
Thermal sensitivity during brood care in workers of two Camponotus ant species: Circadian variation...
11
Pergamon 0022-1910(95)00019-4 J. Insect Physiol. Vol. 41, No. 8, 659-669, pp. 1995 Copyright 0 1995 Elsevier Science Ltd Printed in Great Britain. All rights reserved 0022-1910/95 $9.50 + 0.00 Thermal Sensitivity During Brood Care in Workers of Two Camponotus Ant Species: Circadian Variation and Its Ecological Correlates FLAVIO ROCES*t$ JO?& A. Nfir;TEZ* Received 7 September 1994; revised 9 January 1995 Thermal sensitivity during brood care was analysed in two Neotropical Cumponotus ant species, related phylogenetically but disparate in ecology. Cumponotus mus is dominant in temperate climates, whereas C. rufipes is dominant in subtropical climates. Sensitivity was assessed by first allowing workers to manifest their preference along a thermal gradient, and thereafter increasing the temperature of the nest floor continuously at a rate of 0.2’C/min. The removal of brood by workers was recorded as an indicative response of upper thermal tolerance. Along a thermal gradient, while C. mus nurse workers exhibited a bimodal circadian rhythm of temperature preference to locate the brood, C. rufipes nurses selected an invariant temperature throughout the day. Irrespective of their thermal preferences, sensitivity to temperature changes in both species showed a conspicuous circadian rhythm. Its endogenous nature was supported by a free-run under constant darkness. In C. mus, instantaneous resetting by the light:dark cycle could also be demonstrated, and experiments suggest the involvement of one oscillator controlling thermal responses. At the middle of the light period when extreme environmental temperatures are expected, C. mus nurses exhibited the highest sensitivity (brood removal after an increase of 0.2’(Z), while C. rufipes showed the lowest (removal after an increase of 6.7”C). For both species, the time-course of thermal sensitivity during brood care suggests an adaptation to the wide and slight daily fluctuations undergone by temperature in temperate and tropical climates, respectively. Ant Cumponotus Brood Circadian rhythm Thermal sensitivity INTRODUCTION Environmental temperature plays a decisive role in the metabolism and energy balance of ant societies, because most of them are essentially ectothermic (reviewed in Heinrich, 1993). Temperature primarily controls worker activity by inhibiting or reducing foraging, as well as the speed of brood development, parameters that directly determine the growth rate of a colony (e.g. Porter, 1988; Porter and Tschinkel, 1993). However, excessive temperatures negatively affect colony growth by reducing both brood survival and worker longevity (Calabi and Porter, 1989; Races and Ntifiez, 1989). As a result, one could expect the existence of important adaptations of ant *Departamento de Biologia, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Ciudad Universitaria, 1428 Buenos Aires, Argentina. tTo whom all correspondence should be addressed. JPresent address: Zoologie II. Biozentrum der Universitiit, Am Hubland, 97074 Wiirzburg, Germany. colonies to cope with seasonal variations in environmen- tal temperature, as well as with the naturally occurring daily temperature fluctuations (Steiner, 1929). Long-term thermal regulatory responses of ants have been described at both the individual and colony level. They involve, for instance, the selection of a suitable place to locate the nest by founding queens (Brian, 1952), and the construction of stable nest mounds which function as solar-collecting devices (Steiner, 1929; Scherba, 1962). Short-term responses for temperature control in ants, on the other hand, involve behavioural tracking of preferred temperatures inside the nest by workers, and transport of brood to the selected values (e.g. Ceusters, 1977; Brian, 1973; Races and Ntiiiez, 1989; Porter and Tschinkel, 1993). Thermal preference of ant workers, both for their own location and for that of the brood, has been studied in the laboratory after the establishment of a thermal gradient along the nest. It has been observed that workers tending brood usually have thermal preferences very similar to 659
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Thermal sensitivity during brood care in workers of two Camponotus ant species: Circadian variation...

Pergamon 0022-1910(95)00019-4
J. Insect Physiol. Vol. 41, No. 8, 659-669, pp. 1995 Copyright 0 1995 Elsevier Science Ltd
Printed in Great Britain. All rights reserved 0022-1910/95 $9.50 + 0.00
Thermal Sensitivity During Brood Care in Workers of Two Camponotus Ant Species: Circadian Variation and Its Ecological Correlates FLAVIO ROCES*t$ JO?& A. Nfir;TEZ*
Received 7 September 1994; revised 9 January 1995
Thermal sensitivity during brood care was analysed in two Neotropical Cumponotus ant species, related phylogenetically but disparate in ecology. Cumponotus mus is dominant in temperate climates, whereas C. rufipes is dominant in subtropical climates. Sensitivity was assessed by first allowing workers to manifest their preference along a thermal gradient, and thereafter increasing the temperature of the nest floor continuously at a rate of 0.2’C/min. The removal of brood by workers was recorded as an indicative response of upper thermal tolerance. Along a thermal gradient, while C. mus nurse workers exhibited a bimodal circadian rhythm of temperature preference to locate the brood, C. rufipes nurses selected an invariant temperature throughout the day. Irrespective of their thermal preferences, sensitivity to temperature changes in both species showed a conspicuous circadian rhythm. Its endogenous nature was supported by a free-run under constant darkness. In C. mus, instantaneous resetting by the light:dark cycle could also be demonstrated, and experiments suggest the involvement of one oscillator controlling thermal responses. At the middle of the light period when extreme environmental temperatures are expected, C. mus nurses exhibited the highest sensitivity (brood removal after an increase of 0.2’(Z), while C. rufipes showed the lowest (removal after an increase of 6.7”C). For both species, the time-course of thermal sensitivity during brood care suggests an adaptation to the wide and slight daily fluctuations undergone by temperature in temperate and tropical climates, respectively.
Ant Cumponotus Brood Circadian rhythm Thermal sensitivity
INTRODUCTION
Environmental temperature plays a decisive role in the metabolism and energy balance of ant societies, because most of them are essentially ectothermic (reviewed in Heinrich, 1993). Temperature primarily controls worker activity by inhibiting or reducing foraging, as well as the speed of brood development, parameters that directly determine the growth rate of a colony (e.g. Porter, 1988; Porter and Tschinkel, 1993). However, excessive temperatures negatively affect colony growth by reducing both brood survival and worker longevity (Calabi and Porter, 1989; Races and Ntifiez, 1989). As a result, one could expect the existence of important adaptations of ant
*Departamento de Biologia, Facultad de Ciencias Exactas y Naturales,
Universidad de Buenos Aires, Ciudad Universitaria, 1428 Buenos
Aires, Argentina. tTo whom all correspondence should be addressed.
