
The role of the burrow funnel in feeding processes in the lugworm Arenicola marina (L.)
12
ELSEVIER Journal of Experimental Marine Biology and Ecology, 202 (1996) 107-l 18 JOURNAL OF EXPERIMENTAL MARINE BIOLOGY AND ECOLOGY The role of the burrow funnel in feeding processes in the lugworm Arenicola marina (L.) A.S.W. Retraubun, M. Dawson, S.M. Evans* Department of Marine Sciences and Coastal Management, lJniversit,y of Newcastle upon Tyne. NE1 7RU, UK Received 29 March 1995; revised 21 December 1995; accepted 31 January 1996 Abstract Lugworms (Arenicoh marina) are typical marine deposit feeders (Jumars, 1993). Labile organic matter, notably bacteria, meiofauna and diatoms, is digested from the large volumes of nutritionally-poor sediment which are processed by the gut. Detritus is not evidently digested. However, it is trapped in the funnel of the burrow, and probably enhances the nutritional quality of the food by providing a substrate for bacterial growth. The worm’s irrigation current is also important because, if the headshaft of the burrow is blocked so that the current no longer reaches the funnel, there is a decrease in the numbers of bacteria there. Keywords: Lugworms; Arenicola marina; Feeding 1. Introduction Feeding strategies of marine deposit feeders, with the notable exceptions of the snail Hydrobia, and the fiddler crab Uca, are poorly understood (Jumars, 1993). This is even the case, for instance, in the lugworm Arenicoh marina (L.) which has been relatively intensively studied. Lugworms inhabit characteristically U-shaped burrows and have a substantial impact on the sediment by reworking it. Large volumes of this nutritionally poor food are ingested from the feeding pocket, which is at the base of the headshaft of the burrow, and the undigested remains are subsequently deposited as faecal casts on the surface, adjacent to the exit of the tailshaft. The diet itself has been subject to considerable debate. Thamdrup (1935) suggested that the worms fed unselectively on organic matter delivered to it via the headshaft of the burrow; Hylleberg (1975) thought *Corresponding author. 0022-0981/96/$15.00 0 1996 Elsevier Science B.V. All rights reserved PII SOO22-098 1(96)00017-2
Transcript of The role of the burrow funnel in feeding processes in the lugworm Arenicola marina (L.)

ELSEVIER Journal of Experimental Marine Biology and Ecology,
202 (1996) 107-l 18
JOURNAL OF EXPERIMENTAL MARINE BIOLOGY AND ECOLOGY
The role of the burrow funnel in feeding processes in the lugworm Arenicola marina (L.)
A.S.W. Retraubun, M. Dawson, S.M. Evans*
Department of Marine Sciences and Coastal Management, lJniversit,y of Newcastle upon Tyne. NE1 7RU, UK
Received 29 March 1995; revised 21 December 1995; accepted 31 January 1996
Abstract
Lugworms (Arenicoh marina) are typical marine deposit feeders (Jumars, 1993). Labile organic
matter, notably bacteria, meiofauna and diatoms, is digested from the large volumes of
nutritionally-poor sediment which are processed by the gut. Detritus is not evidently digested. However, it is trapped in the funnel of the burrow, and probably enhances the nutritional quality of the food by providing a substrate for bacterial growth. The worm’s irrigation current is also
important because, if the headshaft of the burrow is blocked so that the current no longer reaches
the funnel, there is a decrease in the numbers of bacteria there.
Keywords: Lugworms; Arenicola marina; Feeding
1. Introduction
Feeding strategies of marine deposit feeders, with the notable exceptions of the snail
Hydrobia, and the fiddler crab Uca, are poorly understood (Jumars, 1993). This is even the case, for instance, in the lugworm Arenicoh marina (L.) which has been relatively intensively studied. Lugworms inhabit characteristically U-shaped burrows and have a
substantial impact on the sediment by reworking it. Large volumes of this nutritionally poor food are ingested from the feeding pocket, which is at the base of the headshaft of the burrow, and the undigested remains are subsequently deposited as faecal casts on the
surface, adjacent to the exit of the tailshaft. The diet itself has been subject to considerable debate. Thamdrup (1935) suggested that the worms fed unselectively on organic matter delivered to it via the headshaft of the burrow; Hylleberg (1975) thought
*Corresponding author.
