


Automatic Population HARDI White Matter Tract Clustering by Label Fusion of Multiple Tract Atlases

B R A I N R E S E A R C H R E V I E W S 6 5 ( 2 0 1 1 ) 1 5 0 – 1 8 3
ava i l ab l e a t www.sc i enced i r ec t . com
www.e l sev i e r . com/ loca te /b ra in res rev
Review
The Retinohypothalamic tract: Comparison of axonalprojection patterns from four major targets
Newton S. Canterasa, Érika Renata Ribeiro-Barbosaa,Marina Gotoa, José Cipolla-Netob, Larry W. Swansonc,⁎aDepartment of Anatomy, Institute of Biomedical Sciences, University of São Paulo, São Paulo, 05508-000, BrazilbDepartment of Physiology and Biophysics, Institute of Biomedical Sciences, University of São Paulo, São Paulo, 05508-900, BrazilcDepartment of Biological Sciences, University of Southern California, Los Angeles, CA 90089-2520, USA
A R T I C L E I N F O
⁎ Corresponding author.E-mail address: [email protected] (L.W.Abbreviations: AAA, anterior amygdalar ar
AHA, anterior hypothalamic area; AHNa, anteAHNd, anterior hypothalamic nucleus, dorsanucleus, ventromedial part; APN, anterioranteroventral preoptic nucleus; AVPV, anterolfactory tract; BLAa, basolateral amygdalar namygdalar nucleus, anterior part; BMAp, basodivision, anterodorsal area; BSTal, bed nucleterminalis, anterior division, anteroventral arBSTdl, bed nuclei of the stria terminalis, anterdorsomedial nucleus; BSTfu, bed nuclei of theposterior division, interfascicular nucleus; BSnuclei of the stria terminalis, anterior divisionucleus; BSTpr, bed nuclei of the stria termindivision, rhomboid nucleus; BSTsc, bed nuclestria terminalis, posterior division, transverseCA1, Ammon's horn; CA3, field CA3, Ammoncapsular part; CEAl, central amygdalar nuclethalamus; CLI, central linear nucleus raphé;cortical amygdalar area, posterior part, lateracerebral peduncle; CSl, superior central nuclnucleus; da, dopamine cell group within zo
0165-0173/$ – see front matter © 2010 Elsevidoi:10.1016/j.brainresrev.2010.09.006
A B S T R A C T
Article history:Accepted 16 September 2010Available online 21 September 2010
The retinohypothalamic tract is one component of the optic nerve that transmitsinformation about environmental luminance levels through medial and lateral branchesto four major terminal fields in the hypothalamus. The spatial distribution and organizationof axonal projections from each of these four terminal fields were analyzed and comparedsystematically with the anterograde pathway tracer PHAL in rats where the terminal fieldshad been labeled with intravitreal injections of a different anterograde pathway tracer, CTb.First, the well-known projections of two medial retinohypothalamic tract targets (theventrolateral suprachiasmatic nucleus and perisuprachiasmatic region) were confirmedand extended. They share qualitatively similar projections to a well-known set of brainregions thought to control circadian rhythms. Second, the projections of a third medial tract
Keywords:Anterior hypothalamic nucleusLateral hypothalamic areaSubparaventricular zoneSuprachiasmatic nucleus
Swanson).ea; ACB, nucleus accumbens; aco, anterior commissure; ADP, anterodorsal preoptic nucleus;rior hypothalamic nucleus, anterior part; AHNc, anterior hypothalamic nucleus, central part;l part; AHNp, anterior hypothalamic nucleus, posterior part; AHNvm, anterior hypothalamicpretectal nucleus; AQ , cerebral aqueduct; ARH, arcuate hypothalamic nucleus; AVP,
oventral periventricular nucleus; B, Barrington's nucleus; BA, Bed nucleus of the accessoryucleus, anterior part; BLAp, basolateral amygdalar nucleus, posterior part; BMAa, basomedialmedial amygdalar nucleus, posterior part; BSTad, bed nuclei of the stria terminalis, anteriori of the stria terminalis, anterior division, anterolateral area; BSTav, bed nuclei of the striaea; BSTcc, bed nuclei of the stria terminalis, anterior division, anterodorsal area, central core;ior division, dorsolateral nucleus; BSTdm, bed nuclei of the stria terminalis, anterior division,stria terminalis, anterior division, fusiform nucleus; BSTif, bed nuclei of the stria terminalis,Tju, bed nuclei of the stria terminalis, anterior division, juxtacapsular nucleus; BSTmg, bedn, magnocellular nucleus; BSTov, bed nuclei of the stria terminalis, anterior division, ovalalis, posterior division, principal nucleus; BSTrh, bed nuclei of the stria terminalis, anteriori of the stria terminalis, anterior division, subcommissural nucleus; BSTtr, bed nuclei of thenucleus; BSTv, bed nuclei of the stria terminalis, anterior division, ventral nucleus; CA1, field's horn; cc, corpus callosum; ccg, genu of corpus callosum; CEAc, central amygdalar nucleus,us, lateral part; CEAm, central amygdalar nucleus, medial part; CL, central lateral nucleusCM, central medial nucleus thalamus; COAa, cortical amygdalar area, anterior part; COApl,l zone; COApm, cortical amygdalar area, posterior part, medial zone; CP, caudoputamen; cpd,eus raphé, lateral part; CSm, superior central nucleus raphé, medial part; CUN, cuneiformna incerta; DGlb, dentate gyrus lateral blade; DMHa, dorsomedial hypothalamic nucleus,
er B.V. All rights reserved.

anterior part; DMHp, dorsomedial hypothalamic nucleus, posterior part; DMHv, dorsomedial hypothalamic nucleus, ventral part; DR,dorsal nucleus raphé; DTN, dorsal tegmental nucleus; fr, fasciculus retroflexus; FS, fundus of the striatum; fx, columns of the fornix; GPl,globus pallidus, lateral segment; GPm, globus pallidus, medial segment; I, internuclear area, hypothalamic periventricular region; IA,intercalated nuclei amygdala; IAD, interanterodorsal nucleus thalamus; IAM, interanteromedial nucleus thalamus; IGL, lateral geniculatecomplex, intergeniculate leaflet; III, oculomotor nucleus; IMD, intermediodorsal nucleus thalamus; int, internal capsule; IPN,interpeduncular nucleus; KF, Kolliker–Fuse subnucleus; LC, locus ceruleus; LDT, laterodorsal tegmental nucleus; LGd, lateral geniculatecomplex, dorsal part; LGvl, lateral geniculate complex, ventral part, lateral zone; LGvm, lateral geniculate complex, ventral part, medialzone; LH, lateral habenula; LHA, lateral hypothalamic area; LHAad, lateral hypothalamic area, anterior region, dorsal zone; LHAai, lateralhypothalamic area, anterior region, intermediate zone; LHAav, lateral hypothalamic area, anterior region, ventral zone; LHAavr, lateralhypothalamic area, anterior region, ventral zone, retinorecipient part; LHAd, lateral hypothalamic area, dorsal region; LHAjd, lateralhypothalamic area, juxtadorsomedial region; LHAjp, lateral hypothalamic area, juxtaparaventricular region; LHAjvd, lateral hypothalamicarea, juxtaventromedial region, dorsal zone; LHAjvv, lateral hypothalamic area, juxtaventromedial region, ventral zone; LHAm, lateralhypothalamic area, magnocellular nucleus; LHAs, lateral hypothalamic area, suprafornical region; LHAsfp, lateral hypothalamic area,subfornical region, posterior zone; LM, lateral mammillary nucleus; LP, lateral posterior nucleus thalamus; LPO, lateral preoptic area; LSc,lateral septal nucleus, caudal (caudodorsal) part; LSc.d, lateral septal nucleus, caudal (caudodorsal) part, dorsal zone; LSc.d.r, lateral septalnucleus, caudal (caudodorsal) part, dorsal zone, rostral region; LSc.v.i, lateral septal nucleus, caudal (caudodorsal) part, ventral zone,intermediate region; LSc.v.l, lateral septal nucleus, caudal (caudodorsal) part, ventral zone, lateral region; LSc.v.m, lateral septal nucleus,caudal (caudodorsal) part, ventral zone, medial region; LSr.dl.l, lateral septal nucleus, rostral (rostroventral) part, dorsolateral zone, lateralregion; LSr.dl.l.d, lateral septal nucleus, rostral (rostroventral) part, dorsolateral zone, lateral region, dorsal domain; LSr.dl.m, lateral septalnucleus, rostral (rostroventral) part, dorsolateral zone, medial region; LSr.dl.m.d, lateral septal nucleus, rostral (rostroventral) part,dorsolateral zone, medial region, dorsal domain; LSr.m, lateral septal nucleus, rostral (rostroventral) part, medial zone; LSr.m.d, lateralseptal nucleus, rostral (rostroventral) part, medial zone, dorsal region; LSr.m.v.c, lateral septal nucleus, rostral (rostroventral) part, medialzone, ventral region, caudal domain; LSr.m.v.r, lateral septal nucleus, rostral (rostroventral) part, medial zone, ventral region, rostraldomain; LSr.vl.d.l, lateral septal nucleus, rostral (rostroventral) part, ventrolateral zone, dorsal region, lateral domain; LSr.vl.d.m, lateralseptal nucleus, rostral (rostroventral) part, ventrolateral zone, dorsal region, medial domain; LSr.vl.v, lateral septal nucleus, rostral(rostroventral) part, ventrolateral zone, ventral region; LSv, lateral septal nucleus, ventral part; MDc, mediodorsal nucleus thalamus,central part; MDl, mediodorsal nucleus thalamus, lateral part; MDm, mediodorsal nucleus thalamus, medial part; ME, median eminence;MEAad, medial amygdalar nucleus, anterodorsal part; MEAav, medial amygdalar nucleus, anteroventral part; MEApd, medial amygdalarnucleus, posterodorsal part; MEApv, medial amygdalar nucleus, posteroventral part; MEex, median eminence external; MEin, medianeminence internal; MEPO, median preoptic nucleus; MEV, mesencephalic nucleus of the trigeminal; MGd, medial geniculate complex,dorsal part; MGv, medial geniculate complex, ventral part; MH, medial habenula; ml, medial lemniscus; mlf, medial longitudinal fascicle;MM,medial mammillary nucleus;mp,mammillary peduncle; MPNc,medial preoptic nucleus, central part; MPNl, medial preoptic nucleus,lateral part; MPNm, medial preoptic nucleus, medial part; MPO, media preoptic area; MPT, medial pretectal area; MRN, midbrain reticularnucleus; MS, medial septal nucleus; mtt, mammillothalamic tract; ND, nucleus of Darkschewitsch; NDB, diagonal band nucleus; NIc,nucleus incertus, compact part; NId, nucleus incertus, diffuse part; NOT, nucleus of the optic tract; NPC, nucleus of the posteriorcommissure; och, optic chiasm; OP, olivary pretectal nucleus; OT, olfactory tubercle; PA, posterior amygdalar nucleus; PAG, periaqueductalgray; PAGdl, periaqueductal gray, dorsolateral division; PAGdm, periaqueductal gray, dorsomedial division; PAGl, periaqueductal gray,lateral division; PAGvl, periaqueductal gray, ventrolateral division; PBlc, parabrachial nucleus, lateral division, central lateral part; PBld,parabrachial nucleus, lateral division, dorsal lateral part; PBle, parabrachial nucleus, lateral division, external lateral part; PBls,parabrachial nucleus, lateral division, superior lateral part; PBlv, parabrachial nucleus, lateral division, ventral part; PBme, parabrachialnucleus, medial division, external medial part; PBmm, parabrachial nucleus, medial division, medial part; PBmv, parabrachial nucleus,medial division, ventral part; pc, posterior commissure; PCG, pontine central gray; PCN, paracentral nucleus thalamus; PF, parafascicularnucleus; PH, posterior hypothalamic nucleus; pm, principal mammillary tract; PMd, dorsal premammillary nucleus; PMv, ventralpremammillary nucleus; PPN, pedunculopontine nucleus; PR, perireuniens nucleus; PRC, periaqueductal gray, precommissural nucleus;PRNr, pontine reticular nucleus, rostral part; PSCH, suprachiasmatic preoptic nucleus; PT, paratenial nucleus; PVa, periventricularhypothalamic nucleus, anterior part; PVHam, paraventricular nucleus hypothalamus,magnocellular division, anteriormagnocellular part;PVHa, paraventricular nucleus hypothalamus, anterior parvicellular part; PVHdp, paraventricular nucleus hypothalamus, dorsalparvicellular part; PVHf, paraventricular nucleus hypothalamus, forniceal part; PVHlp, paraventricular nucleus hypothalamus, lateralparvicellular part; PVHmpd, paraventricular nucleus hypothalamus, medial parvicellular part, dorsal zone; PVHmpv, paraventricularnucleus hypothalamus, medial parvicellular part, ventral zone; PVHpml, paraventricular nucleus hypothalamus, posterior magnocellularpart, lateral zone; PVHpmm, paraventricular nucleus hypothalamus, posterior magnocellular part, medial zone; PVHpv, paraventricularnucleus hypothalamus, periventricular part; PVi, periventricular nucleus hypothalamus, intermediate part; PVp, periventricular nucleushypothalamus, posterior part; PVpo, preoptic periventricular nucleus; PVT, paraventricularnucleus thalamus; RCH, retrochiasmatic area;REa, nucleus reuniens, rostral division, anterior part; REcd, nucleus reuniens, caudal division, dorsal part; REcm, nucleus reuniens, caudaldivision, median part; REcp, nucleus reuniens, caudal division, posterior part; REd, nucleus reuniens, rostral division, dorsal part; REl,nucleus reuniens, rostral division, lateral part; REm, nucleus reuniens, rostral division,median part; REv, nucleus reuniens, rostral division,ventral part; RH, rhomboidnucleus; RN, red nucleus; RR, midbrain reticular nucleus, retrorubral area; SBPV, subparaventricular zone;SBPVps, subparaventricular zone, perisuprachiasmatic region; SCdg, superior colliculus, deep gray layer; SCig, superior colliculus,intermediate gray layer; SCsg, superior colliculus, superficial gray; SCH, suprachiasmatic nucleus; scp, superior cerebellar peduncle; SF,septofimbral nucleus; SFO, subfornical organ; SH, septohippocampal nucleus; SI, substantia innominata; SLD, sublaterodorsal nucleus; sm,stria medullaris; smd, supramammillary decussation; SMT, submedial nucleus of the thalamus; SNc, substantia nigra, compact part; SNr,substantia nigra, reticular part; SO, supraoptic nucleus; SPFm, subparafascicular nucleus thalamus,magnocellular part; st, stria terminalis;SUBv, subiculum, ventral part; SUM, supramammillary nucleus; SUMl, supramammillary nucleus, lateral part; SUMm, supramammillarynucleus, medial part; SUT, supratrigeminal nucleus; TM, tuberomammillary nucleus; TMv, tuberomammillary nucleus ventral part; TRS,triangular nucleus septum; TU, tuberal nucleus; TUl, tuberal nucleus, lateral part; TUsv, tuberal nucleus, subventromedial part; V, motornucleus of the trigeminal nerve; V3, third ventricle; V4, fourth ventricle; VL, lateral ventricle; VMHa, ventromedialnucleus hypothalamus,anterior part; VMHc, ventromedial nucleus hypothalamus, central part; VMHdm, ventromedial nucleus hypothalamus, dorsomedial part;VMHvl, ventromedial nucleus hypothalamus, ventrolateral part; VTA, ventral tegmental area; VTN, ventral tegmental nucleus; ZI, zonaincerta
151B R A I N R E S E A R C H R E V I E W S 6 5 ( 2 0 1 1 ) 1 5 0 – 1 8 3

152 B R A I N R E S E A R C H R E V I E W S 6 5 ( 2 0 1 1 ) 1 5 0 – 1 8 3
target, the ventromedial part of the anterior hypothalamic nucleus, were analyzed for thefirst time and shown to resemble qualitatively those from the suprachiasmatic nucleus andperisuprachiasmatic region. And third, projections from the major lateralretinohypothalamic tract target were analyzed for the first time and shown to be quitedifferent from those associated withmedial tract targets. This target is a distinct core part ofthe ventral zone of the anterior group of the lateral hypothalamic area that lies just dorsal tothe caudal two-thirds of the supraoptic nucleus. Its axonal projections are to neuralnetworks that control a range of specific goal-oriented behaviors (especially drinking,reproductive, and defensive) alongwith adaptively appropriate and complementary visceralresponses and adjustments to behavioral state.
© 2010 Elsevier B.V. All rights reserved.
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1522. Materials and methods . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 153
2.1. Animals . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1532.2. Combined PHAL and intravitreal CTb deposits . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 153
3. Results . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1543.1. Organization of the retinohypothalamic tract . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1543.2. Projections from the SCH: first rhtm target . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 154
3.2.1. Ascending pathway . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1563.2.2. Dorsal pathway . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1593.2.3. Dorsocaudal pathway . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1593.2.4. Lateral pathway . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1623.2.5. Crossed pathway . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 162
3.3. Projections from the perisuprachiasmatic region: second rhtm target . . . . . . . . . . . . . . . . . . . . . . 1623.4. Projections from the AHNvm: third rhtm target . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 165
3.4.1. Ascending pathway . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1653.4.2. Descending pathway . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 166
3.5. Projections from the LHAavr: main rhtl target . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1663.5.1. Ascending pathway . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1673.5.2. Descending pathway . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 169
3.6. Control injections . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1734. Discussion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 173
4.1. The medial retinohypothalamic tract (rhtm) and circadian rhythm control . . . . . . . . . . . . . . . . . . . 1734.2. Circuitry underlying circadian rhythms . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1764.3. Functional roles of the LHAavr–the main target of the rhtl . . . . . . . . . . . . . . . . . . . . . . . . . . . . 177
4.3.1. Goal oriented-behavior control . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1774.3.2. Behavioral state control . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1794.3.3. Viscera-related control . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1794.3.4. Thalamic loops . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 179
4.4. Overview . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 179Acknowledgments . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 179References. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 180
1. Introduction
A direct projection from the retina to the hypothalamus wasfirst demonstrated unequivocally with the autoradiographicanterograde pathway tracing method almost 40 years ago(Moore and Lenn, 1972; Hendrickson et al., 1972). In theseexperiments, an intravitreal injection of tritiated amino acidswas made in one eye and axonally transported tritiatedproteins were detected in a terminal field highly restricted to
ventrolateral regions of the suprachiasmatic nucleus (SCH) onboth sides of the hypothalamus. This discovery was immedi-ately followed by experimental evidence that selective de-struction of the SCH abolishes a wide range of behavioral andneuroendocrine circadian and diurnal rhythms (see Mooreand Eichler, 1972; Stephan and Zucker, 1972; Rusak andZucker, 1979). These results stimulated at least two majorlines of investigation: the nature of mechanisms within theSCH itself that are involved in the photic entrainment and

