
The salamander species assemblage and environment of forested seeps of the Allegheny High Plateau,...
8
Introduction Seeps are shallow, slow-moving outflows of groundwater formed when an aquiclude, an impermeable rock or clay layer, prevents downward percolation of water to the water table, causing it to flow horizontally to the ground surface (Wunz et al., 1983; Welsh and Lind, 1996; Hall et al., 2001; Stein et al., 2004; Mississippi Museum of Natural Science, 2005; McIntyre et al., 2006). Sometimes classified as slope wetlands (e.g., Stein et al., 2004), seeps can occur in a range of landforms but in many regions they tend to be more prevalent in headwater stream systems with moderate to steep terrain (e.g., Mississippi Museum of Natural Science, 2005; McIntyre et al., 2006; Morley and Calhoun, 2009). Usually a minor component of headwater stream systems (e.g., O’Donnell et al., 2007), seeps provide an aquatic environment largely different from that of associated streams. For example, seeps tend to be thermally stable lotic habitats. Water temperature in seeps is usually maintained within a small range year- round, closely approximating the mean annual air temperature of a region, providing cool-water habitat in the summer and an ice-free channel of relatively warmer water during winter, particularly in temperate regions (Wunz et al., 1983; Healy and Casalena, 1996). Likewise, seep flow is usually perennial, augmenting low summer flows common in some temperate headwater streams (e.g., Snodgrass et al., 2007) and providing a water resource for plants and animals during drought (McIntyre et al., 2006). Finally, seeps can be sources of minerals and nutrients, particularly cations like Ca 2+ and Mg 2+ , that can influence the productivity of in-channel and seep-side plant and animal communities while also moderating headwater stream chemistry, particularly pH (Murdoch and Stoddard, 1993; Hall et al., 2001). Seeps provide habitat for a variety of wetland plant species and aquatic and semi-terrestrial invertebrate and Herpetology Notes, volume 8: 99-106 (2015) (published online on 10 April 2015) The salamander species assemblage and environment of forested seeps of the Allegheny High Plateau, Northwestern Pennsylvania, USA Charles E. Williams Williams Ecological, LLC, 103 Hillcrest Lane, Shippenville, PA 16254 Correspondence: [email protected] Abstract. Seeps are shallow, slow-moving outflows of groundwater that can provide habitat and foraging areas for salamanders and other amphibians. Seeps are a largely unstudied component of slope landforms in the Allegheny High Plateau Ecoregion of northwestern Pennsylvania, USA, particularly in regard to their salamander fauna. I conducted a survey of forested seeps (n = 30) in the Allegheny National Forest of northwestern Pennsylvania in order to: 1) identify the salamander species assemblage associated with Allegheny High Plateau seeps; and 2) examine the relationship of the abundance of dominant salamander species, represented as catch-per-unit-effort (CPUE), with key physical and biotic habitat components of the seep environment. Five species of salamanders were encountered during time-constrained surveys of seeps including the northern dusky salamander, Desmognathus fuscus (Rafinesque, 1820), the Allegheny Mountain dusky salamander, D. ochrophaeus Cope, 1859, the northern spring salamander, Gyrinophilus porphyriticus (Green, 1827), the redbacked salamander, Plethodon cinereus (Green, 1818), and the northern two-lined salamander, Eurycea bislineata (Green, 1818). CPUE of post-metamorphs for each of the two dominant salamander species, D. fuscus and D. ochrophaeus, was significantly associated with a small but distinct subset of seep habitat variables: CPUE for D. fuscus was significantly and positively associated with rock cover and pH, whereas CPUE for D. ochrophaeus was significantly and positively associated with vascular plant cover and slope. Keywords: Allegheny High Plateau, Allegheny National Forest, Pennsylvania, salamanders, seeps, USA
-
Upload
columbiasouthern -
Category
Documents
-
view
1 -
download
0
Transcript of The salamander species assemblage and environment of forested seeps of the Allegheny High Plateau,...

Introduction
Seeps are shallow, slow-moving outflows of groundwater formed when an aquiclude, an impermeable rock or clay layer, prevents downward percolation of water to the water table, causing it to flow horizontally to the ground surface (Wunz et al., 1983; Welsh and Lind, 1996; Hall et al., 2001; Stein et al., 2004; Mississippi Museum of Natural Science, 2005; McIntyre et al., 2006). Sometimes classified as slope wetlands (e.g., Stein et al., 2004), seeps can occur in a range of landforms but in many regions they tend to be more prevalent in headwater stream systems with moderate to steep terrain (e.g., Mississippi Museum of Natural Science, 2005; McIntyre et al., 2006; Morley and Calhoun, 2009).