0022-0981/96/$15.00 0 1996 Elsevier Science B.V. All rights reserved
PII SOO22-098 1(96)00017-2

108 A.S.W. Retraubun et al. I J. Exp. Mar. Bid. Ed. 202 (1996) 107-118
that meiofauna were the main source of food but that worms also benefitted from the direct uptake of nutrients; and Cad&e (1976) and Rijken (1979) suggested that diatoms were important food items. More recently, evidence has accumulated that bacteria form a substantial part of the diet (Boon et al., 1978; Rijken, 1979; Grossman and Reichardt,
1991). Any mechanism which enhanced the nutritional quality of the food would be
beneficial and the suggestion by Hylleberg (1975) that A. marina “gardens” by culturing meiofauna in the headshaft of the burrow attracted considerable interest. He
proposed, on the basis of observations of Abarenicolu spp., that high nutrient levels at
the base of the headshaft (from the lugworm’s excretions, decaying plant matter and
diffusion from the surrounding anoxic area), coupled with the supply of oxygen from the irrigation current, stimulated bacterial growth. This increased microbial content then attracted large numbers of nematodes, flagellates and ciliates, which were consumed and digested by the lugworm. Rijken ( 1979) argued that gardening could not occur in A.
marina burrows because ingestion rates of headshaft sediment were too fast to allow for
significant growth of bacteria. However, Hylleberg (1975) believed that a feeding pause of 6 h would be sufficient to allow doubling of many bacteria, and he had observed a suspension of feeding in Aburenicolu p@ficu for periods of as much as 24 h. Similarly,
Cadee ( 1976) described periods of up to five days in which A. marina did not feed.
There has also been disagreement on the definition of gardening. Hylleberg (1975) originally described it as the process of stimulating growth of bacteria, nematodes,
flagellates and ciliates and their subsequent use as food. Similarly, Grossman and Reichardt (1991) considered it to be any growth-promoting effect of burrowing macrofauna on copiotrophic sediment bacteria. However, Plante et al. (1990) suggested
that the nutritional benefit must be from direct consumption of increased microbial abundance and not mediated through the food web (termed “kitchen garden(ing)” by
Fenchel and Finlay (1989)). The objectives of the present study were to: (i) determine the principal components of
the diet of A. marina by taking samples of sediment from stages in its passage along the burrow system and worm’s digestive tract; and (ii) investigate the role of the funnel of
the burrow in enhancing growth of food organisms.
2. Methods
The study was carried out in 1993 and 1994. Field studies were made on a population
of A. marina inhabiting the shore at Goswick Sands, Lindisfarne (Northumberland). It is a low energy shore which supports a large permanent population of lugworms. Worms used in laboratory studies were also collected from this site.
Measurements of the dimensions of burrows and related structures were made in October 1993. Burrows were prised open by inserting a spade into the sediment about 10 cm either from either the headshaft or the tailshaft. The sediment tended to sheer across the burrow’s shafts as pressure was applied to the spade’s handle. In addition, redox potentials were measured using a Jenway 3071 pH meter with appropriate probes. They were measured at 2 cm intervals (n = 10 in each category): (i) vertically down the

A.S.W. Retraubun et al. I .I. Exp. Mar. Bid. Ed. 202 (1996) 107-118 109
headshaft of the burrow, (ii) vertically down sediment approximately 10 cm from the
burrow, (iii) horizontally across sediment and the headshaft of the burrow at a depth of
about 10 cm below the surface and (iv) horizontally across the surface sediment
including the funnel of the burrow.