JPresent address: Zoologie II. Biozentrum der Universitiit, Am Hubland, 97074 Wiirzburg, Germany.
colonies to cope with seasonal variations in environmen- tal temperature, as well as with the naturally occurring daily temperature fluctuations (Steiner, 1929).
Long-term thermal regulatory responses of ants have been described at both the individual and colony level. They involve, for instance, the selection of a suitable place to locate the nest by founding queens (Brian, 1952), and the construction of stable nest mounds which function as solar-collecting devices (Steiner, 1929; Scherba, 1962). Short-term responses for temperature control in ants, on the other hand, involve behavioural tracking of preferred temperatures inside the nest by workers, and transport of brood to the selected values (e.g. Ceusters, 1977; Brian, 1973; Races and Ntiiiez, 1989; Porter and Tschinkel, 1993).
Thermal preference of ant workers, both for their own location and for that of the brood, has been studied in the laboratory after the establishment of a thermal gradient along the nest. It has been observed that workers tending brood usually have thermal preferences very similar to
659

660 FLAW0 ROCES and JOSUI? A. Nk!iEZ
those chosen for the brood itself, and that they are more selective than workers without brood (Kay, 1978). Workers tending brood, in addition, exhibit a distinct thermal preference for different brood developmental stages (Ceusters, 1977; Races and Nuiiez, 1989). These preferences were reported to be stable throughout the day, with no occurrence of further brood translocations under stable thermal conditions. Even along a stable thermal gradient, however, long-term records showed that nurse workers of the ant C. mus exhibit a daily rhythmicity in their temperature preference for the location of brood (Protomastro, 1973; Races and Nuiiez, 1989). Under controlled photoregimes, they selected different temperatures twice a day to translocate the brood: 30.8”C at the middle of the light period, and 27.5”C 8 h later, a thermal behaviour that afforded maximal production of adults (Races and Nuiiez, 1989).
It is worth mentioning that the conditions offered by a thermal gradient, i.e. stable temperatures at any point, strongly contrast with the situation observed in natural nests, where temperature undergoes a daily cycle (e.g. Kato, 1939). Although rhythmically fluctuating tempera- tures are known to affect both development and reproduction in insects (e.g. Messenger, 1964; Ratte, 1985) few studies have addressed the effects of oscillating temperatures on brood development and colony growth in ant societies (Buschinger, 1973; Elmes and Wardlaw, 1983; Races and Nuiiez, 1989). In addition, studies on the workers’ thermal behaviour in relation to the naturally occurring temperature cycle are lacking, because ant thermal preferences have usually been recorded along stable temperature gradients. Do workers inside the nest track the preferred temperature for the brood continu- ously throughout the day as it changes? Or are there critical time “windows” within which workers actively search for their preferred temperatures? If so, how often are workers monitoring the temperature of their nest environments? And how large are the thermal changes tolerated by nurses throughout the daily cycle, before they decide to remove the brood and search for a more suitable place?
In the present study the question of how ant workers cope with changing temperatures along the day in order to regulate brood temperature was addressed. For this, thermal sensitivity during brood care was analysed comparatively in nurse workers of two Neotropical ant species which are close in phylogeny but disparate in ecology. Even though colonies of both species nest in rotting wood or inside holes in twigs, they are dominant in areas which undergo different daily temperature regimes: C. mus in a temperate climate, and C. rujpes in subtropical and tropical climates (Kusnezov, 1951, 1963). Thermal sensitivity was assessed as the threshold of thermal tolerance of workers actually involved in brood tending. Experiments were performed during all phases of the daily cycle as well as under constant photic conditions, in order to account for possible circadian variations, and to relate them to the naturally occurring temperature cycle experienced by ant colonies in their habitats.
Evidence is presented on both the existence of endogenous, self-sustained circadian rhythms of thermal sensitivity in nurse workers of both species, and the role of the light-onset in synchronizing the rhythm.
MATERIALS AND METHODS
Ant species
Two colonies of C. mus and one of C. rujipes were used during this study. Camponotus mus founding queens were captured after the nuptial flight near Buenos Aires, Argentina, and the queen of C. rujipes in Rio Claro, Sao Paulo State, Brazil. They were individually reared in artificial glass nests at 25 &- 1°C and under a 1ight:dark (LD) cycle of 12:12 h (light-on: 8 a.m.; 100 lx). The colonies were fed ad libitum with 50% w/w sucrose solution and freshly-killed Drosophila flies. Voucher specimens of both species were deposited in the Laboratory of Insect Physiology of the University of Buenos Aires (UBA), Argentina.
Thermal sensitivity of C. mus nurse workers
The rationale of the experimental procedure was to allow workers in a laboratory nest to locate the brood at their preferred temperature. Afterwards, a continuous temperature increase (at a rate of 0.2”C/min) was induced experimentally at the brood place, and the maximal thermal increase tolerated by nurses was recorded, using the removal of a brood item as indicative response. For the measurements of thermal sensitivity of C. mus, each colony (consisting of queen, 15-20 workers and brood at the time of the experiments) was located in an artificial plastic nest (11 x 7 x 2.5 cm), and provided with a LD cycle of 12:12 h (light-on: 8 a.m.; 100 lx). A vial with water, attached to a lateral wall of the nest, kept humidity roughly constant at 80-90%. The nest floor consisted of two adjacent aluminium plates, each plate covering one-half of the nest floor. Under these plates, two independent heat sources allowed a constant temperature to be reached on each one. The heat sources consisted of heat wires connected to a electronic circuit which enabled control of temperature to the nearest 0. 1°C and also small temperature increments. To avoid heat conduction between both plates, a narrow plastic strip was placed in between. Temperature measurements were performed on the floor with two calibrated thermistor-thermometers (accuracy O.l”C; time constant 1.6 s), which remained inside the nest throughout the experiments. Even though we recorded floor temperatures, it is likely that the relevant temperatures for the ants might be the air temperature 2-3 mm above the floor. Measurements of the vertical thermogradient inside the nest showed a slight decrease with height, averaging 0.3”C 3 mm above the floor.
