
The role of bacteria and protists in nitrogen turnover in ant nest and forest floor material: A...
8
Original article The role of bacteria and protists in nitrogen turnover in ant nest and forest floor material: A laboratory experiment Veronika Jílkov a a, b, * , Jan Frouz a , Tom a s Cajthaml a, c , Michael Bonkowski b a Institute for Environmental Studies, Charles University in Prague, Ben atsk a 2, CZ-12801 Prague, Czech Republic b Department of Terrestrial Ecology, Zoological Institute, University of Cologne, Zülpicher Str. 47b, D-50674 Cologne, Germany c Institute of Microbiology, Czech Academy of Sciences, Víde nsk a 1083, CZ-14220 Prague, Czech Republic article info Article history: Received 6 January 2015 Received in revised form 14 May 2015 Accepted 18 May 2015 Available online xxx Keywords: Ammonium Microbial loop Bacterial biomass Available carbon Coniferous forest abstract Wood ant nests are hotspots of nutrients and microbial activity in nutrient-limited coniferous forest ecosystems, as ants accumulate honeydew and nutrients in their nests due to foraging and building activities. In this study we carried out a microcosm experiment focussing on the role of bacteria and protozoa on carbon and nitrogen turnover in nutrient-rich and nutrient-poor litter materials. Two types of litter material, (i) ant nest material or (ii) surrounding forest floor material were sterilized and inoculated either with bacteria alone (B treatment) or with bacteria and protozoa in combination (BP treatment). The litter materials were subsequently incubated in laboratory microcosms for 21 days. Respiration of the microcosms was measured during the whole incubation period and leachates were sampled every week and analysed for ammonium and nitrate. Our results showed lower ammonium leaching and increased respiration in the BP treatment, which could be explained by higher microbial biomass in this treatment. The negative effect of protozoa on ammonium leaching was independent of the type of litter material suggesting that available carbon in nest material supported fixation of nitrogen in the bacterial biomass. Our data show that protozoan grazing can play a critical role in retaining ni- trogen in ant nests by increasing microbial activity and biomass, and thereby preventing nitrogen leaching. © 2015 Elsevier Masson SAS. All rights reserved. 1. Introduction Soils of temperate conifer forests that grow on granitic bedrock are acidic and have a low nutrient status [1]. Fulvic acids are released during the decomposition of needle litter material of coniferous trees thus acidifying the soil water and further reducing nutrient availability in litter materials as mineral nutrients are most available at neutral pH [2]. Although conifer forest ecosystems are usually nitrogen (N) limited [3], N availability is increased in nutrient-rich patches that occur in the forest floor, such as wood ant nests. However, N cycling in these nutrient-rich patches have not yet been investigated thoroughly. Wood ant (Formica s. str.) nests are typical for temperate and boreal forest ecosystems [4,5]. They can occur in densities of 10e20 nests per ha [6e8], and the mound nests of wood ants are large and permanent structures, being occupied for decades [5,9]. They are constructed from plant materials, mainly needles, as well as from mineral soil particles [10]. Since ants transport vast amounts of food and building material into their nests [7,11e 13], a part of these nutrient-rich organic materials afterwards succumbs to decompo- sition by microorganisms and, subsequently, mineral forms of nu- trients are released [10,14,15]. Respiration rates per m 2 of nest mounds can be 12 times higher than of the surrounding forest floor [8], and mineral nitrogen accumulates in the nests which consec- utively represent hotspots of carbon and nutrient turnover and significantly increase belowground heterogeneity in the otherwise nutrient-limited forest ecosystem. These nutrients can then be exploited for example by trees growing in nests' surroundings [16]. Although many studies deal with concentrations of nutrients in wood ant nests in comparison to the surrounding forest floor (e.g. Refs. [15,17e19]) or with carbon turnover [6,8,20], there is sur- prisingly little information on the nitrogen turnover in wood ant nest material. According to the general knowledge, one group of soil fauna that could have an impact in this respect are protists. Protists are abundant in soils of coniferous forest ecosystems and * Corresponding author. Institute for Environmental Studies, Charles University in Prague, Ben atsk a 2, CZ-12801 Prague, Czech Republic. E-mail address: [email protected] (V. Jílkov a). Contents lists available at ScienceDirect European Journal of Soil Biology journal homepage: http://www.elsevier.com/locate/ejsobi http://dx.doi.org/10.1016/j.ejsobi.2015.05.004 1164-5563/© 2015 Elsevier Masson SAS. All rights reserved. European Journal of Soil Biology 69 (2015) 66e73
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of The role of bacteria and protists in nitrogen turnover in ant nest and forest floor material: A...

lable at ScienceDirect
European Journal of Soil Biology 69 (2015) 66e73
Contents lists avai
European Journal of Soil Biology
journal homepage: http: / /www.elsevier .com/locate/ejsobi
Original article
The role of bacteria and protists in nitrogen turnover in ant nest andforest floor material: A laboratory experiment
Veronika Jílkov�a a, b, *, Jan Frouz a, Tom�a�s Cajthaml a, c, Michael Bonkowski b
a Institute for Environmental Studies, Charles University in Prague, Ben�atsk�a 2, CZ-12801 Prague, Czech Republicb Department of Terrestrial Ecology, Zoological Institute, University of Cologne, Zülpicher Str. 47b, D-50674 Cologne, Germanyc Institute of Microbiology, Czech Academy of Sciences, Víde�nsk�a 1083, CZ-14220 Prague, Czech Republic
a r t i c l e i n f o
Article history:Received 6 January 2015Received in revised form14 May 2015Accepted 18 May 2015Available online xxx
Keywords:AmmoniumMicrobial loopBacterial biomassAvailable carbonConiferous forest
* Corresponding author. Institute for Environmentin Prague, Ben�atsk�a 2, CZ-12801 Prague, Czech Repub
E-mail address: [email protected] (V. Jíl
http://dx.doi.org/10.1016/j.ejsobi.2015.05.0041164-5563/© 2015 Elsevier Masson SAS. All rights res
a b s t r a c t
Wood ant nests are hotspots of nutrients and microbial activity in nutrient-limited coniferous forestecosystems, as ants accumulate honeydew and nutrients in their nests due to foraging and buildingactivities. In this study we carried out a microcosm experiment focussing on the role of bacteria andprotozoa on carbon and nitrogen turnover in nutrient-rich and nutrient-poor litter materials. Two typesof litter material, (i) ant nest material or (ii) surrounding forest floor material were sterilized andinoculated either with bacteria alone (B treatment) or with bacteria and protozoa in combination (BPtreatment). The litter materials were subsequently incubated in laboratory microcosms for 21 days.Respiration of the microcosms was measured during the whole incubation period and leachates weresampled every week and analysed for ammonium and nitrate. Our results showed lower ammoniumleaching and increased respiration in the BP treatment, which could be explained by higher microbialbiomass in this treatment. The negative effect of protozoa on ammonium leaching was independent ofthe type of litter material suggesting that available carbon in nest material supported fixation of nitrogenin the bacterial biomass. Our data show that protozoan grazing can play a critical role in retaining ni-trogen in ant nests by increasing microbial activity and biomass, and thereby preventing nitrogenleaching.
