
The shadow of forgotten ancestors differently constraints the fate of Alligatoroidea and...
11
DOI: 10.1111/j.1466-8238.2008.00426.x © 2008 The Authors 30 Journal compilation © 2008 Blackwell Publishing Ltd www.blackwellpublishing.com/geb Global Ecology and Biogeography, (Global Ecol. Biogeogr.) (2009) 18, 30–40 RESEARCH PAPER Blackwell Publishing Ltd The shadow of forgotten ancestors differently constrains the fate of Alligatoroidea and Crocodyloidea Paolo Piras 1,2 *, Luciano Teresi 3 , Angela D. Buscalioni 4 and Jorge Cubo 1 * ABSTRACT Aim We tested the hypothesis that the evolutionary fates of two sister groups (Alligatoroidea and Crocodyloidea) are differently constrained by phylogenetic and ecological (functional) factors in the face of climatic change. Location Global. Methods We quantified disparity in skull rostrum shape by means of geometric morphometrics. Mechanical performance of the rostrum was analyzed by applying beam theory calculations to morphological data and experimentally measured bite force. The phylogeny was expressed in the form of principal coordinates, the first ones of which were used as a set of explanatory variables. Extents of species occurrence were computed using species distribution maps. Finally, species maximum skull size were measured and considered as a proxy of maximum body size. We performed variation partitioning analyses in order to compare differential contributions of phylogenetic and ecological factors in Alligatoroidea and Crocodyloidea. Results Alligatoroidea show higher ‘pure’ historical components than Crocodyloidea in explaining both rostrum shape and extent of occurrence (after controlling for body size). On the contrary, geometric variation of skull rostra of Crocodyloidea unequivocally shows a higher ‘pure’ functional component (linked to performance on prey capture) and a higher phylogenetically structured environmental variation than those found in Alligatoroidea. Results obtained for body size variation are con- sistent with these patterns. In Alligatoroidea, body size variation contains a higher phylogenetic signal than in Crocodyloidea. Main Conclusions Our results suggest that Crocodyloidea and Alligatoroidea may react differently when faced with significant environmental changes. We predict that global climatic changes will have a more important effect on Crocodyloidea than in Alligatoroidea by (1) promoting trait shift, adaptation to the new diet and speciation and (2) modifying the geographical range distribution of species (which may track favourable ecological conditions). Keywords Alligatoroidea, climatic change, Crocodyloidea, functional performance, geometric morphometrics, rostral shape, variation partitioning. *Correspondence: Paolo Piras & Jorge Cubo, Université Pierre et Marie Curie, UMR CNRS 7179, 4 Pl Jussieu, BC 19, Paris 75005, France. E-mail: [email protected]; [email protected] 1 Université Pierre et Marie Curie, UMR CNRS 7179, 4 Pl Jussieu, BC 19, Paris 75005, France, 2 Center for Evolutionary Ecology, Largo San Leonardo Murialdo, 1, 00146 Roma, Italy, 3 Department of Studies on Structures, Mathematical Structures of Materials Physics, Universita degli Studi ‘Roma Tre’ Via Corrado Segre 6, I-00146 Roma,Italy, 4 Departamento de Biología, Unidad de Paleontología, Facultad de Ciencias Universidad Autónoma de Madrid, 28049 Cantoblanco, Madrid, Spain INTRODUCTION Diniz-Filho & Bini (2008) published a stimulating paper suggest- ing that the structure of the variation of traits among related species may contain keys for predicting their evolutionary fate. In their own words, ‘the shadow of forgotten ancestors’ could strongly constrain the possibilities of overcoming environmental transformations like the current global climatic change. In this paper we make predictions on how Crocodyloidea and Alli- gatoroidea may react when faced with climatic change by analys- ing rostral shape, rostral structural performance, geographical extent of occurrence and body size in a phylogenetic context. The proposal of Diniz-Filho & Bini (2008) is based on a new (phylogenetic) comparative method allowing quantification of fractions of character variation explained by different factors: a fraction exclusively explained by ecology, a fraction of phylo- genetically structured environmental variation and a fraction exclusively explained by phylogeny. The first fraction refers to
Transcript of The shadow of forgotten ancestors differently constraints the fate of Alligatoroidea and...

DOI: 10.1111/j.1466-8238.2008.00426.x © 2008 The Authors
30
Journal compilation © 2008 Blackwell Publishing Ltd www.blackwellpublishing.com/geb
Global Ecology and Biogeography, (Global Ecol. Biogeogr.)
(2009)
18
, 30–40
RESEARCHPAPER
Blackwell Publishing Ltd
The shadow of forgotten ancestors differently constrains the fate of Alligatoroidea and Crocodyloidea
Paolo Piras
1,2
*, Luciano Teresi
3
, Angela D. Buscalioni
4
and Jorge Cubo
1
*
ABSTRACT
Aim
We tested the hypothesis that the evolutionary fates of two sister groups(Alligatoroidea and Crocodyloidea) are differently constrained by phylogenetic andecological (functional) factors in the face of climatic change.
Location
Global.
Methods
We quantified disparity in skull rostrum shape by means of geometricmorphometrics. Mechanical performance of the rostrum was analyzed by applyingbeam theory calculations to morphological data and experimentally measured biteforce. The phylogeny was expressed in the form of principal coordinates, the firstones of which were used as a set of explanatory variables. Extents of species occurrencewere computed using species distribution maps. Finally, species maximum skull sizewere measured and considered as a proxy of maximum body size. We performedvariation partitioning analyses in order to compare differential contributions ofphylogenetic and ecological factors in Alligatoroidea and Crocodyloidea.
Results
Alligatoroidea show higher ‘pure’ historical components than Crocodyloideain explaining both rostrum shape and extent of occurrence (after controlling forbody size). On the contrary, geometric variation of skull rostra of Crocodyloideaunequivocally shows a higher ‘pure’ functional component (linked to performanceon prey capture) and a higher phylogenetically structured environmental variationthan those found in Alligatoroidea. Results obtained for body size variation are con-sistent with these patterns. In Alligatoroidea, body size variation contains a higherphylogenetic signal than in Crocodyloidea.
Main Conclusions
Our results suggest that Crocodyloidea and Alligatoroideamay react differently when faced with significant environmental changes. We predictthat global climatic changes will have a more important effect on Crocodyloideathan in Alligatoroidea by (1) promoting trait shift, adaptation to the new diet andspeciation and (2) modifying the geographical range distribution of species (whichmay track favourable ecological conditions).
Keywords
Alligatoroidea, climatic change, Crocodyloidea, functional performance, geometric
morphometrics, rostral shape, variation partitioning.