Since it is known that C. mus nurse workers spontaneously prefer two different temperatures for brood location along the day, 27.5 and 30.8”C, which are selected regularly in relation to the 1ight:dark cycle (Races

THERMAL SENSITIVITY IN ANTS 661
and Nuiiez, 1989) one-half of the nest floor (i.e. one aluminium plate) was maintained at 27.5”C, and the other one at 30.8”C. Workers could move freely inside the nest and select at any time one of the two available temperatures to locate the brood. These stable thermal conditions were imposed during a week. After this period, the upper temperature limit to which nurse workers subject the brood, i.e. their threshold of thermal tolerance during brood care, was assessed as follows: the temperature of one aluminium plate, where the brood was actually located, was increased at a rate of 0,2”C/min. Nurse workers, which usually remain inactive over the brood, responded to rising temperature by picking-up brood items and walking loaded around the brood pile, searching for a more suitable place. The temperature at which the first nurse worker began to evacuate a brood item (usually a larvae or cocoon) was recorded as the upper temperature tolerated. This criterion was chosen instead of, for instance, the removal of 50% of the brood, because the latter depends on both the number of nurses and the total amount of brood, which varied throughout the experiments. The assay was then concluded and the temperature of the plate returned to its original value. Assays were performed throughout the day, during both light and dark periods, and began from either 27.5 or 30.8”C, depending on the actual location of brood. They were also carried out under constant darkness, to account for possible endogenously-driven variations. During darkness, observations were performed by using red light (30 lx). Only 3 or 4 assays were performed per day, with an interval of 2-3 h in between, so that colony disturbances due to frequent changes in nest temperature were avoided. A total of 70 assays were carried out. Afterwards, daily threshold curves of thermal sensitivity were obtained by fitting Fourier curves to the empirical data. The Fourier interpolation uses a finite series of cosines and sines to fit the data, in the following form:
f(X) = a0 + C[a,*cos(27rn/L*X) + b,*sin(27rn/L*X)]
where a and b are constants, and [O,L] is the interval in which the function is periodic, i.e. L = 24 h. The number n of sines and cosines to achieve the best fit was determined by observing the percentage of variation explained by the curve (v2), using the program Mathematics (Wolfram, 1991).
Throughout the experimental period, the temporal occurrence of the two spontaneous brood-trans- location events inside the nest was photographically recorded (details in Races and Nuiiez, 1989). Each spontaneous brood-translocation event is manifested as a narrow burst of nurses’ carrying activity, which lasts usually from 5 to 15 min depending on the amount of brood to be moved. The temporal occurrence of each brood-translocation event was therefore defined objec- tively as the point when the onset of brood transport occurred.
Temperature preference and sensitivity of C. rujipes nurse workers
Prior to the measurement of the upper temperature limit for brood care in C. ruJipes nurses, it was necessary to record their thermal preference for the brood, in order to establish the initial conditions for the sensitivity experiments. For this, a temperature gradient was established along the glass nest (40x10~2 cm) where the colony was located (queen, 23 workers and brood). It was achieved by placing a heater with a parabolic screen near one nest-end, and maintaining the other at room temperature. This resulted in a continuous thermal gradient between 40 f 1 and 20 ) 2°C (see details in Races and Nuiiez, 1989). Temperature records were taken on the nest floor with a thermistor-thermometer, which was introduced through equidistant holes (2 cm apart) existing in the nest cover. The colony was provided with a LD cycle of 12: 12 h, and maintained under these conditions 2 weeks prior to recording.
As reported for C. mus (Races and Ntiiiez, 1989) C. rufipes nurse workers concentrated the brood in piles at a given temperature along the gradient. Since the distribution of temperatures was previously evaluated at any point along the nest gradient, nurses’ temperature preference for the brood could be directly derived from the location of the brood along it. Brood location was recorded with a photo-camera located 50 cm over the nest, which took pictures continuously every 10 min throughout 1 week.
After recording thermal preference of nurse workers for the brood, the upper temperature limit at which nurses subject the brood was experimentally determined following the procedure described above for C. mus. This allowed further interspecific comparisons. A total of 40 assays were performed, no more than 2 or 3 each day, and the daily threshold curve of thermal sensitivity was fitted as described for C. mus.
RESULTS
Thermal sensitivity in C. mus
In the artificial nest with two different temperatures (27.5 and 30.8”C), a total of 4-5 nurse workers were observed to translocate the complete brood spon- taneously from one-half of the nest to the other twice a day, following the pattern of thermopreferences described in the Introduction (see also Races and Nufiez, 1989). Measurements of thermal tolerance were initiated from these two temperatures, and the pooled empirical data in relation to the 1ight:dark cycle are depicted in Fig. 1. The figure also presents the periodic daily curve fitted by Fourier interpolation, as well as the confidence intervals of the mean. It was not possible to fit a single periodic curve to the complete data set because of the “jumps” of the curve at 14:00 and 22:00 h, which correspond to the occurrence of the spontaneous brood-translocation events (Fig. 1, arrows). Instead, two different Fourier

662 FLAW0 ROCES and JOSUB A. NofiEZ
interpolations were performed, from 22:00 to 14:00 h, and from 14:00 to 22:00 h, using a periodicity [O,L] of 24 h for each one. These two intervals of the curve were objectively delimited by the spontaneous brood translocations, and characterized by a different initial temperature during the measurements (Fig. 1, dashed horizontal lines). In order to visualize uninterruptedly the two intervals of the curve, the standard plot of circadian times, i.e. 0:OO h coinciding with light onset, was not used.
The threshold of thermal tolerance showed a strong dependence on the time of the day. It reached two minimal values daily, which corresponded to the times at which the two daily spontaneous brood-translocation events regularly occur (Races and NGez, 1989): the first minimum at the middle of the light period, and the second one 8 h later, 2 h after the onset of the dark period (at 22:00 h, Fig. 1). The photophase minimum represents the highest thermal sensitivity of nurses during the daily cycle: shortly after 14:00 h, nurses began to remove the brood as soon as the experimentally induced thermal increase reached 0.1-0.2”C above the self-selected temperature of 30.8”C (Fig. 1, values obtained from the fitted curve). From 14:00 h onwards, the threshold showed a sharp increase up to 3.7”C above the preferred temperature (threshold at 34.5”(Z), and declined thereafter prior to the second spontaneous brood-transport event at 22:00 h.
