
พฤติกรรมที่เป็นปัญหาของสังคม Problematic Behavior of Society
Upload
independentCategory
view
7download
0
The problematic systematic position of Ceratostratiotes Gregor(Hydrocharitaceae ?) – morphological, anatomical
and biochemical comparison with Stratiotes L.
B. Meller1 and P. F. van Bergen2
1University of Vienna, Palaeontological Institute, Geocentre, Vienna, Austria2Geochemistry, Faculty of Earth Sciences, Utrecht University, Utrecht, The Netherlands
Received April 18, 2002; accepted October 18, 2002Published online: February 4, 2003� Springer-Verlag 2003
Abstract. The enigmatic fossil taxon Ceratostrati-otes sinjanus (Kerner) Buzek has been considered aseither a Hydrocharitaceae seed (monocot) or aCeratophyllaceae fruit (dicot). The co-occurrenceof seeds of Stratiotes kaltennordheimensis (Zenker)Keilhack (Hydrocharitaceae) and Ceratostratiotesin Early Miocene sediments in Langau (LowerAustria) enabled comparisons of morphological-anatomical features and lignin compositions, inde-pendent of diagenetic biases.The biochemistry of the Ceratostratiotes seed coatwall is not monocotyledon-like and is dissimilar tothat of the co-occurring Stratiotes testae. In con-trast, the Ceratostratiotes seed coat anatomyresembles that of Hydrocharitaceae genera withthe micropyle and raphe being very similar to thosein seeds of Stratiotes. Although the horizontalarrangement of the Ceratostratiotes spines isdifferent from the longitudinal arrangement of thesurface ridges in Stratiotes kaltennordheimensis andthe spines in Blyxa (Hydrocharitaceae), the spinestructures are very similar. A mixture of similaritiesand differences characterizes the morphological-anatomical features. Thus the systematic affinity ofCeratostratiotes still remains enigmatic.
Key words: Palaeobotany, seed morphology, seedanatomy, flash pyrolysis, Ceratostratiotes sinjanus,Stratiotes kaltennordheimensis.
Introduction
Ceratostratiotes Gregor, a genus of controver-sial systematic position, was established fromfossil propagules in Early Miocene sedimentsof the Langau area in Lower Austria byGregor (1980). The fossil remains were regard-ed as seeds of an aquatic plant based on thespiny appendages, the morphological similar-ities to the Hydrocharitaceae, and the associ-ated fruit and seed assemblages (Gregor 1980).The preserved morphological features havebeen also interpreted as characteristic ofCeratophyllum fruits (e.g. Buzek 1982).The Ceratostratiotes propagules from Lan-
gau were initially identified as Trapa fruits byBerger (1957: Trapa zapfei n. sp.), because oftheir spiny appendages. However, Trapa fruitsare readily distinguished from Ceratostratiotesby their broad obovate globose shape withstrong and thick horn-like appendages termi-nating in thin anchor-like apices, their apicalneck with the germination pore and theirsmooth surface. According to Gregor (1980:108), Ceratostratiotes should be regarded as afossil Hydrocharitaceae genus, or a near rela-tive, because of its general morphological sim-
Plant Syst. Evol. 236: 125–150 (2003)DOI 10.1007/s00606-002-0242-9

ilarities (e.g. dehiscence suture, two symmetricalvalves, dorsal raphe furrow) to Stratiotes eventhough it differs by the spiny appendages and aslightly different surface structure of the seedcoat. Gregor (1980) had compared the spinyappendages with those occurring in the Hy-drocharitaceae genera Blyxa Noronha ex Tho-uars, EnhalusRich. and Thalassia Banks et Sol.ex C.Koenig. In a monographic study of extantand fossil Stratiotes seeds, Cook and Urmi-Konig (1983) followed Gregor and includedCeratostratiotes within the Hydrocharitaceaeand also noted the similarities to the seeds of thegenusBlyxa. The comparisons with the modernStratiotes seeds are complicated by the fact thatthe genus is monotypic today and so there is nopossibility to see modern interspecies varia-tions.In 1982, Buzek identified Ceratostratiotes
diaspores from Podmoli, near Znojmo, inSouth Moravia (Czech Republic). Moreover,he discovered that Ceratostratiotes had alreadybeen described as Ceratophyllum sinjanusKerner from Croatia (Kerner 1905a). Contraryto Gregor’s (1980) results, Buzek regardedthem as a fossil representative of the Cerato-phyllaceae following Kerner (1905a, b, 1916).Les (1988, 1989), who investigated the originand affinities of the Ceratophyllaceae, alsotreated Ceratostratiotes as a fossil genus ofthat family. Anatomical investigations of thefruit wall of Ceratophyllum species have beenstarted and show differences in the fruit wallanatomy of Ceratophyllum demersum L. andCeratophyllum submersum L., as well as differ-ences to the seed coat anatomy of Cerato-stratiotes. However, more Ceratophyllummaterial and species have to be studied beforesystematic conclusions can be made.Up to now, the diaspores have been found in
only a few basins in EarlyMiocene sediments inthe area surrounding the former Central Para-tethys. These are the: Langau area, LowerAustria; Podmoli near Znojmo, S Moravia,Czech Republic; Sinj area NNE Split, Croatia(Kerner 1905b, 1916, 1917; Buzek 1982); Kimi -Aliveri basin, Evia island, Greece. Due to theslightly different morphology of the seeds from
Croatia versus those fromAustria and theCzechRepublic, Buzek (1982) distinguished twoforms: C. sinjanus f. sinjana and C. sinjanus f.zapfei, although accepting that these might onlyrepresent morphological variations caused bydifferent ecological conditions. Gunther andGregor (1989: 88, 1997: 49) regarded the Aus-trian and Greek specimens as different species.It is necessary to investigate more specimensfrom the Czech, Croatian and Greek localitiesto determine whether the specimens from thesouthern and those from the northern localitiesrepresent different species or ecological varietiesof the same species. Fossil species discrimina-tion relies on themorphology and anatomy, butthe already known morphological variability ofseeds and fruits from aquatic plants (Cook1987) complicates this issue.Biochemical investigations of fossil seed
coats of different aquatic plant genera (e.g.Stratiotes, Nymphaea, Nuphar, Potamogeton)have demonstrated that the biochemicalcomposition, despite differences in localitiesand ages, shows characteristic signals oflignin molecules (van Bergen 2001; vanBergen et al. 1994a, b, 1995, 1999). Thus,such investigations can provide a comple-mentary tool, in addition to morphologicaland anatomical comparisons, to determinetaxonomic relationships. An importantadvantage of the current comparative inves-tigation is the co-occurrence of Ceratostra-tiotes and Stratiotes in Langau, whichreduces the influence of different diageneticprocesses on the seed coat morphology,anatomy and biochemistry. In contrast toCeratostratiotes, the morphological preserva-tion of the Stratiotes remains is poorer butits biochemical composition as induced bythe earlier investigations is highly characte-ristic (van Bergen et al. 1994a, b).In this paper, the results of comparative
morphological, anatomical and biochemicalinvestigations of Ceratostratiotes and Strati-otes are presented as a first step to elucidate thesystematic position of Ceratostratiotes. Cera-tophyllum is not included in this investigationbecause of its absence in the Langau area and
126 B. Meller and P. F. van Bergen: Anatomical and biochemical investigations of Ceratostratiotes

the absence of biochemical data of the fruitwall of modern and fossil fruits.
Material
The Ceratostratiotes diaspores studied are from thearea of Langau in Austria, Podmoli near Znojmoin the Czech Republic and Sinj in Croatia (Fig. 1).The records from the Kimi – Aliveri basin (Greece)are currently under investigation by Gregor (pers.comm. 2001). The biochemical investigations havebeen carried out using Ceratostratiotes and Strati-otes propagules from the Langau sample B41/17.The material studied is listed in Table 1.
Geological setting of the Langau area: Langauis situated in the eastern part of the Austrianmolasse foreland basin, at the eastern margin ofthe Bohemian Massif. Brackish – marine to limnic– fluvial sediments were deposited during the
Eggenburgian to Ottnangian (Langau and Rie-gersburg Formation; Early Miocene; 17–19 m. a.).Lignite-producing environments also occurredduring that time (Roetzel 1994, Roetzel and Fuchs1994). The diaspores were found in both brownclayey micaceous silts to sands of the seampartings and in mollusc rich grey-green clayeysands of the basal layers of the Langau Forma-tion. They are often associated with Salvinia andAzolla megaspores, sometimes with seeds ofStratiotes kaltennordheimensis (Zenker) Keilhackand Typha sp., rarely with diaspores from Deco-don sp., Nymphaea sp., Cyperaceae gen. et sp.indet., Umbelliferopsis sp., Vitaceae gen. et sp.indet., Toddalia naviculaeformis (Reid) Gregor velT. latisiliquata (Ludwig) Gregor, Carpolithus haf-niensis Hartz (¼ Carpolithus mettenii (Unger)Mai); according to Gregor (pers. comm. 2001)these diaspores belong to the Boraginaceae genusCordia L.). In addition, Gregor (1980) has foundMyrica ceriferiformoides Buzek & Holy, Myricaboveyana (Heer) Chandler, Chionanthus korniiGregor and Spirematospermum wetzleri (Heer)Chandler. The pollen and spores from this sitewere studied by Draxler (1991), who distinguishedmore than 100 taxa, mainly representatives ofdifferent wetland vegetation types. Pollen grainsfrom Hydrocharitaceae or Ceratophyllum had notbeen discovered in samples from the Langau area.But these are often lacking in the fossil pollenrecord or cannot be recognized (pers. comm.Draxler 2002).
The sample Langau B41/17 originates from adrill core and was taken by Roetzel (1993) (Aus-trian Geological Survey, Vienna) during mappingin that area. Additional material from this layerwas sampled by Daxner-Hock (Natural HistoryMuseum, Vienna) prospecting for vertebrates.Several samples from different layers of that drillcore yielded Ceratostratiotes diaspores (e.g. B41/4)as well as the other numerous bulk samples, alsotaken by Roetzel from that area. The sampleLangau LC5 derives from drilling by the AustrianMineral Oil Company (OMV) in 1981 also provid-ed by Roetzel. Gregor (1980) re-investigated thematerial published by Berger (1957) and numerousnew specimens collected from the heaps at theformer mines in Langau.
The diaspore preservation from the differentlocalities in the area of Langau, as well as from theCzech and Croatian localities differs considerably;
Fig. 1. Geographic position of the investigated local-ities (underlined) yielding seeds of Ceratostratiotessinjanus (Kerner) Buzek
B. Meller and P. F. van Bergen: Anatomical and biochemical investigations of Ceratostratiotes 127