153B R A I N R E S E A R C H R E V I E W S 6 5 ( 2 0 1 1 ) 1 5 0 – 1 8 3
generation of circadian rhythms, andmechanisms underlyingthe transfer of this SCH processing to other parts of the brainfor the expression of specific circadian and diurnal rhythms.The latter problem was first addressed with the autoradio-graphic method in a study where an injection of tritiatedamino acids was placed in the ventrolateral region of the ratSCH and labeled projections were traced dorsally and caudallythrough periventricular regions of the hypothalamus in a pat-tern that suggested innervation of at least the paraventricular,dorsomedial, ventromedial, and arcuate nuclei (Swanson andCowan, 1975).
Since then considerably more sensitive axonal pathwaytracing methods have been introduced. On one hand, theretinohypothalamic tract (rht) has been shown to be muchmore extensive than initially thought, especiallywith the tracercholera toxin subunit b (CTb). In fact, it is now known that theretinohypothalamic tract has two major branches, medial(rhtm) and lateral (rhtl), and that together they innervate fourmajor terminal fields in the hypothalamus (Johnson et al., 1988;Levine et al., 1991; Costa et al., 1999). Specifically, the rhtmgenerates clear terminal fields in the ventrolateral SCH, theperisuprachiasmatic region immediatelydorsal to the SCH itself(SBPVps), and the ventromedial part of the anterior hypotha-lamic nucleus (AHNvm), whereas the rhtl generates a substan-tial terminal field centered in a retinorecipient part of theventral zone of the anterior group of the lateral hypothalamicarea (LHAavr). And on the other hand the extent of axonalprojections from the SCHhas also been shown to bemuchmoreextensive than originally thought, especially with the antero-grade tracer PHAL (Watts et al., 1987;Watts and Swanson, 1987;Buijs et al., 1993; Kalsbeek et al., 1993; Morin et al., 1994;Abrahamson andMoore, 2001; Leak andMoore, 2001; Kriegsfeldet al., 2004). Thus, for example, the densest terminal field is inthe subparaventricular zone, including its ventral end theSBPVps, and there are several important extrahypothalamicterminal fields as well, including the paraventricular thalamicnucleus and the lateral septal nucleus.
Thepresent series of experimentswas designed to comparedirectly and systematically the organization of projectionsfrom the fourmain terminal fields of the rht in the rat using theanterograde pathway tracer PHAL. To control for appropriateinjection site targeting, the rht and its terminal fieldswere alsolabeled in all experimental animals by intravitreal injections ofa different anterograde pathway tracer, cholera toxin subunit b(CTb). To confirm and extend extensive earlier work and toprovide a direct comparison with analyses of other pathwaysusing the same methods, the results of PHAL injections in theSCH are described first, followed by the results of injections inother terminal fields of the rht'smedial (rhtm) and lateral (rhtl)components. These results are meant to provide a frameworkfor more detailed structural analysis and confirmation withretrograde tracer, histochemical, and ultrastructural methods.
2. Materials and methods
2.1. Animals
Adult male Wistar rats (n=31), each weighing approximately250 g and obtained from local breeding facilities, were used in
the present study. The animals were kept under controlledtemperature (23 °C) and illumination (12-h cycle) in the animalquarters and had free access to water and standard laboratorydiet (Purina, Ribeirão Preto, SP, Brazil). Conditions of animalhousing and all experimental procedures were conductedunder institutional guidelines and in accordance with NIHguidelines on animal care.
2.2. Combined PHAL and intravitreal CTb deposits
Animals were anesthetized with a mixture of ketamine andxylazine (volume/volume; 1 ml/kg body weight), and a singleiontophoretic injection of a 2.5% solution of PHAL (VectorLaboratories) in 0.1 M sodium phosphate-buffered saline, pH7.4 (Gerfen and Sawchenko, 1984) was placed in the supra-chiasmatic nucleus (SCH; n=6), the ventromedial part of theanterior hypothalamic nucleus (AHNvm; n=10), or the retinor-ecipient region of the ventral zone of the rostral region of theLHA (LHAavr; n=15). The PHAL deposits were made over a 15-min period through a stereotaxically positioned glass micro-pipette (tip diameter, 10 μm) by applying a +10 μA currentpulsed at 7-s intervals that was provided by a constant-currentsource (model CS3; Midgard Electronics). To label the retino-hypothalamic tract, 1 week after PHAL injection, the animalsreceived into the posterior chamber of the contralateral eye aninjection of 20 μl of a 2% solution of CTb (List BiologicalLaboratories, Inc.) in 2% dimethyl-sulfoxide dissolved inphysiological saline. Survival times after the PHAL depositsranged from 14 to 16 days.
The animals were perfused, and the brains were processedaccording to procedures described in detail elsewhere (Can-teras et al., 1995). Briefly, each animalwas deeply anesthetizedwith sodium pentobarbital (40 mg/kg, i.p.) and perfusedtranscardially with a solution of 4.0% paraformaldehyde in0.1 M sodium borate buffer, pH 9.5; the brains were removedimmediately and postfixed overnight in the same fixativecontaining 20% sucrose. The brains were then frozen and cuton a sliding microtome.
One series of sections was processed for immunohisto-chemistry with an antiserum directed against PHAL (DakoLaboratories, Carpenteria, CA) at a dilution of 1:1,000.Theantigen–antibody complex was localized by using a variationof the avidin–biotin complex system (ABC), with a commer-cially available kit (ABC Elite Kit, Vector Laboratories). Thesections were mounted on gelatin-coated slides and treatedwith osmium tetroxide to enhance the visibility of the reactionproduct. Slides were then dehydrated and cover-slipped withDPX.
Another series of sections was stained to visualize CTbanterogradely labeled fibers of the retinohypothalamic tractwith an anti-CTb serum raised in goat (dilution 1:5,000; ListBiological Laboratories, Inc.). The primary antiserum was lo-calized by using a variation of the avidin–biotin complex (ABC)system. In brief, sections were incubated for 90 min at roomtemperature in a solution of biotinylated donkey anti-goatimmunoglobulin (IgG; Jackson Immunoresearch Laboratories,West Groove, PA) and then placed in the mixed avidin–biotin–horseradish peroxidase (HRP) complex solution (ABC Elite Kit,Vector Laboratories, Burlingame, CA) for the same period oftime. The peroxidase complex was visualized by exposure for

154 B R A I N R E S E A R C H R E V I E W S 6 5 ( 2 0 1 1 ) 1 5 0 – 1 8 3
10 min to a chromogen solution containing 0.02% 3,3'-diami-nobenzidine tetrahydrochloride (DAB) with 0.3% nickel-am-monium sulfate in 0.05 M Tris-buffer, pH 7.6, followed byincubation for 10 min in chromogen solution with hydrogenperoxide (1:3,000) to produce a blue-black product. Thereaction was terminated by extensive washing in potassium/phosphate-buffered saline. Sectionsweremounted on gelatin-coated slides, and thendehydrated and coverslippedwithDPX.
An adjacent series was always stained with thionin to serveas a cytoarchitectonic reference. Sectionswere examinedunderthemicroscope with brightfield and darkfield illumination. ThePHAL-labeled cells in the injection sites, as well as thedistribution of anterogradely labeled fibers, were plotted withthe aid of a camera lucida onto maps prepared from adjacentthionin-stained sections. The figures were prepared for publi-cation by using Adobe Photoshop (version 4.0.1; Adobe Systems,Mountain View, CA) for the photomicrographs, and AdobeIllustrator (version 11.0.1; Adobe Systems) for the line drawings.The original photomicrographs had their contrast and bright-ness adjusted, in addition to background and dust spotsremoved by using the Adobe Photoshop software. Unlessotherwise indicated, parcellation of the rat brain follows thatof Swanson (2004).
3. Results
3.1. Organization of the retinohypothalamic tract
The basic strategy was to inject the anterograde pathwaytracer PHAL into rht terminal fields labeled in the sameanimals with intravitreal CTb injections. The results of thelatter confirm and extend previous similar analyses (seeJohnson et al., 1988; Levine et al., 1991; Costa et al., 1999) andare summarized from our own material as a foundation fordescribing the PHAL experiments. The rht is divided in twocomponents, i.e., the rhtmand the rhtl. The threemostobviousterminal fields of the rhtm include the SCH, SBPVps, andANHvm; whereas in contrast the rhtl projects clearly to thelateral preoptic area and especially the LHAavr. Axons formingthe rhtm emerge directly from the optic chiasm, while the rhtlhas two sources—one group of axons coursing laterally fromthe rhtm, and the other ascending from the optic tract. The rhtlis generated by a group of retinal ganglion cells that is distinctfrom the group that projects to the SCH and adjacent areas(Moore et al., 1995; Leak andMoore, 1997), and develops earlierduring embryogenesis (Speh and Moore, 1993).
The rhtm provides a strikingly dense bilateral input to theSCH with a clear contralateral prevalence (Fig. 1A, B). Theretinal projection targets the ventrolateral part of the SCHcharacterized by small neurons that are less densely packedthan the very small neurons of the dorsomedial part, whichretinal terminals tend to avoid (see Hendrickson et al., 1972;Moore and Lenn, 1972; Johnson et al., 1988; Murakami et al.,1989; Levine et al., 1991; Costa et al., 1999; Abrahamson andMoore, 2001; Morin et al., 2006). The ventrolateral anddorsomedial parts of the SCH correspond, respectively, tothe “core” and “shell” SCH regions as definedmore recently byMoore (for review see Moore et al., 2002). As discussed later,these subdivisions can be characterized by differences in
chemoarchitecture and clock genes expression, as well as bydifferences in afferent and efferent projections, and this re-lationship remains relatively stable among mammals (Leakand Moore, 2001; Moore et al., 2002).
From the SCH, rhtm fibers extend dorsally to innervatesubstantially the small SBPVps (Fig. 1A, B). Finally, theremaining rhtm fibers turn caudally and laterally to generatea dense terminal plexus in the anterior hypothalamic nucleusand a relativelymodest input to the subjacent retrochiasmaticarea (Fig. 1C). The retinal projection to the anterior hypotha-lamic nucleus is mostly contralateral and concentrated in theAHNvm, which presents characteristic strings of moderatelypacked, medium-sized neurons that are easily identified inNissl-stained sections (Fig. 1C'). The input to the AHNvm hasbeen shown in various species (Conrad and Stumpf, 1975;Johnson et al., 1988; Murakami et al., 1989; Levine et al., 1991;Costa et al., 1999) and lies ventromedially in what has beencalled the central part of the AHN (AHNc; see Risold et al., 1994;Swanson, 2004). Based on the present results, the old AHNchas been shrunken to make way for the newly characterizedretinorecipient AHNvm. A small contingent of axons from therhtm may proceed caudally and provide very sparse inputsto the posterior part of the AHN, the dorsal part of thesubparaventricular zone close to the paraventricular nucleus(SBPVd), the anterior part of the dorsomedial nucleus, and theinternuclear area between the dorsomedial and ventromedialnuclei. These seemingly inconsequential distributions havenot been reported before.
The rhtl is heavily labeled on the side contralateral to theCTb-injected eye, with only a few labeled fibers observed onthe ipsilateral side. At preoptic levels, the rhtl provides arelatively sparse input to the lateral preoptic area, in a regionnear the border with the anteroventral preoptic nucleus,which also presents occasional labeling. This projection hasbeen previously observed in the rat (Levine et al., 1991) and themarmoset (Costa et al., 1999). At anterior hypothalamic levels,the rhtl supplies a dense projection to the lateral hypotha-lamic area, centered in a region surrounding the border of theoptic tract and the supraoptic nucleus (Fig. 1A–C). Thisterminal field extends rostrocaudally along the caudal two-thirds of the supraoptic nucleus in what has recently beenidentified as the ventral zone of the rostral region of the LHA(LHAav; Swanson, 2004; Swanson et al., 2005), characterized asrelatively cell-sparse in Nissl-stained sections (Fig. 1A'–C').This target of the rhtl will be referred to as the retinorecipientregion of the LHAav (LHAavr). The retinal projection to theLHA has been previously described in various species (Mai,1979; Johnson et al., 1988; Levine et al., 1991; Costa et al., 1999).Only a few retinal fibers were labeled within the supraopticnucleus itself, as reported before (Mai, 1979; Johnson et al.,1988; Levine et al., 1991). Proceeding caudally, at tuberal levels,fibers from the rhtl provide sparse inputs to the subfornicaland juxtaventromedial regions of the LHA, which have notbeen reported before.
3.2. Projections from the SCH: first rhtm target
In two experiments PHAL-labeled cells are confined al-most entirely to the SCH and the injection sites present acomplementary pattern (Fig. 3). In Experiment SCH#13 the
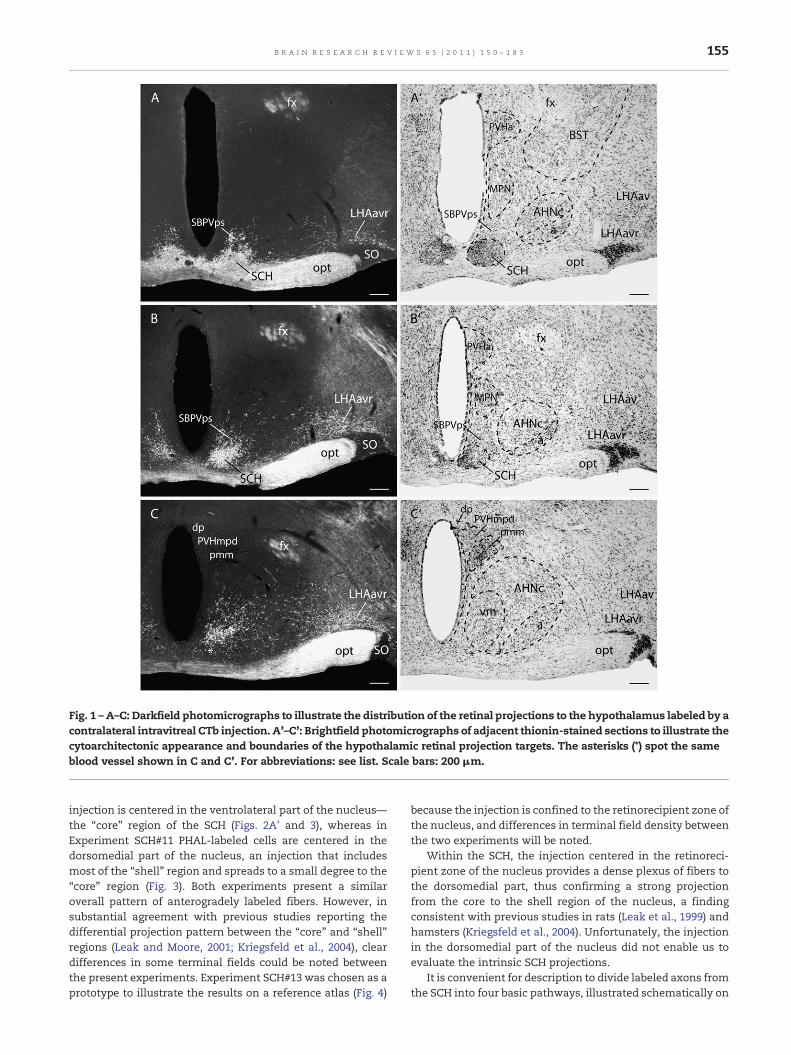
Fig. 1 –A–C: Darkfield photomicrographs to illustrate the distribution of the retinal projections to the hypothalamus labeled by acontralateral intravitreal CTb injection. A'–C': Brightfield photomicrographs of adjacent thionin-stained sections to illustrate thecytoarchitectonic appearance and boundaries of the hypothalamic retinal projection targets. The asterisks (*) spot the sameblood vessel shown in C and C'. For abbreviations: see list. Scale bars: 200 μm.
155B R A I N R E S E A R C H R E V I E W S 6 5 ( 2 0 1 1 ) 1 5 0 – 1 8 3
injection is centered in the ventrolateral part of the nucleus—the “core” region of the SCH (Figs. 2A' and 3), whereas inExperiment SCH#11 PHAL-labeled cells are centered in thedorsomedial part of the nucleus, an injection that includesmost of the “shell” region and spreads to a small degree to the“core” region (Fig. 3). Both experiments present a similaroverall pattern of anterogradely labeled fibers. However, insubstantial agreement with previous studies reporting thedifferential projection pattern between the “core” and “shell”regions (Leak and Moore, 2001; Kriegsfeld et al., 2004), cleardifferences in some terminal fields could be noted betweenthe present experiments. Experiment SCH#13 was chosen as aprototype to illustrate the results on a reference atlas (Fig. 4)
because the injection is confined to the retinorecipient zone ofthe nucleus, and differences in terminal field density betweenthe two experiments will be noted.
Within the SCH, the injection centered in the retinoreci-pient zone of the nucleus provides a dense plexus of fibers tothe dorsomedial part, thus confirming a strong projectionfrom the core to the shell region of the nucleus, a findingconsistent with previous studies in rats (Leak et al., 1999) andhamsters (Kriegsfeld et al., 2004). Unfortunately, the injectionin the dorsomedial part of the nucleus did not enable us toevaluate the intrinsic SCH projections.
It is convenient for description to divide labeled axons fromthe SCH into four basic pathways, illustrated schematically on

Fig. 2 – Photomicrographs to illustrate the appearance of PHAL deposits (A'–C'), and respective adjacent thionin-stained sections(A–C) and CTb-stained sections showing retinal projection fields (A''–C'') in experiments with injections centered in the coreregion of the SCH (A''–A''; experiment SCH#13), in the perisuprachiasmatic region of the SBPV (B–B''; experiment AHNr4), and inthe ventromedial part of the AHN (C–C''; experiment AHNr19). For abbreviations: see list. Scale bars: 200 μm.
156 B R A I N R E S E A R C H R E V I E W S 6 5 ( 2 0 1 1 ) 1 5 0 – 1 8 3
a flatmap in Fig. 9A, B. In addition, Table 1 should be consultedfor a comprehensive, semiquantitative listing of terminal fieldstrength for pathways arising from the fourmajor targets (SCHcore, SCH shell, SCPVps, AHNvm) of the retinohypothalamictract described in the rest of the Results.
3.2.1. Ascending pathwayA group of PHAL-labeled axons from the SCH ascends throughthe preoptic region, where a dense plexus is observed in thesuprachiasmatic preoptic nucleus, as are moderate projectionsto the median preoptic and anterodorsal preoptic nuclei, andsparse inputs to the anteroventral periventricular preoptic andmedial preoptic (all parts) nuclei (Figs. 4A–C and 5B). Themedialpreoptic nucleus and theperiventricular preoptic nuclei containdistinctly more labeled fibers in the experiment with theinjection centered in the dorsomedial part of the SCH (Experi-ment RCH#11), where a clear terminal field was particularlyfound in the anteroventral periventricular preoptic andmedianpreoptic nuclei (Fig. 5A), in addition to labeled fibers andterminals in the vascular organ of the lamina terminalis.
These observations are in line with previous studies showingthat the shell region projects more densely to these preoptictargets (Leak andMoore, 2001; Kriegsfeld et al., 2004). Moreover,both experiments with injections centered either in the core orin the shell partof theSCHlabeledonlyavery sparse input to theanteroventral preoptic nucleus (Fig. 4B, C),which corresponds inpart to the ventrolateral preoptic region, reported to receiveinputs from the core region of the SCH (Novak and Nunez, 2000;Chou et al., 2002).
Rostral to the anterior commissure, some fibers extenddorsally, mostly through (and to) the lateral preoptic area, toenter to the septal region, where they end mostly in the rostraland ventral parts of the lateral septal nucleus, with onlyoccasional labeling elsewhere (Fig. 4A–C). A few labeled axons intheseptal regionthenenter thecolumnof the fornixandcontinuethrough the fimbria to the hippocampus, where occasionallabeling is observed in ventral field CA1. In contrast to what wasobserved for the injection centered in the ventrolateral part of theSCH, the injection centered in the dorsomedial part of the SCH(Experiment SCH#11) labeled only occasional fibers in the lateral

Fig. 3 – Camera lucida plots of labeled neurons following injections of PHAL into the SCH in experiments SCH#11 and #13. Thedrawings for each experiment are arranged from rostral to caudal (top to bottom), and the numbers on the top-right refers toBrain Maps (Swanson, 2004) plate numbers, to indicate the approximate rostro-caudal levels.
157B R A I N R E S E A R C H R E V I E W S 6 5 ( 2 0 1 1 ) 1 5 0 – 1 8 3
septal nucleus, suggesting that the core, but not the shell region,projects to the lateral septal nucleus as reported in previousstudies that injected retrograde tracers in the septal region (Leakand Moore, 2001; Kriegsfeld et al., 2004). Our results on theretinorecipient zone of the SCH generally confirm earlier PHAL
analyses by Watts et al. (1987), which also reported PHALinjections mostly circumscribed to the ventral SCH, except thatin the present study a dense rather than sparse input to thesuprachiasmatic preoptic nucleus was observed, and a sparseinput to the ventral hippocampus was identified.