There it reached 1.1 “C above the preferred value. After the spontaneous brood translocation at 22:00 h, nurses were observed to be highly sensitive to the imposed thermal changes: at 22:30 h, the brood was removed as soon as nest temperature reached 0.8”C above the self-selected value (27.5”(Z). From this time onwards, the thermal tolerance increased up to 5.7”C above the preferred temperature (threshold at 33.2”C), and decreased again prior to the spontaneous brood-transport event, again reaching an absolute value of c. 31.2”C. Thus, the thermal change tolerated, but not the absolute temperature, was strongly dependent on the spontaneous brood-translocation event: while the absolute maximal temperature tolerated near 14:00 h was approximately 3 l”C, nurses tolerated an increase of 3.7”C before, but one of 0.2”C after the spontaneous brood translocation (Fig. 1).
Is the observed daily time-course of the thermal-toler- ance threshold of endogenous nature, i.e. self-sustained? Or is it driven directly by external cues such as the dark-light transitions? If this rhythmicity is based on endogenously generated oscillations, it will persist in the absence of any identifiable periodical external cue. To answer this question, the upper temperature limit for brood location was recorded as described above along 6 days, but the colony was provided with constant
I * - * * * 9. ’ * l * ’ * ’ * I*’ - 1. ‘. *
22 0 2 4 6 6 10 12 14 16 16 20 22
Local time
FIGURE 1. Threshold of thermal tolerance during brood care in Cumponotus mus nurse workers over a day. Each circle represents an independent measurement of the temperature tolerated by nurses prior to brood removal, recorded
throughout the day. Data from 70 independent measurements obtained in two colonies were pooled. Heavy line: threshold-curve fit obtained by Fourier interpolation. For the interval between 22:00 and 14:00 h, the fit is given
by: Y = 30.688 - 0.88*cos(O.262*X) + 2.316*sin(0.262*X) + 0.922*sin(0.524*X), where Y is tolerated temperature and X is
local time (n = 41; rL = 0.88; F = 98.2; P < 0.001). For the interval between 14:OO and 22:00 h, the fit is given by: Y = 28.054 + 0.507*cos(O.262*X) - 6.424*sin(0.262*X) - 0.036*sin(0.524*X)(n = 29; r* = 0.86; F = 51.4; P -e 0.001). Thin
lines indicate the 95%-confidence intervals, calculated as mean k 2SD, SD = 0.484 for the interval between 22:00 and 14:OO h,
and SD = 0.485 between 14:OO and 22:00 h. Dashed lines specify the two different temperatures (mean values) spontaneously selected by nurses to locate the entire brood, from which the experiments to assess thermal sensitivity were started. Arrows mark
the temporal occurrence of these two spontaneous brood translocations. The 1ight:dark cycle (LD 12:12 h) is indicated below the abscissae.

THERMAL SENSITIVITY IN ANTS 663
27 ’ ,
0 12 24 36 48 60 72 94
I Light-on , I I
84 96 108 120 132 144 166 168
FIGURE 2. Evidence for the endogenous, self-sustaining nature of the threshold of thermal tolerance in Cump~n~cu~ MUS nurse
workers. Measurements (solid circles) were performed under constant darkness over 5 days, and plotted together with the
circadian periodic curve (and 95%-confidence intervals) fitted to the measurements performed under a LD 12:12 h (Fig. l),
assuming a free-running period of 24 h. The abscissae show the photoregime used during the experimental period, and the regular
daily occurrence of the two spontaneous brood translocations: open squares for the onset of the brood translocation to 30.8”C and solid squares for that to 27.5”C (see Fig. 1). Dashed curve: hypothetical threshold-curve of thermal tolerance, predicted after
an instantaneous resetting by a 1Zadvanced 1ight:dark regime of 12:12 h, 134 h after the beginning of the experiment.
darkness, and the 1ight:dark cycle was restored thereafter with a 12-h phase-shift. Measurements were then compared with values predicted by the curve of thermal tolerance fitted to the data measured under LD 12:12 h (Fig. l), assuming that it was self-sustained, with a free-running period of 24 h.
The empirical data on tolerated temperature, as well as the hypothetical free-running thermal tolerance, are presented in Fig. 2. Most of the measured values lay in the 95% confidence interval of the hypothetical curve, which strongly supports the assumption of an oscillating, self-sustained thermal tolerance. To allow a comparison of free-running thermal sensitivity and spontaneous thermal preference, the temporal occurrence of the spontaneous brood-translocation events is also presented in Fig. 2 (open and closed squares below the abscissae).
As expected, the spontaneous brood translocations correspond with the two minima in the thermal-tolerance curve (see, for example, the record between 0 and 24 h in Fig. 2). The regular occurrence of two daily brood translocations indicates that nurses’ temperature prefer- ence for the brood was driven by an endogenous oscillator. Both translocation events showed a similar mean free-running period of rDD z 23.4 h, as estimated by eye-fitting a line through daily onsets of translocation activity. If this free-running period is used in Fig. 2 for the hypothetical curve instead of a hypothetical one of 24 h, however, no substantial change in the observed fit arises.
It should be noted that a larger number of measurements would be necessary to fit a curve to the empirical data obtained under constant darkness (Fig. 2). The number of measurements was kept low (usually 3 per

664 FLAW0 ROCES and JOSUfi A. NUfiEZ
day) in order to avoid frequent changes of nest temperature, which could affect the expression of nurses’ responses. Taking into account this restriction, the correspondence between measured and expected values was assessed by comparing them with the corresponding confidence interval of the hypothetical curve. Differences between measured and expected values along the experimental period can be visualized in Fig. 3. Results indicate that the differences between observed and expected values were slight during the first 2 days under constant darkness, and included in the 95%-confi- dence interval of the curve, becoming larger as time advanced.
