
Differential Shh, Bmp and Wnt gene expressions during craniofacial development in mice

The Role of the Sutures inBiomechanical Dynamic Simulation of aMacaque Cranial Finite Element Model:
Implications for the Evolution ofCraniofacial Form
QIAN WANG,1* SARAH A. WOOD,2 IAN R. GROSSE,2 CALLUM F. ROSS,3
URIEL ZAPATA,1 CRAIG D. BYRON,4 BARTH W. WRIGHT,5
AND DAVID S. STRAIT6
1Division of Basic Medical Sciences, Mercer University School of Medicine, Macon, Georgia2Department of Mechanical and Industrial Engineering, University of Massachusetts,
Amherst, Massachusetts3Department of Organismal Biology and Anatomy, University of Chicago, Chicago, Illinois
4Department of Biology, Mercer University, Macon, Georgia5Department of Anatomy, Kansas City University of Medicine and Biosciences,
Kansas City, Missouri6Department of Anthropology, University at Albany, Albany, New York
ABSTRACTThe global biomechanical impact of cranial sutures on the face and
cranium during dynamic conditions is not well understood. It is hypothe-sized that sutures act as energy absorbers protecting skulls subjected todynamic loads. This hypothesis predicts that sutures have a significantimpact on global patterns of strain and cranial structural stiffness whenanalyzed using dynamic simulations; and that this global impact is influ-enced by suture material properties. In a finite element model developedfrom a juvenile Rhesus macaque cranium, five different sets of suturematerial properties for the zygomaticotemporal sutures were tested. Thestatic and dynamic analyses produced similar results in terms of strainpatterns and reaction forces, indicating that the zygomaticotemporalsutures have limited impact on global skull mechanics regardless of load-ing design. Contrary to the functional hypothesis tested in this study, thezygomaticotemporal sutures did not absorb significant amounts of energyduring dynamic simulations regardless of loading speed. It is alterna-tively hypothesized that sutures are mechanically significant only insofaras they are weak points on the cranium that must be shielded fromunduly high stresses so as not to disrupt vitally important growth proc-esses. Thus, sutural and overall cranial form in some vertebrates may beoptimized to minimize or otherwise modulate sutural stress and strain.Anat Rec, 295:278–288, 2012. VC 2011 Wiley Periodicals, Inc.
Key words: vertebrate skulls; elastic properties; loading speed
Abbreviations: CBCT ¼ cone-beam computed tomography; FEA¼ finite element analysis; FEM ¼ finite element model; TMJ ¼temporomandibular joint.*Correspondence to: Dr. Qian Wang, Division of Basic Medical
Sciences, Mercer University School of Medicine, 1550 CollegeStreet, Macon, GA 31207. Fax: 478-301 5489.E-mail: [email protected]
Received 8 July 2011; Accepted 4 October 2011.
DOI 10.1002/ar.21532
Published online 20 December 2011 in Wiley Online Library(wileyonlinelibrary.com).
THE ANATOMICAL RECORD 295:278–288 (2012)
VVC 2011 WILEY PERIODICALS, INC.

Besides being important loci of craniofacial growth(Opperman, 2000), sutures are mechanically ‘‘imper-fecta’’ in an otherwise rigid body (a cranium), and theirmechanical impact has been often discussed and inves-tigated using various experimental and computer-aidedtechniques in both living and fossil species (Behrentset al., 1978; Oudhof and Van Doorenmaalen, 1983;Smith and Hylander, 1985; Herring and Mucci, 1991;Herring, 1993; Jaslow and Biewener, 1995; Raffertyand Herring, 1999; Herring and Rafferty, 2000; Herringand Teng, 2000; Rafferty et al., 2003; Rayfield, 2004,2005; Lieberman et al., 2004; Sun et al., 2004; Kupcziket al., 2007, 2009; Farke, 2008; Wang et al., 2008,2010a; Moazen et al., 2009a,b; Jasinoski et al., 2010a,b;Bright and Groning, 2011; Porro et al., 2011; Reedet al., 2011). Experimental studies (Behrents et al.,1978; Herring and Mucci, 1991; Herring and Teng,2000) have demonstrated that patent sutures affectstrains locally (i.e., near the suture), but practical con-siderations limit the ability of purely experimentalstudies to assess the global influence of these jointsacross the entire cranium. Recently, finite elementanalysis (FEA) has been used to examine the mechani-cal behavior of sutures because sutures can be modeledand assigned varying sets of material properties, andthe process of sutural fusion can be simulated (Jasi-noski et al., 2010a,b; Wang et al., 2010a).
Using static simulations corresponding to incisor,premolar, and molar maximal biting conditions, arecent sensitivity analysis of the mechanical effect ofsutures in FE models of a macaque cranium demon-strates that the presence of sutures does notprofoundly influence global strain patterns, regardlessof their material properties (Wang et al., 2010a). Morespecifically, strain patterns remained relatively unaf-fected away from the suture sites, and bite reactionforce was likewise barely affected. Thus, the biome-chanical significance of sutures in a global contextwould therefore appear to be limited. However, staticanalyses are insufficient to simulate the role suturesmay play under physiological dynamic loading condi-tions. It has been proposed that cranial sutures in
goats are able to absorb 16–100% of energy duringimpact (Jaslow, 1990), but in static analysis, suturesdo not have a high capacity to absorb energy, whichmay be a function of either their limited volume(Wang et al., 2010a), or model resolution and elementsize in in silico, or simplified morphology. Whether ornot sutures provide a significant strain-dampening orshock absorbing function under dynamic loading condi-tions, as suggested by Jaslow and Biewener (1995),has yet to be tested. In this study, a finite elementmodel (FEM) of a primate cranium was analyzedusing dynamic simulations to assess the effect of apair of sutures (zygomaticotemporal) on stress andstrain patterns associated with feeding, a habitualbehavior that has the potential to impose high and/orrepetitive loads on the craniofacial skeleton. It ishypothesized that sutures act as energy absorbers pro-tecting skulls subjected to dynamic loads. Thishypothesis predicts that sutures have a significantimpact on global patterns of strain and cranial struc-tural stiffness when analyzed using dynamicsimulations; and this global impact is influenced bysuture material properties.
MATERIALS AND METHODS
Model Creation
The FEM used in this study was generated followingthe Biomesh protocol described in publications byDumont, Grosse, and colleagues (Dumont and Herrel,2003; Dumont et al., 2005; Grosse et al., 2007; www.bio-mesh.org). The FEM was built from a juvenile Macacamulatta specimen (age 2.5 years, with full set of decidu-ous teeth and the first permanent molar) obtained fromthe Yerkes Primate Research Center (supported by NIHBase Grant RR00165; Fig. 1a). The specimen wasscanned using iCAT cone-beam computed tomography(CBCT) facility (Imaging Sciences International, Hat-filed, PA) at Indiana University. CBCT is a recentlydeveloped technology used to evaluate 3D bone morphol-ogy at high resolution (Cho, 2009). Each volumetricimage was obtained using iCAT CBCT for approximately
Fig. 1. (a) Three-dimensional solid model of a M. mulatta cranium.(b) Loading regime (cranium not scaled as in a). A node on the occlu-sal surface of the left dm1 (1) and a node at the articular eminence ofthe left temporomandibular joint (TMJ) (2) were constrained for dis-placement in three (all) orthogonal directions axes; a node at the emi-
nence of the right TMJ was constrained in the anteroposterior andinferosuperior directions (3). Force of 70 and 50 N were distributedevenly as ten nodal forces (F) in the anterior two-thirds of the inferiorsurface of the left zygomatic arch (working side) and the right zygo-matic arch (balancing side) respectively.