Fig. 4 – Summary of projections from the SCH core. The distribution of PHAL immunoreactive fibers in experiment SCH#13 wasplotted onto a series of standard drawings of the rat brain (Brain Maps—Swanson, 2004) that are arranged from rostral (A) tocaudal (N). The dark gray area in (D) and (E) indicates the injection site, which is illustrated in Figs. 2(A–A'') and 3. Forabbreviations: see list.
159B R A I N R E S E A R C H R E V I E W S 6 5 ( 2 0 1 1 ) 1 5 0 – 1 8 3
3.2.2. Dorsal pathwayAnother fiber group takes a dorsal course from the injection sitethrough the periventricular preoptic nucleus, caudally adjacentanterior part of the periventricular hypothalamic nucleus, andthe anterior parvicellular part of the paraventricular hypotha-lamic nucleus, providing only sparse inputs to these cell groups(Figs. 4D–F and 5D). Note that the projections to theseperiventricular sites were denser in the experiments with aninjection centered in the dorsomedial part of the SCH (Fig. 5C). Afew of these fibers extend laterally to end throughout thejuxtaparaventricular region of the lateral hypothalamic areaand posterior division of the bed nuclei of the stria terminalis(Figs. 4D–E and 5C,D). In addition, a substantial number of fiberscoursing through this pathway continue dorsally to enter thethalamus, where they form a dense terminal field in rostrallevels of the paraventricular nucleus of the thalamus (Figs. 4D–Fand 5C–D), and sparse terminal fields in the paratenial nucleus,
nucleus reuniens, andmore caudal levels of the paraventricularnucleus (Fig. 4D–K).These results generally confirmearlier PHALanalyses in the ratwith the injection centered in theventral partof the SCH (see Watts et al., 1987).
3.2.3. Dorsocaudal pathwayA third labeled fiber group extends dorsocaudally throughanterior and tuberal levels of the hypothalamus, essentially inthe SBPV. On leaving the SCH this pathway forms a denseterminal field in the immediately surrounding, relatively cell-sparse SBPVps (Fig. 4D–E) and that (at least part of which)receives a direct, circumscribed input from the retina in rats(Fig. 1A, B). Notably, the density of projection to the SBPVpswassimilar in injections centered in the core or the shell region ofthe nucleus, a result that is at odds with previous retrogradetract-tracing studies reporting the shell region as themain SCHsource of inputs to the SBPVps (Leak and Moore, 2001).
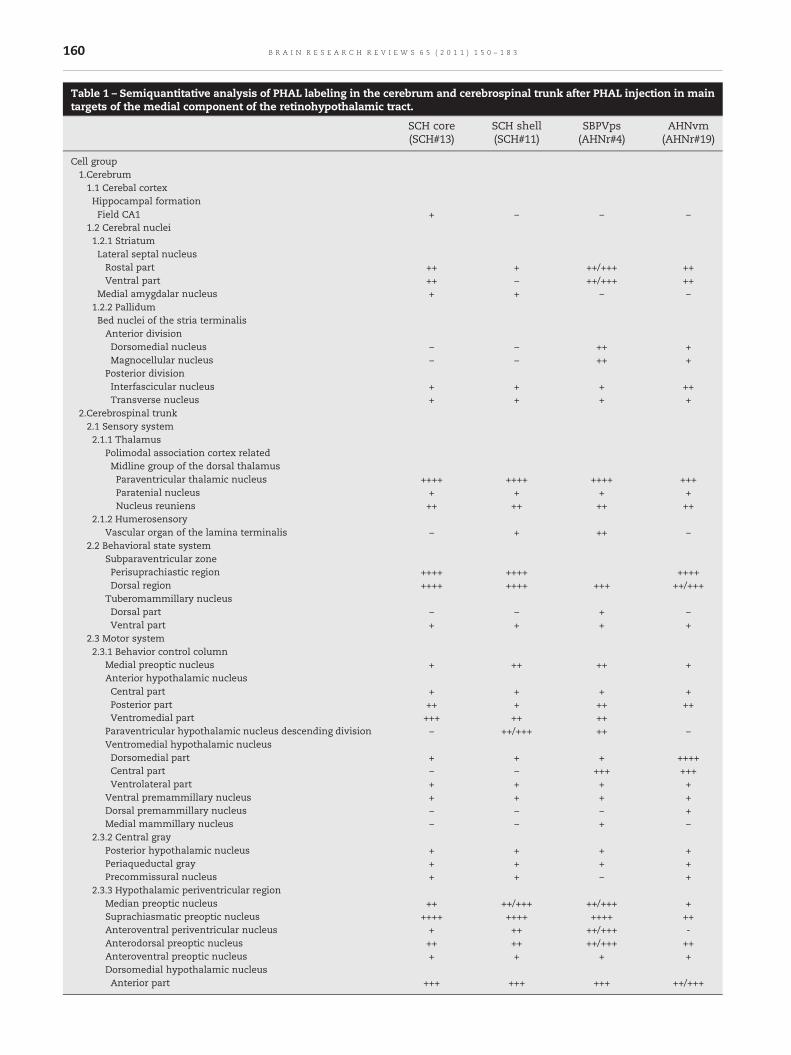
Table 1 – Semiquantitative analysis of PHAL labeling in the cerebrum and cerebrospinal trunk after PHAL injection in maintargets of the medial component of the retinohypothalamic tract.
SCH core SCH shell SBPVps AHNvm(SCH#13) (SCH#11) (AHNr#4) (AHNr#19)
Cell group1.Cerebrum1.1 Cerebal cortexHippocampal formationField CA1 + − − −
1.2 Cerebral nuclei1.2.1 StriatumLateral septal nucleusRostal part ++ + ++/+++ ++Ventral part ++ − ++/+++ ++
Medial amygdalar nucleus + + − −1.2.2 PallidumBed nuclei of the stria terminalisAnterior divisionDorsomedial nucleus − − ++ +Magnocellular nucleus − − ++ +
Posterior divisionInterfascicular nucleus + + + ++Transverse nucleus + + + +
2.Cerebrospinal trunk2.1 Sensory system2.1.1 Thalamus
Polimodal association cortex relatedMidline group of the dorsal thalamusParaventricular thalamic nucleus ++++ ++++ ++++ +++Paratenial nucleus + + + +Nucleus reuniens ++ ++ ++ ++
2.1.2 HumerosensoryVascular organ of the lamina terminalis − + ++ −
2.2 Behavioral state systemSubparaventricular zonePerisuprachiastic region ++++ ++++ ++++Dorsal region ++++ ++++ +++ ++/+++
Tuberomammillary nucleusDorsal part − − + −Ventral part + + + +
2.3 Motor system2.3.1 Behavior control column
Medial preoptic nucleus + ++ ++ +Anterior hypothalamic nucleusCentral part + + + +Posterior part ++ + ++ ++Ventromedial part +++ ++ ++
Paraventricular hypothalamic nucleus descending division − ++/+++ ++ −Ventromedial hypothalamic nucleusDorsomedial part + + + ++++Central part − − +++ +++Ventrolateral part + + + +
Ventral premammillary nucleus + + + +Dorsal premammillary nucleus − − − +Medial mammillary nucleus − − + −
2.3.2 Central grayPosterior hypothalamic nucleus + + + +Periaqueductal gray + + + +Precommissural nucleus + + − +
2.3.3 Hypothalamic periventricular regionMedian preoptic nucleus ++ ++/+++ ++/+++ +Suprachiasmatic preoptic nucleus ++++ ++++ ++++ ++Anteroventral periventricular nucleus + ++ ++/+++ -Anterodorsal preoptic nucleus ++ ++ ++/+++ ++Anteroventral preoptic nucleus + + + +Dorsomedial hypothalamic nucleusAnterior part +++ +++ +++ ++/+++
160 B R A I N R E S E A R C H R E V I E W S 6 5 ( 2 0 1 1 ) 1 5 0 – 1 8 3

Table 1 (continued)
SCH core SCH shell SBPVps AHNvm(SCH#13) (SCH#11) (AHNr#4) (AHNr#19)
Cell groupPosterior part
+ + + +
Ventral part − − −Periventricular hypothalamic nucleusPreoptic part + ++ ++/+++ +Posterior part + + + −
Internuclear area ++ ++ +++ +++2.3.4 Motoneuron groups
Neuroendocrine motor zoneMagnocellularSupraoptic nucleus + + + +
ParvicellularParaventricular hypothalamic nucleusAnterior parvicellular part ++ +++ +++ ++Medial parvicellular part + ++/+++ ++ +Periventricular parvicellular part + ++/+++ ++/+++ +Periventricular hypothalamic nucleus + ++/+++ ++/+++ +Arcuate hypothalamic nucleus − + + −
2.3.5 Reticular formationLateral hypothalamic areajuxtaparaventricular region ++ + ++ ++juxtadorsomedial region − − + +retrochiasmatic area + + ++ ++tuberal nucleus + + ++ ++
Geniculate group, ventral thalamusIntergeniculate leaflet ++ ++ − −Ventral part, lateral geniculate complex + + − −
Pretectal regionOlivary pretectal nucleus + + − −Medial pretectal area + + − −
(−), absence of labeling; (+), light labeling; (++), light to moderate labeling; (++/+++), moderate labeling; (+++) moderate to dense labeling; (++++),dense labeling. The cell group hierarchy is from Swanson (2004).
161B R A I N R E S E A R C H R E V I E W S 6 5 ( 2 0 1 1 ) 1 5 0 – 1 8 3
At anterior hypothalamic levels, some ascending fibersramify throughout the rest of the SBPV (the SBPVd) andprovide a relatively sparse input to the paraventricularnucleus and laterally adjacent juxtaparaventricular region ofthe LHA (Figs. 4G–I and 5F). In addition, lateral fibers in thepathway ascend through, and generate terminal boutons in,immediately adjacent regions of the AHN (Figs. 4F–I and 5F),especially in the AHNvm (Fig. 4F).
As previously suggested in the literature (Watts andSwanson, 1987; Leak and Moore, 2001), the injection centeredin the shell region of the SCH labeled a dense plexus of fiberssituated more medially in the SBPV and provided sparserinputs to immediately adjacent parts of the AHN, includingthe AHNvm (Fig. 5E). In this experiment, a group of fiberscoursing through the SBPV zone extends medially, providinga dense projection to parts of the anterior periventricularnucleus and moderately dense inputs to the paraventricularnucleus, chiefly aimed at the periventricular, dorsal parvi-cellular, andmedial parvicellular parts of the nucleus (Fig. 5E).In line with the present findings, a number of studies havereported that SCH projections to the paraventricular hypo-thalamic nucleus arise chiefly from the shell region of thenucleus (Buijs et al., 1993; Vrang et al., 1995; Teclemariam-Mesbah et al., 1997; Leak and Moore, 2001; Kriegsfeld et al.,2004).
Proceeding caudally, fibers coursing through this pathwayinnervate the dorsomedial nucleus, where they form anespecially prominent terminal field in the anterior part, anda sparse terminal field in the posterior part (Fig. 4J, K). Thedensity and pattern of projections to the dorsomedial nucleuswere similar to those from injections centered in the core orthe shell region of the nucleus, a result that is at odds withprevious retrograde tract-tracing studies reporting the shellregion as the main SCH source of inputs to the dorsomedialnucleus (Leak and Moore, 2001).
At these levels, axons from the SCH also provide amoderate input to the cell-sparse internuclear area betweendorsomedial and central parts of the ventromedial nucleusand the dorsomedial, intermediate periventricular, and arcu-ate nuclei (Fig. 4J, K). Some of these fibers enter the ven-tromedial nucleus, providing a sparse input to dorsomedialpart of the nucleus (Fig. 4J). Moreover, particularly in theexperiment with an injection centered in the dorsomedial partof the SCH, a few labeled fibers project to the arcuate nucleus,and occasional fibers could also be traced to the externallamina of the median eminence.
Fibers coursing through the tuberal region extend caudallyto generate light inputs to the posterior periventricularnucleus, ventral premammillary nucleus, and ventral part ofthe tuberomammillary nucleus (Fig. 4L). In addition, a small

Fig. 5 – Darkfield photomicrographs comparing thedistribution of PHAL immunoreactive axons in theanteroventral periventricular nucleus (AVPV; A, B), theparaventricular thalamic nucleus (PVT; C, D), the anteriorparvicellular part of the paraventricular hypothalamic nucleus(PVHap; C, D), the juxtaparaventricular region of the lateralhypothalamic area (LHAjp;C,D), the subparaventricular zoneofthe hypothalamus (SBPV; E, F), the paraventricularhypothalamic nucleus (PVH; E, F), and the anteriorhypothalamicnucleus (AHN;E, F), following injections centeredrespectively in the SCH shell (experiment SCH#11; A, C and E)and SCH core (experiment SCH#13; B, D and F). Note that theSCH shell provides denser inputs to the periventricular region,including theAVPV,MEPOandPVH;bothcoreandshell regionsof the SCH project equally densely to the PVT and SBPV, and inthis latter region, it is clear that fibers from the SCH core aredistributed more laterally than those from the SCH shell. Forabbreviations: see list. Scale bars: 100 μm.
162 B R A I N R E S E A R C H R E V I E W S 6 5 ( 2 0 1 1 ) 1 5 0 – 1 8 3
number of axons descending through this pathway extendcaudally through (and to) the posterior hypothalamic nucleusinto the periaqueductal gray (PAG) where rostrally, SCH fibersinnervate lightly the precommissural nucleus, and a fewaxons take a dorsolateral course to the pretectum, where theyappear mostly to target the medial pretectal area and olivarypretectal nucleus (Fig. 4M, N). These results generally confirmearlier PHAL analyses in the rat (Watts et al., 1987), except thathere a relatively small number of labeled axons with boutonsare observed caudally in the posterior periventricular hypo-thalamic nucleus, ventral tuberomammillary nucleus, pre-commissural nucleus, medial pretectal nucleus, and olivarypretectal nucleus. Projections to the precommissural nucleusand pretectal region were indicated in a previous PHAL studyin the hamster (Morin et al., 1994).
3.2.4. Lateral pathwayFrom the injection site, a relatively small group of labeledfibers extends caudally and laterally through (and to) theretrochiasmatic area and tuberal nucleus (Fig. 4F–I). A fewof these fibers continue through the ansa peduncularis,providing occasional inputs to the medial amygdalar nucleus,as reported in the hamster by Morin et al. (1994). A largercontingent of fibers coursing through this pathway joins theregion of the supraoptic commissures, and then continuesdorsocaudally between the optic tract and cerebral peduncle.These fibers end as a moderately dense terminal field in theintergeniculate leaflet, along with a sparser input to theventral part of the lateral geniculate complex (Fig. 4M, N). Bothexperiments with injections centered either in the core or inthe shell region of the nucleus labeled an equally denseterminal field in the intergeniculate leaflet, a finding seem-ingly discrepant with previous studies suggesting that thisprojection arises mostly from the core region of the SCH(Kriegsfeld et al., 2004).
3.2.5. Crossed pathwayIn the experiment with a PHAL injection centered in the coreregion of the SCH, only occasional labeled fibers could betraced to the other side of the brain in the medial preopticarea, SBPVps, SCH, and SBPVd, confirming previous PHALanalysis in the rat with an injection centered in the ventralpart of the nucleus (Watts et al., 1987). In contrast, theinjection centered in the dorsomedial part of the SCH labeleda substantial projection to the dorsomedial region in thecontralateral SCH, in addition to a moderate projection to theSBPVps. The present findings on the crossed projections of theSCH are in large agreement with previous observations in therat using either PHAL (Watts et al., 1987; Buijs et al., 1994) orpseudorabies virus transsynaptic tracing, where substantialcrossed projections between the dorsomedial (shell), but notthe ventrolateral (core), regions of the nucleus were reported(Leak et al., 1999).
3.3. Projections from the perisuprachiasmatic region:second rhtm target
In Experiment AHNr4 PHAL-labeled cells are centered in theSBPVps (Fig. 2B'). Labeled fibers from this second majorterminal field of the rhtm course through three of the four

163B R A I N R E S E A R C H R E V I E W S 6 5 ( 2 0 1 1 ) 1 5 0 – 1 8 3
basic pathways described for the SCH (excluding the lateralpathway to the lateral geniculate complex), and the terminalfield distribution is also generally similar, as illustrated sche-matically on a flatmap in Fig. 9C. They will therefore be de-scribed but not formally mapped.
Like the SCH, the SBPVps generates in the septal regionmoderate inputs to the rostral and ventral parts of the lateralseptal nucleus, and only occasional fibers in other parts of theregion. At preoptic levels, axons from the SBPVps ramify verydensely in the suprachiasmatic preoptic nucleus, moderately intheanterodorsal, anteroventral periventricular,medianpreopticnuclei, and vascular organ of the lamina terminalis andrelatively sparsely in the medial and anteroventral preopticnuclei. This projection pattern to the preoptic region moreclosely resembles that found for injections centered in the shellregion of the SCH, and except for the suprachiasmatic preopticnucleus, all these important preoptic targets appear to receive adenser input from the SBPVps than from the SCH itself. Atpreoptic levels, a small group of axons from the SBPVps extendslaterally to end in the anterior division of the bed nuclei of thestria terminalis.
As with the shell region of the SCH, projections from theSBPVps in the dorsal pathway generate a dense input to thepreoptic periventricular nucleus, anterior periventricularnucleus, anterior parvicellular part of the paraventricularhypothalamic nucleus, and rostral levels of the paraventri-cular thalamic nucleus, and sparser inputs to the reuniens andparatenial nuclei, in addition to amoderately dense projectionto the juxtaparaventricular region of the lateral hypothalamicarea (Fig. 6A).
As with the shell region of the SCH, axons from the SBPVpsin the dorsocaudal pathway generate a moderately denseprojection to the SBPV that extends medially to provide a
Fig. 6 – A–C: Representative darkfield photomicrographs showinseveral brain regions following the PHAL deposit, which is illustrsubparaventricular hypothalamic zone (SBPVps; experiment AHNdescribed for the SCH, particularly its shell region, providing densinputs to the periventricular andmedial parvicellular parts of theto project less densely to the rest of the SBPV (B), and provides aabbreviations: see list. Scale bars: 100 μm.
moderate projection to parvicellular parts of the paraventri-cular nucleus, and laterally to provide inputs to adjacent partsof the AHN (Fig. 6B).
At tuberal levels of the hypothalamus, the anterior part ofthe dorsomedial nucleus also receives a substantial inputfrom the SBPVps, and, in contrast to the SCH, this projectionalso provides a substantial number of fibers to the central partof the ventromedial hypothalamic nucleus and the cell-poorinternuclear area between the nucleus and elements of theperiventricular region including the dorsomedial nucleus,intermediate part of the periventricular hypothalamic nucle-us, and arcuate hypothalamic nucleus (Fig. 6C). At these levelsa few axons may extend laterally to provide sparse inputs tothe juxtadorsomedial region of the lateral hypothalamic areathat contains neurons of the medial orexin group. Morecaudally, fibers in this pathway generate inputs to theposterior periventricular nucleus, ventral premammillarynucleus, and dorsal and ventral parts of the tuberomammil-lary nucleus. In addition, a small number of axons descendingthrough this pathway may extend caudally and project to theregion surrounding the medial mammillary nucleus, orcontinue through (and to) the posterior hypothalamic nucleusinto the PAG where a few fibers could be traced up to caudallevels of its ventrolateral division. These results generallyconfirm earlier work by Watts et al. (1987; experiments A43and A34), except in the present experiments dense inputswere also observed in the suprachiasmatic preoptic nucleusand central part of the ventromedial nucleus, in addition torelatively light projections in the medial preoptic nucleus,juxtaparaventricular region of the LHA, posterior periventri-cular nucleus, ventral premammillary nucleus, dorsal andventral parts of the tuberomammillary nucleus, and caudalregions of the ventrolateral division of the PAG. Strong
g the distribution pattern of PHAL immunoreactive axons inated in Figs. 2(B–B''), in the perisuprachiasmatic region of ther4). Note that the SBPVps projection pattern resembles thate inputs to the PVT (A), PVHa (A) and DMHa (C), andmoderatePVH (B). In contrast to the SCH, however, the SBPVps appearsdense terminal field in the central part of the VMH (C). For

