
The role of afferent inputs to supraoptic nucleus oxytocin neurons during naloxone-precipitated...
11
THE ROLE OF AFFERENT INPUTS TO SUPRAOPTIC NUCLEUS OXYTOCIN NEURONS DURING NALOXONE-PRECIPITATED MORPHINE WITHDRAWAL IN THE RAT N. P. MURPHY,*² T. ONAKA,*‡ C. H. BROWN§ and G. LENG*Q *Laboratory for Neuroendocrinology, Babraham Institute, Cambridge CB2 4AT, U.K. ²Department of Physiology, University Medical School, Teviot Place, Edinburgh EH8 9AG, U.K. Abstract––During prolonged exposure to morphine, oxytocin neurons of the rat supraoptic nucleus develop dependence, shown by hyperexcitation following morphine withdrawal. The present study investigated the role of afferent projections to the supraoptic nucleus in this withdrawal excitation. Rats were made morphine-dependent by continuous intracerebroventricular infusion of morphine at increasing doses (up to 50 μg/h). On the sixth day, rats were anaesthetized with pentobarbitone and morphine withdrawal was precipitated by intraperitoneal injection of naloxone (5 mg/kg). Fos-immunoreactivity in the supraoptic nucleus, and also in the median preoptic nucleus, organum vasculosum of the lamina terminalis and subfornical organ, which project to the supraoptic nucleus, increased following morphine withdrawal. However, retrograde tracing from the supraoptic nucleus showed that, of the neurons in these regions which project to the supraoptic nucleus, only 0.4–7.1% expressed Fos in response to morphine withdrawal. Following morphine withdrawal, Fos-immunoreactivity was present in 39.2% and 19.8% of the tyrosine hydroxylase-immunoreactive neurons of the A1/C1 and A2/C2 cell groups. Of the cells in these regions identified as projecting to the supraoptic nucleus, 11.3% in the region of the A2 cell group and 12.7% in the region of the A1 cell group expressed Fos after morphine withdrawal. In a second study, monoamine release was measured in the supraoptic nucleus of urethane-anaesthetized morphine- dependent and -naı ¨ve rats. Retrodialysis of naloxone (10 "5 M) into the supraoptic nucleus induced a small increase in plasma oxytocin concentration in morphine-dependent rats (13.5&4.8 pg/ml increase) but not in naı ¨ve rats (1.2&5.9 pg/ml decrease), with no significant change in monoamine release in either morphine-dependent or -naı ¨ve rats. Intravenous injection of naloxone (5 mg/kg) 1 h later produced a further significant increase in plasma oxytocin concentration in morphine-dependent rats concomitant with a significant increase in noradrenaline release from the supraoptic nucleus. Thus, morphine-withdrawal excitation of supraoptic oxytocin neurons occurs concurrently with a modestly increased activity of their input from the brainstem, and very little activation in other known inputs. ? 1997 IBRO. Published by Elsevier Science Ltd. Key words: opioid, noradrenaline, A2 cell group, median preoptic nucleus, organum vasculosum of the lamina terminalis, subfornical organ. The oxytocin neurons of the supraoptic and para- ventricular nuclei are highly sensitive to inhibition by morphine. 21,23,24 During continuous i.c.v. infusion of morphine, the oxytocin system becomes tolerant to its inhibitory effects, and concurrently develops dependence upon morphine. Acute withdrawal of morphine, by administration of the opiate antagon- ist naloxone, induces a prolonged hyperexcitation of oxytocin neurons leading to hypersecretion of oxytocin. 2 This withdrawal hyperexcitation may reflect an intrinsic hyperexcitability of the oxytocin neuron, or an increased afferent excitation, or a combination of both. Oxytocin neurons express only low levels of mRNA for the μ-opioid receptor through which morphine acts. 30 Despite this, there are indications that the oxytocin neurons themselves are a primary target for the actions of morphine. However, there are also functionally coupled μ-receptors located presynaptically on afferent nerve endings within the supraoptic nucleus, 20 and morphine is also known to act at several of the sites of origin of these afferent nerve endings which possess μ-opioid recep- tors. 15,18,19 Potentially, dependence in the oxytocin system could develop at any or all of these levels. ²Present address: Department of Psychiatry and Biobehav- ioral Sciences, Neuropsychiatric Institute, M.R.L., U.C.L.A., 760 Westwood Plaza, Los Angeles, CA 90024, U.S.A ²Present address: Department of Physiology, Jichi Medical School, Minamikawachi-Machi, Tochigi-Ken, 329-04, Japan. QTo whom correspondence should be addressed. Abbreviations: CCK, cholecystokinin; DAB, 3,3*- diaminobenzidine; FITC, fluorescein isothiocyanate; MnPO, median preoptic nucleus; NTS, nucleus tractus solitarii; OVLT, organum vasculosum of the lamina terminalis; SFO, subfornical organ; TH, tyrosine hydroxylase. Pergamon Neuroscience Vol. 80, No. 2, pp. 567–577, 1997 Copyright ? 1997 IBRO. Published by Elsevier Science Ltd Printed in Great Britain. All rights reserved 0306–4522/97 $17.00+0.00 PII: S0306-4522(97)00142-5 567
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of The role of afferent inputs to supraoptic nucleus oxytocin neurons during naloxone-precipitated...

THE ROLE OF AFFERENT INPUTS TO SUPRAOPTICNUCLEUS OXYTOCIN NEURONS DURING
NALOXONE-PRECIPITATED MORPHINE WITHDRAWALIN THE RAT
N. P. MURPHY,*† T. ONAKA,*‡ C. H. BROWN§ and G. LENG*Q*Laboratory for Neuroendocrinology, Babraham Institute, Cambridge CB2 4AT, U.K.
†Department of Physiology, University Medical School, Teviot Place, Edinburgh EH8 9AG, U.K.
Abstract––During prolonged exposure to morphine, oxytocin neurons of the rat supraoptic nucleusdevelop dependence, shown by hyperexcitation following morphine withdrawal. The present studyinvestigated the role of afferent projections to the supraoptic nucleus in this withdrawal excitation. Ratswere made morphine-dependent by continuous intracerebroventricular infusion of morphine at increasingdoses (up to 50 µg/h). On the sixth day, rats were anaesthetized with pentobarbitone and morphinewithdrawal was precipitated by intraperitoneal injection of naloxone (5 mg/kg). Fos-immunoreactivity inthe supraoptic nucleus, and also in the median preoptic nucleus, organum vasculosum of the laminaterminalis and subfornical organ, which project to the supraoptic nucleus, increased following morphinewithdrawal. However, retrograde tracing from the supraoptic nucleus showed that, of the neurons in theseregions which project to the supraoptic nucleus, only 0.4–7.1% expressed Fos in response to morphinewithdrawal. Following morphine withdrawal, Fos-immunoreactivity was present in 39.2% and 19.8% ofthe tyrosine hydroxylase-immunoreactive neurons of the A1/C1 and A2/C2 cell groups. Of the cells inthese regions identified as projecting to the supraoptic nucleus, 11.3% in the region of the A2 cell groupand 12.7% in the region of the A1 cell group expressed Fos after morphine withdrawal. In a second study,monoamine release was measured in the supraoptic nucleus of urethane-anaesthetized morphine-dependent and -naı̈ve rats. Retrodialysis of naloxone (10"5 M) into the supraoptic nucleus induced asmall increase in plasma oxytocin concentration in morphine-dependent rats (13.5&4.8 pg/ml increase)but not in naı̈ve rats (1.2&5.9 pg/ml decrease), with no significant change in monoamine release in eithermorphine-dependent or -naı̈ve rats. Intravenous injection of naloxone (5 mg/kg) 1 h later produced afurther significant increase in plasma oxytocin concentration in morphine-dependent rats concomitantwith a significant increase in noradrenaline release from the supraoptic nucleus.Thus, morphine-withdrawal excitation of supraoptic oxytocin neurons occurs concurrently with a
modestly increased activity of their input from the brainstem, and very little activation in other knowninputs. ? 1997 IBRO. Published by Elsevier Science Ltd.