EFFECT OF SUTURES IN DYNAMIC SIMULATIONS 279

40 s with a voxel size of 0.25 mm, and saved in the digi-tal imaging and communications in medicine. Then, atessellated surface model was constructed from theseimages in Mimics 13.1 (The Materialise Group, Leuven,Belgium). After the model was surface meshed inMimics, it was transferred as a Binary STL file to Studio11.0 (Geomagic, Research Triangle Park, NC), a digitalshape sampling and processing software, to generate aclean and smooth polygon model. Then the model wasbrought back to Mimics to be converted into a 3D meshof 173,936 solid 4-noded tetrahedral elements andexported as a NASTRAN file. The elements in this modelvaried in size, but typical edge lengths of elements werebetween 0.4 and 1.7 mm. The NASTRAN file wasimported to Strand7 FEA software (v. 2.4.1; Strand7 Pty,Sydney, Australia) for both static and dynamic analysis.Parts for sutures were created by grouping elements(around three elements in thickness) roughly along theprofile of the zygomaticotemporal sutures at the zygo-matic arch at both sides of the cranium. Sutures wereapproximately 1–2 mm in thickness, and the two sutureshad 316 elements, occupying 0.08% of the total volumeof the cranium. Several simplifying assumptions wereused to reduce model complexity and avoid lengthy cal-culation times during dynamic simulation. These includethe lack of modeling of viscoelastic properties of bone,the omission of trabecular bone (trabeculae were mod-eled as cortical bone), teeth and periodontal ligaments,the lack of a complete set of muscle forces, the omissionof all other soft tissues and organs (such as the brainand eyeballs), and the modeling of only one pair ofsutures; only the zygomaticotemporal sutures on bothleft and right sides were modeled (at the age of 2.5years, most craniofacial sutures are open in rhesus mac-aques [Wang et al., 2006b]). This model was notidentical to the previous model with multiple suturesgenerated for static analysis (Wang et al., 2010a).
Elastic properties were the same for the bone yet fivedifferent sets of elastic material properties wereassigned to the sutures (Table 1). Briefly, the elasticmodulus (E) is a measure of material stiffness and is theamount of stress induced per unit of induced strain dueto an applied load. Poisson’s ratio (m) is a measure of thecoupling effect of strain components in transverse direc-tions; it is the negative of the ratio of the strain in thesecondary direction (response direction) divided bystrain in the primary direction (applied load direction).Coefficient of viscous damping measures the dissipation
of energy with time in a dynamic mechanical system.For bone, the nonsutural parts of the cranium wereassigned isotropic elastic properties based on an averageof values obtained from all parts of the skull of adultmacaques (elastic modulus E ¼ 17.3 GPa, Poisson’s ratiom ¼ 0.28; 1 GPa ¼ 109N/m2; Wang and Dechow, 2006).For sutures, there is no consensus on the elastic modu-lus of sutural tissue, as indicated by widely varyingpublished or used data (Wang et al., 2010a). For exam-ple, 1.2 MPa (Kupczik et al., 2009; 1 MPa ¼ 106N/m2),10 MPa (Moazen et al., 2009), 29 MPa (Compression) to74 MPa (Tension; Popowics and Herring, 2007), 46 MPa(Bright and Groning, 2011), 50 MPa (Odame et al.,2005), 0.4 GPa (Farke, 2008), and 0.5–2.5 GPa (Wanget al., 2008) have each been reported or proposed asproper elastic moduli. Five sets of isotropic elastic mate-rial properties were generated based on in vivo and insilico experiments listed above and presumptionsdetailed as follows (Table 1). Models 4 and 5 appliedhigh Poisson’s ratio (m ¼ 0.49). This value assumes thatsutural tissue is nearly incompressible (like a sealedfluid-filled volume) to ensure that bone fronts on oppo-site sides of the suture will not meet each other.Presumably, such contact would interfere with bonegrowth. In Models 2–5, a coefficient of viscous dampingwas assigned (Cv ¼ 0.451 Ns/m, Okazaki et al., 1996) tomodel the shock absorbing capacity of sutures associatedwith their internal friction. Elastic properties datadesigned for sutures were generated from multiple.
Simulation
A constraint regime or reaction loading procedure wasapplied to the model (Fig. 1b). To examine the strainpatterns of the skull incurred from dm1 biting, a node atthe articular eminence of the left temporomandibularjoint (TMJ) was constrained for displacement in three(all) axes, and a corresponding node at the eminence ofthe right TMJ was constrained in the anteroposteriorand inferosuperior directions (i.e., perpendicular to theline defined by the left and right TMJ nodes). A node onthe occlusal surface of the left dm1 (corresponds to theposition of the first permanent premolar/P3 in an adult)was constrained from displacing in three (all) orthogonaldirections.
Only two masticatory muscle forces were applied tothe model, representing the right and left masseters (su-perficial and deep). As calculated from physiological
TABLE 1. Material property values used for simulation models
Model Suture typeElastic
modulus (E)Poisson’sratio (m)
Density(g/cm3)
Coefficient ofviscous damping
(Ns/m)
Model l Suture I – fused suture (bone) 17.3 GPaa 0.28a 2.06a –Model 2 Suture II (static FEA theoretical value) 50 MPab 0.40b 1.06c 0.451d
Model 3 Suture III (experimental value) 1 MPab 0.40b 1.06c 0.451d
Model 4 Suture IV (high elasticity, high Poisson’s ratio) 50 MPab 0.49b 1.06c 0.451d
Model 5 Suture V (higher elasticity, high Poisson’s ratio) 1 MPab 0.49b 1.06c 0.451d
aValues of bone elastic properties and density were calculated from Wang and Dechow (2006) and Strait et al. (2005).bValues elastic properties for sutures were generated based on various in vitro and in silico experiments and further pre-sumptions (see text).cThe value for the density of sutures was a round-up of the value 1.0597 g/cm3 measured from unfixed rabbit and caninemuscle tissue (Mendez and Keys, 1960).dValue of coefficient of viscous damping was from Okazaki et al. (1996).
280 WANG ET AL.

cross-sectional area, the maximum force generatingcapacity of the superficial and deep masseters in adultM. mulatta are 100.8 and 39.0 N, respectively (�140 Nin total; Strait et al., 2005). In this study, a resultantforce of 70 (a coarse estimate for a skull of juvenile size)Newtons (x, y, z vector components: �13.7, 0, �68.6) wasdistributed evenly as 10 nodal forces in the anterior two-thirds of the inferior surface of the left zygomatic arch(working side), and a force of 50 N (9.8, 0, �49.0) wasapplied in a similar manner to the right zygomatic arch(balancing side). The asymmetrical loading pattern (70N vs. 50 N) simulates different activity levels in theWorking Side and Balancing Side muscles during unilat-eral tooth loading. Electromyographic data were notavailable for macaques of this age, so these values arecoarse estimates. We have previously modeled Working-to-Balancing Side asymmetry to be as great as 2-to-1 forpostcanine teeth loadings (Strait et al., 2005; Wanget al., 2010a), but asymmetry is expected to decline inforceful bites, and the forces used in this study likelyrepresent a forceful bite for a juvenile.