*Correspondence: Paolo Piras & Jorge Cubo, Université Pierre et Marie Curie, UMR CNRS 7179, 4 Pl Jussieu, BC 19, Paris 75005, France. E-mail: [email protected]; [email protected]
1
Université Pierre et Marie Curie, UMR CNRS
7179, 4 Pl Jussieu, BC 19, Paris 75005, France,
2
Center for Evolutionary Ecology, Largo San
Leonardo Murialdo, 1, 00146 Roma, Italy,
3
Department of Studies on Structures,
Mathematical Structures of Materials Physics,
Universita degli Studi ‘Roma Tre’ Via Corrado
Segre 6, I-00146 Roma,Italy,
4
Departamento de
Biología, Unidad de Paleontología, Facultad de
Ciencias Universidad Autónoma de Madrid,
28049 Cantoblanco, Madrid, Spain
INTRODUCTION
Diniz-Filho & Bini (2008) published a stimulating paper suggest-
ing that the structure of the variation of traits among related
species may contain keys for predicting their evolutionary fate.
In their own words, ‘the shadow of forgotten ancestors’ could
strongly constrain the possibilities of overcoming environmental
transformations like the current global climatic change. In this
paper we make predictions on how Crocodyloidea and Alli-
gatoroidea may react when faced with climatic change by analys-
ing rostral shape, rostral structural performance, geographical
extent of occurrence and body size in a phylogenetic context.
The proposal of Diniz-Filho & Bini (2008) is based on a new
(phylogenetic) comparative method allowing quantification of
fractions of character variation explained by different factors:
a fraction exclusively explained by ecology, a fraction of phylo-
genetically structured environmental variation and a fraction
exclusively explained by phylogeny. The first fraction refers to

Phylogenetic effects and climatic change
© 2008 The Authors
Global Ecology and Biogeography
,
18
, 30–40, Journal compilation © 2008 Blackwell Publishing Ltd
31
responses of species traits to environmental conditions,
regardless of trait values in closely related species. This fraction
has been considered to match the cladistic concept of auta-
pomorphies at the species level (Cubo, 2004; Cubo
et al
., 2005).
The fraction of phylogenetically structured environmental varia-
tion corresponds to the portion of character variation explained
by ecology
and
phylogeny (Desdevises
et al
., 2003). This fraction
may reflect niche conservatism: closely related species share
character states because they tend to occupy similar ecological
niches (Desdevises
et al
., 2003). Therefore, a high fraction of
character variation explained by ecology
and
phylogeny may sug-
gest that closely related species were tracking the environmental
conditions in which their last common ancestor originally
evolved (Diniz-Filho & Bini, 2008). This fraction matches the
cladistic concept of synapomorphies with functional (ecological)
significance (Cubo, 2004; Cubo
et al
., 2005). Finally, the ‘pure’
phylogenetic fraction refers to character states that were adapted
to the past environment and are now in a suboptimal condition
(this is possible when the feature is not tightly linked to fitness).
In cladistic terminology, this fraction may correspond to syna-
pomorphies without a clear functional significance in extant species
(Cubo, 2004; Cubo
et al
., 2005). According to Diniz-Filho & Bini
(2008), species with a high fraction of character variation
exclusively explained by ecology may react differently to climatic
change (through trait shift and adaptation to the new conditions)
than either species with a high fraction exclusively explained by
phylogeny or species with a high fraction of phylogenetically
structured environmental variation. Although these authors
explicitly used the species level, herein we use their approach at
the level of clades.
Extant species of Crocodylia constitute a suitable group to
which to apply the above model. This clade contains two sister
groups (Crocodyloidea and Alligatoroidea) showing many
specificities (in terms of species richness, ecological diversity and
morphological disparity) that, according to the model proposed
by Diniz-Filho & Bini (2008), may predispose them to react
differently to climatic change. We choose the crocodilian ros-
trum as the best morphological structure to test predictions
made by Diniz-Filho & Bini (2008). It is involved in prey capture,
killing and manipulation, and for this reason it is suitable
for testing the relative importance of functional performance in
the two clades in a phylogenetic context.
MATERIAL AND METHODS
Material
The variation of skull shape explained in terms of functional per-
formance and phylogenetic signal of all extant Crocodyloidea
and Alligatoroidea was used to predict the evolutionary fate of
these clades. Shape data came from a geometric morphometric
analysis of rostra, and mechanical performance was evaluated by
applying beam theory calculations to morphological data and
experimentally measured bite forces (Erickson
et al
., 2003,
2005). Detailed species extents of occurrence were taken from
the IUCN Crocodile Specialist Group official website (http://
iucncsg.org/ph1/modules/Home/), and phylogeny from the
most recent published studies (Brochu, 2000; McAliley
et al
.,
2006; Janke
et al
., 2006). We analysed all living species of
Alligatoroidea (157 individuals, 8 species) and Crocodyloidea
(224 individuals, 14 extant species plus three extinct species –
Crocodylus robustus
from the Pleistocene of Madagascar,
Crocodylus
ossifragus
from the Pleistocene of Java and
Dollosuchus dixoni
from the Early Eocene of Belgium). Perfect three-dimensional
skulls were available for this sample. No undeformed three-
dimensional fossils for Alligatoroidea have been found. Details
about specimens are available in Appendix S1 in Supporting
Information.
Phylogeny
The modern phylogeny of Crocodylia is characterized by a
conflict between molecular studies and morphological/palaeon-
tological studies (see Brochu, 2003, McAliley
et al
., 2006, and
Janke
et al
., 2006, with references for a review). This conflict
regards the position of Gavialoidea within which is placed the
sole extant species belonging to this group,
Gavialis gangeticus
.
In morphological/palaeontological studies, Gavialoidea is placed
outside all other species (as a sister taxon of the remaining
members of the crown group Crocodylia) with a Late Cretaceous
divergence time (Brochu, 2003), while according to molecular
studies it is a sister taxon of
Tomistoma
with their separation
estimated, at the very earliest, in the Late Eocene (Janke
et al
.,
2006).
The remaining extant crocodilian species are unambiguously
grouped into two clades: Alligatoroidea and Crocodyloidea.
Some uncertainties still exist within the genus
Crocodylus
, but the
phylogeny of the group is rather stable. Here we took into
consideration a ‘consensus’ phylogeny mainly deriving from
molecular data (Brochu, 2000; McAliley
et al
., 2006; Janke
et al
.,
2006; Fig. 1). We specifically avoided considering
Gavialis
because of its uncertain position. The consensus tree was
calibrated estimating the branch lengths in millions of
years according to Brochu’s (2003) fossil calibration. We built a
phylogenetic distance matrix for the whole sample (Fig. 1) using
topology and divergence times in Mesquite 2.0 (Maddison &
Maddison, 2007). The matrix was exported using the
Stratigraphic Tools additional module for Mesquite (Josse
et al
.,
2006). Then, following Diniz-Filho
et al
. (1998), phylogeny was
expressed in the form of principal coordinates (PC), which were
subjected to a broken stick analysis (Legendre & Legendre, 1998)
to select a number of them (representative of phylogeny) to be
used in variation partitioning analyses as independent variables.
Geographical extent of occurrence
The geographical extent of occurrence is defined by the IUCN
‘... as the area contained within the shortest continuous imagi-
nary boundary which can be drawn to encompass all the known,
inferred or projected sites of present occurrence of a species,
excluding cases of vagrancy’. We calculated these areas starting
from the maps available for all extant species from the IUCN

P. Piras
et al.