Key words: opioid, noradrenaline, A2 cell group, median preoptic nucleus, organum vasculosum of thelamina terminalis, subfornical organ.
The oxytocin neurons of the supraoptic and para-ventricular nuclei are highly sensitive to inhibition bymorphine.21,23,24 During continuous i.c.v. infusion ofmorphine, the oxytocin system becomes tolerant toits inhibitory effects, and concurrently developsdependence upon morphine. Acute withdrawal of
morphine, by administration of the opiate antagon-ist naloxone, induces a prolonged hyperexcitationof oxytocin neurons leading to hypersecretion ofoxytocin.2
This withdrawal hyperexcitation may reflect anintrinsic hyperexcitability of the oxytocin neuron, oran increased afferent excitation, or a combination ofboth. Oxytocin neurons express only low levels ofmRNA for the µ-opioid receptor through whichmorphine acts.30 Despite this, there are indicationsthat the oxytocin neurons themselves are a primarytarget for the actions of morphine. However, thereare also functionally coupled µ-receptors locatedpresynaptically on afferent nerve endings within thesupraoptic nucleus,20 and morphine is also known toact at several of the sites of origin of these afferentnerve endings which possess µ-opioid recep-tors.15,18,19 Potentially, dependence in the oxytocinsystem could develop at any or all of these levels.
†Present address: Department of Psychiatry and Biobehav-ioral Sciences, Neuropsychiatric Institute, M.R.L.,U.C.L.A., 760 Westwood Plaza, Los Angeles, CA 90024,U.S.A
†Present address: Department of Physiology, Jichi MedicalSchool, Minamikawachi-Machi, Tochigi-Ken, 329-04,Japan.
QTo whom correspondence should be addressed.Abbreviations: CCK, cholecystokinin; DAB, 3,3*-diaminobenzidine; FITC, fluorescein isothiocyanate;MnPO, median preoptic nucleus; NTS, nucleus tractussolitarii; OVLT, organum vasculosum of the laminaterminalis; SFO, subfornical organ; TH, tyrosinehydroxylase.
Pergamon
Neuroscience Vol. 80, No. 2, pp. 567–577, 1997Copyright ? 1997 IBRO. Published by Elsevier Science Ltd
Printed in Great Britain. All rights reserved0306–4522/97 $17.00+0.00PII: S0306-4522(97)00142-5
567

The oxytocin neurons of the supraoptic nucleusreceive major excitatory inputs from anterior struc-tures adjacent to the wall of the third ventricle: themedian preoptic nucleus (MnPO), the organum vas-culosum of the lamina terminalis (OVLT) and thesubfornical organ (SFO), and from the nucleus trac-tus solitarii (NTS) in the brainstem,4,7 all of whichare possible sites mediating the actions of morphineon oxytocin neurons. In particular, neurons in theNTS develop morphine dependence and undergowithdrawal excitation.29 While the NTS principallyprojects to oxytocin neurons,5,22 the MnPO, OVLTand SFO project to both oxytocin and vasopressinneurons in the supraoptic nucleus, but it is not clearwhether individual neurons in these structures projectto both oxytocin and vasopressin neurons, orwhether different subpopulations project to the twocell types. Vasopressin neurons are not consistentlyinhibited by morphine nor do they show a consistentresponse to morphine withdrawal. In addition, al-though some neurons in the MnPO, OVLT and SFOexpress µ-receptors, it is by no means certain that theneurons which project to the magnocellular systemdo so.This study investigated the activity of the afferent
projections to the supraoptic nucleus at the time ofwithdrawal excitation of oxytocin neurones. To thisend, we determined the withdrawal-induced expres-sion of Fos protein in neurons identified by retro-grade tracing as projecting to the supraoptic nucleus.Fos is an immediate-early gene product which, insome systems at least, has proved an efficient markerof neuronal activation. The possible involvement ofnoradrenergic projections was further investigated bymicrodialysis studies of noradrenaline release withinthe supraoptic nucleus.
EXPERIMENTAL PROCEDURES
Induction of morphine tolerance/dependence
Adult virgin female Wistar rats of the Babraham Institutecolony (180–255 g) were anaesthetized with Avertin (2% w/vtribromoethanol /8% ethanol/1.2% amyl alcohol in 0.9%saline, 10 ml/kg, i.p.) and placed in a stereotaxic frame. Anosmotic minipump (Alzet 2001, Charles River, U.K.) wasimplanted subcutaneously over the scapula, to deliver anincreasing dose (10 µg/h for 40 h, 20 µg/h for 40 h, 50 µg/hthereafter) of morphine sulphate (Sigma., U.K. dissolved insterile water) into the right lateral ventricle (3.0 mm caudal,2.0 mm lateral to bregma) through a stainless steel cannula(21G, length 4.5 mm from surface of skull) for five days.23
Animals were housed individually after surgery intemperature- and light-controlled conditions (21–23)C; 13/11 h light–dark), and were allowed food (standard breedermix) and water ad libitum. All procedures were carried outin accordance with the U.K. Animals (Scientific Procedures)Act, 1986 and the minimum number of animals were usedthat would allow for statistical analyses of the results.
