
CASA MARINA HOTEL: 1924-2014 By Don Mabry djmabry.org/casa.pdf
Upload
independentCategory
view
1download
0
Journal of Experimental Marine Biology and Ecology 410 (2011) 45–52
Contents lists available at SciVerse ScienceDirect
Journal of Experimental Marine Biology and Ecology
j ourna l homepage: www.e lsev ie r .com/ locate / jembe
Burial of seeds and seedlings by the lugworm Arenicola marina hampers eelgrass(Zostera marina) recovery
Thomas Valdemarsen ⁎, Kim Wendelboe, Jonas T. Egelund, Erik Kristensen, Mogens R. FlindtInstitute of Biology, University of Southern Denmark, Campusvej 55, 5230 Odense M, Denmark
⁎ Corresponding author. Tel.: +45 26 39 94 44.E-mail addresses: [email protected], vald
(T. Valdemarsen).
0022-0981/$ – see front matter © 2011 Elsevier B.V. Alldoi:10.1016/j.jembe.2011.10.006
a b s t r a c t
a r t i c l e i n f oArticle history:Received 22 June 2011Received in revised form 4 October 2011Accepted 4 October 2011Available online 27 October 2011
Keywords:Ecosystem engineerEutrophicationHabitat lossRecolonizationSeagrassSediment reworking
Eelgrass (Zostera marina) used to dominate the vegetation in Odense Fjord, Denmark, and covered >17 km2
of the shallow fjord in 1983. Decades of excessive nutrient loading has lead to decreased eelgrass distribution,and only ~2 km2 is covered at present. The state of low eelgrass coverage has not changed despite significantimprovements of water quality in the past >10 years, and lugworms, Arenicola marina, have colonized theformer eelgrass areas (1–8 ind.m−2). It was hypothesized that the lack of eelgrass recoverywas due to A.marina,which was investigated by a combined field and laboratory approach. At a study site where eelgrass used todominate, a seasonal study of lugworm population dynamics and sediment reworking activity wasperformed. Additionally, density dependent burial of eelgrass seeds and seedlings due to sediment rework-ing by A. marina was investigated in mesocosm experiments. Our results indicate that A. marina may nega-tively impact eelgrass recovery, since sediment reworking lead to rapid burial of eelgrass seeds andseedlings; within 1–2 months, 95% of seeds and 75% of seedlings were buried below critical depth. Consid-erations based on empirical modeling suggest that negative impact occur even at low A. marina density(5–10 ind.m−2). Therefore the spread of A. marina into former eelgrass areas is critical, since eelgrass recov-ery may be severely impaired, even when water quality favors eelgrass recolonization.
© 2011 Elsevier B.V. All rights reserved.
1. Introduction
The lugworm, Arenicola marina L., is a common polychaete alongEuropean Atlantic coasts where it inhabits intertidal and subtidalsandy sediments at typical densities of 3–80 individuals m−2
(Cadée, 1976; Jones and Jago, 1993; Volkenborn and Reise, 2006).A. marina is an upward conveyor that feeds head-down at depth inthe sediment and it has an impressive capacity for particle reworking.For example, Cadée (1976) estimated thatA.marina is capable ofmixingthe upper 6–33 cm of the sediment per year in the Dutch WaddenSea, and similar estimates (1–18 cm y−1) are reported from manyother areas (e.g. Retraubun et al., 1996a; Riisgard and Banta, 1998;Valdemarsen et al., 2010b).
Arenicola marina is considered an important “ecosystem engineer”,since its intense particle mixing and associated physicochemicalchanges affect the biological properties of sediments. Since particleslarger than 1–2 mm are not ingested by A. marina (Jones and Jago,1993), the intensely mixed sediment inhabited by A. marina is oftencharacterized by a zone of homogeneous sandoverlying a zone of coars-er particles such as gravel and shells (Andresen and Kristensen, 2002).As a consequence of the constant disturbance of surface sediment, A.marina modifies the composition and abundance of other infauna by
rights reserved.
excluding for instance tube building polychaetes and crustaceans(Flach, 1992; Volkenborn and Reise, 2006, 2007). Additionally,A. marina interacts with the distribution of rooted marine macro-phytes. In areas where lugworms are present near beds of seagrasses(Zostera noltii and Z. marina) or cordgrass (Spartina anglica), almostno transition zone is observed between areas with vegetation andbare A. marina inhabited sediment. Such a distribution pattern mayonly occur if roots and rhizomes prevent the settling of A. marina orif sediment reworking by A. marina prevents growth of vegetativeshoots and seedlings (Philippart, 1994; Valdemarsen et al., 2010b;van Wesenbeeck et al., 2007). Competition between rooted macro-phytes and A. marina was also indicated by the spread of seagrassesinto large scale lugworm exclusion plots in the German WaddenSea (N. Volkenborn, personal comm.).
Does Arenicola marina also affect the recovery of eelgrass follow-ing its eutrophication driven decline along European and NorthAmerican coasts during the 20th century? Excessive nutrient loadingled to poor growth conditions for eelgrass, by lowering light availabil-ity and by stimulating hypoxic events and poor sediment conditions(Greve et al., 2005; Hauxwell et al., 2001; Mascaro et al., 2009;Valdemarsen et al., 2010a). This situation has been somewhat reversedduring the last decades, due to restrictions on nutrient loading(Petersen et al., 2009). The enhanced environmental quality, evi-denced by for instance reduced levels of phytoplankton and improvedsediment conditions, was expected also to facilitate eelgrass recovery.However, this appears not to be the case or only occurs slowly in

46 T. Valdemarsen et al. / Journal of Experimental Marine Biology and Ecology 410 (2011) 45–52
many areas, which indicates that other parameters than environmentalquality, for instance disturbance by benthic invertebrates, are criticalduring seagrass recovery (Dumbauld and Wyllie-Echeverria, 2003;Valdemarsen et al., 2010b).
We speculate that Arenicola marina enter/invade former eelgrassareas and effectively prevent eelgrass recovery.A.marinahas a high dis-persive potential with 50–400 juveniles m−2 during settling events andis therefore an effective colonizer of areas formerly vegetated by eel-grass (Flach and Beukema, 1994, Reise et al., 2001). This has been ob-served in Odense Fjord, Denmark, where 13 km2 that was covered byeelgrass in 1983 is now bare sediment inhabited by 1–8 A. marinam−2
(Fig. 1; E. Kristensen, Unpublished results). Moreover A. marina densityis up to 80 m−2 in other areas of Odense Fjord,where eelgrass dominat-ed in the beginning of the 20th century (Ostenfeldt, 1908).
The seed bank is critical for the reproductive cycle of seagrassessince it represents the recolonization potential (Jarvis and Moore,2010; Orth et al., 2006) and eelgrass recovery after large scale distur-bance may occur exclusively from seeds (Greve et al., 2005; Lee et al.,2007; Plus et al., 2003). Eelgrass seeds are, however, sensitive toburial, since the ability of seeds to germinate and develop into self-sustaining plants depends on the sediment depth where they are po-sitioned (Greve et al., 2005; Probert and Brenchley, 1999). If seeds ofZostera marina are buried below 5–6 cm depth due to for instancesediment reworking by Arenicola marina, the germination process willfail because the limited energy storage in seeds does not allow the hy-pocotyls to reach the sediment surface (Greve et al., 2005; Harrison,1993). Similarly, A. marina may lead to burial of eelgrassseedlings (Philippart, 1994; Valdemarsen et al., 2010b).