For each model, in addition to linear static simula-tions, two sets of linear transient dynamic simulationswere run using Strand7. Forces were applied to the mod-els at two different loading rates (Table 2). The firstloading rate (Slow Loading) used 10 time steps spreadover 100 ms, with the peak loading force occurring at 30ms. The second loading rate (Fast Loading) used 10 timesteps spread over 10 ms, with the peak force occurringat 3 ms. In adult rhesus monkeys, the time-to-peak ofbiting forces are reached from 31.0 ms (male) to 37.4 ms(female; Dechow and Carlson, 1990). In this sense, theSlow Loading design adopted a loading speed compara-ble to the accumulation speed of normal biting forces,thus it could be viewed as a simulation of the normalbiting speed (as a shorthand, the authors refer to thisloading regime as ‘‘normal’’ biting), yet the Fast Loadingwas an order of magnitude faster, simulating a quicksudden force loading, to examine the capacity of thesutures to absorb energy at different loading rates.
Sutural integrity was assessed by examining the prin-cipal stresses within the elements assigned suturematerial properties. In the pig nasofrontal sutures,Popowics and Herring (2007) measured the peak failurestresses in pigs (5–6 months old) to be 0.9 6 0.5 MPa(mean 6 SD, N ¼ 4) in tension and 3.4 6 1.4 MPa(mean 6 SD, N ¼ 4) in compression. Thus a principalstress bracket from �3.4 MPa (compressive) to 0.9 MPa
(tensile) was used as a ‘‘safe zone’’ to represent the rangeof safe stress values for typical cranial sutures. Obvi-ously, the boundaries of this safe zone are not meant toindicate that values outside of range will necessarilylead to suture failure, but rather that as one exceedsthese limits the risk of suture failure might reasonablybe expected to increase. Following the initial analyses,models whose principal stresses in the sutural elementsfall in the safe zone were subjected to increased forcesthat simulate very powerful bites. In this case, forceswere first doubled (140 N left, 100 N right), then maxi-mum forces were applied at both sides (140 N left, 140N right), and more loading cases were solved. The maxi-mum forces were calculated from the musclearchitecture of adult specimens, so they represent abnor-mally high loads for a juvenile.
RESULTS
Element Principal Stresses at theZygomaticotemporal Sutures
Element principal stresses (both compressive and ten-sile) of all sutural elements were examined in all fourmodels that include patent zygomaticotemporal sutures.The magnitude of the element principal stresses at someelements within the sutures fell outside the safe zone(from �3.4 to 0.9 MPa; Fig. 2), indicating that thesesutures are at varying degrees of risk of failure during‘‘normal’’ biting. In Model 2, 104 of 632 sutural elements(16.5%) fell out of the safe zone. This number rose to164 out of 632 (25.9%) in Model 4. These two modelsapply an elastic modulus of 50 MPa to sutural tissue. Incontrast, Models 3 and 5 applied a modulus value of 1MPa. In Model 5, 42 elements (6.6%) were outside of thesafe zone, but in Model 3, which used a lower Poisson’sratio (0.40 vs. 0.49), only one element (0.1%) was slightlyover 0.9 MPa in tensional stress. When the loadingforces were doubled in magnitude in Model 3, 15 ele-ments (2.4%) were out of the safe zone, and when a forceof 140 N was applied on both left and right sides, thatnumber rose to 41 elements (6.5%). These results wereunaffected by the nature of the simulations (Static vs.Dynamic).
Overall Strain Patterns
In general, global strain patterns on craniofacial bonesurfaces are comparable for each FEA simulationregardless of assigned material property values or load-ing design (Fig. 3). Suture patency affected strainslocally insofar as decreasing sutural stiffness was associ-ated with local reductions in strain on the zygomaticarch, an area that normally is highly strained owing tothe fact that the muscle loads are applied there.
Strain Energy Absorbed by theZygomaticotemporal Sutures
The percentages of strain energy absorbed by thezygomaticotemporal suture in dynamic simulations areidentical to those in the static simulations for a givenset of material properties. However, these absorbedstrain energies are different depending on which suturemodel is being loaded, with Models 2 and 5 absorbingslightly more energy than Model 3 (Table 3). The
TABLE 2. Profiles of fast and slow dynamic transientlinear simulations
StepSlow
loading (ms)Fast
loading (ms)Percentage
of total peak force
0 0 0 01 10 1 402 15 1.5 603 20 2 804 30 3 1005 40 4 1006 50 5 1007 60 6 808 70 7 609 80 8 4010 90 9 0
EFFECT OF SUTURES IN DYNAMIC SIMULATIONS 281

increase in the amount of strain energy absorbed by thezygomaticotemporal sutures lagged behind the increasein force, as in the bone from the rest of the cranium(Table 4). Compared with bone, the zygomaticotemporalsutures do not play a greater role in absorbing energy atany stage during the loading process.
Biting Efficiency
In all five models, the reaction force stepwise profile isidentical regardless of loading speed (Fig. 4), and the dif-ferences in biting efficiency (¼reaction forces divided bythe loading forces along Z axis) due to the loadingdesigns and material property values are small (Table5), indicating insignificant or no effect on biting effi-
ciency by the presence of the zygomaticotemporalsutures, as also observed in a previous analysis (Wanget al., 2010a).
DISCUSSION
Limitations of the Study
The FEM used in this analysis is limited in certainrespects. First, it was simplified structurally to reducemodel complexity and avoid lengthy calculation timesduring dynamic simulation; thus, it was impractical tomodel all of the sutures. Second, this FEM simplifiesthe viscoelastic properties of the sutures, which arenormally a combination of several mechanical behaviors(including both elastic and viscous components), such
Fig. 2. Distribution of element principal stresses (maximum and minimum combined) in right and leftzygomaticotemporal sutures (total number of element principal stresses: 632 ¼ 316 � 2) during SlowLoading Dynamic simulations. A ‘‘safe zone’’ was identified between �3.4 and 0.9 MPa based on datafrom Popowics and Herring (2007).
282 WANG ET AL.

that only a single coefficient of viscous damping wasassigned in addition to a linear elastic modulus. Inaddition, the sutural deformation behavior and capacityfor absorbing energy would be different with inclusionof many factors not considered in this article, such assize of model elements, loading regimes more similar toa physiological reality, suture morphology (i.e., intra-and inter-species variations in terms of ontogeny andfusion patterns), and particularly an effective valida-tion with consideration of micro-anatomical features in
sutures (i.e., interdigitations and interactions with cov-ering linings, such as dura matter or periosteom).Nonetheless, the models are usefully serve the purposeof examining the global effects of zygomaticotemporalsutures under dynamic simulation, since comparisonsamong results from different experiments (with sutureor without suture) using identical loading regimes mayprovide a useful framework for understanding the bio-mechanical behavior and global influence of sutures ingeneral.