Immunohistochemistry
Morphine-dependent rats were anaesthetized with so-dium pentobarbitone (60 mg/kg, i.p.) and 15–90 min later,were injected with naloxone hydrochloride (Sigma, 5 mg/kg,0.5 ml/kg in 0.9% saline, i.p.) or vehicle. After a further
90 min, the rats were killed by sodium pentobarbitoneoverdose and perfused transcardially with heparinizedsaline (50 mU/ml) followed by 4% paraformaldehyde in0.1 M phosphate buffer. Brains were removed and post-fixed for 2 h in the same fixative containing 15% sucrose.They were then transferred to a 30% sucrose solution in0.1 M phosphate buffer until sectioning. 35 µm coronalsections were cut through the hypothalamus and brainstemon a freezing microtome and collected into 0.1 M phosphatebuffer. The sections were then processed for detection ofFos protein using a rabbit polyclonal antibody raisedagainst the N-terminal [2–17] of rat Fos protein (diluted1:10000; kindly donated by Drs G. I. Evans and D. C.Hancock, Imperial Cancer Research Fund, UK), and incu-bated overnight at 4)C. After a second overnight incubationat 4)C with peroxidase-labelled goat anti-rabbit IgG (VectorLaboratories, Peterborough, U.K. diluted 1:500), Fosimmunoreactivity was visualized as a black nuclear precipi-tate using a glucose oxidase-based, nickel-intensified, 3,3*-diaminobenzidine (DAB) procedure (adapted from Shuet al.27). Sections were mounted on gelatinized slides andviewed under a Leitz DM RB microscope.Some brainstem sections were also labelled for tyrosine
hydroxylase (TH). Fos-immunolabelled sections were incu-bated in a mouse monoclonal antibody directed against TH(Chemicon, U.S.A., 1:4000) overnight at 4)C, followed byincubation with biotinylated horse anti-mouse IgG (Vector,U.K.; 1:2000) for 2 h, and then with streptavidin–biotinylated horseradish peroxidase complex (Amersham,U.K.; 1:200) for 2 h at room temperature. TH immunore-activity was visualized as a brown cytoplasmic precipitateusing a glucose oxidase-based, DAB procedure.27
For Fos immunohistochemistry, five sections each wereprocessed from the supraoptic nucleus and five to 10sections from the brainstem. The number of Fos-immunoreactive profiles per section were counted for eacharea and the mean per section calculated for each animal.For Fos and TH double-labelled brainstem sections, allFos-, TH- and double-labelled neurons in the A1/C1 andA2/C2 region were counted between "1680 and +720 µmrelative to the obex.
Retrograde tracing studies
Six rats anaesthetized with Avertin were placed in astereotaxic frame and the dorsal surface of the skull ex-posed. Two 1 mm holes were drilled in the skull (1.7 mm leftand right lateral, 0.9 mm posterior of bregma) throughwhich 0.5 µl of a suspension of fluorescently-labelled latexmicrospheres (Lumafluor Inc., USA, at concentration sup-plied) was injected into each supraoptic nucleus over 5 minusing a 1 µl Hamilton syringe lowered 9.1 mm from thesurface of the skull; rhodamine (red)- and fluorescein iso-thiocyanate (FITC, green)- labelled microspheres (0.02–0.2 µm diameter) were used for right and left micro-injections, respectively. The cannula was left in place for5 min after each injection, and after the second injectionthe skin over the skull was sutured and the rats housedindividually for 84–91 days.All rats injected with tracer were subsequently treated
with morphine to induce dependence as described above,and, after naloxone-induced withdrawal, brain sectionswere cut and processed for immunohistochemical identifi-cation of Fos protein as previously described. Retrogradelabelling was observed on at least three sections per area perrat (n=3–6) using a microscope (Leitz DM RB, Germany)equipped with an ultraviolet light source.
Microdialysis and blood sampling
Rats were anaesthetized with urethane (ethyl carbamate,1.25 g/kg, i.p.), tracheotomized and the right jugular veincannulated for administration of drugs and withdrawal ofblood samples. The right supraoptic nucleus was exposedby transpharyngeal surgery as previously described.13 A
568 N. P. Murphy et al.

concentric microdialysis probe (0.5 mm o.d.#1 mm mem-brane length, CMA/microdialysis, Sweden) was placed inthe exposed supraoptic nucleus, immediately rostral to thedecussation of the internal penetrating artery, and left for1 h before sample collection. The probes had in vitro recov-ery rates of 16&3% for GABA measured at the perfusionrates used in the experiments. Probe placement was verifiedhistologically at the end of the experiment. In five controlexperiments the probe was placed approximately 1 mmrostral to the supraoptic nucleus to determine whether anyresponses found were specific to the supraoptic nucleus.Ringer’s solution (147 mM NaCl, 4 mM KCl, 2.5 mM
CaCl2) was filtered, degassed and pumped through theprobe at 0.6 µl/min using a syringe pump (Harvard appara-tus, USA). Samples were collected over consecutive 20 minperiods into 0.5 ml microfuge tubes on ice, into which 2 µlof 0.05 M perchloric acid had been placed to minimizeoxidation of noradrenaline. After three 20 min samples, thedialysate was switched to Ringer’s solution containing10"5 M naloxone. Five minutes before and after thischange, 0.3 ml blood samples were removed. After collec-tion of a further three 20 min microdialysate samples,naloxone was injected (5 mg/kg, i.v.). A further bloodsample was removed 5 min later. Three more 20 min dialysissamples were collected prior to the termination of theexperiment.Dialysis samples were frozen on dry ice and stored at
"80)C for future analysis of monoamine content by highperformance liquid chromatography coupled to an electro-chemical detector as described previously.12 The detectionlimit for noradrenaline, dopamine and serotonin were 10, 20and 40 pg/ml, respectively. The mean monoamine concen-trations in the three samples for each treatment period werecalculated and used for subsequent statistical analyses.The plasma (separated by centrifugation) was stored at
"20)C before determination of oxytocin content by radio-immunoassay (RIA), using a method modified from thatof Higuchi et al.,11 as described elsewhere.14 The interassayand intraassay coefficients of variation were 11.8% and9.9%, respectively. Blood cells were suspended in 0.9%saline and injected through the jugular catheter followingremoval of the subsequent sample.
Statistics
All statistical tests were completed on the SigmaStatsoftware package (Jandel Scientific GmbH, 40699 Erkrath,Germany).
RESULTS
Effects of morphine withdrawal on Fos expression inthe hypothalamus and brainstem
Fos-immunoreactivity in morphine-dependent rats(n=6) was virtually absent from all of the areas underinvestigation, as has been previously demonstrated.24
By contrast, the brains of morphine-withdrawn ratsshowed extensive induction of intense nuclear Fos-like immunoreactivity in the supraoptic and paraven-tricular nuclei. In the supraoptic nucleus, 67.9&22.5(mean&S.E.M.) Fos-immunoreactive nuclei/sectionwere detected in morphine-withdrawn rats (n=6),compared with 1.4&0.9 in morphine-dependent rats(P<0.01; Mann–Whitney U-test).Fos expression in the NTS of these animals showed
a similar pattern; in dependent rats (n=4) Fos-immunoreactivity was present in 8.2&4.1 nuclei/section while in withdrawn rats (n=4) significantlymore Fos-immunoreactive nuclei were present
(43.7&10.8 nuclei/section; P<0.01; Mann–WhitneyU-test). Other brainstem regions at this level showedno conspicuous Fos expression in either group ofrats, with the exception of the area postrema and A1cell group in the ventrolateral medulla, which werenot counted systematically but which showed Fosexpression in all withdrawn rats, but not in controlrats (morphine-dependent rats treated with morphinebut not withdrawn with naloxone).In the sections double-labelled for Fos protein and
TH (Fig. 1), Fos was detected in 53 neurons in theA2/C2 region of control, morphine-dependent rats,of which 30 contained TH, comprising 2.1% of theTH cell population counted (1457 neurons counted in21–31 sections per rat). In morphine-withdrawn rats,Fos was detected in 1430 neurons of which 242contained TH, comprising 19.8% of the TH neurons(1224 neurons counted). Fos was also detected in 47neurons in the A1/C1 region of morphine-dependentrats, of which 33 contained TH, comprising 2.1% ofthe TH neurons (1561 neurons counted in 22–30sections per rat). In morphine-withdrawn rats, Foswas detected in 616 neurons of which 319 con-tained TH, comprising 39.2% of the TH neurons(814 neurons). The rostrocaudal distribution of Fosexpression is mapped in Fig. 2.