In this studywe evaluate the impact of lugworms (Arenicolamarina)on eelgrass (Zostera marina) reestablishment by a combined field andlaboratory approach. The A. marina population dynamics and seasonalsediment reworking activity were quantified at a study site in thenorthern part of Odense Fjord, where eelgrass is diminishing despite fa-vorable light conditions andwater quality. Additionally, A.marina burialof eelgrass seeds and seedlings was investigated in laboratory meso-cosm experiments at a fixed worm density (60 m−2). The resultsobtained from field and laboratory experimentswere combined to eval-uate the importance of A. marina for the reestablishment of eelgrass atthe study site. Furthermore, a simple empirical model for the assess-ment of A. marina impact during eelgrass reestablishment was devel-oped with worm size and density as input variables.
2. Materials and methods
2.1. Study site
The population characteristics and activity of Arenicola marina wasfollowed during 2009–2010 at Enebærodde in the outer part of OdenseFjord, Denmark (consult Valdemarsen et al., 2010b for location of study
Fig. 1. Left; in Odense Fjord, Denmark, the lugworm, Arenicola marina, has invaded formerfound in the same areas as A. marina. Right; when considering the size of fecal casts and fe
site). The area was extensively covered with eelgrass in the past and in1983 it consisted of a 7–8 km2 continuous eelgrass meadow thatcovered ~50% of the outer Odense Fjord (Fyns Amt, 2006). Eelgrasscoverage has since then declined dramatically in Odense Fjord (FynsAmt, 2006; Greve et al., 2005) andwas at the study site in 2009 reducedto a 400 mwide zone that extended ~100 m from the shore to 1–1.5 mwater depth. The remaining eelgrass vegetation in the area was patchyand covered only half of the surface, and small to intermediate eelgrasspatches (1–100 m2) were fractionated with areas of bare sediment.The infauna on bare sediment was dominated by the polychaetesA. marina (3–5 m−2) and Marenzelleria viridis (Quintana et al.,2011; Valdemarsen et al., 2010b).
2.2. Population characteristics and activity of Arenicola marina
The dynamics, size and distribution of the Arenicola marina popu-lation and its in situ reworking activity at the study site were evaluat-ed by combining various approaches. A laboratory experiment wasfirst conducted to obtain the relationship between A. marina sizeand feces diameter. This relationship was later used to estimate thesize distribution of the A. marina population based on underwaterphotographs of fecal casts taken during a 13 months survey. Thesephotographs were also used to monitor the seasonal variation of A.marina density. Finally, the seasonality of in situ sediment reworkingactivity was monitored during the same period to obtain an estimateof the sediment mixed by A. marina at the study site.
2.2.1. Relationship between Arenicola marina size and feces diameterA. marina (n=31) with different sizes were collected and acclima-
tized for ~4 days at 15 °C in buckets with sediment. The worms werethen carefully retrieved from the sediment and kept in aerated seawaterfor 24 h to ensure that they had voided their guts. Subsequently, theywere gently dried with a paper tissue and weighed to determine wetweights. The worms were hereafter placed individually in 25 cm indiameter (i.d.) plastic buckets containing 20 cm sediment and~2 cm overlying seawater. After 10–12 h, the newly produced fecalcasts were photographed together with a known scale. Photographswere registered as non-earth projections in GIS software (MapInfo)and the average diameter of feces was determined with 0.1 mm preci-sion. The data were used to generate a relationship between A. marinaweight and feces diameter, which formed the basis for evaluating thesize distribution of A.marinapopulation at the study site. The size distri-bution was based on photographs of fecal casts taken during the13 months survey of in situ reworking activity (n=142).
2.2.2. Arenicola marina densityThe density of active Arenicola marina on bare sediment in the vicin-
ity of eelgrass vegetation (b5 m distance) was determined on severaloccasions during 2009–2010 by photographing and counting the
eelgrass areas. Middle; eelgrass seedlings, which are critical for eelgrass recovery, areeding funnels negative impact of A. marina during eelgrass recovery seem likely.

47T. Valdemarsen et al. / Journal of Experimental Marine Biology and Ecology 410 (2011) 45–52
number of fecal casts using 0.25 m2 quadrants (n=50). This methodhas been frequently applied to study the seasonality of A.marinadensity(e.g. Cadée, 1976; Reise et al., 2001). It typically leads to 5–10% underes-timates, since a proportion of theworms are inactive at the time ofmea-surement (Flach and Beukema, 1994; Retraubun et al., 1996a).
2.2.3. Sediment reworking by Arenicola marinaIn situ sediment reworking activity of Arenicola marina in the vi-
cinity of eelgrass patches (0–5 m distance) was measured by themethod described in Valdemarsen et al. (2010b). On every samplingoccasion five 0.25 m2 quadrants were placed on bare sediment at~1 m water depth. The sediment surface framed by the quadrantswas smoothened to erase signs of old fecal casts. After 1–2 h incuba-tion, underwater photographs of newly deposited fecal casts wererecorded together with a known scale. Photographs were registeredin GIS software (MapInfo) and the length and width of feces were de-termined with 0.1 mm precision. The worm size was obtained fromthe correlation of feces diameter vs. worm size, and time specific sed-iment reworking activity normalized to worm wet weight(cm3 g d−1) was calculated assuming a cylindrical shape of feces.Reworking activity and water temperature was measured on 12 occa-sions from July 2009 to July 2010. No measurements were made fromJanuary to March 2010 due to ice cover at the study site. The dataobtained during the field survey formed the basis for a general corre-lation of reworking activity as a function of worm size. This correla-tion was normalized to the standard temperature of 15 °C as used insubsequent laboratory experiments.
2.3. Sampling of sediment and worms for laboratory experiments
Well sorted, organic poor sediment (~200 μmmedian grain size andb0.7% organic matter) for use in laboratory experiments was collectedat the study site on several occasions during 2009–2010. Sedimentwas collected at b1 m water depth, sieved through a 1 mm mesh toremove large particles and macrofauna, and homogenized prior toexperiments. The Arenicola marina used in laboratory experimentswere collected at the study site or Bregnør Bay in the Eastern part ofOdense Fjord (Papaspyrou et al., 2007). A. marina were acclimatizedfor 2–3 days in 15 °C aerated seawater before use in experiments.
2.4. Set-up for laboratory experiments
Laboratory experiments were conducted in an annular flume con-sisting of two transparent acrylic cylinders with diameter of 1.8 and1.4 m, respectively. These were fixed to a solid base forming an85 cm high circular tank with a 20.5 cm wide channel, a bottomarea of 1.03 m2 and a total volume of 740 L. During all experimentsthe flume was filled with 20 and 54 cm sediment and seawater (salin-ity of 20), respectively (corresponding to ~160 L sediment and ~470 Lwater). The sediment layer in the active current channel thus formeda 5.5 m long, 0.2 m wide and 0.2 m deep sediment layer, whichallowed the lugworms to move freely in the horizontal plane. Watercurrent was driven by a rotating lid with 8 equally distanced paddles.Experiments were conducted at constant velocity (~20 cm s−1) and
Table 1The average density, biomass (g wet weight) and reworking activity of Arenicola marina durby A. marina every month is shown (values from September 2009 to April 2010 are based on2010. Error values represent standard error.