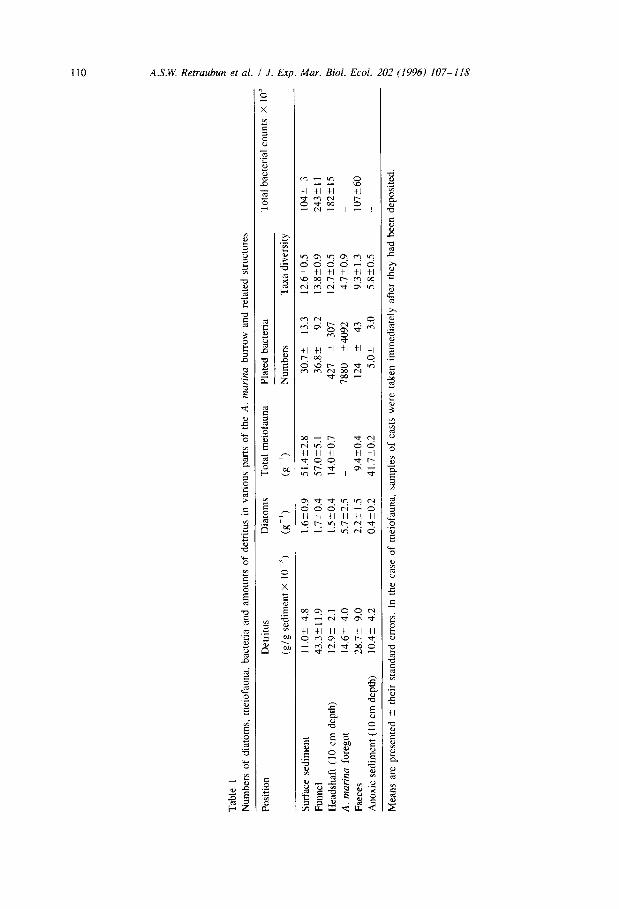
Diet was investigated by measuring changes that occurred in the organic content of sediment as it passed through the burrow system and the worm’s gut. Samples of sediment (n = 10 in each category; see Table 1) were collected in November 1992 by
using a spatula. As far as possible, samples were restricted entirely to the part of the burrow indicated (funnel or headshaft) and were not contaminated with any surrounding sediment. Each sample consisted of about 5-10 g of sediment. Foregut samples were
taken from worms (n = IO) which had been killed by plunging them into boiling water. All samples were processed within 24 h of collection. Procedures for treating them are
described below.
2.1. Diatoms
Diatoms were extracted from 1 g sediment subsamples (one from each sample) on glass microscope slides by covering them first with filter paper and then a glass cover
slip. These samples were left overnight under illumination. Light was provided by a standard 60 W bulb in a bench lamp 30 cm above the slides. Diatoms migrated upwards
and became attached to the cover slip. The following morning coverslips were removed, mounted on clean slides and the numbers of diatoms in ten microscopic fields of view
(magnification X 400) counted for each of them.
2.2. Maiofauna
1 g subsamples of sediment (one from each sample) were flooded with solutions of magnesium chloride in order to anaesthetise the meiofauna, which were then preserved and stained with Rose Bengal in 4% formalin in seawater. They were stirred in a
magnetic stirrer for 10 min, and the supernatant liquid filtered through a 63 km sieve.
This procedure was repeated three times for each sample. Counts were made of meiofauna in the sediment, observed under a binocular microscope.
2.3. Detritus
Subsamples of approximately 1 g (one from each sample) sediment were immersed in seawater in petri dishes and observed under a binocular microscope. Pieces of detritus were picked out of them during ten one-minute searches. Subsequently dry weights of
extracted detritus were measured.
2.4. Bacteria
Plate counts of aerobic heterotrophic bacteria were obtained on a marine agar
containing 0.7 g of neopeptone, 0.2 g of glucose, 0.1 g of yeast extract and 1.5 g of agar in 1 litre of artificial seawater. Using a plate spread technique, 0.1 cm3 portions of serial
Tab
le
1 N
umbe
rs
of
diat
oms,
m
eiof
auna
, ba
cter
ia
and
amou
nts
of
detr
itus
in
vari
ous
part
s of
th
e A
. m
arin
a bu
rrow
an
d re
late
d st
ruct
ures
Posi
tion
Det
ritu
s D
iato
ms
Tot
al
mei
ofau
na
Plat
ed
bact
eria
T
otal
ba
cter
ial
coun
ts
X I
O’
(g/g
se
dim
ent
X I
O-‘
) (g
-l)
(g-l
) N
umbe
rs
Tax
a di
vers
ity
Surf
ace
sedi
men
t 1
I.05
4.
8 1.
650.
9 51
.4k2
.8
30.7
5 13
.3
12.6
kO.5
10
4+
3 Fu
nnel
43
.3”
11.9
1.
720.