Usually a minor component of headwater stream systems (e.g., O’Donnell et al., 2007), seeps provide an aquatic environment largely different from that of associated streams. For example, seeps tend to be thermally stable lotic habitats. Water temperature in seeps is usually maintained within a small range year-round, closely approximating the mean annual air temperature of a region, providing cool-water habitat in the summer and an ice-free channel of relatively warmer water during winter, particularly in temperate regions (Wunz et al., 1983; Healy and Casalena, 1996). Likewise, seep flow is usually perennial, augmenting low summer flows common in some temperate headwater streams (e.g., Snodgrass et al., 2007) and providing a water resource for plants and animals during drought (McIntyre et al., 2006). Finally, seeps can be sources of minerals and nutrients, particularly cations like Ca2+ and Mg2+, that can influence the productivity of in-channel and seep-side plant and animal communities while also moderating headwater stream chemistry, particularly pH (Murdoch and Stoddard, 1993; Hall et al., 2001).
Seeps provide habitat for a variety of wetland plant species and aquatic and semi-terrestrial invertebrate and
Herpetology Notes, volume 8: 99-106 (2015) (published online on 10 April 2015)
The salamander species assemblage and environment of forested seeps of the Allegheny High Plateau,
Northwestern Pennsylvania, USA
Charles E. Williams
Williams Ecological, LLC, 103 Hillcrest Lane, Shippenville, PA 16254
Correspondence: [email protected]
Abstract. Seeps are shallow, slow-moving outflows of groundwater that can provide habitat and foraging areas for salamanders and other amphibians. Seeps are a largely unstudied component of slope landforms in the Allegheny High Plateau Ecoregion of northwestern Pennsylvania, USA, particularly in regard to their salamander fauna. I conducted a survey of forested seeps (n = 30) in the Allegheny National Forest of northwestern Pennsylvania in order to: 1) identify the salamander species assemblage associated with Allegheny High Plateau seeps; and 2) examine the relationship of the abundance of dominant salamander species, represented as catch-per-unit-effort (CPUE), with key physical and biotic habitat components of the seep environment. Five species of salamanders were encountered during time-constrained surveys of seeps including the northern dusky salamander, Desmognathus fuscus (Rafinesque, 1820), the Allegheny Mountain dusky salamander, D. ochrophaeus Cope, 1859, the northern spring salamander, Gyrinophilus porphyriticus (Green, 1827), the redbacked salamander, Plethodon cinereus (Green, 1818), and the northern two-lined salamander, Eurycea bislineata (Green, 1818). CPUE of post-metamorphs for each of the two dominant salamander species, D. fuscus and D. ochrophaeus, was significantly associated with a small but distinct subset of seep habitat variables: CPUE for D. fuscus was significantly and positively associated with rock cover and pH, whereas CPUE for D. ochrophaeus was significantly and positively associated with vascular plant cover and slope.
Keywords: Allegheny High Plateau, Allegheny National Forest, Pennsylvania, salamanders, seeps, USA

Charles E. Williams100
vertebrate species, including species of conservation interest (Fike, 1999; Hall et al., 2001; Stein et al., 2004; Miller et al., 2005; Mitchell et al., 2006; Morley and Calhoun, 2009; Hager et al., 2012). For terrestrial vertebrates such as eastern wild turkey (Meleagris gallopavo) and white-tailed deer (Odocoileus virginianus), seeps can be important foraging areas, especially in winter when they are generally snow-free (Wunz et al., 1983; Healy and Casalena, 1996). Many amphibian species are associated with seeps in North America (Mississippi Museum of Natural Science, 2005; O’Donnell et al., 2007), and they can be key habitats for salamander species that require clean, cold water and a fish-free environment to thrive (Mitchell et al., 2006). For example, the northern spring salamander, Gyrinophilus porphyriticus (Green, 1827), often reaches its greatest abundance in cold-water seeps, springs, and
headwater streams where predatory fish are absent (Petranka, 1998; Lowe and Bolger, 2002).
On the Allegheny High Plateau Ecoregion of northwestern Pennsylvania, USA, seeps have a patchy occurrence within and across headwater stream systems and are usually associated with mid to lower slope landforms dominated by colluvial soils (Figure 1; Williams et al., 2005). Allegheny High Plateau seeps have received little ecological study, particularly the species composition and habitat relations of the in-channel salamander fauna. The only detailed study of regional seeps (Wunz et al., 1983) noted the presence of Desmognathus spp. or dusky salamanders; however, information on species composition or abundance was not collected. To provide basic information on the seep-inhabiting salamander fauna of the region, I conducted a survey of forested seeps in the Allegheny National Forest of northwestern Pennsylvania. The goals of the survey were to: 1) identify the salamander species assemblage associated with Allegheny High Plateau seeps; and 2) examine the relationship of the abundance of dominant salamander species, represented as catch-per-unit-effort, with key physical and biotic habitat components of the seep environment.
