse zygotes (Fig. 9g: actin; Fig. 9h: fodrin), actin and fodrin are colocalized with an apparent accumulation at the furrow, and in blastocysts fodrin delineates each cell border (Fig. 9i), as previously re- ported by Sobel and Alliegro (1985) using antispectrin immunofluorescence microscopy. Figure 9j is the DNA fluorescence image corresponding to the blastocyst shown in Fig. 9i.
DISCUSSION
In erythrocytes, spectrin interacts with actin oligo- mers to form a cortical gel which is associated with the

SCHATTEN ET AL. Fodrin during Fertilization 463
FIG. 8. Immunoelectron microscopy of fodrin labeling of a fertilized sea urchin egg surface at 50 min after insemination. Immunogold labeling of antifodrin antibodies is restricted to the egg cortex. (a) Survey image showing gold particles restricted to the cell periphery near demembranated microvilli. (b) A higher magnification image demonstrating that the gold particles are found at the cell surface. Bars = lrm.
plasma membrane (reviewed by Branton et ak, 1981). Spectrin-related proteins, such as fodrin, are concen- trated in the corresponding cortical regions in many other cells (Goodman et ah, 1981; Levine and Willard, 1981; Burridge et al, 1982; Reparsky et al., 1982; Cheney et al., 1983; Glenney and Glenney, 1983; Lazarides et aZ., 1984; Virtanen et al, 1984; Sobel and Alliegro, 1985; Sobel et aL, 1985). In this study, the location of fodrin-like proteins in gametes and their rearrangements during fertilization and early development are examined.
A protein which shares antigenic determinants with mammalian a-fodrin and has a similar molecular weight (approximately 240 kDa) is observed here to be a com- ponent of sea urchin eggs. Consistent with this obser- vation, Fishkind et al. (1985) have reported the isolation of a spectrin-like protein from sea urchin eggs. Previ- ously Kane (1980) described a high molecular weight protein that could be related to fodrin in homogenates of unfertilized sea urchin eggs induced to form gels. Til- ney (1976a,b) described a high molecular weight protein from sea urchin sperm, and speculated that such a pro- tein might be related to spectrin. The presence of a fod- rin-like polypeptide in sea urchins indicates that certain physical proteins (e.g., molecular weight) and antigenic determinants of these proteins have been conserved in organisms whose evolutionary pathways diverged over 550 million years ago.
In sea urchins, fodrin is found at the contact region between the gametes. We observed that fodrin-specific fluorescence is highly concentrated in the acrosomal tip of the sperm and is distributed through the egg cortex, very likely in association with the microvilli. The later accumulation of fodrin in the fertilization cone, which is particularly pronounced after detergent extraction in Figs. 5a-c, supports the interpretation that fodrin is found in or around the site of sperm-egg fusion and incorporation in sea urchins.