© 2008 The Authors
32
Global Ecology and Biogeography
,
18
, 30–40, Journal compilation © 2008 Blackwell Publishing Ltd
Crocodile Specialist Group official website (http://iucncsg.org/
ph1/modules/Home/). We used a single map source to avoid
problems linked to heterogeneity of error measurements.
Another source of uncertainty may be linked to the fact that all
species inhabit fresh to brackish waters. Only land areas have
been computed to partially circumvent this problem.
Geometric morphometrics (analysis of rostrum shape disparity)
Twenty-one unilateral three-dimensional landmarks were digi-
tized on the left side of skull rostra to quantify morphospace
occupation for all living Alligatoroidea and Crocodyloidea. Data
collection was performed with an Immersion Microscribe G2®.
Rostrum configuration and landmark definitions are shown in
Fig. 2. These landmark configurations accurately reflect subtle
shape differences among Alligatoroidea and Crocodyloidea over
other typological rostral categories (e.g. slender-snout, blunt-snout
or generalized as used by other authors; see Brochu, 2001).
Landmarks allow precise tests on whether rostral variation is
constricted by phylogeny, by ecology or by both, since they
capture the overall contour, shape details and size.
Generalized Procrustes analysis (Bookstein, 1991) has been
selected for the analysis of shape. This technique can perform a
full investigation of shape differences eliminating the effect of
position, rotation and size. All homologous landmarks of any
configuration are superimposed, minimizing the Procrustes
distance, which is defined as ‘the square root of the sum of
squared differences between the positions of the landmarks in
two optimally (by least-squares procedure) superimposed
configurations at centroid size.’ (Bookstein, 1991). The generalized
least squares method has been used for the superimposition
procedure. All configurations are scaled at the same size unit, i.e.
the centroid size, which is defined as ‘the square root of the sum
of squared distances of a set of landmarks from their centroid.’
The centroid is ‘that point, in a given configuration, whose
coordinates of any dimension are represented by the arithmetic
mean of corresponding dimension coordinates of all landmarks
of that configuration’ (Bookstein, 1991). Procrustes residuals
were subjected to a principal components analysis and the scores
(PCs) were subjected to a broken stick analysis (Legendre &
Legendre, 1998) to select a number of them (representative of
shape) to use in successive comparative analyses. Procrustes
analysis and successive principal components analyses were
performed in Morphologika 2.5 (O’Higgins & Jones, 2007).
Two separate Procrustes superimpositions (Rohlf & Slice,
1990) were performed for Alligatoroidea and Crocodyloidea.
In order to eliminate intraspecific allometric effects, for each
species all individuals were standardized at their maximum
recorded size using IMP series software (Sheets, 2006). The
goodness of standardization was tested by per species multivariate
regressions of original log CS (log
10
centroid size) against shape
variables of standardized data.
Beam theory (analysis of rostral mechanical performance)
Beam theory was adopted to analyse rostral mechanical performance.
The rostrum was modelled as a hollow semi-circular conic trunk
cantilevered at its larger section, corresponding to the antorbital
diameter (Fig. 3). This geometry was considered as a good
approximation of the crocodilian rostrum, probably better
than that of Metzger
et al
. (2005) that used a full tapered ellipsoid
as a model. Dimensions of the model came directly from the
Figure 1 Phylogeny of Crocodylia used in this work. Geological time scale in millions of years.

Phylogenetic effects and climatic change
© 2008 The Authors
Global Ecology and Biogeography
,
18
, 30–40, Journal compilation © 2008 Blackwell Publishing Ltd
33
height and width of each single specimen at three precise coronal
section levels: premaxillary constriction diameter, fourth
alveolus diameter and pre-orbital diameter. To build the final
geometry, sagittal distances between sections were calculated
and the thickness of the whole object was assumed to represent
10% of the vertical axis of the corresponding section. A Matlab
routine (Trave; Teresi, 2006) read our geometric database,
implemented and solved a beam problem under different load
conditions and eventually computed the stress states in the beam
wall. Linear measurements were computed directly from geo-
metric morphometrics data, projecting on the proper planes
appropriate landmarks through the program Jcocco (Iocchi &
Piras, 2006). The geometric properties of the solid were specified
before any calculation under load conditions.
Bite force was calculated and scaled for each specimen
according to the equation of Erickson
et al
. (2003): log
10
(biteforce) =
−
0.46 + 2.57 log
10
(head length). Head length was calculated by
projecting the distance between the tip of the premaxilla and the
posterior-most tip of the quadrate articular surface on the
sagittal plane. This equated the measurements used by Erickson
et al
. (2003), namely the projected jaw length from jaw tip to
articular surface. Erickson
et al
. (2003) measured the maximum
bite force on one side only for
Alligator mississippiensis
at the level
of fourth maxillary alveolus. Then, a symmetric bite was inferred
with a double force. Moreover, given that this model is a beam
simulation based on steady force distribution, the values of bite force
corresponding to the first section (at the premaxillo-maxillary
Figure 2 Landmark configuration. Landmarks are: 1, medial dorsal anterior projection of frontal; 2, medial dorsal anterior projection of nasals; 3, posterolateral tip of external nares opening; 4, anterior tip of external nares opening (in Alligator and Osteolaemus it just corresponds to anterior profile); 5, posterolateral tip of premaxilla; 6, lateral skull margin at antero-orbital diameter; 7, distal tip of premaxilla; 8, distal tip of external nares contour; 9, anterior tip of premaxilla; 10, posterior-most ventral tip of maxillary–premaxillary contact; 11, anterior-most tip maxillo-palatine contact; 12, premaxillary constriction just on premaxillary–maxillary suture, along the alveolar margin; 13, lateral tip of fourth alveolus; 14, lateral tip of fifth alveolus; 15, medial tip of fifth alveolus; 16, medial tip of fourth alveolus; 17, lateral tip of seventh alveolus; 18, lateral tip of eighth alveolus; 19, lateral tip of eleventh alveolus; 20, lateral tip of twelfth alveolus; 21, anterior tip of incisive foramen.
Figure 3 Geometric model for beam theory analysis built to analyse rostral mechanical performance using the actual skull morphology.

P. Piras
et al.
© 2008 The Authors
34
Global Ecology and Biogeography
,
18
, 30–40, Journal compilation © 2008 Blackwell Publishing Ltd
constriction) were calculated as F1 = F2
×
L_23/(L_12 + L_23),
where L_
ij
is the distance between section
i
and
j
, with section
1 the shortest and F2 is the force calculated experimentally by
Erickson
et al
. (2003). Erickson
et al
. (2005) directly measured
the maximal bite force from all extant crocodilian species.
These authors did not publish quantitative results but they stated
that, for all species, both absolute values and values scaled to
allometric change followed the pattern published by Erickson
et al
. (2003) for
A. mississippiensis
. They also stated that this
was true both for brevirostral and longirostral forms.