Fig. 3. Von Mises strain (mm/mm) in five suture models under simplified left dm1 biting point with forceof 70 N in the left side and 50 N in the right side. White areas on the zygomatic arch indicated von Misesstrain values exceeds the scale minimum (500 le).
EFFECT OF SUTURES IN DYNAMIC SIMULATIONS 283
Influence of Zygomaticotemporal Sutures onGlobal Skull Mechanics
Results obtained in this study are inconsistent withthe hypotheses that (1) simple sutures such as the
zygomaticotemporal behave differently under dynamicrather than static loads, and (2) sutures have a profoundimpact on global strain patterns. It is unclear whetheror not other craniofacial sutures or variation in morpho-logical complexity would influence strain patternsduring these dynamic simulations. Results of this analy-sis do not exclude the possibility that highly complexsutures (such as those seen in head-butting vertebrates),or those with proportionally large volumes may performthis role. It would appear that simple, linear, and butt-ended sutures do not possess advantageous properties inthe context of feeding biomechanics.
Findings of this analysis differ from those of Moazenet al. (2008, 2009a,b), in which the presence/absence ofsutures affected strains both near the suture site and atother regions across the cranium. Previously, a staticstudy by Wang et al. (2010a) examining many suturessimultaneously, found that suture patency had only alimited effect on strain magnitudes at sites across thecranium. It is notable that the size and architecture ofthe lizard skull examined by Moazen et al. (2008,2009a,b) differs from that of macaques. The lizard skullis at least an order of magnitude smaller in size and iscomposed primarily of bony struts. Although the maca-que skull used in this study is a juvenile, it is still largerand composed of bony plates. These differences mayaffect the manner in which sutures mediate cranialstrains. Findings of this analysis are also contrary tothose by Kupczik et al. (2007) who also used macaque
TABLE 3. Percentage of total strain energy absorbed by the sutures during static and slow loading simula-tions (peak force – 70 N left, 50 N right) relative to the total strain energy in cranium at corresponding steps
Model 1(17.3 GPa, 0.28)
Model 2(50 MPa, 0.40)
Model 3(1 MPa, 0.40)
Model 4(50 MPa, 0.49)
Model 5(1 MPa, 0.49)
Static (%) 4.3 9.9 8.3 6.5 9.5DS1 (%) 4.4 9.9 8.2 6.5 9.6DS2 (%) 4.3 9.8 8.2 6.4 9.5DS3 (%) 4.4 9.9 8.2 6.5 9.6DS4 (%) 4.3 9.8 8.2 6.4 9.5DS5 (%) 4.4 9.9 8.3 6.5 9.6DS6 (%) 4.3 9.8 8.2 6.4 9.5DS7 (%) 4.3 9.9 8.3 6.5 9.6DS8 (%) 4.3 9.9 8.2 6.5 9.5DS9 (%) 4.3 9.8 8.2 6.4 9.5DS10 (%) 0.2 0.7 0.5 0.3 1.3
DS, dynamic steps.
TABLE 4. Stepwise energy absorption in bones and sutures relative to peak values during dynamic simula-tions in Model 3 (E 5 1 MPa, m 5 0.40; Peak force – 70 N left, 50 N right)
Dynamic stepsForce relative
to peak force (%)
Model 3 (1 MPa, 0.40) slowloading
Model 3 (1 MPa, 0.40) fastloading
Bone (%) Suture (%) Bone (%) Suture (%)
Step 1 40 16.0 16.0 15.7 15.9Step 2 60 36.0 36.0 36.0 36.0Step 3 80 64.0 63.9 62.5 63.4Step 4 100 100.0 100.0 100.0 100.0Step 5 100 100.0 100.0 97.8 99.1Step 6 100 100.0 100.0 99.8 100.0Step 7 80 63.9 64.4 62.8 63.3Step 8 60 36.0 36.0 35.8 36.1Step 9 40 16.0 16.0 15.8 15.7Step 10 0 0.00 0.00 0.02 0.00
Peak forces were reached at Step 4 and remained unchanged at Steps 5 and 6. Forces were canceled completely at Step 10.
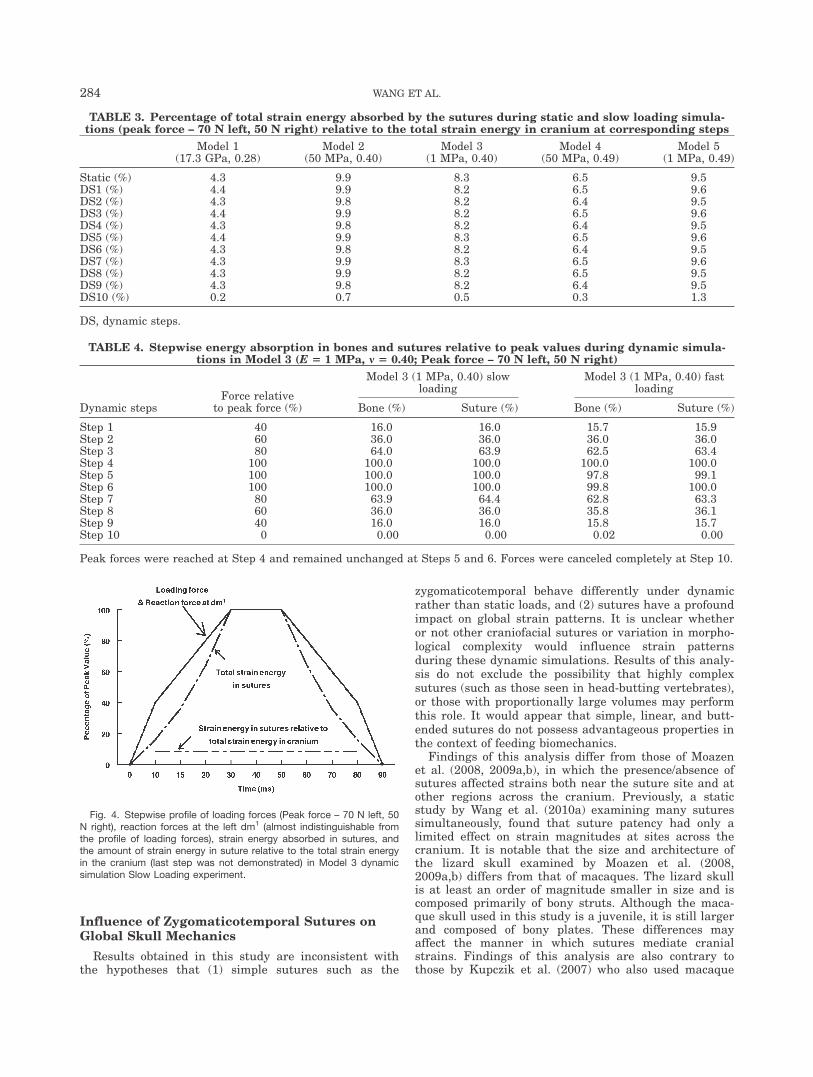
Fig. 4. Stepwise profile of loading forces (Peak force – 70 N left, 50N right), reaction forces at the left dm1 (almost indistinguishable fromthe profile of loading forces), strain energy absorbed in sutures, andthe amount of strain energy in suture relative to the total strain energyin the cranium (last step was not demonstrated) in Model 3 dynamicsimulation Slow Loading experiment.