© 2015 Elsevier Masson SAS. All rights reserved.
1. Introduction
Soils of temperate conifer forests that grow on granitic bedrockare acidic and have a low nutrient status [1]. Fulvic acids arereleased during the decomposition of needle litter material ofconiferous trees thus acidifying the soil water and further reducingnutrient availability in litter materials as mineral nutrients aremostavailable at neutral pH [2]. Although conifer forest ecosystems areusually nitrogen (N) limited [3], N availability is increased innutrient-rich patches that occur in the forest floor, such as woodant nests. However, N cycling in these nutrient-rich patches havenot yet been investigated thoroughly.
Wood ant (Formica s. str.) nests are typical for temperate andboreal forest ecosystems [4,5]. They can occur in densities of 10e20nests per ha [6e8], and the mound nests of wood ants are large and
al Studies, Charles Universitylic.kov�a).
erved.
permanent structures, being occupied for decades [5,9]. They areconstructed from plant materials, mainly needles, as well as frommineral soil particles [10]. Since ants transport vast amounts offood and building material into their nests [7,11e13], a part of thesenutrient-rich organic materials afterwards succumbs to decompo-sition by microorganisms and, subsequently, mineral forms of nu-trients are released [10,14,15]. Respiration rates per m2 of nestmounds can be 12 times higher than of the surrounding forest floor[8], and mineral nitrogen accumulates in the nests which consec-utively represent hotspots of carbon and nutrient turnover andsignificantly increase belowground heterogeneity in the otherwisenutrient-limited forest ecosystem. These nutrients can then beexploited for example by trees growing in nests' surroundings [16].
Although many studies deal with concentrations of nutrients inwood ant nests in comparison to the surrounding forest floor (e.g.Refs. [15,17e19]) or with carbon turnover [6,8,20], there is sur-prisingly little information on the nitrogen turnover in wood antnest material. According to the general knowledge, one group ofsoil fauna that could have an impact in this respect are protists.Protists are abundant in soils of coniferous forest ecosystems and

V. Jílkov�a et al. / European Journal of Soil Biology 69 (2015) 66e73 67
according to food web models, play a major role in carbon andnitrogen cycling in these systems [21]. Within coniferous forestecosystems, protozoa and bacteria are particularly abundant in antnests [15,19,22], and a mechanism that could be crucial for nutrientliberation are predatoreprey interactions between protists andbacteria as protists are important microbiota grazers [21,23,24].This interaction is generally known as a microbial loop and isconsidered important for the release of mineral nitrogen fromconsumed microbial biomass, since protozoa excrete much of theingested nitrogen as ammonium [25]. Overgrazing can decreasemicrobial activity, but more often grazing effects result in enhancedmicrobial turnover [26,27]. Protists have been found to boost thedecomposing activity of bacteria, because bacterial populationsthat are grazed are metabolically more active [24,28,2]. Moreover,protozoan grazing can change composition of bacterial communitybecause gram-positive (Gþ) bacteria are generally much harder todigest by protists than gram-negative (G�) bacteria [30e32].Bearing this in mind, ant nests are useful model systems to studymicrobial mechanisms involved in carbon and nutrient dynamics oforganic hotspots in the forest floor.
In this study we focused on different roles of bacteria and pro-tists and especially on their interaction affecting nitrogen turnoverand nitrogen flow in nutrient-rich (ant nest) and nutrient-poor(forest floor) materials. We carried out an experiment using mi-crocosms with sterilized ant nest or forest floor material, eitherinoculated with litter bacteria alone or with bacteria and protiststogether. We subsequently sampled leaching water from the mi-crocosms and analysed the leachates for mineral forms of nitrogenat regular intervals. Also, respiration of the microcosms wasmeasured during the whole incubation period. Our hypotheseswere that 1) higher microbial activity (i.e. respiration) will occur inthe ant nest material, especially when inoculated with bacteria andprotists together, 2) higher amounts of mineral nitrogen will beleached from the ant nest material, 3) even more nitrogen will beleached in presence of protists, and 4) bacterial communitycomposition will be shifted towards Gþ bacteria in presence ofprotists.
2. Materials and methods
2.1. Sampling and preparation of the materials
Litter materials were sampled in a forest stand dominated byNorway spruce (Picea abies (L.) H. Karst.) with rare beech (Fagus sp.)and oak (Quercus sp.) situated at 600m abovem.s.l. on the southernslope of the Klet Mountain in South Bohemia (Czech Republic) inAugust 2012. Approximately 200 cm3 of ant nest material wassampled from the centre of five randomly selected ant (Formicaaquilonia Yarrow) nests of a similar size. The centre of the nests wassampled as there is the highest microbial activity [33,34]. Forcomparison, the same volume of litter material was sampled fromthe forest floor at a depth of 0e5 cm where microbial activity ishighest [35]. The forest floor material (L horizon), mainly repre-sented by conifer needles, was sampled in the nests' surroundings,approximately five meters from each nest. Materials were thentransferred to the laboratory and thoroughly mixed. Ants wereseparated from the ant nest material using pincers. Fresh ant nestand forest floor material was then put in thin layers into plasticbags and g-ray sterilized (isotope Co60, total dose 40 kGy) [36].Afterwards, the materials were kept at 4 �C until the incubationstarted. Total carbon and nitrogen content was determined in driedand crushed materials on a CN analyser. Material pH was measuredin a 1:10 material:distilled water solution using a glass electrode.Ammonia and nitrate were extracted from the materials byKAl(SO4)2 solution and determined using the Kjeldahl method [37].
Properties of the sterile materials are summarized in Table 1.
2.2. Preparation of the inocula
To prepare bacterial inocula, ca. 30 g of fresh ant nest or forestfloor material were mixed with 200 ml of sterile water and incu-bated for two hours. Bacteria were isolated from the litter materialsby a sequence of filtering steps according to Rosenberg et al. [32].First, the suspension was filtered through a paper filter to separatethe organic materials. Two subsequent filtering steps with 3 and1.2 mm filters (Cellulose Acetat Filters (Minisart®), Sartorius) understerile conditions resulted in a filtrate with a mixed diverse bac-terial community. The filtrates were incubated for three days atroom temperature in Neffs Modified Amoebae Saline (NMAS) me-dium [38] and then checked for contaminations by protists underthe microscope. The protozoa-free bacterial inocula from ant nestand litter material were pooled and thoroughly mixed together forsubsequent litter inoculation.
To prepare protozoan inocula, app. 5 g of fresh ant nest or forestfloor material were mixed with 20 ml of NMAS medium supple-mented with 1:9 v:v sterile nutrient broth (NB-NMAS, Oxoid) in aPetri dish [38]. The suspension was incubated for seven days atroom temperature. Then, protists (amoebae, flagellates, ciliates)were identified at 100� magnification using an inverted micro-scope (Nikon TE2000) only to be sure that all the groups werepresent in the inoculum. Tenml of inoculumwere transferred usinga Pasteur pipette into a new Petri dish with 20 ml of NB-NMASmedium and further incubated for two days at room temperature.The inocula from ant nest and litter material were pooled andthoroughly mixed together for subsequent litter inoculation.