Upon restoring the LD 12:12 h cycle with a 12 h-phase shift (134 h after the beginning of the experiment; see Fig. 2), two lines of evidence suggest that this rhythmicity was set in phase by the alternation of light and darkness. First, measurements of thermal tolerance approximate the expected values if the rhythm is assumed to be instantaneously re-entrained by the first onset of the light period (Fig. 2, dashed curve). This pattern can also be visualized by comparing the differences between measurements and expectations from the re-entrained curve (Fig. 3, open circles after light-on), or expectations from the free-running curve (Fig. 3, closed circles after light-on). Second, no brood translocations were observed during the first restored light phase (Figs 2 and 3, squares below the abscissae after light-on). Note that during the first light phase, the free-running rhythm (continuous
5
o^ 4
0 3
G $ * $1
2 w 0
$ -’
L -2 $
5:
-5
curve) would predict the occurrence of a spontaneous brood translocation to 27.5”C 2 h after the onset of the subjective dark period (Fig. 2, see minimal value of the continuous, free-running curve during the restored light period, 142 h after beginning of the experiments). The re-entrained rhythm (dashed curve), conversely, would predict the occurrence of a brood translocation to 30.8”C at the middle of the light period. Records indicated that no brood translocations at all were observed during the first restored light period, i.e. the brood remained at 30.8”C, where they had been located during the preceding dark period. The acceptance of 30.8”C by nurses at this time, and their lack of preference for 27.5”C, are consistent with the idea of an instantaneous phase-setting by the onset of the light period. This conclusion is further supported by both the subsequent brood-translocation event to 27.5”C, and the measurements of thermal tolerance during the first restored 12 h dark period (Fig. 2, after 146 h), which corresponded with the re-entrained rhythm (dashed curve).
Temperature preference and sensitivity in C. rujipes
Contrary to C. mus, C. rufipes nurse workers showed no rhythmicity in their temperature preference for brood location along the thermal gradient. Long-term photo- graphic records indicated that they selected a stable value of 25.0 + 0.4”C (mean + SE, n = 52) for the brood, irrespective of the time of day.
, Light-on
i . I .
. I
0 12 24 36 46 60 72 94 96 106 120 132 144 156 166
Time (hs)
FIGURE 3. Correspondence between measurements ofthe upper thermal tolerance under constant darkness, and the hypothetical
temperatures under the assumption of a self-sustained circadian thermal tolerance in C. mus nurse workers. Heavy line (centred
at zero) specifies the ideal fit of measurements to the expected curve. Dashed lines mark the 95%-confidence intervals, as in Fig. 1.
Solid circles correspond to the differences between measurements and a free-running (ODD = 24 h) hypothetical curve, whereas open circles specify those between measurements and a hypothetical curve, instantaneously reset by a 12-h advanced phase-shift
(after light-on). The abscissae show the photoregime used during the experimental period, and the regular daily occurrence of the two spontaneous brood translocations: open squares for the onset of the brood translocation to 30.8”C and solid squares
for that to 27.5”C.

THERMAL SENSITIVITY IN ANTS 665
0 2 4 6 6 10 12 14
Local time
16 16 20 22 24
FIGURE 4. Threshold of thermal tolerance during brood care in Camponotus rufpes nurse workers, as recorded for C. rnus
(Fig. 1). Data from one colony were pooled (n = 40). Heavy line: threshold-curve fit obtained after Fourier interpolation, given
by: Y = 28.403 - 2.194*cos(O.262*X) + 0.767*cos(O.524*X) - 0.961*sin(0.262*X) + 0.631*sin(0.524*X), where Y is tolerated
temperature and X is local time (n = 40; r* = 0.85; F = 49.3; P < 0.001). Thin lines indicate the 95%-confidence intervals,
calculated as mean + 2SD, SD = 0.761. Dashed lines indicate the stable temperature selected by nurses to locate the entire brood
throughout the day (mean = 25°C). The 1ight:dark cycle (LD 12:12 h) is indicated below the abscissae.
Does this unchanging preference indicate that nurse workers will continuously track 25.O”C inside the nest if environmental temperature varies during the day, as it occurs in field nests? To answer this question, the upper temperature tolerated by nurses prior to removal of brood was measured in this species as described for C. mus. Due to the lack of variable thermopreference, all assays were initiated by increasing the temperature from 25.O”C, i.e. both plates of the nest floor were maintained at this value prior to the experiments.
Even though C. rufipes nurse workers showed a stable thermal preference throughout the daily cycle, the upper temperature tolerated until brood removal showed a marked dependence on the time of day (Fig. 4). At the middle of the light period, when environmental temperature normally peaks, it reached the maximal values. Nurse workers tolerated a thermal increase as large as 6.7-6.8”C above the mean preferred temperature for brood location, i.e. 25°C (Fig. 4, value obtained from the fitted curve). During the dark period, conversely, nurses proved to be highly sensitive to thermal increases, starting to remove the brood as soon as temperature averaged 1.9”C above their preferred temperature (Fig. 4).