Material and Methods
Study area. The 200,000+ ha Allegheny National Forest (ANF) (41° 45' N, 79° 00' W) is located within the non-glaciated Allegheny High Plateau Ecoregion of northwestern Pennsylvania (Figure 2). The ANF occurs in a transition zone between the eastern hemlock-white pine-northern hardwood forest and Appalachian oak forest types (Braun, 1950). The landscape is typified by relatively flat to gently rolling plateaus dissected by deeply eroded, dendritic stream valleys (Hough and Forbes, 1943). Plateau elevations range from 500 to 700 m above sea level (asl); stream valley floors generally range from 300 to 400 m asl (Whitney, 1990). Summers are cool with an average temperature of 18.9°C and precipitation is distributed evenly throughout the year, averaging between 100 and 110 cm (Cerutti, 1985). Forty-four seep study sites were located in the Allegheny National Forest from 2000 to 2001, largely using vehicular “windshield surveys” conducted in winter when seeps are snow-free and visibly apparent. Thirty seeps were randomly selected for salamander surveys, which were conducted from June through August 2002. All study sites were located in second-growth forest with a tree overstory (stems > 10 cm dbh) of approximately 60 to 80 years in age. Common
Figure 1. Typical seeps of the Allegheny High Plateau, McKean County, Pennsylvania, USA. A = Well-vegetated seep channel; B = Sparsely-vegetated seep channel.

overstory species at study sites included eastern hemlock (Tsuga canadensis), red maple (Acer rubrum), sugar maple (A. saccharum), yellow birch (Betula alleghaniensis), white ash (Fraxinus americana), northern red oak (Quercus rubra) and basswood (Tilia americana). Dominant herbaceous plants in seeps were largely of wetland affinity and included touch-me-not (Impatiens capensis), common blue violet (Viola sororia), slender mannagrass (Glyceria melicaria), foamflower (Tiarella cordifolia), dwarf enchanter’s nightshade (Circaea alpina) and mountain watercress (Cardamine rotundifolia). Shrubs were generally absent from the study sites. Scientific and common names of plants follow Rhoads and Block (2007).Sampling strategy and data analyses. At each study site, a 20-m transect was established in the middle of, and parallel to, the seep channel. Transect ends were permanently marked with iron rebar and seep location was geo-referenced with a hand-held GPS unit. Channel width and pH were measured at the top, middle, and bottom of each seep transect. Slope (%) and aspect (degrees N of S) were measured with a clinometer and compass at the middle of each transect.
Prior to salamander surveys, percent cover of in-channel habitat components (vascular plants, bryophytes, rock, and leaf litter) was visually estimated in each of ten 1-m2 plots located in a stratified random manner along each seep transect. All large woody debris (LWD; logs > 10.0 cm basal diameter and > 1 m in length) located within the area delineated by the length of the transect line and the upper and lower channel widths of each
seep study site was tallied and measured (total length of logs within the seep channel and upper and lower log diameters).
Salamanders were sampled at each seep study site using time-constrained touch searches (Moore et al., 2000, 2001), during which moveable cover items within the channel were turned over or dug into by hand and examined for salamanders, which were captured and placed in holding containers for identification. Touch searches have been found to be an effective and repeatable methodology for sampling the salamander fauna within seeps (O’Donnell et al., 2007). Three 10-minute searches were conducted at each seep study site by one or two trained persons for a total of 30 minutes search per site. Escaped salamanders that could be identified were included in final species tallies. Cover items were returned to their original positions, and salamanders were released as closely as possible to their capture sites, when searches were completed. Captured salamanders were categorized as either larvae (having visible gills) or post-metamorphs. Due to time constraints caused by concurrent projects at the study sites, neither size nor mass of captured salamanders were measured during this study.
Salamander captures were converted to catch-per-unit effort (CPUE per hour) by species for analysis. LWD measurements were converted to total volume using the equation of Lienkamper and Swanson (1986). Mean LWD volume (m3) and density were expressed on a 100-m basis (Williams and Cook, 2010). Values for pH were converted to hydrogen ion concentration to minimize
Salamander species assemblage and environment of forested seeps, USA 101
Figure 2. Location of the Allegheny National Forest, northwestern Pennsylvania, USA.
errors associated with standard pH transformation (Murphy, 1982). Data other than pH were transformed prior to analysis to ensure normality [cos(45 - aspect) + 1 for aspect data; square root (x + 0.5) for count data; arcsin transformation for percentage data; log(x + 0.5) for other continuous data] and back-transformed for tabular presentation (Beers et al., 1966; Zar, 1996).
Stepwise multiple regression was used to identify a predictive multivariable model of CPUE of dominant salamander species with the ten physical and biotic habitat variables measured at each seep (Table 1; Zar, 1996). Stepwise multiple regression was done using the backward option of SYSTAT version 7.0, with P to enter and remove habitat variables set at 0.10 (Wilkinson, 1997). Prior to multiple regression analysis, multicollinearity among habitat variables was assessed
using pairwise Pearson correlations (Lowe and Bolger, 2002) and by variance inflation factors (VIF) calculated for each habitat variable (O’Brien, 2007; Robinson and Schumacker, 2009; Evangelista et al., 2011). Correlation analysis was done using SYSTAT version 7.0 and VIFs were calculated using NCSS version 8.0 (Hintze, 2014) with a threshold of 10. Low r-values and non-significant correlations (P > 0.05) as well as VIFs less than 10 for all habitat variables, suggested that multicollinearity was not a major concern, thus all variables were retained for multiple regression analysis.