During the incubation period microbial and protozoan com-munities adapted to the available resources in the respective littertreatments could establish.
2.3. Microcosm experiment
The design of the microcosms corresponded to those used byBonkowski et al. [29]. Microcosm chambers consisted of perspextubes (150 mm height, 60 mm diam) placed on ceramic lysimeterplates. The chambers could be drained under natural conditions ofsoil matric potential by lowering the atmospheric pressure in a boxbelow the plates. Removable, air-tight lids on top of the chambersallowed watering of microcosms. Small vials attached to the lidscontained 5 ml 1 N KOH to trap CO2 for respiration measurements.All microcosm chambers had been sterilized by soaking in 70%ethanol.
Into each of the microcosms, 3 g of either ant nest or forest floormaterial was put and the materials were consecutively wateredwith 3 ml of sterile distilled water. All microcosms were inoculatedwith 1 ml of bacterial inoculum. Moreover, one half of the micro-cosms was inoculated with 0.5 ml of the protozoan inoculum (BPtreatment). To the other bacteria-treated half, 0.5 ml of NB-NMASmedium was added to get the same moisture and nutrient condi-tions (B treatment). We had 12 replicates for each treatment plus 12blanks (empty microcosms) for respiration measurements, in total60 microcosms. The incubation started at the beginning ofSeptember 2012 and continued for 21 days at room temperatureunder sterile conditions in a sterile bench.
Measurements of microcosm respiration started on the thirdday of incubation to avoid the first burst of respiration after inoc-ulation and then were done regularly, daily at the beginning and atan interval of two days after six days of incubation. Respiration wasdetermined by titration of 1 N KOH with 0.01 M hydrochloric acid,after precipitation by barium chloride according to Schinner et al.[37]. Microcosms were leached weekly with 20 ml of sterile

Table 1Properties of the sterile materials before the incubation. Means ± SD are shown.
Material NH4þ (mg g�1 DW) NO3
� (mg g�1 DW) C (%) N (%) pHH2O
Ant nest 2827 ± 138 42 ± 40 44 ± 2.2 1.21 ± 0.07 4.44 ± 0.03Forest floor 1623 ± 80 58 ± 48 47 ± 0.3 1.15 ± 0.02 4.33 ± 0.01
V. Jílkov�a et al. / European Journal of Soil Biology 69 (2015) 66e7368
distilled water that was subsequently drained through lysimeterplates at 40 kPa into beakers underneath to monitor nutrientrelease by microbial mineralization and protozoan feeding (seeBonkowski et al. [29]). The leachates were analysed for mineralforms of nitrogen, i.e. ammonia and nitrate, using the Kjeldahlmethod [37].
At the end of the incubation, the materials from the microcosmswere sampled and divided for analyses of basal respiration, mi-crobial biomass, and the composition of phospholipid fatty acids.
Fig. 1. Cumulative respiration of the materials during the incubation. B means inoc-ulated with bacteria, BP means inoculated with bacteria and protists.
2.4. Microbial analyses
Basal respiration and microbial biomass (substrate inducedrespiration, SIR) were determined using an automated respirom-eter based on electrolytic O2 microcompensation described byScheu [39]. For basal respiration, a sample corresponding to 0.5 gdry weight of litter was incubated, and oxygen consumption rateswere measured at 22 �C every hour. Basal respiration was calcu-lated from the average O2 consumption rate (ml g�1 h�1) in samplesduring hours 15e20 when oxygen consumption had reached asteady-state. The same samples were subsequently used for SIRmeasurements after addition of 80mg glucose g�1 dry weight as anaqueous solution. The mean of the three lowest respiration mea-surements during the first 10 h after glucose addition was taken asmaximum initial respiratory response (MIRR). Microbial biomass C(Cmic, mg g�1) was calculated as 38 �MIRR [40,41]. The respiratoryquotient (qO2, ml O2 mg Cmic�1 h�1) was determined as ameasure ofmicrobial carbon turnover by dividing microbial respiration bymicrobial biomass.
Litter material for phospholipid fatty acid (PLFA) analyses wasextracted with a mixture of chloroformemethanolephosphatebuffer (1:2:0.8) [42]. Phospholipids were separated using solid-phase extraction cartridges (LiChrolut Si 60, Merck), and the sam-ples were subjected to mild alkaline methanolysis. The free methylesters of phospholipid fatty acids were analysed by gas chroma-tographyemass spectrometry (450-GC, 240-MS ion trap detector,Varian, Walnut Creek, CA, USA). The GC instrument was equippedwith a split/splitless injector, and a DB-5MS column was used forseparation (60 m, 0.25 mm i.d., 0.25 mm film thickness). Thetemperature program started at 60 �C and was held for 1 min insplitless mode. Then the splitter was opened, and the oven washeated to 160 �C at a rate of 25 �C min�1. The second temperatureramp was up to 280 �C at a rate of 2.5 �C min�1, and this finaltemperature was maintained for 10 min. The solvent delay timewas 8 min. The transfer line temperature was 280 �C. Mass spectrawere recorded at 1 scan s�1 under electron impact at 70 eV, massrange 50e350 amu. Methylated fatty acids were identified ac-cording to their mass spectra and by comparison with a mixture ofchemical standards obtained from Sigma. Relative bacterialbiomass was quantified as a sum of 10Me-16:0, 10Me-17:0, 10Me-18:0 (actinobacteria), i14:0, i15:0, a15:0, i16:0, i17:0, a17:0 (gram-positive (Gþ) bacteria), 16:1u7t, 18:1u7, cy17:0, cy19:0, 16:1u5(gram-negative (G�) bacteria), and relative fungal biomass wasquantified based on 18:2u6,9 content. During stationary growth ofG� bacteria, the fatty acid cy17:0 is converted from its precursor16:1u7 [43,44], consequently, the ratio of cy17:0 to 16:1u7 can beused as an indicator of the growth phase of G� bacteria with lowervalues indicating log phase growth and higher values indicating
stationary phase growth [45,46].
2.5. Statistical analyses
Datawere analysed by two-factorial ANOVAwith Litter (ant nestmaterial vs. forest floormaterial) and Protozoa (without vs. with) asfactors. If required, data were log-transformed to maintain homo-geneity of variance. Microbial contributions of PLFA data werecalculated as relative proportions of total PLFAs in a sample, andarcsine square root transformed to obtain homogeneity of variance[47]. In the litter material, although sterilized by gamma-radiation,a slight contamination by fungi occurred in some treatments.Therefore, values of 18:2u6,9 > 1 ppm were used as covariate inANOVA to correct for the influence of fungi. Means of the individualtreatments were compared using Tukey's honestly significant dif-ference (HSD) at the 5% probability level. Linear regressions for thedependence of mobilized nitrogen on microbial parameters werecalculated using the Linear Model procedure in the statisticalpackage R. The changes in soil respiration and nutrient leachingthrough time were analysed by a multivariate analysis of variance(MANOVA), with the explanatory variables litter type and presence/absence of protozoa; proceeding to the ANOVA (analysis of vari-ance) only if theMANOVAyielded a significant result (i.e. ‘protectedANOVA’ [48]). Roy's Greatest Root was chosen as test of significantdifferences among groups in the MANOVA procedure. All statisticalanalyses were calculated in R version 3.0.2.