Retrograde tracing studies
In morphine-withdrawn rats, but not morphine-dependent rats, there was consistently high Fos ex-pression in both dorsal and ventral portions of theMnPO, in the OVLT (Fig. 3), and, more variably, inthe SFO. These regions were analysed in detail fromsix rats in which retrograde tracer had been placed inone or both of the supraoptic nuclei prior to theinduction of morphine-dependence. In each case thisresulted in extensive retrograde labelling in the threeregions.In each brain, three to four sections were counted
in the region of the OVLT, revealing between 48 and107 Fos-positive nuclei per section in the differentrats, scattered throughout the OVLT (total of 1604Fos-positive neurons examined). Between 12 and65.5 retrogradely-labelled neurons per section weredetected in the different rats, the variability reflectingdifferences in the density of filling of the supraopticnucleus with tracer, and in some cases resulting frombilateral injection sites. The labelled neurons wereprincipally distributed around the dorsal perimeter ofthe OVLT without apparent lateral bias. Of the 752retrogradely-labelled neurons found in this area, only53 were also Fos positive (7.1%, range 3.3–8.4%between rats).Similarly in the MnPO, three to four sections were
counted in each brain, revealing between 3.7 and 31.7Fos-positive nuclei per section in the different rats(total of 315 Fos-positive neurons examined). Be-tween 17.3 and 58.7 retrogradely-labelled neuronsper section were detected between different rats. Of
Oxytocin afferent activity in morphine withdrawal 569
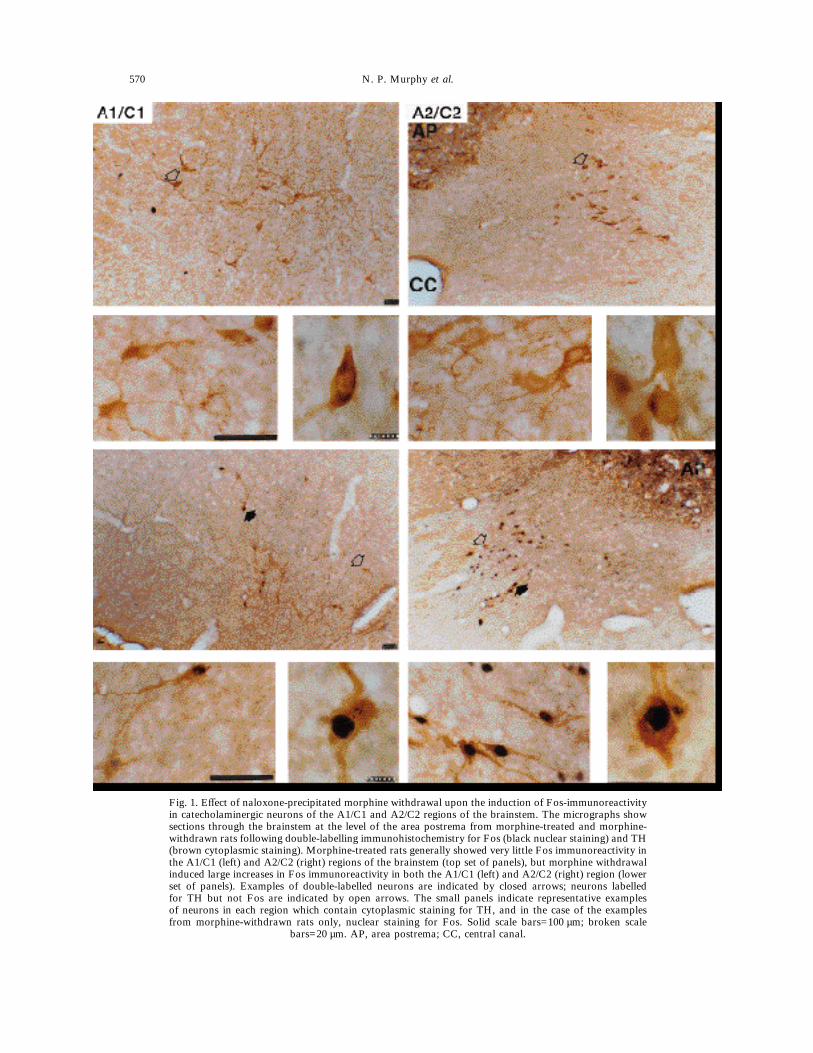
Fig. 1. Effect of naloxone-precipitated morphine withdrawal upon the induction of Fos-immunoreactivityin catecholaminergic neurons of the A1/C1 and A2/C2 regions of the brainstem. The micrographs showsections through the brainstem at the level of the area postrema from morphine-treated and morphine-withdrawn rats following double-labelling immunohistochemistry for Fos (black nuclear staining) and TH(brown cytoplasmic staining). Morphine-treated rats generally showed very little Fos immunoreactivity inthe A1/C1 (left) and A2/C2 (right) regions of the brainstem (top set of panels), but morphine withdrawalinduced large increases in Fos immunoreactivity in both the A1/C1 (left) and A2/C2 (right) region (lowerset of panels). Examples of double-labelled neurons are indicated by closed arrows; neurons labelledfor TH but not Fos are indicated by open arrows. The small panels indicate representative examplesof neurons in each region which contain cytoplasmic staining for TH, and in the case of the examplesfrom morphine-withdrawn rats only, nuclear staining for Fos. Solid scale bars=100 µm; broken scale
bars=20 µm. AP, area postrema; CC, central canal.
570 N. P. Murphy et al.

the 630 retrogradely-labelled neurons examined inthis area, 30 were also Fos positive (4.7%, range0–8.5% between rats).In each SFO, four to five sections were counted,
revealing between 2.3 and 9.5 Fos-positive nuclei persection in the different rats, scattered throughout theSFO. Between 12.3 and 76.3 retrogradely-labelledneurons per section were detected in the differentrats. The labelled neurons were principally distrib-uted around the perimeter of the SFO without appar-ent lateral bias. Of the 898 retrogradely-labelledneurons examined, only four were also Fos positive(0.4%, range 0–2.3%).
