
A comparative analysis of phloem exudate proteins from Cucumis melo, Cucumis sativus and Cucurbita...
8
Planta (Berl.) 130, 211-218 (1976) Planta by Springer-Verlag 1976 A Comparative Analysis of Phloem Exudate Proteins from Cucumis melo, Cucumis sativus and Cucurbita maxima by Polyacrylamide Gel Electrophoresis and Isoelectric Focusing * Dinkar D. Sabnis and James W. Hart Department of Botany, University of Aberdeen, Aberdeen AB9 2UD, U.K. Summary. Proteins in sieve tube exudate from Cucu- mis melo L., Cucumis sativus L. and Cucurbita maxima Duch. were analysed by gel electrophoresis and iso- electric focusing. Estimated molecular weights and iso- electric points for the major and minor proteins from each plant species are presented. Electrophoresis re- vealed striking differences between the protein complements of exudate from the two genera investi- gated. Similarly, although a few exudate proteins from the two species of Cucumis possessed identical molecular weights, several major proteins were pecu- liar to each species. Isoelectric focusing of proteins in exudate samples from the three plants confirmed the marked differences in their protein complements. Furthermore, focusing also revealed differences be- tween cultivars of Cueumis sativus. Both Cucumis sati- vus and Cucurbita maxima possessed relatively large amounts of basic proteins; these were absent in exu- date from Cucumis melo. The implications of these results are discussed in relation to present concepts regarding the interrelationships and possible function- al roles of P-proteins. Introduction Sieve elements in the phloem of angiosperms charac- teristically contain filamentous and tubular material, which collectively has been termed "P-protein" (Esau and Cronshaw, 1967; Cronshaw and Esau, 1968). De- spite the abundance and highly organized nature of the various types of P-protein, their functional roles within the sieve tube are still unknown. Ultrastructu- ral studies have described tubules ranging in diameter from 135 to 240 A in different species, and filaments ranging in diameter from 60 to 140 A (Cronshaw and * Abbreviations: DTT = dithiothreitol ; Bis = N,N'-methylenebis- acrylamide; SDS = sodium dodecyl sulphate; TCA = trichloroacetic acid Esau, 1968; Behnke, 1969; Parthasarathy and Mtihle- thaler, 1969; Evert and Deshpande, 1969; Wergin and Newcomb, 1970; Palevitz and Newcomb, 1971; Sabnis and Hart, 1973). The electron microscopic evi- dence has been interpreted as showing that the various forms of P-protein consist of the same or similar subunits, which can aggregate into different structures representing different stages of organization and de- velopment. Further, it has been proposed that the equilibria between states of P-protein may be gov- erned by factors in the internal environment, including pH, temperature and co-factors (Wergin and New- comb, 1970; Palevitz and Newcomb, 1971). Filaments, similar in appearance and dimensions to native P-protein filaments, have been seen in exu- date collected from the transversely cut phloem of Ricinus communis (Williamson, 1972), Cucurbita pepo (Walker and Thaine, 1971 ; Williamson, 1972), Cucur- bita maxima (Kollmann et al., 1970; Kleinig et al., 1971 ; Cronshaw et al., 1973), Cucumis sativus (Cron- shaw et al., 1973), Nicotiana glauca x suaveolens (Koll- mann et al., 1970), and Nicotiana tabacum (Cronshaw et al., 1973). The protein content of exudate ranges between 2 mg/ml in Ricinus (Hall and Baker, 1972) to 20 mg/ml in Cucurbita pepo (Walker and Thaine, 1971) and it is assumed that the "exudate represents the whole contents of living sieve cells rather than a pure stream of translocates" (Kollmann et al., 1970). Biochemical studies on the proteins in sieve tubes have been limited to analyses of exudate from C. maxima (Weber and Kleinig, 1971; Kleinig et al., 1971; Eschrich et al., 1971; Beyenbach et al., 1974; Weber et al., 1974) and C. pepo (Walker and Thaine, 1971 ; Walker, 1972). Each of these two plant species contains a basic protein which gels in the absence of sulphydryl-protecting agents such as 2-mercapto- ethanol and dithioerythritol in the collecting medium. However, in C. maxima the gelling protein has a mo- lecular weight of 116,000 daltons and constitutes ca. 40% of total protein in the exudate, whereas in C.
-
Upload
sunderland -
Category
Documents
-
view
3 -
download
0
Transcript of A comparative analysis of phloem exudate proteins from Cucumis melo, Cucumis sativus and Cucurbita...

Planta (Berl.) 130, 211-218 (1976) Planta �9 by Springer-Verlag 1976
A Comparative Analysis of Phloem Exudate Proteins from Cucumis melo, Cucumis sativus and Cucurbita maxima by Polyacrylamide Gel Electrophoresis and Isoelectric Focusing *
Dinkar D. Sabnis and James W. Hart
Department of Botany, University of Aberdeen, Aberdeen AB9 2UD, U.K.
Summary. Proteins in sieve tube exudate from Cucu- mis melo L., Cucumis sativus L. and Cucurbita maxima Duch. were analysed by gel electrophoresis and iso- electric focusing. Estimated molecular weights and iso- electric points for the major and minor proteins from each plant species are presented. Electrophoresis re- vealed striking differences between the protein complements of exudate from the two genera investi- gated. Similarly, although a few exudate proteins from the two species of Cucumis possessed identical molecular weights, several major proteins were pecu- liar to each species. Isoelectric focusing of proteins in exudate samples from the three plants confirmed the marked differences in their protein complements. Furthermore, focusing also revealed differences be- tween cultivars of Cueumis sativus. Both Cucumis sati- vus and Cucurbita maxima possessed relatively large amounts of basic proteins; these were absent in exu- date from Cucumis melo. The implications of these results are discussed in relation to present concepts regarding the interrelationships and possible function- al roles of P-proteins.