3. Results
3.1. Respiration measurements
Microbial respiration was highest during the first six days of theincubation in all treatments, and then decreased (Fig. 1). Microbialrespiration was affected by an interaction of litter material andpresence of protozoa (MANOVA Litter� Prot: Roy's GreatestRoot9,33 ¼ 0.88, P ¼ 0.007), and this was due to the fact that by farthe highest respiration occurred in the ant nest material in the BP

V. Jílkov�a et al. / European Journal of Soil Biology 69 (2015) 66e73 69
treatment (Litter� Prot: F1,44 ¼ 17.14, P < 0.001). However, after sixdays of incubation, respiration in this treatment also decreased, andrespiration had converged in all treatments to similar rates after 19days of incubation. At the end of the incubation, actual respirationrates did not differ between treatments (F1,44 ¼ 0.058, P ¼ 0.81).Total respiration was increased on average by a factor of 1.48 inpresence of protozoa (Prot: F1,41¼30.43, P < 0.001), and on average,it was also higher in ant nest material (�1.17, Litter: F1,41 ¼ 10.80,P ¼ 0.002), but total respiration of the ant nest material in presenceof protozoa appeared to be increased by 41, 39, and 22% comparedto the Forest floor B, Ant nest B, and Forest floor BP treatments,respectively (Litter� Prot: F1,41 ¼ 3.91, P ¼ 0.0549).
3.2. Nitrogen leaching
Nitrogen leaching was strongly dependent on litter materials,but the effects were exclusively caused by high amounts ofammonia, while no differences were found for nitrate (Litter:F1,39 ¼ 1.56, P ¼ 0.22; Protozoa: F1,39 ¼ 0.03, P ¼ 0.87), as almostthree times more ammonia than nitrate was liberated from thematerials during thewhole incubation (Table 2). Ammonia leachingdecreased strongly after the first week of incubation, but the totalamounts of nitrogen leaching depended on litter materials (MAN-OVA, Roy's Greatest Root3,36 ¼ 1.32, P < 0.001). After 7 days of in-cubation, when the major part of nitrogen was leached, about 1.25times more ammonium was released from the ant nest litter incomparison to the forest floor litter (Litter: F1,43 ¼ 14.61, P < 0.001).However, in the second leachates sampled after 14 days of incu-bation, ammonium concentrations in leachates of ant nest materialhad decreased by more than half and were now even 35% lowerthan in leachates from the forest floor material (Litter: F1,44 ¼ 31.36,P < 0.001). Surprisingly, 20% less ammonia was leached in presenceof protozoa (Prot: F1,44 ¼ 8.39, P < 0.01). In the third leachatessampled after 21 days of the incubation, when concentrations ofavailable nutrients likely had strongly decreased, no significantdifferences were found between the litter materials (Litter:F1,41 ¼ 2.75, P ¼ 0.11), nor of protozoa treatments (Prot: F1,41 ¼ 0.91,P ¼ 0.35). Over the whole incubation period, total ammonialeaching was not affected by the types of litter material (Litter:F1,38 ¼ 1.77, P ¼ 0.19), but it was still significantly reduced (�7%) inpresence of protozoa (Prot: F1,38 ¼ 5.40, P < 0.05). It is noteworthythat only in the ant nest material with protozoa, total leaching ofammonium was negatively correlated with total microbial respi-ration (F1,7 ¼ 6.91, P ¼ 0.034, R2 ¼ 0.50), showing that less nitrogenwas leached when microbial respiration increased. In the sametreatment, total ammonia leaching also was slightly negativelycorrelated with SIR microbial biomass at the end of the incubation(F1,35¼ 6.5, P < 0.05, R2 ¼ 0.15), indicating that increasing microbialbiomass may lead to lower rates of ammonia leaching.
3.3. Microbial analyses
At the end of the incubation, basal respiration was on average
Table 2Mineral nitrogen amounts (mg g�1 DW) leached from the materials during incubation.Different letters show significant differences (P < 0.05) between the mean values of respewere not significant.
Material Length of incubation 1 week 2 w
Treatment NH4þ NO3
� NH
Forest floor B 127 ± 14 ab 35 ± 8 10BP 110 ± 27 b 36 ± 12 95
Ant nest B 153 ± 21 a 43 ± 10 78BP 143 ± 39 a 43 ± 11 53
33% higher in the forest floor material than in the ant nest material(Litter: F1,40 ¼ 1713, P < 0.01), protozoa did not influence basalrespiration (Prot: F1,40 ¼ 2.34, P ¼ 0.13, Fig. 2a). Microbial biomass(SIR) did not differ between the litter materials (Litter: F1,36 ¼ 0.24,P ¼ 0.63), but it appeared to be 1.36-fold enhanced in presence ofprotozoa (Prot: F1,36 ¼ 3.57, P ¼ 0.067). Particularly in ant nestmaterial of the BP treatment, microbial biomass was almost twofold (�1.77) higher compared to the B treatment, although this wasnot significant (Litter� Prot: F1,36 ¼ 3.19, P ¼ 0.08) (Fig. 2b). At theend of the incubation, the respiratory quotient was 42% higher inforest floor material compared to ant nest material (Litter:F1,33 ¼ 18.37, P < 0.001) and it was particularly low in the protozoantreatment of ant nest material compared to the protozoan treat-ment of forest floor material (Fig. 2c).