Fig. 7 – Summary of projections from the AHNvm. The distribution of PHAL immunoreactive fibers in experiment AHNr19 wasplotted onto a series of standard drawings of the rat brain (Brain Maps—Swanson, 2004) that are arranged from rostral (A) tocaudal (N). The dark gray area in (F) indicates the injection site, which is illustrated in Fig. 2(C–C''). For abbreviations: see list.
165B R A I N R E S E A R C H R E V I E W S 6 5 ( 2 0 1 1 ) 1 5 0 – 1 8 3
projections from the SBPVps to the ventromedial nucleus havealso been reported in the hamster (Kriegsfeld et al., 2004).
The vast majority of projections from the SBPVps are ob-served ipsilateral to the injection site, with only very sparselabeling on the contralateral side.
3.4. Projections from the AHNvm: third rhtm target
In five experiments, PHAL-labeled neurons of the injection siteare centered in the third major terminal field of the rhtm, theAHNvm (Experiments AHNr7, 10, 11, 16, and 19), as confirmedby the CTb-labeled input from the contralateral retina. Ex-periment AHNr19 is described in detail because the injectionsite is confined almost entirely to the AHNvm (Figs. 2C–C'', 7,and 8), and because the projection pattern is typical of thoselabeled in the other experiments with an injection centered inthe AHNvm. Projections from this part of the AHN have notbeen studied before in the rat (see Risold et al., 1994), and onlyvery fragmentary results on the projections of this area have
been previously described in the hamster (Kriegsfeld et al.,2004).
Labeled axons from the AHNvm may be divided into anascending group that innervates targets rostral to the nucleusin the rostral hypothalamus, thalamus, and septal region, anda descending group to caudal hypothalamus and midbrain, asillustrated schematically on a flatmap in Fig. 9D.
3.4.1. Ascending pathwayThe ascending projection from the AHNvm divides into twobranches, ventral and dorsal. Axons in the ventral branch ini-tially form a dense terminal field surrounding the SCH in theSBPVps (Fig. 7D–E). A small group of these fibers continuesrostrally through the preoptic region, where it provides moder-ate inputs to the anterodorsal preoptic nucleus and suprachias-matic preoptic nucleus, as well as a very sparse input to themedial preoptic area, anteroventral preoptic nucleus, andmedian preoptic nucleus (Fig. 7A–C). At preoptic levels, a smallgroup of axons from the AHNvm extends laterally to project to

Fig. 8 – A–C: Representative darkfield photomicrographs showing the distribution pattern of PHAL immunoreactive axons inseveral brain regions following the PHAL deposit (illustrated in Figs. 2C–C'') in the ventromedial part of the anteriorhypothalamic nucleus (AHNvm; experiment AHNr19). Note that the AHNvmprojection pattern resembles that described for theSCH. Themain SCH targets, including the PVT, SBPV andDMHa, receive clear, but less dense inputs, from theAHNvm,which, inturn, provides a robust projection to the dorsomedial and central parts of the VMH. For abbreviations: see list. Scale bars:100 μm.
166 B R A I N R E S E A R C H R E V I E W S 6 5 ( 2 0 1 1 ) 1 5 0 – 1 8 3
the anterior division of the bed nuclei of the stria terminalis,where light inputs are observed primarily in the dorsomedialand magnocellular nuclei (Fig. 7C). Rostral to the anteriorcommissure, some fibers coursing through (and to) the lateralpreoptic area turn dorsally into the septal region, where theycould be traced to the rostral and ventral parts of the lateralseptal nucleus (Fig. 7A–C).
The other ascending branch of axons from the AHNvmcourses dorsally through (and to) the AHN, anterior hypotha-lamic area, and then rostral medial preoptic area to innervatethe anterior parvicellular part of the paraventricular hypotha-lamic nucleus, the posterior division of the bed nuclei of thestria terminalis, juxtaparaventricular region of the lateralhypothalamic area, paraventricular thalamic nucleus, andnucleus reuniens (Fig. 7C–F). Some of these ascending fibersthen extend laterally to innervate relatively moderately thejuxtaparaventricular region of the lateral hypothalamic areaand rostrodorsally to it the posterior division of the bednuclei ofthe stria terminalis, where a somewhat lighter terminal field isobserved throughout, although it is most obvious in theinterfascicular nucleus (Fig. 7D, E). A group of fibers in thedorsal pathway also enters the thalamus, providingmoderatelydense inputs to rostral levels of the paraventricular thalamicnucleus (Figs. 7D, E and 8A), and lighter inputs to the ventral,rostral, and median parts of the nucleus reuniens (Fig. 7F).
3.4.2. Descending pathwayThese axons from the AHNvm descend through the medialhypothalamic nuclei, and to a lesser extent through theperiventricular region. At anterior hypothalamic levels, theAHNvm provides a clear plexus of fibers to the SBPV but only avery sparse projection to nearby periventricular hypothalamicsites, including the paraventricular hypothalamic nucleus
(Figs. 7G–I and 8B). At these levels, the AHNvm also projects toother parts of the anterior hypothalamic nucleus (Figs. 7G–I and8B) and then caudally to other medial hypothalamic nuclei aswell. Thus, at tuberal levels, descending axons fromtheAHNvmgenerate a particularly dense terminal field in the dorsomedialand central parts of the ventromedial nucleus, a projectionpreviously suggested in the hamster with BDA depositsincluding the AHNvm (Kriegsfeld et al., 2004), and then extendinto the internuclear area between the nucleus and the inter-mediate periventricular nucleus (Figs. 7I–K and 8C). At theselevels, a moderate input is also observed to the dorsomedialnucleus, where labeled fibers end mostly in the anterior part(Figs. 7J, K and 8C). Proceeding caudally, a few labeled axonswere traced to and through premammillary levels and then toand through theposterior hypothalamic nucleus into the rostralend of the PAG, where scattered fibers and terminals could beobserved along its entire rostrocaudal extent (Fig. 7L–N).
As with the SCH and perisuprachiasmatic region, the vastmajority of projections from the AHNvm are ipsilateral, withonly a scattered labeled fibers extending to the other side ofthe brain.
3.5. Projections from the LHAavr: main rhtl target
In four experiments (LHAr15, 21, 23, 34), the PHAL injectionlabeled neurons that are centered in the LHAavr, as confirmedby the CTb-labeled input from the contralateral retina (seeFig. 10 A, B). Curiously, all PHAL injections centered in theLHAavr also label a few neurons in the subjacent supraopticnucleus, presumably via dendrites extending dorsally into theinjection site. A very similar pattern of anterogradely labeledfibers is observed in all four experiments. Experiment LHAr21was chosen as a prototype to illustrate the results (Figs. 10

LSr,v DMHaRCH/
TUPVa
AVPVPVpo
PSCH
MEPO
AVPADP PVHMPN
PVHaAHNvm
LHAjpBST
PVT
PT RE
SBPVps SBPVd
I (VMH)
LHAjp
PVp
PRC
PAG
PH
TMv
PMv
MPT / OP
IGL / LGv
LSr DMHa
AVPVPVpo
PSCH
MEPO
AVPADP PVH
MPNAHNvm
LHAjpBST
PVT PT
RE
SBPVps SBPVd
I (VMH)
LHAjp
PVp
PRC
PAG
PH
TMv
PMv
MPT / OP
IGL / LGv
ov
LSr,v DMHa
AVPVPVpo
PSCH
MEPO
AVPADP PVH
MPN
AHN
LHAjpBSTpost
PVT
PT RE
SBPVd I (VMH)/ VMHc
LHAjp
PVp
PRC
PAG
PH
TM
PMv
ov
BSTant
LHAjdMM
LSr,v DMHaPSCH
MEPO
AVPADP PVHMPO
PVHa
AHNLHAjpBSTpost
PVT
RE
SBPVps
SBPVd I (VMH)/
VMHdm, c
LHAjp
PAG
PH
PMd BSTant
SCHc
SCHs
PVaPVHa
SBPVps
PVa
PVHa
AHNvm
AHNc,p
AHNc,p
PT
RCH/TU
RCH/TU
RCH/TU
SCHcore
SCHshell
SBPVps
AHNvm
A
B
C
D
Fig. 9 – The general organization of projections from shell region of the SCH (B) and from the main targets of the medialcomponent of the retino-hypothalamic tract, namely the core region of the SCH (A), the perisuprachiasmatic region of thesubparaventricular zone (SBPVps; C), and the ventromedial part of the AHN (AHNvm; D). The magnitude of each pathway isroughly proportional to the thickness of the line representing it. For abbreviations: see list.
167B R A I N R E S E A R C H R E V I E W S 6 5 ( 2 0 1 1 ) 1 5 0 – 1 8 3
and 11) because the injection here labels the most extensivepopulation of cells in the LHAavr (Fig. 10A, B). The projectionsof the LHAavr have not been previously explored, and some ofthe projection fields here described could be confirmed inearlier retrograde tract-tracing studies.
Labeled axons from the LHAavr may be divided into anascending group that innervates rostral targets in the forebrain,
anda descending group to caudal hypothalamus,midbrain, andpons, illustrated schematically on a flatmap in Fig. 12.
3.5.1. Ascending pathwayMost ascendingaxons from the LHAavr course rostrally throughthe medial hypothalamic nuclei and laterally adjacent regionsof themedial forebrain bundle. At anterior hypothalamic levels,

168 B R A I N R E S E A R C H R E V I E W S 6 5 ( 2 0 1 1 ) 1 5 0 – 1 8 3
the LHAavr provides a dense fiber plexus to the juxtaparaven-tricular region of the lateral hypothalamic area andmoderatelydense projections to the anterior hypothalamic nucleus (allparts) and ventromedially adjacent retrochiasmatic region, andthe SBPV (Figs. 10D and 11E–G). In sharp contrast, only a smallnumber of fibers at these levels innervate the periventricularregion, where they appear to end in the paraventricular nucleus(anterior, anterior magnocellular, and periventricular parts;
Fig. 10 – A: Brightfield photomicrograph to illustrate the appearalocalized in the retinorecipient region of the ventral zone of the rB: Darkfield photomicrograph to illustrate the adjacent CTb-stainD–H: Representative darkfield photomicrographs showing the dibrain regions following a PHAL deposit in the LHAavr (experimenbars: 100 μm.
Figs. 10D and 11E–G). At preoptic levels, LHAavr fibers generatea dense input to undifferentiated parts of the medial preopticarea and adjacent lateral part of the medial preoptic nucleus,whereas only a few appear to project to, or through, the medialand central parts of this nucleus (Fig. 11C–E). In addition, denseprojections from the LHAavr are observed in the medianpreoptic and anteroventral preoptic (Figs. 10C and 11C, D)nuclei. Projections from the LHAavr to themedian preoptic and
nce of a PHAL injection site for a representative injectionostral region of the LHA (LHAavr; experiment LHAr21).ed section showing the retinal projection field to the LHAavr.stribution pattern of PHAL immunoreactive axons in severalt LHAr21). See text for details. For abbreviations: see list. Scale

Fig. 10 (continued).
169B R A I N R E S E A R C H R E V I E W S 6 5 ( 2 0 1 1 ) 1 5 0 – 1 8 3
anteroventral preoptic (corresponding in part to the ventrolat-eral preoptic region) nuclei have been suggested in earlierretrograde-tract-tracing studies (Saper and Levisohn, 1983;Chou et al., 2002).
Agroupof ascendingLHAavr fibersextendsdorsally fromtheregion of the juxtaparaventricular region of the lateral hypo-thalamic area toenter the thalamuswheredense terminal fieldsare established in the lateral part of the rostral nucleus reuniensand rostral levels of the paraventricular nucleus (Fig. 11E–G). Arelatively small number of axons also enter the stria medullarisand then generate light terminal fields in the lateral habenulaand caudal levels of the paraventricular nucleus (Fig. 11H, I). Inaddition, a small group of LHAavr fibers descending through theposteriorhypothalamicnucleusalsoextend into caudal levelsofthe thalamus, where they end in the intermediodorsal nucleus(Fig. 11I).
Ascending axons from the LHAavr also provide substantialinputs to restricted parts of the telencephalon, including thelateral septal nucleus, bed nuclei of the stria terminalis, and
amygdalar region. At preoptic levels, many fibers leave thehypothalamus by taking either a dorsolateral route toward theBST or a dorsal route rostral to the anterior commissuretoward the lateral septal nucleus, basically coursing through(and to) the medial and lateral preoptic areas (Fig. 11C–E).Within the posterior division of the BST, the LHAavr suppliesmoderately dense inputs to the transverse and interfascicularnuclei (Fig. 11E). At this level, a group of labeled axons alsoenters the substantia innominata (in the ansa peduncularis),or to a much lesser extent the stria terminalis, to access theamygdalar region, as described below. Within the anteriordivision of the BST, the LHAavr supplies moderate to denseinputs to the magnocellular, dorsomedial, and ventral nuclei,and the anteromedial area (Fig. 11D). In the lateral septalnucleus, rather dense inputs are generated in the medial andventrolateral zones of the rostral part (Fig. 11A–D). Projectionsto the lateral septal nucleus have also been suggested inprevious retrograde tracer studies (Risold and Swanson, 1997).A few labeled axons in the septal region enter the column ofthe fornix and then continue through the fimbria to thehippocampus, where occasional labeling is observed in ventralfield CA1.
Ascending axons from the LHAavr also provide inputs toseveral parts of the amygdalar region, reaching it through twodistinct pathways, the stria terminalis and ansa peduncularis,with the latter representing by far the main route (Fig. 11D–G).Two distinct bundles of labeled axons course through the ansapeduncularis to the amygdalar region. The first consists offibers that leave the posterior division of the BST and coursethrough the substantia innominata (Fig. 11E–G), branchingsomewhat and generating terminal boutons along the way.The second bundle consists of a smaller number of fibers thatreach the amygdalar region by traveling directly through thelateral preoptic and lateral hypothalamic areas (Fig. 11D–G). Inthe amygdalar region, a strikingly dense projection was tracedto the posterior nucleus, which represents one of the maintargets of the LHAavr (Figs. 10H and 11I–J), a projection alsodescribed in previous retrograde tracing studies (Canteraset al., 1992). The LHAavr also provides sparser, thoughmoderate inputs to other amygdalar sites, including theanterodorsal part of the medial nucleus and medial part ofthe central nucleus, and light inputs to the posterolateralcortical nucleus and posterior basomedial nucleus (Fig. 11F–H).
3.5.2. Descending pathwayDescending axons from the LHAavr course through themedialhypothalamic nuclei and adjacent medial regions of themedial forebrain, with a dorsal branch through the midbrainperiventricular system. At tuberal levels, the LHAavr providessubstantial inputs to the ventromedial and dorsomedialnuclei, as well as to adjacent parts of the lateral hypothalamicarea (Fig. 11G, H). In the ventromedial nucleus, a dense ter-minal field extends from the anterior part to rostral levels ofthe central part of the nucleus, whereas a relatively sparseprojection is found in the ventrolateral part of the nucleus(Figs. 10D, E and 11G, H). In the dorsomedial nucleus, theLHAavr appears to innervate densely the anterior part of thenucleus (Figs. 10E and 11H), and only very sparsely the othertwo parts. The projection to the dorsomedial nucleus has alsobeen suggested in earlier retrograde tracer studies (Thompson

170 B R A I N R E S E A R C H R E V I E W S 6 5 ( 2 0 1 1 ) 1 5 0 – 1 8 3
and Swanson, 1998). Furthermore, at these levels the LHAavralso generates dense terminal fields in particular componentsof the lateral hypothalamic area including the suprafornical,
ccg
CP
VL
LSr
r.m
r.vl.d
.m r.v
l.d.l
SH
OT
ACB aco
NDB
fx
SI
int
cc
LPO
CP
VL
MPO
BST
NDB
aco
act
PS
ME
PO
SH
dm
OT
AD
P
AVP
och
MS
fu
ov
ju
cc
AV
PV
av
al LPO
LSv
LSc
LSr
r.m.v
.c
r.vl.v
c.d
c.v.
m c.
v.i c.
v.l
LH Aav
LHAai
LSc.d.r
fx
opt
SI
int
st
fi
AHNa
pv
PVpo
SO
sm PVT
V3
GPl
CP
PT
VL
SCH
SFO
MA
MPN
l m
MPO
BST pr
pr
if
if
v
SF
tr am am
NDB
PVHa
A B
C D
E F
Fig. 11 – Summary of projections from the LHAavr. The distributioplotted onto a series of standard drawings of the rat brain (Braincaudal (O). The dark gray area in (F) indicates the injection site, w
subfornical, juxtaventromedial, and juxtadorsomedial regions(Figs. 10E and 11H). A few of these axons continue dorsally toprovide a light input to rostral levels of the zona incerta.
SI
ccg
LPO
CP
VL
NDB
SH
OT
MS
FS
ACB
aco
LSc
LSr
r.dl.m.d r.dl.l.d
r.m.d
r.m.v
.r r.vl.d
.m
r.vl.d
.l
opt
SI
int
st
cc
LPO
PVpo
V3
V3
GPl
CP
SCH
MPN
l m
MPO
BST
pr
v
SF
NDB
c
mg
dl rh
al
aco act
PD
PS
TRS
LSc.d
LHAjp LHAad
LHAav
d
l
LHAai
fx
opt
SI
int
st
ZI
MDm
RE
AHNc
PVH pmm mpd
PVa
V3
ME
Aad st
CEAm
LA
sm
PVT
V3
RT
1 2
BMAa
IAD
sup
COAa
c
c
CM IAM
MDl PT
p
RCH
dp
SO
BA
a
AAA
SB
PV
m
n of PHAL immunoreactive fibers in experiment LHAr21 wasMaps—Swanson, 2004) that are arranged from rostral (A) tohich is illustrated in Fig. 10 (A). For abbreviations: see list.
fx
pm
cpd
LHA
ZI
PF
OP
COApm
BMAp
PHSNr
PA
DGlbMM
TMv
fr
SUMl
FF
APN
PRC
PAG
PAG
MRN
mp
LM
VTA
SUBv
CA3
NPC
ml
MRN
cpd
AQPAGl
MRN
VTA
ml
ND
RN
SC
sg
ig
dg
dm
dl
mlf
CLI
IPN
RR
III
scp
AQ
Dmlf
scp
MEV
NLL
ll
CUN
IC e
VTNPPN
PRNrCSm
CSl
LDT
PAGl
PAGvl
PBlcls
le
KF
PBmmR
fxopt
SI
mtt
int
st
fi
VMHdm
LHA
ZI
PCN
MD
RE
ARH
V3
ME
GPm
MEApv
IA
CEAl
BLAp
SMT
RH
smLH
CLPVT
ml
BMAa
PR
sup
MEA
pd
vl
m
c
c
CM
c
c
DMHa
BLAa
BMAp
da
IMD
cpd
c
cb
a
TUsv
TUi
LHAm
LHAjvvLHAsfpLHAjvd
LHAjd
LHAsLHAd
fx
opt
pm cpd
fi
ZI
PF
MD
m
PVp
BLAp
sm
MH
LH
CL
PVT
COAplCOApm
CM
BMAp
IMD
LP
PH
STN
PA
DGlb
PMv
PMd
TMv
SPFm
SUBv
fr
SU
Ml
MM
me
FF
ml
CA1
CA3
LHAp
PSTN
fx
opt
SI
int
st
fi
VMHa
ZI
MDm
RE
AHNc
ARH
PVH pmlmpdpv
PV
i
V3
ME
GPm
MEAad
IA
CE
Am
BLAa
RH
sm
PVT
GPl
CP
1
2
BMAa
IAD
COAa
av
CEAl
CEAc
c
CM
IAM
MDl
PT
p
dp
mpv
BA
SB
PV
m
LHAjpLHAad
LHAai
LH Aav
TUsv
d
l
SUMm
PAGdm
dl
m
G H
I J
K L
Fig. 11 (continued).
171B R A I N R E S E A R C H R E V I E W S 6 5 ( 2 0 1 1 ) 1 5 0 – 1 8 3
At premammillary levels, descending axons from theLHAavr provide a rather dense bilateral projection to ventralregions of the dorsal premammillary nucleus, as well as lightinputs to the ventral premammillary and ventral tuberomam-
millary nuclei (Figs. 10F and 11I). The projection to the dorsalpremammillary nucleus has also been noted in previousretrograde tracer studies (Comoli et al., 2000). Proceedingcaudally, at mammillary levels, a few axons are observed in

