
The pharmacology of Malo maxima jellyfish venom extract in isolated cardiovascular tissues: A...
10
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of The pharmacology of Malo maxima jellyfish venom extract in isolated cardiovascular tissues: A...

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Toxicology Letters 201 (2011) 221–229
Contents lists available at ScienceDirect
Toxicology Letters
journa l homepage: www.e lsev ier .com/ locate / tox le t
The pharmacology of Malo maxima jellyfish venom extract in isolatedcardiovascular tissues: A probable cause of the Irukandji syndromein Western Australia
Ran Lia,b,1, Christine E. Wrighta,∗,1, Kenneth D. Winkelb, Lisa-Ann Gershwinc, James A. Angusa
a Cardiovascular Therapeutics Unit, Department of Pharmacology, The University of Melbourne, Grattan Street, Parkville, Victoria 3010, Australiab Australian Venom Research Unit, Department of Pharmacology, The University of Melbourne, Victoria 3010, Australiac Queen Victoria Museum, Launceston, Tasmania 7250, Australia
a r t i c l e i n f o
Article history:Received 16 September 2010Received in revised form 5 January 2011Accepted 6 January 2011Available online 13 January 2011
Keywords:IrukandjiMalo maximaCarukia barnesiCatecholaminesCalcitonin gene-related peptideJellyfish venom
a b s t r a c t
The in vitro cardiac and vascular pharmacology of Malo maxima, a newly described jellyfish suspected ofcausing Irukandji syndrome in the Broome region of Western Australia, was investigated in rat tissues. Inleft atria, M. maxima crude venom extract (CVE; 1–100 �g/mL) caused concentration-dependent inotropicresponses which were unaffected by atropine (1 �M), but significantly attenuated by tetrodotoxin (TTX;0.1 �M), propranolol (1 �M), Mg2+ (6 mM) or calcitonin gene-related peptide antagonist (CGRP8–37;1 �M). CVE caused no change in right atrial rate until 100 �g/mL, which elicited bradycardia. Thiswas unaffected by atropine, TTX, propranolol or CGRP8–37. In the presence of Mg2+, CVE 30–100 �g/mLcaused tachycardia. In small mesenteric arteries CVE caused concentration-dependent contractions(pEC50 1.03 ± 0.07 �g/mL) that were unaffected by prazosin (0.3 �M), �-conotoxin GVIA (0.1 �M) orMg2+ (6 mM). There was a 2-fold increase in sensitivity in the presence of CGRP8–37 (3 �M). TTX (0.1 �M),box jellyfish Chironex fleckeri antivenom (92.6 U/mL) and benextramine (3 �M) decreased sensitivity by2.6, 1.9 and 2.1-fold, respectively. CVE-induced maximum contractions were attenuated by C. fleckeriantivenom (−22%) or benextramine (−49%). M. maxima CVE appears to activate the sympathetic, but notparasympathetic, nervous system and to stimulate sensory nerve CGRP release in left atria and resistancearteries. These effects are consistent with the catecholamine excess thought to cause Irukandji syndrome,with additional actions of CGRP release.
© 2011 Elsevier Ireland Ltd. All rights reserved.
1. Introduction
The Irukandji syndrome is a jellyfish-related envenomationthat has been attributed to the sting of certain species of small,four-tentacled box jellyfish (Barnes, 1964; Fenner et al., 1988;Flecker, 1952; Gershwin, 2007, 2008; Gershwin and Alderslade,2005; Little et al., 2006; Southcott, 1967). The syndrome has threerecognised clinical components consisting of (i) acute muscularchest and back pain; (ii) catecholamine-like effects notably sweat-ing, anxiety, nausea, vomiting, headache, tachycardia, potentiallylife-threatening hypertension with supraventricular tachyarrhyth-mias; and (iii) cardiopulmonary decompensation. Most patientspresent with milder, non-life threatening symptoms including gen-eralised pain, hypertension, nausea, vomiting and distress. About
∗ Corresponding author. Tel.: +61 3 8344 8219; fax: +61 3 8344 0241.E-mail addresses: [email protected] (R. Li), [email protected]
(C.E. Wright), [email protected] (K.D. Winkel), [email protected](L.-A. Gershwin), [email protected] (J.A. Angus).
1 These authors contributed equally to this work.
half require hospital admission and a small number need advancedlife support, usually because of cardiac failure (Huynh et al., 2003;Macrokanis et al., 2004). The deaths in 2002 of two tourists inFar North Queensland, Australia, highlighted the potential risk andseverity of this syndrome (Fenner and Hadok, 2002; Huynh et al.,2003). It is now appreciated to occur elsewhere in the Indo-Pacificand the Caribbean (de Pender et al., 2006; Grady and Burnett, 2003;Pommier et al., 2005; Yoshimoto and Yanagihara, 2002).
Only a single species – Carukia barnesi – has been definitivelyconfirmed as a cause of Irukandji syndrome, however many dif-ferent species have been linked to it (Barnes, 1964; Burnett et al.,1996; Fenner et al., 1985; Gershwin, 2005, 2007; Little et al., 2001,2006; Little and Seymour, 2003; O’Reilly et al., 2001; Southcott,1967; Tibballs et al., 2001). One species suspected of causingsevere Irukandji syndrome near the Broome region of WesternAustralia is the recently described Malo maxima (Gershwin, 2005).The syndrome is a major occupational health and safety hazard forBroome’s pearling industry and a concern for locals and tourists(Macrokanis et al., 2004). However, aside from the original taxo-nomical description of M. maxima, no research has been conductedon this species. Consequently, treatment of Irukandji syndrome
0378-4274/$ – see front matter © 2011 Elsevier Ireland Ltd. All rights reserved.doi:10.1016/j.toxlet.2011.01.003

Author's personal copy
222 R. Li et al. / Toxicology Letters 201 (2011) 221–229
in Western Australia is based purely on the assumption that thevenom of M. maxima has similar, if not identical, mechanisms ofaction to that of C. barnesi (Macrokanis et al., 2004).
Current understanding of the pharmacological mechanismsunderlying Irukandji syndrome is rather limited. Amongst sus-pected Irukandji species, only the venoms of C. barnesi and Alatinamordens, both thought to be limited to Queensland waters, havebeen studied to any extent (Ramasamy et al., 2005; Tibballs et al.,2001; Winkel et al., 2005; Winter et al., 2008). Clinically, some fea-tures of Irukandji syndrome resemble that of adrenal medullary orcatecholamine excess, such as seen in phaeochromocytoma and inacute autonomic overactivity conditions (Baguley, 2008; Burnettet al., 1998). Indeed, elevated serum catecholamine levels havebeen found in vivo when animals were injected with C. barnesivenom (Ramasamy et al., 2005; Tibballs et al., 2001; Winkel et al.,2005; Winter et al., 2008). The most effective therapy for Irukandjisyndrome to date seems to be an intravenous infusion of magne-sium (MgSO4 or MgCl2) (Corkeron et al., 2004; Corkeron, 2003). Therationale for the use of magnesium therapy is based on the in vivoevidence of catecholamine excess, sympathetic and parasympa-thetic involvement in Irukandji syndrome, combined with theapparent utility of magnesium in reducing evoked release of sym-pathetic neurotransmitters (Ohtsuka et al., 2002; Tibballs et al.,2001). Anecdotal evidence suggests that it is useful in decreasingboth the hypertension and severe pain of the syndrome (Corkeronet al., 2004; Corkeron, 2003). Therefore, the aims of this study wereto assess whether a species other than C. barnesi can cause effectsconsistent with Irukandji syndrome in cardiac and vascular tis-sues in vitro, and to determine the specific properties of M. maximavenom. Specifically, the roles of sympathetic and parasympathetic,as well as of the sensory, neurotransmitters in the actions of M. max-ima venom were ascertained. Further, the effects of magnesium onthe responses to the venom extract were assessed with a view totreatment of such a potential Irukandji syndrome.