A relatively greater proportion of bacterial PLFAs was found inthe ant nest material (85%) in comparison to the forest floor ma-terial (76%, Litter: F1,44 ¼ 22.62, P < 0.001) but the effect mainlyresulted from high bacterial biomass in ant nest material in pres-ence of protozoa (Fig. 3a). The significant increase of bacterial PLFAsin the BP (83%) compared to the B treatment (78%) confirmed ourfindings from microbial biomass SIR measurements (Prot:F1,44 ¼ 7.81, P < 0.01). Actinobacteria made up only a minor fractionof PLFAs, but they were significantly more abundant in ant nestmaterial in absence of protozoa (17%) compared to all other treat-ments (~14%) (Litter� Prot: F1,44 ¼ 4.46, P < 0.05) (Fig. 3b). MoreGþ bacteria occurred in the forest floor material (48%) in compar-ison to the ant nest material (43%) (Litter: F1,44¼ 6.39, P < 0.05), andprotozoa also generally favoured Gþ bacteria in the forest floormaterial (50%), while decreasing the abundance of Gþ bacteria inant nest material (42%) (Litter� Prot: F1,44 ¼ 4.79, P < 0.05, Fig. 3c).G� bacteria PLFAs were relatively more abundant in the ant nestmaterial (43%) compared to the forest floor material (34%)(F1,42 ¼ 56.37, P < 0.001, Fig. 3d). Although the abundance of G�bacteria PLFAs was not affected by protozoa at the end of the in-cubation period (F1,42 ¼ 0.14, P ¼ 0.71), the ratio of cy17:0/16:1u7might indicate that growth of G� bacteria in the ant nest materialin the B treatment was in a stationary phase, compared to a logphase growth in all other treatments (Litter� Prot: F1,39 ¼ 21.04,P < 0.001) (Fig. 4). Contaminations by fungi occurred in both, forestfloor material (24%) and in ant nest material (15%). Although theeffect of fungal contaminations on total respirationwas statisticallysignificant (F1,41 ¼ 5.42, P < 0.05), it explained only a minor part ofthe overall variation (10%). Fungal contaminations tended to have aminor effect on the total amount of bacterial PLFAs (F1,43 ¼ 3.74,P ¼ 0.06), which explained 12% of the overall variation, which wasbased on an overall negative correlation between fungal and bac-terial PLFAs in the ant nest material (F1,22 ¼ 6.11, p ¼ 0.022), but notin the forest floor material (F1,22 ¼ 1.31, p ¼ 0.27).
4. Discussion
Wood ants transport not only nutrients, but also vast amounts ofeasily-available carbon in form of honeydew into their mounds. For
B means inoculated with bacteria, BP means inoculated with bacteria and protists.ctive treatments in each week. Means ± SD are shown. Differences in NO3
� leaching
eeks 3 weeks Total
4þ NO3
� NH4þ NO3
� NH4þ NO3
�
7 ± 33 a 39 ± 14 64 ± 10 29 ± 6 297 a 103± 24 ab 37 ± 8 62 ± 12 28 ± 6 267 b 100± 8 b 30 ± 7 59 ± 14 25 ± 7 290 a 97± 14 c 24 ± 4 55 ± 11 26 ± 7 250 b 92
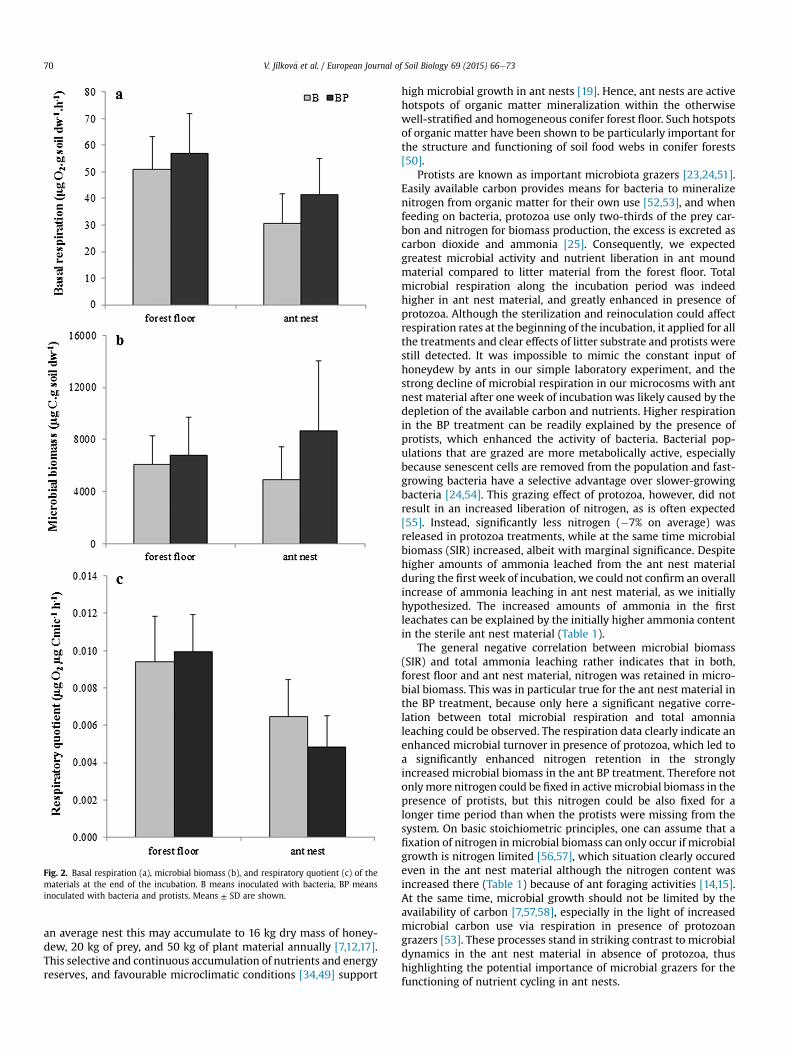
Fig. 2. Basal respiration (a), microbial biomass (b), and respiratory quotient (c) of thematerials at the end of the incubation. B means inoculated with bacteria, BP meansinoculated with bacteria and protists. Means ± SD are shown.
V. Jílkov�a et al. / European Journal of Soil Biology 69 (2015) 66e7370
an average nest this may accumulate to 16 kg dry mass of honey-dew, 20 kg of prey, and 50 kg of plant material annually [7,12,17].This selective and continuous accumulation of nutrients and energyreserves, and favourable microclimatic conditions [34,49] support
high microbial growth in ant nests [19]. Hence, ant nests are activehotspots of organic matter mineralization within the otherwisewell-stratified and homogeneous conifer forest floor. Such hotspotsof organic matter have been shown to be particularly important forthe structure and functioning of soil food webs in conifer forests[50].
Protists are known as important microbiota grazers [23,24,51].Easily available carbon provides means for bacteria to mineralizenitrogen from organic matter for their own use [52,53], and whenfeeding on bacteria, protozoa use only two-thirds of the prey car-bon and nitrogen for biomass production, the excess is excreted ascarbon dioxide and ammonia [25]. Consequently, we expectedgreatest microbial activity and nutrient liberation in ant moundmaterial compared to litter material from the forest floor. Totalmicrobial respiration along the incubation period was indeedhigher in ant nest material, and greatly enhanced in presence ofprotozoa. Although the sterilization and reinoculation could affectrespiration rates at the beginning of the incubation, it applied for allthe treatments and clear effects of litter substrate and protists werestill detected. It was impossible to mimic the constant input ofhoneydew by ants in our simple laboratory experiment, and thestrong decline of microbial respiration in our microcosms with antnest material after one week of incubation was likely caused by thedepletion of the available carbon and nutrients. Higher respirationin the BP treatment can be readily explained by the presence ofprotists, which enhanced the activity of bacteria. Bacterial pop-ulations that are grazed are more metabolically active, especiallybecause senescent cells are removed from the population and fast-growing bacteria have a selective advantage over slower-growingbacteria [24,54]. This grazing effect of protozoa, however, did notresult in an increased liberation of nitrogen, as is often expected[55]. Instead, significantly less nitrogen (�7% on average) wasreleased in protozoa treatments, while at the same time microbialbiomass (SIR) increased, albeit with marginal significance. Despitehigher amounts of ammonia leached from the ant nest materialduring the first week of incubation, we could not confirm an overallincrease of ammonia leaching in ant nest material, as we initiallyhypothesized. The increased amounts of ammonia in the firstleachates can be explained by the initially higher ammonia contentin the sterile ant nest material (Table 1).