SLD
scp MEV
NLL
ll
CUN
DTN
PPN
PRNr
CSm CSl
LDT
V4
PCG DR PBmm
KF
lc
lv ld
le
SLD
scp
MEV
DTN SUT
LDT
V4
PCG
c
PBmm me
lc lv
le LC
V
B
mv
NId
SLC
mlf
scp
MEV
DTN PCG
V4
c
PBmm
lc lv
LC
V
NId
M N
O
Fig. 11 (continued).
172 B R A I N R E S E A R C H R E V I E W S 6 5 ( 2 0 1 1 ) 1 5 0 – 1 8 3
the medial and lateral parts of the supramammillary nucleus,and in the capsule surrounding ventrolaterally the medialmammillary nucleus (Fig. 11J). A few axons in this pathwaycontinue through (and to) the ventral tegmental area,apparently to end in the rostral and central linear nuclei ofthe raphe (Fig. 11K).
Many descending axons from the LHAavr travel caudallythrough (and generate terminal boutons in) the posteriorhypothalamic nucleus to enter the PAG (Figs. 11I, J). Rostrallythey generate a rather dense terminal field in the precom-missural nucleus (Fig. 11J). At the level of the oculomotornucleus, a dense terminal field is observed in the lateraldivision of the PAG (Figs. 10G and 11K), a terminal fieldidentified in earlier retrograde tracer studies (Mota-Ortiz et al.,2009). At this level a relatively small group of axons is alsoobserved coursing to and through the dorsomedial division ofthe PAG (Figs. 10G and 11K). At caudal PAG levels, fiberscoursing through themidbrain periventricular system providea moderately dense input to the ventrolateral division(Fig. 11L). A small number of fibers coursing through the
LH
VMHa AVP
MPO MPNl
PVH SBPV
PVT
RE
LHAjp LHA
AHN
BSTdm, mg, if, tr
MEPO LSr
LSc
ap, s
t
SI
PT
sm
ZIda
VMH
LHAavr ADP
LPO
RCH
Fig. 12 – The general organization of projections from themain tanamely the retinorecipient region of the LHAav. Themagnitude oline representing it. For abbreviations: see list.
midbrain periventricular system enter the superior colliculus(Fig. 11K), the midbrain reticular nucleus (Fig. 11J, K), and thesuperior central nucleus (Fig. 11L, M), providing sparse inputsto these gray matter differentiations. At the caudal pole of thePAG, a group of fibers leaves the ventrolateral division to enterthe cuneiform nucleus and parabrachial nucleus, where aclear terminal field is observed in the external lateral part(Fig. 11M). A relatively small number of labeled axons extendcaudally from the PAG into the pontine central gray, wherethey provide a circumscribed, moderate input to Barrington'snucleus (which input is bilateral, with an ipsilateral predom-inance), a projection also suggested in a previous retrogradetracing study (Valentino et al., 1994), and light inputs tothe laterodorsal tegmental nucleus and locus ceruleus(Fig. 11M–O).
Except for the dense contralateral projection to the dorsalpremammillary nucleus (see Fig. 11F), all other terminal fieldsestablished by LHAavr are very predominantly ipsilateral, withonly a few axons crossing themidline to the contralateral side.In general, the very sparse crossed projections are a mirror
IMD
DMHa PMd PMv MM TM SUM
PH
VTA
CLi
PAG B
LC CUN PBle
CS PRC
s, jd, jv, sfp
PA
CEA MEA
vl
BMAp
COAp
SC
MRN LDT
LHAavr
rget of the lateral component of the retino-hypothalamic tract,f each pathway is roughly proportional to the thickness of the

173B R A I N R E S E A R C H R E V I E W S 6 5 ( 2 0 1 1 ) 1 5 0 – 1 8 3
image of the ipsilateral projections. In the forebrain, labeledfibersmay cross themidline either dorsal to the third ventricleor through the supraoptic commissures. In themidbrain, mostfibers cross the midline dorsal to the cerebral aqueduct in thePAG.
3.6. Control injections
Two PHAL injections centered in the LHA just dorsal to theLHAavr were obtained (Experiments LHA28 and 60) and serveas excellent controls for the specificity of LHAavr projections,which are quite different. Labeled neurons in the injection siteof these two experiments are centered in the ventral andintermediate zones of the anterior region of the LHA (Figs. 13and 14), and the projections of these LHA differentiations willbe described fully in a subsequent publication. Here majordifferences will be outlined to document specificity of LHAavrprojections. In both experiments with an injection just dorsalto the LHAavr clear terminal fields are labeled in the medialseptal complex and caudal levels of the lateral septal nucleus(Fig. 14A, B; contrast with Fig. 11B–C), and there are sparse atbest projections to the bed nuclei of the stria terminalis andamygdalar region (Fig. 14A, B; contrast with Fig. 11D, E). In thethalamus, there is little input to the paraventricular nucleus,but instead amassive input the lateral habenula (Fig. 14D), anddense inputs to the lateral part of the mediodorsal nucleus(Fig. 14C) and the paratenial nucleus (contrast with Fig. 11F, G).In the hypothalamus there is a dense terminal field in thelateral preoptic area and light input to the medial preopticarea (Fig. 14A, B; contrast with Fig. 11C–D), and very little inputto the dorsomedial nucleus and juxtadorsomedial region ofthe LHA (contrast with Fig. 11H). More caudally, there aremoderate terminal fields in the dorsal premammillary(Fig. 14E) and supramammillary (Fig. 14F) nuclei. Finally,there is only a very sparse input to the PAG (contrast withFig. 11J–L). Interestingly, a 3H-amino acid injection centered
LHAavr
LHAav
SCH
MPN
AHNc
a
BST
PVH
opt
SO
LHANI #28
23
Fig. 13 – Camera lucida plots of labeled neurons following a contrzone of the anterior region of the LHA, immediately dorsal to theThe drawings are arranged from rostral to caudal (left to right), an2004) plate numbers, to indicate the approximate rostro-caudal l
just rostral to the LHAavr, just over the rostral third of thesupraoptic nucleus in a transitional region between the lateralpreoptic area and the LHAav, labeled a general projectionpattern very similar to that just described for the region justdorsal to the LHAavr (Swanson, 1976; see Fig. 7 there).
4. Discussion
4.1. The medial retinohypothalamic tract (rhtm) andcircadian rhythm control
From the literature it is clear that the three most obviousterminal fields of the rhtm include the SCH, SBPVps, andANHvm, and the present results demonstrate that all threeproject to elements of the network controlling circadianrhythms. Moore et al. (1995), using retrograde tract-tracingmethods and transneuronal transport of the Bartha strain ofthe swine herpes virus, showed that the retinal ganglion cellsgenerating the rhtm are widely distributed over the retina andcorrespond to a subset of type III, or W, cells. These retinalneurons presumably exhibit firing rate changes in response tochanges in luminance, and are probably unresponsive to othercharacteristics of visual stimuli.
The ventrolateral part of the SCH represents themain targetof the rhtm. Extensive research on the SCHdemonstrates that itisheterogeneous in termsof extrinsic axonal input patternsanddistinct neuron types. As noted in the Introduction, the SCHhasbeen conceptualized as having at least two parts, named core(ventrolateral part) and shell (dorsomedial part). Phenotypically,the SCH shell is delineated by vasopressin (VAS)-, enkephalin-,angiotensin II-, andCAR-expressingneurons (Vandesandeetal.,1975; Card and Moore, 1984; van den Pol and Tsujimoto, 1985;Watts and Swanson, 1987; Jacobowitz and Winsky, 1991),whereas the SCH core contains a variety of other cell pheno-types, including vasoactive intestinal polypeptide (VIP)-,
LHAavr
LHAaiAHN
SCH
PVH
opt
SO
fx
24
ol injection of PHAL centered in the ventral and intermediateretinorecipient region of the LHAav (experiment LHANI #28).d the numbers on the top-right refer to Brain Maps (Swanson,evels.
Fig. 14 – A–F: Representative darkfield photomicrographs showing the distribution pattern of PHAL immunoreactive axons inseveral brain regions following a control PHAL injection, which is illustrated in Fig. 13, centered in the ventral and intermediatezone of the anterior region of the LHA, immediately dorsal the LHAavr (experiment LHANI #28). See text for details. Forabbreviations: see list. Scale bars: 200 μm in (A) and (B), 100 μm in (C-F).
174 B R A I N R E S E A R C H R E V I E W S 6 5 ( 2 0 1 1 ) 1 5 0 – 1 8 3

175B R A I N R E S E A R C H R E V I E W S 6 5 ( 2 0 1 1 ) 1 5 0 – 1 8 3
neurotensin-, gastrin-releasing peptide (GRP)-, and calbindin-expressing cells (Card et al., 1981; Watts and Swanson, 1987;Stopa et al., 1988; Mikkelsen et al., 1991; Arvanitogiannis et al.,2000). The SCH contains self-sustained rhythmically firingneurons driven by the autoregulatory transcription-translationfeedback that regulates expression of the Period (Per) andCryptochrome genes (Reppert and Weaver, 2001). These neuronsare largely confined to the SCH shell (Hamada et al., 2001, 2004;Yan and Okamura, 2002). Conversely, the SCH core receivesdirect retinal input and expresses Per1 and Per2 in response tophase-shifting light pulses (Romijn et al., 1996; Silver et al.,1996b; Hamada et al., 2001; Yan and Okamura, 2002; Yan andSilver, 2002). The SCH core region also receives other visualinputs from the geniculo- and pretectohypothalamic tracts(Swanson et al., 1974; Card and Moore, 1989; Mikkelsen andVrang, 1994; Moga and Moore, 1997), and terminals of seroto-nergic fibers (Moore et al., 1978; van den Pol and Tsujimoto,1985).
For the circadian clock to respond to light-induced phaseshift, photic information must be relayed from light-respond-ing to rhythmic neurons. Thus, following phase-shifting lightpulses, Per expression is first induced in the SCH core, which isfollowed by Per expression in the SCH shell (for review, seeAntle and Silver, 2005). As we describe here, the core projectsdensely to the shell region, and the neurotransmitters thathave been implicated in intra-SCH communication of photicsignals include GRP and VIP (for review, see Antle and Silver,2005). In addition, communication between the core and shellregions of the SCH may involve nitric oxide (NO) signaling,which has been shown to support circadian phase-shiftsrequired for synchronization to light–dark (LD) cycles (Planoet al., 2010). Therefore, an important function of the coremight be to use primary and secondary visual information toentrain the pacemaker mechanisms in the shell region.Importantly, we have found strong commissural projectionsbetween the shell, but not the core, regions of the SCH, whichis supported by other studies in the rat (Watts et al., 1987; Buijset al., 1994; Leak et al., 1999).
Confirming the literature (Leak and Moore, 2001; Kriegsfeldet al., 2004), we found that the SCH core and shell regions mayalso interact at the level of projection targets. By and large, thesetwo SCH regions generate similar projection patterns, withsubstantial inputs to the anterior paraventricular thalamicnucleus, suprachiasmatic preoptic nucleus, SBPVps, SBPVd,and dorsomedial nucleus. In the SBPV, the shell region projectsmoremedially,whereas fibers fromthe core regionwere locatedmore laterally, spreading to a larger degree to adjacent parts ofthe anterior hypothalamic nucleus, including the AHNvm.Similar findings have also been reported in studies usingretrograde tracers (Leak and Moore, 2001) and immunostainingfor VAS and VIP (Watts and Swanson, 1987). In line withprevious reports, we have also found that the shell regionprovides denser inputs to a number of preoptic targets,particularly the vascular organ of the lamina terminalis andthe anteroventral periventricular preoptic nucleus (Leak andMoore, 2001; Kriegsfeld et al., 2004), in addition to denserprojections to a number of periventricular sites, including theperiventricular preoptic nucleus, anterior part of the periven-tricular hypothalamic nucleus, and the paraventricular hypo-thalamic nucleus (Buijs et al., 1993; Vrang et al., 1995;
Teclemariam-Mesbah et al., 1997; Leak and Moore, 2001;Kriegsfeld et al., 2004).
The SBPVps is another important rhtm target. Our findingsindicate that, in addition to direct retinal projections, theSBPVps integrates inputs from other rhtm targets, includingthe SCH core and shell and the AHNvm. Confirming previousstudies by Watts et al. (1987), we also found that the SBPVpsgenerates a projection pattern similar to that described for theSCH itself. In particular, the SBPVps has a projection patternmuch closer to that described for the shell than for the corepart of the SCH. However, in contrast to the SCH shell, which islargely avoided by axonal inputs from outside the SCH, theSBPVps integrates inputs from a variety of telencephalic,diencephalic, and pontine sites, including the infralimbiccortical area, ventral subiculum, lateral septal nucleus, andmedial preoptic, ventromedial hypothalamic, and posteriorhypothalamic nuclei, as well as the pontine parabrachialnucleus (Canteras and Swanson, 1992; Vertes et al., 1995;Canteras et al., 1994; Moga and Moore, 1997; Risold andSwanson, 1997). This distribution of axonal inputs to theSCH shell and SBPVps helps explainwhy the daily SCH activitycycle remains locked to the light–dark cycle, whereas essentialenvironmental cues, such as food availability, ambienttemperature, and social interactions, are able to entrainadaptive rhythmic daily activities like body temperature,sleep–wakefulness, feeding, locomotor activity, and cortico-steroid secretion, thus maximizing chances of survival andreproduction.
We showhere that the AHNvm is an important target of therhtm, aprojectionpreviously shown in the rat, and in anumberof other species as well (Conrad and Stumpf, 1975; Johnson etal., 1988; Murakami et al., 1989; Levine et al., 1991; Costa et al.,1999). Like the SBPVps, the AHNvm receives inputs from theSCH core, and to a lesser degree, from the shell regions, inaddition to the brain regions just listed for the SBPVps(Canteras and Swanson, 1992; Vertes et al., 1995; Canteraset al., 1994; Moga and Moore, 1997; Risold and Swanson, 1997).On the efferent side, like the other targets of the rhtm,we showhere that the AHNvm generates a projection pattern similar tothat described for the SCH, particularly its core region. Thus,like the SBPVps, the AHNvm is positioned to entrain rhythmicdaily activities in response to a wide range of environmentalcues, from the light–dark cycle to a number of survival relevantcues, including food availability, ambient temperature, andsocial interactions. However, at the moment, the exact role ofthe SBPVps and AHNvm in entraining circadian rhythmsfollowing exposure to these environmental cues remains tobe demonstrated experimentally.
Interestingly, both the SBPVps and AHNvm share strongconnections with the ventromedial hypothalamic nucleus,which may be a component of the neural system regulatingenvironmental entrainment of circadian rhythms. In line withthis view, massive lesions centered in (but not restricted to)the ventromedial nucleus abolish the prefeeding corticoste-rone peak in rats whose access to food is restricted to a fixedtime of day (Krieger et al., 1977; Krieger, 1980).
In summary, the present findings support the hypothesisthat rhtm targets are primarily involved in controllingcircadian rhythms. First, retinal inputs to the SCH core regionare critical for controlling photic induced phase-shift