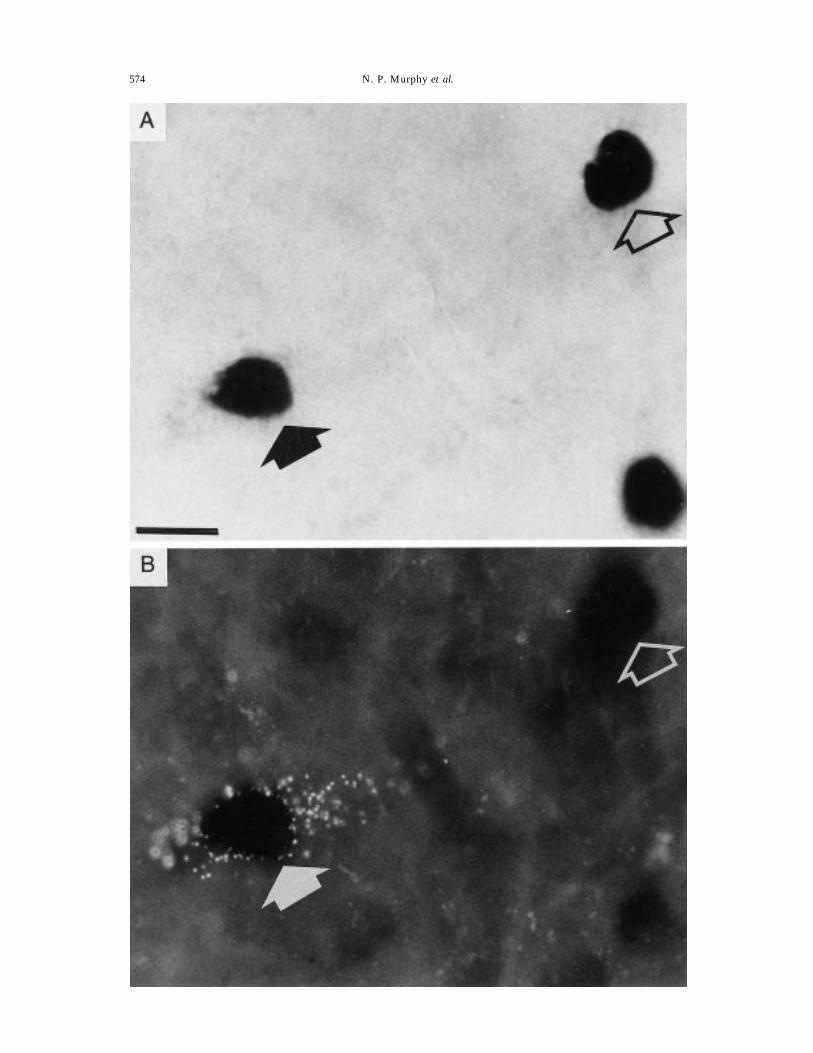
Detailed counts were made in the brainstem of threerats, in each of which injections of tracer were princi-pally confined to the supraoptic nucleus on both sides.In each rat, 14 brainstem sections were analysed. Inthe A2 region, 18, 24 and 55 retrogradely-labelledneurons were detected in the three rats (Fig. 4), ofwhich 87–96% projected to the ipsilateral supraopticnucleus, and of which 11.3% expressed nuclear Fos-like immunoreactivity. In the A1 region for the samerats, 34, 64 and 52 retrogradely-labelled neurons weredetected of which 67–78% projected to the ipsilateralsupraoptic nucleus, and of which 12.7% expressednuclear Fos-like immunoreactivity.
Fig. 2. Effect of naloxone-precipitated morphine withdrawal upon the number of catecholaminergicneurons of the brainstem expressing Fos immunoreactivity. The number of neurons showing Fos-(closedcircles) and TH-(open circles) immunoreactivity were counted in the A1/C1 and A2/C2 regions of thebrainstem (between "1680 and 720 µm relative to the obex) in morphine-treated and withdrawn rats.Each panel represents the means&S.E.M. for 11 sections through a single region of the brainstem for eachgroup. Note that morphine withdrawal induced increases in Fos expression in TH-immunoreactive
neurons in both the A1/C1 and A2/C2 regions (triangles).
Oxytocin afferent activity in morphine withdrawal 571
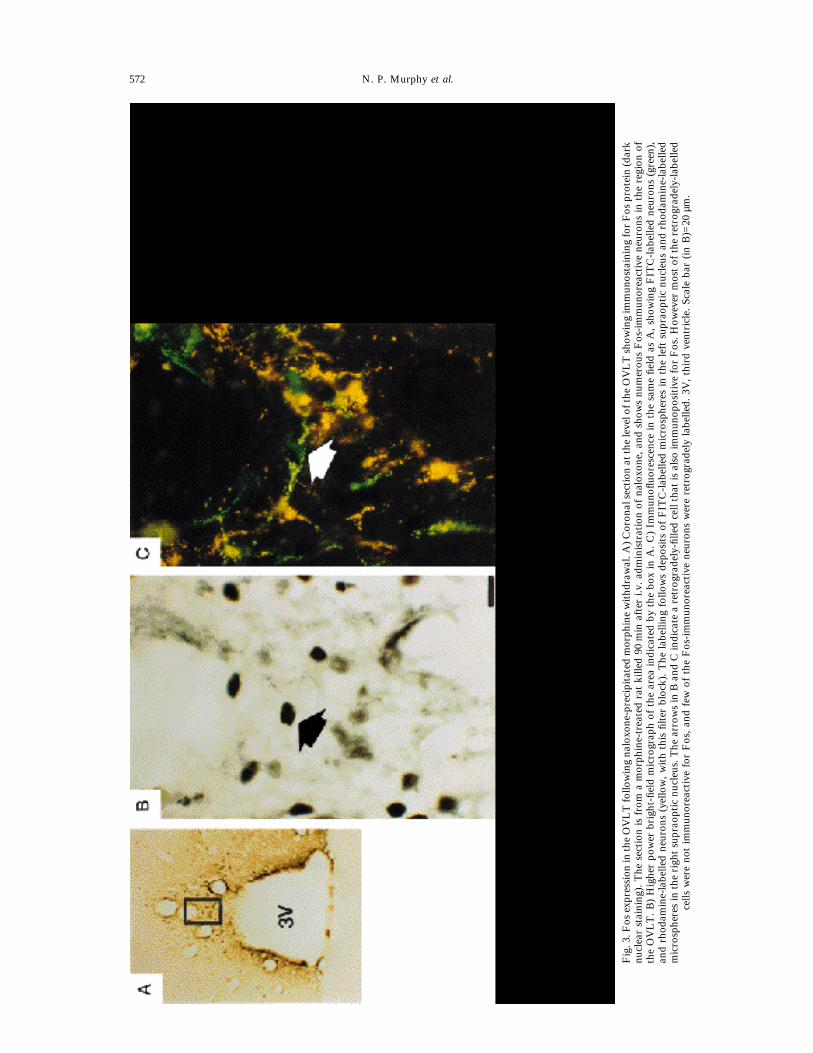
Fig.3.Fosexpression
intheOVLTfollowingnaloxone-precipitatedmorphinewithdrawal.A)CoronalsectionattheleveloftheOVLTshowingimmunostainingforFosprotein(dark
nuclearstaining).Thesectionisfrom
amorphine-treatedratkilled90minafteri.v.administrationofnaloxone,andshowsnumerousFos-immunoreactiveneuronsintheregion
oftheOVLT.B)Higherpowerbright-fieldmicrographoftheareaindicatedby
theboxinA.C)ImmunofluorescenceinthesamefieldasA,showingFITC-labelledneurons(green),
andrhodamine-labelledneurons(yellow,withthisfilterblock).Thelabelling
followsdepositsofFITC-labelledmicrospheresintheleftsupraopticnucleusandrhodamine-labelled
microspheresintherightsupraopticnucleus.ThearrowsinBandCindicatearetrogradely-filledcellthatisalsoimmunopositiveforFos.H
owevermostoftheretrogradely-labelled
cellswerenotimmunoreactiveforFos,andfewoftheFos-immunoreactiveneuronswereretrogradelylabelled.3V,thirdventricle.Scalebar(inB)=20µm
.