Six loading conditions were simulated (Fig. 4). They were
chosen as representatives of a great variety of biting circum-
stances. Basically, we can distinguish between symmetrical and
asymmetrical positions that imply very different torsional and
bending responses for various rostral morphologies. Bites with
force exerted on the first and/or the second section level were
considered. Details about mechanical quantities and their
computation are given in Appendix S2.
Material properties were set with a Poisson’s ratio of 0.3 and
with a Young elastic modulus of 10 GPa. These values correspond
to the properties of trabecular bone reported in the reference list
of McGowan (1999). However, given the comparative aim of this
investigation (that is not strictly ‘validative’), possible differences
for these constants with other studies are not relevant and should
not affect the results. The basic idea was to correlate the shape
variables obtained from geometric morphometric analysis with
the simulated rostral mechanical response. Even though six
loading conditions were simulated, sigmas of simulation 5
were only considered as a proxy of functional performance. In
fact, simulation 5 represents a somewhat more realistic bite.
Other conditions revealed identical results relative to species
differences, just scaled in intensity. Thus they would be
redundant in successive statistical analyses. Sigma represents
the maximum normal stress on the section. This parameter is
often used to predict the failure of brittle material; thus, we
considered it to be linked to the overall mechanical performance
of the rostrum.
Comparative methods
The use of classical regression methods requires that the values of
each variable be independent of each other but, in an interspecific
study, the phylogenetic relationships between species imply that
these values are non-independent (Felsenstein, 1985; Harvey &
Pagel, 1991; Garland
et al
., 1992). To overcome this problem, we
used variation partitioning analyses including the phylogeny as
an explanatory factor (Desdevises
et al
., 2003). This method
allows an assessment of the portions of variation of a dependent
variable: (1) explained exclusively by phylogeny; (2) explained
exclusively by ecology (function); or (3) explained by the overlap
of these two sets of variables. It is an extension of the partitioning
method proposed in ecology (Borcard
et al
., 1992; Borcard &
Legendre, 1994; Legendre & Legendre, 1998). Similarly, a method
of variation partitioning among three factors, including the
phylogeny as a set of explanatory variables (Cubo
et al
., 2008) has
been used as an extension of previous developments in ecology
(Anderson & Gribble, 1998; Cushman & McGarigal, 2002;
Økland, 2003). Details about steps of the variation partitioning
method can be found in Desdevises
et al
. (2003) and Cubo
et al
.
(2008). We first applied variation partitioning analysis to a model
involving rostrum shape as a matrix of dependent variables and
two sets of explanatory variables: log
10
(sigmas) (as proxy of
biomechanical performance or rostrum function) and the
selected phylogenetic principal coordinates (as proxy of phylogeny).
This analysis was performed separately in Alligatoroidea and
Crocodyloidea to check the differential contribution of function
and phylogeny to variation of rostrum shape in the two groups.
In a second analysis, we tested the influence of rostrum shape
(as a proxy for degree of trophic specialization) on geographical
extent of species occurrence, controlling for phylogenetic
background, and maximum recorded body size. This exploratory
analysis was also performed separately for Alligatoroidea and
Crocodyloidea. Lastly we quantified the fraction of interspecific
maximum body size variation explained by phylogeny. This
analysis aimed to see whether, in the two groups, this trait
contained a differential degree of phylogenetic signal.
RESULTS
Analysis of rostrum shape disparity using geometric morphometrics
Our standardization procedure eliminated any allometric effect.
Per species multivariate regressions between the original log CS
and shape variables coming from standardized data returned
neither significant Wilk’s lambda nor singular correlations.
The broken stick model returned the first two PCs as significant
for Crocodyloidea, explaining, respectively, 65.99% and 10.83%
of total variation, and the first three for Alligatoroidea,
explaining, respectively, 39.21%, 19.79% and 8.43% of total
variation.
Figure 4 Loading conditions studied for the beam analysis. Schematic drawings show sections of rostrum corresponding to the described anatomical levels (see text). Arrows indicate forces (F) applied.
Phylogenetic effects and climatic change
© 2008 The Authors
Global Ecology and Biogeography
,
18
, 30–40, Journal compilation © 2008 Blackwell Publishing Ltd
35
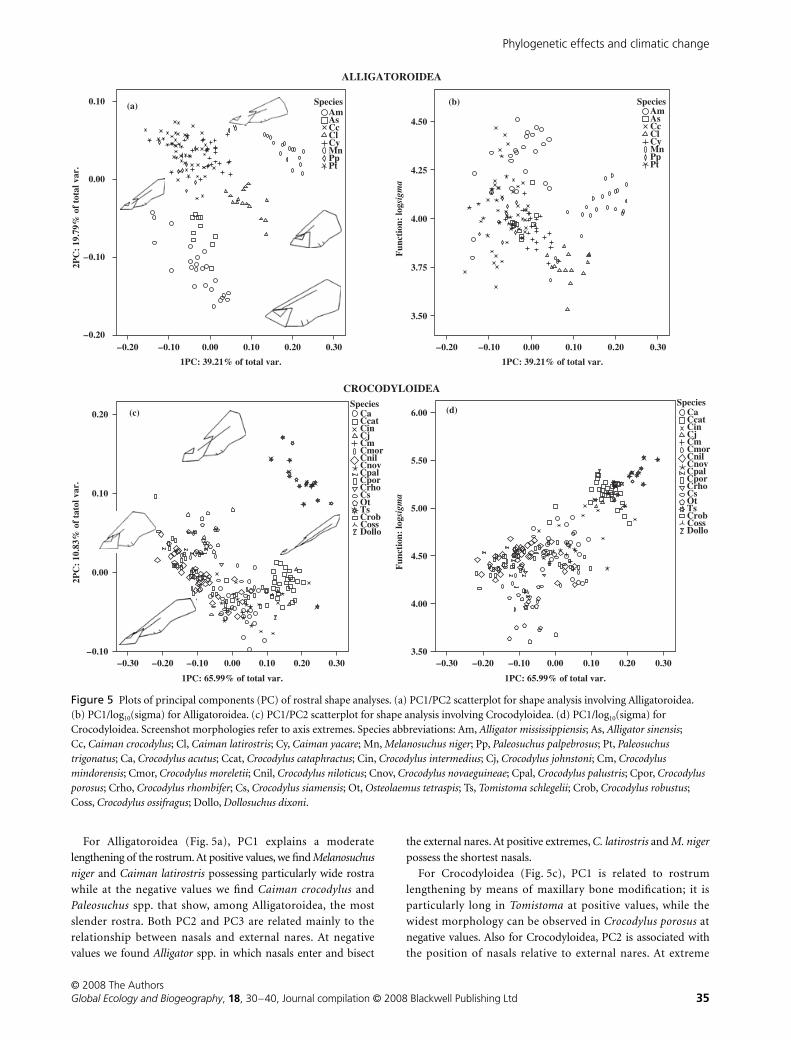
For Alligatoroidea (Fig. 5a), PC1 explains a moderate
lengthening of the rostrum. At positive values, we find
Melanosuchus
niger
and
Caiman latirostris
possessing particularly wide rostra
while at the negative values we find
Caiman crocodylus
and
Paleosuchus
spp. that show, among Alligatoroidea, the most
slender rostra. Both PC2 and PC3 are related mainly to the
relationship between nasals and external nares. At negative
values we found
Alligator
spp. in which nasals enter and bisect
the external nares. At positive extremes,
C. latirostris
and
M. niger
possess the shortest nasals.