Results
The in-channel seep environment showed a range of variation in physical and biotic habitat components
1
Table 1. Summary of physical and biotic habitat variables measured across Allegheny
High Plateau seeps (n = 30) in the Allegheny National Forest, northwestern
Pennsylvania, USA. LWD = large wood debris (logs > 10.0 cm basal diameter
and > 1 m in length).
Habitat variables by category Mean + 1 SE Range Physical variables
Aspect (degrees N of S) 163.2 + 18.6 6.0 – 354.0 Slope (%) 21.7 + 2.2 4.0 – 45.0 Channel width (m) 5.3 + 0.5 1.7 – 10.1 pH 7.01 + 0.11 4.93 – 7.96 Rock cover (%) 22.2 + 3.3 4.7 – 65.5
Biotic variables Leaf litter cover (%) 20.2 + 2.2 4.7 – 59.0 Bryophyte cover (%) 23.2 + 1.8 2.5 – 43.5 Vascular plant cover (%) 24.4 + 4.3 0.1 – 43.5 LWD volume (m3/100-m) 11.3 + 2.1 0 – 5.5 LWD density (no. pieces/100-m) 1.4 + 0.3 0 – 45.0
Table 1. Summary of physical and biotic habitat variables measured across Allegheny High Plateau seeps (n = 30) in the Allegheny National Forest, northwestern Pennsylvania, USA. LWD = large wood debris (logs > 10.0 cm basal diameter and > 1 m in length).
2
Table 2. Summary of salamander species encountered (n = 257 individuals) during time-
constrained surveys of Allegheny High Plateau seeps (n = 30) in the Allegheny National
Forest, northwestern Pennsylvania, USA. The majority of Gyrinophilus porphyriticus
individuals captured were larvae (n = 19; 79.2% of captures for the species); no larval
salamanders of other species were encountered during surveys. % captured = number of
individuals of a species captured divided by the total individual salamanders captured
during the study; no. seeps present = total number of seeps where a species was
encountered during time-constrained searches; % occurrence = proportion of surveyed
seeps where a species was encountered.
Species No. captured % captured No. seeps present % occurrence Desmognathus ochrophaeus 117 45.5 24 80.0 D. fuscus 109 42.4 25 83.3 Gyrinophilus porphyriticus 24 9.3 11 36.7 Plethodon cinereus 6 2.3 5 16.7 Eurycea bislineata 1 0.4 1 3.3
Table 2. Summary of salamander species encountered (n = 257 individuals) during time-constrained surveys of Allegheny High Plateau seeps (n = 30) in the Allegheny National Forest, northwestern Pennsylvania, USA. The majority of Gyrinophilus porphyriticus individuals captured were larvae (n = 19; 79.2% of captures for the species); no larval salamanders of other species were encountered during surveys. % captured = number of individuals of a species captured divided by the total individual salamanders captured during the study; no. seeps present = total number of seeps where a species was encountered during time-constrained searches; % occurrence = proportion of surveyed seeps where a species was encountered.
Charles E. Williams102

across study sites (Table 1). Seeps occurred across a diversity of aspects from 6 to 354o N of S with a mean aspect (163.2o) that tended toward south-southeast. Percent slope of seep study sites ranged from gentle (4%) to relatively steep (45%) with a mean slope of 21.7%. Channel width ranged from 1.7 to 10.1 m with a mean of 5.3 m. The mean in-channel pH across study sites was circumneutral (7.01) but conditions ranged from acidic (4.93) to alkaline (7.96) (sensu USGS, no date). Two seep study sites had a pH of less than 6.0 (7.7% of the total) while 8 sites had pH values between 6.4 and 6.97 (26.7% of the total). The majority of study sites (20 sites; 66.7% of the total) had in-channel pH values of 7.0 or greater. Cover items (i.e., rock, leaf litter, vascular plants and bryophytes) exhibited similar mean values across study sites, near 20% cover. Overall,
LWD was sparse across study sites in both mean density and volume (Table 1).