Table1.Listofthestudiedmaterial(Abbreviations:NMA=NatureMuseumAugsburg,Germany;CUP-GP=CharlesUniversityPrague,
departmentofgeologyandpalaeontology,CzechRepublic;NMNHP=NationalMuseumofNaturalHistoryPrague,palaeobotanicalcol-
lection,CzechRepublic;NHMW=NaturalHistoryMuseumVienna,Austria;IPUW=
InstituteofPalaeontology,UniversityofVienna,
Austria;GBA=AustrianGeologicalSurvey;OMV=AustrianMineralOilCompany,Austria;BSGPM=BavarianStateCollectionof
GeologyandPalaeontologyMunich,Germany;leg.=sampledby)
Taxa
(originallabel)
Locality
Sample
number
Area,
country
Collection
Kindof
sampling
Sediment
Formation,
Paratethysstage,age
Stratiotes
kaltennordheimensis
Ceratostratiotes
sinjanus
Langau
B41/17
Lower
Austria
IPUWno.3114
drilling(GBA,
leg.Roetzel)
green-grey
sandyclay
silttosilty
clay
LangauFm.,
Eggenburgian-
Ottnangian
EarlyMiocene
Ceratostratiotes
sinjanus
Langau
B41/4
Lower
Austria
IPUWno.3113
drilling(GBA,
leg.Roetzel)
claysiltto
siltyclay
LangauFm.,
Eggenburgian-
Ottnangian
EarlyMiocene
Ceratostratiotes
sinjanus
Langau
LC5:
9.8–9.7m
Lower
Austria
IPUWno.3112
drilling(OMV)
sandwith
crystalline
components
LangauFm.,
Eggenburgian-
Ottnangian
EarlyMiocene
Ceratostratiotes
sinjanus
(Ceratostratiotes
zapfei)
Langau
Lower
Austria
BSGPMno.
2002I7-14;
NMAno.1384
looseblocks
(leg.Gregor)
siltymicaceous
clayrock
?LangauFm.,
Eggenburgian-
Ottnangian
EarlyMiocene
Ceratostratiotes
sinjanus
(Ceratostratiotes
zapfei)
Langau
Lower
Austria
NHMW
1966/674,675
Pb453-469
(leg.Zapfe)
reddish-brown
grey,silty
micaceous
clayrock
LangauFm.,
Eggenburgian-
Ottnangian
EarlyMiocene
Ceratostratiotes
sinjanus
Podmoli
Z-103:
15.1–16m
near
Znojmo,S
Moravia,
Czech
Republic
CUP-GP;
NMNHP-
G7905-7906
drilling
(leg.Ctyroky)
greysiltyclay
EarlyMiocene,
presumably
synchronouswith
Langau(from
Buzek1982)
Ceratostratiotes
sinjanus
Stupalo
St
Sladoja
nearSinj,
typearea;
Croatia
NMNHP6/93,
K392-395;
(K394partly
dissolved)
outcrop(leg.
Buzek)
strongsilty
marlstone
Miocene;probably
‘‘Ceratophyllumschichten’’,
accordingKerner
(1914,1916)
(fromBuzek1982)
Ceratostratiotes
sinjanus
(Ceratostratiotes
sinjanuszapfei)
Sinjsko
polje
Glavica
nearHana;
nearSinj,
typearea;
Croatia
NMAno.1580
outcrop
(leg.Jurisic-
Polsack)
siltymarlstone
EarlyMiocene;?
Eggenburgian-
Ottnangian;type
areaofKerner
(accordingtothe
label)
128 B. Meller and P. F. van Bergen: Anatomical and biochemical investigations of Ceratostratiotes

preservation may vary between specimens from thesame locality or even within one sample. Highquality morphological preservation does not ne-cessarily correlate with well-preserved anatomicalstructures and vice versa. Sample Langau B41/17includes numerous fragments, often isolated valvesand several nearly complete diaspores of Cerato-stratiotes, with at least 50 specimens in total. Theyare mainly secondarily compressed (laterally, dor-sal-ventrally or oblique) and flattened, appearingthin-walled with very shallow pits (compressionand/ or abrasion) and their surface is brown andshiny. Some fragments appear less flattened, with arough surface and a grey-green colour probablydue to the content of iron-sulphide. The spines arebroken off or totally missing, exposing either theiranatomical structure or a black shiny wall. Strati-otes is documented by fragments only, which arenot secondarily flattened. Sample Langau B41/4also yielded numerous fragmentary Ceratostrati-otes specimens, sometimes isolated valves, rarelynearly complete specimens. The outer wall surfacesappear often abraded and the seed coat is thin.Sample Langau LC5: 9.8–9.7 m yielded a similarnumber of specimens, often isolated valves orfragments. In general these are not flattened,appearing thicker-walled and the pits are muchdeeper. Stratiotes remains have not been found inthis sample. Altogether, the anatomical structuresare sometimes nicely preserved. The specimens ofGregor’s samples from the spoil heaps are alsopreserved in 3-D, with a well-preserved wallstructure. However, some diaspores are smoothwithout any sculpturing or ornamentation butpitted, already exposing the anatomical structure,due to mechanical abrasion or biological degrada-tion. All samples include specimens where the innersurface is covered by a thin translucent skinny-likelayer but no isolated complete tegmens wereobserved.
The Ceratostratiotes specimens from Langaufigured by Berger (1957: Fig. 1A–M) are stillincorporated within small pieces of sediment. Theirseed coats show ruptures and cracks and a few ofthe figured specimens are less complete than inBerger’s drawings. Anatomical structures are sel-dom observable but the inner thin translucent layeris sometimes preserved. Beside the specimensfigured by Berger (1957), there are additional seedswithin the sediment pieces which had been partlyfigured by Gregor (1980).
The Podmoli locality is situated near Znojmo,in the Czech Republic, near the Austrian borderand is part of the same depositional system as theLangau area (Buzek 1982). The sample Z103: 15.1–16 m originates from a drill core. The museum inPrague houses a few pieces of sediment from whicha few nearly complete valves and several fragmentswere extracted for this investigation.
The diaspore-bearing sediments from Stupalonear Sladoje and Glavica near Hana (Sinjarea¼ Sinjsko polje, Dalmatia, Croatia) are marlysandstones or silty sandy marls. The sediments areregarded as limnic – fluvial freshwater deposits ofMiocene age. An Early Miocene, probably Ottnan-gian, age of the layers with Ceratostratiotes fromthe Sinj area is indirectly dated by the partlyoverlying mollusc-bearing layers and by compari-sons with vertebrate-bearing sites. The dating is apreliminary result of the present extensive researchproject covering the geology and palaeontology ofthe Miocene freshwater basins of Croatia (pers.comm. Jurisic-Polsack 2001; Croatian NaturalHistory Museum, Zagreb).
The surrounding of Sinj, 20 km NE Split,represents the type area of Ceratostratiotes sinjanus(Kerner) Buzek. An exact type locality was notmentioned by Kerner (1905a) and the figureexplanations lack the name of the exact locality.Due to the absence of Kerner’s material ofCeratophyllum sinjanum from the Sinj area, Buzekchose a Neotype from that material, which he hadcollected in Stupalo near Sladoja. It is stored in thepalaeontological collection of the Czech GeologicalSurvey in Prague but had not been available duringthis investigation due to the re-organisation of theSurvey. Small pieces of sediment from Stupalo withdiaspores are housed in the collection of theNational Museum of Natural History in Prague.The collection of the Nature Museum Augsburg(Germany) includes several diaspores and one pieceof sediment from Glavica near Hana, sampled byJurisic-Polsack.
Methods
Sample preparation for morphological and anatomi-
cal analyses. The diaspore-bearing sediments fromLangau were dried and disaggregated in a weak (3–4 %) H2O2 solution, then washed and sieved (sievemesh diameters 5 mm, 2.5 mm, 1 mm, 0.5 mm).The residual sediment was studied with a binocular
B. Meller and P. F. van Bergen: Anatomical and biochemical investigations of Ceratostratiotes 129

microscope for seeds and fruits. The photos(negatives) and digital images were taken with aWild-Leitz binocular microscope (M420, LZ 12)and a Nikon digital camera. For the SEM-studies(Jeol scanning electron microscope), the diasporeswere cleaned with HCl and HF to remove mineraldeposits, mounted on SEM-stubs with a stickycarbon tape or plastic conductive carbon cement,coated with a thin, about 50 nm thick, layer of goldand studied with 10 kV. Negatives were alsodigitalized and all digital images were manipulatedin contrast and brightness. A small piece ofsediment from Podmoli was also treated with adiluted H2O2 solution and one small piece of marlysediment from Stupalo was partly dissolved withHCl only.
Sample preparation for biochemical analyses.
Single sclerotic valves of Ceratostratiotes andStratiotes sclerotic valve fragments, extracted fromsample Langau B41/17 with H2O2, were crushedand extracted ultrasonically using dichlorometh-ane and methanol (1:1 v/v; 5x) to remove solublesolvent compounds. Based on the colouration ofthe extract alone, few soluble compounds werepresent in either of the two taxa studied. Extractswere removed and the residues were dried. Thesepowdered residues were studied using flash pyrol-ysis-gas chromatography (Py-GC) and Py-GC-mass spectrometry (Py-GC-MS) to determine thechemical composition of the resistant walls of thediaspores. Some sclerotic parts may have con-tained non-sclerotic tissue but its contribution interms of total mass would have been negligible.Each taxon was pyrolysed at least 3 times todetermine chemical variation within the samples.
Py-GC and Py-GC-MS. One technique torapidly obtain detailed molecular information onsmall quantities of modern and fossil plant materialis using flash pyrolysis (For additional informationabout this technique and its application for chemo-systematics the reader is referred to Stankiewiczet al. 1997, 1998; van Bergen 1999, 2001; vanBergen et al. 1999). Flash pyrolysis-GC was per-formed using a Curie-point FOM-3LX unit forpyrolysis connected to a Hewlett Packard 5890 gaschromatograph. The samples were pressed onto aferromagnetic wire with a Curie temperature of610�C. The pyrolysis time was 10 sec. The pyrolysisinterface and injector temperature were set at200�C. The GC (equipped with a cryogenic unit)was programmed from 0�C (5 min) to 300�C
(10 min) at a rate of 3�C min)1. Separation wasachieved using a fused-silica capillary column(25 m · 0.32 mm) coated with CP Sil-5 (film thick-ness 0.4 lm). Helium was used as the carrier gas.Compounds were detected using a flame ionizationdetector (FID) set at 325�C.
Py-GC-MS was performed also using anidentical pyrolysis unit attached to a HP 5890 GCcoupled to a VG Autospec Ultima mass spectrom-eter. The temperature programme and generalconditions were the same as for the Py-GCanalyses. The VG Autospec Ultima operated at70eV with a mass range m/z 45-800 and a cycle timeof 1.7 sec. Compound identifications were based onmass spectral data and retention time comparisonswith reference samples and data reported in theliterature (van Bergen et al. 1994b, 1995, 1996,1999).
Terminology. The terminology used in thedescription and results treats the Ceratostratiotesdiaspores as seeds for two reasons: their morpho-logical-anatomical similarity to Stratiotes seeds andthe necessity to describe the features with unam-biguous terms.
Results
Morphology and anatomy of Ceratostratiotessinjanus from the Langau area. (samples Lan-gau B41/17, B41/4, LC5: 9.7–9.8 m, andGregor’s (1980) material).Description: The seeds are of obovate
shape with a wide (rarely narrow) roundedapex and a narrow rounded base (Figs. 5, 7–8)and are composed of two symmetrical valves(Figs. 3, 35–36). The base of the seeds isslightly thickened (Figs. 3–4, 39–40) and bearstwo spines, one on each valve (Figs. 3, 35). Theseed lengths (without spines) vary from4–6 mm, rarely 8 mm (without spines), theirwidths are 2.5–3 mm, rarely 4 mm. The outermorphology is generally characterized by 0.5–1(?) cm long spines positioned rarely in two(Fig. 6), generally in three planes (Figs. 2–4,12): basal spines (Figs. 2–4, 7), lateral spines inthe upper part (Figs. 3–5) and main spines inthe middle part of the seed (Figs. 2–3, 7, 12,17, 20, 25). The spines are generally broken offor appear abraded. Their complete length is
130 B. Meller and P. F. van Bergen: Anatomical and biochemical investigations of Ceratostratiotes