2. Methods
2.1. Collection of specimens
All specimens of M. maxima were caught offshore from 80 Mile Beach, in thepearling grounds south of Broome, Kimberley coast, Western Australia, from April toMay, 2004. The specimens were captured at night, using lights to attract the animalsand collected either with a custom-designed hand-net or drift-net (Uninet Enclo-sure Systems; Cairns, QLD, Australia). Upon collection, specimens were immediatelyplaced in small containers of seawater, pending identification via light microscopy.Specimens were examined by microscopy and formally identified, prior to pro-cessing, as M. maxima according to Gershwin’s description of bell, tentacle andnematocyst morphology (Gershwin, 2005).
Thereafter they were individually placed into plastic zip-lock bags and imme-diately frozen on dry ice (−80 ◦C). The frozen specimens were then transportedat −80 ◦C to the Australian Venom Research Unit (Department of Pharmacology,The University of Melbourne, Victoria, Australia) and stored at −70 ◦C as wholespecimens prior to processing.
2.2. Extraction of venom and protein determination
The protocol used to obtain M. maxima crude venom extract (CVE) was adaptedfrom that used for C. barnesi CVE (Tibballs et al., 2001; Wiltshire et al., 2000; Winkelet al., 2005). Briefly, thawed specimens of M. maxima were manually ground for30 min in a siliconized glass mortar and pestle surrounded by an ice slurry. The resul-tant mixture was centrifuged at 2292 × g for 15 min at 4 ◦C (Sigma Laborzentrifugen3K15; Osterode am Herz, Germany). The supernatant was decanted, separated into1 mL aliquots and frozen at −70 ◦C until needed.
Protein concentration of CVE was determined using the Bradford protein assayand compared to a bovine serum albumin (BSA) standard curve of known con-centration (0–0.2 mg/mL) (Bradford, 1976). Bio-Rad® Protein Assay Dye ReagentConcentrate (Hercules, CA, USA) was added to both the CVE sample and the BSAstandard and the absorbance at 590 nm was measured with a spectrophotometer(Wallac 1420 Victor; Perkin Elmer, Boston, MA, USA).
2.3. Biochemical analysis
Sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE) wasused to characterise the protein content of M. maxima CVE. All CVE samples were
first diluted to 5 �L aliquots of 2 mg/mL CVE. Non-reducing sample buffer (0.125 MTris–HCl, pH 6.8, 4% SDS, 20% glycerol) was then added and the samples were boiledfor 1 min and centrifuged for 20 s at 17,968 × g. BenchMarkTM Pre-Stained ProteinLadder (InvitrogenTM; Carlsbad, CA, USA) was used as a molecular weight markerfor the range of 181.8–6 kDa.
Electrophoresis gels (12.5% acrylamide; 85 × 60 × 0.75 mm) were prepared atleast a day in advance from 40% acrylamide/bis solution, 37.5:1 (Bio-Rad Labora-tories, Hercules, CA, USA), stock solutions of resolving gel buffer (1.5 M Tris–HCl,pH 8.8), stacking gel buffer (0.5 M Tris–HCl, pH 6.8) and 10% SDS (Sigma–AldrichPty. Ltd., Castle Hill, NSW, Australia), TEMED (Sigma–Aldrich) and freshly made 10%ammonium persulphate (Sigma–Aldrich). The gels were stored at 4 ◦C until required.
The gels were run in a stock solution of tank buffer (0.025 M Tris, pH 8.3, 0.192 Mglycine, 0.1% SDS), at 50 V, 200 mA (constant voltage) for 30 min, then resolved at200 V, 10 mA (constant current) for 120 min. Visualisation of the protein bands wasachieved by a 30 min stain with Brilliant Blue R concentrate (Sigma–Aldrich), thentreatment with stock Coomassie destain solution (10% acetic acid, 20% methanol)until satisfactory protein banding visualisation was achieved.
2.4. Catecholamine assays
CVE catecholamine levels were determined with alumina adsorption, separatedby HPLC and the amounts quantified by electrochemical detection, as describedpreviously (Lambert and Jonsdottir, 1998). Analysis was performed at 24 ◦C withthe operating potentials set at +0.35 V for the guard cell and −0.35 and +0.29 V fordetectors 1 and 2, respectively. All measurements were made using the oxidizingpotential applied at detector 2 and compounds in M. maxima CVE were identifiedby their retention behaviour compared with that of authentic standard solutions.
2.5. In vitro pharmacological analysis
Animal tissue experiments were approved by The University of Melbourne Ani-mal Ethics Committee in accordance with the Australian code of practice for thecare and use of animals for scientific purposes (7th edition, 2004, National Healthand Medical Research Council, Australian Government). Male Sprague–Dawley rats(250–450 g) were anaesthetised via spontaneous inhalation of 5% halothane (Vet-erinary Companies of Australia Pty. Ltd.; Artarmon, NSW, Australia) and 95% O2
for 5 min before exsanguination. Tissues were immediately excised and placed ina beaker of Krebs’ physiological salt solution (PSS) with the following composi-tion (mM): NaCl 119; KCl 4.69; MgSO4·7H2O 1.17; KH2PO4 1.18; glucose 11 (5.5 formesenteric artery); NaHCO3 25; CaCl2·6H2O 2.5; EDTA 0.026. The PSS was continu-ously saturated with carbogen (95% O2; 5% CO2) at pH 7.4.
2.6. Rat left and right atria
The left atrium and the spontaneously beating right atrium were dissected whilesubmerged in PSS (Nakashima et al., 1982). Each atrium was pierced at opposingends by 30G stainless steel hooks; one attached to a platinum loop embedded in theorgan bath leg and the other to an isometric force transducer (FT03C, Grass Instru-ments; Warwick, RI, USA). Data were recorded via a transducer amplifier and Chartv.5.5.6 for Mac (ADInstruments Pty. Ltd.; Bella Vista, NSW, Australia). Mounted atria,in 35–37 ◦C PSS-filled jacketed organ baths, were stretched to a passive resting forceequivalent to 1 g (9.81 mN), left to equilibrate for 10 min and then re-stretched to 1 g.The left atrium was then stimulated electrically (1 Hz, 0.3 ms, approx. 120% thresholdvoltage) via two punctate platinum electrodes connected to a Grass S88 stimulator(Winkel et al., 2005). After 30 min stabilisation, tissues were then pre-treated for60 min with either PSS (vehicle), atropine (1 �M), propranolol (1 �M), tetrodotoxin(TTX; 0.1 �M), calcitonin gene related peptide 8–37 (CGRP8–37; 1 �M) or magnesiumsulphate (6 mM). M. maxima CVE was added to each bath in increasing cumulativeconcentrations (1–100 �g/mL), allowing sufficient time between additions for anychange in response to reach a plateau (approx. 5–10 min).
2.7. Rat mesenteric arteries
A 2 mm long segment of mesenteric artery was mounted on a pair of 40 �mdiameter stainless steel wires in a Mulvany-Halpern isometric myograph (610M;Danish Myo Technology; Aarhus, Denmark) connected to an amplifier (Mulvanyand Halpern, 1977). The tissues were stretched to a passive tension equivalentto a transmural pressure of 100 mmHg, as generated by a passive length-tensioncurve, and allowed to equilibrate for 30–60 min (Angus and Wright, 2000). Ves-sel viability was tested by simultaneous exposure to 10 �M noradrenaline andpotassium-depolarising solution (KPSS; PSS with an equimolar substitution of KClfor NaCl; K+ 124 mM) for 2 min.