4 57
.0t5
.1
36.8
t 9.
2 13
.8k0
.9
243k
ll H
eads
haft
(1
0 cm
de
pth)
12
.92
2.1
1.52
0.4
14.O
kO.7
42
7 t
307
12.7
kO.5
18
2t15
A
. m
arin
a fo
regu
t 14
.62
4.0
5.72
2.5
- 78
80
k409
2 4.
720.
9 _
Faec
es
28.7
2 9.
0 2.
22
1.5
9.4k
O.4
12
4 2
43
9.32
1.3
1072
60
Ano
xic
sedi
men
t ( 1
0 cm
de
pth)
10
.4+
4.2
0.4k
O.2
41
.7Z
0.2
5.O
k 3.
0 5.
850.
5
Mea
ns
are
pres
ente
d 2
thei
r st
anda
rd
erro
rs.
In t
he
case
of
m
eiof
auna
, sa
mpl
es
of
cast
s w
ere
take
n im
med
iate
ly
afte
r th
ey
had
been
de
posi
ted.

A.S.W. Retraubun et al. I J. Exp. Mar. Bid. Ed. 202 (1996) 107-118 III
solutions of sediment suspensions ( 1 g sediment shaken in 10 cm3 of water) were
applied to the agar in duplicate. Colonies of bacteria were counted after 3 wk of
incubation at 25°C. The most frequent bacterial types were identified on the basis of the
shape and colour of colonies, and standard Gram stain, catalase and oxidase tests. In view of the paucity of information on marine bacteria, no attempt was made to identify
individual taxa. Total bacterial counts were made from 2 cm3 sediment subsamples (one from each
sample) taken with a plastic syringe. They were preserved on collection in 2% formalin in filtered seawater. The sample was shaken by hand, put into a universal sterile bottle and 10 cm3 of 0.1% sodium cholate was added, together with 5 glass beads (2.0-3.0
mm in diameter) and 2 cm3 of dewex. The sample was shaken in an orbital shaker at 100 t-pm for 2 h, and then centrifuged at 2500 rpm for 10 min. 1 cm3 of the supernatant fluid was placed in an eppendorf tube and microfuged at 1300 rpm for 10 min, before 0.3 cm3 of the detergent NP-40 (in PBS) was added. Three 0.5 cm3 subsamples were taken and placed on gelatin-treated slides. They were dehydrated successively in 50, 80 and 95%
ethanol, allowing three min for each treatment. Numbers of bacteria were counted, using
an epifluorescent microscope, after staining them with 0.01% acridine orange. Counts were made in ten microscopic fields for each subsample at a magnification of X 1000.
A field experiment was carried out to investigate the effects of preventing the lugworm’s irrigation current from reaching the sediment in the funnel of the headshaft of
the burrow on the growth of bacteria in it. This was done by pushing Perspex sheets 10 cm X 5 cm, obliquely into the sediment adjacent to burrow funnels so that they cut across the headshaft separating the funnel from the rest of the burrow. There were eight experimental burrows and eight controls which were not treated in this way. Experimen-
tal and control burrows were paired on the basis of their proximity to one another. Sediment samples were taken for total bacterial counts from the funnels of burrows of
both experimental and control groups when the experiment was set-up and approximate-
ly 48 h later. Means are quotedktheir standard errors (SE.) throughout this paper. Unless stated
otherwise, the t-test has been used to determine levels of significance between means.
3. Results
The headshafts of A. marina burrows were characterised by cone-shaped depressions forming funnels in the surface sediment, and the tailshafts by mounds of faecal casts
(Fig. 1). Although these surface structures were often destroyed by wave action, they had a substantial impact on the topography of the surface during calm weather. Algal debris, such as pieces of frond or thallus of macroalgae, tended to collect in the funnels. The dry weight of algal debris collected from them (X = 0.0163+0.0020 g per g
sediment, n = 22) was significntly greater than that from equivalent areas of surface sediment 10 cm away from burrows (X = 0.0008-+0.0003 g per g sediment; n = 22) (P < 0.001, Wilcoxon matched-pairs signed-ranks test).