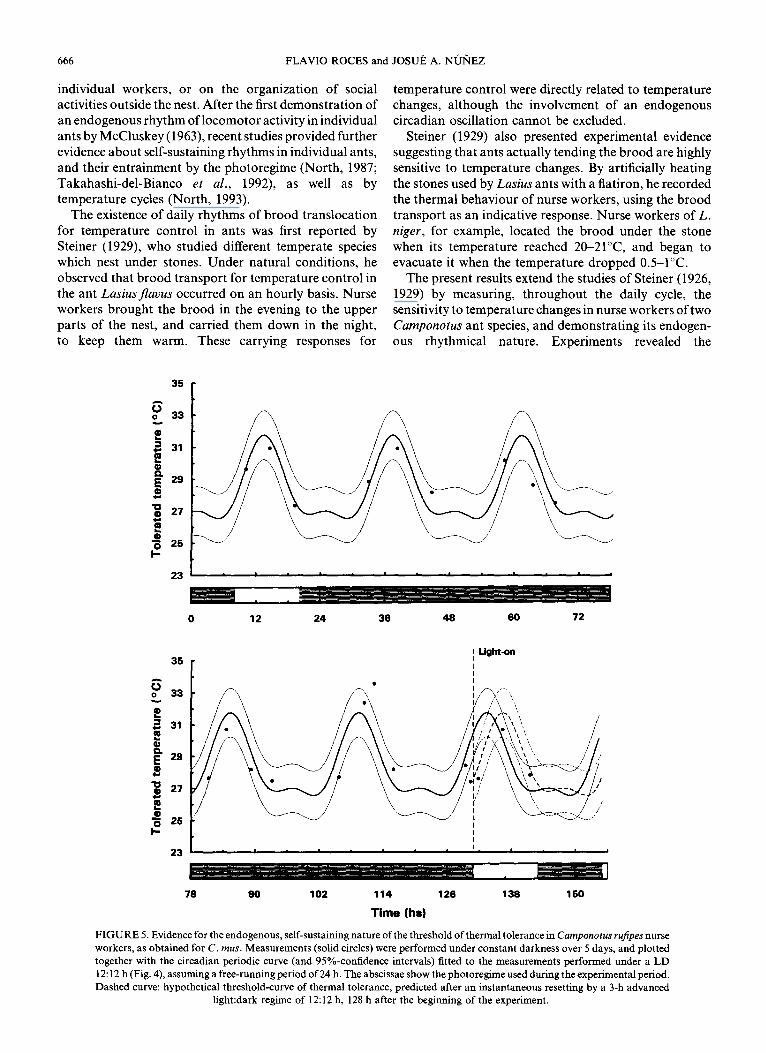
As for C. mus, the observed daily modulation of thermal tolerance during brood care raises the question of whether these daily changes are based on endogenously generated oscillations. To answer this question, the upper temperature limit for brood location was recorded in another experimental series by presenting the colony with constant darkness for 5 days, and later with the restored, 3-h phase-shifted 1ight:dark cycle. The results are IP 4118-B
summarized in the plot presented in Fig. 5, which is analogous to that of Fig. 2 for C. mus. The empirical data (circles) can be compared with the curve expected if the threshold of thermal tolerance is endogenously driven, assuming a free-running period of 24 h. Figure 5 shows that most of the measured values lay in the 95% confidence interval of the hypothetical curve, which supports the assumption of an oscillating, self-sustained thermal tolerance. As for C. mus, it was not possible to obtain a large number of measurements to fit a curve. Instead, the correspondence between measured and expected values was assessed by comparing their difference with the confidence interval of the hypothetical curve, as presented in Fig. 6. Results indicate that these differences were slight during the period under constant darkness, and contained in the 95%-confidence interval of the curve. Regrettably, the initial LD cycle was restored with an advance of only 3 h (light-on 131 h after the beginning of the assays, Figs 5 and 6) which makes clear distinction between values adjusted either to the free-running or to the re-entrained curves due to overlap of confidence intervals difficult. A single interesting measurement which can be mentioned is the value recorded shortly-after light-on, and corresponds to the instantaneously re-entrained threshold (dashed curve), but of course it does not suffice to demonstrate instantaneous phase-resetting by light-on.
DISCUSSION
Studies concerning daily rhythmical responses in ants have been focused either on the locomotor activity of
666 FLAVIO ROCES and JOSUe A. NOAEZ
individual workers, or on the organization of social activities outside the nest. After the first demonstration of an endogenous rhythm of locomotor activity in individual ants by McCluskey (1963), recent studies provided further evidence about self-sustaining rhythms in individual ants, and their entrainment by the photoregime (North, 1987; Takahashi-del-Bianco et al., 1992), as well as by temperature cycles (North, 1993).
The existence of daily rhythms of brood translocation for temperature control in ants was first reported by Steiner (1929) who studied different temperate species which nest under stones. Under natural conditions, he observed that brood transport for temperature control in the ant Lasiusjlavus occurred on an hourly basis. Nurse workers brought the brood in the evening to the upper parts of the nest, and carried them down in the night, to keep them warm. These carrying responses for
temperature control were directly related to temperature changes, although the involvement of an endogenous circadian oscillation cannot be excluded.
Steiner (1929) also presented experimental evidence suggesting that ants actually tending the brood are highly sensitive to temperature changes. By artificially heating the stones used by Lasius ants with a flatiron, he recorded the thermal behaviour of nurse workers, using the brood transport as an indicative response. Nurse workers of L. niger, for example, located the brood under the stone when its temperature reached 20-21”C, and began to evacuate it when the temperature dropped OS-1°C.
The present results extend the studies of Steiner (1926, 1929) by measuring, throughout the daily cycle, the sensitivity to temperature changes in nurse workers of two Camponotus ant species, and demonstrating its endogen- ous rhythmical nature. Experiments revealed the
35 r
23
78 SO 102 114 126 138 160
Time (hs)
FIGURE 5. Evidence for the endogenous, self-sustaining nature of the threshold of thermal tolerance in Camponotus rufipes nurse
workers, as obtained for C. mus. Measurements (solid circles) were performed under constant darkness over 5 days, and plotted
together with the circadian periodic curve (and 95%-confidence intervals) fitted to the measurements performed under a LD
12: 12 h (Fig. 4), assuming a free-running period of 24 h. The abscissae show the photoregime used during the experimental period.
Dashed curve: hypothetical threshold-curve of thermal tolerance, predicted after an instantaneous resetting by a 3-h advanced hght:dark regime of 12:12 h, 128 h after the beginning of the experiment.

THERMAL SENSITIVITY IN ANTS 667
5
G 4
0 3
7; al 2
g 1 $ G 0
i -’
f -2
a;
-5
f Light-on r I
I I I
. I I I I I I-------------
. . 9
I
. 1.’ .
. .
l . . I 0
.
l . %
I 0
. i------------------ l
.
4
b I I
‘
0 12 24 36 48 60 72 84 96 108 120 132 144 156
Time (hs)
FIGURE 6. Correspondence between measurements ofthe upper thermal tolerance under constant darkness, and the hypothetical
temperatures under the assumption of a self-sustained circadian thermal tolerance in C. rujipes nurse workers. Heavy and dashed
lines as in Fig. 3. Solid circles correspond to the differences between measurements and a free-running (TDD = 24 h) hypothetical
curve, whereas open circles specify those between measurements and a hypothetical curve, instantaneously reset by a 3-h advanced
phase-shift (after light-on). The abscissae show the photoregime used during the experimental period.
existence of a daily rhythm of maximal thermal tolerance in both species (Figs 1 and 4) which was driven by a circadian oscillation (Figs 2 and 5). For both species, the endogenous nature of the threshold of thermal tolerance was supported by a free-run under constant darkness, and, at least for C. mus, synchronization by a reliable environmental cue as the 1ight:dark cycle was experimen- tally demonstrated. In this species, the nurses’ threshold of thermal tolerance showed two minima along the day: one at noon, when environmental temperature usually reaches its maximum, and the second one at night, 8 h later. In C. rujipes, conversely, the thermal threshold reached its maximum at noon. Even when our experiments were performed in small laboratory colonies, direct measurements in field nests suggest that similar thermoregulatory responses actually occur in larger colonies (Races, 1988).