The myograph was washed out with PSS and the tissue exposed to KPSS a secondtime to provide a reference contraction. After a second washout with PSS the ves-sel was treated for 10 min with one of PSS (vehicle), prazosin (0.3 �M), �-conotoxinGVIA (�-CTX GVIA; 0.1 �M), TTX (0.1 �M), CGRP8–37 (3 �M) or magnesium sulphate(6 mM). For benextramine pre-treatment, the arteries were first incubated for 5 minwith 0.3 �M prazosin to bind to �1-adrenoceptors, then 3 �M benextramine, an irre-versible non-selective �1- and �2-adrenoceptor antagonist, was added for a further5 min. The myograph was then washed out 3 times at 5 min intervals. This pro-

Author's personal copy
R. Li et al. / Toxicology Letters 201 (2011) 221–229 223
tocol effectively prepares a tissue with �2-adrenoceptors irreversibly blocked but�1-adrenoceptors fully functional (Angus et al., 1988).
The vessel was then actively precontracted with sufficient arginine vasopressin(AVP; approximately 0.1–0.2 nM) to elicit a small contraction equivalent to 5–10%of the maximum contraction to KPSS. Pre-contraction with AVP was necessary tounmask contractile effects of any weak vasoconstrictor agents.
2.8. Drugs
Drugs (and suppliers) were as follows: atropine SO4 (Sigma, St. Louis, MO,USA); box jellyfish antivenom (CSL, Parkville, Vic., Australia); CGRP8–37 (rat; Aus-pep, Parkville, Vic., Australia); �-conotoxin GVIA (Auspep); noradrenaline bitartrate(Sigma); prazosin HCl (Sigma); propranolol HCl (Sigma); and tetrodotoxin (TTX; Sap-phire Bioscience, Sydney, NSW, Australia). CVE aliquots were stored at −70 ◦C untiluse and 0.1 mmol/L aliquots of CGRP8–37, TTX and �-conotoxin GVIA at −20 ◦C.
2.9. Statistical analyses
Data in text, tables and figures are the mean ± standard error of the mean(S.E.M.) of n experiments. For rat isolated mesenteric artery data, sigmoidalconcentration–response curves were fitted by Prism 5 (GraphPad Software, San
Diego, California, USA). The negative logarithm of the CVE concentration required tocause 50% of the maximal response (pEC50) was compared in the absence (control)and presence of pre-treatments using one-way analysis of variance (ANOVA), withDunnett’s post test for multiple comparisons as appropriate (Prism 5). Rat isolatedleft atrial force, or right atrial rate, data were compared within and between groupsby repeated measures ANOVA with Greenhouse–Geisser correction for correlation(Ludbrook, 1994), using SuperANOVA 1.11 for Mac (Abacus Concepts, Berkeley, CA,USA). Atrial baselines or maximal changes in force or rate were compared betweengroups using one-way ANOVA with Dunnett’s post test for multiple comparisons.The effect of pre-treatment on baseline atrial values were compared within group bypaired Student’s t test. In all cases, P < 0.05 was accepted as statistically significant.
3. Results
3.1. Jellyfish morphology, CVE yield, protein content andcatecholamines
For these experiments, 5 batches of M. maxima CVE were pre-pared. The overall appearance of a typical M. maxima jellyfish isshown in Fig. 1a. A close-up of the dominant tentacle nematocysts
Fig. 1. (a) Photograph of a Malo maxima jellyfish in situ offshore Broome, Western Australia, showing the characteristic tall, narrow and robust body with a flattened apexof the medusa or bell (5 cm diameter at base), and the four retracted tentacles. (b) A close-up of the dominant nematocysts along the length of the tentacles. Originalmagnification 40× objective (i.e. 400× magnification). (c) Two isolated examples of these tentacular nematocysts showing the spines orientated away from the capsule (thenematocyst is approximately 40 × 15 �m wide; lower left white scale bar represents 10 �m). (d) Another isolated nematocyst, without obvious spine, from a tentacle squashusing the same magnification (lower left white scale bar represents 10 �m).

Author's personal copy
224 R. Li et al. / Toxicology Letters 201 (2011) 221–229
Fig. 2. SDS-PAGE (non-reducing conditions) comparison of each batch of Malo maxima and Chironex fleckeri venom extract used in these experiments. Lane M, BenchMarkTM
Pre-Stained Protein Ladder; Lanes 1–5, M. maxima CVE corresponding to batches 1–5; Lane 6, C. fleckeri nematocyst extract.
is shown in Fig. 1b (photographed using a Leica DMLB compoundmicroscope and Leica IM-50 Image Manager v.1.20 for Windows),with the morphology of isolated nematocysts presented at highermagnification (Fig. 1c and d). The protein yield of the typical batchof 5 pooled M. maxima jellyfish was 11 mg/mL prepared in 5 mL PBS.
SDS-PAGE analysis revealed that the CVE contained at least 6major constituent proteins and a large number of minor proteinbands (Fig. 2). The major protein species occurred at approximately40, 50, 60 and 80 kDa, with at least one major band between 115.5and 181.8 kDa. Minor protein bands were present at 20–30 kDa and35 kDa; less well differentiated minor protein bands occurred at<14.8 kDa. Each of the 5 batches of M. maxima CVE had consistentSDS-PAGE profiles and contained similar proportions of each of theprotein components.
Concerning catecholamines, neither noradrenaline nor itsmethylation product adrenaline were detected by electrochemicalassessment of the M. maxima CVE itself, but the precursor, DOPA(dihydroxyphenylalanine) was detected at a level of approximately20 ng/mL venom extract (data not shown; personal communica-tion, Dr Gavin Lambert, Baker IDI Heart and Diabetes Institute).
3.2. Rat isolated left atria
None of the pre-treatments significantly affected baseline forceof contraction in rat isolated electrically driven left atria and valueswere similar between groups prior to addition of M. maxima CVE(P > 0.05); the pooled baseline was 0.48 ± 0.03 g (n = 34). M. maximaCVE caused concentration-dependent positive inotropic responsesin the vehicle group (n = 6; Fig. 3a); contractile force increasedfrom baseline by 0.46 ± 0.07 g at 100 �g/mL. Atropine (1 �M; n = 6)did not affect CVE-induced inotropic responses (P > 0.05; Fig. 3a).However, pre-treatment with either propranolol (1 �M; n = 7), TTX(0.1 �M; n = 6), an increased concentration of magnesium (6 mMcompared to 1.2 mM in normal PSS; n = 4) or CGRP8–37 (1 �M; n = 5)significantly attenuated the inotropic responses to CVE (P = 0.0001compared with the vehicle group; Fig. 3).
3.3. Rat isolated right atria
Sixty min pre-treatment with propranolol (1 �M) causeda decrease in rat spontaneously beating right atrial rate of
1 3 10 30 100
0.0
0.2
0.4
0.6a
*
*
Lef
tatr
ialf
orc
e g
1 3 10 30 100
0.0
0.2
0.4
0.6b
**
Malomaxima CVE (µg/mL)
Lef
tatr
ialf
orc
e g
Fig. 3. Concentration-dependent effects of Malo maxima crude venom extract (CVE;�g/mL) on rat left atrial force. Data are shown as change (�) from baseline force ofcontraction after 60 min pre-treatment with (a) vehicle (�, PSS, n = 6); 1 �M atropine(©, n = 6); 1 �M propranolol (�, n = 7); or 1 �M CGRP8–37 (�, n = 5); or (b) vehicle (�,PSS, n = 6); 0.1 �M TTX (�, n = 6); or 6 mM Mg2+ (♦, n = 4). Error bars are ±1 S.E.M.;where no error bar is visible the S.E.M. is within the symbol representing the mean.*P < 0.05 compared to vehicle control group (repeated measured ANOVA). n, numberof atria.