Headshafts of burrows extended deep into the dark anoxic sediment but, since they were usually filled with the light coloured sediment from the surface, they were

112 A.S.W. Retraubun et al. I J. Exp. Mar. Biol. Ecol. 202 (1996) 107-118
RPD _ I_---_ ---------a_- -m
ts -
- hs 0.33*0*03
0~35*0~03
__
21.0 27.6
Fig. I. Measurements (n = 10 in each case) of various parts of the burrow system of A. marina. Abbreviations:
f = funnel; hs = headshaft; fp = feeding pocket; hg = horizontal gallery; ts = tailshaft; fc = faecal cast; RPD =
redox discontinuity layer.
conspicuous when burrows were prised open (see Section 2). The surface sediment had high redox potentials, which declined gradually with increasing depth beneath the
surface, until the Redox Potential Discontinuity layer (RPD), the boundary between the light coloured surface and the dark anoxic sediment, was reached. Here redox values
decreased rapidly with increasing depth (Fig. 2). Redox values in burrow funnels were higher than those in adjacent surface sediment. They were also higher down the headshaft of the burrow (remaining at surface levels) than in adjacent sediment (Fig. 2).
Sediment in the study area was rich in diatoms, meiofauna, detritus and bacteria, all of
which were potentially available as food for A. marina. Evidence that they were utilised as such is as follows:
3.1. Diatoms
Diatoms belonging primarily to the genera Navicula, Rhizoselenia, Raphoneis,
Amphora, Nitzchia and Caloneis were common in surface sediment. They were
evidently transported down the headshaft of burrows with the descending column of sediment because they were present at similar densities in samples from the headshaft as
in those from the funnel and surface (Table 1). There is evidence of selective ingestion by the worms because the density in the foregut was significantly higher than that in the headshaft of the burrow (P < 0.05). There was a decline in the abundance of diatoms between the foregut and faecal casts. Furthermore, most diatoms in casts were empty, the contents evidently had been digested.

A.S.W. Retraubun et al. I J. Exp. Mar. Bid. Ed. 202 (1996) 107-11X
Depth profales Horrzontal profiles
200 1 WDown redlment l(c) locm below surface
2oo 1 (bIDown headshaft
Redox 100
In”
0 5 10
Depth (cm)
3210123
(d)At surface
20 10 0 10 20
Distance from burrow(O)(cm)
II3
Fig. 2. Mean redox potentials at various positions in the headshaft and funnel of the burrow and in adjacent
sediment. (a) Vertical profile in sediment, (b) vertical profile down headshaft of burrow, (c) horizontal profile at
10 cm depth across headshaft of burrows, (d) horizontal profile at surface across funnel of burrow.
3.2. Meiofauna
Copepods, nematodes, ciliates and ostracods were the most numerous members of the meiofauna in the surface sediment but gastrotrichs, turbellarians and polychaetes were also recorded in substantial numbers (Table 2). Meiofauna originating from surface
sediment were unlikely to have been a major component of the diet, however, because relatively few of them were transported down the headshaft of the burrow. Nevertheless,
Table 2
Numbers (means ? standard errors, II= IO) of meiofauna (g ’ sediment) in major taxonomic groups taken from
A. marina burrows and related areas
Group Burrow
Funnel Headshaft( 10 cm depth)
Sediment depth Cast
Surface IO cm
Ciliata 2.92 1.2
Copepoda 42.322.1
Gastrotricha 0.620.2
Nematoda 4.92 I .4
Ostracoda 3.91 I.5
Polychaeta 0.320.2
Turbellaria 2.120.8
Total 57.025. I
1.8+-I.2
4.l?l.l
0.220.2
5.52 1.6 I .9?0.8
0.3kO.3
0.220.2
14.OkO.7
9.0*4.4
11.9+1.7
0.lto.I
22.7k7.0
4.6+ I.2
0.620.3
2.551.1
51.4+-2.8
I .9%0.8 0.7 kO.4
0. I 50. I 1.2-tO.5
0 0
4.1*1.4 3.720.8
0.2”0.2 1.7?l.l
0.21-0.2 0.220.2
1.0+0.1 1.820.6
7.520.5 9.420.4

114 A.S.W. Retraubun et al. I .I. Exp. Mar. Biol. Ecol. 202 (1996) 107-118
a significant decrease in numbers of meiofauna in the faecal casts compared with the
headshaft suggest that some of those present were digested (Table 1).