The general negative correlation between microbial biomass(SIR) and total ammonia leaching rather indicates that in both,forest floor and ant nest material, nitrogen was retained in micro-bial biomass. This was in particular true for the ant nest material inthe BP treatment, because only here a significant negative corre-lation between total microbial respiration and total amonnialeaching could be observed. The respiration data clearly indicate anenhanced microbial turnover in presence of protozoa, which led toa significantly enhanced nitrogen retention in the stronglyincreased microbial biomass in the ant BP treatment. Therefore notonlymore nitrogen could be fixed in activemicrobial biomass in thepresence of protists, but this nitrogen could be also fixed for alonger time period than when the protists were missing from thesystem. On basic stoichiometric principles, one can assume that afixation of nitrogen in microbial biomass can only occur if microbialgrowth is nitrogen limited [56,57], which situation clearly occuredeven in the ant nest material although the nitrogen content wasincreased there (Table 1) because of ant foraging activities [14,15].At the same time, microbial growth should not be limited by theavailability of carbon [7,57,58], especially in the light of increasedmicrobial carbon use via respiration in presence of protozoangrazers [53]. These processes stand in striking contrast to microbialdynamics in the ant nest material in absence of protozoa, thushighlighting the potential importance of microbial grazers for thefunctioning of nutrient cycling in ant nests.

Fig. 3. PLFA composition of the materials at the end of the incubation. Biomass of the bacteria (a), actinobacteria (b), Gþ bacteria (c), and G� bacteria (d) is shown. B meansinoculated with bacteria, BP means inoculated with bacteria and protists. Different letters indicate significant differences between means of respective treatments in each group ofmicrobial community (P < 0.05, Tukey's HSD test). Means ± SD are shown.
Fig. 4. Ratio of cy17:0 to 16:1u7 of G� bacteria. B means inoculated with bacteria, BPmeans inoculated with bacteria and protists. Different letters indicate significant dif-ferences between means at P < 0.05 (Tukey's HSD test). Means ± SD are shown.
V. Jílkov�a et al. / European Journal of Soil Biology 69 (2015) 66e73 71
The PLFA data were helpful to differentiate microbial groupsbetween ant nest and litter material, and in particular to shed lighton some of the changes in microbial community composition dueto protozoan grazing. It is well known that some bacterial taxaevolved grazing tolerances against protozoa [59], that bacteria
respond to grazing with physiological changes in functioning[53,60,61], and that protozoan grazing is selective, thus funda-mentally changing the community composition of bacteria [54]. Ageneral pattern in forest floor material, although not significant inant nest material, appeared to be an increase of Gþ bacteria inpresence of protozoan grazers in soil, which is in accordance withresults of Griffiths et al. [30], Rønn et al. [31], and Rosenberg et al.[32].
The relatively high biomass of Actinobacteria in the ant nestmaterial does not correspond to the field situation in the forestecosystem [19]. At a closer look, this result was mainly caused bythe high abundance of Actinobacteria in the treatment with antnest material in absence of protozoa (Fig. 3b). Although the ratio ofcy17:0/16:1u7 might indicate a shift in bacterial communitycomposition, this might also indicate a shift in the physiologicalstate of gram-negative bacteria [62]. Therefore we assume that theratio of cy17:0/16:1u7 rather indicated that growth of G� bacteriain the ant nest material in the B treatment was in a stationaryphase, whereas G� bacterial populations in the other treatmentsshowed a log phase growth [43e45], suggesting growth-limitingconditions at the end of the incubation period in the B treatment.Higher nutrient losses and low respiration in this treatment alsoindicate that the microbial community was not able to use theavailable nutrients in absence of protozoa, while the same initialcommunity showed completely different dynamics in presence ofprotozoa. Unfortunatelly, no specific bacterial community data forthe ant nest material have been published so far and our experi-mental set up did not allow to reveal the specific mechanisms any

V. Jílkov�a et al. / European Journal of Soil Biology 69 (2015) 66e7372
further. Most bacteria produce antibiotics, and strong antagonisticinteractions with mutual inhibitions, i.e. ‘bacteriostasis’, are com-mon among bacterial communities in nature as well as in experi-mental systems [63e67]. We can only speculate that similarprocesses hampered microbial growth in ant nest material and thatthese interactions fundamentally changed with protozoan grazing.More research into the specific mechanisms of microbial in-teractions and its modifications by protozoa in ant nest material isdefinitely needed, notably since these interactions at the base ofsoil food webs can be expected to propagate to higher trophiclevels.
5. Conclusion
The research described here presents one of the first experi-ments dealing with nitrogen turnover in the ant nest material.Although this experiment was quite artificial in nature and short-term, our results provide a first step to uncover the potential mi-crobial key players, and some fundamental mechanisms influ-encing carbon and nitrogen cycling in nutrient-poor conifer forestlitter, and nutrient-rich ant nest material. Protists appeared crucialfor retaining ammonia in the ant nest material. They did so byincreasing microbial activity and biomass, which locked up nitro-gen in microbial biomass and prevented it from leaching, at least ina short-term perspective. Although this mechanism might benegligible at an ecosystem level due to the relatively low number ofant nests per ha, it is likely important for organisms living in or inthe vicinity of ant nests. Undoubtedly, further research is requiredto assess the absolute rate of nitrogenmineralization in the ant nestmaterial and the specific interactions between the microbialplayers involved, but our data clearly show that protists are a sig-nificant component to control the amount of nutrient availability inthe nutrient-rich organic hotspots that ant nests create in other-wise nutrient-limited litter layers of conifer forests.
Acknowledgement
This study was supported by a scholarship from the Universityof Cologne in terms of the partnership between this university andCharles University in Prague. We would like to thank Anna Herzog,Cornelia Thielen, and Irene Brockhaus for technical help in the lab.We are grateful to Stefan Geisen who helped with inocula prepa-ration, Alena Filipova for her help with PLFA analyses, and to Dr.Robert Koller who made usefull suggestions for the experimentalset-up.
References
[1] N.C. Brady, R.R. Weil, The Nature and Properties of Soils, Prentice-Hall, UpperSaddle River, New Jersey, 2002.
[2] W.D. Bowman, H. Steltzer, T.N. Rosenstiel, C.C. Cleveland, C.L. Meier, Littereffects of two co-occurring alpine species on plant growth, microbial activityand immobilization of nitrogen, Oikos 104 (2004) 336e344.