572 N. P. Murphy et al.

Effect of retrodialysis of naloxone into the supraopticnucleus of morphine-dependent and naı̈ve rats
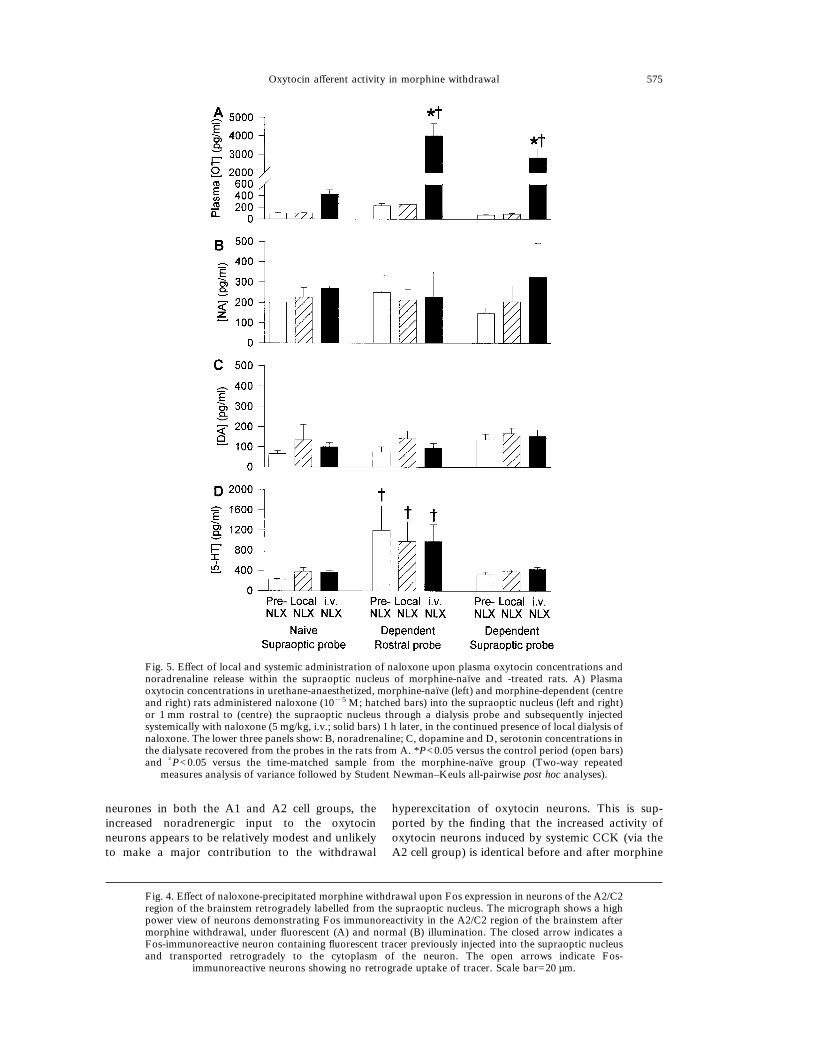
Retrodialysis of naloxone into the supraoptic nu-cleus of morphine-dependent rats (n=11) produced asmall increase in plasma oxytocin (+13.5&4.8 pg/ml)which was significantly greater than the changeobserved in morphine-naı̈ve controls (n=12,"1.2&5.9 pg/ml, P<0.05 Mann–Whitney U-test).Local administration of naloxone into the supraopticnucleus produced a small, transient and non-significant increase of noradrenaline in the supraopticnucleus in both morphine-treated and morphine-naı̈ve rats. There were no significant changes in therelease of dopamine or serotonin in either group, andno significant differences between groups, except thatthe serotonin levels in the probes placed rostral to thesupraoptic nucleus were elevated with respect tothose placed within the nucleus.Following systemic administration of naloxone
(5 mg/kg, i.v.), there was a further non-significantincrease in the release of noradrenaline measured inmorphine-naı̈ve rats (to 146.1&30.1% of the controllevels) while in morphine-dependent rats the furtherincrease (to 183.3&64.5% of the control levels) wassignificantly elevated above the pre-i.v. naloxoneconcentration (P<0.05, Wilcoxon signed rank test).The increase in noradrenaline detected followingwithdrawal in the five rats in which the microdialysisprobe was placed rostral to the supraoptic nucleuswas to only 137.0&88.8% of control levels (Fig. 5).Systemic injection of naloxone induced, as expected,a significant increase in plasma oxytocin in morphinenaı̈ve rats, and a significantly larger increase inmorphine-dependent rats.