For Crocodyloidea (Fig. 5c), PC1 is related to rostrum
lengthening by means of maxillary bone modification; it is
particularly long in
Tomistoma
at positive values, while the
widest morphology can be observed in
Crocodylus porosus
at
negative values. Also for Crocodyloidea, PC2 is associated with
the position of nasals relative to external nares. At extreme
Figure 5 Plots of principal components (PC) of rostral shape analyses. (a) PC1/PC2 scatterplot for shape analysis involving Alligatoroidea. (b) PC1/log10(sigma) for Alligatoroidea. (c) PC1/PC2 scatterplot for shape analysis involving Crocodyloidea. (d) PC1/log10(sigma) for Crocodyloidea. Screenshot morphologies refer to axis extremes. Species abbreviations: Am, Alligator mississippiensis; As, Alligator sinensis; Cc, Caiman crocodylus; Cl, Caiman latirostris; Cy, Caiman yacare; Mn, Melanosuchus niger; Pp, Paleosuchus palpebrosus; Pt, Paleosuchus trigonatus; Ca, Crocodylus acutus; Ccat, Crocodylus cataphractus; Cin, Crocodylus intermedius; Cj, Crocodylus johnstoni; Cm, Crocodylus mindorensis; Cmor, Crocodylus moreletii; Cnil, Crocodylus niloticus; Cnov, Crocodylus novaeguineae; Cpal, Crocodylus palustris; Cpor, Crocodylus porosus; Crho, Crocodylus rhombifer; Cs, Crocodylus siamensis; Ot, Osteolaemus tetraspis; Ts, Tomistoma schlegelii; Crob, Crocodylus robustus; Coss, Crocodylus ossifragus; Dollo, Dollosuchus dixoni.

P. Piras et al.
© 2008 The Authors36 Global Ecology and Biogeography, 18, 30–40, Journal compilation © 2008 Blackwell Publishing Ltd
positive values we find Tomistoma, in which nasals are extremely
short, while at the opposite lies Crocodylus acutus, which
possesses the longest nasals. PC2 is associated also with a
maxillary lengthening at negative values, so that two longirostral
forms (as Tomistoma and C. acutus) occupy (just along PC2)
opposite positions. Nevertheless they are visibly close along PC1.
Analysis of mechanical performance of the rostrum using beam theory
Log10(sigmas) coming from simulation 5 were correlated with
dominant dimensions of shape analysis of Alligatoroidea and
Crocodyloidea, respectively. Bivariate relationships between
log10(sigmas) and dominant dimensions of Alligatoroidea and
Crocodyloidea are shown in Fig. 5(b) and 5(d) respectively.
Obviously, longirostral forms present higher log10(sigmas)
values, but the correlation between performance and shape is
clearly different in the two groups, being visibly stronger in
Crocodyloidea. In fact, while in this group some species are
unequivocally longirostral, no such morphologies are present in
Alligatoroidea; only a minor variability is present, spanning from
Melanosuchus to Paleosuchus.
Analysis of rostrum shape, extent of occurrence and maximum size using variation partitioning
All variation partitioning analyses returned different quantitative
results for Alligatoroidea and Crocodyloidea. In the analysis of
rostrum shape, as shown in Fig. 6, both the functional fraction
and overlapping fraction are higher in Crocodyloidea than in
Alligatoroidea. Conversely, the phylogeny explains much more
variation in Alligatoroidea. All R2 are significant. Partition
variation applied to the extent of occurrence (as the dependent
variable), phylogeny, rostrum shape and maximum body size
(as independent variables) shows that Crocodyloidea possesses
a functional factor (expressed by rostrum shape) higher than
Alligatoroidea (Fig. 7). Analyses performed on maximal body
size (as the dependent variable) and phylogeny (as independent
variables) showed that the phylogenetic signal is higher in
Alligatoroidea (61.4% of size variation explained by phylogeny)
than in Crocodyloidea (54.3%).
DISCUSSION
Diniz-Filho & Bini (2008) suggested that the structure of the
variation of characters in clades may contain keys for predicting
their evolutionary fate. We are aware that a proper analysis of
fitness involves testing, at the level of populations, the effect of
morphological variation on functional performance, and the
effect of performance on fitness (Arnold, 1983; Kingsolver &
Huey, 2003). Thus comparative methods designed to deal with
Figure 6 Results from variation partitioning analyses involving significant principal coordinates of rostral shape analysis (as the dependent variable matrix), performance and phylogeny (as independent variable matrices). Fraction ‘a’ corresponds to the ‘pure’ functional component, fraction ‘c’ to the ‘pure’ phylogenetic component, fraction ‘b’ to the phylogenetically structured functional variation and fraction ‘d’ to the unexplained component.
Figure 7 Results from variation partitioning analyses involving extent of occurrence (as the dependent variable), phylogeny, rostrum shape principal coordinates and maximum species size (as independent variable matrices). Fraction ‘A’ corresponds to ‘pure’ shape, fraction ‘B’ to ‘pure’ species maximal size and fraction ‘C’ to pure phylogeny. For simplicity, fractions not discussed in the text (D, E, F and G) were omitted.

Phylogenetic effects and climatic change
© 2008 The Authors Global Ecology and Biogeography, 18, 30–40, Journal compilation © 2008 Blackwell Publishing Ltd 37
variation at the level of species, such as those used in the present
study, cannot be used to analyse fitness (i.e. morphological disparity
cannot be used as a surrogate of fitness). Here we only tested the
first component of Arnold’s (1983) model: the relationship
between morphological disparity (quantified using geometric
morphometrics) and functional performance (assessed using the
biomechanical approach of beam theory). Following Diniz-Filho
& Bini (2008), the results of these tests were used to make the
following predictions on the evolutionary fate of clades. (1)
Confronted with climatic change, clades showing characters with
high ‘pure’ ecological (functional) components may quickly
respond through trait shift and adaptation to the new conditions.
(2) Characters showing a high fraction explained by ecology and
phylogeny may be tightly linked to environmental conditions
because this variation is the outcome of niche conservatism
(Desdevises et al., 2003). Thus, confronted with climatic change,
species showing this kind of character may track the favourable
ecological conditions by geographical range shifts, generating
phylogenetic signal by persistence of ancestral phenotypes
(Diniz-Filho & Bini, 2008). (3) Finally, characters showing a high
‘pure’ phylogenetic fraction are in a suboptimal condition. In the
past, these characters did not track environmental changes (this
was possible because they were not tightly linked to fitness).