176 B R A I N R E S E A R C H R E V I E W S 6 5 ( 2 0 1 1 ) 1 5 0 – 1 8 3
mechanisms, acting in the shell region, and perhaps in otherSCH targets as well. Second, the other two rhtm targets–SBPVps and AHNvm–also project to the SCH circuitry and arethus likely to modulate circadian rhythms. In addition toretinal inputs, both the SBPVps and AHNvm receive stronginputs from a variety of brain sites, and are thus likely to servea unique function in entraining circadian rhythms followingexposure to a wide range of environmental cues, varying fromthe light–dark cycle to a number of environmental cues criticalfor survival.
4.2. Circuitry underlying circadian rhythms
The SCH is classically known as the primary circadianpacemaker locus in mammals. SCH lesions abolish circadianrhythms (Moore and Eichler, 1972; Stephan and Zucker, 1972),whereas transplants of donor SCH tissue restore motorbehavior circadian rhythms, despite a lack of axonal connec-tions between graft and host brain. Thus, locomotor behaviorcircadian rhythms do not necessarily require neural outputfrom the SCH, and can be supported instead by diffusiblesignals in intact animals and by transplants in SCH-lesionedanimals (Silver et al., 1996a; Kramer et al., 2001; Cheng et al.,2002; Kraves and Weitz, 2006).
However, SCH grafts do not restore melatonin or cortico-steroid rhythms, which thus require SCH neural output asconfirmed by evidence that endocrine rhythms are abolishedafter knife cuts of SCH projections (Silver et al., 1996a,b;Nunez and Stephan, 1977; Meyer-Berstein et al., 1999). Thus,SCH participation in most circadian rhythm control involvesits axonal projections, and even the paracrine effects ofdiffusible factors from the SCH must rely on nearby receptivebrain tissue involved in the circuitry underlying circadianrhythms.
The central components of circuitry controlling circadianrhythms are the SCH and SBPV, which present a column oftissue that arches dorsally and caudally, occupying the regionjust above the SCH (the SBPVps) and the region ventral to theparaventricular hypothalamic nucleus (the SBPVd). As previ-ously discussed, the SCH and SBPVps share virtually the sametargets, and this dual stage in conveying circadian rhythminformation is important to understand how light entrain-ment of circadian rhythms could be separable from entrain-ment by a number of environmental cues critical for survival.In the present work, we have added a new element to thiscontrol, the AHNvm, whose axonal projections, as documen-ted, resemble in many ways those of the SBPVps (and SCH).Thus, daily cycles of activity in the SCH remain locked to thelight–dark cycle, whereas adaptability to external eventsshould rely, at least in part, on the second stage of thecircadian integrator, which includes the SBPVps and perhapsthe AHNvm as well.
The SBPVps and SBPVd have a differential role in circadianrhythm control. Ibotenic acid lesions involving the SBPVpsreduce the amplitude of circadian rhythms associated withsleep–wakefulness, locomotor activity, and corticosteroid se-cretion, but seemnot to correlatewith theamplitudeof thebodytemperature circadian rhythm (Lu et al., 2001; Saper et al., 2005).Conversely, lesions involving the SBPVd significantly influencetheamplitudeof thebody temperature circadian rhythm, butdo
not affect sleep or corticosteroid secretion rhythms (Lu et al.,2001).
As pointed out in the Results, the anterior part of thedorsomedial hypothalamic nucleus is heavily targeted by boththe SCH and SBPVps. A number of studies suggest that thedorsomedial nucleus is critical for maintainingmost circadianrhythms. Lesions of the nucleus cause severe loss of circadianrhythms associated with sleep–wakefulness, feeding, locomo-tor activity, and corticosteroid secretion, but not body temper-ature (Chou et al., 2003). Comparedwith the SBPV and SCH, thedorsomedial nucleus has more extensive outputs to themajoreffector sites for these functions (Thompson et al., 1996).Moreover, like the SBPV, the dorsomedial nucleus is alsotargeted by widely distributed brain sites, located in thetelencephalon, hypothalamus, and lower brainstem (Thomp-son and Swanson, 1998), and therefore should also serve as akey interface for circadian rhythm entrainment by environ-mental cues other than light and darkness.
Much has been learned about mechanisms converting theclock signal into patterning of physiological and behavioralrhythms. As mentioned, sleep–wake, costicosteroid, andlocomotor rhythms (and also probably feeding cycles) dependon theprojection fromSCH to SBPVps, and then todorsomedialnucleus. As far as corticosteroid secretion is concerned, allthree of these hypothalamic neuron groups provide directaxonal inputs to the dorsal zone of the medial parvicellularpart of the paraventricular nucleus containingneuroendocrineCRH neurons, with the dorsomedial nucleus providing partic-ularly dense, and the SCH and SBPVps providing sparse tomoderate, terminal fields. Similarly, effectors governing sleep–wake activity may also receive relatively sparse direct projec-tions from the SCH and SBPVps, and more substantial directprojections from the dorsomedial nucleus. It has been shownthat the ventrolateral preoptic region (VLPO) receives sparsedirect projections from the SCH and SBPVps, andmuch denserinputs from the dorsomedial hypothalamic nucleus (Thomp-son et al., 1996). The VLPO has been implicated in theregulation of both rapid-eye movement (REM) and non-REMsleep (Sherin et al., 1996; Lu et al., 2000). Moreover, the SCH andSBPVps appear to provide relatively weak inputs to wakepromotor nuclei, including the orexinergic neuron groupcentered in dorsal tuberal levels of the hypothalamus andthe histaminergic neuron group centered in the tuberomam-millary nucleus. The regions containing both of these neurongroups receive much stronger projections from the dorsome-dial hypothalamic nucleus, which also projects to a number ofother wake-regulatory lower brainstem sites, including thedorsal raphe nucleus, cholinergic laterodorsal tegmentalnucleus, and locus ceruleus (Thompson et al., 1996). Curiously,all of these neuron groups implicated in sleep–wake controlshow activity diurnal rhythms that persist under constantdarkness, suggesting that they receive endogenous circadiansignals from the SCH (for review, see Deurveilher and Semba,2005). Dorsomedial nucleus neurons provideGABAergic inputsto theVLPOandglutamatergic inputs to theorexinergic neurongroup. Therefore, the dorsomedial nucleus probably increaseswakefulness and arousal, exciting orexinergic arousal neuronsand inhibiting sleep neurons in the VLPO (Chou et al., 2003).
Melatonin secretion, in contrast, is seemingly not affectedby lesions of the SBPV or dorsomedial nucleus (Mota et al.,

177B R A I N R E S E A R C H R E V I E W S 6 5 ( 2 0 1 1 ) 1 5 0 – 1 8 3
2001; Saper et al., 2005). The melatonin rhythm depends on adirect projection from the SCH to the paraventricular hypo-thalamic nucleus. Pseudorabies virus injected into the pinealgland of rats labeled the SCH shell, following the appearanceof infected neurons in the dorsal and lateral parvicellular partsof the paraventricular hypothalamic nucleus (Larsen et al.,1998). The SCH is more active during the light cycle, and itsGABAergic neurons inhibit the paraventricular premotorneurons that promote melatonin secretion (Munch et al.,2002). During the dark part of the cycle, this inhibition isreleased and melatonin secretion peaks, locked to the light–dark cycle and seemingly not entrained by other environmen-tal cues.
A commonly used strategy to manipulate circadian (or atleast diurnal) rhythms of a wide range of behaviors andphysiological functions is to restrict food availability to asingle period at a fixed time of day (restricted feeding).Animals adapt to this condition within a few days, andbecome active about an hour before food is presented. Suchanticipatory behavior is accompanied by increased bodytemperature, corticosterone secretion, and gastrointestinalmotility (Mistlberger, 1994; Stephan, 2002), whereas SCHrhythmicity remains phase-locked to the light–dark cycle(Damiola et al., 2000; Stokkan et al., 2001). Unfortunately, theneural and molecular substrates controlling food-entrainedbehavioral rhythms have proven difficult to identify and arethe subject of controversy (for review, see Mistlberger, 2009).
As shown here, the paraventricular thalamic nucleus isanother important target of the SCH, SBPVps, and AHNvm.However, its role in the control of circadian rhythms is stillpoorly understood. Lesions of the paraventricular thalamicnucleus seemingly induce increased locomotor activity duringthe latter half of the dark period (Landry et al., 2007), an effectassociatedwith longer circadian period in blind rats (Moga andMoore, 2000) and animals maintained in constant dark (Edgaret al., 1991). The paraventricular thalamic nucleus also seemsto play a role in modulating circadian rhythm entrainment tolight (Salazar-Juarez et al., 2002).
4.3. Functional roles of the LHAavr–the main target ofthe rhtl
The rhtl provides a very sparse input to the lateral preopticarea and a massive input to the LHA, centered in a regionsurrounding the border of the optic tract and the caudal twothirds of the supraoptic nucleus, in the core of what hasrecently been identified as the ventral zone of the rostralregion of the LHA (LHAav–Swanson, 2004; Swanson et al.,2005), and referred to here specifically as the retinorecipientregion (or core) of the LHAav (LHAavr). This region corre-sponds, at least in part, to a perinuclear zone of the supraopticnucleus that has been suggested as a source of GABAergicinputs to the supraoptic nucleus, playing a role in theinhibition of vasopressin release following increased bloodpressure and blood volume (Grindstaff and Cunningham,2001). However, we observed here only occasional labeledaxons projecting to the supraoptic nucleus itself, confirmingprevious studies using PHAL deposits centered in the supra-optic perinuclear zone (Roland and Sawchenko, 1993). Instead,our PHAL data indicate that the pattern of axonal projections
from the LHAavr is exceptionally complex and involves anumber of different targets that fall into fairly obviousfunctional categories that suggest interesting possibilities forexperimental analysis.
Ganglion cells that innervate the LHAavr are locatedalmost exclusively in the superior temporal quadrant of theretina, and apparently correspond to a homogenous subset oftype III (also known as W) cells that, as discussed for ganglioncells projecting through the rhtm, change firing rate inresponse to altered luminance, but probably not to othertypes of visual stimuli (Leak and Moore, 1997). In addition tothe retina, the LHAavr receives particularly dense projectionsfrom the lateral septal nucleus (Risold and Swanson, 1997) andthe medial group of the anterior division of the bed nuclei ofthe stria terminalis (Dong and Swanson, 2006a,b), both ofwhich are clearly related to brain regions innervated by theLHAavr. In addition, the LHAavr is targeted by a number ofarousal-related neuron groups in the brainstem, including thepedunculopontine nucleus (Hallanger and Wainer, 1988),laterodorsal tegmental nucleus (Satoh and Fibiger, 1986),median raphe nucleus (Vertes et al., 1999), and locus ceruleus(ER Ribeiro-Barbosa and NS Canteras, personal observation).The LHAavr is thus uniquely positioned to integrate levels ofenvironmental luminescence with information from wake-related centers, and may therefore play an important role inarousal mechanisms.
Brain regions directly and substantially innervated by theLHAavr can be sorted, at least provisionally, into fourfunctional categories: goal-oriented behavior control, behav-ioral state control, viscera-related control, and thalamic loops.
4.3.1. Goal oriented-behavior controlTo sort out the different networks controlling goal-orientedbehaviors, Swanson (2000) developed the concept of a general“behavior control column” formed by distinct hypothalamiccircuitry involved in the control of the three basic classes ofgoal-oriented behaviors: ingestive (eating and drinking), repro-ductive (sexual and parental), and defensive (agonistic) beha-viors. Thus, the medial preoptic, anterior hypothalamic,ventromedial, and premammillary nuclei are interconnectedinahighlydifferentiatedway, and functional evidence indicatesthat they form critical parts of circuitry underlying theexpression of reproductive and defensive behaviors assuringsurvival of the species and the individual. In addition, a thirdcomponent of the behavior control column is the descendingdivision of the paraventricular nucleus (PVHd), dealing withingestive (eating and drinking) behaviors. The identification ofcircuitry involving the behavior control column is a usefulstarting point to understand how the hypothalamus deals witha wealth of sensory, hormonal, behavioral state, and cognitiveinputs, and organizes the different classes of goal-orientedmotor responses or behaviors (Swanson, 2000).
As we shall now see, the LHAavr generates substantialprojections to hypothalamic elements of the behavior controlcolumn, as well as telencephalic and lower brainstem sitesrelated to these elements. Thus, the LHAavr is positioned toexert important influences on the expression of a wide rangeof goal-oriented behaviors.
At rostral preoptic levels, the LHAavr projects densely tothemedian preoptic nucleus (MEPO). TheMEPO plays a central

178 B R A I N R E S E A R C H R E V I E W S 6 5 ( 2 0 1 1 ) 1 5 0 – 1 8 3
role in organizing drinking behavior. In addition to containingosmoreceptors (Denton et al., 1996; McKinley et al., 1996, 1998;Bourque and Oliet, 1997; Fitzsimons, 1998), the MEPO receivesinputs from catecholaminergic cell groups in the brainstemrelaying information related to cardiovascular volume recep-tors, and from the subfornical organ (Saper and Levisohn,1983; Swanson and Lind, 1986). MEPO influences on drinkingbehavior are likely mediated through its projections to theLHA and PVHd. The MEPO projects to the LHA region contain-ing two neuron populations (expressing melanin-concentrat-ing hormone and orexin/hypocretin) that is positioned tomediate the foraging behavior of animals actively seeking asource of water (Kelly andWatts, 1996). Conversely, the PVHd–another main target of the MEPO–is apparently related to theconsummatory phase of drinking behavior (for review, seeSwanson, 2000). The LHAavr may also influence drinkingbehavior through its projections to the magnocellular anddorsomedial nuclei of the anterior division of the BST, both ofwhich project densely to the MEPO and innervate other“humerosensory” brain regions related to drinking control—the subfornical organ and vascular organ of the laminaterminalis (Dong and Swanson, 2006a,b). It is interesting tospeculate that bright light reflected from a natural watersupply might preferentially influence (via the rhtl) the firingrate of LHAavr neurons that play a role in drinking behavior.
The LHAavr does not seem to innervate hypothalamicelements of the behavior control column related to feeding perse, but is able to influence brain regions critically related tofood-seeking behavior. The present results suggest that theLHAavr has a major influence on the lateral division of thePAG, either directly or indirectly, via dense projections to theanterior part of the ventromedial nucleus, a major source ofprojection to the lateral division of the PAG (Canteras et al.,1994). This PAG division seems to be a critical node in theneural system involved inmaking decisions between hunting,foraging, and other behavioral responses (Sukikara et al.,2006). Therefore, it probably represents an important link forthe LHAavr to influence the motivational drive to hunt andforage. Previous studies have shown that NMDA excitotoxiclesions of the lateral division of the PAG, but not other parts ofthe PAG, dramatically inhibit insect hunting, an effect thoughtto be mediated through its projections to the LHA regioncontaining melatonin-concentrating hormone and orexinneurons (Sukikara et al., 2006).
Our PHAL data reveal that the LHAavr provides sparse tomoderate projections to the medial preoptic, ventrolateral partof theventromedial, tuberal, andventral premammillarynuclei.They are a core part of the sexually dimorphic circuit thatcorresponds to the behavior control column division mediatingreproductive behaviors. In addition, the LHAavr projects ratherdensely to telencephalic regions including the medial andventrolateral zones of the rostral part of the lateral septalnucleus and the posterior amygdalar nucleus that generatemajor descending inputs to reproduction-related componentsof the behavior control column (Canteras et al., 1992; Risold andSwanson, 1997). All neuron groups of the reproduction-relatedbehavior control column express high levels of gonadalhormone receptors (Simerly et al., 1990), and have beenidentified as important sites for regulation of reproductivebehaviors, includingmale and female sexual behavior (Pfaff and
Sakuma, 1979a,b; Pfaus and Phillips, 1991; Coolen et al., 1996;Paredes et al., 1993, 1998; Hull et al., 2005; Hull and Dominguez,2006;García-Horsmanetal., 2008;Guarraci et al., 2008),maternalbehavior (for review, see Numan, 2006), and social agonisticbehaviors (Kollack-Walker and Newman, 1995; Kollack-Walkeret al., 1997, 1999; Motta et al., 2009).
Here it is important to note that the magnocellular,dorsomedial, and ventral nuclei of the anterior division ofthe BST, also targeted by the LHAavr, seem to correspond atleast in part to what Numanhas called the “ventral part” of theBST, which is apparently critical for the neural control ofmaternal behavior (see Numan, 2006). The LHAavr alsoprovides moderate inputs to the ventrolateral division of thePAG, a midbrain region targeted by elements of the reproduc-tive neural system that mediates the consummatory phase ofmale and female sexual behavior (Lonstein and Stern, 1998;Murphy and Hoffman, 2001), as well as the kyphosis postureassumed during nursing behavior (Lonstein and Stern, 1998).Taken together, the evidence suggests a role for the LHAavr inmodulating a wide array of reproductive responses, anhypothesis that requires experimentally analysis.
The LHAavr also projects to elements of the defensive(agonistic) behavior-related nuclei of the behavior controlcolumn: the anterior hypothalamic, dorsomedial part of theventromedial, and dorsal premammillary (PMd) nuclei (Swan-son, 2000; Canteras, 2002). Our PHAL data shows that theLHAavr projects densely to the PMd and provides relativelysparse inputs to the anterior hypothalamic nucleus as well.Notably, the projection to the PMd targets mostly its ventralregion, the region most responsive to predatory threats(Cezario et al., 2008; Motta et al., 2009). The PMd is the mostsensitive brain region responding to a predator or its cues, andlesions therein have been most effective in reducing anti-predator defensive responses (Canteras et al., 1997; Dielenberget al., 2001; Blanchard et al., 2003; Cezario et al., 2008). Thishypothalamic site appears to act as an amplifier for neuralprocessing in the medial hypothalamic defensive circuit—theinterconnections between defense-related nuclei of the be-havior control column.
The LHAavr also projects to brain sites that providesubstantial inputs to the medial hypothalamic defense circuit,including the rostral part of the lateral septal nucleus (Risoldand Swanson, 1997), transverse and interfascicular nuclei of theBST (Dong and Swanson, 2004), and juxtaparaventricular regionof the LHA (Hahn and Swanson, 2010), all of which are likely toexert key roles in the control of anti-predatory defensiveresponses. Moreover, the precommissural nucleus of the PAG,one of the main targets of the medial hypothalamic defensecircuit (Canteras and Goto, 1999), receives dense projectionsfromtheLHAavr.Considering theputative rolesof theLHAavr inthe context of anti-predator defensive responses, it seemsreasonable to suggest that different behavioral strategies mightbe expressed depending on environmental luminescence. Forexample, instead of flight behaviors, freezing immobility seemsto be particularly effective as a camouflage in darkness becausefreezing greatly reduces prey visibility and noise generation. Italso seems reasonable that this influence of the LHAavr on thecircuit underlying defensive responses at least partly mediatesthe well-documented anxiogenic profile induced by high levelsof environmental luminescence (for review, see Canteras, 2002).