Introduction
Sieve elements in the phloem of angiosperms charac- teristically contain filamentous and tubular material, which collectively has been termed "P-protein" (Esau and Cronshaw, 1967; Cronshaw and Esau, 1968). De- spite the abundance and highly organized nature of the various types of P-protein, their functional roles within the sieve tube are still unknown. Ultrastructu- ral studies have described tubules ranging in diameter from 135 to 240 A in different species, and filaments ranging in diameter from 60 to 140 A (Cronshaw and
* A b b r e v i a t i o n s : DTT = dithiothreitol ; Bis = N,N'-methylenebis- acrylamide; SDS = sodium dodecyl sulphate; TCA = trichloroacetic acid
Esau, 1968; Behnke, 1969; Parthasarathy and Mtihle- thaler, 1969; Evert and Deshpande, 1969; Wergin and Newcomb, 1970; Palevitz and Newcomb, 1971; Sabnis and Hart, 1973). The electron microscopic evi- dence has been interpreted as showing that the various forms of P-protein consist of the same or similar subunits, which can aggregate into different structures representing different stages of organization and de- velopment. Further, it has been proposed that the equilibria between states of P-protein may be gov- erned by factors in the internal environment, including pH, temperature and co-factors (Wergin and New- comb, 1970; Palevitz and Newcomb, 1971).
Filaments, similar in appearance and dimensions to native P-protein filaments, have been seen in exu- date collected from the transversely cut phloem of Ricinus communis (Williamson, 1972), Cucurbita pepo (Walker and Thaine, 1971 ; Williamson, 1972), Cucur- bita maxima (Kollmann et al., 1970; Kleinig et al., 1971 ; Cronshaw et al., 1973), Cucumis sativus (Cron- shaw et al., 1973), Nicotiana glauca x suaveolens (Koll- mann et al., 1970), and Nicotiana tabacum (Cronshaw et al., 1973). The protein content of exudate ranges between 2 mg/ml in Ricinus (Hall and Baker, 1972) to 20 mg/ml in Cucurbita pepo (Walker and Thaine, 1971) and it is assumed that the "exudate represents the whole contents of living sieve cells rather than a pure stream of translocates" (Kollmann et al., 1970).
Biochemical studies on the proteins in sieve tubes have been limited to analyses of exudate from C. maxima (Weber and Kleinig, 1971; Kleinig et al., 1971; Eschrich et al., 1971; Beyenbach et al., 1974; Weber et al., 1974) and C. pepo (Walker and Thaine, 1971 ; Walker, 1972). Each of these two plant species contains a basic protein which gels in the absence of sulphydryl-protecting agents such as 2-mercapto- ethanol and dithioerythritol in the collecting medium. However, in C. maxima the gelling protein has a mo- lecular weight of 116,000 daltons and constitutes ca. 40% of total protein in the exudate, whereas in C.

212 D.D. Sabnis and J.W. Hart: Comparative Analysis of Phloem Exudate Proteins
pepo the gelling protein has an estimated molecular weight of 136,000 daltons and comprises 1~4% of total protein. Further, in C. maxima, a second basic protein constitutes another 40% of total exudate protein, while seven acidic proteins represent only 5% of the total. Conversely, in C. pepo, the numerous acidic proteins comprise the bulk of total protein in the exudate.
These results from the literature summarise the major qualitative and quantitative differences be- tween the phloem proteins of the plant species so far investigated. If such variance represents real and fundamental differences in the protein complements of the sieve tubes of different species, this has pro- found implications regarding any functional role of P-protein. Therefore, it was thought to be important to analyse the phloem proteins from different species under comparable conditions.
The proteins in the phloem exudate from three species of the family Cucurbitaceae have been frac- tionated by gel electrophoresis and gel isoelectric fo- cusing. The present report compares data on the esti- mated molecular weights and isoelectric points of the major and minor protein subunits from each species. The results indicate that striking differences do exist in the protein complements of phloem exudate from closely related plants.
Materials and Methods
Plants of Cucumis melo L. "Blenheim Orange" (melon), Cucumis sativus L. "Telegraph" (cucumber) and Cucurbita maxima Duches- ne "Golden Delicious" (giant marrow) were grown from seed under greenhouse conditions. Preliminary experiments were also conducted with Cucumis sativus L, "Bedfordshire Prize". Phloem exudate was collected from the transversely cut lower internodes of 2-morith-old plants of each species, and from the petioles of C. maxima. The cut surfaces were rinsed with the collecting medium (0.01 M Tris-HCl, pH 7.5, containing 1 mM DTT), blotted on filter paper to minimize contamination from damaged cells, and dipped into small volumes of the collecting solution (0.5 ml/plant). Urea, to a final concentration of 6 M, was added immediately to all samples. After further specified treatments, the samples were sub- jected to gel electrophoresis or isoelectric focusing. All reagents used were of "Analar" grade.
Procedures for isoelectric focusing in polyacrylamide gel cylin- ders were adapted from those described by Righetti and Drysdale (1974) and Davies (1975). Gels were prepared using the following stock solutions in the specified amounts:
All stock solutions (A-E) contained 4 M urea. Gels (5% acryl- amide; 0.2% Bis) were cast to a length of 8.5 cm in glass tubes 95 x 4 mm i.d. In some experiments ammonium persulphate was replaced by 0.004% riboflavin and the gels were photopolymerized.