Thus they do not reflect current conditions but past ecological
conditions. In this case, the response to climatic change will
depend upon whether characters are more or less linked to
fitness. When they are not tightly linked to fitness, characters will
persist out of their optimum (reflecting the past ecological
conditions in which they originated). On the contrary, when
they are tightly linked to fitness, organisms will either track
favourable ecological conditions by geographical range shifts
(when possible) or become extinct (Diniz-Filho & Bini, 2008).
This model can be used in two ways. First, it can be used to make
predictions about how a given clade may react faced with a given
environmental perturbation. Our study is an example of this
kind of approach: we have analysed skull shape, skull performance,
geographical extent of occurrence and body size of all extant
species of Crocodyloidea and Alligatoroidea in a phylogenetic
context, in order to make predictions about how these clades
may react when confronted with climatic change. Second, it can
be used retrospectively, by making predictions about how
different clades (e.g. crocodiles and dinosaurs) reacted to a given
environmental perturbation (e.g. the Cretaceous–Tertiary crisis).
Although in this second case predictions can be easily tested
(by analysing morphological disparity, species richness and
geographical distribution of crocodiles and dinosaurs before and
after the biological crisis), in the first case (and in our study),
unfortunately, predictions cannot be tested.
Global climatic change has an important effect on the
geographical range distribution of many animal species, e.g.
modifying the species composition of communities, and there-
fore the potential prey of alligators and crocodiles. Thus climatic
change may have an indirect effect on the diet of these animals,
and may affect both their performance in prey capture (which
depends on the geometry of the skull rostrum), and their fitness.
In Crocodyloidea, geometric variation of skull rostra shows a
‘pure’ functional (biomechanical) component (linked to
performance in prey capture) more than four times higher
than that of Alligatoroidea (Fig. 6). The fraction of geometric
variation of skull rostra explained by phylogeny and function is
also more than four times higher in Crocodyloidea than in
Alligatoroidea (Fig. 6). According to the hypothesis of Diniz-
Filho & Bini (2008), these results suggest that the geometry of
skull rostra is tightly linked to performance in prey capture (an
important component of fitness) in Crocodyloidea. Species of
this clade may quickly respond to climatic changes through
either (1) trait shift, adaptation to a new diet and speciation or
(2) geographical range shifts (tracking favourable ecological
conditions), as suggested, respectively, by the high ‘pure
functional’ component, and the high ‘phylogenetically
structured functional variation’ component of skull rostrum
variation (Fig. 6). In contrast, in Alligatoroidea, geometric
variation of skull rostra shows a high ‘pure’ historical (phylo-
genetic) component (more than four times higher than in
Crocodyloidea, Fig. 6). According to the hypothesis by Diniz-
Filho & Bini (2008), this result suggests that in Alligatoroidea,
geometric variation of skull rostra is not tightly linked to
fitness; therefore, this character will persist out of the optimum
and generate a phylogenetic signal.
Rostral shape in Alligatoroidea is associated with an overall
change in skull morphology along its evolutionary history.
A basal cladogenesis that can be traced back to the Palaeocene
gave rise to two major clades, Alligatorinae and Caimaninae;
the latter comprises the recent genera Paleosuchus, Caiman
and Melanosuchus. Rostral and skull shapes differ in both major
clades in the cranio-madibular articulation, maxillary festooning,
pattern dentition, palatine curvature, presence of rostral canthi,
presence of palpebral ossification, frontal ossification and
robustness of the cranial table (see also Brochu, 1999). On the
contrary, in Crocodyloidea rostral morphology (relative length
and shape) is extremely variable along their evolutionary history.
Longirostry has come out recurrently as a homoplasic feature
linked to dietary adaptation and fishing capabilities even within
the clade Crocodylinae (containing the recent genus Crocodylus)
since their origin near the Eocene–Oligocene boundary (Aoki,
1983; Tchernov, 1986; Brochu, 2003). Two extreme cases include
the longirostral Euthecodon (from the Mio-Pliocene of East
Africa) and the brevirostral extant genus Osteolaemus.
One of our predictions suggests that Crocodyloidea may
undergo geographical range shifts to track favourable ecological
conditions under climatic change (see above). Although we are
unable to test this prediction, we can check whether a relationship
exists between home range and the geometric variation of skull
rostra, controlling for phylogeny and body size (two factors
known to influence home range). The results do not contradict
our hypothesis. ‘Pure’ skull rostrum shape explains a significant
portion of home range variation in both Crocodyloidea and
Alligatoroidea, and the portion of variation explained in the first
group is higher than that explained in the second one (Fig. 7).
Therefore, we predict that global climatic change will have a
more important effect on Crocodyloidea (promoting trait
shift, adaptation to a new diet, and speciation, and modifying the

P. Piras et al.
© 2008 The Authors38 Global Ecology and Biogeography, 18, 30–40, Journal compilation © 2008 Blackwell Publishing Ltd
geographical range distribution of species, which may track
favourable ecological conditions) than in Alligatoroidea.
The evolutionary history of the clade Alligatoroidea is marked
by its tightness to the ecosystems, as a steneotopic organism
adapted to a narrow range of environmental conditions,
associated with continental freshwater ecosystems. Lingual salt
glands capable of excreting hyperosmotic sodium chloride (salt
water) are absent from the alligatorids (A. mississippiensis and
C. crocodylus) (Taplin & Grigg, 1989). This imposes a physiological
constraint in the dispersal capabilities of Alligatoroidea, and thus
the disjoint distribution of A. sinensis and A. mississippiensis
could be better explained in terms of habitat fidelity theory
(Vrba, 1995) than by dispersal (see Brochu, 2003, for the dispersive
hypothesis), considering fragmentation and drift of their ecosystems
during the Oligocene–Miocene temperate climate at high
latitude in North America and Asia (see Markwick, 1998a). On
the contrary, Crocodylinae are better adapted through a suite of
morphological specializations for life in hyperosmotic environments
(Jackson et al., 1996). Salinity discrimination in Crocodylinae
has been used to explain the distribution of living crocodilians by
means of their dispersive capability. Furthermore, speciation
phenomena in Crocodylinae have also been associated with the
increase in variation of rostral shapes (see Tchernov, 1986, for
the area of Great Rudolf Basin where Crocodylus lloidi, Crocodylus
niloticus, Crocodylus cataphractus and Euthecodon coexisted
during the Pliocene–Pleistocene time interval).