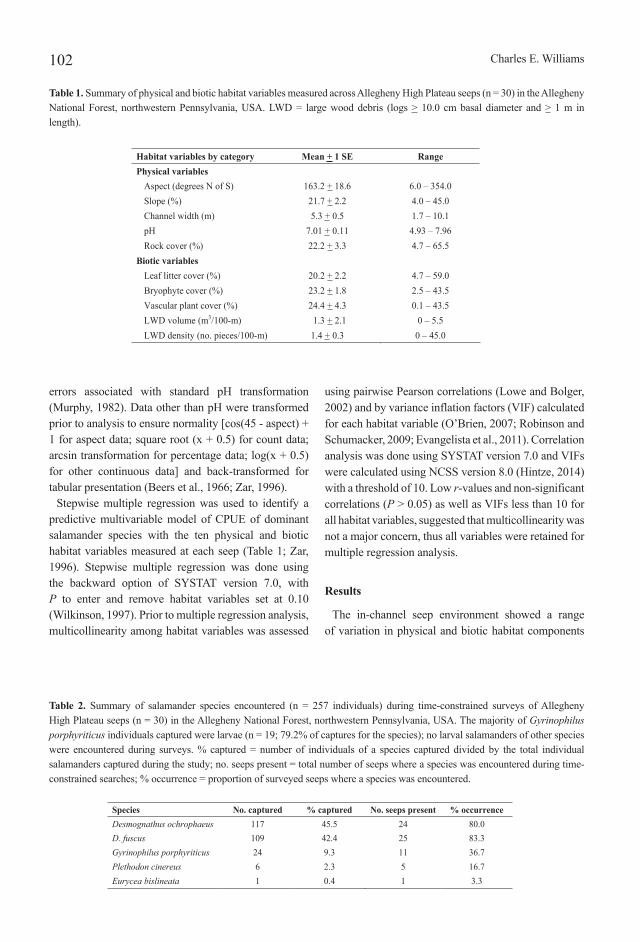
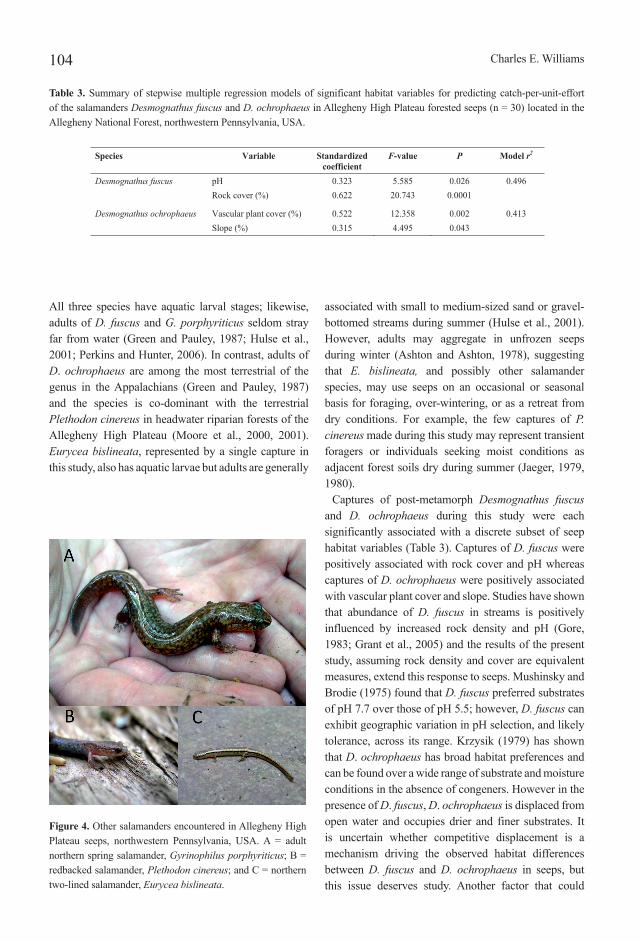
Two hundred-fifty-seven salamanders of five species were encountered during in-channel surveys of the 30 Allegheny High Plateau seep study sites (Table 2). Two species were dominant in numbers captured and occurrence across seeps: the Allegheny Mountain dusky salamander, Desmognathus ochrophaeus Cope, 1859 (Figure 3A), and the northern dusky salamander, D. fuscus (Rafinesque, 1820) (Figure 3B). Together these two species accounted for 87.9% of total salamander captures across the study sites. The northern spring salamander, Gyrinophilus porphyriticus (Green, 1827) (Figure 4A), ranked third in total captures and occurrence across study sites. Most of the G. porphyriticus individuals captured across seeps were larvae (n = 19; 79.2% of captures for the species); no larval salamanders of any other species were encountered during surveys. This discrepancy in larval captures across salamander species may be an artifact of sampling methods (e.g., Grant et al., 2005). The redbacked salamander, Plethodon cinereus (Green, 1818) (Figure 4B), with six captures across study sites, and the northern two-lined salamander, Eurycea bislineata (Green, 1818) (Figure 4C), with a single capture, were uncommonly encountered during surveys compared to the other three salamander species. Overall, salamanders were captured at 29 of the 30 (96.7%) study sites during timed searches.
Stepwise multiple regression identified distinct predictive models of CPUE for the two dominant salamander species at seeps, Desmognathus fuscus and D. ochrophaeus, each retaining a small subset of the ten habitat variables measured at each study site (Table 3). The multiple regression model of CPUE for D. fuscus was significantly and positively associated with pH and rock cover, whereas CPUE for D. ochrophaeus was significantly and positively associated with vascular plant cover and slope. On the basis of larger standardized coefficients (Schroeder et al. 1986), rock cover and vascular plant cover had greater relative influence on predicting CPUE for D. fuscus and D. ochrophaeus, respectively, than did the companion variables of pH (D. fuscus) or slope (D. ochrophaeus).