Author's personal copy
R. Li et al. / Toxicology Letters 201 (2011) 221–229 225
1 3 10 30 100
-40
-30
-20
-10
0
10
20
30
*
Malomaxima CVE (µg/mL)
Ra
trig
hta
tria
lra
te(b
/min
)
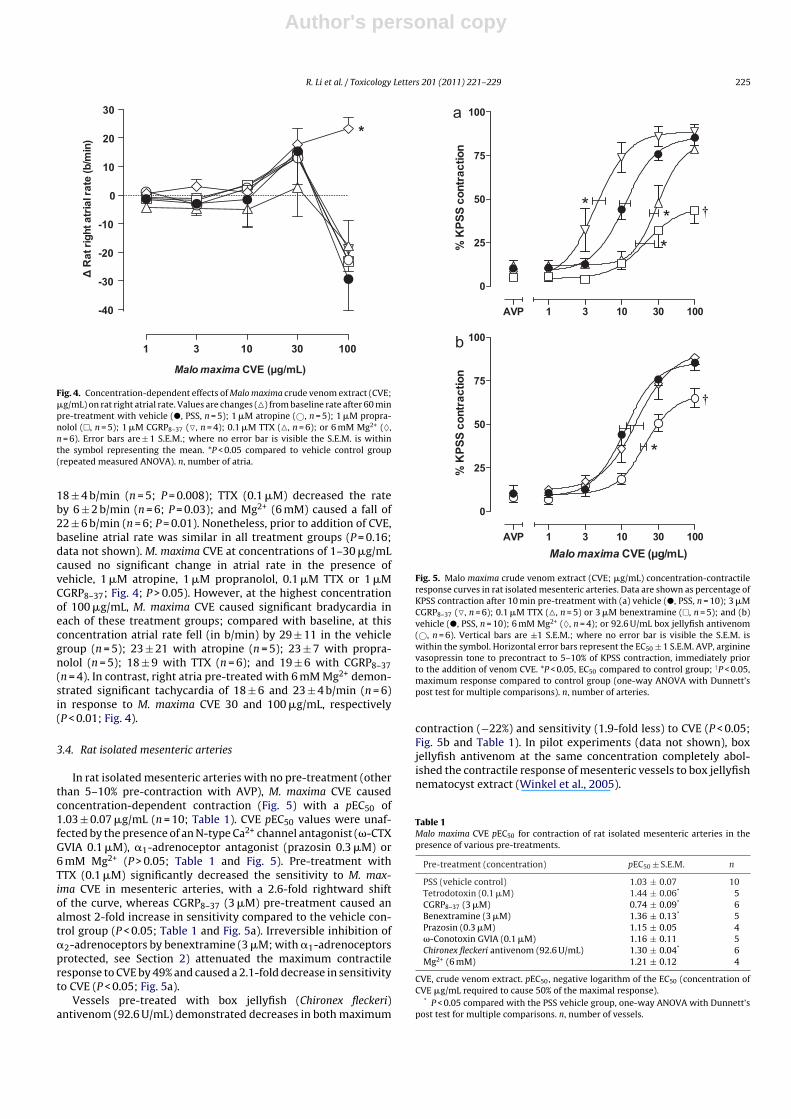
Fig. 4. Concentration-dependent effects of Malo maxima crude venom extract (CVE;�g/mL) on rat right atrial rate. Values are changes (�) from baseline rate after 60 minpre-treatment with vehicle (�, PSS, n = 5); 1 �M atropine (©, n = 5); 1 �M propra-nolol (�, n = 5); 1 �M CGRP8–37 (�, n = 4); 0.1 �M TTX (�, n = 6); or 6 mM Mg2+ (♦,n = 6). Error bars are ± 1 S.E.M.; where no error bar is visible the S.E.M. is withinthe symbol representing the mean. *P < 0.05 compared to vehicle control group(repeated measured ANOVA). n, number of atria.
18 ± 4 b/min (n = 5; P = 0.008); TTX (0.1 �M) decreased the rateby 6 ± 2 b/min (n = 6; P = 0.03); and Mg2+ (6 mM) caused a fall of22 ± 6 b/min (n = 6; P = 0.01). Nonetheless, prior to addition of CVE,baseline atrial rate was similar in all treatment groups (P = 0.16;data not shown). M. maxima CVE at concentrations of 1–30 �g/mLcaused no significant change in atrial rate in the presence ofvehicle, 1 �M atropine, 1 �M propranolol, 0.1 �M TTX or 1 �MCGRP8–37; Fig. 4; P > 0.05). However, at the highest concentrationof 100 �g/mL, M. maxima CVE caused significant bradycardia ineach of these treatment groups; compared with baseline, at thisconcentration atrial rate fell (in b/min) by 29 ± 11 in the vehiclegroup (n = 5); 23 ± 21 with atropine (n = 5); 23 ± 7 with propra-nolol (n = 5); 18 ± 9 with TTX (n = 6); and 19 ± 6 with CGRP8–37(n = 4). In contrast, right atria pre-treated with 6 mM Mg2+ demon-strated significant tachycardia of 18 ± 6 and 23 ± 4 b/min (n = 6)in response to M. maxima CVE 30 and 100 �g/mL, respectively(P < 0.01; Fig. 4).
3.4. Rat isolated mesenteric arteries
In rat isolated mesenteric arteries with no pre-treatment (otherthan 5–10% pre-contraction with AVP), M. maxima CVE causedconcentration-dependent contraction (Fig. 5) with a pEC50 of1.03 ± 0.07 �g/mL (n = 10; Table 1). CVE pEC50 values were unaf-fected by the presence of an N-type Ca2+ channel antagonist (�-CTXGVIA 0.1 �M), �1-adrenoceptor antagonist (prazosin 0.3 �M) or6 mM Mg2+ (P > 0.05; Table 1 and Fig. 5). Pre-treatment withTTX (0.1 �M) significantly decreased the sensitivity to M. max-ima CVE in mesenteric arteries, with a 2.6-fold rightward shiftof the curve, whereas CGRP8–37 (3 �M) pre-treatment caused analmost 2-fold increase in sensitivity compared to the vehicle con-trol group (P < 0.05; Table 1 and Fig. 5a). Irreversible inhibition of�2-adrenoceptors by benextramine (3 �M; with �1-adrenoceptorsprotected, see Section 2) attenuated the maximum contractileresponse to CVE by 49% and caused a 2.1-fold decrease in sensitivityto CVE (P < 0.05; Fig. 5a).
Vessels pre-treated with box jellyfish (Chironex fleckeri)antivenom (92.6 U/mL) demonstrated decreases in both maximum
AVP 1 3 10 30 100
0
25
50
75
100a
*
** †
%K
PS
Sc
on
tra
ctio
n
AVP 1 3 10 30 100
0
25
50
75
100b
*
†
Malomaxima CVE ( g/mL)
%K
PS
Sc
on
tra
ctio
n
Fig. 5. Malo maxima crude venom extract (CVE; �g/mL) concentration-contractileresponse curves in rat isolated mesenteric arteries. Data are shown as percentage ofKPSS contraction after 10 min pre-treatment with (a) vehicle (�, PSS, n = 10); 3 �MCGRP8–37 (�, n = 6); 0.1 �M TTX (�, n = 5) or 3 �M benextramine (�, n = 5); and (b)vehicle (�, PSS, n = 10); 6 mM Mg2+ (♦, n = 4); or 92.6 U/mL box jellyfish antivenom(©, n = 6). Vertical bars are ±1 S.E.M.; where no error bar is visible the S.E.M. iswithin the symbol. Horizontal error bars represent the EC50 ± 1 S.E.M. AVP, argininevasopressin tone to precontract to 5–10% of KPSS contraction, immediately priorto the addition of venom CVE. *P < 0.05, EC50 compared to control group; †P < 0.05,maximum response compared to control group (one-way ANOVA with Dunnett’spost test for multiple comparisons). n, number of arteries.
contraction (−22%) and sensitivity (1.9-fold less) to CVE (P < 0.05;Fig. 5b and Table 1). In pilot experiments (data not shown), boxjellyfish antivenom at the same concentration completely abol-ished the contractile response of mesenteric vessels to box jellyfishnematocyst extract (Winkel et al., 2005).