never preserved. The proximal part of the mainspines resembles fish fins (dorsal margin cor-responding to the fish back) and is horizontally
flattened. These two main spines (one fromeach valve) are fused in their proximal part(Fig. 3; compare also the specimen from
2 3 4 5 9
lsplsp
lsp
lsplsp
lsp
lsp
lsp
msp
msp
msp
bsp
bsp
bsp
bsp bspbsp
Figs. 2–11. Outer morphology of Ceratostratiotes sinjanus (Kerner) Buzek (Figs. 2–18) and Stratioteskaltennordheimensis (Zenker) Keilhack (Figs. 9–10) from Langau studied with the LM (Figs. 2–5, 9) and theSEM (Figs. 6–8, 10–11); scales always 1 mm if not otherwise inscribed; (r¼ raphe furrow, msp¼main spine,bsp¼ basal spine, lsp¼ lateral spine). Fig. 2. Lateral view on a seed valve surface; the main spine is nearlytotally preserved, the basal and lateral spines are preserved with the proximal part only (IPUW 3112/02).Figs. 3–4. Dorsal (3) and ventral (4) view on a seed, which is slightly compressed in longitudinal direction. Themain spines are located at the dorsal margin in the lower part of the seed due to the compression; the basalspines are more fragmentary. Fig. 4 shows the wing-like structure in the upper part of the seed built up by thefused lateral spine bases (IPUW 3113/06). Fig. 5. Seed valve with distinct rows of pits. The main and basalspines are broken off, one of the lateral spines is curved downwards (IPUW 3112/23). Fig. 6. Elongate obovateseed valve; note that the main spine is in the upper third part of the valve in the same level as the lateral spines(BSGPM 2002 I 4). Fig. 7. Broad obovate seed valve; the spines are located in the middle part of the seed; allspines are broken off (IPUW 3112/12). Fig. 8. Small spineless seed valve (IPUW 3112/11). Fig. 9. Fragmentaryseed valve of Stratiotes kaltennordheimensis; note the slightly curved basal micropylar end, the prominent keel-like dorsal (left) margin, and the longitudinal ridges (IPUW 3114/16). Fig. 10. The most complete Stratiotesseed valve with distinct longitudinal surface ridges (IPUW 3114/10). Fig. 11. Detail of the seed surface with thepits and ridges (upper left part of Fig. 10). (Figs. 2–76. Numbers and symbols in parentheses represent thecollection inventory number; explanations of the abbreviations see Table 1; LM¼ binocular microscope;SEM¼ scanning electron microscope; other abbreviations are explained in the explanations of Figs. 2–11 andFigs. 12–19)
B. Meller and P. F. van Bergen: Anatomical and biochemical investigations of Ceratostratiotes 131

Croatia, Figs. 65–67). The three lateral spinesin the upper part of the seed are mainlylongitudinally flattened, straight or curved invarying directions; their proximal parts aresometimes fused, forming a wing-like structure(Fig. 4). A few specimens lack all the spines(Fig. 8), others have only short rounded spinesor lack the lateral spines.The seed surface sculpturing is built up by
distinct pits (Figs. 6–8, 41–42), arranged in
nearly regular rows in the middle lateral partof the seed (Fig. 6), but irregularly arrangedaround the appendages, near the valve marginsand near the apex and base (Fig. 7). Themargins between the pits are mainly rounded(Fig. 42), sharp andwith thin edges in the apicalor marginal part (Figs. 7, 41) of the valves andare built up by additional cells (Fig. 46). Thediameter of the pits is about 100–300 lm, theirdepths vary between 100 and 150 lm.
12 13 15 18
16
14
17 19
r
r
r
r
rr
r
r
r
r
r
r
r
r
r
r
r
r
r
rmsp
msp
msp
m m
msp
msp
bsp
bsp
m
lsp
lsp lsp
Figs. 12–19. Inner morphology of Ceratostratiotes sinjanus (Kerner) Buzek (Figs. 12–17) and Stratioteskaltennordheimensis (Zenker) Keilhack (Figs. 18–19) from Langau studied with the LM; scales 1 mm if nototherwise inscribed; (r¼ raphe furrow, m¼micropyle). Figs. 12–14. Same seed valve as in Fig. 2; the raphefurrow is indicated by the pale sediment filling. Fig. 13 shows a detail of the left apical part, Fig. 14 displays theraphe furrow in the main spine and in the lower part of the dorsal seed wall (IPUW 3112/02). Fig. 15. Seedvalve exposing a long raphe in the dorsal wall (IPUW 3112/03). Fig. 16. Seed valve fragment with a distinctdeep and long raphe furrow in the dorsal wall (details see Figs. 29–31) (IPUW 3112/24). Fig. 17. Seed valvewith a long raphe furrow, which is also visible in the basal seed part (details see Figs. 25–28) (IPUW 3112/05).Fig. 18. Fragment of the upper part of a Stratiotes seed valve with a distinct raphe furrow in the thickeneddorsal wall, running from the inner testa margin to the outer testa margin (IPUW 3114/17). Fig. 19. Fragmentof the lower part of a seed valve (the same as in Fig. 9) with the raphe furrow at the thickened dorsal wall. Theouter part of the dorsal wall is preserved in the upper part but broken off in the lower one, producing anapparently marginal position of the raphe furrow. Below the lowermost raphe arrow traces of the furrow arenot visible (IPUW 3114/16)
132 B. Meller and P. F. van Bergen: Anatomical and biochemical investigations of Ceratostratiotes

The micropyle is indicated at the seedsurface in the slightly thickened supra-basalventral part of the base by a very small deepen-ing (Figs. 39–40) or distinct hole which is rarelyobservable (Gregor 1980: Pl. 2, Fig. 3). Theinner valve shows the flat micropylar furrowrunning from the locule straight through theseed coat in a supra-basal position at an angle tothe main seed axis (Figs. 21, 26).The seed coatwall is often slightly thicker in this area.In the middle part to upper third of the
dorsal wall margin, where the main spineoriginates, the seed coat is distinctly thickened
and forms a triangular extension of varyingsize (Figs. 12, 14, 16–17, 20, 25, 29). Thisextension forms the proximal part of the mainspine in the area, where the spine of each valveis still attached to the corresponding one of theother valve (see above). The dorsal seed wallis generally thicker than the ventral one(Figs. 12, 16–17, 20, 25) forming a weak keel-like structure along the dorsal seed margin(Figs. 3, 35–36).The raphe furrow leaves the locule sub-
apical at the dorsal side (Figs. 12–13, 22, 28),bends immediately downwards and runs within
Figs. 20–24. Inner morphology of a narrow seed valve of Ceratostratiotes sinjanus (Kerner) Buzek fromLangau; scales 1 mm if not otherwise inscribed; SEM (IPUW 3112/16). Fig. 20. Seed valve with well preservedinner morphology; the raphe furrow is visible at the dorsal wall in the plane of dehiscence and symmetrybetween apex and base; the main and basal spines are partly broken off. Fig. 21. Basal part of the seed exposingclearly the raphe furrow in the dorsal wall and the furrow of the micropyle in the ventral wall. Fig. 22. Dorsal-apical seed part with the distinct raphe furrow. Fig. 23. The base of the main spine with the curved raphefurrow. Fig. 24. The distinct raphe furrow in the lower part of the seed
B. Meller and P. F. van Bergen: Anatomical and biochemical investigations of Ceratostratiotes 133