2009
July August September
Density (m−2) 4.5±0.3 5.1±0.2 n.a.Worm size (g) 4.3±0.5 5.8±0.6 6.1±0.5Reworking activity (cm3 g−1 d−1) 3.5±0.3 4.0±0.3 3.0±0.3Daily reworking (cm3 m−2 d−1) 68±5.6 118±2.5 91±9.1Depth of reworked layer (cm month−1) 0.21 0.36 0.27
temperature (15 °C) at 12/12 h light/dark cycles (PAR during lightperiods was 100–120 μmol photons m−2 s−1).
Burial of Z. marina seeds by lugworms was investigated using seedmimics of different colors produced from nylon cord (1.5 mm diame-ter and density of 1 g cm−3). The nylon cord was cut into ~3 mmpieces (3.3±0.2 mm, n=100). The mimics thus simulated the size,shape and density (3.3–3.8 mm length and density of ~1.1 g cm−3)reported for natural Z. marina seeds (Wyllie-Echeverria et al., 2003).More than 9000 seed mimics with different colors (white, blue andred) were prepared.
2.5. Burial of eelgrass seeds by Arenicola marina
The impact of lugworms on seed burial was investigated in theannular flume with seed mimics of different colors randomly distrib-uted at 0, 7 and 15 cm sediment depth before addition of worms.While sediment was added to the flume, ~3000 white seed mimicswere randomly distributed at 15 cm depth and then another ~3000blue seed mimics were distributed at 7 cm depth. Finally, ~3000 redseed mimics were randomly distributed at the sediment surface(0 cm depth) before water was gently added. The sediment in theflume was consolidated for 6 days before 60 individuals of A. marinawere added, which corresponded to a density of 60 ind.m−2. Theflume was then incubated for 65 days while sediment cores weresampled initially and after 36 and 65 days (n=5, 8 and 8, respective-ly). Care was taken to not disturb the surrounding sediment duringinitial and intermediate sampling of sediment cores. During sampling,a 10 cm i.d. tube was inserted into the sediment and the enclosed sur-face was sampled with an 8 cm i.d. core liner. The hole left after coresampling was filled with homogenized sediment to ambient heightbefore the 10 cm i.d. tube was retrieved. The positions of initial andintermediate samplings were marked on the flume wall and wereavoided during the final sampling. The vertical distribution of seedmimics was determined by sectioning sediment cores into 1 cmslices. Every slice was sieved through a 1 mm mesh and retainedseed mimics were sorted into colors. Since A. marina had a dramaticimpact on the surface topography (±1–4 cm relative to the initial20 cm sediment depth), the sampled sediment cores had unequalheight. To correct for this fauna induced bathymetric effect, the verti-cal seed distribution was aligned relative to the bottom of the sedi-ment cores.
The density of live Arenicola marina was monitored regularly bycounting the number of fecal casts in the flume. Additionally, the A. ma-rina sediment reworking activity was determined for 10 worms after30 days. The flume was stopped and old fecal casts were erased withthe least possible disturbance of the surrounding sediment. After1–2 h, newly produced fecal casts were photographed and the rework-ing activity and A. marina size was determined by digital photographicanalysis. Hereafter the water velocity was restored to ~20 cm s−1.
2.6. Burial of eelgrass seedlings by Arenicola marina
Zostera marina seedlings (n=19) were collected at the study sitein June 2010, which was roughly 1 month after germination.
ing 2009–2010 at Enebærodde in Odense Fjord. The depth of the sediment layer mixeda 5 m−2 density of A. marina). Density was not measured from September 2009 to April
2010
December April May June July
n.a. n.a. 5.0±0.5 10.9±0.6 10.7±0.36.1±0.5 10.1±0.5 10.1±0.5 8.7±0.4 9.6±0.32.0±0.2 3.0±0.3 2.8±0.2 3.5±0.3 3.6±0.461±7.3 151±14 141±11 332±33 370±390.19 0.45 0.44 0.99 1.15
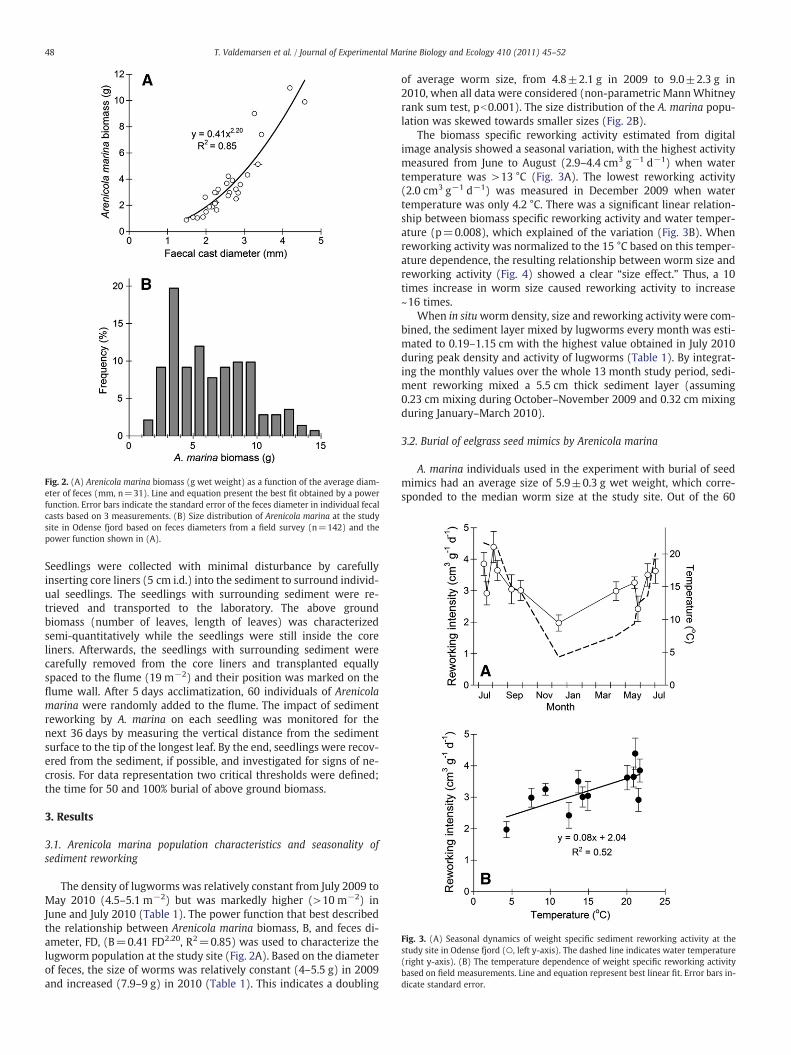
Fig. 2. (A) Arenicola marina biomass (g wet weight) as a function of the average diam-eter of feces (mm, n=31). Line and equation present the best fit obtained by a powerfunction. Error bars indicate the standard error of the feces diameter in individual fecalcasts based on 3 measurements. (B) Size distribution of Arenicola marina at the studysite in Odense fjord based on feces diameters from a field survey (n=142) and thepower function shown in (A).
Fig. 3. (A) Seasonal dynamics of weight specific sediment reworking activity at thestudy site in Odense fjord (○, left y-axis). The dashed line indicates water temperature(right y-axis). (B) The temperature dependence of weight specific reworking activitybased on field measurements. Line and equation represent best linear fit. Error bars in-dicate standard error.