Table 1Malo maxima CVE pEC50 for contraction of rat isolated mesenteric arteries in thepresence of various pre-treatments.
Pre-treatment (concentration) pEC50 ± S.E.M. n
PSS (vehicle control) 1.03 ± 0.07 10Tetrodotoxin (0.1 �M) 1.44 ± 0.06* 5CGRP8–37 (3 �M) 0.74 ± 0.09* 6Benextramine (3 �M) 1.36 ± 0.13* 5Prazosin (0.3 �M) 1.15 ± 0.05 4�-Conotoxin GVIA (0.1 �M) 1.16 ± 0.11 5Chironex fleckeri antivenom (92.6 U/mL) 1.30 ± 0.04* 6Mg2+ (6 mM) 1.21 ± 0.12 4
CVE, crude venom extract. pEC50, negative logarithm of the EC50 (concentration ofCVE �g/mL required to cause 50% of the maximal response).
* P < 0.05 compared with the PSS vehicle group, one-way ANOVA with Dunnett’spost test for multiple comparisons. n, number of vessels.

Author's personal copy
226 R. Li et al. / Toxicology Letters 201 (2011) 221–229
Table 2Comparison of Malo maxima and Carukia barnesi CVE in vitro.
Pre-treatment Malo maxima CVE Carukia barnesi CVE
Rat isolated right atriaVehicle (control) Nil or bradycardiaa Tachycardia (++)Atropine (1 �M) No change from control Tachycardia (+++)Propranolol (1 �M) No change from control BradycardiaTTX (0.1 �M) No change from control Inhibition (+)CGRP8–37 (1 �M) No change from control Not testedMg2+ (6 mM) Tachycardiab Not testedRat isolated left atriaVehicle (control) Inotropic (++) Inotropic (++)Atropine (1 �M) Inotropic (++) Inotropic (+++)Propranolol (1 �M) Inotropic (+) Not testedTTX (0.1 �M) Inotropic (+) Not testedCGRP8–37 (1 �M) Inotropic (+) Not testedMg2+ (6 mM) Inotropic (+) Not testedRat isolated mesenteric arteriesVehicle (control) Contraction ContractionPrazosin (0.3 �M) No change from control No change from controlTTX (0.1 �M) 2.6-fold rightward shift No change from controlCGRP8–37 (3 �M) 2.0-fold leftward shift Not testedMg2+ (6 mM) No change from control Not tested�-Conotoxin GVIA (0.1 �M) No change from control No change from controlChironex fleckeri antivenom (92.6 U/mL) 1.9-fold rightward shift; 22% decrease in maximum contraction No change from controlBenextramine (3 �M) 2.1-fold rightward shift; 49% decrease in maximum contraction Not tested
The relative degree of chronotropic or inotropic response is represented by ‘+’, where (++) is the response in the control group, (+++) represents a more pronounced responseand (+) represents a less pronounced response.
a Bradycardia only occurred at the highest concentration of M. maxima CVE (100 �g/mL).b Tachycardia only occurred at 30–100 �g/mL. C. barnesi results are from (Winkel et al., 2005).
4. Discussion
This study provides the first experimental evidence that M. max-ima may be responsible for Irukandji syndrome in the Broomeregion of Western Australia. Specifically these data support thehypothesis that M. maxima CVE causes catecholamine releasein vitro, a characteristic effect of the other Irukandji venoms exam-ined to date – C. barnesi and A. mordens (Ramasamy et al., 2005;Winkel et al., 2005; Winter et al., 2008). As such it is the firstexperimental evidence implicating a jellyfish species from outsideQueensland as a cause of the Irukandji syndrome.
Further, the findings of this first biochemical and pharmacolog-ical analysis of the venom from the jellyfish M. maxima allow fora comparison of its effects on cardiac and vascular tissues in vitrowith those of the archetype Irukandji jellyfish, C. barnesi (Table 2).Considering the findings in detail, it was observed that the M. max-ima CVE-induced positive inotropic response in rat left atria waspartially inhibited by the �1-adrenoceptor antagonist propranolol,suggesting release of neuronal noradrenaline, as only DOPA wasdetected in the venom sample itself and this precursor of nora-drenaline had no effect on either �- or �-adrenoceptors (data notshown). This positive inotropic response was more markedly atten-uated by the voltage-gated sodium channel inhibitor tetrodotoxinand sensory peptide antagonist CGRP8–37. So, in left atrial tissues,the concentration-dependent increase in the force of contractionappears to be the net result of the positive inotropic effects ofboth noradrenaline and CGRP. Mg2+ (6 mM) was also very effec-tive at attenuating the positive inotropic effects of M. maxima CVE.However, the current guidelines for the emergency management ofthe Irukandji syndrome recommend the use of the �-adrenoceptorantagonist, phentolamine, rather than propranolol (Pereira et al.,2007). Further, in an examination of the cardiovascular effectsof the other Queensland Irukandji jellyfish, A. mordens, �- butnot �-adrenoceptor blockade significantly attenuated the pressorresponse to intravenous administration of the venom in anaes-thetised rats (Winter et al., 2008). M. maxima CVE does not appearto stimulate the release of significant amounts of acetylcholine, dif-fering from the effects of C. barnesi CVE which demonstrated clear
parasympathetic involvement in atrial tissues (Table 2) (Winkelet al., 2005).
Given that the concentration–response curve to M. maxima CVEwas attenuated (or slope depressed) by propranolol alone and morestrongly by the CGRP antagonist (alone), it could be assumed thatthe toxin is causing the release of neural transmitter noradrenalineand neural peptide CGRP. If we consider the force response as 100%at CVE 100 �g/mL (with PSS pre-treatment), then crudely speakingnoradrenaline release might be responsible for 40% of the maxi-mum inotropic response and CGRP release for 60% of the maximumresponse. How the toxin causes the release of transmitter nora-drenaline from sympathetic nerves and CGRP from sensory nerveendings is uncertain, but the TTX and Mg2+ experiments show thatmuch of the inotropic response to the toxin (≈80%) is decreasedby these treatments. Thus, a working hypothesis may suggest thatM. maxima CVE causes a Na+ channel-opening response on theadrenergic and peptidergic sensory varicosities releasing endoge-nous noradrenaline and CGRP (and possibly neuropeptide Y; nottested). The residual (≈20%) inotropic response to the M. maximaCVE in the presence of TTX or Mg2+ could be explained by somedirect post-junctional stimulation of �1-adrenoceptors or CGRPreceptors.
In the right atrial tissue, the weak chronotropic response toM. maxima CVE (30 �g/mL) of about 15 beats/min contrasts withthe ≈80 beats/min tachycardia caused by 10 �g/mL C. barnesi CVE(Winkel et al., 2005). This weak tachycardic response to M. maximaCVE (30 �g/mL) was inhibited by TTX pre-treatment. Importantly,M. maxima CVE showed a significant bradycardia at 100 �g/mL thatwas not significantly affected by TTX. However, this bradycardiawas completely blocked by Mg2+ suggesting that in severe enven-omation with M. maxima, Mg2+ administration would protect theheart rate and thus support cardiac output.