134 B. Meller and P. F. van Bergen: Anatomical and biochemical investigations of Ceratostratiotes

the seed coat wall in the plane of dehiscenceinto the triangular extension (= proximal partof the main spine), then turns down sometimes(Figs. 14, 16, 23, 25, 27, 29, 30). Below the mainspine, a furrow is clearly observable in somespecimens (Figs. 14, 17, 20–21, 25, 26, 29, 30).Often a furrow has not been clearly observed inthe lower part of the seed valve and the raphefurrow appears to finish in the main spine. Onefragment of a basal seed part shows the hilumfurrow along the outer dorsal margin in thelower part of the seed; the valves gape there(Figs. 36–38). Above the gap, the dorsalmargin is built like a rounded ridge or weakkeel. Other seed fragments show also a distinctfurrow between the dorsal keel-like margins ofthe valves (Fig. 35).The cross-section of the 100–250 lm thick
seed coat displays a spongy structure(Figs. 46–47, 49) with numerous large holesand thickened pitted stone cells arranged inirregular cell layers. The pits represent theopenings of the tunnels passing through thecell walls. The spines are also built up by pittedstone cells (Figs. 48, 50–51) which are distinct-ly aligned. The stone cell walls of the basalspines are thicker (Fig. 51) than those of thelateral spines (Figs. 48, 50).The inner surface of the seed coat is
covered by remains of a thin translucent layerof a shiny gold-colour (compare also Gregor1980: Pl. 1, Fig. 7). A fine longitudinal stria-tion is often visible, also with the SEM(Figs. 20, 25, 54–55). Below that surface,narrow rectangular longitudinal elongatedsometimes pitted cells build the innermost seed
coat layer (endotesta ?; Fig. 55). One fragmen-tary specimen bears longitudinal rows of flattubercle-like warts (Figs. 56–57) on a smooththin surface (tegmen or nucellus cuticle). Thewarts are 1–2 lm in diameter.
Stratiotes kaltennordheimensis from Lan-
gau (sample Langau B41/17). The specimensare all fragmentary (Figs. 9, 18–19, 32, 43, 44).The most complete specimen is one single seedvalve (Fig. 10) with an apparently well pre-served surface (Fig. 11) and a poorly preservedinner morphology. Nevertheless the fragmentsshow all the characteristic features as describedfor that species (Cook and Urmi-Konig 1983;Holy and Buzek 1966; Knobloch 1989; Pala-marev 1979; Shaffer-Fehre 1991a, b; vanBergen et al. 1994a, b): outer surface sculp-tured by longitudinal ridges (Figs. 9–10), testasurface built by small pits (Figs. 11, 45), seedbase neck-like and slightly curved (Figs. 9–10,43), testa thick-walled (Figs. 18–19), raphefurrow deep and distinct running along thethickened dorsal valve wall (building a keel-like extension) in the plane of dehiscence andsymmetry (Figs. 18–19, 32), micropyle situatedsupra-basal in the ventrally curved neck-likebase (Fig. 44), testa built up by irregularlyarranged layers of stone cells (Fig. 52), stonecells thick-walled and irregularly pitted(Fig. 53). Remains of the mucilaginous cellsas described from well preserved Stratiotesseed surfaces (Hooker et al. 1995, van Bergenet al. 1995) have not been found on theLangau specimens.The morphology and anatomy of modern
and fossil Stratiotes seeds has already been
Figs. 25–34. Comparison of the inner morphology of Ceratostratiotes sinjanus (Kerner) Buzek (Figs. 25–31)and Stratiotes kaltennordheimensis (Zenker) Keilhack (Figs. 32–34) from Langau studied with the SEM; scales1 mm if not otherwise inscribed. Figs. 25–28. Same seed valve as in Fig. 17 after cleaning with HCl and HF(IPUW 3112/05). Fig. 26. The seed base, exposing the furrow of the micropyle in the ventral slightly damagedwall and the raphe furrow in the dorsal wall. Fig. 27. The base of the main spine, showing the distinct raphefurrow in the upper part and the less distinct furrow in the lower part of the spine base. Fig. 28. The apical-dorsal seed part with the raphe furrow. Figs. 29–31. Details of the very distinct raphe furrow in a fragmentaryseed valve, the same as in Fig. 16 (IPUW 3112/24). Fig. 29. The raphe furrow is distinct and wide in the mainspine and below. Fig. 30. Detail of the raphe furrow in the main spine. Fig. 31. Surface of the raphe furrow.Figs. 32–34. Seed fragment of Stratiotes with the distinct and deep raphe furrow (IPUW 3114/07).Fig. 33. Detail of the apical seed part. Fig. 34. Detail of the raphe furrow surface
b
B. Meller and P. F. van Bergen: Anatomical and biochemical investigations of Ceratostratiotes 135

described elsewhere and we refer the reader tothese references for more details (references seeabove). Some less well-known anatomicaldetails of the Langau specimens are presentedhere. The raphe furrow is covered with a layerof longitudinally elongated cells (Figs. 32, 34).In the plane of symmetry on the keel the testashows a pattern of elongated strongly undulatecells. Generally the inner valve surfaceappears with a fine longitudinal striationbuild (Figs. 44, 58). At a larger magnification,the testa cells display their irregular shape(Fig. 52) in section and thick-walled stone cellswith small lumina and tunnels crossing thewalls (Fig. 53). One testa fragment is coveredby a thin smooth skinny layer, without any celloutlines but locally with a few flat circulartubercle-like warts of 3–4 lm in diameter(Fig. 59).
Ceratostratiotes from the Czech and
Croatian localities. Ceratostratiotes from Pod-
moli near Znojmo (Figs. 60–64): The seeds aremainly poorly preserved and the pits are oftenshallow. In sections, the seed coat usuallydisplays a shiny black wall without cell out-lines. With the SEM, cell structures remainindistinct. The characteristic seed coat struc-ture is only exposed in a few specimens at thebroken base of the spines. The raphe furrow isobservable sometimes (Figs. 61–62, 64) alongthe thickened dorsal wall and the micropyle atthe supra-basal ventral position (Fig. 63).Within the sediment, two well-preserved
specimens are situated parallel to each other(Fig. 60). This might indicate the position ofthe seeds within the fruit, but as there are noother specimens supporting this hypothesis, itis more probably that this positioning isfortuitous.The specimens figured by Buzek (1982, Pls.
II-IV) show often well preserved long spines orlack any spine (Buzek 1982: Pl. IV, Figs. 5–6).
Figs. 35–38. Ceratostratiotes sinjanus (Kerner) Buzek from Langau. Fig. 35. Dorsal-lateral view on a basalseed fragment with well preserved, but not complete, basal spines. The dorsal margins of the valves are slightlyprominent with a distinct furrow between them. Shallow pits build the seed surface sculpturing; scale 1 mm;LM (IPUW 3113/10). Fig. 36. Dorsal view on a basal seed fragment with a distinct furrow between theprominent valve margins indicating the course of the raphe. The seed surface is abraded producing a nearlysmooth surface. The basal spines are only fragmentary preserved; scale 100 lm; SEM (IPUW 3113/03).Fig. 37. Detail of Fig. 36, where the vascular bundles leave the seed coat; scale 10 lm. Fig. 38. Detail ofFig. 36, lower part of the gap; the elongate narrow furrows in the left side of the raphe furrow are probably theimpressions of the vascular bundles (vb); scale 10 lm
136 B. Meller and P. F. van Bergen: Anatomical and biochemical investigations of Ceratostratiotes
He figured the specimens in turned positionaccording his interpretation, calling the basalspines apical spines, the lateral spines basalspines, and used ventral and dorsal in theopposite meaning. According his drawings(Buzek 1982: Fig. 1b) he noticed the thickeneddorsal margin (= ventral suture) and recog-nized an inner and outer seed coat wallindicated by a line there (compare also Buzek1982: Pl. III, Fig. 8). The outer part of the seedcoat wall is called a winged margin.
Ceratostratiotes from Glavica (Figs. 65–75): The preservation of the anatomical struc-
tures of the seeds appears to be poorer than insamples from Podmoli or Langau. However,the main spines of a few specimens are nearlycompletely and three-dimensionally preserved(Figs. 65–70) and a long raphe furrow is oftendistinctly visible along the thickened dorsalvalve wall, partly indicated by the palersediment in filling the furrow (Figs. 68,70–71). The specimens all have a thin seedcoat with shallow pits mainly arrangedin longitudinal rows and a narrow elongatedto narrow obovate shape. With the SEM,the seed surfaces display varying shapes of the
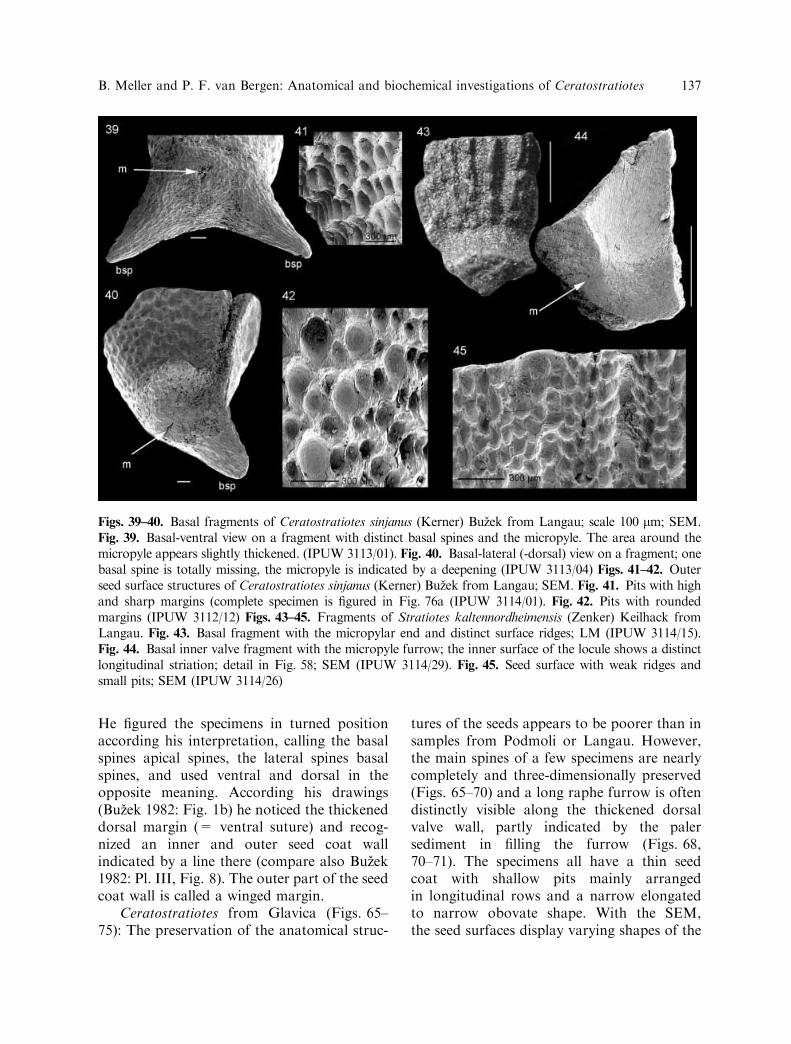
Figs. 39–40. Basal fragments of Ceratostratiotes sinjanus (Kerner) Buzek from Langau; scale 100 lm; SEM.Fig. 39. Basal-ventral view on a fragment with distinct basal spines and the micropyle. The area around themicropyle appears slightly thickened. (IPUW 3113/01). Fig. 40. Basal-lateral (-dorsal) view on a fragment; onebasal spine is totally missing, the micropyle is indicated by a deepening (IPUW 3113/04) Figs. 41–42. Outerseed surface structures of Ceratostratiotes sinjanus (Kerner) Buzek from Langau; SEM. Fig. 41. Pits with highand sharp margins (complete specimen is figured in Fig. 76a (IPUW 3114/01). Fig. 42. Pits with roundedmargins (IPUW 3112/12) Figs. 43–45. Fragments of Stratiotes kaltennordheimensis (Zenker) Keilhack fromLangau. Fig. 43. Basal fragment with the micropylar end and distinct surface ridges; LM (IPUW 3114/15).Fig. 44. Basal inner valve fragment with the micropyle furrow; the inner surface of the locule shows a distinctlongitudinal striation; detail in Fig. 58; SEM (IPUW 3114/29). Fig. 45. Seed surface with weak ridges andsmall pits; SEM (IPUW 3114/26)
B. Meller and P. F. van Bergen: Anatomical and biochemical investigations of Ceratostratiotes 137