48 T. Valdemarsen et al. / Journal of Experimental Marine Biology and Ecology 410 (2011) 45–52
Seedlings were collected with minimal disturbance by carefullyinserting core liners (5 cm i.d.) into the sediment to surround individ-ual seedlings. The seedlings with surrounding sediment were re-trieved and transported to the laboratory. The above groundbiomass (number of leaves, length of leaves) was characterizedsemi-quantitatively while the seedlings were still inside the coreliners. Afterwards, the seedlings with surrounding sediment werecarefully removed from the core liners and transplanted equallyspaced to the flume (19 m−2) and their position was marked on theflume wall. After 5 days acclimatization, 60 individuals of Arenicolamarina were randomly added to the flume. The impact of sedimentreworking by A. marina on each seedling was monitored for thenext 36 days by measuring the vertical distance from the sedimentsurface to the tip of the longest leaf. By the end, seedlings were recov-ered from the sediment, if possible, and investigated for signs of ne-crosis. For data representation two critical thresholds were defined;the time for 50 and 100% burial of above ground biomass.
3. Results
3.1. Arenicola marina population characteristics and seasonality ofsediment reworking
The density of lugworms was relatively constant from July 2009 toMay 2010 (4.5–5.1 m−2) but was markedly higher (>10 m−2) inJune and July 2010 (Table 1). The power function that best describedthe relationship between Arenicola marina biomass, B, and feces di-ameter, FD, (B=0.41 FD2.20, R2=0.85) was used to characterize thelugworm population at the study site (Fig. 2A). Based on the diameterof feces, the size of worms was relatively constant (4–5.5 g) in 2009and increased (7.9–9 g) in 2010 (Table 1). This indicates a doubling
of average worm size, from 4.8±2.1 g in 2009 to 9.0±2.3 g in2010, when all data were considered (non-parametric MannWhitneyrank sum test, pb0.001). The size distribution of the A. marina popu-lation was skewed towards smaller sizes (Fig. 2B).
The biomass specific reworking activity estimated from digitalimage analysis showed a seasonal variation, with the highest activitymeasured from June to August (2.9–4.4 cm3 g−1 d−1) when watertemperature was >13 °C (Fig. 3A). The lowest reworking activity(2.0 cm3 g−1 d−1) was measured in December 2009 when watertemperature was only 4.2 °C. There was a significant linear relation-ship between biomass specific reworking activity and water temper-ature (p=0.008), which explained of the variation (Fig. 3B). Whenreworking activity was normalized to the 15 °C based on this temper-ature dependence, the resulting relationship between worm size andreworking activity (Fig. 4) showed a clear “size effect.” Thus, a 10times increase in worm size caused reworking activity to increase~16 times.
When in situworm density, size and reworking activity were com-bined, the sediment layer mixed by lugworms every month was esti-mated to 0.19–1.15 cm with the highest value obtained in July 2010during peak density and activity of lugworms (Table 1). By integrat-ing the monthly values over the whole 13 month study period, sedi-ment reworking mixed a 5.5 cm thick sediment layer (assuming0.23 cm mixing during October–November 2009 and 0.32 cm mixingduring January–March 2010).
3.2. Burial of eelgrass seed mimics by Arenicola marina
A. marina individuals used in the experiment with burial of seedmimics had an average size of 5.9±0.3 g wet weight, which corre-sponded to the median worm size at the study site. Out of the 60
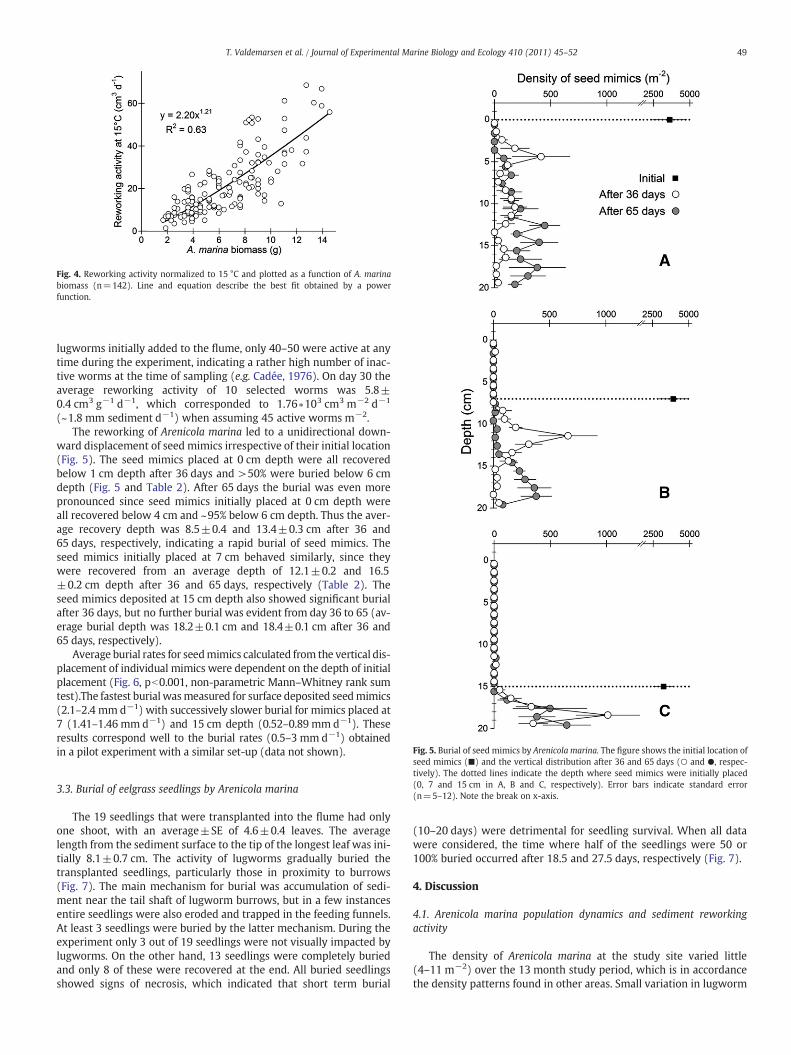
Fig. 4. Reworking activity normalized to 15 °C and plotted as a function of A. marinabiomass (n=142). Line and equation describe the best fit obtained by a powerfunction.
Fig. 5. Burial of seed mimics by Arenicola marina. The figure shows the initial location of
49T. Valdemarsen et al. / Journal of Experimental Marine Biology and Ecology 410 (2011) 45–52
lugworms initially added to the flume, only 40–50 were active at anytime during the experiment, indicating a rather high number of inac-tive worms at the time of sampling (e.g. Cadée, 1976). On day 30 theaverage reworking activity of 10 selected worms was 5.8±0.4 cm3 g−1 d−1, which corresponded to 1.76∗103 cm3 m−2 d−1
(~1.8 mm sediment d−1) when assuming 45 active worms m−2.The reworking of Arenicola marina led to a unidirectional down-
ward displacement of seed mimics irrespective of their initial location(Fig. 5). The seed mimics placed at 0 cm depth were all recoveredbelow 1 cm depth after 36 days and >50% were buried below 6 cmdepth (Fig. 5 and Table 2). After 65 days the burial was even morepronounced since seed mimics initially placed at 0 cm depth wereall recovered below 4 cm and ~95% below 6 cm depth. Thus the aver-age recovery depth was 8.5±0.4 and 13.4±0.3 cm after 36 and65 days, respectively, indicating a rapid burial of seed mimics. Theseed mimics initially placed at 7 cm behaved similarly, since theywere recovered from an average depth of 12.1±0.2 and 16.5±0.2 cm depth after 36 and 65 days, respectively (Table 2). Theseed mimics deposited at 15 cm depth also showed significant burialafter 36 days, but no further burial was evident from day 36 to 65 (av-erage burial depth was 18.2±0.1 cm and 18.4±0.1 cm after 36 and65 days, respectively).