Proteins were focused in a specially constructed apparatus with anode and cathode electrode solution chambers designed to contain a minimal volume of electrolyte (40 ml/chamber; cf. Rig- hetti and Drysdale, 1974). The lower electrode chamber was sealed so that the apparatus could be packed with ice. Reproducibility of protein patterns within and between experiments was high.
Ampholine (pH range 3.5 to 10) was added to all samples to a final concentration of 2%. Samples containing up to 100 gg protein were applied to the gels in volumes of 10 to 100 gl, and covered with a layer of buffer containing 2% Ampholine (pH 3.5 to 10) and 1% ascorbic acid to minimize possible anodic oxida- tion of proteins. The acid electrolyte (upper electrode chamber) was 4 drops concn. HzSOJ40 ml H20; the alkaline electrolyte was 0.2 g NaOH/40 ml H20. Focusing was carried out at an initial voltage of 100 v, increasing to a maximum of 300 v. Maximum power employed was 3 w. The duration of the run was approxi- mately 4 h.
Gels were fixed, stained with Coomassie blue and destained at 50 ~ (Vesterberg, 1972). Densitometric tracings of the gels were made with a Chromoscan 200 (Joyce-Loebl & Co.) fitted with filter No. 5-040. For determination of the gel pH gradient, duplicate gels from each experiment were sliced into 5.0 mm segments, and each segment was eluted with 1.0 ml of H20 for 4 h at 4 ~ The pH of the eluates was measured at 4 ~ with a calibrated microelec- trode.
Gel electrophoresis was carried out in 7.5% acrylamide gels containing 0.1% SDS (Weber and Osborn, 1969). Gels were cast to a length of 6.5 to 6.8 cm in glass tubes 75 x 6 mm i.d. Urea (4 M) and DTT (1 raM) were incorporated into the gels for most experiments. Electrophoresis was performed at a constant current of 6.5 mA/gel, using Bromophenol blue as the tracking dye. After electrophoresis, the gels were fixed in 10% TCA for 1 h at 50 ~ stained with Coomassie blue (Weber and Osborn, 1969) and de- stained according to Vesterberg (1972).
Discontinuous urea-SDS-gel electrophoresis (with 8 M urea in all gels, samples and buffers) was performed as described by Shelanski (1974). The discontinuous SDS system developed by Laemmli (1970) was also tested.
Samples of exudate in collection buffer were prepared for elec- trophoresis by mixing with equal volumes of 1% SDS in 0.01 M phosphate buffer, pH 7.1, containing 1% mercaptoethanol, and heating to 100 ~ for 1.5 rain. Samples to be fractionated on gels containing 4 M urea were then mixed with equal volumes of 8 M urea (final urea concentration 5.5 M).
The molecular weights of exudate proteins were estimated from a standard graph of the relative migration of known proteins plot- ted against the logs of their molecular weights (Shapiro et al., 1967). Standard proteins employed were bovine serum albumin, catalase, ?, -globulin H and L chains, trypsin, myoglobin and cyto- chrome C, treated in the same manner as the exudate samples.
Reduction and alkylation of exudate proteins from Cucumis sativus "Bedfordshire Prize" were carried out according to the procedures of Renaud et al. (1968).
A. Acrylamide (B.D.H.) 23.1% 6.25 ml B. N,N'-methylenebisacrylamide (B.D.H.) 0.9% 7.0 ml C. Ammonium persulphate 1.5% 1.5 ml D. Glycerol 10.0% 10.0 ml E. Dithiothreitol (Sigma) 10.0 mM 3.0 ml Ampholine (L.K.B.), pH range 3.5 to 10.0 1.5 ml Ampholine (L.K.B.), pH range 4.0 to 6.0 0.2 ml Ampholine (L.K.B.), pH range 5.0 to 8.0 0.25 ml Ampholine (L.K.B.), pH range 8.0 to 9.5 0.3 ml