284 WANG ET AL.

models and who observed that sutures had a globaleffect, but whose material property values may havebeen influenced by chemical preservative soaked intothe tissues of the skulls examined by them. Moreover,Kupczik et al. (2007) modeled a loading regime corre-sponding to an in vitro experiment, and those loadingconditions do not fully reproduce mastication. Assess-ment of the inconsistencies between Moazen et al. (2008,2009a, b), Kupczik et al. (2007) and the present studyrequires detailed sensitivity studies focusing on bonematerial properties, constraints, and a consideration ofstrut versus plate mechanics.
Sensitivity of Sutural Element Stress toVariation in Material Properties
The zygomaticotemporal sutures exhibited consider-able sensitivity to variation in material properties.Under ‘‘normal’’ biting conditions that correspond toslightly elevated loads in juveniles, comparatively minorvariations in elastic modulus and Poisson’s ratio led tostress levels that could have potentially threatenedsuture integrity. In general, the potential risk to integ-rity increased with increasing elastic modulus andPoisson’s ratio. Thus, as suture connective tissue growsstiffer, it is more likely to injure. This analysis does notallow one to conclude that the material properties usedin Model 3 are accurate, but it does imply that thoseused in the other models could be unrealistic. Anotherimplication of this study is that suture connective tissuemay not behave as a perfectly incompressible material.
Even when using the most favorable material proper-ties, suture integrity still may be threatened duringmaximal biting conditions (Fig. 3). Moreover, suture ma-terial properties almost certainly change duringontogeny, and those changes might influence the degreeto which stress accumulates in the suture (Popowics andHerring, 2007). Thus, one might infer that sutures aremechanically vulnerable structures. Given that suturesare important loci of bone growth, it appears evidentthat natural selection had to favor mechanisms to shieldsutures from stress. This possibility has potentiallybroad implications.
Sutures and the Evolution of Craniofacial Form
The primary function of sutures in young individualsis to allow the growth of the skull. The finding that ele-vated feeding loads have the potential to threatensuture integrity at certain material properties stands incontrast to experimental data demonstrating that the fa-
cial skeleton in several mammalian species normallyexperiences relatively low strains during feeding (thebehavior that most regularly exposes the cranium tohigh or repetitive loads; Lieberman et al., 2004; Rossand Metzger, 2004). This has led to the hypothesis thatsome aspects of the form of the facial skeleton in manyclades may not be optimized to dissipate feeding loads(Ross and Metzger, 2004), despite the fact that compara-tive analyses seem to suggest a relationship betweencraniofacial form and feeding behavior (Hylander,1977a,b; Rak, 1983; Anton, 1990, 1993; Ravosa, 1991a,b;Spencer and Demes, 1993; Daegling, 1996; Ross, 2001;Trinkaus, 2003; Vinyard et al., 2003; Wright, 2005; Bas-tir et al., 2007; Strait et al., 2008, 2009; Gilbert et al.,2009; Williams et al., 2009; Wang et al., 2010b; Wang,2012).
One reason underlying this apparent contradictionmay be that craniofacial form in many mammals and,perhaps, other vertebrates as well, might include designfeatures that minimize or otherwise modulate stressesin sutures rather than in bone. The authors hypothesizethat the primary function of cranial sutures in subadultindividuals is intramembranous bone growth. Becausesutures are zones of structural weakness, they must beshielded from high stresses so that the vertebrate cra-nium can grow a shape appropriate for properfunctioning of sensory, respiratory, and feeding systems.The result in this article that elevated feeding loadshave the potential to damage sutures with certain mate-rial properties, suggests that craniofacial form inmammals, and perhaps other vertebrates, might includedesign features that reduce stresses in sutures ratherthan in bone. These design features might be found notonly in high stress areas of the facial skeleton but alsoin areas characterized by lower stresses (Hylander et al.,1991; Ross, 2001; Ross and Metzger, 2004).
Several overlapping predictions could be derived fromthis hypothesis: (1) cranial sutures (via the guidingforces of natural selection) when possible, had to befound in areas where stresses do not concentrate in thevertebrate cranium; (2) when cranial sutures are foundin areas that experience relatively high stress concentra-tions, suture complexity forms as a protectivemechanism to preserve the growth center; (3) the facialskeleton had to adapt by taking a form that minimizesstress across the face, or in particular facial regions.Consequently, craniofacial form may be optimized forfeeding while growing throughout ontogeny, but not sim-ply by minimizing stress and strain in bones.
Many of the components of the hypothesis describedabove can be found in prior studies, but to our knowl-edge, they have not previously been consolidated in acomprehensive framework. This hypothesis can be testedusing comparative, experimental, and modeling meth-ods. For example, one mechanism being considered is anage-related increase in cranial suture complexityobserved in mice (Byron, 2006) and humans (Saito et al.,2002) that helps to preserve flat bone growth fronts.Moreover, it will be important to establish the degree towhich the hypothesis applies across vertebrates. It isconceivable that it applies only to certain vertebrategroups (e.g., mammals), body sizes, or that the hypothe-sis manifests itself differently in different groups.Craniofacial strain magnitudes recorded during in vivoexperiments in alligators are approximately twice those
TABLE 5. Nodal reaction forces at the left dm1 bitingpoint in Z axis (up and down direction relative to theocclusal plane. See Fig. 1) compared with total load-ing forces (Peak force – 70 N left, 50 N right) in Z axis
Static(%)
Dynamic –slow
loading (%)
Dynamic –fast
loading (%)
Model 1 (17.3 GPa, 0.28) 42.1 42.1 43.2Model 2 (50 MPa, 0.40) 43.8 43.8 44.9Model 3 (1 MPa, 0.40) 45.1 45.1 46.2Model 4 (50 MPa, 0.49) 43.1 43.1 44.2Model 5 (1 MPa, 0.49) 44.8 44.9 46.0
EFFECT OF SUTURES IN DYNAMIC SIMULATIONS 285

observed during analogous experiments in primates,swine, and hyraxes (Lieberman et al., 2004; Ross andMetzger, 2004), and strain magnitudes in FEA simula-tions of lizards resemble those of alligators (Moazenet al., 2008, 2009a), so it is possible that broad patternsof cranial biomechanics differ between mammals andnonmammal vertebrates. Moreover, some sutures in cer-tain taxa may be specially adapted to accommodatemovements (cranial kinesis), and in those taxa the abovehypothesis might not apply.
Challenges in Suture Biology andBiomechanics
Although the global impact of sutures is negligible inskulls of relatively large mammals such as monkeys andpigs (Wang et al., 2010a; Bright and Groning, 2011),their local effect and interactions with bone fronts andcovering tissues might be of special interest in studies ofbone growth and adaptation, given that sutures are mor-phologically diverse in terms of complexity, location (i.e.,cranial vs. facial), ontogeny, fusion, sexual dimorphism,and intra- and inter-species differences. The diversity insutural morphology poses a tremendous challenge toanalyses of suture biology and biomechanics.