The mechanism by which the sea urchin sperm is in- corporated into the egg after membrane fusion is still obscure. Sperm incorporation is prevented by cytochal- asins (Longo, 1978, 1980; Schatten and Schatten, 1980, 1981) and latrunculin (Schatten et aZ., 1986), suggesting that actin polymers are important for this process. Al- though this apparent requirement for actin during in- corporation could be explained if the sperm were drawn into the egg by an actin-myosin interaction analogous to the sliding filament mechanism of muscle contraction, the polarity of the actin both in the sperm acrosomal process and in the egg microvilli is the reverse of that predicted for such a mechanism (Tilney and Jaffe, 1980). The current observations that fodrin-like proteins and actin are present during fusion and incorporation sug- gest the possibility that the formation of an actin-spec- trin gel at the site of sperm-egg membrane fusion could
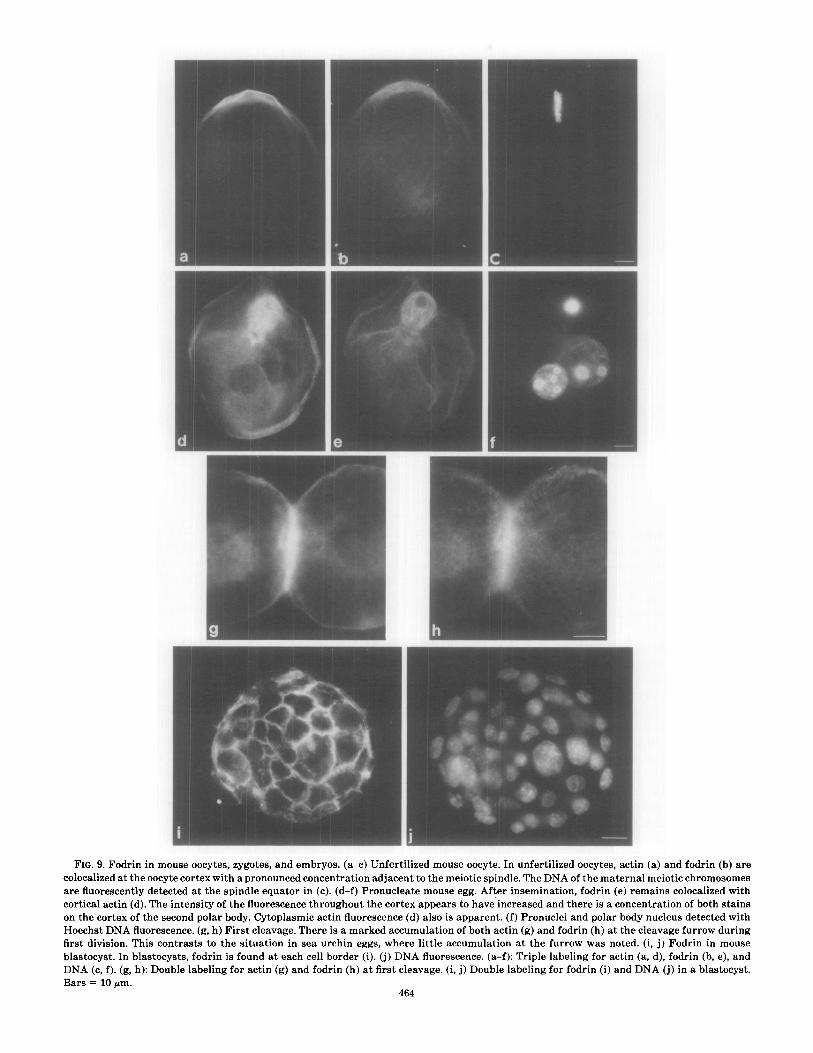
FIG. 9. Fod colocalized at are fluoresce1 cortical actin on the cortex Hoechst DNA first division. blastocyst. In DNA Cc, 0 k Bars = 10 pm
rin in mouse oocytes, zygotes, and embryos. (a-c) Unfertilized mouse oocyte. In unfertilized oocytes, actin (a) and fodrin the oocyte cortex with a pronounced concentration adjacent to the meiotic spindle. The DNA of the maternal meiotic : chrom itly detected at the spindle equator in (c). (d-f) Pronucleate mouse egg. After insemination, fodrin (e) remains cc blocalizS (d). The intensity of the fluorescence throughout the cortex appears to have increased and there is a concentration of botl of the second polar body. Cytoplasmic actin fluorescence (d) also is apparent. (f) Pronuclei and polar body nucleus detect
L fluorescence. (g, h) First cleavage. There is a marked accumulation of both actin (g) and fodrin (h) at the cleavage furrow This contrasts to the situation in sea urchin eggs, where little accumulation at the furrow was noted. (i, j) Fo drin in
, blastocysts, fodrin is found at each cell border (i). (j) DNA fluorescence. (a-f): Triple labeling for actin (a, d), foe bin (b, :, h): Double labeling for actin (g) and fodrin (h) at first cleavage. (i, j) Double labeling for fodrin (i) and DNA (j) i n a bla: I.
464
(b) are
ed with 1 stains ed with during mouse e), and
stocyst.

SCHAWEN ET AL. Fodrin during Fertilization 465
serve to anchor the sperm to the egg surface, perhaps via direct interactions between the sperm head or at- tached acrosomal microfilaments and the egg cortical gel. This anchoring function might account for the re- quirement for polymerized actin for successful sperm incorporation.