138 B. Meller and P. F. van Bergen: Anatomical and biochemical investigations of Ceratostratiotes

pits dependent on the state of preservation(Figs. 74–75). Cross-sections of the seed coatexpose black shiny walls showing no cellstructures. The specimens from Glavica differfrom those of the Czech and Austrian localitiesin their generally larger length, which is up to9 mm. The main spines are located in theupper third of the seeds or are even sub-apical.None of the few specimens show a main spinein the middle part of the seed, as do specimensfrom Langau and Podmoli.
Ceratostratiotes from Stupalo: The extrac-ted specimens are all fragmentary and show thesame kind of preservation as the other speci-mens from Croatia, with shallow pits, thin-walled seed coat, and black shiny cross-sections.More complete specimens from that localitywith nicely preserved spines were investigatedby Buzek (1982: Pl. I).The very poor anatomical preservation of
the Croatian specimens might have been causedby the different mineralogical and chemicalcomposition of the marly sediment, in turnpossibly indicating a different water chemistryin which the plants grew. This might also havebeen responsible for the different position of thespines on the Croatian specimens, compared tothe Austrian and Czech ones.
Discussion of the morphological and anatom-
ical features. The most striking feature arguingfor apositionwithin theHydrocharitaceae is thecell structure of the seed coat. Shaffer-Fehre(1991b) proposed that layers of stone cells withlarge lumina and tunnels crossing the stone cellwalls, are characteristic of mesotestae ofHydrocharitaceae taxa. Within the Hydrocha-ritaceae genera, the shape and the frequency of
the tunnels and pits, as well as the number of celllayers, vary considerably. Their patterns arecharacteristic on generic and specific level(Shaffer-Fehre 1991b). For example, transversesections of Blyxa testae show no tunnels, butthey are observable in longitudinal section. Incontrast transverse sections of Stratiotes testaedisplay thick-walled stone cells with smalllumina and irregular pits and tunnels. Thedifferent species of Blyxa and Najas are notonly characterized bydifferent numbers of stonecell layers, the size of the cells and the lumina(Shaffer-Fehre 1991b: Figs. 22–23) but also bytheir outer surface morphology. The structureof the mesotesta and the endotegmen tubercleslead Shaffer-Fehre (1991b) to include Najas inthe Hydrocharitaceae. The endo- and exotestaof the seeds of modern Hydrocharitaceae con-sists of a thinmembranous single cell layer (pers.comm. Shaffer-Fehre 2002).The position and course of the raphe furrow
corresponds with that in fossil Stratiotes seedsand different lengths of the raphe (= hilumposition) characterize the different fossil Strati-otes species (Palamarev 1979: Table 1). InStratiotes kaltennordheimensis the raphe furrowis located within the seed coat at the plane ofdehiscence and symmetry along the thickeneddorsal valve wall (Figs. 18–19, 22) and leavesthe seed coat in the lower part, near the base.None of the fragmentary Stratiotes specimensfrom Langau displays the raphe furrow com-pletely preserved, therefore, we refer the readerto references, where such complete seeds arefigured (Holy and Buzek 1966,Knobloch 1989).Only one basal Stratiotes fragment from Lan-gau (Fig. 19) distinctly shows the lower part of
Figs. 46–53. Seed coat and spine anatomy of Ceratostratiotes sinjanus (Kerner) Buzek (Figs. 46–51) andStratiotes kaltennordheimensis (Zenker) Keilhack (Figs. 52–53) from Langau; SEM. Fig. 46. View on a seedcoat section and the pitted structure of the seed surface (IPUW 3112/07). Figs. 47, 49. Longitudinal section ofthe seed coat, showing the irregularly spongy seed coat structure with large holes (IPUW 3112/09). Figs. 48,50. Section of a lateral spine, exposing the stone cells; the detail in Fig. 50 shows a tunnel passing the stone cellwall (IPUW 3112/08). Fig. 51. Thick-walled stone cells of a basal spine; the stone cell walls are composed ofnumerous layers in contrast to the few layers of the lateral spine in Figs. 48, 50 (BSGPM 2002 I 4)Fig. 52. Longitudinal section of the seed coat of Stratiotes; note the innermost thin bright skin-like layer(IPUW 3114/09). Fig. 53. Thick-walled pitted stone cells of the Stratiotes testa; the cells are partly crushed (leftside) and compressed (IPUW 3114/08)
b
B. Meller and P. F. van Bergen: Anatomical and biochemical investigations of Ceratostratiotes 139

the raphe furrow. The raphe course is variableand changes between continuously transversalfrom apex to base (Fig. 18, 32; see also Knob-loch 1989: Pl. 1, Fig. 2) and transversal in theupper part, then near the outer margin inthe lower part (Knobloch 1989: Pl. 3, Fig. 1).The raphe furrowofCeratostratiotes is less deepand wide than in Stratiotes seeds (Figs. 12–20,25, 29, 32). Its course is easily observed in theupper part of the seed and in the proximal partof themain spine but often less clear in the lowerseed part, proposing, that the vascular bundlesleave the seed coat at the main spines. How-ever, a few specimens from Langau (Figs. 12,14–17, 20, 25, 29) and also from Podmoli
(Figs. 61–62, 64) and Glavica (Figs. 68, 70–73)display shallow furrows or at least weakimpressions along the dorsal seed coat wall,indicating the course of the vascular bundles.One basal fragment from Langau with a poorlypreserved abraded surface proves by the pres-ervation of the hilum that the raphe furrowleaves the seed coat in the lower part of the seedand runs within the dorsal gap between the twovalves (Figs. 36–38). Other specimens also dis-play such gap at the dorsal margin below themain spine (Fig. 35). Possibly, the length of theraphe within the seed coat could have beenvariable (as in Stratiotes kaltennordheimensis;compare Holy & Buzek 1966: Pl. 5) and/ or the
Figs. 54–59. Details of the inner valve surface of Ceratostratiotes sinjanus (Kerner) Buzek (Figs. 54–57) andStratiotes kaltennordheimensis (Zenker) Keilhack (Figs. 58–59) from Langau; SEM. Fig. 54. Inner valvesurface, exposing weak ridges (IPUW 3112/11). Fig. 55. A part of the innermost cell wall is turned aroundexposing straight cell margins and pitted anticlinal walls of the endotesta probably (BSGPM 2002 I 10).Fig. 56. Inner surface, exposing a smooth wrinkled skinny layer with rows of tubercle-like warts. The arrowspoint to the rows of tubercle-like warts (IPUW 3113/02). Fig. 57. Detail of Fig. 56 with the tubercle-like warts.Fig. 58. Inner valve surface of Stratiotes, exposing the longitudinal striation (IPUW 3114/29). Fig. 59.Tubercle-like warts on a smooth skinny inner layer of Stratiotes (IPUW 3119/09)
140 B. Meller and P. F. van Bergen: Anatomical and biochemical investigations of Ceratostratiotes