Discussion
The seep-inhabiting salamander fauna of the Allegheny High Plateau of northwestern Pennsylvania appears to be largely dominated by three species: Desmognathus fuscus, D. ochrophaeus, and Gyrinophilus porphyriticus.
Figure 3. Dominant salamanders of Allegheny High Plateau seeps, northwestern Pennsylvania, USA. A = northern dusky salamander, Desmognathus fuscus; and B = Allegheny Mountain dusky salamander, Desmognathus ochrophaeus.
Salamander species assemblage and environment of forested seeps, USA 103
All three species have aquatic larval stages; likewise, adults of D. fuscus and G. porphyriticus seldom stray far from water (Green and Pauley, 1987; Hulse et al., 2001; Perkins and Hunter, 2006). In contrast, adults of D. ochrophaeus are among the most terrestrial of the genus in the Appalachians (Green and Pauley, 1987) and the species is co-dominant with the terrestrial Plethodon cinereus in headwater riparian forests of the Allegheny High Plateau (Moore et al., 2000, 2001). Eurycea bislineata, represented by a single capture in this study, also has aquatic larvae but adults are generally
associated with small to medium-sized sand or gravel-bottomed streams during summer (Hulse et al., 2001). However, adults may aggregate in unfrozen seeps during winter (Ashton and Ashton, 1978), suggesting that E. bislineata, and possibly other salamander species, may use seeps on an occasional or seasonal basis for foraging, over-wintering, or as a retreat from dry conditions. For example, the few captures of P. cinereus made during this study may represent transient foragers or individuals seeking moist conditions as adjacent forest soils dry during summer (Jaeger, 1979, 1980).
Captures of post-metamorph Desmognathus fuscus and D. ochrophaeus during this study were each significantly associated with a discrete subset of seep habitat variables (Table 3). Captures of D. fuscus were positively associated with rock cover and pH whereas captures of D. ochrophaeus were positively associated with vascular plant cover and slope. Studies have shown that abundance of D. fuscus in streams is positively influenced by increased rock density and pH (Gore, 1983; Grant et al., 2005) and the results of the present study, assuming rock density and cover are equivalent measures, extend this response to seeps. Mushinsky and Brodie (1975) found that D. fuscus preferred substrates of pH 7.7 over those of pH 5.5; however, D. fuscus can exhibit geographic variation in pH selection, and likely tolerance, across its range. Krzysik (1979) has shown that D. ochrophaeus has broad habitat preferences and can be found over a wide range of substrate and moisture conditions in the absence of congeners. However in the presence of D. fuscus, D. ochrophaeus is displaced from open water and occupies drier and finer substrates. It is uncertain whether competitive displacement is a mechanism driving the observed habitat differences between D. fuscus and D. ochrophaeus in seeps, but this issue deserves study. Another factor that could
Figure 4. Other salamanders encountered in Allegheny High Plateau seeps, northwestern Pennsylvania, USA. A = adult northern spring salamander, Gyrinophilus porphyriticus; B = redbacked salamander, Plethodon cinereus; and C = northern two-lined salamander, Eurycea bislineata.
Table 3. Summary of stepwise multiple regression models of significant habitat variables for predicting catch-per-unit-effort of the salamanders Desmognathus fuscus and D. ochrophaeus in Allegheny High Plateau forested seeps (n = 30) located in the Allegheny National Forest, northwestern Pennsylvania, USA.
1
Table 3. Summary of stepwise multiple regression models of significant habitat variables for predicting catch-per-unit-effort of the
salamanders Desmognathus fuscus and D. ochrophaeus in Allegheny High Plateau forested seeps (n = 30) located in the Allegheny National
Forest, northwestern Pennsylvania, USA.
Species Variable Standardized coefficient
F-value P Model r2
Desmognathus fuscus pH 0.323 5.585 0.026 0.496 Rock cover (%) 0.622 20.743 0.0001
Desmognathus ochrophaeus Vascular plant cover (%) 0.522 12.358 0.002 0.413 Slope (%) 0.315 4.495 0.043
Charles E. Williams104

influence abundance or differential habitat use by D. fuscus and D. ochrophaeus is predation by Gyrinophilus porphyriticus, an important predator of Desmognathus species in the Appalachians (Petranka, 1998). Beachy (1994) showed that predation by G. porphyriticus was the key factor influencing the structure of experimental stream salamander communities.