Results
When exudate was collected in Tris buffer containing 1 mM DTT, the solutions frequently became turbid quite rapidly. We have not ascertained whether higher concentrations of DTT or mercaptoethanol reduce such aggregation and precipitation of exuded corn-
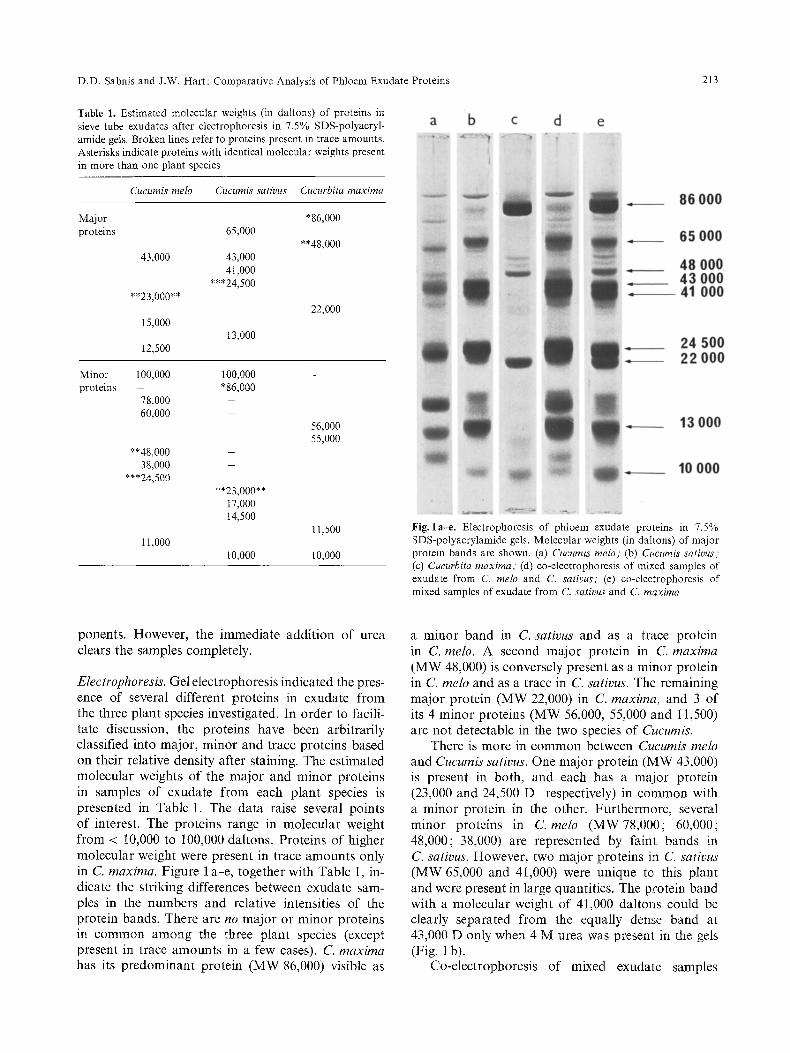
D.D. Sabnis and J.W. Hart: Comparative Analysis of Phloem Exudate Proteins 213
Table 1. Estimated molecular weights (in dattons) of proteins in sieve tube exudates after electrophoresis in 7.5% SDS-polyacryl- amide gels. Broken lines refer to proteins present in trace amounts. Asterisks indicate proteins with identical molecular weights present in more than one plant species
Cucumis melo Cucumis sativus Cucurbita maxima
M~or *86,000 proteins 65,000
**48,000 43,000
**23,000**
15,000
12,500
43,000 41,000
***24,500
13,000
Minor 100,000 100,000 protNns - *86,000
78,000 60,000
22,000
**48,000 38,000
***24,500
11,000
**23,000** 17,000 14,500
10,000
56,000 55,000
11,500
I0,000
Fig. la-e. Electrophoresis of phloem exudate proteins in 7.5% SDS-polyacrylamide gels. Molecular weights (in daltons) of major protein bands are shown. (a) Cucumis melo; (b) Cucumis sativus; (c) Cueurbita maxima; (d) co-electrophoresis of mixed samples of exudate from C. melo and C. sativus," (e) co-electrophoresis of mixed samples of exudate from C. sativus and C. maxima
ponents. However, the immediate addition of urea clears the samples completely.
Electrophoresis. Gel electrophoresis indicated the pres- ence of several different proteins in exudate from the three plant species investigated. In order to facili- tate discussion, the proteins have been arbitrarily classified into major, minor and trace proteins based on their relative density after staining. The estimated molecular weights of the major and minor proteins in samples of exudate f rom each plant species is presented in Table 1. The data raise several points of interest. The proteins range in molecular weight from < 10,000 to 100,000 daltons. Proteins of higher molecular weight were present in trace amounts only in C. maxima. Figure 1 a-e, together with Table 1, in- dicate the striking differences between exudate sam- ples in the numbers and relative intensities of the protein bands. There are no major or minor proteins in common among the three plant species (except present in trace amounts in a few cases). C. maxima has its predominant protein (MW 86,000) visible as
a minor band in C. sativus and as a trace protein in C. melo. A second major protein in C. maxima
(MW 48,000) is conversely present as a minor protein in C. melo and as a trace in C. sativus. The remaining major protein (MW 22,000) in C. maxima, and 3 of its 4 minor proteins (MW 56,000, 55,000 and 11,500) are not detectable in the two species of Cucumis.
There is more in common between Cucumis melo and Cucumis sativus. One major protein (MW 43,000) is present in both, and each has a major protein (23,000 and 24,500 D respectively) in common with a minor protein in the other. Furthermore, several minor proteins in C. melo (MW78,000; 60,000; 48,000; 38,000) are represented by faint bands in C. sativus. However, two major proteins in C. sativus (MW 65,000 and 41,000) were unique to this plant and were present in large quantities. The protein band with a molecular weight of 41,000 daltons could be clearly separated from the equally dense band at 43,000 D only when 4 M urea was present in the gets (Fig. 1 b).