complete length is rarely preserved and observ-able. The vascular bundles might have beenlocated near the outer dorsal seed coat wall inthe lower part of the seed and their course isprobably not observable due to the positionnear to the margin and the abrasion of theoutermost seed coat layer. Similarly, the dorsalkeel on seedsofStratiotes kaltennordheimensis isfavoured for abrasion by its prominent position(Fig. 19). The thick-walled, uncompressed spec-imens fromLangau lack a distinct raphe furrowin the lower part of the seeds and exposes onlyweak impressions along the lower dorsal seedcoat wall whichmight be interpreted as to be theraphe furrow. More complete specimens nearlyalways gape at the dorsal margin between thetwo valves, which might be the reason for theabsenceof a distinct furrow in the seed coatwall.A slightly variable length of the vascular bun-dles might correspond with the position of theseeds within the fruit. The curved course of theraphe appears to be very odd and unusual. Butseveral specimens prove this feature.
msp
60
lsp
lsp
bspbsp
Fig. 60. Ceratostratiotes sinjanus (Kerner) Buzekfrom Podmoli, two seeds are situated parallel to eachother within the sediment, which might be fortuitousor due to the original position within the fruit; scale1 mm; LM (NMNHP G7905)
Figs. 61–64. Ceratostratiotes sinjanus (Kerner) Buzek from Podmoli; SEM (NMNHP G7906). Fig. 61. Innermorphology of a valve where raphe furrow and micropyle are well preserved; scale 1 mm. Figs. 62–64 show thedetails of this specimen. Fig. 62. View on the raphe furrow in the apical part of the seed. Fig. 63. Basal partwith the distinct micropyle furrow; scale 100 lm. Fig. 64. The curved raphe furrow in the main spine
B. Meller and P. F. van Bergen: Anatomical and biochemical investigations of Ceratostratiotes 141
Both Gregor (1980: Fig. 2) and Buzek(1982: Fig. 1) reconstructed the seed or fruitwith a short furrow, which represents eitherthe raphe furrow or the funicular canal. Inboth reconstructions, the furrow ends at thetop of the triangular extension at the mainspine. As already mentioned, Buzek’s (1982)figured specimens display the same features asthe Langau specimens but were interpreted in adifferent way.The supra-basal position of the micropyle
in Ceratostratiotes seeds (Figs. 21, 26, 39–40)fits very well with the micropyle position inseeds of Stratiotes kaltennordheimensis(Fig. 44). The micropylar thickened andcurved basal end of Stratiotes is slightlydifferent to the base of the Ceratostratiotesseeds, where this area is only slightly thickened(Figs. 9–10, 43) and less prominent but notcurved.The horizontal arrangement of the Cerato-
stratiotes spines represents a distinguishing
feature to Stratiotes and the other Hydrocha-ritaceae. Spines are absent in modern andfossil Stratiotes seeds but occur in some speciesof Blyxa. However, the spines are arranged inlongitudinal rows on the seed (Cook andLuond 1983, Shaffer-Fehre 1991b) as do theridges on the seed surface of Stratiotes kalten-nordheimensis (Figs. 9–11, 43–44). The surfaceof the seed of the extant Stratiotes aloides L. issmooth or shows weak longitudinal ridgeswhich are built by persistent hair bases of theunicellular hairs from the mucilaginous exo-testa layer (Cook and Urmi-Konig 1983: 219,Shaffer-Fehre 1991b: 195). The ridges of thefossil Stratiotes species apparently includeadditional stone cells (Gregor 1980).In contrast to the arrangement of the
Hydrocharitaceae spines and ridges, the struc-ture of the Ceratostratiotes spines is similar tothat in Blyxa (Shaffer-Fehre, pers. comm.2002) and the stone cell shape and structureof the Ceratostratiotes spines are very similar
65 66 67msp msp msp msp msp
mspapex
lsp
lsp lsplsp
lsp
lsp
lsp
lsplsp
lsp
lsp
bspbspbspbspbsp
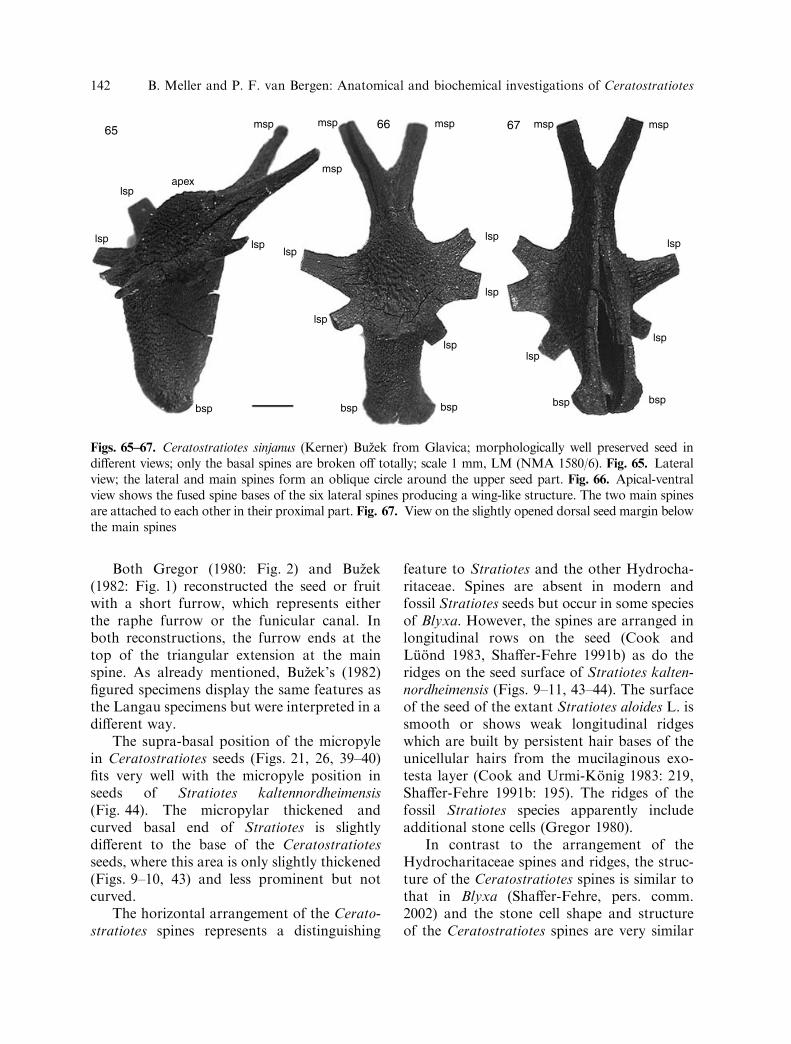
Figs. 65–67. Ceratostratiotes sinjanus (Kerner) Buzek from Glavica; morphologically well preserved seed indifferent views; only the basal spines are broken off totally; scale 1 mm, LM (NMA 1580/6). Fig. 65. Lateralview; the lateral and main spines form an oblique circle around the upper seed part. Fig. 66. Apical-ventralview shows the fused spine bases of the six lateral spines producing a wing-like structure. The two main spinesare attached to each other in their proximal part. Fig. 67. View on the slightly opened dorsal seed margin belowthe main spines
142 B. Meller and P. F. van Bergen: Anatomical and biochemical investigations of Ceratostratiotes

to the stone cells of the Stratiotes testa(Figs. 51, 53). Morphological variability ofCeratostratiotes is indicated by the absence ofspines in some seeds (Fig. 8) which, in thisaspect, closely resemble Stratiotes.
Gregor (1980) observed a thick spongylayer on fossil Stratiotes seeds which covers theridges and does not appear to be comparablewith the thin membranous single cell layer ofthe exotesta of modern Stratiotes seeds. He
68 69
70
72
71
73
74 75
r
r
rr
r
msp lsp
msp
r
m
m
bsp
bsp
Figs. 68–75. Ceratostratiotes sinjanus (Kerner) Buzek from Glavica. Figs. 68–73. Different views of the sameseed valve (NMA 1580/1); all SEM, apart from Figs. 68–69. Figs. 68, 70. Inner valve morphology; note thelong raphe furrow in the dorsal (right) wall margin scale 1 mm. Fig. 69. Outer valve morphology; the wellpreserved main spine is in the same plane as the lateral spines; the lateral spines and the basal spine are brokenoff. Fig. 71. Detail of the distinct raphe furrow in the main spine. Fig. 72. The basal seed part exposesdistinctly the micropyle furrow in the ventral wall and the raphe furrow in the dorsal margin; scale 100 lm.Fig. 73. Detail of the apical part with the distinct raphe furrow. Fig. 74. Poorly preserved outer seed surfacewithout distinct pit margins; SEM (NMA 1580/10). Fig. 75. Poorly preserved outer seed surface with undulatemargins of the pits; SEM (NMA 1580/12)
B. Meller and P. F. van Bergen: Anatomical and biochemical investigations of Ceratostratiotes 143

144 B. Meller and P. F. van Bergen: Anatomical and biochemical investigations of Ceratostratiotes

proposed that the fossil Stratiotes species donot belong to the modern genus Stratiotes. Theexact anatomical structure of this spongy layerwas not described by Gregor (1980) and ourSEM-investigations did not reveal any struc-ture there. This might be due to either thepreservation quality or a real absence ofstructures.A pitted surface sculpturing also occurs in
the Miocene Stratiotes kaltennordheimensis(Figs. 11, 45) as well as in modern Stratiotes,Blyxa and Najas species (Shaffer-Fehre 1991b).The shape and size vary within the moderngenera and are characteristic on species level.The pits of the Ceratostratiotes seeds aredistinctly wider than the pits of Stratioteskaltennordheimensis, the shape is rounder andthe deepening oblique in- and upwards direct-ed which has not been observed in Stratiotesseeds (Figs. 41–42, 45).The inner valve surfaces of Ceratostratiotes
and Stratiotes kaltennordheimensis are similarwith fine longitudinal striations and the occur-rence of an translucent thin inner layer.Endotegmen tubercles are a characteristic
feature of the Hydrocharitaceae (Shaffer Fehre1991a, b) at both generic and specific levels.Exotegmen tubercles have been only describedfor modern seeds of Limnobium, Hydrilla andHydrocharis (Shaffer-Fehre 1991a: Figs. 17–20). The position and shape of the tubercle-likewarts observable in one Ceratostratiotes speci-men from Langau (Figs. 56–57) does notappear to be comparable to the endotegmentubercles of the Hydrocharitaceae. Only thediameter fits with the size of the Hydrochari-taceae endotegmen tubercles (Shaffer-Fehre1991a: Table 2). The wart-bearing smoothlayer without any cell outlines might representa part of the nucellus cuticle (according to
Shaffer-Fehre, pers. comm. 2002: the nucelluscuticle and tegmen could not be separated inmodern Stratiotes seeds). There is only onespecimen with these structures and withoutchemical and structural investigations of theseparticular warts, it cannot be excluded thatthey are remains of mineral crystals or bacte-ria.The observed tubercle-like warts on a
smooth layer in Stratiotes kaltennordheimensis(Figs. 40–41) are larger than the endotegmentubercles in Stratiotes aloides L. (length ·width¼ 0.7 · 1.4 lm; Shaffer-Fehre 1991a:Table 2). The Stratiotes aloides seed investi-gated by Shaffer-Fehre derived from a 70 yearold herbarium sheet and displayed only a fewtubercles and it was assumed (Shaffer-Fehre1991a: 177) that fresh material might display adifferent pattern. More fossil and extantmaterial needs to be investigated but seeds ofStratiotes aloides are extremely rare in herbariaseed collections.In conclusion, there seems to be a high
degree of morphological-anatomical similaritybetween the Ceratostratiotes seeds and theseeds of Stratiotes and Blyxa.
Biochemistry. To determine the molecularrelationship between the two taxa, the macro-molecular compositions were examined usingflash-pyrolysis. The different pyrolysis analysesof samples within each taxon were virtuallyidentical indicating little chemical variabilitywithin the two species. The pyrolysates of theCeratostratiotes and Stratiotes sclerotic prop-agule wall samples (Langau B41/17) are bothdominated by phenols (Fig. 76) but the typesof phenols [i.e. simple alkylphenols (P, 2P,3+4P), catechols (C), methoxyphenols (G)]and the distribution patterns of these aredifferent between the two taxa.
Fig. 76. Total ion currents (TIC) of Curie-point (610�C) pyrolysates of sclerotic propagule walls of aCeratostratiotes sinjanus (IPUW 3114/26) and b Stratiotes kaltennordheimensis (IPUW 3114/01). Note that aportion of the TIC (Fig. 76b) has been enlarged (x5). P, phenol; 2P, 2-methylphenol; 3+4P, coeluting 3-methyl-and 4-methylphenol; C , catechol; G, guaiacol. Side chains indicated are attached to carbon atom number 4(opposite the OH group). LG, Levoglucosan (i.e. cellulose marker). The two figured specimens derive from thesame sample as the biochemical investigated specimens. The spines of Ceratostratiotes sinjanus (a) are brokenoff
b
B. Meller and P. F. van Bergen: Anatomical and biochemical investigations of Ceratostratiotes 145