Habitat differences between Desmognathus fuscus and D. ochrophaeus in seeps may also have potential management implications. Thinning of the forest overstory, or “day-lighting”, is a management practice that is sometimes used to increase vegetation productivity in and around seeps to improve wildlife forage (Wunz et al., 1983; Healy and Casalena, 1996). The results of this study suggest that day-lighting could have unintended effects on seep salamander communities: increased vegetative cover may cause a shift toward D. ochrophaeus dominance in treated areas. A detailed study of salamander species responses to day-lighting of seeps is needed to test this hypothesis, especially in regard to the larger issues of interspecific competition and predation as discussed above.
Acknowledgments. Billy Moriarity, Tim Jeffcoat, and Heather Whittier assisted with seep surveys. This project was supported in part by a Challenge-Cost Share Agreement between the USDA Forest Service, Allegheny National Forest and Clarion University of Pennsylvania. This paper is dedicated to the memory of the late William J. “Billy” Moriarity who helped facilitate this and many other projects on the Allegheny National Forest.
References
Ashton, R.E., Jr, Ashton, P.S. (1978): Movements and winter behavior of Eurycea bislineata (Amphibia, Urodela, Plethodontidae). Journal of Herpetology 12: 295-298.
Beachy, C.K. (1994): Community ecology in streams: effects of two species of predatory salamanders on a prey species of salamander. Herpetologica 50: 129-136.
Beers, T.W., Dress, P.E., Wensel, L.C. (1966): Aspect transformation in site productivity research. Journal of Forestry 64: 691-692.
Braun, E.L. (1950): Deciduous Forests of Eastern North America. Philadelphia, The Blakiston Company.
Cerutti, J.R. (1985): Soil Survey of Warren and Forest Counties, Pennsylvania. Washington, DC, USDA Soil Conservation Service.
Evangelista, P.H., Kumar, S., Stohlgren, T.J., Young, N.E. (2011): Assessing forest vulnerability and the potential distribution of pine beetles under current and future climate scenarios in the Interior West of the US. Forest Ecology and Management 262: 307-316.
Fike, J. (1999): Terrestrial and Palustrine Plant Communities of Pennsylvania. Harrisburg, PA, Pennsylvania Department of Conservation and Natural Resources.
Gore, J.A. (1983): The distribution of desmognathine larvae (Amphibia: Plethodontidae) in coal surface mine impacted streams of the Cumberland Plateau, USA. Journal of Freshwater Ecology 2: 13-23.
Grant, E.H.C., Jung, R.E., Rice, K.E. (2005): Stream salamander species richness and abundance in relation to environmental factors in Shenandoah National Park, Virginia. American Midland Naturalist 153: 348-356.
Green, N.B., Pauley, T.K. (1987): Amphibians and Reptiles in West Virginia. Pittsburgh, PA, University of Pittsburgh Press.
Hager, B.J., Kalantari, N.J., Scholten, V.A. (2012): The distribution of Cordulegaster (Odonata: Cordulegastridae) nymphs in seeps and springs of Nelson Swamp (Madison County, NY). Northeastern Naturalist 19: 67-76.
Hall, B.R., Raynal, D.R., Leopold, D.J. (2001): Environmental influences on plant species composition in ground-water seeps in the Catskill Mountains of New York. Wetlands 21: 125-134.
Healy, W.M., Casalena, M.J. (1996): Spring Seep Management for Turkeys and Other Wildlife. Edgefield, SC, National Wild Turkey Federation Wildlife Bulletin No. 30.
Hintze, J. (2014): NCSS 8. Available at: http://www.ncss.com. Last accessed on 10 December 2014.
Hough, A.F., Forbes, R.D. (1943): The ecology and silvics of forests in the high plateaus of Pennsylvania. Ecological Monographs 13: 301–320.
Hulse, A.C., McCoy, C.J., Censky, E. (2001): Amphibians and Reptiles of Pennsylvania and the Northeast. Ithaca, NY, Cornell University Press.
Jaeger, R.G. (1979): Seasonal spatial distributions of the terrestrial salamander Plethodon cinereus. Herpetologica 35: 90-93.
Jaeger, R.G. (1980): Microhabitats of a terrestrial forest salamander. Copeia 1980: 265-268.
Krzysik, A.J. (1979): Resource allocation, coexistence, and the niche structure of a streambank salamander community. Ecological Monographs 49: 173-194.
Lienkamper, G.W., Swanson, F.J. (1986): Dynamics of large woody debris in streams in old-growth Douglas-fir forests. Canadian Journal of Forest Research 17: 150-156.
Lowe, W.H., Bolger, D.T. (2002): Local and landscape-scale predictors of salamander abundance in New Hampshire headwater streams. Conservation Biology 16: 189-193.
McIntyre, A.P., Schmitz, R.A., Crisafulli, C.M. (2006): Associations of Van Dyke’s salamander (Plethodon vandykei) with geomorphic conditions in headwall seeps of the Cascade Range, Washington State. Journal of Herpetology 40: 309-322.
Miller, N.G., Fryday, A.M., Hinds, J.W. (2005): Bryophytes and lichens of a calcium-rich spring seep isolated on the granitic terrain of Mt. Katahdin, Maine, U.S.A. Rhodora 107: 339-358.