Co-electrophoresis of mixed exudate samples
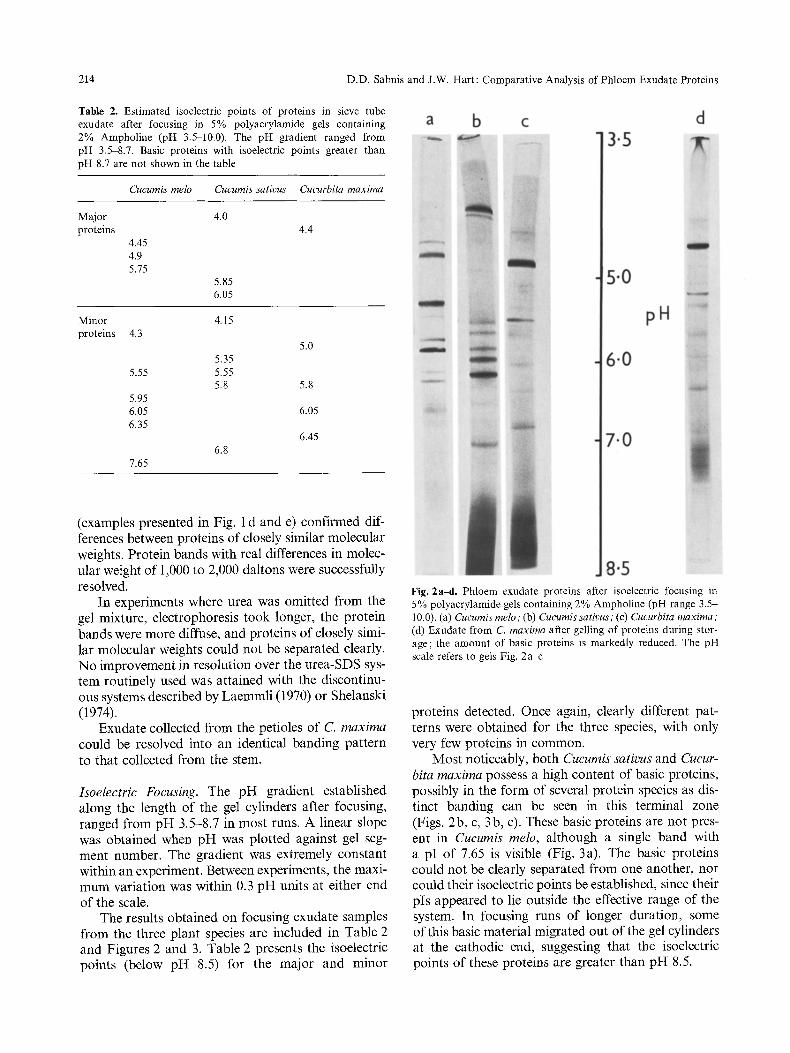
214 D.D. Sabnis and J.W. Hart: Comparative Analysis of Phloem Exudate Proteins
Table 2. Estimated isoelectric points of proteins in sieve tube exudate after focusing in 5% polyacrylamide gels containing 2% Ampholine (pH 3.5-10.0). The pH gradient ranged from pH 3.5-8.7. Basic proteins with isoelectric points greater than pH 8.7 are not shown in the table
Cucumis melo Cucumis sativus Cucurbita maxima
Major 4.0 proteins
4.45 4.9 5.75
5.85 6.05
4.4
Minor 4.15 proteins 4.3
5.0 5.35
5.55 5.55 5.8 5.8
5.95 6.05 6.05 6.35
6.45 6.8
7.65
(examples presented in Fig. 1 d and e) confirmed dif- ferences between proteins of closely similar molecular weights. Protein bands with real differences in molec- ular weight of 1,000 to 2,000 daltons were successfully resolved.
In experiments where urea was omitted from the gel mixture, electrophoresis took longer, the protein bands were more diffuse, and proteins of closely simi- lar molecular weights could not be separated clearly. No improvement in resolution over the urea-SDS sys- tem routinely used was attained with the discontinu- ous systems described by Laemmli (1970) or Shelanski (1974).
Exudate collected from the petioles of C. maxima could be resolved into an identical banding pattern to that collected from the stem.
Isoelectric Focusing. The pH gradient established along the length of the gel cylinders after focusing, ranged from pH 3.5-8.7 in most runs. A linear slope was obtained when pH was plotted against gel seg- ment number. The gradient was extremely constant within an experiment. Between experiments, the maxi- mum variation was within 0.3 pH units at either end of the scale.
The results obtained on focusing exudate samples from the three plant species are included in Table 2 and Figures 2 and 3. Table 2 presents the isoelectric points (below pH 8.5) for the major and minor
Fig. 2a-d. Phloem exudate proteins after isoelectric focusing in 5% polyacrylamide gels containing 2% Ampholine (pH range 3.5- 10.0). (a) Cucumis melo ; (b) Cucumis sativus ; (c) Cucurbita maxima; (d) Exudate from C. maxima after gelling of proteins during stor- age; the amount of basic proteins is markedly reduced. The pH scale refers to gels Fig. 2a~;
proteins detected. Once again, clearly different pat- terns were obtained for the three species, with only very few proteins in common.
Most noticeably, both Cucumis sativus and Cucur- bita maxima possess a high content of basic proteins, possibly in the form of several protein species as dis- tinct banding can be seen in this terminal zone (Figs. 2b, c, 3b, c). These basic proteins are not pres- ent in Cucumis melo, although a single band with a pI of 7.65 is visible (Fig. 3a). The basic proteins could not be clearly separated from one another, nor could their isoelectric points be established, since their pIs appeared to lie outside the effective range of the system. In focusing runs of longer duration, some of this basic material migrated out of the gel cylinders at the cathodic end, suggesting that the isoelectric points of these proteins are greater than pH 8.5.

D.D. Sabnis and J.W. Hart: Comparative Analysis of Phloem Exudate Proteins 215
Fig. 4a and b. Exudate proteins from Cucumis sativus "Bedfordshire Prize" after isoelectric focusing in 5% polyacrylamide gels containing 2% Ampholine (pH range 3.5-10.0). (a) Exudate proteins collected in Tris buffer containing 1% mercaptoethanol ; (b) exudate proteins after reduction and alkylation
,!
y I
l ~ , /
8:5 720 6[0 510 315 pH
Fig. 3a-e. Densitometric scans of gels after isoelectric focusing of exudate proteins from (a) Cucumis melo, (b) Cucumis sativus, and (c) Cucurbita maxima. Peaks between pH 7.5 and 8.5 in Fig. 2b and c represent unfocused basic proteins
At least 21 protein bands are visible in focused samples of C. melo exudate. Three of these proteins are present in major amounts, together with ca. six distinct minor proteins. C. sativus displays more than 23 bands, including three major acidic proteins and five minor proteins, in addition to the unresolved basic proteins. Approximately 23 bands are also pres- ent in exudate from C. maxima. However, in this case, only one major acidic protein (pI 4.5) is visible, in addition to the unresolved basic proteins. Identical results to those shown in Figure 2 were obtained when ammonium persulphate was replaced by riboflavin as the catalyst for acrylamide gel polymerization.