DISCUSSION
Naloxone-precipitated morphine withdrawal in-duces a profound increase in the electrical activity ofmagnocellular oxytocin neurons resulting in hyper-secretion of oxytocin from the neurohypophysisinto the systemic circulation,21 and also inducesincreased secretion of oxytocin from dendrites intothe supraoptic nucleus itself.25 The present studydemonstrates that morphine-withdrawal hyperexcita-tion of supraoptic nucleus oxytocin neurons isaccompanied by increased expression of Fos immu-noreactivity in the supraoptic nucleus and in somebrain areas which project to the supraoptic nucleus.The oxytocin neurons of the supraoptic nucleus
receive major direct excitatory inputs from the NTS,SFO, OVLT and MnPO,7 and all of these regionsexhibited increased expression of Fos protein follow-ing withdrawal. This did not simply reflect indiscrimi-nate activation of all brain structures; indeed, at thebrain levels studied, the increased expression of Fosappeared to be principally confined to these areasmentioned, with the exception that extensive Foslabelling was also present in the A1/C1 region of theventrolateral medulla, which projects to vasopressin
neurons rather than oxytocin neurons.22 However,although morphine withdrawal thus seemed to in-duce Fos expression throughout the magnocellularsystem and its afferent inputs, detailed analysisshowed that this superficial appearance was mis-leading.In the anterior structures (MnPO, OVLT and
SFO), very few of the neurons which could beidentified as projecting to the supraoptic nucleus wereactivated to express Fos following withdrawal. Thisinferred lack of involvement of the anterior periven-tricular structures is consistent with our previousobservations that oxytocin neurons still undergo hy-perexcitation following morphine withdrawal evenafter electrolytic lesion of the entire region adjacentto the anterior and ventral walls of the thirdventricle.26
Only in the NTS was there notable activationamongst retrogradely-labelled neurons, and evenhere the activation was relatively unimpressive. Ofneurons that projected to the supraoptic nucleus,only about 15% were activated by morphine with-drawal; by comparison, as we found using similartechniques in a recent study, more than 50% ofprojecting neurons in the NTS are activated follow-ing systemic administration of cholecystokinin(CCK),19 a stimulus which leads to a relatively muchsmaller and more transient activation of oxytocinneurons than is seen during morphine withdrawal.10
Of the neurons in the NTS that project to thesupraoptic nucleus,19,22 approximately 80% arenoradrenergic. Clonidine (á2-noradrenergic agonist)relieves the behavioural symptoms evident in opiateaddicts upon withdrawal,8 suggesting that noradren-ergic systems may be important in withdrawal exci-tation of their efferent neurones. Similarly, thebehavioural signs of withdrawal in rats are reducedby clonidine administration into the locus coeru-leus.31 Clonidine also reduces morphine-withdrawal-induced increases in noradrenaline release in thehippocampus6,28 and i.c.v. clonidine prevents thereduction in noradrenaline content in the hypo-thalamus which occurs after naloxone-precipitatedwithdrawal.9,16
Here, morphine withdrawal appeared to cause arise in noradrenaline release into the supraopticnucleus of morphine-dependent rats. The magnitudeof this release was equivalent to, or less than, thatinduced in similar studies by systemic administrationof CCK,12 but the release appeared to be moreprolonged. However, whereas CCK activates the A2,but not A1, neurons which project to the supraopticnucleus,19 morphine withdrawal activated a similarproportion of projecting neurons in both brainstemregions. Since the A1 projection is to vasopressinneurons which are little affected by morphine with-drawal, these microdialysis studies therefore addsupport to the conclusion that although mor-phine withdrawal is accompanied by activation ofnoradrenaline release resulting from activation of
Oxytocin afferent activity in morphine withdrawal 573
574 N. P. Murphy et al.
neurones in both the A1 and A2 cell groups, theincreased noradrenergic input to the oxytocinneurons appears to be relatively modest and unlikelyto make a major contribution to the withdrawal
hyperexcitation of oxytocin neurons. This is sup-ported by the finding that the increased activity ofoxytocin neurons induced by systemic CCK (via theA2 cell group) is identical before and after morphine
Fig. 5. Effect of local and systemic administration of naloxone upon plasma oxytocin concentrations andnoradrenaline release within the supraoptic nucleus of morphine-naı̈ve and -treated rats. A) Plasmaoxytocin concentrations in urethane-anaesthetized, morphine-naı̈ve (left) and morphine-dependent (centreand right) rats administered naloxone (10"5 M; hatched bars) into the supraoptic nucleus (left and right)or 1 mm rostral to (centre) the supraoptic nucleus through a dialysis probe and subsequently injectedsystemically with naloxone (5 mg/kg, i.v.; solid bars) 1 h later, in the continued presence of local dialysis ofnaloxone. The lower three panels show: B, noradrenaline; C, dopamine and D, serotonin concentrations inthe dialysate recovered from the probes in the rats from A. *P<0.05 versus the control period (open bars)and †P<0.05 versus the time-matched sample from the morphine-naı̈ve group (Two-way repeated
measures analysis of variance followed by Student Newman–Keuls all-pairwise post hoc analyses).
Fig. 4. Effect of naloxone-precipitated morphine withdrawal upon Fos expression in neurons of the A2/C2region of the brainstem retrogradely labelled from the supraoptic nucleus. The micrograph shows a highpower view of neurons demonstrating Fos immunoreactivity in the A2/C2 region of the brainstem aftermorphine withdrawal, under fluorescent (A) and normal (B) illumination. The closed arrow indicates aFos-immunoreactive neuron containing fluorescent tracer previously injected into the supraoptic nucleusand transported retrogradely to the cytoplasm of the neuron. The open arrows indicate Fos-
immunoreactive neurons showing no retrograde uptake of tracer. Scale bar=20 µm.
Oxytocin afferent activity in morphine withdrawal 575

withdrawal3 and by the failure of noradrenergicablation to prevent morphine withdrawal-inducedhypersecretion of oxytocin.17 No changes were ap-parent in the release of either of the other mono-amines measured (dopamine and serotonin) in thesupraoptic nucleus over the course of the experiment.Retrodialysis of a small concentration of naloxone
into a single supraoptic nucleus produced a smalland transient rise in noradrenaline release inboth morphine-dependent and -naı̈ve rats, and inmorphine-dependent rats only, naloxone resulted in asmall, non-significant rise in plasma oxytocin concen-trations. This rise thus appeared to be independent ofany directly evoked noradrenaline release, and indeedthere was no detected correlation between the mag-nitude of the measured oxytocin release and themagnitude of the observed noradrenaline releasefrom the data for individual rats (r=0.136, P=0.674).The magnitude of oxytocin release in response to
intranuclear naloxone was very much smaller thanthat which followed systemic naloxone. However, itshould be recalled that one supraoptic nucleus con-tains in all only about 20% of the oxytocin neuronswhich project to the neural lobe and it is unlikely thatsubstances administered through the dialysis probeaccess all oxytocin neurons. Moreover, the observedincrease in systemic release reflects not only acti-
vation of oxytocin neurons, but also an enhancementof stimulus-secretion coupling at the neural lobe itselfas a consequence of antagonism of endogenousê-opioid auto-inhibitory mechanisms which operateat the level of the nerve terminals.1 This peripheralopioid antagonism leads typically to a doublingin the efficiency of stimulus-secretion coupling inboth morphine-naı̈ve and morphine-dependent ratsequally.24
CONCLUSION
Thus, this locally-evoked oxytocin release appearslikely to reflect an action of naloxone at neuronalelements within the supraoptic nucleus itself thatdisplay dependence. The present observations suggestthat withdrawal hyperexcitation of oxytocin neuronsprincipally reflects a withdrawal response intrinsic tothe oxytocin neurons themselves.
Acknowledgements—Supported by the BBSRC. N.P.M. wassupported by a BBSRC scholarship and T.O. by the BritishCouncil and Ciba-Geigy Foundation (Japan). We are grate-ful to Drs G. I. Evans and D.C. Hancock for their kind giftof anti-Fos and anti-Jun antibodies, and T. Higuchi for theanti-oxytocin antibody. The authors wish to thank Dr K.Kendrick, Mr C. Chapman and Ms S. Dye for their adviceand assistance.
REFERENCES
1. Bicknell R. J. (1988) Optimizing release from peptide hormone secretory nerve terminals. J. exp. Biol. 139, 51–65.2. Bicknell R. J., Leng G. and Russell J. A. (1987) Oxytocin neurones: tolerance to prolonged morphine exposure and
hypersecretion following naloxone following withdrawal. In Inactivation of Hypersensitive Neurones (ed. ChalazonitisN. and Gola M.), pp. 187–194. Alan R. Liss Inc., New York.