However, during mammalian fertilization, fodrin is found at regions seemingly uninvolved in sperm-egg fu- sion in both gametes. In mouse sperm the midpiece stains most intensely with antifodrin antibodies (data not shown), and in human sperm spectrin-like antigens have been detected at the acrosomal cap and at the principal piece (Virtanen et al., 1984).
In the mouse oocyte, fodrin fluorescence is brilliant at the cortical domain adjacent to the meiotic spindle (Figs. 9a-c), which is devoid of microvilli (Eager et al, 1975) and which cannot support sperm-oocyte mem- brane fusion (Nicosia et al, 1977). This suggests that there may be different specialized roles for the fodrin- related molecules in the unfertilized mouse oocyte, per- haps involved in maintaining the meiotic spindle in its correct position (Long0 and Chen, 1985), in eliminating the second polar body, or perhaps even in determining the axis for the first cleavage (Howlett and Bolton, 1985). However, the absence of fodrin staining of the mouse oocyte cortical regions which can support sperm fusion, coupled with the findings that microfilament inhibitors such as the cytochalasins (Longo, 19’78; Maro et al., 1984) and latrunculin (Schatten et ak, 1986) do not prevent mouse sperm incorporation, raises questions as to whether actin or spectrin-like proteins serve any similar role during mammalian sperm incorporation.
As the fertilized mouse egg continues to develop, an- tifodrin reactivity redistributes to become most concen- trated in the cleavage furrow, and later in the region of cell-cell contact in the blastocyst, as orginally reported by Sobel and Alliegro (1985). It is interesting that Sobel and Alliegro (1985), using an antibody against eryth- rocyte spectrin on cumulus- and zona-invested oocytes, did not detect antispectrin binding. The recent findings by Sobel et al. (1985) that a 240-kDa protein in mouse oocytes cross-reacts with antibodies against the P-chain of spectrin, but that antibody binding is not detected in unfertilized oocytes by immunocytochemical methods (S. Sobel and F. Longo, personal communication), provide additional indications that spectrin-like proteins may be differentially expressed, exposed, or more tightly bound to the cytomatrix during mouse development.
Although all of the regions of gametes, zygotes, and embryos which contained high levels of antifodrin reac- tivity also appeared to contain high concentrations of polymerized actin, the association of fodrin with actin polymers appears more selective. Most strikingly, the cytoplasmic shells of perinuclear actin which form
around the adjacent pronuclei in fertilized mouse oocytes (Maro et aZ., 1984) are not stained by antifodrin, indi- cating that the cytoplasmic functions performed by these actin filaments in the mouse require fodrin at lower con- centrations (if at all) compared to the functions per- formed by cortical actin filaments.
In summary, these findings of spectrin-like proteins in mouse and sea urchin sperm and eggs suggest roles for fodrin-actin arrays during the membrane events of fertilization and cleavage. In sea urchins, the fusigenic regions of both the sperm and the egg have fodrin and actin at the respective plasma membranes. During sperm incorporation, fodrin appears to accumulate around the sperm and is colocalized with actin in the fertilization cone; this actin-spectrin interaction during incorpora- tion might well anchor the sperm to the egg surface. In mice, fodrin may play a role in the formation of the second polar body, but might not be involved in sperm incorporation. During later development, fodrin is found at the entire zygote surface and, in the mouse, it con- centrates at the cleavage furrow during cytokinesis. These findings support the suggestions that fodrin, by its interactions with the plasma membrane and cortical actin, serves to stabilize cytoskeletal-membrane inter- actions during fertilization and early development.
We thank Dr. Sahina Sobel of the State University of New York at Buffalo for helpful suggestions and for sharing unpublished results, Dr. George Erdos and Dr. Henry Aldrich of the University of Florida for the generous donation of protein A-gold conjugate, Professor Theodor Wieland of the Max-Planck-Institut fiir Medizinische For- schung, Heidelberg, for the generous donation of rhodaminyl-phal- loidin, and Dr. Gerard Coffe of the University of Paris and of this institution for helpful discussions. The support of this research by the National Institutes of Health (M.W. and G.S.) and by the National Science Foundation (G.S.) is gratefully acknowledged.
REFERENCES
BALCZON, R., and SCHATTEN, G. (1983). Microtubule-containing deter- gent-extracted cytoskeletons in sea urchin eggs from fertilization through cell division. Cell Motil. 3,213-226.