[3] M.E. McGroddy, T. Daufresne, L.O. Hedin, Scaling of C: N:P stoichiometry inforests worldwide: implications of terrestrial redfield-type ratios, Ecology 85(2004) 2390e2401.
[4] G.M. Dlusskij, Muravji Roda Formica, Nauka, Moskva, 1967.[5] B. Seifert, Ameisen: Beobachten, Bestimmen, Naturbuch Verlag, Ausburg,
1996.[6] T. Domisch, L. Finer, M. Ohashi, A.C. Risch, L. Sundstr€om, P. Niemel€a,
M.F. Jurgensen, Contribution of red wood ant mounds to forest floor CO2 effluxin boreal coniferous forests, Soil Biol. Biochem. 38 (2006) 2425e2433.
[7] V. Jílkov�a, M. �Sebek, J. Frouz, Mechanisms of pH change in wood ant (Formicapolyctena) nests, Pedobiologia 55 (2012) 247e251.
[8] A.C. Risch, M.F. Jürgensen, D.S. Page-Dumroese, M. Schütz, The contribution ofred wood ants to soil C and N pools and CO2 emissions in subalpine forests,Ecology 86 (2005) 419e430.
[9] B. H€olldobler, E.O. Wilson, The Ants, Springer Verlag, Berlin, 1990.[10] S.M. Kristiansen, W. Amelung, Abandoned anthills of Formica polyctena and
soil heterogeneity in a temperate deciduous forest: morphology and organic
matter composition, Eur. J. Soil Sci. 52 (2001) 355e363.[11] K. Horstmann, Untersuchungen über den Nahrungswerb der Waldameisen
(Formica polyctena Foerster) im Eichenwald. III. Jahresbilanz, Oecologia 15(1974) 187e204.
[12] A.D. Pokarzhevskij, The distribution and accumulation of nutrients in nests ofant Formica polyctena (Hymenoptera, Formicidae), Pedobiologia 21 (1981)117e124.
[13] D.J. Stradling, Food and feeding habits of ants, in: M.V. Brian (Ed.), ProductionEcology of Ants and Termites, Cambridge University Press, Cambridge, 1978,pp. 81e106.
[14] T. Domisch, M. Ohashi, L. Finer, A.C. Risch, L. Sundstr€om, J. Kilpel€ainen,P. Niemel€a, Decomposition of organic matter and nutrient mineralisation inwood ant (Formica rufa group) mounds in boreal coniferous forests ofdifferent age, Biol. Fert. Soils 44 (2008) 539e545.
[15] L. Lenoir, T. Persson, J. Bengtsson, Wood ant nests as potential hot spots forcarbon and nitrogen mineralisation, Biol. Fert. Soils 34 (2001) 235e240.
[16] J. Frouz, M. Rybní�cek, P. Cudlín, E. Chmelíkov�a, Influence of the wood ant,Formica polyctena, on soil nutrient and the spruce tree growth, J. Appl.Entomol. 132 (2008) 281e284.
[17] J. Frouz, H. �Santr�u�ckov�a, J. Kal�cík, The effect of wood ants (Formica polyctenaFoerst.) on the transformation of phosphorus in a spruce plantation, Pedo-biologia 41 (1997) 437e447.
[18] J. Frouz, J. Kal�cík, P. Cudlín, Accumulation of phosphorus in nests of red woodants Formica s. str, Ann. Zool. Fenn. 42 (2005) 269e275.
[19] J. Laakso, H. Set€al€a, Nest mounds of red wood ants (Formica aquilonia): hotspots for litter-dwelling earthworms, Oecologia 111 (1997) 565e569.
[20] M. Ohashi, L. Finer, T. Domisch, A.C. Risch, M.F. Jürgensen, CO2 efflux from redwood ant mound in a boreal forest, Agr. For. Meteorol. 130 (2005) 131e136.
[21] D. Schr€oter, V. Wolters, P.C. De Ruiter, C and N mineralisation in thedecomposer food webs of a European forest transect, Oikos 102 (2003)294e308.
[22] G.A. Korganova, Ecological aspects of development of testacean complexes innests of Formica lugubris and Formica exsecta (Hymenoptera, Formicidae),Entomol. Rev. 88 (2008) 1075e1087.
[23] M. Bonkowski, W. Cheng, B.S. Griffiths, J. Alphei, S. Scheu, Microbial-faunalinteractions in the rhizosphere and effects on plant growth, Eur. J. Soil Biol. 36(2000) 135e147.
[24] R. Rønn, M. Vestergard, F. Ekelund, Interactions between bacteria, protozoaand nematodes in soil, Acta Protozool. 51 (2012) 223e235.
[25] B.S. Griffiths, Soil nutrient flow, in: J.F. Darbyshire (Ed.), Soil Protozoa, CABInternational, Wallingford, 1994, pp. 65e92.
[26] E.R. Ingham, J.A. Trofymow, R.N. Ames, H.W. Hunt, C.R. Morley, J.C. Moore,D.C. Coleman, Trophic interactions and nitrogen cycling in a semiarid grass-land soil. 1. Seasonal dynamics of the natural populations, their interactionsand effects on nitrogen cycling, J. Appl. Ecol. 23 (1986) 597e614.
[27] H. Petersen, M. Luxton, A comparative analysis of soil fauna populations andtheir role in decomposition processes, Oikos 39 (1982) 287e388.
[28] J. Alphei, M. Bonkowski, S. Scheu, Protozoa, Nematoda and Lumbricidae in therhizosphere of Hordylemus europaeus (Poaceae): faunal interactions, responseof microorganisms and effects on plant growth, Oecologia 106 (1996)111e126.
[29] M. Bonkowski, B. Griffiths, C. Scrimgeour, Substrate heterogeneity andmicrofauna in soil organic ‘hotspots’ as determinants of nitrogen capture andgrowth of ryegrass, Appl. Soil Ecol. 14 (2000) 37e53.
[30] B.S. Griffiths, M. Bonkowski, G. Dobson, S. Caul, Changes in soil microbialcommunity structure in the presence of microbial-feeding nematodes andprotozoa, Pedobiologia 43 (1999) 297e304.
[31] R. Rønn, A.E. McCaig, B.S. Griffiths, J.I. Prosser, Impact of protozoan grazing onbacterial community structure in soil microcosms, Appl. Environ. Microbiol.68 (2002) 6094e6105.
[32] K. Rosenberg, J. Bertaux, K. Krome, A. Hartmann, S. Scheu, M. Bonkowski, Soilamoebae rapidly change bacterial community composition in the rhizosphereof Arabidopsis thaliana, ISME J. 3 (2009) 675e684.
[33] D. Coenen-Stab, B. Schaarschmidt, I. Lamprecht, Temperature distribution andcalorimetric determination of heat production in the nest of the wood ant,Formica polyctena (Hymenoptera, Formicidae), Ecology 61 (1980) 238e244.
[34] J. Frouz, M. Holec, J. Kal�cík, The effect of Lasius niger (Hymenoptera, For-micidae) ant nest on selected soil chemical properties, Pedobiologia 47 (2003)205e212.