Circadian bimodal activity patterns in animals, as observed in C. mus, have been explained as the expression of either one oscillator controlling both peaks, or two independent oscillators individually controlling the two peaks (Wiedenmann 1980, 1983). While the split sensitivity curve in C. mus nurse workers (Fig. 1) suggests the involvement of two independent endogenous oscillations, the observed similar free-running periods of both spontaneous brood translocations, as well as previous data under regimes with light periods of different lengths (Races and Nufiez, 1989), support the idea of coupled brood translocations controlled by a single oscillator. Future measurements of ant thermal responses after single light pulses (phase response curves) should also help in understanding the unexpected re-setting
without transient cycles in C. mus, and clarify the major role of one or more oscillators. More interesting, since nurses in field colonies normally perform their tasks within the nest where light cannot penetrate, and have a priori no direct information about the light conditions outside the nest, how do they assess the environmental photoregime responsible for entrainment? Do nurses periodically abandon the nest, so that a light-sampling behaviour is responsible for photoentrainment, as observed in nocturnal rodents (DeCoursey, 1986)? Or are other cyclical environmental cues such as thermocycles important as entraining agents?
What is the importance of a circadian rhythm of thermal tolerance for brood care? Why do related species show such different time-courses of thermal tolerance along the day? As mentioned above, brood temperature should be carefully regulated because it directly affects development and ultimately colony growth rate (Porter, 1988). Camponotusmus is a dominant species in temperate climates in South America (Kusnezov, 1951, 1963). Since these ants nest under stones or occupy rotting wood on the ground, the colony is daily exposed to the natural changes in environmental temperature. The daily temperature cycle in temperate climates is characterized by a marked difference between maximal and minimal values. In C. mus, one of the two brood-transporting processes occurs roughly at the time where the highest environmental temperatures are expected (noon), and the other during the night. Although the latter is manifested prior to the occurrence of the daily minimum of environmental temperature (expected near dawn), there is no marked temperature difference between these times,

668 FLAVIO ROCES and JOSUE A. NUNEZ
especially inside the nest where thermal fluctuations are reduced (Races, 1988). It is therefore suggested that the daily variation in thermal preference would represent a strategy adapted to the marked daily changes in environmental temperature: brood temperature would be properly controlled if nurses are highly sensitive and pay attention to temperature only at the critical times of the day, when it approximates the extreme (maximal and minimal) daily values. Beyond these times, the immobile brood will remain at suitable thermal values because nest temperature will vary between these critical values. In other words, after the brood translocation at noon, nest temperature will be expected to decrease, and after another translocation event, it will not decrease much more. Measurements of temperature inside brood chambers in natural field nests support this assumption (Races, 1988). As a result, rather than continuously tracking their preferred temperature inside the nest as environmental temperature changes (which would need continuous transport of the brood), C. mus nurse workers control brood temperature by selecting their preferred values at two fixed times of the day, which correlate with the occurrence of daily extreme temperatures. If the daily thermal cycle at the selected place fits the thermal expectations of nurses, brood translocations will not be necessary at all, as previously observed during simulations of a daily thermal cycle (Races and Nuiiez, 1989). If it does not fit, brood translocations will be expected to occur either at the two critical times of the day, i.e. when the thermal sensitivity of nurses is maximal, or whenever temperature exceeds the upper limit determined by the nurses’ threshold of thermal tolerance.
Nurse workers of C. rujipes, on the other hand, showed no rhythm of brood translocation when a thermal gradient was established along the nest. They preferred the brood at a mean value of 25.O”C throughout the daily cycle. Their threshold of thermal tolerance for brood care, however, showed a marked daily variation of endogenous nature (circadian rhythm), being higher at the times when environmental temperature reaches its maximal values. Note that the threshold of thermal tolerance in this species, as opposed to C. mus, roughly resembles the daily course of environmental temperature (Fig. 4). Campono- tus rujipes is a dominant species in subtropical and tropical climates (Kusnezov, 195 1, 1963), where temperature differences between day and night are less pronounced than in temperate, more rigorous climates. Inside the nest, nurse workers remain mostly inactive over the brood as temperature rises during the light period, and engage in brood care particularly during the night, when foragers are active in bringing food for the colony (Jaffe and Sanchez, 1984). Therefore, if nurses select a suitable temperature during the night, when they are more sensitive (Fig. 4), brood temperature would thereafter remain under control, due to the expected slight fluctuations in environmental temperature. No further brood translocations would be needed, because the decreasing sensitivity (i.e. increasing tolerance) of nurses
during the day would allow them to tolerate the naturally-occurring thermal increase.
In conclusion, related C. mus and C. rufpes species have adopted different strategies to cope with the naturally-occurring daily temperature cycle. The time- course of their endogenously-driven thermal sensitivities suggest that their responses have been adapted in the course of evolution to the wide and slight daily fluctuations undergone by environmental temperature in temperate and tropical climates, respectively. The synchronization of the rhythmical thermal sensitivity with the photoregime assures that nurse workers are better prepared at the critical times of the day to react appropriately to environmental temperature. At noon, whereas nurses of C. mus are highly sensitive to temperature and show regulatory responses, i.e. brood translocations, C. rujipes nurses are less sensitive and tolerate the occurring thermal increases, remaining inactive over the brood. Interestingly, similar strategies for control of brood temperature have been reported for related honey bee species from different climates. At noon, temperate European honey bees (Apis mellifera Zigustica) respond to rising temperature by increasing their activity and collecting water to cool the nest. Tropical Africanized honey bees (Apis mellifera adan- sonii), on the other hand, tolerate the thermal increase and show at this time a marked decrease in activity (Nufiez, 1979). The involvement of endogenous rhythms of thermal sensitivity in honey bees, however, remains to be investigated.
REFERENCES
Brian M. V. (1952) The structure of a dense natural ant population.