The main effect of climatic change has been the increase
in mean annual temperatures registered in recent years. For
ectothermic animals like crocodiles and alligators, global
warming has important effects on metabolic rates. These effects
are mediated by body mass, because both the surface to volume
ratio and the caloric transmission per mass unit decrease as body
mass increases (in geometrically similar animals, while the surface
is proportional to length raised to the second power, volume and
mass are proportional to length raised to the third power). Thus
global warming may promote mass homeothermy in species of
large body size: although these animals generate little physiological
heat (compared with endotherms), high external temperatures
during daytime and the low surface to volume ratio (allowing
retention of much heat at night) promotes the conservation of a
roughly constant temperature. Mass homeothermy (linked to
large size) may increase fitness because it allows high growth
rates, which are energetically costly because of high protein turn-
over (Montes et al., 2007) and a shortening of the juvenile period
of growth (during which individuals undergo high predation
pressure). In conclusion, we expect that global warming may
promote an increase in species-specific body size because both
variables contribute to reaching mass homeothermy, which may
increase the fitness both by increasing growth rates and by
shortening the juvenile period of growth. However, what is the
evolutionary plasticity of maximal body size in the clades
studied? Variation partitioning analyses of maximum skull size
(as an indicator of maximum body mass), using the phylogenetic
PCs as a single set of independent variables, show that the
fraction of variance independent of phylogeny is higher in
Crocodyloidea than in Alligatoroidea. These results strongly
suggest that maximum body mass is evolutionarily more
plastic in the first clade than in the second one (in other words,
Crocodyloidea may appear more reactive to selection for changing
body mass than Alligatoroidea). These results agree with those
obtained in variation partitioning of rostrum shape, suggesting
that Crocodyloidea may track potential environmental changes.
Extant alligatoroid taxa exhibit rather similar rostral shape
and size; most retain a ‘generalist’ rostrum (Brochu, 2001). When
body size notably increased along the evolutionary history of
Alligatoroidea (which includes the giant genus Purrusaurus from
the Miocene of South America, a member of the alligatoroid clade
Nettosuchidae), selection favoured changes in the rostrum width
towards a duck-shaped form. This also occurs in the extant
Melanosuchus. However, the most conspicuous evolutionary
trend within the clade is dwarfism. Cenozoic North American and
European genera such as Allognathosuchus, Arambourgia, Baryphracta,
among others show miniaturized skulls, transforming their rostrum
to what has been denominated ‘blunt snouts’ (see also Brochu,
2001). None of the extant taxa bear such a skull configuration.
Considering that mostly extant species were analysed in this
study (only three fossil Crocodyloidea were included in the
sample), it could be argued that recent diversity of crocodilians
is relict in comparison to the past (Marcwick, 1994a,b, 1995,
1998a,b; Brochu, 2003; Kotsakis et al., 2004). However, as the
aim of the paper is to predict how two contemporaneously living
groups (Alligatoroidea and Crocodyloidea) may respond to an
environmental change, the use of a sample containing mainly
extant species is not a limit but a necessary condition. Moreover,
a greater taxonomic diversity does not necessarily imply a greater
morphological disparity. Nevertheless, looking at the fossil
record of Crocodyloidea and Alligatoroidea helps to understand
their past diversity and disparity (some qualitative considerations
have been made here, see above).
It should be kept in mind that our predictions about the
possible effects of global environmental changes on the biodiversity
(species richness and morphological disparity) of Alligatoridae
and Crocodylidae are macroevolutionary and more linked to
climatic change than to quick changes derived from human
activity such as habitat loss, illegal trade, etc. Data on life-history
traits of Alligatoridae and Crocodylidae, such as life span, age at
sexual maturity and offspring size, among others, may be necessary
to make predictions on how these animals may respond to
anthropic environmental changes.
ACKNOWLEDGEMENTS
Paolo Piras had access to the collections of the Natural History
Museum in London, to the Institut Royal des Sciences Naturelles
in Bruxelles and to the Museum d’Histoire Naturelle in Paris
thanks to the SINTHESYS program, and to the collection of the
Field Museum in Chicago thanks to the Visiting Scholarship Pro-
gram. Part of this research was conducted during a post-doctoral
stay of Paolo Piras, supervised by Jorge Cubo, at the Université
Pierre et Marie Curie. We thank Professor Diniz-Filho, two
anonymous referees and Michel Laurin for their comments.
Michel Laurin also improved the English of the manuscript.

Phylogenetic effects and climatic change
© 2008 The Authors Global Ecology and Biogeography, 18, 30–40, Journal compilation © 2008 Blackwell Publishing Ltd 39
REFERENCES
Anderson, M.J. & Gribble, N.A. (1998) Partitioning the variation
among spatial, temporal and environmental components in a
multivariate dataset. Australian Journal of Ecology, 23, 158–
167.
Aoki, R. (1983) A new generic allocation of Tomistoma
machikanense, a fossil crocodilian from the Pleistocene of
Japan. Copeia, 1983, 89–95.
Arnold, S.J. (1983) Morphology, performance, and fitness.
American Zoologist, 23, 347–361.
Bookstein, F.L. (1991) Morphometric tools for landmark data.
Geometry and biology. Cambridge University Press, New York.
Borcard, D. & Legendre, P. (1994) Environmental control and
spatial structure in ecological communities: an example using
oribatid mites (Acari, Oribatei). Environmental and Ecological
Statistics, 1, 37–61.
Borcard, D., Legendre, P. & Drapeau, P. (1992) Partialling out the
spatial component of ecological variation. Ecology, 73, 1045–1055.
Brochu, C.A. (1999) Phylogeny, systematics, and historical
biogeography of Alligatoroidea. Society of Vertebrate Paleontology
Memoirs, 6, 9–100.
Brochu, C.A. (2000) Phylogenetic relationships and divergence
timing of Crocodylus based on morphology and the fossil
record. Copeia, 2000, 657–673.
Brochu, C.A. (2001) Crocodilian snouts in space and time:
phylogenetic approaches toward adaptive radiation. American
Zoologist, 41, 564–585.
Brochu, C.A. (2003) Phylogenetic approaches toward crocodilian
history. Annual Review of Earth and Planetary Science, 31, 357–397.
Cubo, J. (2004) Pattern and process in constructional morphology.
Evolution and Development, 6, 131–133.
Cubo, J., Ponton, F., Laurin, M., de Margerie, E. & Castanet, J.
(2005) Phylogenetic signal in bone microstructure of Sauropsids.
Systematic Biology, 54, 562–574.
Cubo, J., Legendre, P., de Ricqlès, A., Montes, L., de Margerie, E.,
Castanet, J. & Desdevises Y. (2008) Phylogenetic, functional,
and structural components of variation in bone growth rate of
amniotes. Evolution and Development, 10, 217–227.
Cushman, S.A. & McGarigal, K. (2002) Hierarchical, multiscale
decomposition of species–environment relationships.
Landscape Ecology, 17, 637–646.
Desdevises, Y., Legendre, P., Azouzi, L. & Morand, S. (2003)
Quantifying phylogenetically structured environmental
variation. Evolution, 57, 2647–2652.
Diniz-Filho, J.A.F. & Bini, L.M. (2008) Macroecology, global
change and the shadow of forgotten ancestors. Global Ecology
and Biogeography, 17, 11–17.
Diniz-Filho, J.A.F., de Sant’ Ana, C.E.R. & Bini, L.M. (1998) An
eigenvector method for estimating phylogenetic inertia.
Evolution, 52, 1247–1262.
Erickson, G.M., Lappin, A.K. & Vliet, K. (2003) The ontogeny of
bite-force performance in American alligator (Alligator
mississippiensis). Journal of Zoology, 260, 317–327.