BARAK, L. S., YOCUM, R. R., and WEBB, W. W. (1981). In wivo staining of cytoskeletal actin by autointernalization of nontoxic concentra- tions of nitrobenzoxadiazole-phallacidin. J. Cell Bill 89,368-372.
BRANTON, D., COHEN, C. M., and TYLER, J. (1981). Interaction of cyto- skeletal proteins on the human erythrocyte membrane. Cell 24,24- 32.
BURRIDGE, K., KELLY, T., and MANGEAT, P. (1982). Non-erythrocyte spectrins: Actin-membrane attachment proteins occurring in many cell types. J. Cell Biol 95,478-486.
CHENEY, R., HIROKAWA, N., LEVINE, J., and WILLARD, M. (1983). In- tracellular movement of fodrin. Cell MOM 3, 649-655.
CLARKE, G. N., and YANAGIMACHI, R. (1978). Actin in mammalian sperm heads. J. Exp. Zool. 205,125-132.
CLINE, C., and SCHATTEN, G. (1986). Microfilaments during sea urchin fertilization: Fluorescence detection with rhodaminyl phalloidin. Gamete Res. 14,277-291.

466 DEVELOPMENTAL BIOLOGY VOLUME 118, 1986
DEMEY, J. (1983). Colloidal gold probes in immunocytochemistry. In “Immunocytochemistry” (J. M. Polak and S. van Norden, eds.), pp. 82-112. Wright, Bristol.
EAGER, D., JOHNSON, M. H., and THURLEY, K. W. (1975). Ultrastructural studies on the surface membrane of the mouse egg. J. CeU Sci. 22, 345-353.
EDDY, E. M., and SHAPIRO, B. M. (1976). Changes in the topography of the sea urchin egg after fertilization. J. CeU Biol. 71.35-48.
ERDOS, G. W. and WHITAKER, D. (1983). Failure to detect immunocy- tochemically reactive endogenous lectin on the cell surface of Die- tyostelium diswideum. J. Cell Biol. 97,993-1000.
FISHKIND, D. J., BONNER, E. M., and BEGG, D. A. (1985). Isolation and characterization of sea urchin spectrin: A novel Ca++-sensitive actin crosslinker. J. Cell Biol. 101,285a.
GOODMAN, S. R., ZAGON, I. S., and KULIKOWSKI, R. R. (1981). Identifi- cation of a spectrin-like protein in nonerythroid cells. Proc N&L Acad Sci. USA 73,7570-7574.
GLENNEY, J. R., JR., and GLENNEY, P. (1983). Fodrin is the general spectrin-like protein found in most cells whereas spectrin and the TW protein have a restricted distribution. Cell 34,503-512.
HIROKAWA, N., CHENEY, R. E., and WILLARD, M. (1983). Localization of a protein of the fodrin-spectrin TW2601240 family in the mouse intestinal brush border. Cell 32,953-965.
HOWLER, S. K., and BOLTON, V. N. (1985). Sequence and regulation of morphological and molecular events during the first cell cycle of mouse embryogenesis. J. Embyol Exp. Morphol. 87,175-206.
KANE, R. E. (1980). Induction of either contractile or structural actin- based gels in sea urchin egg cytoplasmic extracts. J. Cell Biol. 86, 803-809.
LAEMMLI, U. K. (1970). Cleavage of structural proteins during the as- sembly of the head of bacteriophage T4. Nature (London) 227,680- 685.
LAZARIDES, E., NELSON, W. J., and KASAMATSU, T. (1984). Segregation of two spectrin forms in the chicken optic system. Cell 36,269-278.
LEVINE, J., and WILLARD, M. (1981). Fodrin: Axonally transported polypeptides associated with the internal periphery of many cells. J. Cell Biol. 90,631-643.
LEVINE, J., and WILLARD, M. (1983). Redistribution of fodrin accom- panying capping of cell surface molecules. Proc. Natl. Acad. Sci. USA 80,191-195.
LONGO, F. J. (1978). Effects of cytochalasin B on sperm-egg interactions. Dev. Biol. 67.249-364.
LONGO, F. J. (1980). Organization of microfilaments in sea urchin (Ar- bucia punctti) eggs at fertilization: Effects of cytochalasin B. Dev. Biol. 74,422-431.