In small mesenteric arteries the neural innervation ofsympathetic and sensory nerves cause vasoconstriction and vasodi-latation, respectively, in contrast to the similar inotropic responsesfrom these neural networks in the heart. With this in mind, if M.maxima CVE causes release of both sympathetic (noradrenaline andneuropeptide Y) and sensory transmitters (CGRP) as in the heart,

Author's personal copy
R. Li et al. / Toxicology Letters 201 (2011) 221–229 227
the “control” curve to the toxin will be the algebraic sum of bothconstrictor and dilator stimuli noradrenaline and neuropeptide Ycompeting with CGRP, respectively. This hypothesis is borne outwhen the “control” curve to M. maxima CVE in mesenteric arter-ies was significantly shifted to the left by the CGRP antagonist,CGRP8–37. This curve represents the effects (without CGRP) of con-strictor stimuli (noradrenaline and neuropeptide Y) where 80% ofthe maximum effect to the toxin occurs at 10 �g/mL. Further, TTXabolishes the contraction of 10 �g/mL M. maxima CVE suggestinga neural prejunctional Na+ channel release-dependency. Similarly,Mg2+ would block the release of CGRP and, to a degree, the release ofnoradrenaline. This interpretation of a nil effect of Mg2+ comparedwith PSS pretreatment must bear in mind the functional antag-onism of the dilator and constrictor stimuli. The neural releaseof both noradrenaline and CGRP in vascular tissues is dependentupon N-type calcium channels (Brain and Grant, 2004; De Meyet al., 2008; Pruneau and Angus, 1990). Inhibition of this channelwith �-conotoxin GVIA appeared to equally inhibit the respec-tive agonist contractile and relaxant effects, resulting in the sameconcentration-dependent M. maxima CVE contraction of mesen-teric arteries as observed in the control group. Thus, mesentericartery contraction by M. maxima CVE seems to be N-type calciumchannel-independent.
To elucidate the role of noradrenaline we tested prazosin toantagonise �1-adrenoceptors with no effect while the irreversible�2-adrenoceptor antagonist benextramine was effective in shift-ing the mesenteric artery M. maxima CVE concentration–responsecurve to the right and markedly attenuating the maximum plateauresponse. These experiments suggest that perhaps the major com-ponent to sympathetic vasoconstrictors is via �2-adrenoceptorswith neuropeptide Y co-release making up the powerful contrac-tion response. We used a specific pretreatment regimen to protect�1-adrenoceptors when applying the benextramine so that theantagonist only irreversibly blocked �2-adrenoceptors. In anec-dotal experiments, we have since tested benextramine (withoutprazosin protection of �1-adrenoceptors, i.e. to block both �1- and�2-adrenoceptors irreversibly) alone, as well as in combinationwith CGRP8–37; the contractile responses to M. maxima CVE (datanot shown) were similar to those observed with �2-adrenoceptorblockade (Fig. 5a).
This result reconfirms the potential role for �-adrenoceptorantagonists in the clinical management of this syndrome. How-ever, given severe cases can be associated with a late phase ofcardiac failure, hypotension and pulmonary oedema, a reversible�-adrenoceptor antagonist, with a short half-life, phentolamine,rather than phenoxybenzamine, an irreversible �1 and �2-adrenoceptor antagonist, is currently recommended for thetreatment of Irukandji syndrome-induced persistent hypertensionthat is otherwise uncontrolled by nitrates (Pereira et al., 2007).Phentolamine, a non-specific �1- and �2-adrenoceptor antagonist,has also been used successfully in the treatment of sympatheticallymediated pain syndromes (Hord et al., 2001; Phillips et al., 2006).This characteristic may contribute an anti-nociceptive function inits role in the medical management of the Irukandji syndrome(Pereira et al., 2007).
In comparison to the efficacy of box jellyfish (C. fleckeri)antivenom on C. fleckeri venom, it appears that M. maxima CVEis poorly neutralised by C. fleckeri antivenom. Similarly, C. fleckeriantivenom has little, if any, effect on C. barnesi CVE (Winkel et al.,2005) nor on A. mordens venom in vitro (Winter et al., 2008) or intreating Irukandji stings (Fenner et al., 1986; Winkel et al., 2003).The decreased maximum and sensitivity of contraction observedin rat mesenteric arteries pre-treated with C. fleckeri antivenommay be due to non-specific binding of M. maxima CVE to the ovineproteins constituent in C. fleckeri antivenom, rather than specifichomology between M. maxima and C. fleckeri venom.
Due to the absence of an Irukandji antivenom, and the evidenceof catecholamine excess, magnesium therapy has been used as atherapeutic approach to treating Irukandji syndrome (Corkeronet al., 2004; Corkeron, 2003). These results support the anecdo-tal clinical evidence that intravenous magnesium infusion may beeffective in treating the pain and hypertension that occurs in thissyndrome. Specifically, the in vitro evidence suggests that mag-nesium acts to decrease cardiac contractility, rather than causevasodilatation, at a clinically relevant concentration (6 mM).
These results also offer a plausible explanation for the anal-gesic effects of magnesium infusion in Irukandji syndrome. Thesestudies are the first to specifically investigate the role of sensoryneurotransmitters in an experimental model of the cardiovascu-lar effects of the syndrome. A logical rationale would suggest thatmagnesium, as a divalent cation, acts as a competitive antago-nist to calcium ions, decreasing pre-synaptic calcium influx insensory nerves, thereby attenuating the release of CGRP. Addition-ally, CGRP itself increases post-synaptic l-type calcium channelcurrent through an adenylate cyclase and cyclic AMP pathway,a process that may be attenuated by an increased concentrationof magnesium ions (Bell and McDermott, 1994). Influx of Ca2+
through TRPV1 channels may also cause CGRP release, providing yetanother mechanism that might be antagonised by magnesium ions(Docherty et al., 1996). Indeed recent studies have provided evi-dence for desensitization-dependent TRPV1 activation caused bysome cnidarian venoms, including the cubozoan C. fleckeri (Cuyperset al., 2006).
The extraction technique used to produce the crude venomis relevant. Grinding whole jellyfish specimens may produce anextract that includes components with clinically irrelevant, butpharmacologically interesting effects, such as those from tenta-cles. Previous studies have found that although C. barnesi tentacularextracts appear to have distinct pharmacological actions, the mainpharmacological activity present in tentacle extracts reflects thatof nematocyst contents (Calton and Burnett, 1986; Ramasamyet al., 2005). However, anecdotal evidence suggests that C. barnesibell nematocyst venom appears to have at least local effects andobtaining venom from tentacular nematocysts ignores the possibleinvolvement of bell nematocysts which are present on all describedIrukandji species (Barnes, 1964; Gershwin, 2005, 2007; Gershwinand Alderslade, 2005; Southcott, 1967). Clearly future studies of M.maxima should assess whether all of the activities described hereare attributable to pure nematocyst-derived venom.
The M. maxima SDS-PAGE analysis revealed many similaritieswith the Irukandji species C. barnesi. SDS-PAGE protein profiles ofC. barnesi CVE showed a number of major protein bands, occur-ring at approximately 40, 45, 50, 80, 106 and >106 kDa (Wiltshireet al., 2000). M. maxima CVE contains major bands coinciding withall major proteins in C. barnesi CVE, except at 50 kDa where a minorprotein species exists. However, since these two venoms havenot been simultaneously run on the same gel these comparisonsare qualitative and SDS-PAGE alone is insufficient to determinewhether chemical or functional similarities exist. These results areconsistent with the complexity and spread of proteins extracted byothers from A. mordens and C. fleckeri venoms (Winter et al., 2008).