If samples of exudate, prepared for analysis by isoelectric focusing, were stored in 6 M urea and 1 mM DTT for a week at 4 ~ considerable gelling took place in the sample from C. sativus, rather less gelling in the sample from C. maxima, and no gelling
whatsoever in the sample from C. melo. This gelling is most probably due to the presence of the basic proteins in the two former species. Samples of exudate from C. maxima were focused, both before and after gelling, and the results showed that the pattern of acidic and neutral proteins remained similar in the two equivalent samples. However, there was a marked reduction in the amounts of basic proteins that entered the gels from the stored samples, with the concomitant accumulation of precipitated material on the upper surface of these gels (Fig. 2d).
In preliminary experiments with Cucumis sativus L. "Bedfordshire Prize ", the pattern of protein bands visible after focusing showed some major differences from that obtained with the cultivar "Telegraph" (cf. Figs. 2b and 4a). When exudate from the former cultivar was focused after reduction and alkylation of the proteins, as many as 33 distinct bands were detected (Fig. 4b). A more detailed comparison of proteins in the exudate of different cultivars is in progress.

216 D.D. Sabnis and J.W. Hart: Comparative Analysis of Phloem Exudate Proteins
Discussion
The most significant finding in the present work was the wide variation in the numbers, relative quantities and properties of the major proteins in exudate from the three cucurbit species examined. SDS-electro- phoresis indicated that no major or minor proteins were common to all three species, except for trace amounts in some cases. The greater part of the total protein in exudate from C. maxima consisted of a single band with a molecular weight of 86,000 daltons. This band could not be split into smaller components by the addition of 6 M urea and 1 mM DTT to both sample and polyacrylamide gel. The protein was pres- ent in minor amounts in C. sativus and as a trace in C. melo. Furthermore, at least two major proteins (MW 65,000 and 41,000) in exudate from C. sativus were not present in the other plant exudates.
While neither molecular weight nor isoelectric point alone is sufficient to establish the identity of protein bands in acrylamide gels of different exudate samples, we conclude from Table 1 that although only one major protein in C. melo was also present in C. sativus, several minor proteins in the former were represented by faint bands in the latter. Thus, the two species of Cueumis possess more phloem proteins in common than either compared with C. maxima. Even so, of the two Cucumis species only C. sativus possesses the predominant basic proteins probably responsible for the gelling behaviour of its exudate. We have no clear evidence that the basic proteins of Cucumis sativus and Cucurbita maxima are identical (although both species possess basic proteins in large amounts; cf. Fig. 2b and c), particularly since we could find no correspondence among the major proteins in exudate from both plants fractionated by gel electrophoresis. These discrepancies are not due to variation between exudate samples taken from the same plant species, as aliquots from the same exudate sample were used for both electrophoresis and isoelec- tric focusing. For this preliminary report, we have made no attempt to assign isoelectric points to any of the proteins separated by electrophoresis.
Fractionation of exudate proteins by isoelectric focusing reinforces the electrophoresis data in indicat- ing marked differences in the protein complement of exudate from the three related plant species. Here again, with the possible exception of the basic proteins in C. sativus and C. maxima, no major or minor exudate proteins were present in all three species. The preliminary results with a second cultivar of C. sativus (" Bedfordshire Prize") demonstrate that even between cultivars of the same species prominent dif- ferences may exist.
The apparent differences between cultivars of cu-
cumber in the present work, make it easier to accept the discrepancies between our results with Cucurbita maxima "Golden Delicious" and the results reported by Kleinig and co-workers (Kleinig et al., 1971; Weber and Kleinig, 1971; Beyenbach, Weber and Kleinig, 1974)using Cucurbita maxima "Gelber Zent- ner". We have attempted to ensure that we are deal- ing with protein subunits, by collecting exudate direct- ly into buffer containing sulphydryl protecting agents and by immediate treatment of samples with 6 M urea. The absence of proteins with molecular weights over 100,000 daltons would tend to support this con- tention. Nevertheless, we have obtained a completely different protein pattern for exudate from C. maxima from that reported by the above-mentioned workers. Kleinig and co-workers report the presence of five major proteins, two of which are basic and represent 40% each of total protein in the exudate. The molecu- lar weights of these two basic proteins were estimated to be 30,000 D and 116,000 D. Treatment of the 116,000 D protein with 8 M urea or 6 M guanidine hydrochloride did not split it into smaller components (Beyenbach, et al., 1974). Although basic proteins were also present in large amounts in our exudate samples from C. maxima, SDS-electrophoresis indi- cated only one predominant protein, with a molecular weight of 86,000 daltons, together with much smaller amounts of proteins with molecular weights of 48,000 and 22,000 daltons. The fact that the basic gelling protein of Cucurbita pepo has a molecular weight of 136,000 D and comprises only 4% of total protein in exudate from this plant (Walker and Thaine, 1971 ; Walker, 1972), only emphasizes the problem that has arisen.
Heyser et al. (1974) subjected sieve tube exudate from C. maxima to isoelectric focusing on a sucrose density gradient, pH range 3-10. The elution profile was continuously monitored at 280 nm. Their results demonstrate prominent peaks between pH 9-10 ;" how- ever, several large peaks were recorded between pH 3.5 and 6, possibly representing some of the acidic proteins that we have detected in this species.