Historically, important experimental work by Moss inthe mid-twentieth century demonstrated that suturalmorphologies are somewhat labile when the mechanicalenvironment is perturbed, which led to a synthesis ofsuture concepts that defined intrinsic and extrinsic fac-tors that direct the fate of cranial suture formation(Moss, 1954, 1957, 1961, 1962; Moss and Young, 1960).An important intrinsic factor that has been the focus ofmuch research in the last 20 years is the outermost cra-nial meninx, the dura mater, which regulates significantsuture growth events such as fusion or maintenance ofpatency (Opperman et al., 1993, 1995, 1998). Within thelast 10 years, Byron and colleagues (Byron et al., 2004;Byron, 2006) have hypothesized that small-scale suturegrowth events such as adding (or resorbing) bone min-eral to (from) the leading edge of a suture front areinfluenced by extrinsic, mechanical events that oftenrelate to masticatory biomechanics. This assertion issupported by comparative evidence in primates (Byron,2009) and in caimans (Monteiro and Lessa, 2000) wheredurophagous feeders have enhanced cranial suturecomplexity.
Unlike cranial sutures, facial sutures do not overliedura and thus do not receive the kinds of fate-determin-ing cytokine secretions that the braincase suturesreceive. This is a fundamental difference between twokinds of suture, which suggests that facial sutures maybe more directed by extrinsic mechanical forces than cal-varial sutures. Traditionally, suture complexities arenormally modeled in two dimensions using fractal geom-etry (Long, 1985; Hartwig, 1991; Long and Long, 1992;Monteiro and Lessa, 2000; Skrzat and Walocha, 2003a,b;Lynnerup and Jacobsen, 2003; Yu et al., 2003; Byronet al., 2004; Byron, 2006). However, if facial sutures pro-vide a more ‘‘pure’’ indication of biomechanical loadinghistory, then a detailed study on sutural three-dimen-sional complexity measures will be necessary.
Ultimately, the competing interests of protection andgrowth, and their variation throughout ontogeny andphylogeny warrant more study at both macro- and
micro-anatomical levels. FEA will continue to be a usefultool for addressing these challenges, along withadvanced imaging modalities (such as micro-CT) andhistological studies (i.e., Stadler et al., 2006; Reinholtet al., 2009; Holliday et al., 2010; Smith et al., 2010;Payne et al., 2011), growth, complexity, and fusion pat-terns ( i.e., Yu et al., 2003; Sun et al., 2004; Byron, 2006,2009; Wang et al., 2006b), genetics and evolution of su-tural configuration patterns (Sidor, 2001; Wang et al.,2006a), and in vivo and in vitro experiments ( i.e., Beh-rents et al., 1978; Henry and Teng, 2000; Wang et al.,2008).
CONCLUSION
FEA was used to evaluate the roles of zygomaticotem-poral sutures in biomechanics of a macaque cranium.Static and dynamic analyses produced similar results interms of strain patterns and reaction forces, indicatingthat zygomaticotemporal sutures play a limited role inmodulating global skull mechanics regardless of loadingdesign. The zygomaticotemporal sutures did not absorbsignificant amounts of energy during dynamic simula-tions, regardless of loading speed. It is thushypothesized that sutures are mechanically significantonly insofar as they are weak points on the craniumthat must be shielded from unduly high stresses so asnot to disrupt vitally important growth processes.Sutures are morphologically diverse in terms of location,ontogeny, fusion, sexual dimorphism, and intra- andinter-species variations. This hypothesis predicts thatsutural protection is an important selective pressurethat may have influenced diverse aspects of sutural andcraniofacial form.
ACKNOWLEDGEMENT
This project was funded by grants from the National Sci-ence Foundation Physical Anthropology HOMINID pro-gram (BCS 0725183, 0725078, 0725126, 0725136, and0725147). Animal tissues were obtained from the YerkesPrimate Research Center at the Emory University,Atlanta, Georgia (Supported by NIH Base GrantRR00165). Dr. Sean Shih-Yao Liu helped provide the CT-scanning of the specimen. Dr. Betsy Dumont, Mr. DanPulaski, Dr. Jul Davis, and Dr. Ming Chen are sincerelythanked for help during the generation of the FEM of ajuvenile macaque at the 4th Annual Workshop on ‘‘FiniteElement Modeling in Biology’’ May 29 – June 5, 2010, atUniversity of Massachusetts, Amherst (NSF DBI0743460; http://www.biomesh.org/workshops). Theauthors are also grateful to Ms. Anne Delvaux, Dr. JerryTift, Dr. Michael Horst, Dr. Daniel Hagan, Dr. SandraLeeper-Woodford, Dr. Padmanabhan Rengasamy, Dr.Bob Moon, Eng. Carlos Lopez, Mrs. Denise Collins, Mrs.Ernestine Waters, Mrs. Marianne Watkins, and Mrs. LiSun for help of various kinds. The authors sincerelythank the editors and reviewers for providing valuableadvice for improving the manuscript.
LITERATURE CITED
Anton SC. 1990. Neandertals and the anterior dental hypothesis: abiomechanical evaluation of bite force production. Kroeber An-thropological Society Papers 71–72:67–76.
286 WANG ET AL.

Anton SC. 1993. Internal masticatory muscle architecture in theJapanese macaque and its influence on bony morphology(Abstract). Am J Phys Anthropol S16:50.
Bastir M, O’Higgins P, Rosas A. 2007. Facial ontogeny in Neander-thals and modern humans. Proc Biol Sci 274:1125–1132.
Behrents RG, Carlson DS, Abdelnour T. 1978. In vivo analysis ofbone strain about the sagittal suture in Macaca mulatta duringmasticatory movements. J Dent Res 57:904–908.
Bright JA, Groning F. 2011. Strain accommodation in the zygomaticarch of the pig: a validation study using digital speckle patterninterferometry and finite element analysis. J Morphol 272:1388–1398.
Byron CD. 2006. Role of the osteoclast in cranial suture waveformpatterning. Anat Rec 288A:552–563.
Byron CD. 2009. Cranial suture morphology and its relationship todiet in Cebus. J Hum Evol 57:649–655.
Byron CD, Borke J, Yu J, Pashley D, Wingard CJ, Hamrick M.2004. Effects of increased muscle mass on mouse sagittal suturemorphology and mechanics. Anat Rec 279A:676–684.
Cho HJ. 2009. A three-dimensional cephalometric analysis. J ClinOrthod 43:235–252.
Daegling DJ. 1996. Growth in the mandibles of African apes. JHum Evol 30:315–341.
Dechow PC, Carlson DS. 1990. Occlusal force and craniofacial bio-mechanics during growth in rhesus monkeys. Am J Phys Anthro-pol 83:219–237.
Dumont ER, Herrel A. 2003. The effects of gape angle and bitepoint on bite force in bats. J Exp Biol 206:2117–2123.
Dumont ER, Piccirillo J, Grosse IR. 2005. Finite-element analysis ofbiting behavior and bone stress in the facial skeletons of bats.Anat Rec 283A:319–330.
Farke AA. 2008. Frontal sinuses and head-butting in goats: a finiteelement analysis. J Exp Biol 211:3085–3094.
Gilbert CC, Frost SR, Strait DS. 2009. Allometry, sexual dimor-phism, and phylogeny: a cladistic analysis of extant Africanpapionins using craniodental data. J Hum Evol 57:298–320.