The potential significance of the detection of DOPA in theM. maxima extract is unclear as this amino acid is very widelydistributed in the tissues and fluids of invertebrates, includ-ing cnidarians (Waite, 1992). Interestingly, only DOPA andrelated metabolites (5-hydroxy-DOPA and cysteinyl-DOPA) havebeen identified in the nervous systems of some cnidarians andctenophores (Carlberg, 1988; Carlberg and Elofsson, 1987). Cer-tainly no pharmacological activity was attributable to l-DOPA wheninvestigated using the rat isolated right and left atria and themesenteric artery at the concentrations detected in the M. maximavenom extract (data not shown).

Author's personal copy
228 R. Li et al. / Toxicology Letters 201 (2011) 221–229
While the in vitro assays in this study utilised mammalian car-diac and vascular tissues, the results would be further validated byextending this investigation to more clinically relevant tissues suchas human atrial trabeculae preparations in vitro or anaesthetisedlaboratory animals in vivo. The clinical significance of the appar-ently lower in vitro potency of M. maxima venom extract, comparedto that of C. barnesi, on cardiac and vascular tissues in vitro, remainsto be determined. A similar, lesser potency of A. mordens venom,compared with that of C. barnesi, was seen in studies of anaes-thetised rats by others (Winter et al., 2008).
Current evidence suggests that Irukandji syndrome occurring inthe Broome region of Western Australia may differ from that seen innorth Queensland (Macrokanis et al., 2004). For example, clinicalaudits of the presenting features of the syndrome in both local-ities suggest a higher rate of narcotic analgesia requirements forthe Broome patients (Little and Mulcahy, 1998; Macrokanis et al.,2004). This also appeared to be accompanied by the highest pre-sentation rate, in Broome, for Irukandji syndrome yet defined inAustralia. On the other hand, the same audit found a near identi-cal rate of hypertension (around 50% of patients) and pulmonaryoedema (around 3%) in both localities and a higher rate of admis-sion for the Cairns hospital cases (28%) compared with Broome(17%) (Little and Mulcahy, 1998; Macrokanis et al., 2004). Hence,until there are high quality, long-term prospective clinical studiesusing similar outcome measures, it cannot be said that WesternAustralian Irukandji syndrome differs significantly from that seenin north Queensland. Likewise our studies that appear to confirmthe probability that M. maxima is a cause of Irukandji syndrome inWestern Australia and that similar treatment protocols as currentlyrecommended for the syndrome, based on Queensland experience(Pereira et al., 2007), should apply to cases presenting to Broomehospital. Future studies of all Irukandji venoms should focus ondefining the precise molecular nature of the toxins and the relevanteffector pathways underlying the syndrome.
Conflict of interest statement
None.
Funding
Lisa-Ann Gershwin is primarily funded by EnvironmentAustralia (ABRS Grant # 208-82) for taxanomy. These specimenswere collected in association with the Pearl Producers Association& Paspaley Pearling Co.
Acknowledgements
The authors wish to thank Mr Mark Ross-Smith for his technicalexpertise in the isolated mesenteric artery myograph experiments.We also thank Dr Gavin Lambert and Ms Reena Chopra of theHuman Neurotransmitters Laboratory, Baker IDI Heart and Dia-betes Institute, Melbourne, for undertaking the catecholamineassays. We thank Dr Angel Yanagihara, Bekesy Laboratory of Neu-robiology, and Asia Pacific Institute of Tropical Medicine, Universityof Hawaii at Manoa, Hawaii, for the photographs of M. maxima iso-lated nematocysts as well as Mark Alexander, Paspaley PearlingCompany, for his photograph of M. maxima. The Australian VenomResearch Unit is funded by the Australian Government Departmentof Health and Ageing.
References
Angus, J.A., Broughton, A., Mulvany, M.J., 1988. Role of alpha-adrenoceptors in con-strictor responses of rat, guinea-pig and rabbit small arteries to neural activation.J. Physiol. 403, 495–510.
Angus, J.A., Wright, C.E., 2000. Techniques to study the pharmacodynamics ofisolated large and small blood vessels. J. Pharmacol. Toxicol. Methods 44,395–407.
Baguley, I.J., 2008. The excitatory:inhibitory ratio model (EIR model): an integrativeexplanation of acute autonomic overactivity syndromes. Med. Hypotheses 70,26–35.
Barnes, J.H., 1964. Cause and effect in Irukandji stingings. Med. J. Aust. 1, 897–904.
Bell, D., McDermott, B.J., 1994. Calcitonin gene-related peptide stimulates a positivecontractile response in rat ventricular cardiomyocytes. J. Cardiovasc. Pharmacol.23, 1011–1021.
Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of micro-gram quantities of protein utilizing the principle of protein–dye binding. Anal.Biochem. 72, 248–254.
Brain, S.D., Grant, A.D., 2004. Vascular actions of calcitonin gene-related peptide andadrenomedullin. Physiol. Rev. 84, 903–934.
Burnett, J.W., Currie, B., Fenner, P., Rifkin, J., Williamson, J., 1996. Cubozoans (‘Boxjellyfish’). In: Williamson, J., Fenner, P., Burnett, J.W., Rifkin, J. (Eds.), Venomousand Poisonous Marine Animals. University of New South Wales Press, Sydney,pp. 236–283.
Burnett, J.W., Weinrich, D., Williamson, J.A., Fenner, P.J., Lutz, L.L., Bloom, D.A., 1998.Autonomic neurotoxicity of jellyfish and marine animal venoms. Clin. Auton.Res. 8, 125–130.
Calton, G.J., Burnett, J.W., 1986. Partial purification of Chironex fleckeri (sea wasp)venom by immunochromatography with antivenom. Toxicon 24, 416–420.
Carlberg, M., 1988. Localisation of catechol compounds in the ctenophore Mnemiop-sis leidyi. Comp. Biochem. Physiol. 91C, 69–74.
Carlberg, M., Elofsson, R., 1987. Presence of 3,4-dihydroxyphenylalanine and 3,4,5-trihydroxyphenylalanine in a coelenterate nervous system: possible tyrosinase-mediated accumulation. Neurochem. Int. 11, 161–167.
Corkeron, M., Pereira, P., Makrocanis, C., 2004. Early experience with magnesiumadministration in Irukandji syndrome. Anaesth. Intensive Care 32, 666–669.
Corkeron, M.A., 2003. Magnesium infusion to treat Irukandji syndrome. Med. J. Aust.178, 411.
Cuypers, E., Yanagihara, A., Karlsson, E., Tytgat, J., 2006. Jellyfish and other cnidar-ian envenomations cause pain by affecting TRPV1 channels. FEBS Lett. 580,5728–5732.
De Mey, J.G., Megens, R., Fazzi, G.E., 2008. Functional antagonism betweenendogenous neuropeptide Y and calcitonin gene-related peptide in mesentericresistance arteries. J. Pharmacol. Exp. Ther. 324, 930–937.
de Pender, A.M., Winkel, K.D., Ligthelm, R.J., 2006. A probable case of Irukandjisyndrome in Thailand. J. Travel Med. 13, 240–243.
Docherty, R.J., Yeats, J.C., Bevan, S., Boddeke, H.W., 1996. Inhibition of calcineurininhibits the desensitization of capsaicin-evoked currents in cultured dorsal rootganglion neurones from adult rats. Pflugers Arch. 431, 828–837.
Fenner, P.J., Fitzpatrick, P.F., Hartwick, R.J., Skinner, R., 1985. Morbakka, anothercubomedusan. Med. J. Aust. 143, 550–551, 554–555.
Fenner, P.J., Hadok, J.C., 2002. Fatal envenomation by jellyfish causing Irukandjisyndrome. Med. J. Aust. 177, 362–363.