Mississippi Museum of Natural Science. 2005. Mississippi’s Comprehensive Wildlife Conservation Strategy. Jackson, MS, Mississippi Department of Wildlife, Fisheries and Parks, Mississippi Museum of Natural Science.
Mitchell, J.C., Breisch, A.R., Buhlmann K.A. (2006): Habitat Management Guidelines for Amphibians and Reptiles of the Northeastern United States. Montgomery, AL, Partners in Amphibian and Reptile Conservation, Technical Publication HMG-3.
Salamander species assemblage and environment of forested seeps, USA 105

Accepted by Hendrik Müller
Moore, A.L., Williams, C.E., Martin, T.H., Moriarity, W.J. (2001): Influence of season, geomorphic surface and cover item on capture, size and weight of Desmognathus ochrophaeus and Plethodon cinereus in Allegheny Plateau riparian forests. American Midland Naturalist 145: 39-45.
Moore, A.L., Williams, C.E., Moriarity, W.J. (2000): Species composition of salamander assemblages in Allegheny Plateau riparian forests. Journal of the Pennsylvania Academy of Science 74: 24-28.
Morley, T.R., Calhoun, A.J.K. (2009): Vegetation characteristics of forested hillside seeps in eastern Maine, USA. Journal of the Torrey Botanical Society 136: 520-531.
Murdoch, P.S., Stoddard, J.L. (1993): Chemical characteristics and temporal trends in eight streams of the Catskill Mountains, New York. Water, Air, Soil Pollution 67: 367–395.
Murphy, M.R. (1982): Analyzing and presenting pH data. Journal of Dairy Science 65: 161-163.
Mushinsky, H.R., Brodie E.D., Jr. (1975): Selection of substrate pH by salamanders. American Midland Naturalist 93: 440-443.
O’Brien, R.M. (2007): A caution regarding rules of thumb for variance inflation factors. Quality & Quantity 41: 673-690
O’Donnell, R.P., Quinn, T., Hayes, M.P., Ryding, K.E. (2007): Comparison of three methods for surveying amphibians in forested seep habitats in Washington State. Northwest Science 81: 274-283.
Perkins, D.W., Hunter, M.L., Jr. (2006): Use of amphibians to define riparian zones of headwater streams. Canadian Journal of Forest Research 36: 2124-2130.
Petranka, J.W. (1998): Salamanders of the United States and Canada. Washington, DC, Smithsonian Institution Press.
Rhoads, A.F., Block, T.A. (2007): The Plants of Pennsylvania: An Illustrated Manual, 2nd edition. Philadelphia, PA, University of Pennsylvania Press.
Robinson, C., Schumacker, R.E. (2009): Interaction effects: centering, variance inflation factor, and interpretation issues. Multiple Linear Regression Viewpoints 35: 6-11.
Schroeder, L.D., Sqoquist, D.L., Stephan, P.E. (1986): Understanding Regression Analysis. London, Sage Publications.
Snodgrass, J.W., Forester, D.C., Lahti, M., Lehman, E. (2007): Dusky salamander (Desmognathus fuscus) nest-site selection over multiple spatial scales. Herpetologica 64: 441-449.
Stein, E.D., Mattson, M.., Fetscher, A.E., Halama K.J. (2004): Influence of geologic setting on slope wetland hydrodynamics. Wetlands 24: 244-260.
[USGS] United States Geological Survey. (No date) National Water Summary on Wetland Resources. U.S. Geological Survey Water-Supply Paper 2425. Available at: http://water.usgs.gov/nwsum/WSP2425/index.html. Last accessed on 12 December 2014.
Welsh, H.H., Jr., Lind, A.J. (1996): Habitat correlates of the Southern Torrent Salamander Rhyacotriton variegatus (Caudata: Rhyacotritonidae), in northwestern California. Journal of Herpetology 30: 385-398.
Whitney, G.G. (1990): The history and status of the hemlock-hardwood forest of the Allegheny Plateau. Journal of Ecology 78: 443–458.
Wilkinson, L. (1997): SYSTAT 7.0. Statistics. Chicago, SPSS, Inc.
Williams, C.E., Baumer, G.E., Moriarity, W.J. (2005): Ecological Land Type Mapping Units for the Allegheny National Forest. Warren, PA, Project report, Allegheny National Forest.
Williams, C.E., Cook, R.E. (2010): Forest seral stage and large woody debris abundance in an Allegheny High Plateau forest-stream system. Natural Areas Journal 30: 20-26.
Wunz, G.A., Hayden, A.H., Potts, R.R. (1983): Spring seep ecology and management. Transactions of the Northeast Fish and Wildlife Conference 40: 19-30.
Zar, J.H. (1996): Biostatistical Analysis, 3rd edition. Upper Saddle River, NJ, Prentice-Hall, Inc.
Charles E. Williams106




