Grosse IR, Dumont ER, Coletta C, Tolleson A. 2007. Techniques formodeling muscle-induced forces in finite element models of skele-tal structures. Anat Rec 290:1069–1088.
Hartwig WC. 1991. Fractal analysis of sagittal suture morphology.J Morphol 210:289–298.
Herring SW. 1993. Epigenetic and functional influences on skullgrowth. Skull 1:153–206.
Herring SW, Mucci RJ. 1991. In vivo strain in cranial sutures: thezygomatic arch. J Morphol 207:225–239.
Herring SW, Rafferty KL. 2000. Cranial and facial sutures: func-tional loading in relation to growth and morphology. In: Davido-vitch Z, Mah J, editors. Biological mechanisms of tooth eruption,resorption and replacement by implants. Boston: Harvard SocAdv Orthodontics. p 269–276.
Herring SW, Teng S. 2000. Strain in the braincase and its suturesduring function. Am J Phys Anthropol 112:575–593.
Holliday CM, Gardner NM, Paesani SM, Douthitt M, Ratliff JL.2010. Microanatomy of the mandibular symphysis in lizards: pat-terns in fiber orientation and Meckel’s cartilage and their signifi-cance in cranial evolution. Anat Rec 293:1350–1359.
Hylander WL. 1977a. The adaptive significance of Eskimo craniofa-cial morphology. In: Dahlberg AA, Graber TM, editors. Orofacialgrowth and development. The Hague: Mouton. p 129–169.
Hylander WL. 1977b. In vivo bone strain in the mandible of Galagocrassicaudatus. Am J Phys Anthropol 46:309–326.
Hylander WL, Picq PG, Johnson KR. 1991. Masticatory-stress hy-pothesis and the supraorbital region of primates. Am J PhysAnthropol 86:1–36.
Jasinoski SC, Rayfield EJ, Chinsamy A. 2010a. Functional implica-tions of dicynodont cranial suture morphology. J Morphol 271:705–728.
Jasinoski SC, Reddy BD, Louw KK, Chinsamy A. 2010b. Mechanicsof cranial sutures using the finite element method. J Biomech 43:3104–3111.
Jaslow CR. 1990. Mechanical properties of cranial sutures. J Bio-mech 23:313–321.
Jaslow CR, Biewener AA. 1995. Strain patterns in the horncores,cranial bones and sutures of goats (Capra hircus) during impactloading. J Zool 235:193–210.
Kupczik K, Dobson CA, Crompton RH, Phillips R, Oxnard CE,Fagan MJ, O’Higgins P. 2009. Masticatory loading and bone adap-tation in the supraorbital torus of developing macaques. Am JPhys Anthropol 139:193–203.
Kupczik K, Dobson CA, Fagan MJ, Crompton RH, Oxnard CE,O’Higgins P. 2007. Assessing mechanical function of the zygo-matic region in macaques: validation and sensitivity testing of fi-nite element models. J Anat 210:41–53.
Lieberman DE, Krovitz GE, Yates FW, Devlin M, St. Claire M.2004. Effects of food processing on masticatory strain and cranio-facial growth in a retrognathic face. J Hum Evol 46:655–677.
Long CA. 1985. Intricate sutures as fractal curves. J Morphol 185:285–295.
Long CA, Long JE. 1992. Fractal dimensions of cranial sutures andwaveforms. Acta Anat (Basel) 145:201–206.
Lynnerup N, Jacobsen JC. 2003. Brief communication: age and frac-tal dimensions of human sagittal and coronal sutures. Am J PhysAnthropol 121:332–336.
Mendez J, Keys A. 1960. Density and composition of mammalianmuscle. Metabolism 9:184–188.
Moazen M, Curtis N, Evans SE, O’Higgins P, Fagan MJ. 2008.Combined finite element and multibody dynamics analysis of bit-ing in a Uromastyx hardwickii lizard skull. J Anat 213:499–508.
Moazen M, Curtis N, O’Higgins P, Evans SE, Fagan MJ. 2009a. Bio-mechanical assessment of evolutionary changes in the lepidosau-rian skull. Proc Natl Acad Sci USA 106:8273–8277.
Moazen M, Curtis N, O’Higgins P, Jones MEH, Evans SE, FaganMJ. 2009b. Assessment of the role of sutures in a lizard skull: acomputer modelling study. Proc R Soc Lond B Biol Sci 276:39–46.
Monteiro LR, Lessa LG. 2000. Comparative analysis of cranialsuture complexity in the genus Caiman (Crocodylia, Alligatori-dae). Braz J Biol 60:689–694.
Moss ML. 1954. Growth of the calvaria in the rat. The determina-tion of osseous morphology. Am J Anat 94:333–362.
Moss ML. 1957. Experimental alteration of sutural area morphol-ogy. Anat Rec 127:569–589.
Moss ML. 1961. Extrinsic determination of sutural area morphologyin the rat calvaria. Acta Anat (Basel) 44:263–272.
Moss ML. 1962. The functional matrix. In: Kraus B, Reidel R, edi-tors. Vistas in orthodontics. Philadelphia: Lea & Febiger. p 85–98.
Moss ML, Young RW. 1960. A functional approach to craniology. AmJ Phys Anthropol 18:281–292.
Odame P, Yu JC, Zhang G. 2005. 3D Finite Element study of stressdistribution at the cranial bone/suture interface. Transactions ofthe Society for Biomaterials 26:556.
Opperman LA. 2000. Cranial sutures as intramembranous bonegrowth sites. Dev Dyn 219:472–485.
Opperman LA, Chhabra A, Nolen AA, Bao Y, Ogle RC. 1998. Duramater maintains rat cranial sutures in vitro by regulating suturecell proliferation and collagen production. J Craniofac Genet DevBiol 18:150–158.
Opperman LA, Passarelli RW, Morgan EP, Reintjes M, Ogle RC.1995. Cranial sutures require tissue interactions with dura materto resist osseous obliteration in vitro. J Bone Miner Res 10:1978–1987.
Opperman LA, Sweeney TM, Redmon J, Persing JA, Ogle RC. 1993.Tissue interactions with underlying dura mater inhibit osseousobliteration of developing cranial sutures. Dev Dyn 198:312–322.
Oudhof HAJ, Van Doorenmaalen WJ. 1983. Skull morphogenesisand growth: hemodynamic influence. Acta Anat 117:181–186.
Okazaki M, Fukumoto M, Takahashi J. 1996. Damped oscillationanalysis of natural and artificial periodontal membranes. AnnBiomed Eng 24:234–240.
Payne SL, Holliday CM, Vickaryous MK. 2011. An osteological andhistological investigation of cranial joints in geckos. Anat Rec294:399–405.
Popowics TE, Herring SW. 2007. Load transmission in the nasofron-tal suture of the pig, Sus scrofa. J Biomech 40:837–844.
EFFECT OF SUTURES IN DYNAMIC SIMULATIONS 287

Porro LB, Holliday CM, Anapol F, Ontiveros LC, Ontiveros LT, RossCF. 2011. Free body analysis, beam mechanics, and finite elementanalysis of the mandible of Alligator mississippiensis. J Morphol272:910–937.