Fenner, P.J., Williamson, J., Callanan, V.I., Audley, I., 1986. Further understanding of,and a new treatment for, “Irukandji” (Carukia barnesi) stings. Med. J. Aust. 145,569, 572–574.
Fenner, P.J., Williamson, J.A., Burnett, J.W., Colquhoun, D.M., Godfrey, S., Gunawar-dane, K., Murtha, W., 1988. The “Irukandji syndrome” and acute pulmonaryoedema. Med. J. Aust. 149, 150–156.
Flecker, H., 1952. Irukandji sting to North Queensland bathers without productionof weals but with severe general symptoms. Med. J. Aust. 2, 89–91.
Gershwin, L., 2005. Two new species of jellyfishes (Cnidaria: Cubozoa: Caryb-deida) from tropical Western Australia, presumed to cause Irukandji Syndrome.Zootaxa, 1–30.
Gershwin, L., 2007. Malo kingi: a new species of Irukandji jellyfish (Cnidaria:Cubozoa: Carybdeida), possibly lethal to humans, from Queensland, Australia.Zootaxa, 55–68.
Gershwin, L., Alderslade, P., 2005. A new genus and species of box jellyfish (Cubozoa:Carybdeida) from tropical Australian waters. The Beagle 21, 26–27.
Gershwin, L.A., 2008. Morbakka fenneri, a new genus and species of Irukandji jelly-fish (Cnidaria: Cubozoa). Mem. Qld Mus. – Nat. 54, 23–33.
Grady, J.D., Burnett, J.W., 2003. Irukandji-like syndrome in South Florida divers. Ann.Emerg. Med. 42, 763–766.
Hord, A.H., Denson, D.D., Stowe, B., Haygood, R.M., 2001. �-1 and �-2 adrener-gic antagonists relieve thermal hyperalgesia in experimental mononeuropathyfrom chronic constriction injury. Anesth. Analg. 92, 1558–1562.
Huynh, T.T., Seymour, J., Pereira, P., Mulcahy, R., Cullen, P., Carrette, T., Little, M.,2003. Severity of Irukandji syndrome and nematocyst identification from skinscrapings. Med. J. Aust. 178, 38–41.
Lambert, G.W., Jonsdottir, I.H., 1998. Influence of voluntary exercise on hypothala-mic norepinephrine. J. Appl. Physiol. 85, 962–966.
Little, M., Mulcahy, R.F., 1998. A year’s experience of Irukandji envenomation in farnorth Queensland. Med. J. Aust. 169, 638–641.
Little, M., Mulcahy, R.F., Wenck, D.J., 2001. Life-threatening cardiac failure in ahealthy young female with Irukandji syndrome. Anaesth. Intensive Care 29,178–180.
Little, M., Pereira, P., Carrette, T., Seymour, J., 2006. Jellyfish responsible for Irukandjisyndrome. Qld J. Med. 99, 425–427.
Little, M., Seymour, J., 2003. Another cause of “Irukandji stingings”. Med. J. Aust. 179,654.

Author's personal copy
R. Li et al. / Toxicology Letters 201 (2011) 221–229 229
Ludbrook, J., 1994. Repeated measurements and multiple comparisons in cardiovas-cular research. Cardiovasc. Res. 28, 303–311.
Macrokanis, C.J., Hall, N.L., Mein, J.K., 2004. Irukandji syndrome in northern WesternAustralia: an emerging health problem. Med. J. Aust. 181, 699–702.
Mulvany, M.J., Halpern, W., 1977. Contractile properties of small arterial resistancevessels in spontaneously hypertensive and normotensive rats. Circ. Res. 41,19–26.
Nakashima, A., Angus, J.A., Johnston, C.I., 1982. Chronotropic effects of angiotensin I,angiotensin II, bradykinin and vasopressin in guinea pig atria. Eur. J. Pharmacol.81, 479–485.
O’Reilly, G.M., Isbister, G.K., Lawrie, P.M., Treston, G.T., Currie, B.J., 2001. Prospectivestudy of jellyfish stings from tropical Australia, including the major box jellyfishChironex fleckeri. Med. J. Aust. 175, 652–655.
Ohtsuka, S., Oyake, Y., Seo, Y., Eda, K., Yamaguchi, I., 2002. Magnesium sulphateinfusion suppresses the cardiac release of noradrenaline during a handgrip stresstest. Can. J. Cardiol. 18, 133–140.
Pereira, P., Corkeron, M., Little, M., Farlow, D., 2007. Irukandji taskforce guidelines forthe emergency management of Irukandji syndrome. Prevention and ResponseWorking Group of the Irukandji Taskforce, Queensland Government.
Phillips, W.J., Tollefson, B., Johnson, A., Abell, T., Lerant, A., 2006. Relief of acutepain in chronic idiopathic gastroparesis with intravenous phentolamine. Ann.Pharmacother. 40, 2032–2036.
Pommier, P., Coulange, M., De Haro, L., 2005. Systemic envenomation by jellyfish inGuadeloupe: Irukandji-like syndrome? Med. Trop. (Mars) 65, 367–369.
Pruneau, D., Angus, J.A., 1990. �-Conotoxin GVIA is a potent inhibitor of sympatheticneurogenic responses in rat small mesenteric arteries. Br. J. Pharmacol. 100,180–184.
Ramasamy, S., Isbister, G.K., Seymour, J.E., Hodgson, W.C., 2005. The in vivo cardio-vascular effects of the Irukandji jellyfish (Carukia barnesi) nematocyst venomand a tentacle extract in rats. Toxicol. Lett. 155, 135–141.
Southcott, R.V., 1967. Revision of some Carybdeidae (Scyphozoa: Cubomedusae),including a description of the jellyfish responsible for the “Irukandji syndrome”.Aust. J. Zool. 15, 651–671.
Tibballs, J., Hawdon, G., Winkel, K., 2001. Mechanism of cardiac failure in Irukandjisyndrome and first aid treatment for stings. Anaesth. Intensive Care 29, 552.
Waite, J.H., 1992. The DOPA ephemera: a recurrent motif in invertebrates. Biol. Bull.183, 178–184.
Wiltshire, C.J., Sutherland, S.K., Fenner, P.J., Young, A.R., 2000. Optimization andpreliminary characterization of venom isolated from 3 medically importantjellyfish: the box (Chironex fleckeri), Irukandji (Carukia barnesi), and blubber(Catostylus mosaicus) jellyfish. Wilderness Environ. Med. 11, 241–250.
Winkel, K., Hawdon, G., Fenner, P., Gershwin, L., Collins, A.G., Tibballs, J., 2003. Jelly-fish antivenoms: past, present, and future. J. Toxicol. 22, 115–127.
Winkel, K.D., Tibballs, J., Molenaar, P., Lambert, G., Coles, P., Ross-Smith, M., Wilt-shire, C., Fenner, P.J., Gershwin, L.A., Hawdon, G.M., Wright, C.E., Angus, J.A., 2005.Cardiovascular actions of the venom from the Irukandji (Carukia barnesi) jelly-fish: effects in human, rat and guinea-pig tissues in vitro and in pigs in vivo. Clin.Exp. Pharmacol. Physiol. 32, 777–788.
Winter, K.L., Isbister, G.K., Schneider, J.J., Konstantakopoulos, N., Seymour, J.E., Hodg-son, W.C., 2008. An examination of the cardiovascular effects of an ‘Irukandji’jellyfish, Alatina nr mordens. Toxicol. Lett. 179, 118–123.
Yoshimoto, C.M., Yanagihara, A.A., 2002. Cnidarian (coelenterate) envenomations inHawai’i improve following heat application. Trans. R. Soc. Trop. Med. Hyg. 96,300–303.




