Erickson, G., Lappin, A.K., Vliet, K., Brueggen, J., Kledzik, D. &
Webb, G. (2005) Comparative bite-force performance of extant
and fossil crocodilians: a window into evolutionary feeding
biomechanics. Journal of Vertebrate Paleontology, 26A, 46.
Felsenstein, J. (1985) Phylogenies and the comparative method.
The American Naturalist, 125, 1–15.
Garland, T. Jr, Harvey, P.H. & Ives, A.R. (1992) Procedures for
the analysis of comparative data using phylogenetically
independent contrasts. Systematic Biology, 41, 18–32.
Harvey, P.H. & Pagel, M.D. (1991) The comparative method in
evolutionary biology. Oxford University Press, Oxford, UK.
Iocchi, L. & Piras, P. (2006) Jcocco 1.0. http://host.uniroma3.it/
laboratori/paleontologia/piras_vert.htm
Jackson, K., Butler, D.G. & Brooks, D.R. (1996) Habitat and
phylogeny influence salinity discrimination in crocodilians:
implications for osmoregulatory physiology and historical
biogeography. Biological Journal of the Linnean Society, 58,
371–383.
Janke, A., Gullberg, A., Hughes, S., Aggarwal, R.K. & Arnason, U.
(2006) Mitogenomic analyses place the gharial (Gavialis
gangeticus) on the crocodile tree and provide pre-K/T
divergence times for most crocodilians. Journal of Molecular
Evolution, 61, 620–626.
Josse, S., Moreau, T. & Laurin, M. (2006) Stratigraphic tools for
Mesquite. http://mesquiteproject.org/packages/stratigraphicTools/
Kingsolver, J.G. & Huey, R.B. (2003) Introduction: The evolution
of morphology, performance, and fitness. Integrative Com-
parative Biology, 43, 361–366.
Kotsakis T., Delfino M. & Piras P. (2004) Italian Cenozoic
crocodilians: taxa, timing and palaeobiogeographic implications.
Palaeogeography, Palaeoclimatology, Palaeoecology, 210, 67–87.
Legendre, P. & Legendre, L. (1998) Numerical ecology, 2nd Engl.
edn. Elsevier Science BV, Amsterdam.
McAliley, L.R., Willis, R.E., Ray, D.A., Scott White, P., Brochu, C.A.
& Densmore, L.D. III (2006) Are crocodiles really monophyletic?
Evidence for subdivisions from sequence and morphological
data. Molecular Phylogenetics and Evolution, 39: 16–32.
McGowan, C. (1999) A practical guide to vertebrate mechanics.
Cambridge University Press, New York, USA.
Maddison, W.P. & Maddison, D.R. (2007) Mesquite: a modular system
for evolutionary analysis, version 2.01. http://mesquiteproject.org
Markwick, P.J. (1994a) Crocodilian distribution and diversity
across the Cretaceous–Tertiary boundary. Implications for
climatically induced extinction. Geological Society of America,
Abstracts with Programs, 26, 395.
Markwick, P.J. (1994b) Equability continentality and Tertiary
‘climate’: the crocodilian perspective. Geology, 22, 613–616.
Markwick, P.J. (1995) A new method for examining paleoclimatology
and paleoecology using present day faunas assigned to climate
stations. Geol. Soc. Am. Meeting, New Orleans.
Markwick, P.J. (1998a) Crocodilian diversity in space and time:
the role of climate in paleoecology and its implication for
understanding K/T extinctions. Paleobiology, 24, 470–497.
Markwick, P.J. (1998b) Fossil crocodilians as indicators of Late
Cretaceous and Cenozoic climates: implications for using
palaeontological data in reconstructing palaeoclimate.
Palaeogeography, Palaeoclimatology, Palaeoecology, 137, 205–271.
Metzger, K.A., Daniel, W.J.T. & Ross, C.F. (2005) Comparison of

P. Piras et al.
© 2008 The Authors40 Global Ecology and Biogeography, 18, 30–40, Journal compilation © 2008 Blackwell Publishing Ltd
beam theory and finite-element analysis with in vivo bone
strain data from the Alligator cranium. The Anatomical Record
Part A, 283A, 331–348.
Montes, L., Le Roy, N., Perret, M., de Buffrénil, V., Castanet, J. &
Cubo, J. (2007) Relationship between bone growth rate, body
mass and resting metabolic rate in growing amniotes: a phylogenetic
approach. Biological Journal of the Linnean Society, 92: 63–76.
O’Higgins, P. & Jones, N. (2007) Morphologika 2.5. http://
hyms.fme.googlepages.com/downloadmorphologica
Økland, R.H. (2003) Partitioning the variation in a plot-by species
data matrix that is related to n sets of explanatory variables.
Journal Vegetation Science, 14, 693–700.
Rohlf, F.J. & Slice, D.E. (1990) Extensions of the Procrustes
method for the optimal superimposition of landmarks.
Systematic Zoology, 39, 40–59.
Sheets, H.D. (2006) IMP software package. http://www3.canisius.
edu/~sheets/moremorph.html
Taplin, L.E. & Grigg, G.C. (1989) Historical zoogeography of the
eusuchian crocodilians: a physiological perspective. American
Zoologist, 29, 885–901.
Tchernov, E. (1986) Evolution of the crocodiles in East and North
Africa. CNRS, Cahiers de Paléontologie, Travaux de Paléonto-
logie Est-Africaine, Paris.
Vrba, E. (1995) Species as habitat-specific, complex systems.
Speciation and the recognition concept (ed. by D.M. Lambert and
H.G. Spencer), pp. 3–44. Johns Hopkins University Press,
Baltimore, MD.
SUPPORTING INFORMATION
Additional Supporting Information may be found in the online
version of this article:
Appendix S1 List of materials used in this study.
Appendix S2 Beam theory computation detail.
Please note: Wiley-Blackwell is not responsible for the content
or functionality of any supporting materials supplied by the
authors. Any queries (other than missing material) should be
directed to the corresponding author for the article.
Editor: José Alexandre F. Diniz-Filho
BIOSKETCHES
Paolo Piras is a post-doctoral researcher at Université
Pierre et Marie Curie, Paris. He is interested in
macroevolutianry process, and is involved in the study
of Eusuchian systematics and evolution, as well as the
evolution of Plio-Pleistoicene faunas of Europe.
Luciano Teresi earned a PhD in theoretical and applied
mechanics at Sapienza, Università di Roma (Italy) and
is now an assistant professor at the School of Engineering,
Università Roma Tre, Italy. Main researches include
finite element methods in nonlinear elasticity, biomechanics
of growth and electro-mechanical modeling of the heart.
Angela Delgado-Buscalioni is assistant professor at
the Universidad Autónoma de Madrid. She is interested
on skull geometry, integrating neontological and
palaeobiological data, as well as in early taphonomic
alterations in recent and fossil archosaurs.
Jorge Cubo is Maitre de Conferences at the Université Pierre
et Marie Curie, Paris. He is interested in the quantification
of phylogenetic, functional and structural components of
variation in morphological characters in tetrapods.




