3.3. Detritus
At least some of the organic debris which collects in the funnels of burrows (see above) became incorporated into the sediment in the funnel. This was because there
were significantly larger quantities of it in samples of sediment from funnels than in those from surface sediment nearby (P < 0.02, Table 1). The detritus was not transported down the headshaft in substantial quantities because samples from the headshaft were
poorer in it than those taken from funnels (P < 0.01).
3.4. Bacteria
Bacteria are almost certainly a major source of food for A. marina. There is also evidence that they are cultured in burrows. This is as follows:
1. Bacteria were numerous in surface sediments, particularly that which was rich in detritus. Numbers of (plated) bacteria correlated with the amount of detritus in
sediment samples from both burrow funnels and nearby surface areas (Fig. 3).
Funnels acted as traps for detritus (see above) and, as would be expected if they were used as substrates for growth, bacteria were more abundant in funnels than in other
parts of the surface sediment (f’ < 0.01, Table 1).
Bacteria
x 10’ 0.004 0.008
. Futlnel rro~ss’ .
. .’ * 1
0.005 0 01 O-015 0 020
Detritus (g per g sediment)
Fig. 3. Spearman Rank correlations between numbers of plated bacteria and detritus in samples of (a) surface
sediment and (b) sediment from funnels of burrows. *P < 0.05.

A.S.W. Retraubun et al. / J. Exp. Mar. Bid. Ed. 202 (1996) 107-118 I15
2. Bacteria were evidently transported down the headshaft of the burrow with the
descending sediment, because densities (total counts) of them in headshafts were high
and not significantly different from those in funnels (P > 0.05, Table 1). Furthermore,
the population of bacteria in the headshaft was apparently the same as that at the surface. A total of 20 bacterial taxa were identified by plating techniques and each was recorded in samples from the headshaft, funnel and surface sediment (Fig. 4).
They were also recorded in similar relative proportions of abundance at each of these sites. The Kendall coefficient of concordance for their rank orders of abundance at them was 0.9022 (P < 0.001). The diversity of bacteria (i.e., number of taxa) was
also similar in samples taken from these places (i.e., numbers of taxa, Table 1). 3. The population of bacteria in the anoxic sediment was different from that in the
surface sediment (and almost certainly included anaerobic species which were not
identified by the plating techniques used here). The Spearman rank coefficient of
correlation between the rank orders of abundance of taxa in anoxic and surface
sediments was 0.3 122 (P > 0.05).
4. Bacteria were evidently digested in substantial numbers because abundances of plated bacteria in samples taken from the ingestive side of the burrow system (headshaft or foregut) were much higher than those on the egestive side (faecal casts, Table 1). Similarly, counts of total bacteria from the headshaft were higher than those from the
faeces.
Percent Head shaft I
of lb&
population
Anoxic sediment
5 i0 Ii 21 Taxon number
Fig. 4. Relative proportions of twenty bacterial taxa at different positions in the burrow system of A. marina.

116 A.S.W. Retraubun et al. / J. Exp. Mar. Biol. Ecol. 202 (1996) 107-I 18
Table 3
Mean numbers (5standard errors) of bacteria (total counts), in samples taken from the funnels of burrows
(n = 8 in each case) before and after the headshaft had been blocked by the insertion of a plastic sheet
Sample group Number of bacteria X 10’
Before After
Experimental
Control
187.8+12.4 135.4k7.6 P io.01
208.5? 6.3 21.5.0Z6.3 n.s.
Controls did not receive this treatment. Statistical comparisons were made using the Wilcoxon matched-pairs
signed-ranks test. n.s. = not significant.
There is evidence that the worm’s irrigation current is important in maintaining the
abundance of bacteria in the headshaft. There was a significant decrease in the density of bacteria in funnels of burrows 48 h after the insertion of a plastic sheet between the funnel and headshaft, but no significant change in densities in controls over the same period (Table 3).
4. Discussion
Feeding in the lugworm is characteristic of that of marine deposit feeders (Jumars,
1993). Large volumes of nutritionally-poor food (sediment) are processed in the gut, but since its residency time in the gut is short, only labile organic matter can be digested.