Average burial rates for seedmimics calculated from the vertical dis-placement of individual mimics were dependent on the depth of initialplacement (Fig. 6, pb0.001, non-parametric Mann–Whitney rank sumtest).The fastest burial wasmeasured for surface deposited seedmimics(2.1–2.4 mm d−1) with successively slower burial for mimics placed at7 (1.41–1.46 mm d−1) and 15 cm depth (0.52–0.89 mm d−1). Theseresults correspond well to the burial rates (0.5–3 mm d−1) obtainedin a pilot experiment with a similar set-up (data not shown).
seed mimics (■) and the vertical distribution after 36 and 65 days (○ and ●, respec-tively). The dotted lines indicate the depth where seed mimics were initially placed(0, 7 and 15 cm in A, B and C, respectively). Error bars indicate standard error(n=5–12). Note the break on x-axis.
3.3. Burial of eelgrass seedlings by Arenicola marina
The 19 seedlings that were transplanted into the flume had onlyone shoot, with an average±SE of 4.6±0.4 leaves. The averagelength from the sediment surface to the tip of the longest leaf was ini-tially 8.1±0.7 cm. The activity of lugworms gradually buried thetransplanted seedlings, particularly those in proximity to burrows(Fig. 7). The main mechanism for burial was accumulation of sedi-ment near the tail shaft of lugworm burrows, but in a few instancesentire seedlings were also eroded and trapped in the feeding funnels.At least 3 seedlings were buried by the latter mechanism. During theexperiment only 3 out of 19 seedlings were not visually impacted bylugworms. On the other hand, 13 seedlings were completely buriedand only 8 of these were recovered at the end. All buried seedlingsshowed signs of necrosis, which indicated that short term burial
(10–20 days) were detrimental for seedling survival. When all datawere considered, the time where half of the seedlings were 50 or100% buried occurred after 18.5 and 27.5 days, respectively (Fig. 7).
4. Discussion
4.1. Arenicola marina population dynamics and sediment reworkingactivity
The density of Arenicola marina at the study site varied little(4–11 m−2) over the 13 month study period, which is in accordancethe density patterns found in other areas. Small variation in lugworm
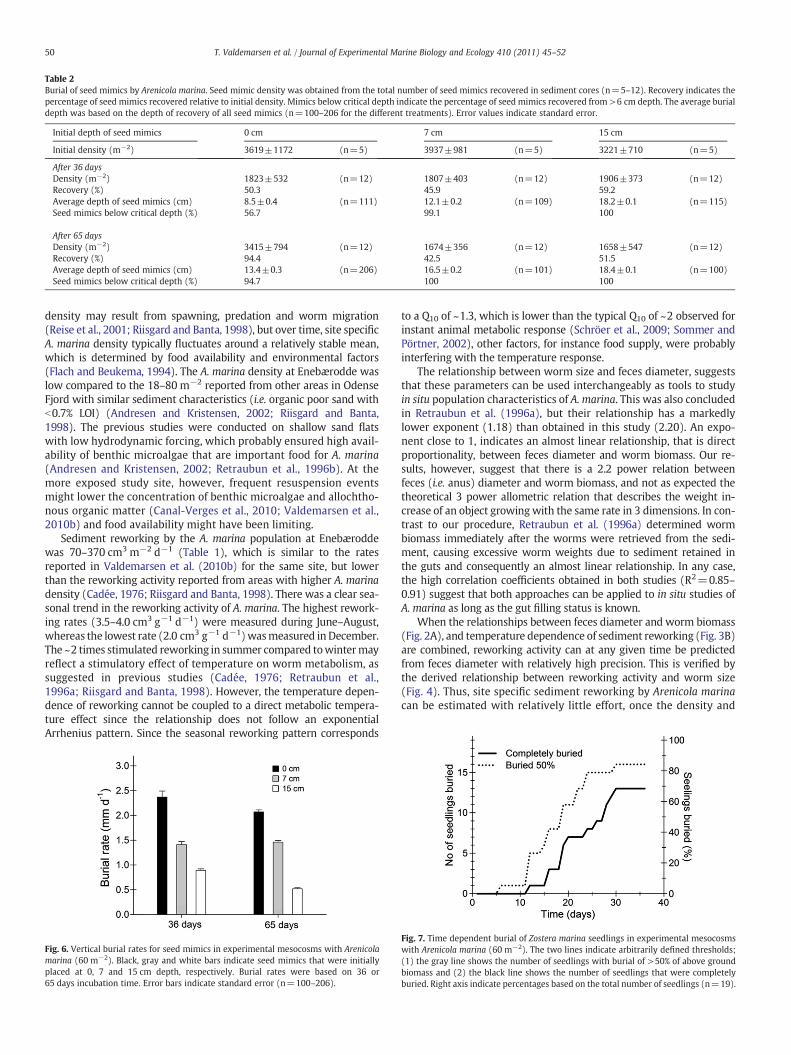
Table 2Burial of seed mimics by Arenicola marina. Seed mimic density was obtained from the total number of seed mimics recovered in sediment cores (n=5–12). Recovery indicates thepercentage of seed mimics recovered relative to initial density. Mimics below critical depth indicate the percentage of seed mimics recovered from >6 cm depth. The average burialdepth was based on the depth of recovery of all seed mimics (n=100–206 for the different treatments). Error values indicate standard error.
Initial depth of seed mimics 0 cm 7 cm 15 cm
Initial density (m−2) 3619±1172 (n=5) 3937±981 (n=5) 3221±710 (n=5)
After 36 daysDensity (m−2) 1823±532 (n=12) 1807±403 (n=12) 1906±373 (n=12)Recovery (%) 50.3 45.9 59.2Average depth of seed mimics (cm) 8.5±0.4 (n=111) 12.1±0.2 (n=109) 18.2±0.1 (n=115)Seed mimics below critical depth (%) 56.7 99.1 100
After 65 daysDensity (m−2) 3415±794 (n=12) 1674±356 (n=12) 1658±547 (n=12)Recovery (%) 94.4 42.5 51.5Average depth of seed mimics (cm) 13.4±0.3 (n=206) 16.5±0.2 (n=101) 18.4±0.1 (n=100)Seed mimics below critical depth (%) 94.7 100 100
50 T. Valdemarsen et al. / Journal of Experimental Marine Biology and Ecology 410 (2011) 45–52
density may result from spawning, predation and worm migration(Reise et al., 2001; Riisgard and Banta, 1998), but over time, site specificA. marina density typically fluctuates around a relatively stable mean,which is determined by food availability and environmental factors(Flach and Beukema, 1994). The A. marina density at Enebærodde waslow compared to the 18–80 m−2 reported from other areas in OdenseFjord with similar sediment characteristics (i.e. organic poor sand withb0.7% LOI) (Andresen and Kristensen, 2002; Riisgard and Banta,1998). The previous studies were conducted on shallow sand flatswith low hydrodynamic forcing, which probably ensured high avail-ability of benthic microalgae that are important food for A. marina(Andresen and Kristensen, 2002; Retraubun et al., 1996b). At themore exposed study site, however, frequent resuspension eventsmight lower the concentration of benthic microalgae and allochtho-nous organic matter (Canal-Verges et al., 2010; Valdemarsen et al.,2010b) and food availability might have been limiting.