Rafferty KL, Herring SW. 1999. Craniofacial sutures: Morphology,growth, and in vivo masticatory strains. J Morphol 242:167–179.
Rafferty KL, Herring SW, Marshall CD. 2003. Biomechanics of therostrum and the role of facial sutures. J Morphol 257:33–44.
Rak Y. 1983. The Australopithecine face. New York: AcademicPress.
Ravosa MJ. 1991a. Interspecific perspective on mechanical and non-mechanical models of primate circumorbital morphology. Am JPhys Anthropol 86:369–396.
Ravosa MJ. 1991b. Ontogenetic perspective on mechanical and non-mechanical models of primate circumorbital morphology. Am JPhys Anthropol 85:95–112.
Rayfield EJ. 2004. Cranial mechanics and feeding in Tyrannosaurusrex. Proc R Soc Lond B Biol Sci 271:1451–1459.
Rayfield EJ. 2005. Using finite-element analysis to investigatesuture morphology: a case study using large carnivorous dino-saurs. Anat Rec 283A:349–365.
Reed DA, Porro LB, Iriarte-Diaz J, Lemberg JB, Holliday CM, Ana-pol F, Ross CF. 2011. The impact of bone and suture materialproperties on mandibular function in Alligator mississippiensis:testing theoretical phenotypes with finite element analysis. JAnat 218:59–74.
Reinholt LE, Burrows AM, Eiting TP, Dumont ER, Smith TD. 2009.Histology and micro CT as methods for assessment of facialsuture patency. Am J Phys Anthropol 138:499–506.
Ross CF. 2001. In vivo function of the craniofacial haft: the interor-bital ‘‘pillar.’’ Am J Phys Anthropol 116:108–139.
Ross CF, Metzger KA. 2004. Bone strain gradients and optimizationin vertebrate skulls. Ann Anat 186:387–396.
Saito K, Shimizu Y, Ooya K. 2002. Age-related morphologicalchanges in squamous and parietomastoid sutures of human cra-nium. Cells Tissues Organs 170:266–273.
Sidor CA. 2001. Simplification as a trend in synapsid cranial evolu-tion. Evolution 58:1419–1442.
Skrzat J, Walocha J. 2003a. Application of fractal dimension inevaluation of cranial suture complexity. Harmonic Fractal ImageAnal 1:39–41.
Skrzat J, Walocha J. 2003b. Fractal dimensions of the sagittal(interparietal) sutures in humans. Folia Morphol (Warsz) 62:119–122.
Smith KK, Hylander WL. 1985. Strain gauge measurement of meso-kinetic movement in the lizard Varanus exanthematicus. J ExpBiol 114:53–70.
Smith TD, Burrows AM, Dumont ER. 2010. Microanatomicalassessment of nasomaxillary suture patency. Anat Rec 293:651–657.
Spencer MA, Demes B. 1993. Biomechanical analysis of masticatorysystem configuration in Neandertals and Inuits. Am J PhysAnthropol 91:1–20.
Stadler JA, Cortes W, Zhang L-L, Hanger CC, Gosain AK. 2006. Areinvestigation of murine cranial suture biology: microcomputedtomography versus histologic technique. Plast Reconstr Surg 118:626–634.
Strait DS, Wang Q, Dechow PC, Ross CF, Richmond BG, SpencerMA, Patel BA. 2005. Modeling elastic properties in finite element
analysis: how much precision is needed to produce an accuratemodel? Anat Rec 283A:275–287.
Strait DS, Weber GW, Neubauer S, Chalk J, Richmond BG, LucasPW, Spencer MA, Schrein C, Dechow PC, Ross CF, Grosse IR,Wright BW, Constantino P, Wood BA, Lawn B, Hylander WL,Wang Q, Byron C, Slice DE, Smith AL. 2009. The feeding biome-chanics and dietary ecology of Australopithecus africanus. ProcNatl Acad Sci U S A 106:2124–2129.
Strait DS, Wright BW, Richmond BG, Ross CF, Dechow PC, SpencerMA, Wang Q. 2008. Craniofacial strain patterns during premolarloading: implications for human evolution. In: Vinyard C, RavosaMJ, Wall C, editors. Primate craniofacial function and biology.New York: Springer. p 173–198.
Sun Z, Lee E, Herring SW. 2004. Cranial sutures and bones: growthand fusion in relation to masticatory strain. Anat Rec 276A:150–161.
Trinkaus E. 2003. Neandertal faces were not long; modern humanfaces are short. Proc Natl Acad Sci USA 100:8142–8145.
Vinyard CJ, Wall CE, Williams SH, Hylander WL. 2003. Compara-tive functional analysis of skull morphology of tree-gouging pri-mates. Am J Phys Anthropol 120:153–170.
Wang Q. 2012. Dental maturity and the ontogeny of sex-baseddifferences in the dentofacial complex of rhesus macaques fromCayo Santiago. In: Wang Q, editor. Bones, Genetics, and Behaviorof Rhesus Macaques: Macaca mulatta of Cayo Santiago andBeyond. Springer: New York. p 177–194.
Wang Q, Dechow PC. 2006. Elastic properties of external corticalbone in the craniofacial skeleton of the rhesus monkey. Am JPhys Anthropol 131:402–415.
Wang Q, Dechow PC, Wright BW, Ross CF, Strait DS, RichmondBG, Spencer MA, Byron CD. 2008. Surface strain on bone andsutures in a monkey facial skeleton: an in vitro method and itsrelevance to Finite Element Analysis. In: Vinyard CJ, Ravosa MJand Wall CE, editors. Primate craniofacial function and biology.New York: Springer. p 149–172.
Wang Q, Opperman LA, Havill LM, Carlson DS, Dechow PC.2006a. Inheritance of sutural patterns at pterion in rhesus mon-keys skulls. Anat Rec 288A:1042–1049.
Wang Q, Strait DS, Dechow PC. 2006b. Fusion patterns of craniofa-cial sutures in rhesus monkey skulls of known age and sex fromCayo Santiago. Am J Phys Anthropol 131:469–485.
Wang Q, Smith AL, Strait DS, Wright BW, Richmond BG, GrosseIR, Byron CD, Zapata U. 2010a. The global impact of suturesassessed in a Finite Element Model of a macaque cranium. AnatRec 293:1477–1491.
Wang Q, Wright BW, Smith A, Chalk J, Byron CD. 2010b. Mechani-cal impact of incisor loading on the primate midfacial skeletonand its relevance to human evolution. Anat Rec 293:607–617.
Williams SH, Vinyard CJ, Wall CE, Hylander WL. 2009. Mandibu-lar corpus bone strain in goats and alpacas: implications forunderstanding the biomechanics of mandibular form in seleno-dont artiodactyls. J Anat 214:65–78.
Wright BW. 2005. Craniodental biomechanics and dietary toughnessin the genus Cebus. J Hum Evol 48:473–492.
Yu JC, Wright RL, Williamson MA, Braselton JP, Abell ML. 2003. Afractal analysis of human cranial sutures. Cleft Palate CraniofacJ 40:409–415.
288 WANG ET AL.
Copyright © 2022 FDOKUMEN




