There is insufficient time to digest refractory components of the food. The throughput of sediment in A. marina is 4.7-80 cm3 per day (Cadee, 1976) and the residency time in
the gut is about 15 min (Kermack, 1955). Bacteria and diatoms are components of the diet because their numbers decreased during their passage through the gut (see also Grossman and Reichardt ( 1991)). They were also effective foods when they were supplied as artificial diets in the laboratory. Rijken (1979) found that either bacteria or
diatoms enhanced the growth of lugworms which were fed on them. Meiofauna trapped by the funnel, and then transported down the headshaft to the feeding pocket, are unlikely to form a significant part of the diet because they occur in such low numbers in
the headshaft. They probably escape ingestion by migrating upwards against the flow of the descending column of sediment. Nevertheless, a rich (and probably self-sustaining)
population of meiofauna occurs in the feeding pocket (Scherer, 198.5), and this is presumably ingested by lugworms. Detritus is evidently not digested and, since
lugworms do not secrete cellulase (Longbottom, 1970) it probably passes largely unaltered through the gut. Minced Ulva, fed to lugworms as artificial detritus, was utilised ineffectively by them (Rijken, 1979).
Pits in the surface substrate are sites of selective deposition of particles of low specific gravity (Yager et al., 1993). They could therefore improve the diets of deposit-feeding benthic organisms such as A. marina by, for example, trapping organic matter, including bacteria and/or detritus. Indeed, it was shown here that algal debris is trapped by the funnels of A. marina burrows. Its effect is evidently indirect, because although it is not digested, it probably forms a substrate for bacterial growth in the burrow system. As

A.S.W. Retraubun et al. I J. Exp. Mar. Bid. Ed. 202 (1996) 107-118 117
would be expected if this is the case, bacteria were present at higher densities in the
funnel than in adjacent sediment. This could be due to selective deposition of bacteria in the funnel but the reduction in the quantity of detritus between the funnel and headshaft
suggests that it is being utilised by bacteria. Furthermore, when the worm’s irrigation current was prevented from reaching it by inserting a Perspex sheet into the headshaft, the numbers of bacteria growing there decreased. This current, which involves the passage of about 40 cm3 of oxygenated water h-’ through the burrow in a posterior- anterior direction (Baumfalk, 1979) effectively extends the RPD layer deep into
otherwise anoxic sediment. It creates highly favourable conditions for the growth of
bacteria and the meiofauna which feeds on them (Scherer, 1985; Andersen and Kristensen, 1992; Kristensen et al., 1992). This system by which the lugworms trap indigestible detritus and then convert it into digestible bacteria (and/or meiofauna which consume the bacteria), can be described as “gardening” according to any of the
definitions which have been used previously (see Section 1). Selective ingestion of nutritive particles by the lugworm is an additional means of
improving the quality of its food. Baumfalk (1979) showed that small particles are
ingested at the expense of larger ones, which collect at the base of the headshaft, often forming a recognisable layer there. These small particles evidently include bacteria
which are present in the foregut in larger numbers than in the sediment of the headshaft (Grossman and Reichardt, 1991; present investigation). Similarly, diatoms appear to be ingested selectively because they too are concentrated in the foregut.
Biogenic structures, such as those created by A. marina burrows, have substantial impacts on the “neighbouring” interstitial fauna (e.g., Reise and Ax, 1979; Reise, 198 1;
Scherer, 1985). As Reise (1981) had shown earlier, the meiofauna inhabiting the funnels of A. marina burrows and casts differ from those occurring in nearby surface sediment. Recently-deposited casts are poor in meiofaunal abundance (present study), although
organisms undoubtedly invade them as they age (e.g., Reise, 198 1; Reise, 1985). It was shown both here, and by Reise (1981) that copepods aggregate in funnels which represent tiny pools when the tide is out; nematodes tend to avoid funnels and casts.
Acknowledgments
ASWR would like to express his thanks to the Indonesian Government’s Marine Sciences Education Project for funding. He also thanks the former Rector, Professor J.L.
Nanere, the former Dean of Fisheries Mr J.M. Nanlohy and the present Rector, Dr M. Huliselan, of Pattimura University (Indonesia) for their constant support and encourage- ment. Thanks are also due to Dr A.G. O’Donnell for his help in estimating numbers of bacteria.