3. Brown C. H., Munro G., Murphy N. P., Leng G., Russell J. A. (1996) Activation of oxytocin neurones by systemiccholecystokinin is unchanged by morphine-dependence or withdrawal excitation in the rat. J. Physiol., Lond. 496,787–794.
4. Cunningham E. T. Jr., Sawchenko P. E. (1988) Anatomical specificity of noradrenergic inputs to the paraventricularand supraoptic nuclei of the rat hypothalamus. J. comp. Neurol. 274, 60–76.
5. Day T. A., Sibbald J. R. (1988) Direct catecholaminergic projection from nucleus tractus solitarii to supraopticnucleus. Brain Res. 454, 387–392.
6. Done C., Silverstone P., Sharp T. (1992) Effect of naloxone-precipitated morphine withdrawal on noradrenalinerelease in rat hippocampus in vivo. Eur. J. Pharmac. 215, 333–336.
7. Garten L. L., Sofroniew M. V., Dyball R. E. (1989) A direct catecholaminergic projection from the brainstem to theneurohypophysis of the rat. Neuroscience 33, 149–155.
8. Gold M. S., Redmond D. E. Jr., Kleber H. D. (1978) Clonidine blocks acute opiate-withdrawal symptoms. Lancet 2,599–602.
9. Gonzalvez M. L., Milanes M. V., Martinez-Pinero M. G., Marin M. T., Vargas M. L. (1994) Effects ofintracerebroventricular clonidine on the hypothalamic noradrenaline and plasma corticosterone levels of opiate naiverats and after naloxone-induced withdrawal. Brain Res. 647, 199–203.
10. Hamamura M., Leng G., Emson P. C., Kiyama H. (1991) Electrical activation and c-fos mRNA expression in ratneurosecretory neurones after systemic administration of cholecystokinin. J. Physiol., Lond. 444, 51–63.
11. Higuchi T., Honda K., Fukuoka T., Negoro H., Wakabayashi K. (1985) Release of oxytocin during suckling andparturition in the rat. J. Endocrinol. 105, 339–346.
12. Kendrick K., Leng G., Higuchi T. (1991) Noradrenaline, dopamine and serotonin release in the paraventricular andsupraoptic nuclei of the rat in response to intravenous cholecystokinin injection. J. Neuroendocrinol. 3, 139–144.
13. Leng G. (1981) The effects of neural stalk stimulation upon firing patterns in rat supraoptic neurones. Expl Brain Res.41, 135–145.
14. Leng G., Mansfield S., Bicknell R. J., Blackburn R. E., Brown D., Chapman C., Dyer R. G., Hollingsworth S.,Shibuki K., Yates J. O. (1988) Endogenous opioid actions and effects of environmental disturbance on parturition andoxytocin secretion in rats. J. Reprod. Fert. 84, 345–356.
15. Mansour A., Khachaturian H., Lewis M. E., Akil H., Watson S. J. (1988) Anatomy of CNS opioid receptors. TrendsNeurosci. 11, 308–314.
16. Martinez-Pinero M. G., Milanes M. V., Vargas M. L. (1994) Modulation by catecholamine of hypothalamus-pituitary-adrenocortical (HPA) axis activity in morphine-tolerance and withdrawal. Gen. Pharmac. 25, 187–192.
17. Murphy N. P., Brown C. H., Leng G., Russell J. A. (1995) Morphine-withdrawal excitation of oxytocin neurones inthe absence of their noradrenergic input. Analgesia 1, 594–597.
576 N. P. Murphy et al.

18. North R. A., Williams J. T. (1985) On the potassium conductance increased by opioids in rat locus coeruleus neurones.J. Physiol., Lond. 364, 265–280.
19. Onaka T., Luckman S. M., Antonijevic I., Palmer J. R., Leng G. (1995) Involvement of the noradrenergic afferentsfrom the nucleus tractus solitarii to the supraoptic nucleus in oxytocin release after peripheral cholecystokininoctapeptide in the rat. Neuroscience 66, 403–412.
20. Onaka T., Luckman S. M., Guevara-Guzman R., Ueta Y., Kendrick K., Leng G. (1995) Presynaptic actions ofmorphine: blockade of cholecystokinin-induced noradrenaline release in the rat supraoptic nucleus. J. Physiol., Lond.482, 69–79.
21. Pumford K. M., Leng G., Russell J. A. (1991) Morphine actions on supraoptic oxytocin neurones in anaesthetizedrats: tolerance after i.c.v. morphine infusion. J. Physiol., Lond. 440, 437–454.
22. Raby W. N., Renaud L. P. (1989) Dorsomedial medulla stimulation activates rat supraoptic oxytocin and vasopressinneurones through different pathways. J. Physiol., Lond. 417, 279–294.
23. Rayner V. C., Robinson I. A. C. F., Russell J. A. (1988) Chronic intracerebroventricular morphine and lactation inrats: dependence and tolerance in relation to oxytocin neurones. J. Physiol., Lond. 396, 319–347.
24. Russell J. A., Leng G., Bicknell R. J. (1995) Opioid tolerance and dependence in the magnocellular oxytocin system:a physiological mechanism?. Expl Physiol. 80, 307–340.
25. Russell J. A., Neumann I., Landgraf R. (1992) Oxytocin and vasopressin release in discrete brain areas after naloxonein morphine-tolerant and -dependent anesthetized rats: push-pull perfusion study. J. Neurosci. 12, 1024–1032.
26. Russell J. A., Pumford K. M., Bicknell R. J. (1992) Contribution of the region anterior and ventral to the thirdventricle to opiate withdrawal excitation of oxytocin secretion. Neuroendocrinology 55, 183–192.
27. Shu S. Y., Ju G., Fan L. Z. (1988) The glucose oxidase-DAB-nickel method in peroxidase histochemistry of thenervous system. Neurosci. Lett. 85, 169–171.
28. Silverstone P. H., Done C., Sharp T. (1992) Clonidine but not nifedipine prevents the release of noradrenalineduring naloxone-precipitated opiate withdrawal: an in vivo microdialysis study in the rat. Psychopharmacology 109,235–238.
29. Stornetta R. L., Norton F. E., Guyenet P. G. (1993) Autonomic areas of rat brain exhibit increased Fos-likeimmunoreactivity during opiate withdrawal in rats. Brain Res. 624, 19–28.
30. Sumner B. E., Coombes J. E., Pumford K. M., Russell J. A. (1990) Opioid receptor subtypes in the supraoptic nucleusand posterior pituitary gland of morphine-tolerant rats. Neuroscience 37, 635–645.
31. Taylor J. R., Elsworth J. D., Garcia E. J., Grant S. J., Roth R. H., Redmond D. E. Jr. (1988) Clonidine infusions intothe locus coeruleus attenuate behavioral and neurochemical changes associated with naloxone-precipitated with-drawal. Psychopharmacology 96, 121–134.
(Accepted 19 March 1997)
Oxytocin afferent activity in morphine withdrawal 577




