Yapa and Spanner (1972) claim to have focused phloem exudate as well as extracts of whole phloem tissue from Heracleum mantegazzianum using sucrose density gradients and polyacrylamide gels. Extracts of isolated phloem strands in NaCI, NaOH and urea contained a filamentous component that focused at a pH of 4.9. However, we have serious reservations regarding the validity of these results. It is astonishing that aqueous or 8 M urea extracts of whole phloem tissue yielded only one or two protein bands after gel electrofocusing. Secondly, the filaments in their extracts (cf. their Fig. 2a) seem identical in appear- ance and dimensions to filaments that we have isolat-

D.D. Sabnis and J.W. Hart: Comparative Analysis of Phloem Exudate Proteins 217
ed and partially characterized from extracts of phloem, xylem and cortical tissues of Heracleum, and which are positively not a form of P-protein (Hart and Sabnis, 1975). These unusually stable, possibly glycoprotein filaments focused at a similar pH (4.9) on a sucrose density gradient column. Finally, it seems remarkable that any form of P-protein should remain in a filamentous form in the presence of 8 M urea. Even if it did, it could not then enter 5% poly- acrylamide gels.
Several workers have proposed that P-proteins may have an active role in the functioning of sieve tubes (Thaine, 1969; Hejnowicz, 1970; Lee, 1971; MacRobbie, 1971). Others have suggested that P- proteins merely represent products of ribosomal de- gradation (Weber et al., 1974; Behnke, 1974). In ei- ther case, the widely differing protein patterns reported for exudate from different species is difficult to understand. The relatively large amounts of P- proteins in the sieve elements of many species, and their highly organized structure, render the possibility of their being degradation product unlikely. The tubu- lar P-protein elements develop early during the differ- entiation of sieve elements, before any signs of nuclear or cytoplasmic degeneration are visible, and, particu- larly in the legumes, the proteins are organized into elaborate crystals with definite shapes and orientation within the cell (see recent reviews by Cronshaw, 1975 ; Kollmann, 1975). P-protein elements and bodies have also been seen in the parenchyma and companion cells of phloem (Cronshaw and Esau, 1968; Zee, 1969; Steer and Newcomb, 1969; Esau, 1973; Wergin et al., 1975).
Whether or not exudate from the sieve elements of different plant species truly represents their native protein complement, nevertheless, one would still ex- pect some similarities among those proteins, soluble or structural, that do appear in the phloem sap. Bio- chemical analyses of the proteins in exudate have revealed only one significant feature so far. With the exception of Cucumis melo, the remaining three cucur- bit species contain basic proteins (in widely differing amounts) that are likely to be responsible for the gelling reaction of exudate and for the formation of filaments in vitro. Whether the filaments are identical in different species has yet to be determined. The assumption, on the basis of electron microscopic evi- dence, that the various tubular, filamentous, crystal- line and granular forms of P-protein reported in the literature represent different conformations of the same protein subunits is now also open to question. While the possibility exists that the species-based dif- ferences in exudate proteins that we report here may be due to different physiological and developmental states of the three plant species, the protein subunits
themselves are clearly different. The identical pattern obtained in exudate collected from the oldest inter- nodes and from relatively young petioles of C. maxi- ma, would tend to support the view that the phys- iological state of the plant is unlikely to be responsible for the different protein patterns obtained.
The limited information available makes further speculation regarding the nature or function of "P- protein" unwarranted. Obviously, further work of a detailed and comparative nature is urgently needed before any general picture is likely to emerge.
We are grateful to Mr I. Skene for technical assistance and to Mr E. Middleton for the photography.
References
Behnke, H.-D.: fiber den Feinbau und die Ausbreitung der Siebr6hren-Plasmafilamente und fiber Bau und Differenzierung der Siebporen bei einigen Monocotylen und bei Nuphar. Proto- plasma (Wien) 68, 377~402 (1969)
Behnke, H.-D. : Comparative ultrastructural investigations of an- giosperm sieve elements: aspects of the origin and early develop- ment of P-protein. Z. Pflanzenphysiol. 74, 2-34 (1974)
Beyenbach, J., Weber, C., Kleinig, H.: Sieve tube proteins from Cucurbita maxima. Planta (Berl.) 119, t13-124 (1974)
Cronshaw, J.: P-proteins. In: Phloem transport, pp. 79-115. Eds. : Aronoff, S., Dainty, J., Gorham, P.R., Srivastava, L.M., Swan- son, C.A., New York-London: Plenum Press 1975
Cronshaw, J., Esau, K.: P-protein in the phloem of Cucurbita. I. The development of P-protein bodies. J. Cell Biol. 38, 25-39 (1968)