Sediment reworking by the A. marina population at Enebæroddewas 70–370 cm3 m−2 d−1 (Table 1), which is similar to the ratesreported in Valdemarsen et al. (2010b) for the same site, but lowerthan the reworking activity reported from areas with higher A. marinadensity (Cadée, 1976; Riisgard and Banta, 1998). There was a clear sea-sonal trend in the reworking activity of A. marina. The highest rework-ing rates (3.5–4.0 cm3 g−1 d−1) were measured during June–August,whereas the lowest rate (2.0 cm3 g−1 d−1)wasmeasured in December.The ~2 times stimulated reworking in summer compared towintermayreflect a stimulatory effect of temperature on worm metabolism, assuggested in previous studies (Cadée, 1976; Retraubun et al.,1996a; Riisgard and Banta, 1998). However, the temperature depen-dence of reworking cannot be coupled to a direct metabolic tempera-ture effect since the relationship does not follow an exponentialArrhenius pattern. Since the seasonal reworking pattern corresponds
Fig. 6. Vertical burial rates for seed mimics in experimental mesocosms with Arenicolamarina (60 m−2). Black, gray and white bars indicate seed mimics that were initiallyplaced at 0, 7 and 15 cm depth, respectively. Burial rates were based on 36 or65 days incubation time. Error bars indicate standard error (n=100–206).
to a Q10 of ~1.3, which is lower than the typical Q10 of ~2 observed forinstant animal metabolic response (Schröer et al., 2009; Sommer andPörtner, 2002), other factors, for instance food supply, were probablyinterfering with the temperature response.
The relationship between worm size and feces diameter, suggeststhat these parameters can be used interchangeably as tools to studyin situ population characteristics of A. marina. This was also concludedin Retraubun et al. (1996a), but their relationship has a markedlylower exponent (1.18) than obtained in this study (2.20). An expo-nent close to 1, indicates an almost linear relationship, that is directproportionality, between feces diameter and worm biomass. Our re-sults, however, suggest that there is a 2.2 power relation betweenfeces (i.e. anus) diameter and worm biomass, and not as expected thetheoretical 3 power allometric relation that describes the weight in-crease of an object growing with the same rate in 3 dimensions. In con-trast to our procedure, Retraubun et al. (1996a) determined wormbiomass immediately after the worms were retrieved from the sedi-ment, causing excessive worm weights due to sediment retained inthe guts and consequently an almost linear relationship. In any case,the high correlation coefficients obtained in both studies (R2=0.85–0.91) suggest that both approaches can be applied to in situ studies ofA. marina as long as the gut filling status is known.
When the relationships between feces diameter and worm biomass(Fig. 2A), and temperature dependence of sediment reworking (Fig. 3B)are combined, reworking activity can at any given time be predictedfrom feces diameter with relatively high precision. This is verified bythe derived relationship between reworking activity and worm size(Fig. 4). Thus, site specific sediment reworking by Arenicola marinacan be estimated with relatively little effort, once the density and
Fig. 7. Time dependent burial of Zostera marina seedlings in experimental mesocosmswith Arenicola marina (60 m−2). The two lines indicate arbitrarily defined thresholds;(1) the gray line shows the number of seedlings with burial of >50% of above groundbiomass and (2) the black line shows the number of seedlings that were completelyburied. Right axis indicate percentages based on the total number of seedlings (n=19).
51T. Valdemarsen et al. / Journal of Experimental Marine Biology and Ecology 410 (2011) 45–52
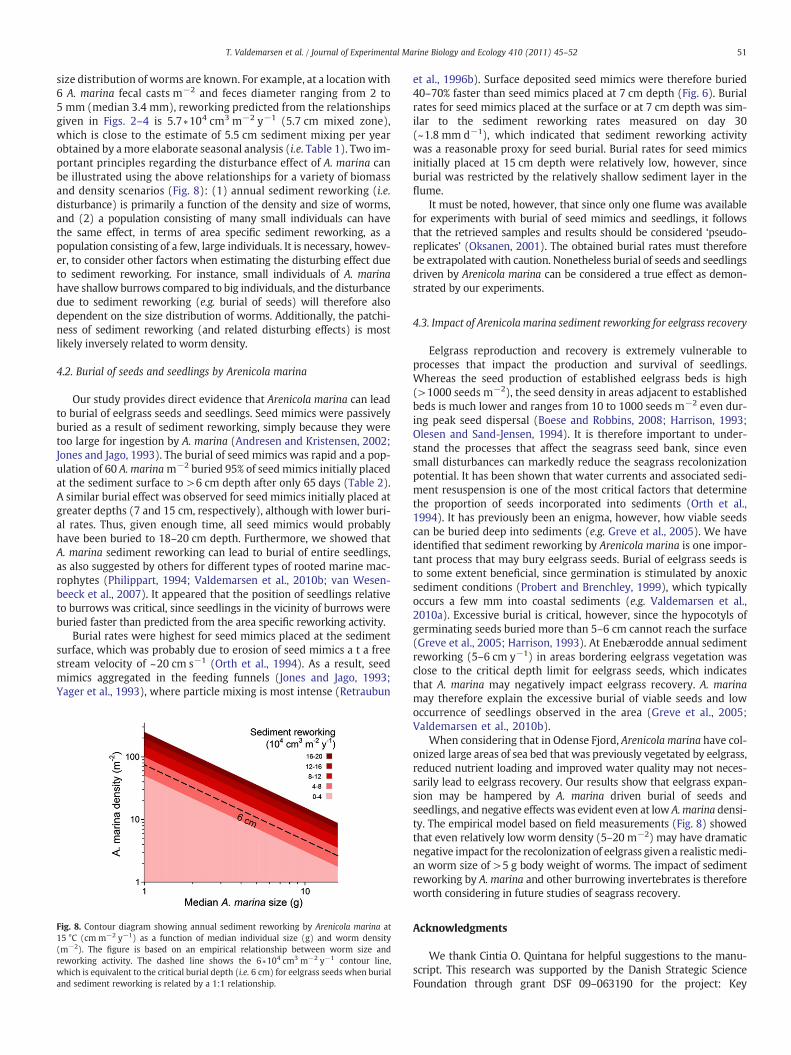
size distribution of worms are known. For example, at a location with6 A. marina fecal casts m−2 and feces diameter ranging from 2 to5 mm (median 3.4 mm), reworking predicted from the relationshipsgiven in Figs. 2–4 is 5.7 ∗104 cm3 m−2 y−1 (5.7 cm mixed zone),which is close to the estimate of 5.5 cm sediment mixing per yearobtained by amore elaborate seasonal analysis (i.e. Table 1). Two im-portant principles regarding the disturbance effect of A. marina canbe illustrated using the above relationships for a variety of biomassand density scenarios (Fig. 8): (1) annual sediment reworking (i.e.disturbance) is primarily a function of the density and size of worms,and (2) a population consisting of many small individuals can havethe same effect, in terms of area specific sediment reworking, as apopulation consisting of a few, large individuals. It is necessary, howev-er, to consider other factors when estimating the disturbing effect dueto sediment reworking. For instance, small individuals of A. marinahave shallow burrows compared to big individuals, and the disturbancedue to sediment reworking (e.g. burial of seeds) will therefore alsodependent on the size distribution of worms. Additionally, the patchi-ness of sediment reworking (and related disturbing effects) is mostlikely inversely related to worm density.