179B R A I N R E S E A R C H R E V I E W S 6 5 ( 2 0 1 1 ) 1 5 0 – 1 8 3
4.3.2. Behavioral state controlSeveral brain structures innervated by the LHAavr playimportant roles in controlling circadian rhythms, sleep/wakecycle, and arousal level in the waking state. In sharp contrastto targets of the rhtm, the LHAavr provides only very sparseinputs to the SBPV, but projects densely to the dorsomedialhypothalamic nucleus, which, as discussed above, is heavilytargeted by the SCH and SBPV and is critical for maintainingcircadian rhythms of most functions (Chou et al., 2003).
Several brain regions involved in regulating the sleep/wakecycle and behavioral arousal are innervated by the LHAavr.Thus, the LHAavr provides dense inputs to the VLPO and to thejuxtadorsomedial region of the lateral hypothalamic area thatcontains the perifornical group of orexin neurons (Swansonet al., 2005). As previously discussed, the VLPO has beenimplicated in the regulation of both rapid-eye movement(REM) and non-REM sleep, and contains GABA/galaninergicneurons that are active during sleep (Sherin et al., 1996, 1998;Gaus et al., 2002; Lu et al., 2002). The perifornical group oforexin neurons shows diurnal changes consistent with a rolein the production or maintenance of arousal (Harris et al.,2005). In addition, the LHAavr provides sparse to moderateprojections to the posterior hypothalamic and supramammil-lary nuclei, which play an important role in modulatinghippocampal theta rhythm, which in turn is present duringexploratory behavior and the rapid eye movement stage ofsleep (see Vertes and Kocsis, 1997). The LHAavr also projects tothe dendritic and core region of the locus ceruleus, a majorwakefulness-promoting nucleus (Aston-Jones and Cohen,2005).
4.3.3. Viscera-related controlThe LHAavr provides substantial projections to a number ofbrain sites involved in visceromotor control, including themedial group of the anterior division of the BST (particularlythe dorsomedial and magnocellular nuclei), MEPO, dorsome-dial hypothalamic nucleus, ventrolateral division of the PAG,and Barrington's nucleus. The medial group of the anteriordivision of the BST has been shown to project to a numbersites related to the central autonomic control network in-cluding the central amygdalar nucleus, BST anterolateralgroup, descending paraventricular hypothalamic nucleus,retrochiasmatic area, ventrolateral PAG, and Barrington'snucleus (Dong and Swanson, 2006a,b). The MEPO and thedorsomedial hypothalamic nucleus are integral elements ofthe hypothalamic visceromotor pattern generator, a complexnetwork that is in a position to coordinate complex autonomicand neuroendocrine responses (Thompson and Swanson,2003). The ventrolateral division of the PAG has extensiveprojections to medullary “cardiorespiratory” regions thatprovide major sources of excitatory or inhibitory drive onsympathetic vasomotor tone, and contain also the dorsal andventral respiratory neuron groups (Van Bockstaele et al., 1991;Cameron et al., 1995; Farkas et al., 1997, 1998). Therefore,stimulation in this region produces a variety of autonomiceffects including changes in respiratory rate and cardiovas-cular-related responses (see Carrive and Bandler, 1991;Behbehani, 1995). Barrington's nucleus projects to lumbosa-cral regions of the spinal cord that innervate parasympatheticcomponents of the pelvic nerves to the urinary bladder, distal
colon, and genitals (Satoh et al., 1978; Loewy et al., 1979;Pavcovich et al., 1998; Valentino et al., 2000; Blok and Holstege,1996), and is critical for controlling micturition, defecation,and penile erection (see Dong and Swanson, 2006b).
Interestingly, we have also found that the LHAavr providesa moderately dense plexus of fibers that tends to center in theexternal lateral part of the parabrachial nucleus, which isknown to be involved in conveying general visceral sensoryinformation associated with cardiovascular, respiratory, andnociceptive inputs (Herbert et al., 1990). Like the LHAavr, thispart of the parabrachial nucleus also receives direct inputsfrom the retina (Fite and Janusonis, 2002).
4.3.4. Thalamic loopsThe present results indicate that the LHAavr projects substan-tially to the paraventricular thalamic nucleus and nucleusreunions. These two nuclei generate somewhat complemen-tary projection patterns to the telencephalon, where thenucleus reuniens provides substantial inputs to a number ofcortical fields (anterior piriform, medial and ventral orbital,anterior cingulate, prelimbic, infralimbic, insular, and peri-rhinal areas, aswell as field CA1, dorsal and ventral subiculum,parasubiculum, and entorhinal area of the hippocampalformation), whereas the paraventricular thalamic nucleusprojects especially densely to the nucleus accumbens (coreand shell), the central, posterior basolateral, and posteriorbasolateral amygdalar nuclei, and the lateral group of theanterior division of the BST (Vertes et al., 2006; Vertes andHoover, 2008).
4.4. Overview
Perhaps themost basic conclusion tobedrawn fromthepresentwork is that axonal outputs of the three major direct targets ofthe rhtm–the ventrolateral SCH, perisuprachiasmatic region,andAHNvm–arequite similar andquitedistinct fromtheaxonaloutputs of the direct major target of the rhtl—the LHAavr.Specifically, targets of the rhtm have a rather limited set ofprojections that, based on the current literature, are primarilyinvolved in the control of circadian rhythms. Conversely, themajor targetof the rhtl doesnot seemtobedirectly related to thecontrol of circadian rhythms. Instead, theLHHavr is inapositionto combine inputs from the retina about current environmentalluminance levels and from brainstem nuclei about currentarousal state, and then send axonal projections to neuralnetworks that control a wide range of goal-oriented behaviors(especially drinking, reproductive, and defensive) along withadaptively appropriate and complementary visceral responsesand adjustments to behavioral state.
Acknowledgments
The research described here was funded by the Fundação deAmparo à Pesquisa do Estado de São Paulo: FAPESP - 05/59286-4 (to N.S.C) and 09/52920-0 (to JCN); Conselho Nacional deDesenvolvimento Científico e Tecnologico (to N.S.C. and JCN);FAPESP fellowships (to ERRB and MG); and the NationalInstitutes of Health grant R01NS16686 (to L.W.S).

180 B R A I N R E S E A R C H R E V I E W S 6 5 ( 2 0 1 1 ) 1 5 0 – 1 8 3
R E F E R E N C E S
Abrahamson, E.E., Moore, R.Y., 2001. Suprachiasmatic nucleus inthe mouse: retinal innervation, intrinsic organization andefferent projections. Brain Res. 916, 172–191.
Antle, M.C., Silver, R., 2005. Orchestrating time: arrangements ofthe brain circadian clock. Trends Neurosci. 28, 145–151.
Arvanitogiannis, A., Robinson, B., Beaulé, C., Amir, S., 2000.Calbindin-D28k immunoreactivity in the suprachiasmaticnucleus and the circadian response to constant light in the rat.Neuroscience 99, 397–401.
Aston-Jones, G., Cohen, J.D., 2005. Adaptive gain and the role of thelocus coeruleus–norepinephrine system in optimalperformance. J. Comp. Neurol. 493, 99–110.
Behbehani, M.M., 1995. Functional characteristics of the midbrainperiaqueductal gray. Prog. Neurobiol. 46, 575–605.
Blanchard, D.C., Li, C.I., Hubbard, D., Markham, C.M., Yang, M.,Takahashi, L.K., Blanchard, R.J., 2003. Dorsal premammillarynucleus differentially modulates defensive behaviors induceddifferent threat stimuli in rats. Neurosci. Lett. 345, 145–148.
Blok, B.F., Holstege, G., 1996. The neuronal control of micturitionand its relation to the emotional motor system. Prog. Brain Res.107, 113–126.
Bourque, C.W., Oliet, S.H., 1997. Osmoreceptors in the centralnervous system. Annu. Rev. Physiol. 59, 601–619.
Buijs, R.M., Markman, M., Nunes-Cardoso, B., Hou, Y.X., Shinn, S.,1993. Projections of the suprachiasmatic nucleus tostress-related areas in the rat hypothalamus: a light andelectron microscopic study. J. Comp. Neurol. 335, 42–54.
Buijs, R.M., Hou, Y.X., Shinn, S., Renaud, L.P., 1994. Ultrastructuralevidence for intra- and extranuclear projections of GABAergicneurons of the suprachiasmatic nucleus. J. Comp. Neurol. 340,381–391.
Cameron, A.A., Khan, I.A., Westlund, K.N., Willis, W.D., 1995. Theefferent projections of the periaqueductal gray in the rat: aPhaseolus vulgarisleucoagglutinin study. II. Descendingprojections. J. Comp. Neurol. 351, 585–601.
Canteras, N.S., 2002. The medial hypothalamic defensive system:hodological organization and functional implications.Pharmacol. Biochem. Behav. 71, 481–491.
Canteras, N.S., Goto, M., 1999. Connections of the precommissuralnucleus. J. Comp. Neurol. 408, 23–45.
Canteras, N.S., Swanson, L.W., 1992. Projections of the ventralsubiculum to the amygdala, septum, and hypothalamus: aPHAL anterograde tract-tracing study in the rat. J. Comp.Neurol. 324, 180–194.
Canteras, N.S., Simerly, R.B., Swanson, L.W., 1992. Connections ofthe posterior nucleus of the amygdala. J. Comp. Neurol. 324,143–179.
Canteras, N.S., Simerly, R.B., Swanson, L.W., 1994. Organization ofprojections from the ventromedial nucleus of thehypothalamus: a Phaseolus vulgaris-leucoagglutinin study inthe rat. J. Comp. Neurol. 348, 41–79.
Canteras, N.S., Simerly, R.B., Swanson, L.W., 1995. Organization ofprojections from the medial nucleus of the amygdala: a PHALstudy in the rat. J. Comp. Neurol. 360, 213–245.
Canteras, N.S., Chiavegatto, S., Valle, L.E., Swanson, L.W., 1997.Severe reduction of rat defensive behavior to a predator bydiscrete hypothalamic chemical lesions. Brain Res. Bull. 44,297–305.
Card, J.P., Moore, R.Y., 1984. The suprachiasmatic nucleus of thegolden hamster: immunohistochemical analysis of cell andfiber distribution. Neuroscience 13, 415–431.
Card, J.P., Moore, R.Y., 1989. Organization of lateralgeniculate-hypothalamic connections in the rat. J. Comp.Neurol. 284, 135–147.
Card, J.P., Brecha, N., Karten, H.J., Moore, R.Y., 1981.Immunocytochemical localization of vasoactive intestinal
polypeptide-containing cells and processes in thesuprachiasmatic nucleus of the rat: light and electronmicroscopic analysis. J. Neurosci. 1, 1289–1303.
Carrive, P., Bandler, R., 1991. Viscerotopic organization of neuronssubserving hypotensive reactions within the midbrainperiaqueductal grey: a correlative functional and anatomicalstudy. Brain Res. 541, 206–215.
Cezario, A.F., Ribeiro-Barbosa, E.R., Baldo, M.V., Canteras, N.S.,2008. Hypothalamic sites responding to predator threats—therole of the dorsal premammillary nucleus in unconditionedand conditioned antipredatory defensive behavior. Eur. J.Neurosci. 28, 1003–1015.
Cheng, M.Y., Bullock, C.M., Li, C., Lee, A.G., Bermak, J.C., Belluzzi, J.,Weaver, D.R., Leslie, F.M., Zhou, Q.Y., 2002. Prokineticin 2transmits the behavioural circadian rhythm of thesuprachiasmatic nucleus. Nature 417, 405–410.
Chou, T.C., Bjorkum, A.A., Gaus, S.E., Lu, J., Scammell, T.E., Saper,C.B., 2002. Afferents to the ventrolateral preoptic nucleus. J.Neurosci. 22, 977–990.
Chou, T.C., Scammell, T.E., Gooley, J.J., Gaus, S.E., Saper, C.B., Lu, J.,2003. Critical role of dorsomedial hypothalamic nucleus in awide range of behavioral circadian rhythms. J. Neurosci. 23,10691–10702.
Comoli, E., Ribeiro-Barbosa, E.R., Canteras, N.S., 2000. Afferentconnections of the dorsal premammillary nucleus. J. Comp.Neurol. 423, 83–98.
Conrad, C.D., Stumpf, W.E., 1975. Endocrine-optic pathways to thehypothalamus. Anat. Neuroendocrinology, Int. Conf.Neurobiology of CNS-Hormone Interactions, Chapel Hill 1974.Karger, Basel, pp. 62–68.
Coolen, L.M., Peters, H.J., Veening, J.G., 1996. Fos immunoreactivityin the rat brain following consummatory elements of sexualbehavior: a sex comparison. Brain Res. 738, 67–82.
Costa, M.S., Santee, U.R., Cavalcante, J.S., Moraes, P.R., Santos, N.P.,Britto, L.R., 1999. Retinohypothalamic projections in thecommon marmoset (Callithrix jacchus): a study using choleratoxin subunit B. J. Comp. Neurol. 415, 393–403.
Damiola, F., Le Minh, N., Preitner, N., Kornmann, B., Fleury-Olela,F., Schibler, U., 2000. Restricted feeding uncouples circadianoscillators in peripheral tissues from the central pacemaker inthe suprachiasmatic nucleus. Genes Dev. 14, 2950–2961.
Denton, D.A., McKinley, M.J., Weisinger, R.S., 1996. Hypothalamicintegration of body fluid regulation. Proc. Natl Acad. Sci. USA93, 7397–7404.
Deurveilher, S., Semba, K., 2005. Indirect projections from thesuprachiasmatic nucleus to major arousal-promoting cellgroups in rat: implications for the circadian control ofbehavioural state. Neuroscience 130, 165–183.
Dielenberg, R.A., Hunt, G.E., McGregor, I.S., 2001. “When a ratsmells a cat”: the distribution of c-fos expression in rat brainfollowing exposure to a predator odor. Neuroscience 104,1085–1097.
Dong, H.W., Swanson, L.W., 2004. Projections from bed nuclei ofthe stria terminalis, posterior division: implications forcerebral hemisphere regulation of defensive and reproductivebehaviors. J. Comp. Neurol. 471, 396–433.
Dong, H.W., Swanson, L.W., 2006a. Projections from bed nuclei ofthe stria terminalis, dorsomedial nucleus: implications forcerebral hemisphere integration of neuroendocrine,autonomic, and drinking responses. J. Comp. Neurol. 494,75–107.
Dong, H.W., Swanson, L.W., 2006b. Projections from bed nuclei ofthe stria terminalis, magnocellular nucleus: implications forcerebral hemisphere regulation of micturition, defecation, andpenile erection. J. Comp. Neurol. 494, 108–141.
Edgar, D.K., Martin, C.E., Dement, W.C., 1991. Activity feedbackto the mammalian circadian pacemaker: influence onobservedmeasures of rhythm period length. J. Biol. Rhythms 6,185–199.

181B R A I N R E S E A R C H R E V I E W S 6 5 ( 2 0 1 1 ) 1 5 0 – 1 8 3
Farkas, E., Jansen, A.S., Loewy, A.D., 1997. Periaqueductal graymatter projection to vagal preganglionic neurons and thenucleus tractus solitarius. Brain Res. 764, 257–261.
Farkas, E., Jansen, A.S., Loewy, A.D., 1998. Periaqueductal graymatter input to cardiac-related sympathetic premotorneurons. Brain Res. 792, 179–192.
Fite, K.V., Janusonis, S., 2002. Optic afferents to the parabrachialnucleus. Brain Res. 943, 9–14.
Fitzsimons, J.T., 1998. Angiotensin, thirst, and sodium appetite.Physiol. Ver. 78, 583–686.
García-Horsman, S.P., Agmo, A., Paredes, R.G., 2008. Infusions ofnaloxone into the medial preoptic area, ventromedial nucleusof the hypothalamus, and amygdala block conditioned placepreference induced by paced mating behavior. Horm. Behav.54, 709–716.
Gaus, S.E., Strecker, R.E., Tate, B.A., Parker, R.A., Saper, C.B., 2002.Ventrolateral preoptic nucleus contains sleep-active,galaninergic neurons in multiple mammalian species.Neuroscience 115, 285–294.
Gerfen, C.R., Sawchenko, P.E., 1984. An anterogradeneuroanatomical tracing method that shows the detailedmorphology of neurons, their axons and terminals:immunohistochemical localization of an axonally transportedplant lectin, Phaseolus vulgaris leucoagglutinin (PHA-L). BrainRes. 290, 219–238.
Grindstaff, R.R., Cunningham, J.T., 2001. Cardiovascular regulationof vasopressin neurons in the supraoptic nucleus. Exp. Neurol.171, 219–226.
Guarraci, F.A., Frohardt, R.J., Hines, D., Navaira, E., Smith, J.,Wampler, L., 2008. Intracranial infusions of amphetamine intothe medial preoptic area but not the nucleus accumbens affectpaced mating behavior in female rats. Pharmacol. Biochem.Behav. 89, 253–262.
Hahn, J.D., Swanson, L.W., 2010. Distinct patterns of neural inputsand outputs of the juxtaparaventricular andsuprafornical regions of the lateral hypothalamic area in themale rat. Brain Res. Rev. 64, 14–103.
Hallanger, A.E., Wainer, B.H., 1988. Ascending projections from thepedunculopontine tegmental nucleus and the adjacentmesopontine tegmentum in the rat. J. Comp. Neurol. 274,483–515.
Hamada, T., LeSauter, J., Venuti, J.M., Silver, R., 2001. Expression ofPeriod genes: rhythmic and nonrhythmic compartments of thesuprachiasmatic nucleus pacemaker. J. Neurosci. 21, 7742–7750.
Hamada, T., Antle, M.C., Silver, R., 2004. Temporal and spatialexpression patterns of canonical clock genes andclock-controlled genes in the suprachiasmatic nucleus. Eur. J.Neurosci. 19, 1741–1748.
Harris, G.C., Wimmer, M., Aston-Jones, G., 2005. A role for lateralhypothalamic orexin neurons in reward seeking. Nature 437,556–559.
Hendrickson, A.E., Wagoner, N., Cowan, W.M., 1972. Anautoradiographic and electron microscopic study ofretino-hypothalamic connections. Z. Zellforsch. Mikrosk. Anat.135, 1–26.
Herbert, H., Moga, M.M., Saper, C.B., 1990. Connections of theparabrachial nucleus with the nucleus of the solitary tract andthe medullary reticular formation in the rat. J. Comp. Neurol.293, 540–580.
Hull, E.M., Dominguez, J.M., 2006. Getting his act together: roles ofglutamate, nitric oxide, and dopamine in the medial preopticarea. Brain Res. 1126, 66–75.
Hull, E.M., Wood, R.I., McKenna, K.E., 2005. The neurobiology ofmale sexual behavior, In: Neill, J.D. (Ed.), Knobil and Neill'sPhysiology of Reproduction, third ed. Academic Press, SanDiego, New York, pp. 1729–1824.
Jacobowitz, D.M., Winsky, L., 1991. Immunocytochemicallocalization of calretinin in the forebrain of the rat. J. Comp.Neurol. 304, 198–218.
Johnson, R.F., Morin, L.P., Moore, R.Y., 1988. Retinohypothalamicprojections in the hamster and rat demonstrated using choleratoxin. Brain Res. 462, 301–312.
Kalsbeek, A., Teclemariam-Mesbah, R., Pevet, P., 1993. Efferentprojections of the suprachiasmatic nucleus in the goldenhamster (Mesocricetus auratus). J. Comp. Neurol. 332, 293–314.
Kelly, A.B., Watts, A.G., 1996. The mediation ofdehydration-induced peptidergic gene expression in the ratlateral hypothalamic area by forebrain afferent projections.J. Comp. Neurol. 370, 231–246.
Kollack-Walker, S., Newman, S.W., 1995. Mating and agonisticbehavior produce different patterns of Fos immunolabeling inthe male Syrian hamster brain. Neuroscience 66, 721–736.
Kollack-Walker, S., Watson, S.J., Akil, H., 1997. Social stress inhamsters: defeat activates specific neurocircuits within thebrain. J. Neurosci. 17, 8842–8855.
Kollack-Walker, S., Don, C., Watson, S.J., Akil, H., 1999. Differentialexpression of c-fos mRNA within neurocircuits of malehamsters exposed to acute or chronic defeat.J. Neuroendocrinol. 11, 547–559.
Kramer, A., Yang, F.C., Snodgrass, P., Li, X., Scammell, T.E., Davis,F.C.,Weitz, C.J., 2001. Regulation of daily locomotor activity andsleep by hypothalamic EGF receptor signaling. Science 294,2511–2515.
Kraves, S., Weitz, C.J., 2006. A role for cardiotrophin-like cytokinein the circadian control of mammalian locomotor activity. Nat.Neurosci. 9, 212–219.
Krieger, D.T., 1980. Ventromedial hypothalamic lesions abolishfood-shifted circadian adrenal and temperature rhythmicity.Endocrinology 106, 649–654.
Krieger, D.T., Hauser, H., Krey, L.C., 1977. Suprachiasmatic nuclearlesions do not abolish food-shifted circadian adrenal andtemperature rhythmicity. Science 197, 398–399.
Kriegsfeld, L.J., Leak, R.K., Yackulic, C.B., LeSauter, J., Silver, R.,2004. Organization of suprachiasmatic nucleus projections inSyrian hamsters (Mesocricetus auratus): an anterograde andretrograde analysis. J. Comp. Neurol. 468, 361–379.
Landry, G.J., Yamakawa, G.R., Mistlberger, R.E., 2007. Robust foodanticipatory circadian rhythms in ratswith complete ablation ofthe thalamic paraventricular nucleus. Brain Res. 1141, 108–118.
Larsen, P.J., Enquist, L.W., Card, J.P., 1998. Characterization of themultisynaptic neuronal control of the rat pineal gland usingviral transneuronal tracing. Eur. J. Neurosci. 10, 128–145.
Leak, R.K., Moore, R.Y., 1997. Identification of retinal ganglion cellsprojecting to the lateral hypothalamic area of the rat. Brain Res.770, 105–114.
Leak, R.K., Moore, R.Y., 2001. Topographic organization ofsuprachiasmatic nucleus projection neurons. J. Comp. Neurol.433, 312–334.
Leak, R.K., Card, J.P., Moore, R.Y., 1999. Suprachiasmaticpacemaker organization analyzed by viral transynaptic transport. Brain Res. 819, 23–32.
Levine, J.D., Weiss, M.L., Rosenwasser, A.M., Miselis, R.R., 1991.Retinohypothalamic tract in the female albino rat: a studyusing horseradish peroxidase conjugated to cholera toxin.J. Comp. Neurol. 306, 344–360.
Loewy, A.D., Saper, C.B., Baker, R.P., 1979. Descending projectionsfrom the pontine micturition center. Brain Res. 172, 533–538.
Lonstein, J.S., Stern, J.M., 1998. Site and behavioral specificity ofperiaqueductal gray lesions on postpartum sexual, maternal,and aggressive behaviors in rats. Brain Res. 804, 21–35.
Lu, J., Greco, M.A., Shiromani, P., Saper, C.B., 2000. Effect of lesionsof the ventrolateral preoptic nucleus on NREM and REM sleep.J. Neurosci. 20, 3830–3842.
Lu, J., Zhang, Y.H., Chou, T.C., Gaus, S.E., Elmquist, J.K., Shiromani,P., Saper, C.B., 2001. Contrasting effects of ibotenate lesions ofthe paraventricular nucleus and subparaventricular zone onsleep–wake cycle and temperature regulation. J. Neurosci. 21,4864–4874.