Ceratostratiotes sinjanus. The total ioncurrent of the pyrolysate of the insolubleresidue of the sclerotic propagule wall ofCeratostratiotes is dominated by 2-methoxy-phenols (=guaiacyl derivatives; G inFig. 76a). The main derivatives include2-methoxyphenol, 4-methyl-2-methoxyphenol,4-vinyl-2-methoxyphenol and trans-4-prope-nyl-2-methoxyphenol. Additional importantproducts are eugenol, cis-4-propenyl-2-meth-oxyphenol, two O-containing guaiacyl deriva-tives and 4-(2-(E)-propenal)-2-methoxyphenolclearly showing intact C3 side-chains. Otherrelatively abundant pyrolysis products includephenol (P), methylphenols (2P and 3+4P), 1,2-benzenediol (C) and 4-methyl-1,2-benzenediol.Only very few polysaccharide pyrolysis prod-ucts were detected of which only levoglucosan(LG, Fig. 76a) is annotated.These pyrolysis results clearly reveal the
presence of lignin based on the characteristic 2-methoxyphenols (Saiz-Jimenez and de Leeuw1986, van Bergen et al. 1994b). In particular,the distribution of the main products and thepresence of many derivatives with intact C3side-chains (including 4-(2-(E)-propenal)-2-methoxyphenol) show a relatively well-pre-served lignin macromolecule (van Bergen et al.1994b, 2000). Further evidence of high qualitychemical preservation is revealed by the pres-ence of levoglucosan, a marker for cellulose(Pouwels et al. 1989). However, the absence ofadditional abundant polysaccharide pyrolysisproducts, including hemicellulose markers,indicates that a certain degree of chemicalalteration has affected these seed coats. This isfurther substantiated by the presence of 1,2-benzenediols (C) and simple phenols both ofwhich are known as lignin degradation prod-ucts (Hatcher and Clifford 1997).Characteristic angiosperm lignin moieties,
i.e. 2,6-dimethoxyphenol (= syringyl), werenot detected. Chemical alteration may also bethe explanation for the absence of thesecharacteristic angiosperm moieties. To date,lignin from propagules of all modern angio-sperms have yielded 2,6-dimethoxyphenolsupon pyrolysis (Boon et al. 1989; van Bergen
1994, 2001; van Bergen et al. 1994a, b, 1995,1996, 1997; Bland et al. 1998), but fossilangiosperm propagules rarely yield 2,6-dimethoxyphenols (e.g. Boon et al. 1989).Dimethoxyphenols are well known to be pre-ferentially lost from the lignin macromoleculeupon the very early stages of fossilization,often as early as 10,000 years (van Bergen et al.2000 and references cited therein). Notwith-standing a certain degree of alteration, theoverall pyrolysis data suggest that the fossilseed coat of Ceratostratiotes was originallycomposed of a ligno-cellulose based biomac-romolecule similar to that normally found inmodern angiosperms. Unfortunately thesedata allow no differentiation between dicoty-ledonous and monocotyledonous lignin whichare chemically different in modern plants butbecome more similar in fossil specimens (dueto the preferential loss of 2,6-dimethoxyphe-nols and the monocotyledonous-specificp-coumaric and ferulic acids; van Bergen et al.1994b, 1996, 1997).
Stratiotes kaltennordheimensis. In con-trast, the pyrolysate of the insoluble residueof the seed coat of Stratiotes kaltennordheim-ensis is dominated by simple phenols, i.e.phenol (P) and methylphenols (2P and 3+4P;Fig. 76b), as opposed to the methoxyphenols(G) in the pyrolysate of Ceratostratiotes. Allother pyrolysis products are much less im-portant (note the magnification) but include anumber of characteristic products such as 1,4-benzenediol and formylphenol (= 4-hydroxy-benzaldehyde). In addition, low relativeamounts of guaiacyl derivatives, C2-phenols,and (alkyl) dibenzofurans are detectedwhilst polysaccharide pyrolysis products areabsent.Pyrolysis data of seed coats of modern
Stratiotes aloides (van Bergen et al. 1994a),fossil s. a. S. kaltennordheimensis (Miocene;van Bergen et al. 1995) and S. headonenensisChandler (Eocene and Oligocene, van Bergenet al. 1994a, b), have revealed a distinctchemical composition of the lignin in thisaquatic monocotyledonous angiosperm. Todate, all pyrolysates of Stratiotes seed coats
146 B. Meller and P. F. van Bergen: Anatomical and biochemical investigations of Ceratostratiotes

have been shown to be dominated by phenol(P). Van Bergen et al. (1994b) suggested thatphenol in these pyrolysates is a decarboxyla-tion product of 4-hydroxybenzoic acid. Thislatter compound appears to be a Stratiotesspecific lignin building block (van Bergen et al.1994b) and has been detected in pyrolysates ofmodern, one Miocene and two Eocene speci-mens (van Bergen et al. 1994a, b, 1995).However, in pyrolysates of fossil specimensthis precursor acid is only present in very smallrelative amounts (in one Miocene and twoEocene samples) or even absent all together (inall other fossil Stratiotes specimens). Othercharacteristic Stratiotes pyrolysis products are1,4-benzenediol and 4-formylphenol, both ofwhich are only minor pyrolysis products innormal angiosperm lignin (i.e. both di- andmonocotyledons). Despite the absence of thediagnostic compound 4-hydroxybenzoic acid,the presence of other taxon specific productsand the distinct abundance of the (alkyl)phe-nols implies that the chemical composition ofthe seed coat of these Stratiotes kaltennord-heimensis is similar if not identical to that inpreviously studied Stratiotes seeds. Thus, thegenus-specific chemical signals are still retainedin these specimens.Overall, the biochemical data reveal evi-
dence of a certain degree of chemical altera-tion, i.e. removal of polysaccharides, but witha recognizable lignin macromolecule stillpreserved. Furthermore, the pyrolysis resultsshow that the chemical composition of thetwo taxa differ considerably. The biochemist-ry of the Stratiotes seed coat can be clearlyassigned to the genus Stratiotes whereas thechemical composition of the Ceratostratiotesseed coats points towards macromolecularmaterial similar to that of other fossilnon-monocotyledonous angiosperms (e.g.Brasenia sp., Sabrenia sp., Nymphaea sp.;van Bergen et al. 1994a, b, 1996). Despitethe lack of characteristic dicotyledonous com-pounds in the pyrolysates of Ceratostratiotesthe abundance of the guaiacyl compounds issimilar to those found in pyrolysates of dico-tyledons (as e.g. Nymphaeaceae and Nyssa
(Nyssaceae)) and not like that foundin monocotyledons.
Conclusions
The organic chemical composition of theCeratostratiotes outer seed coat wall differssignificantly from the lignin composition of theStratiotes seed coat wall.A mixture of similarities and differences
characterize the anatomical and morphologicalfeatures. The detailed investigation of the3-dimensional anatomically preserved speci-mens of Ceratostratiotes confirms that aspectsof Ceratostratiotes anatomy and morphologystrongly resemble those of Stratiotes seeds:sclereid anatomy in section, raphe position,micropylar organisation and internal surfacetexture are all similar. Large spines, formed asextensions of the outer seed coat wall inCeratostratiotes are not found in Stratiotes.Longitudinal surface ridges are characteristicof Stratiotes kaltennordheimensis but Cerato-stratiotes displays no longitudinally arrangedsurface extensions. Ceratostratiotes and Strati-otes both are characterized by a pitted surface,although the shape and size of the pits differ.The basal expanded micropylar end in Strati-otes differs from the narrow, rounded and onlyslightly expanded base in Ceratostratiotes. Theweak dorsal keel in Ceratostratiotes is similarto the prominent keel in Stratiotes.Assuming that both the morphological-
anatomical and the biochemical methods arereliable approaches in systematic-taxonomicinvestigations, then the answer to the criticalquestion about the systematic position ofCeratostratiotes is not as simple as originallyanticipated. The distinct morphological-ana-tomical evidence suggests that the systematicposition of Ceratostratiotes is within theHydrocharitaceae. Several monocots includedicotyledonous features (Chase et al. 2000)and the non-monocotyledonous biochemicalcomposition might be explained by this fact.To date, biochemical information of the mac-romolecular composition of seed- and fruitcoat walls of the numerous plant genera and
B. Meller and P. F. van Bergen: Anatomical and biochemical investigations of Ceratostratiotes 147