4.2. Burial of seeds and seedlings by Arenicola marina
Our study provides direct evidence that Arenicola marina can leadto burial of eelgrass seeds and seedlings. Seed mimics were passivelyburied as a result of sediment reworking, simply because they weretoo large for ingestion by A. marina (Andresen and Kristensen, 2002;Jones and Jago, 1993). The burial of seed mimics was rapid and a pop-ulation of 60 A. marinam−2 buried 95% of seed mimics initially placedat the sediment surface to >6 cm depth after only 65 days (Table 2).A similar burial effect was observed for seed mimics initially placed atgreater depths (7 and 15 cm, respectively), although with lower buri-al rates. Thus, given enough time, all seed mimics would probablyhave been buried to 18–20 cm depth. Furthermore, we showed thatA. marina sediment reworking can lead to burial of entire seedlings,as also suggested by others for different types of rooted marine mac-rophytes (Philippart, 1994; Valdemarsen et al., 2010b; van Wesen-beeck et al., 2007). It appeared that the position of seedlings relativeto burrows was critical, since seedlings in the vicinity of burrows wereburied faster than predicted from the area specific reworking activity.
Burial rates were highest for seed mimics placed at the sedimentsurface, which was probably due to erosion of seed mimics a t a freestream velocity of ~20 cm s−1 (Orth et al., 1994). As a result, seedmimics aggregated in the feeding funnels (Jones and Jago, 1993;Yager et al., 1993), where particle mixing is most intense (Retraubun
Fig. 8. Contour diagram showing annual sediment reworking by Arenicola marina at15 °C (cm m−2 y−1) as a function of median individual size (g) and worm density(m−2). The figure is based on an empirical relationship between worm size andreworking activity. The dashed line shows the 6∗104 cm3 m−2 y−1 contour line,which is equivalent to the critical burial depth (i.e. 6 cm) for eelgrass seeds when burialand sediment reworking is related by a 1:1 relationship.
et al., 1996b). Surface deposited seed mimics were therefore buried40–70% faster than seed mimics placed at 7 cm depth (Fig. 6). Burialrates for seed mimics placed at the surface or at 7 cm depth was sim-ilar to the sediment reworking rates measured on day 30(~1.8 mm d−1), which indicated that sediment reworking activitywas a reasonable proxy for seed burial. Burial rates for seed mimicsinitially placed at 15 cm depth were relatively low, however, sinceburial was restricted by the relatively shallow sediment layer in theflume.
It must be noted, however, that since only one flume was availablefor experiments with burial of seed mimics and seedlings, it followsthat the retrieved samples and results should be considered ‘pseudo-replicates’ (Oksanen, 2001). The obtained burial rates must thereforebe extrapolated with caution. Nonetheless burial of seeds and seedlingsdriven by Arenicola marina can be considered a true effect as demon-strated by our experiments.
4.3. Impact of Arenicola marina sediment reworking for eelgrass recovery
Eelgrass reproduction and recovery is extremely vulnerable toprocesses that impact the production and survival of seedlings.Whereas the seed production of established eelgrass beds is high(>1000 seeds m−2), the seed density in areas adjacent to establishedbeds is much lower and ranges from 10 to 1000 seeds m−2 even dur-ing peak seed dispersal (Boese and Robbins, 2008; Harrison, 1993;Olesen and Sand-Jensen, 1994). It is therefore important to under-stand the processes that affect the seagrass seed bank, since evensmall disturbances can markedly reduce the seagrass recolonizationpotential. It has been shown that water currents and associated sedi-ment resuspension is one of the most critical factors that determinethe proportion of seeds incorporated into sediments (Orth et al.,1994). It has previously been an enigma, however, how viable seedscan be buried deep into sediments (e.g. Greve et al., 2005). We haveidentified that sediment reworking by Arenicola marina is one impor-tant process that may bury eelgrass seeds. Burial of eelgrass seeds isto some extent beneficial, since germination is stimulated by anoxicsediment conditions (Probert and Brenchley, 1999), which typicallyoccurs a few mm into coastal sediments (e.g. Valdemarsen et al.,2010a). Excessive burial is critical, however, since the hypocotyls ofgerminating seeds buried more than 5–6 cm cannot reach the surface(Greve et al., 2005; Harrison, 1993). At Enebærodde annual sedimentreworking (5–6 cm y−1) in areas bordering eelgrass vegetation wasclose to the critical depth limit for eelgrass seeds, which indicatesthat A. marina may negatively impact eelgrass recovery. A. marinamay therefore explain the excessive burial of viable seeds and lowoccurrence of seedlings observed in the area (Greve et al., 2005;Valdemarsen et al., 2010b).
When considering that in Odense Fjord, Arenicola marina have col-onized large areas of sea bed that was previously vegetated by eelgrass,reduced nutrient loading and improved water quality may not neces-sarily lead to eelgrass recovery. Our results show that eelgrass expan-sion may be hampered by A. marina driven burial of seeds andseedlings, and negative effects was evident even at low A.marina densi-ty. The empirical model based on field measurements (Fig. 8) showedthat even relatively low worm density (5–20 m−2) may have dramaticnegative impact for the recolonization of eelgrass given a realisticmedi-an worm size of >5 g body weight of worms. The impact of sedimentreworking by A. marina and other burrowing invertebrates is thereforeworth considering in future studies of seagrass recovery.
Acknowledgments
We thank Cintia O. Quintana for helpful suggestions to the manu-script. This research was supported by the Danish Strategic ScienceFoundation through grant DSF 09–063190 for the project: Key

52 T. Valdemarsen et al. / Journal of Experimental Marine Biology and Ecology 410 (2011) 45–52
parameters and processes affecting the re-establishment of eelgrass inestuaries and coastal waters. [SS]
References
Andresen, M., Kristensen, E., 2002. The importance of bacteria and microalgae in thediet of the deposit-feeding polychaete Arenicola marina. Ophelia 56, 179–196.
Boese, B.L., Robbins, B.D., 2008. Effects of erosion and macroalgae on intertidal eelgrass(Zostera marina) in a northeastern Pacific estuary (USA). Bot. Mar. 51, 247–257.
Cadée, G.C., 1976. Sediment reworking by Arenicola marina on tidal flats in the DutchWadden Sea. Neth. J. Sea Res. 10, 440–460.
Canal-Verges, P., Vedel, M., Valdemarsen, T., Kristensen, E., Flindt, M.R., 2010. Resus-pension created by bedload transport of macroalgae: implications for ecosystemfunctioning. Hydrobiologia 649, 69–76.
Dumbauld, B.R., Wyllie-Echeverria, S., 2003. The influence of burrowing thalassinidshrimps on the distribution of intertidal seagrasses in Willapa Bay, Washington,USA. Aquat. Bot. 77, 27–42.
Flach, E.C., 1992. Disturbance of benthic infauna by sediment-reworking activities ofthe lugworm Arenicola marina. Neth. J. Sea Res. 30, 81–89.
Flach, E.C., Beukema, J.J., 1994. Density-governing mechanisms in populaiton of thelugworm Arenicola marina on tidal flats. Mar. Ecol. Prog. Ser. 115, 139–149.