182 B R A I N R E S E A R C H R E V I E W S 6 5 ( 2 0 1 1 ) 1 5 0 – 1 8 3
Lu, J., Bjorkum, A.A., Xu, M., Gaus, S.E., Shiromani, P.J., Saper, C.B.,2002. Selective activation of the extended ventrolateralpreoptic nucleus during rapid eyemovement sleep. J. Neurosci.22, 4568–4576.
Mai, J.K., 1979. Distribution of retinal axons within the lateralhypothalamic area. Exp. Brain Res. 34, 373–377.
McKinley, M.J., Pennington, G.L., Oldfield, B.J., 1996. Anteroventralwall of the third ventricle and dorsal lamina terminalis:headquarters for control of body fluid homeostasis? Clin. Exp.Pharmacol. Physiol. 23, 271–281.
McKinley, M.J., Allen, A.M., Burns, P., Colvill, L.M., Oldfield, B.J.,1998. Interaction of circulating hormones with the brain: theroles of the subfornical organ and the organum vasculosum ofthe lamina terminalis. Clin. Exp. Pharmacol. Physiol. Suppl. 25,S61–S67.
Meyer-Berstein, E.L., Jetton, A.E., Matsumoto, S.I., Markuns, J.F.,Lehman, M.N., Bittman, E.L., 1999. Effects of suprachiasmatictransplants on circadian rhythms of neuroendocrine functionin golden hamsters. Endocrinology 140, 207–218.
Mikkelsen, J.D., Vrang, N., 1994. A direct pretectosuprachiasmaticprojection in the rat. Neuroscience 62, 497–505.
Mikkelsen, J.D., Larsen, P.J., O'Hare, M.M., Wiegand, S.J., 1991.Gastrin releasing peptide in the rat suprachiasmatic nucleus:an immunohistochemical, chromatographic andradioimmunological study. Neuroscience 40, 55–66.
Mistlberger, R.E., 1994. Circadian food-anticipatory activity: formalmodels and physiological mechanisms. Neurosci. Biobehav.Rev. 18, 171–195.
Mistlberger, R.E., 2009. Food-anticipatory circadian rhythms:concepts and methods. Eur. J. Neurosci. 30, 1718–1729.
Moga, M.M., Moore, R.Y., 1997. Organization of neural inputs to thesuprachiasmatic nucleus in the rat. J. Comp. Neurol. 389,508–534.
Moga, M.M., Moore, R.Y., 2000. Paraventricular thalamus lesionsalter circadian period and activity distribution in the blindedrat. Biol. Rhythm Res. 31, 212–219.
Moore, R.Y., Eichler, V.B., 1972. Loss of a circadian adrenalcorticosterone rhythm following suprachiasmatic lesions inthe rat. Brain Res. 42, 201–206.
Moore, R.Y., Lenn, N.J., 1972. A retinohypothalamic projection inthe rat. J. Comp. Neurol. 146, 1–14.
Moore, R.Y., Halaris, A.E., Jones, B.E., 1978. Serotonin neurons ofthe midbrain raphe: ascending projections. J. Comp. Neurol.180, 417–438.
Moore, R.Y., Speh, J.C., Card, J.P., 1995. The retinohypothalamictract originates from a distinct subset of retinal ganglion cells.J. Comp. Neurol. 352, 351–366.
Moore, R.Y., Speh, J.C., Leak, R.K., 2002. Suprachiasmatic nucleusorganization. Cell Tissue Res. 309, 89–98.
Morin, L.P., Goodless-Sanchez, N., Smale, L., Moore, R.Y., 1994.Projections of the suprachiasmatic nuclei, subparaventricularzone and retrochiasmatic area in the golden hamster.Neuroscience 61, 391–410.
Morin, L.P., Shivers, K.Y., Blanchard, J.H., Muscat, L., 2006. Complexorganization of mouse and rat suprachiasmatic nucleus.Neuroscience 137, 1285–1297.
Mota, S.R., Canteras, N.S., Bartol, I., Skorupa, A.L., Scialfa, J.H.,Terra, I.M., Afeche, S.C., Cipolla-Neto, J., 2001. Lesions of thedorsomedial hypothalamic nucleus do not influence the dailyprofile of pineal metabolism in rats. Neuroendocrinology 73,123–128.
Mota-Ortiz, S.R., Sukikara, M.H., Felicio, L.F., Canteras, N.S., 2009.Afferent connections to the rostrolateral part of theperiaqueductal gray: a critical region influencing the motivationdrive to hunt and forage. Neural Plast. 2009, 612698.
Motta, S.C., Goto, M., Gouveia, F.V., Baldo, M.V., Canteras, N.S.,Swanson, L.W., 2009. Dissecting the brain's fear system revealsthe hypothalamus is critical for responding in subordinateconspecific intruders. Proc. Natl Acad. Sci. USA 106, 4870–4875.
Munch, I.C., Moller, M., Larsen, P.J., Vrang, N., 2002. Light-inducedc-Fos expression in suprachiasmatic nuclei neurons targetingthe paraventricular nucleus of the hamster hypothalamus:phase dependence and immunochemical identification.J. Comp. Neurol. 442, 48–62.
Murakami, D.M., Miller, J.D., Fuller, C.A., 1989. Theretinohypothalamic tract in the cat: retinal ganglion cellmorphology and pattern of projection. Brain Res. 482, 283–296.
Murphy, A.Z., Hoffman, G.E., 2001. Distribution of gonadal steroidreceptor-containing neurons in the preoptic-periaqueductalgray-brainstem pathway: a potential circuit for the initiation ofmale sexual behavior. J. Comp. Neurol. 438, 191–212.
Novak, C.M., Nunez, A.A., 2000. A sparse projection from thesuprachiasmatic nucleus to the sleep-active ventrolateralpreoptic area in the rat. NeuroReport 11, 93–96.
Numan, M., 2006. Hypothalamic neural circuits regulatingmaternal responsiveness toward infants. Behav. Cogn.Neurosci. Rev. 5, 163–190.
Nunez, A.A., Stephan, F.K., 1977. The effects of hypothalamic knifecuts on drinking rhythms and the estrous cycle of the rat.Behav. Biol. 20, 224–234.
Paredes, R.G., Highland, L., Karam, P., 1993. Socio-sexual behaviorin male rats after lesions of the medial preoptic area: evidencefor reduced sexual motivation. Brain Res. 618, 271–276.
Paredes, R.G., Tzschentke, T., Nakach, N., 1998. Lesions of themedial preoptic area anterior hypothalamus (MPOA/AH)modify partner preference in male rats. Brain Res. 813, 1–8.
Pavcovich, L.A., Yang, M., Miselis, R.R., Valentino, R.J., 1998. Novelrole for the pontine micturition center, Barrington's nucleus:evidence for coordination of colonic and forebrain activity.Brain Res. 784, 355–361.
Pfaff, D.W., Sakuma, Y., 1979a. Facilitation of the lordosis reflex offemale rats from the ventromedial nucleus of thehypothalamus. J. Physiol. 288, 189–202.
Pfaff, D.W., Sakuma, Y., 1979b. Deficit in the lordosis reflex offemale rats caused by lesions in the ventromedial nucleus ofthe hypothalamus. J. Physiol. 288, 203–210.
Pfaus, J.G., Phillips, A.G., 1991. Role of dopamine in anticipatoryand consummatory aspects of sexual behavior in the male rat.Behav. Neurosci. 105, 727–743.
Plano, S.A., Golombek, D.A., Chiesa, J.J., 2010. Circadianentrainment to light–dark cycles involves extracellular nitricoxide communication within the suprachiasmatic nuclei. Eur.J. Neurosci. 31, 876–882.
Reppert, S.M.,Weaver, D.R., 2001. Molecular analysis ofmammaliancircadian rhythms. Annu. Rev. Physiol. 63, 647–676.
Risold, P.Y., Swanson, L.W., 1997. Connections of the rat lateralseptal complex. Brain Res. Brain Res. Rev. 24, 115–195.
Risold, P.Y., Canteras, N.S., Swanson, L.W., 1994. Organization ofprojections from the anterior hypothalamic nucleus: aPhaseolus vulgaris-leucoagglutinin study in the rat. J. Comp.Neurol. 348, 1–40.
Roland, B.L., Sawchenko, P.E., 1993. Local origins of someGABAergic projections to the paraventricular and supraopticnuclei of the hypothalamus in the rat. J. Comp. Neurol. 332,123–143.
Romijn, H.J., Sluiter, A.A., Pool, C.W., Wortel, J., Buijs, R.M., 1996.Differences in colocalization between Fos and PHI, GRP, VIPand VP in neurons of the rat suprachiasmatic nucleus after alight stimulus during the phase delay versus the phaseadvance period of the night. J. Comp. Neurol. 372, 1–8.
Rusak, B., Zucker, I., 1979. Neural regulation of circadian rhythms.Physiol. Rev. 59, 449–526.
Salazar-Juarez, A., Escobar, C., Aguilar-Roblero, R., 2002. Anteriorparaventricular thalamus modulates light-induced phaseshifts in circadian rhythmicity in rats. Am. J. Physiol. Regul.Integr. Comp. Physiol. 283, R897–R904.
Saper, C.B., Levisohn, D., 1983. Afferent connections of the medianpreoptic nucleus in the rat: anatomical evidence for a

183B R A I N R E S E A R C H R E V I E W S 6 5 ( 2 0 1 1 ) 1 5 0 – 1 8 3
cardiovascular integrative mechanism in the anteroventralthird ventricular (AV3V) region. Brain Res. 288, 21–31.
Saper, C.B., Lu, J., Chou, T.C., Gooley, J., 2005. The hypothalamicintegrator for circadian rhythms. Trends Neurosci. 28, 152–157.
Satoh, K., Fibiger, H.C., 1986. Cholinergic neurons of thelaterodorsal tegmental nucleus: efferent and afferentconnections. J. Comp. Neurol. 253, 277–302.
Satoh, K., Shimizu, M., Tohyama, M., Maeda, T., 1978. Localizationof the micturition reflex center at dorsolateral pontinetegmentum of the rat. Neurosci. Lett. 8, 27–33.
Sherin, J.E., Shiromani, P.J., McCarley, R.W., Saper, C.B., 1996.Activation of ventrolateral preoptic neurons during sleep.Science 271, 216–219.
Sherin, J.E., Elmquist, J.K., Torrealba, F., Saper, C.B., 1998.Innervation of histaminergic tuberomammillary neurons byGABAergic and galaninergic neurons in the ventrolateralpreoptic nucleus of the rat. J. Neurosci. 18, 4705–4721.
Silver, R., LeSauter, J., Tresco, P.A., Lehman, M.N., 1996a. Adiffusible coupling signal from the transplantedsuprachiasmatic nucleus controlling circadian locomotorrhythms. Nature 382, 810–813.
Silver, R., Romero, M.T., Besmer, H.R., Leak, R., Nunez, J.M.,LeSauter, J., 1996b. Calbindin-D28K cells in the hamster SCNexpress light-induced Fos. NeuroReport 7, 1224–1228.
Simerly, R.B., Chang, C., Muramatsu, M., Swanson, L.W., 1990.Distribution of androgen and estrogen receptormRNA-containing cells in the rat brain: an in situ hybridizationstudy. J. Comp. Neurol. 294, 76–95.
Speh, J.C., Moore, R.Y., 1993. Retinohypothalamic tractdevelopment in the hamster and rat. Brain Res. Dev. Brain Res.76, 171–181.
Stephan, F.K., 2002. The “other” circadian system: food as aZeitgeber. J. Biol. Rhythms 17, 284–292.
Stephan, F.K., Zucker, I., 1972. Circadian rhythms in drinkingbehavior and locomotor activity of rats are eliminated byhypothalamic lesions. Proc. Natl Acad. Sci. USA 69, 1583–1586.
Stokkan, K.A., Yamazaki, S., Tei, H., Sakaki, Y., Menaker, M., 2001.Entrainment of the circadian clock in the liver by feeding.Science 291, 490–493.
Stopa, E.G., Minamitani, N., Jonassen, J.A., King, J.C., Wolfe, H.,Mobtaker, H., Albers, H.E., 1988. Localization of vasoactiveintestinal peptide and peptide histidine isoleucineimmunoreactivity and mRNA within the rat suprachiasmaticnucleus. Brain Res. 464, 319–325.
Sukikara, M.H., Mota-Ortiz, S.R., Baldo, M.V., Felicio, L.F., Canteras,N.S., 2006. A role for the periaqueductal gray in switchingadaptive behavioral responses. J. Neurosci. 26, 2583–2589.
Swanson, L.W., 1976. An autoradiographic study of the efferentconnections of the preoptic region in the rat. J. Comp. Neurol.167, 227–256.
Swanson, L.W., 2000. Cerebral hemisphere regulation ofmotivatedbehavior. Brain Res. 886, 113–164.
Swanson, L.W., 2004. Brain Maps: Structure of the Rat Brain, 3rdedition. Academic Press, pp. 1–215.
Swanson, L.W., Cowan, W.M., 1975. The efferent connections ofthe suprachiasmatic nucleus of the hypothalamus. J. Comp.Neurol. 160, 1–12.
Swanson, L.W., Lind, R.W., 1986. Neural projections subservingthe initiation of a specific motivated behavior in the rat:new projections from the subfornical organ. Brain Res. 379,399–403.
Swanson, L.W., Cowan, W.M., Jones, E.G., 1974. Anautoradiographic study of the efferent connections of theventral lateral geniculate nucleus in the albino rat and the cat.J. Comp. Neurol. 156, 143–163.
Swanson, L.W., Sanchez-Watts, G., Watts, A.G., 2005. Comparisonof melanin-concentrating hormone and hypocretin/orexinmRNA expression patterns in a new parceling scheme of thelateral hypothalamic zone. Neurosci. Lett. 387, 80–84.
Teclemariam-Mesbah, R., Kalsbeek, A., Pevet, P., Buijs, R.M., 1997.Direct vasoactive intestinal polypeptide-containing projectionfrom the suprachiasmatic nucleus to spinal projectinghypothalamic paraventricular neurons. Brain Res. 748, 71–76.
Thompson, R.H., Swanson, L.W., 1998. Organization of inputs tothe dorsomedial nucleus of the hypothalamus: areexamination with Fluorogold and PHAL in the rat. Brain Res.Brain Res. Rev. 27, 89–118.
Thompson, R.H., Swanson, L.W., 2003. Structural characterizationof a hypothalamic visceromotor pattern generator network.Brain Res. Brain Res. Rev. 41, 153–202.
Thompson, R.H., Canteras, N.S., Swanson, L.W., 1996.Organization of projections from the dorsomedial nucleus ofthe hypothalamus: a PHA-L study in the rat. J. Comp. Neurol.376, 143–173.
Valentino, R.J., Page, M.E., Luppi, P.H., Zhu, Y., Van Bockstaele, E.,Aston-Jones, G., 1994. Evidence for widespread afferents toBarrington's nucleus, a brainstem region rich in corticotrophinreleasing hormone neurons. Neuroscience 62, 125–143.
Valentino, R.J., Kosboth, M., Colflesh, M., Miselis, R.R., 2000.Transneuronal labeling from the rat distal colon: anatomicevidence for regulation of distal colon function by a pontinecorticotropin-releasing factor system. J. Comp. Neurol. 417,399–414.
Van Bockstaele, E.J., Aston-Jones, G., Pieribone, V.A., Ennis, M.,Shipley, M.T., 1991. Subregions of the periaqueductal graytopographically innervate the rostral ventral medulla in therat. J. Comp. Neurol. 309, 305–327.
van den Pol, A.N., Tsujimoto, K.L., 1985. Neurotransmitters of thehypothalamic suprachiasmatic nucleus: immunocytochemicalanalysis of 25 neuronal antigens. Neuroscience 15, 1049–1086.
Vandesande, F., Dierickx, K., De Mey, J., 1975. Identification ofseparate vasopressin-neurophysin II and oxytocin-neurophysinI containing nerve fibres in the external region of the bovinemedian eminence. Cell Tissue Res. 158, 509–516.
Vertes, R.P., Hoover, W.B., 2008. Projections of the paraventricularand paratenial nuclei of the dorsal midline thalamus in the rat.J. Comp. Neurol. 508, 212–237.
Vertes, R.P., Kocsis, B., 1997. Brainstem-diencephalo-septohippocampal systems controlling the theta rhythm of thehippocampus. Neuroscience 81, 893–926.
Vertes, R.P., Crane, A.M., Colom, L.V., Bland, B.H., 1995. Ascendingprojections of the posterior nucleus of the hypothalamus:PHA-L analysis in the rat. J. Comp. Neurol. 359, 90–116.
Vertes, R.P., Fortin, W.J., Crane, A.M., 1999. Projections of themedian raphe nucleus in the rat. J. Comp. Neurol. 407, 555–582.
Vertes, R.P., Hoover,W.B., DoValle, A.C., Sherman, A., Rodriguez, J.J.,2006. Efferent projections of reuniens and rhomboidnuclei of thethalamus in the rat. J. Comp. Neurol. 499, 768–796.
Vrang, N., Larsen, P.J., Mikkelsen, J.D., 1995. Direct projection fromthe suprachiasmatic nucleus to hypophysiotrophiccorticotropin-releasing factor immunoreactive cells in theparaventricular nucleus of the hypothalamus demonstrated bymeans of Phaseolus vulgarisleucoagglutinin tract tracing. BrainRes. 684, 61–69.
Watts, A.G., Swanson, L.W., 1987. Efferent projections of thesuprachiasmatic nucleus: II. Studies using retrogradetransport of fluorescent dyes and simultaneous peptideimmunohistochemistry in the rat. J. Comp. Neurol. 258, 230–252.
Watts, A.G., Swanson, L.W., Sanchez-Watts, G., 1987. Efferentprojections of the suprachiasmatic nucleus: I. Studies usinganterograde transport of Phaseolus vulgaris leucoagglutinin inthe rat. J. Comp. Neurol. 258, 204–229.
Yan, L., Okamura, H., 2002. Gradients in the circadian expressionof Per1 and Per2 genes in the rat suprachiasmatic nucleus. Eur.J. Neurosci. 15, 1153–1162.
Yan, L., Silver, R., 2002. Differential induction and localization ofmPer1 and mPer2 during advancing and delaying phase shifts.Eur. J. Neurosci. 16, 1531–1540.
Copyright © 2022 FDOKUMEN




