[35] E.A. Paul, F.E. Clark, Soil Microbiology and Biochemistry, Academic Press, SanDiego, 1996.
[36] J. Frouz, A. Nov�akov�a, The potential effect of high atmospheric CO2 on soilfungi-invertebrate interactions, Glob. Change Biol. 8 (2002) 339e344.
[37] F. Schinner, R. €Ohlinger, E. Kandeler, R. Margesin, Methods in Soil Biology,Springer Verlag, Berlin, Heidelberg, New York, 1996.
[38] F.C. Page, An Illustrated Key to Freshwater and Soil Amoebae, FreshwaterBiological Association, The Ferry House, Ableside, Cumbria, 1976.
[39] S. Scheu, Automated measurement of the respiratory response of soilmicrocompartments: active microbial biomass in earthworm faeces, Soil Biol.Biochem. 24 (1992) 1113e1118.
[40] T. Beck, R.G. Joergensen, E. Kandeler, F. Makeschin, E. Nuss, H.R. Oberholzer,S. Scheu, An inter-laboratory comparison of ten different ways of measuringsoil microbial biomass C, Soil Biol. Biochem. 29 (1997) 1023e1032.
[41] A.V. Tiunov, S. Scheu, Microbial biomass, biovolume and respiration in Lum-bricus terrestris L. cast material of different age, Soil Biol. Biochem. 32 (2000)

V. Jílkov�a et al. / European Journal of Soil Biology 69 (2015) 66e73 73
265e275.[42] J. �Snajdr, V. Val�a�skov�a, V. Merhautov�a, T. Cajthaml, P. Baldri�an, Activity and
spatial distribution of lignocellulose-degrading enzymes during forest soilcolonization by saprotrophic basidiomycetes, Enzyme Microb. Tech. 43 (2008)186e192.
[43] V.A. Knivett, J. Cullen, Some factors affecting cyclopropane acid formation inEscherichia coli, Biochem. J. 96 (1965) 771e776.
[44] J.H. Law, H. Zalkin, T. Kaneshiro, Transmethylation reactions in bacterial lipids,Biochim. Biophys. Acta 70 (1963) 143e151.
[45] D. Bossio, K. Scow, Impacts of carbon and flooding on soil microbial com-munities: phospholipid fatty acid profiles and substrate utilization patterns,Microb. Ecol. 35 (1998) 265e278.
[46] D. Bossio, K. Scow, N. Gunapala, K. Graham, Determinants of soil microbialcommunities: effects of agricultural management, season, and soil type onphospholipid fatty acid profiles, Microb. Ecol. 36 (1998) 1e12.
[47] R.R. Sokal, F.J. Rohlf, Biometry e the Principles and Practice of Statistics inBiological Research, W.H. Freeman & Co, New York, 1995.
[48] C.N. van Ende, Repeated-mesures analysis: growth and other time-dependentmeasures, in: S. Scheiner, J. Gurevitch (Eds.), Design and Analysis of EcologicalExperiments, Chapman and Hall, New York, 2001, pp. 134e157.
[49] J. Frouz, The effect of nest moisture on daily temperature regime in the nestsof Formica polyctena wood ants, Insect. Soc. 47 (2000) 229e235.
[50] M.P. Berg, J. Bengtsson, Temporal and spatial variability in soil food webstructure, Oikos 116 (2007) 1789e1804.
[51] M. Clarholm, Bacteria and protozoa as integral components of the forestecosystem e their role in creating a naturally varied soil fertility, Antonie VanLeeuw. J. Microb. 81 (2002) 309e318.
[52] S. Blagodatsky, E. Blagodatskaya, T. Yuyukina, Y. Kuzyakov, Model of apparentand real priming effects: linking microbial activity with soil organic matterdecomposition, Soil Biol. Biochem. 42 (2010) 1275e1283.
[53] M. Bonkowski, M. Clarholm, Stimulation of plant growth through interactionsof bacteria and protozoa: testing the auxiliary microbial loop hypothesis, ActaProtozool. 51 (2012) 237e247.
[54] M. Bonkowski, Protozoa and plant growth: the microbial loop in soil revisited,
New. Phytol. 162 (2004) 617e631.[55] F. Ekelund, S. Saj, M. Vestergard, J. Bertaux, J. Mikola, The “soil microbial loop”
is not always needed to explain protozoan stimulation of plants, Soil Biol.Biochem. 41 (2009) 2336e2342.
[56] A. Hodge, D. Robinson, A. Fitter, Are microorganisms more effective thanplants at competing for nitrogen? Trends Plant Sci. 5 (2000) 1360e1385.
[57] M. Mooshammer, W. Wanek, I. H€ammerle, L. Fuchslueger, F. Hofhansl,A. Knoltsch, J. Schnecker, M. Takriti, M. Watzka, B. Wild, K.M. Keiblinger,S. Zechmeister-Boltenstern, A. Richter, Adjustment of microbial nitrogen useefficiency to carbon:nitrogen imbalances regulates soil nitrogen cycling, Nat.Commun. 5 (2014) 3694.
[58] R.G. Joergensen, S. Scheu, Response of soil microorganisms to the addition ofcarbon, nitrogen and phosphorus in a forest Rendzina, Soil Biol. Biochem. 31(1999) 859e866.
[59] C. Matz, S. Kjelleberg, Off the hook e how bacteria survive protozoan grazing,Trends Microbiol. 13 (2005) 302e307.
[60] A. Jousset, Ecological and evolutive implications of bacterial defences againstpredators, Environ. Microbiol. 14 (2012) 1830e1843.
[61] A. Jousset, M. Bonkowski, The model predator Acanthamoeba castellanii in-duces the production of 2,4, DAPG by the biocontrol strain Pseudomonas flu-orescens Q2-87, Soil Biol. Biochem. 42 (2010) 1647e1649.
[62] Å. Frostegård, A. Tunlid, E. Bååth, Use and misuse of PLFA measurements insoils, Soil Biol. Biochem. 43 (2011) 1621e1625.
[63] J. Becker, N. Eisenhauer, S. Scheu, A. Jousset, Increasing antagonistic in-teractions cause bacterial communities to collapse at high diversity, Ecol. Lett.15 (2012) 468e474.
[64] M.E. Brown, Soil bacteriostasis limitation in growth of soil and rhizospherebacteria, Can. J. Microbiol. 19 (1973) 195e199.
[65] W.C. Ho, W.H. Ko, Soil microbiostasis: effects of environmental and edaphicfactors, Soil Biol. Biochem. 17 (1985) 167e170.
[66] W.H. Ko, F.K. Chow, Characteristics of bacteriostasis in natural soils, J. Gen.Microbiol. 102 (1977) 295e298.
[67] K. Vetsigian, R. Jajoo, R. Kishony, Structure and evolution of Streptomycesinteraction networks in soil and in silico, PLoS Biol. 9 (2011).




