Fyns Amt, 2006. Miljøfarlige stoffer og Ålegræs i Odense Fjord. Fyns Amt, Natur- ogVandmiljøafdelingen, Odense.
Greve, T.M., Krause-Jensen, D., Rasmussen, M.B., Christensen, P.B., 2005. Means of rapideelgrass (Zostera marina L.) recolonisation in former dieback areas. Aquat. Bot. 82,143–156.
Harrison, P.G., 1993. Variations in demography of Zostera marina and Z. noltii on an in-tertidal gradient. Aquat. Bot. 45, 63–77.
Hauxwell, J., Cebrian, J., Furlong, C., Valiela, I., 2001. Macroalgal canopies contribute toeelgrass (Zostera marina) decline in temperate estuarine ecosystems. Ecology 82,1007–1022.
Jarvis, J.C., Moore, K.A., 2010. The role of seedlings and seed bank viability in the recov-ery of Chesapeake Bay, USA, Zostera marina populations following a large-scale de-cline. Hydrobiologia 649, 55–68.
Jones, S.E., Jago, C.F., 1993. In situ assessment of modification of sediment properties byburrowing invertebrates. Mar. Biol. 115, 133–142.
Lee, K.S., Park, J.I., Kim, Y.K., Park, S.R., Kim, J.H., 2007. Recolonization of Zostera marinafollowing destruction caused by a red tide algal bloom: the role of new shoot re-cruitment from seed banks. Mar. Ecol. Prog. Ser. 342, 105–115.
Mascaro, O., Valdemarsen, T., Holmer, M., Perez, M., Romero, J., 2009. Experimental ma-nipulation of sediment organic content and water column aeration reduces Zosteramarina (eelgrass) growth and survival. J. Exp. Mar. Biol. Ecol. 373, 26–34.
Oksanen, L., 2001. Logic of experiments in ecology: is pseudoreplication a pseudoissue?Oikos 94, 27–38.
Olesen, B., Sand-Jensen, K., 1994. Patch dynamics of eelgrass Zostera marina. Mar. Ecol.Prog. Ser. 106, 147–156.
Orth, R.J., Luckenbach, M., Moore, K.A., 1994. Seed dispersal in a marine macrophyte —implications for colonization and restoration. Ecology 75, 1927–1939.
Orth, R.J., Harwell, C.H., Inglis, G.J., 2006. Ecology of seagrass seeds and seagrass dis-persal process. In: Larkum, A.W.D., Orth, R.J., Duarte, C.M. (Eds.), Seagrasses: Biol-ogy, Ecology and Conservation. Springer, Dordrecht, pp. 111–133.
Ostenfeld, C.H., 1908. On the growth and distribution of eelgrass (Zostera marina) inDanish waters. Rep. Dan. Biol. Stn. 16, 1–62.
Papaspyrou, S., Kristensen, E., Christensen, B., 2007. Arenicola marina (Polychaeta) andorganic matter mineralisation in sandy marine sediments: in situ and microcosmcomparison. Estuar. Coast. Shelf Sci. 72, 213–222.
Petersen, J.D., Rask, N., Madsen, H.B., Jorgensen, O.T., Petersen, S.E., Nielsen, S.V.K.,Pedersen, C.B., Jensen, M.H., 2009. Odense Pilot River Basin: implementation ofthe EU Water Framework Directive in a shallow eutrophic estuary (OdenseFjord, Denmark) and its upstream catchment. Hydrobiologia 629, 71–89.
Philippart, C.J.M., 1994. Interactions between Arenicola marina and Zostera noltii on atidal flat in the Wadden Sea. Mar. Ecol. Prog. Ser. 111, 251–257.
Plus, M., Deslous-Paoli, J.M., Dagault, F., 2003. Seagrass (Zostera marina L.) bed recolo-nisation after anoxia-induced full mortality. Aquat. Bot. 77, 121–134.
Probert, R.J., Brenchley, J.L., 1999. The effect of environmental factors on field and lab-oratory germination in a population of Zostera marina L. from southern England.Seed Sci. Res. 9, 331–339.
Quintana, C.O., Hansen, T., Delefosse, M., Banta, G., Kristensen, E., 2011. Burrow venti-lation and associated porewater irrigation by the polychaete Marenzelleria viridis.J. Exp. Mar. Biol. Ecol. 397, 179–187.
Reise, K., Simon, M., Herre, E., 2001. Density-dependent recruitment after winter dis-turbance on tidal flats by the lugworm Arenicola marina. Helgol. Mar. Res. 55,161–165.
Retraubun, A.S.W., Dawson, M., Evans, S.M., 1996a. Spatial and temporal factors affect-ing sediment turnover by the lugworm Arenicola marina (L). J. Exp. Mar. Biol. Ecol.201, 23–35.
Retraubun, A.S.W., Dawson, M., Evans, S.M., 1996b. The role of the burrow funnel infeeding processes in the lugworm Arenicola marina (L). J. Exp. Mar. Biol. Ecol.202, 107–118.
Riisgard, H.U., Banta, G.T., 1998. Irrigation and deposit feeding by the lugworm Areni-cola marina, characteristics and secondary effects on the environment. A reviewof current knowledge. Vie Milieu 48, 243–257.
Schröer, M., Wittmann, A.C., Gruner, N., Steeger, H.U., Bock, C., Paul, R., Portner, H.O.,2009. Oxygen limited thermal tolerance and performance in the lugworm Areni-cola marina: a latitudinal comparison. J. Exp. Mar. Biol. Ecol. 372, 22–30.
Sommer, A.M., Pörtner, H.O., 2002. Metabolic cold adaptation in the lugworm Arenicolamarina: comparison of a North Sea and a White Sea population. Mar. Ecol. Prog.Ser. 240, 171–182.
Valdemarsen, T., Kristensen, E., Holmer, M., 2010a. Sulfur, carbon, and nitrogen cyclingin faunated marine sediments impacted by repeated organic enrichment. Mar.Ecol. Prog. Ser. 400, 37–53.
Valdemarsen, T., Canal-Verges, P., Kristensen, E., Holmer, M., Kristiansen, M.D., Flindt,M.R., 2010b. Vulnerability of Zostera marina seedlings to physical stress. Mar.Ecol. Prog. Ser. 418, 119–130.
van Wesenbeeck, B.K., van de Koppel, J., Herman, P.M.J., Bakker, J.P., Bouma, T.J., 2007.Biomechanical warfare in ecology; negative interactions between species by habi-tat modification. Oikos 116, 742–750.
Volkenborn, N., Reise, K., 2006. Lugworm exclusion experiment: responses by depositfeeding worms to biogenic habitat transformations. J. Exp. Mar. Biol. Ecol. 330,169–179.
Volkenborn, N., Reise, K., 2007. Effects of Arenicola marina on polychaete functional di-versity revealed by large-scale experimental lugworm exclusion. J. Sea Res. 57,78–88.
Wyllie-Echeverria, S., Cox, P.A., Churchill, A.C., Brotherson, J.D., Wyllie-Echeverria, T.,2003. Seed size variation within Zostera marina L. (Zosteraceae). Bot. J. Linn. Soc.142 (3), 281–288.
Yager, P.L., Nowell, A.R.M., Jumars, P.A., 1993. Enhanced deposition to pits — a localfood source for benthos. J. Mar. Res. 51, 209–236.
Copyright © 2022 FDOKUMEN




