LONGO, F. J., and CHEN, D.-Y. (1985). Development of cortical polarity in mouse eggs: Involvement of the meiotic apparatus. Dew. Bid 107, 382-394.
MARO, B., JOHNSON, M., PICKERING, S. J., and FLACH, G. (1984). Changes in actin distribution during fertilization of the mouse egg. J. Embrgol Exp. Mwphol. 81,211-237.
MAZIA, D., SCHATTEN, G., and SALE, W. (1975). Adhesion of cells to surfaces coated with polylysine. J. Cell Biol 66,198200.
NICOSIA, S. V., WOLF, D. P., and INOUE, M. (1977). Cortical granule distribution and cell surface characteristics in mouse eggs. Dev. Biol 57,56-74.
REPARSKY, E. A., GRANGER, B. L., and LAZARIDES, E. (1982). Widespread occurrence of avian spectrin in nonerythroid cells. Cell 29,821~833.
SCHATTEN, G., and SCHA~EN, H. (1981). Effects of motility inhibitors during sea urchin fertilization. Exp. Cell Res. 135,311-330.
SCHATTEN, G., SCHA~EN, H., SPECTOR, I., CLINE, C., PAWELETZ, N., SIMERLY, C., and PETZELT, C. (1986). Latrunculin inhibits the mi- crofilament-mediated processes during fertilization, cleavage and early development in sea urchins and mice. Exp. Cell Res., in press.
SCHATTEN, G., SIMERLY, C., and SCHATTEN, H. (1985). Microtubule con- figurations during mouse fertilization, mitosis and embryogenesis. Proc. Natl. Acad. Sci. USA 82,4152-4156.
SCHATTEN, H., and SCHATTEN, G. (1980). Surface activity at the egg plasma membrane during sperm incorporation and its cytochalasin B sensitivity. Dev. Biol. 78,435-449.
SCHROEDER, T. (1981). Interrelations between the cell surface and the cytoskeleton in cleaving sea urchin eggs. In “Cytoskeletal Elements and Plasma Membrane Organization” (G. Poste and G. L. Nicolson, eds.), pp. 169-216. Elsevier/North Holland, Amsterdam.
SOBEL, J. S., and ALLIEGRO, M. A. (1985). Changes in the distribution of a spectrin-like protein during development of the preimplantation mouse embryo. J. Cell Biol. 100,333-336.
SOBEL, J. S., OPAS, M., and KALNINS, V. I. (1985). Spectrin and cell contacts in the early mouse embryo. J. Cell Biol. 101,1766a.
TILNEY, L. G. (1976a). Polymerization of Actin. II. How nonfilamentous actin becomes non-randomly distributed in sperm: Evidence for the association of this actin with membranes. J. Cell Biol. 69,51-72.
TILNEY, L. G. (197613). Polymerization of Actin. III. Aggregates of non- fllamentous actin and its associated proteins: A storage form of actin. .I. Cell Biol. 69,73-89.
TILNEY, L. G. (1978). Polymerization of actin. V. A new organelle, the actomere, that initiates the assembly of actin filaments in Thyone sperm. J. Cell BioL 69,53-72.
TILNEY, L. G., and JAFFE, L. R. (1980). Actin, microvilli and the fer- tilization cone of sea urchin eggs. J. Cell Bill. 67,771-782.
VACQUIER, V. D. (1981). Dynamic changes of the egg cortex. Dev. Biol. 84, l-26.
VIRTANEN, I., BADLEY, R. A., PAASIVUO, R., and LEHTO, V.-P. (1984). Distinct cytoskeletal domains revealed in sperm cells. J. CeU Bid 99,1083-1091.
WHIT~INGHAM, D. G. (1968). Fertilization of mouse eggs in vitro. Nature (London) 220,592-593.
WIELAND, TH., and FAULSTICH, H. (1977). Amatoxins, phallotoxins, phallolysin, and antamanide. CRC 0%. Rev. Biochem 5,185-260.
YANAGIMACHI, R., and NODA, Y. D. (1976). Ultrastructural changes in the hamster sperm head during fertilization. J. UZtrastruct. Res. 31, 465-485.