J. Animal Ecol. 38, 12-24. Brian M. V. (1973) Temperature choice and its relevance to brood
survival and caste determination in the ant Myrmica rubra L. Physiol. Zool. 46, 245-252.
Buschinger A. (1973) The role of daily temperature rhythms in brood
development of ants of the tribe Leptothoracini (Hymenoptera;
Formicidae). In Effects of Temperature on Ectothermic Organisms. Ecological Implications and Mechanisms of Compensation (Ed. Wieser
W.), pp. 229-232. Springer, Berlin. Calabi P. and Porter S. D. (1989) Worker longevity in the fire ant,
Solenopsis invicta: ergonomics considerations of correlations between
temperature, size and metabolic rates. J. Insect Physiol. 35,643-650. Ceusters R. (1977) Social homeostasis in colonies of Formicapolyctena
Foerst (Hymenoptera: Formicidae): nestform and temperature
preferences. In Proceedings 8th International Congress IUSSI, pp. 11 l-l 12, Wageningen, The Netherlands.
DeCoursey P. J. (1986) Light-sampling behaviour in photoentrainment
of a rodent circadian rhythm. J. Camp. Physiol. A 159, 161-169. Elmes G. W. and Wardlaw J. C. (1983) A comparison of the effect of
temperature on the development of large hibernated larvae of four
species of Myrmica (Hymenoptera: Formicidae). Insect Sot. 30, 106118.
Heinrich B. (1993) The Hot-blooded Insects. Springer, Berlin. Jaffe K. and Sanchez C. (1984) Comportamiento alimentario y sistema
de reclutamiento en la hormiga Camponotus rufipes (Hymenoptera:
Formicidae). Acta Zool. Venez. 35, 2X&277.
Kato M. (1939) The diurnal rhythm of temperature in the mound of an
ant, Formica truncorum truncorum var. yessensis Forel, widely distributed at Mt Hakkoda. Sci. Rep. Tohoku Univ. 14, 53-64.

THERMAL SENSITIVITY IN ANTS 669
Kay C. A. R. (1978) Preferred temperatures of desert honey ants
(Hymenoptera: Formicidae). J. Thermal Biol. 3, 2 13-2 17. Kusnezov N. (1951) El genera Camponotus en la Argentina. Acta Zool.
Lilloana 12, 1833252. Kusnezov N. (1963) Zoogeografia de las hormigas de Sud America. Actu
Zool. Lilloana 19, 25-186. McCluskey E. S. (1963) Rhythms and clocks in harvester and Argentine
ants. Physiol. Zool. 36, 273-292. Messenger P. S. (1964) The influence of rhythmically fluctuating
temperatures on the development and reproduction of the spotted alfalfa aphid, Therioaphis maculata. J. Econ. Ent. 57, 71-76.
North R. D. (1987) Circadian rhythm of locomotor activity in
individual workers of the wood ant Formica rufu. Physiol. Ent. 12, 445454.
North R. D. (1993) Entrainment of the circadian rhythm of
locomotor activity in wood ants by temperature. Animal Behau. 45, 393-397.
Ntiiiez J. A. (1979) Comparative study of thermoregulation between
European and Africanized Apis mellifera in Brazil. J. Apic. Res. 18, 116121.
Porter S. D. (1988) Impact of temperature on colony growth and
developmental rates of the ant, Solenopsis invicta. J. Insect Physiol. 34, 1127-l 133.
Porter S. D. and Tschinkel W. R. (1993) Fire ant thermal preferences:
behavioural control of growth and metabolism. Behav. Ecol. Sociobiol. 32, 321-329.
Protomastro J. J. (1973) Relation entre el comportamiento de crianza
y un ritmo diario de preferencia termica en la hormiga Camponotus mus Roger. Physis (Buenos Aires) 32, 123-128.
Ratte H. T. (1985) Temperature and insect development. In
Environmental Physiology and Biochemistry of Insects (Ed. Hoffmann
K. H.), pp. 33-66. Springer, Berlin.
Races F. (1988) Disponibilidad de presas y comportamiento de crianza
en la hormiga Cumponotus mus Roger. Graduate thesis, University of
Buenos Aires, Argentina.
Races F. and Nuiiez J. A. (1989) Brood translocation and circadian
variation of temperature preference in the ant Camponotus mus. Oecologia 81, 33-37.
Scherba G. (1962) Mound temperatures of the ant Formica ulkei Emery.
Am. Midl. Nat. 67, 373-385. Steiner A. (1926) Temperaturmessungen in den Nestern der Waldameise
(Formica rufa var. rufa-pratensis) und der Wegameise (Lasius niger) wahrend des Winters. Mitt. Naturforsch. Ges. Bern, pp. l-19.
Steiner A. (1929) Temperaturuntersuchungen in Ameisennestern mit
Erdkuppeln, im Nest von Formica exsecta Nyl. und in Nestern unter
Steinen. Z. vergl. Physiol. 9, l-66. Takahashi-del-Bianco M., Benedito-Silva A. A., Hebling M. J. A.,
Marques N. and Marques M. D. (1992) Circadian oscillatory patterns
of oxygen uptake in individual workers of the ant Camponotus r@pes. Physiol. Em. 17, 377-383.
Wiedenmann G. (1980) Two peaks in the activity rhythm ofcockroaches
controlled by one circadian pacemaker. J. Comp. Physiol. 137, 249-254.
Wiedenmann G. (1983) Splitting in a circadian activity rhythm: the
expression of bilaterally paired oscillators. J. Comp. Physiol. 150, 51-60.
Wolfram S. (1991) Mathematics: a System for Doing Mathematics by Computer, 2nd edn. Addison-Wesley, Redwood City, Calif.
Acknowledgements-We are deeply indebted to B. Lopez Alvaredo and
I. Haubitz for help with Fourier analysis and statistics. We also thank
W. Farina, M. Giurfa, C. Lazzari and J. Tautz for critically reading the
manuscript and G. Wiedenmann for general comments on biological
rhythms. This research was conducted at the University of Buenos Aires,
Argentina, with funding from CONICET (grant to J.A. Nunez). This
paper was written at the University of Wilrzburg, Germany, with
funding from the Deutsche Forschungsgemeinschaft (Leibniz Prize to
B. Holldobler and SFB 251, Project 18 of the University of Wiirzburg).




