families are still fragmentary and thus detailedsystematic conclusions are affected by this.Additionally biochemical and morphological-anatomical analyses of the other Hydrochari-taceae genera have to be done in order toelucidate the systematic position of this taxon.If the testae of all other Hydrocharitaceaegenera show monocotyledon-like composi-tions, Ceratostratiotes might be a representa-tive of a separate family. If the seed coatwall of some genera are monocotyledon-like,others dicotyledon-like, the classificationwithin the family has to be reconsidered. Alsoof interest would be then the seed- and fruitcoat wall compositions of the other familiesof Alismatales, the Acorales and also theCeratophyllales.At present the enigma of the affinity of
Ceratostratiotes remains unsolved with thebiochemistry suggesting a non-monocot affin-ity. The current combined investigation on thesystematic position of Ceratostratiotes appearsto have risen even more questions clearlyindicating the essential need for further inte-grated studies of fossil and modern plantpropagules.
We thank numerous colleagues whose interestand helpful comments or references have supportedthis investigation: M.E. Collinson (University ofLondon, Royal Holloway; Great Britain), D.K.Ferguson, C.-C. Hofmann (University of Vienna,Austria), H.-J. Gregor (Nature Museum Augsburg,Germany), Z. Jurisic-Polsack (Croatian NaturalHistory Museum Zagreb, Croatia), M. Shaffer-Fehre, P. Rudall (Royal Botanical Gardens, Kew;Great Britain). In Prague (Czech Republic),Z. Kvacek and J. Kvacek were very thorough intheir search for Buzek’s material in the NationalMuseum of Natural History and in the CharlesUniversity. Specimens were also studied in theMuseum of Nature in Augsburg, as they werementioned previously by H.-J. Gregor, in theBavarian State Collection of Geology and Pala-eontology in Munich (Germany) and the NaturalHistory Museum in Vienna (Austria). This inves-tigation has been supported by the AustrianScience Fund project P-13742. Special thanks toA. H. N. Rice for correcting the English and to
M. E. Collinson for her review, which improved theclarity of the manuscript.
References
Berger W. (1957) Eine neue fossile Wassernuß ausden untermiozanen Braunkohlenablagerungenvon Langau bei Geras in Niederosterreich.Phyton 7,1–3: 152–158.
Bland H. A., van Bergen P. F., Carter J. F.,Evershed R. P. (1998) Early diagenetic transfor-mations of proteins and polysaccharides inarchaeological plant remains. In: StankiewiczB. A., van Bergen P. F. (eds.) Nitrogen-Con-taining Macromolecules in the Bio- and Geo-sphere. Oxford University Press, New York, pp.113–131. (ACS Symposium Series, 707).
Boon J. J., Stout S. A., Genuit W., Spackman W.(1989) Molecular paleobotany of Nyssa endo-carps. Acta Bot. Neerl. 38: 391–404.
Buzek C. (1982) Ceratostratiotes Gregor, an extinctwater plant of uncertain affinity from the Euro-pean Miocene. Vest. Ustred. ust. geol. 57, 5:285–294.
Chase M. W., Soltis D. E., Soltis P. S., Rudall P. J.,Fay M. F., Hahn W. H., Sullivan S., Joseph J.,Molvray M., Kores P. J., Givnish T. J., SytsmaK. J., Pires J. C. (2000) Higher-Level systematicsof the Monocotyledons: an assessment of currentknowledge and a new classification. In: WilsonK. L., Morrison D. A. (eds.) Monocots: Syste-matics and Evolution. CSIRO, Collingwood,pp. 3–16.
Cook C. D. K. (1987) Dispersion in aquaticand amphibious vascular plants. In: CrawfordR. M. M. (ed.) Plant life in aquatic andamphibious habitats: produced as a tribute toD. H. N. Spence, Oxford, pp. 179–190.
Cook C. D. K., Luond R. (1983) A revision of thegenus Blyxa (Hydrocharitaceae). Aquat. Bot. 15:1–52.
Cook C. D. K., Urmi-Konig K. (1983) A revisionof the genus Stratiotes (Hydrocharitaceae).Aquat. Bot. 16: 213–249.
Draxler I. (1991) Die untermiozane Mikrofloraaus dem Raum Eggenburg – Horn – Geras.In: Roetzel R. (ed.) Geologie am Ostrand derBohmischen Masse, Schwerpunkt Blatt 21 Horn.Geol. Bundesanst. Wien, pp. 109–113.
Gregor H.-J. (1980) Trapa zapfei Berger aus demUntermiozan von Langau bei Geras (NO.) – eine
148 B. Meller and P. F. van Bergen: Anatomical and biochemical investigations of Ceratostratiotes

Hydrocharitaceae. Ann. Naturhist. Mus. Wien83: 105–118.
Gunther T., Gregor H.-J. (1989) Computeranalyseneogener Frucht- und Samenfloren Europas.Band 1: Fundorte und deren Florenlisten. Do-cumenta naturae 50, 1:1–180.
Gunther T., Gregor H.-J. (1997) Computeranalyseneogener Frucht- und Samenfloren Europas.Band 5: Artennachweise und stratigraphischeProblematik. Documenta naturae 50, 5: 1–180.
Hatcher P. G., Clifford D. J. (1997) The organicgeochemistry of coal: from plant materials tocoal. Organic Geochem. 27: 251–274.
Holy F., Buzek C. (1966) Seeds Stratiotes L.(Hydrocharitaceae) in the Tertiary of Czecho-slovakia. Sborn. geol. ved., Paleont. 8: 108–135.
Hooker J. J., Collinson M. E., van Bergen P. F.,Singer R. L., de Leeuw J. W., Jones T. P. (1995)Reconstruction of land and freshwater palaeoen-vironments near the Eocene-Oligocene bound-ary, southern England. J. Geol. Soc. London152: 449–468.
Kerner F. v. (1905a) Neogenpflanzen vom Nord-rande des Sinjsko polje in Mitteldalmatien. Jb.kaiser konigl geol. Reichsanst. 55: 593–612.
Kerner F. v. (1905b) Gliederung der SinjanerNeogenformation. Verh. kaiserl. konigl. geol.Reichsanst. 1905, 6: 127–165.
Kerner F. v. (1916) Alt- und jungtertiare Pflanzen-reste aus dem oberen Cetinatale. Verh. kaiserl.konigl. geol. Reichsanst. 1916, 8: 181–191.
Kerner F. v. (1917) Die Lignitformation im Vraba-tale (Mitteldalmatien). Verh. kaiserl. konigl.geol. Reichsanst. 1917, 12: 202–213.
Knobloch E. (1989) Biometrie und Morphologieder Samen von Stratiotes kaltennordheimensisand S. tuberculatus aus dem mitteleuropaischenNeogen. Sborn. geol. ved., Paleont. 30: 159–173.
Les D. H. (1988) The origin and affinities of theCeratophyllaceae. Taxon 37: 326–345.
Les D. H. (1989) The Evolution of AcheneMorphology in Ceratophyllum (Ceratophylla-ceae), IV. Summary of Proposed Relation-ships and Evolutionary Trends. Syst. Bot. 14,2: 254–262.
Les D. H., Schneider E. L. (1995) The Nymphae-ales, Alismatidae, and the theory of an aquaticmonocotyledon origin. In: Ruddall P. J., CribbP. J., Cutler D. F., Humphires C. J. (eds.)Monocotyledons: systematics and evolution.Royal Botanical Gardens, Kew, pp. 23–42.
Palamarev E. H. (1979) Die Gattung Stratiotes L.in der Tertiarflora Bulgariens und ihre Ent-wicklungsgeschichte in Eurasien. Phytology 12:3–36.
Pouwels A. D., Eijkel G. B., Boon J. J. (1989) Curiepoint pyrolysis capillary gas chromatographyhigh resolution mass spectrometry of microcrys-talline cellulose. J. Anal. Appl. Pyrolysis 14: 237–280.
Roetzel R. (1993) Bericht 1992 uber geologischeAufnahmen im Tertiar und Quartar auf Blatt 8Geras und Bemerkungen zur Lithostratigraphiedes Tertiars in diesem Raum. Jb. Geol. Bundes-anst. Wien 136: 542–546.
Roetzel R. (1994) Der Braunkohlenbergbau vonLangau. In: Brandtner A. J. (ed.) Langau imWaldviertel. Community of Langau, pp. 299–319.
Roetzel R., Fuchs G. (1994) Der geologischeAufbau der Landschaft um Langau. In: Brandt-ner A. J. (ed.) Langau im Waldviertel. Commu-nity of Langau, pp. 284–298.
Saiz-Jimenez C., de Leeuw, J. W. (1986) Ligninpyrolysis products: Their structures and theirsignificance as biomarkers. Organic Geochem.10: 869–876.
Shaffer-Fehre M. (1991a) The endotegmen tuber-culae: an account of little-known structures fromthe seed coat of the Hydrocharitoideae (Hy-drocharitaceae) and Najas (Najadaceae). Bot. J.Linn. Soc. 107: 169–188.
Shaffer-Fehre M. (1991b) The position ofNajas within the subclass Alismatidae (Mono-cotyledones) in the light of new evidence fromseed coat structures in the Hydrocharitoideae(Hydrocharitales). Bot. J. Linn. Soc. 107: 189–209.
Stankiewicz B. A., Mastalerz M., Kruge M. A., vanBergen P. F., Sadowska A. (1997) A compara-tive study of modern and fossil cone scales andseeds of conifers: a geochemical approach. NewPhytologist 135: 375–393.
Stankiewicz B. A., van Bergen P. F., Smith M. B.,Carter J. F., Briggs D. E. G., Evershed R. P.(1998) Comparison of analytical performance offilament and Curie-point pyrolysis devices.J. Anal. Appl. Pyrolysis. 45: 133–151.
van Bergen P. F. (1994) Palaeobotany of Propa-gules: An Investigation combining Microscopyand Chemistry. Ph. D. thesis, University ofLondon, UK.
B. Meller and P. F. van Bergen: Anatomical and biochemical investigations of Ceratostratiotes 149

van Bergen P. F. (1999) Pyrolysis and chemolysis offossil plant remains: applications to palaeobota-ny. In: Jones T. P., Rowe N. P. (eds.) Fossilplants and spores: modern techniques. TheGeological Society, London, pp. 143–148.
van Bergen P. F. (2001) Resistant plant tissues –Cuticles and Propagules. In: Briggs D. E. G.,Crowther P. R. (eds.) Palaeobiology II. Black-well Science, Oxford, pp. 256–259.
van Bergen P. F., Collinson M. E., Hatcher P. G.,de Leeuw J. W. (1994a) Lithological control onthe state of preservation of fossil seed coats ofwater plants. Organic Geochem. 22: 683–702.
van Bergen P. F., Goni M., Collinson M. E., BarrieP. J., Sinninghe Damste J. S., de Leeuw J. W.(1994b) Chemical and microscopic characteriza-tion of outer seed coats of fossil and extant waterplants. Geochim. Cosmochim. Acta 58: 3823 –3844.
van Bergen P. F., Collinson M. E., Briggs D. E. G.,de Leeuw J. W., Scott A. C., Evershed R. P.,Finch P. (1995) Resistant biomacromolecules inthe fossil record. Acta Bot. Neerl. 44: 319–342.
van Bergen P. F., Collinson M. E., de Leeuw J. W.(1996) Characterization of the insoluble constit-uents of propagule walls of fossil and extant
water lilies; implications for the fossil record.Ancient Biomolecules 1: 55–81.
van Bergen P. F., Bland H. A., Horton M. C.,Evershed R. P. (1997) Chemical and morpho-logical changes in archaeological seeds and fruitsduring preservation by desiccation. Geochim.Cosmochim. Acta 61: 1919–1930.
van Bergen P. F., Collinson M. E., Stankiewicz B.A. (1999) The importance of molecular palae-obotany. Acta Palaeobot. Supplement 2: 653–657.
van Bergen P. F., Poole I., Ogilvie T. M. A., CapleC., Evershed R. P. (2000) Evidence for Deme-thylation of Syringyl Moieties in ArchaeologicalWood using Pyrolysis/Gas Chromatography/Mass Spectrometry. Rapid Communications inMass Spectrometry 14: 71–79.
Addresses of the authors: Barbara Meller(e-mail: [email protected]), Universityof Vienna, Palaeontological Institute, Geocentre,Althanstrasse 14, A-1090 Vienna. Pim F. vanBergen, Geochemistry, Faculty of Earth Sciences,Utrecht University, P.O. Box 80021, NL-3508 TAUtrecht, The Netherlands.
150 B. Meller and P. F. van Bergen: Anatomical and biochemical investigations of Ceratostratiotes
Copyright © 2022 FDOKUMEN




