References
Andersen, F.O. and E. Kristensen, 1992. The importance of benthic macrofauna in decomposition of
microalgae in a coastal marine sediment. Limnol. Oceanogr., Vol. 37, pp. 1392- 1403.

118 A.S.W. Retraubun et al. I J. Exp. Mar. Biol. Ecol. 202 (1996) 107-118
Baumfalk, Y.A., 1979. Heterogeneous grain size distribution in tidal flat sediment caused by bioturbation
activity of Arenicolu marina (Polychaeta). Neth. J. Sea Res., Vol. 13, pp. 428-440.
Boon, J.J., W. Liefkens, W.I.C. Rijpstra, M. Baas and J.W. De Leeuw, 1978. Fatty acids of Desulfovibrio
desulfuricans as marker molecules in sedimentary environments. In, Environmental Biogeochemisrry and
Geomicrobiology. Ann Arbor Science, Ann Arbor, MI, Vol. 1, pp. 355-372.
Cadte, G.C., 1976. Sediment reworking by Arenicola marina on tidal flats in the Dutch Wadden Sea. Neth. J. Sea Res., Vol. 10, pp. 440-460.
Fenchel, T. and B.J. Finlay, 1989. Kenrrophoros: a mouthless ciliate with a symbiotic kitchen garden.
Opehelia, Vol. 30, pp. 75-93.
Grossman, S. and W. Reichardt, 1991. Impact of Arenicola marinu on bacteria in intertidal sediments. Mar.
Ecol. frog. Ser., Vol. 77, pp. 85-93.
Hylleberg, J., 1975. Selective feeding by Abarenicolu pacijcu with notes on Abaraenicola vagabunda and a
concept of gardening in lugworms. Ophelia, Vol. 14, pp. 113-137.
Jumars, P.A., 1993. Gourmands of mud: diet selection in marine deposit feeders. In, Diet Selecrion: An
Inter-disciplinaq Approach to Foraging Behaviour, edited by R.N. Hughes, Blackwell, Oxford, pp.
124-156.
Kermack, D.M., 1955. The anatomy and physiology of the gut of the polychaete Arenicolu marina (L.). Proc.
Zool. Sot. Lond., Vol. 125, pp. 347-381.
Kristensen, E., F.O. Andersen and T.M. Blackbum, 1992. Effects of benthic macrofauna and temperature on
degradation of macroalgal detritus: the fate or organic carbon. Limnol. Oceanogr., Vol. 37, pp. 1404- 1419.
Longbottom, M.R., 1970. Distribution of the digestive enzymes in the gut of Arenicola marina. J. Mar. Biol.
Ass. U.K.,Vol. 50, pp. 121-128.
Plante, C.J., P.A. Jumars and J.A. Baross, 1990. Digestive associations between marine detritivores and
bacteria. Ann. Rev. Ecol. Syst.,Vol. 21, pp. 93-127.
Reise, K., 1981. High abundance of small zoobenthos around biogenic structures in tidal sediments of the
Wadden Sea. Helgol. wiss. Meeresunlers., Vol. 34, pp. 423-425.
Reise, K., 1985. Tidal Flat Ecology, Springer-Verlag, Berlin, 191 pp.
Reise, K. and P. Ax, 1979. A macrofaunal “thiobios” limited to the anaerobic sulfide system of marine sand
does not exist. Mar. Biol., Vol. 54, pp. 225-237.
Rijken, M., 1979. Food and food uptake in Arenicola marina. Neth. J. Sea Res., Vol. 13, pp. 406-421.
Scherer, B., 1985. Annual dynamics of a meiofaunal community from the sulphide layer of a North Sea sand
flat. Microfuunu marina., Vol. 2, pp. 117-161.
Thamdrup, H.M., 1935. Beitrage zur Gkologie der wattenfauna auf experimenteller Grundlage. Meddr. Kommn.
Danm. Fisk.og. Havunders. Serie Fisk& Vol. 10, pp. l-12.5.
Yager, P.L., A.R.M. Nowell and P.A. Jumars, 1993. Enhanced deposition to pits: a local food source for
benthos. .I. Mar. Res., Vol. 51, pp. 209-236.




