Cronshaw, J., Gilder, J., Stone, D.: Fine structural studies of P-proteins in Cucurbita, Cucumis and Nicotiana. J. Ultrastruct. Res. 45, 192-205 (1973)
Davies, H. : Thin layer gel isoelectric focusing. In: Isoelectric focus- ing. Eds. : Arbuthnott, J.P., Beeley, J.A. London: Butterworths 1975
Eschrich, W., Evert, R.F., Heyser, W. : Proteins of the sieve tube exudate of Cucurbita maxima. Planta (Berl.) 100, 208-221 (1971)
Esau, K.: Comparative structure of companion cells and phloem parenchyma cells in Mimosa pudica L. Ann. Bot. 37, 625-632 (1973)
Esau, K., Cronshaw, J.: Tubular components in cells of healthy and tobacco mosaic virus-infected Nicotiana. Virology 33, 26 35 (1967)
Evert, R.F., Deshpande, B.P.: Electron microscope investigation of sieve-element ontogeny and structure in Ulmus americana.: Protoplasma (Wien) 68, 403432 (1969)
Hall, S.M., Baker, D.A.: The chemical composition of Ricinus phloem exudate. Planta (Berl.) 106, 131-140 (1972)
Hart, J.W., Sabnis, D.D.: The occurrence and some properties of a new filamentous component of plant extracts. Ann. Bot. 39, 153-159 (1975)
Hejnowicz, Z.: Propagated disturbances of transverse potential gradient in intracellular fibrils as the source of motive forces for longitudinal transport in cell. Protoplasma (Wien) 71, 343 364 (1970)
Heyser, W., Eschrich, W., Htittefmann, A., Evert, R.F., Burchardt, R., Fritz, E., Heyser, R. : Phosphodiesterase in sieve-tube exu- date of Cucurbita maxima. Z. Pflanzenphysiol. 71, 413-423 (1974)

218 D.D. Sabnis and J.W. Hart: Comparative Analysis of Phloem Exudate Proteins
Kleinig, H., D6rr, I., Weber, C., Kollmann, R.: Filamentous proteins from plant sieve tubes. Nature 229, 152 153 (1971)
Kollmann, R.: Sieve element structure in relation to function. In: Phloem transport, pp. 225-242. Eds. : Aronoff, S., Dainty, J., Gorham, P.R., Srivastava, L.M., Swanson, C.A., New York- London: Plenum Press 1975
Kollmann, R., D6rr, I., Kleinig, H. : Protein filaments-structural components &the phloem exudate. I. Observations with Cucur- bita and Nicotiana. Planta (Berl.) 95, 8644 (1970)
Laemmli, U.K. : Cleavage of structural proteins during the assemb- ly of the head of bacteriophage T4. Nature 680-685 (1970)
Lee, D.R. : Possible significance of filaments in sieve elements. Nature 235, 286 (1971)
MacRobbie, E.A.C.: Phloem translocation. Facts and mecha- nisms: a comparative survey. Biol. Rev. 46, 429-481 (1971)
Palevitz, B.A., Newcomb, E.H. : The ultrastructure and develop- ment of tubular and crystalline P-protein in the sieve elements of certain papilionaceous legumes. Protoplasma (Wien) 72, 399 426 (1971)
Parthasarathy, M.V., Mtihlethaler, K.: Ultrastructure of protein tubules in differentiating sieve elements. Cytobiol. 1, 17-36 (1969)
Renaud, F.L., Rowe, A.J., Gibbons, I.R. : Some properties of the protein forming the outer fibers of cilia. J. Cell Biol. 36, 79 90 (1968)
Righetti, P.G., Drysdale, J.W. : Isoelectric focusing in gels. J. Chro- matography 98, 271-321 (1974)
Sabnis, D.D., Hart, J.W. : P-protein in sieve elements. I. Ultrastruc- ture after treatment with vinbIastine and colchicine. Planta (Berl.) 109, 127-133 (1973)
Shapiro, A.L., Vifiuela, E., Maizel, J.V. : Molecular weight estima- tion of polypeptide chains by electrophoresis in SDS-polyacryl- amide gels. Biochim. Biophys. Res. Commun. 28, 815-820 (1967)
Shelanski, M.L. : Methods for the neurochemical study of micro- tubules. In: Research methods in neurochemistry. Eds. : Marks, N., Rodnight, R., vol. 2. New York-London: Plenum Press 1974
Steer, M.W., Newcomb, E.H.: Development and dispersal of P- protein in the phloem of Coleus blumei Benth. J. Cell Sci. 4, 155-169 (1969)
Thaine, R.: Movement of sugars through plants by cytoplasmic pumping. Nature 222, 873-875 (1969)
Vesterberg, O. : Isoelectric focusing of proteins in polyacrylamide gels. Biochim. biophys. Acta (Amst.) 257, 11-19 (1972)
Walker, T.S.: The purification and some properties of a protein causing gelling in phloem sieve tube exudate from Cucurbita pepo. Biochim. biophys. Acta (Amst.) 257, 433-444 (1972)
Walker, T.S., Thaine, R. : Proteins and fine structural components in exudate from sieve tubes in Cucurbita pepo stems. Ann. Bot. 35, 773-790 (1971)
Weber, C., Franke, W.W., Kartenbeck, J. : Structure and biochem- istry of phloem proteins isolated from Cucurbita maxima. Exp. Cell. Res. 87, 79-106 (1974)
Weber, C., Kleinig, H. : Molecular weights of Cucurbita sieve tube proteins. Planta (Bed.) 99, 179-182 (1971)
Weber, K., Osborn, M.: The reliability of molecular weight de- terminations by dodecyl sulfate-polyacrylamide gel electro- phoresis. J. biol. Chem. 244, 4406-4412 (1969)
Wergin, W.P., Newcomb, E.H. : Formation and dispersal of crys- talline P-protein in sieve elements of soybean (Glycine max. L.). Protoplasma (Wien) 71, 365-388 (1970)
Wergin, W.P., Palevitz, B.A., Newcomb, E.H. : Structure and de- velopment of P-protein in phloem parenchyma and companion cells of legumes. Tissue Cell 7, 227-242 (1975)
Williamson, R.E. : An investigation of the contractile protein hypo- thesis of phloem translocation. Planta (Berl.) 106, 127-133 (1973)
Yapa, P.A.J., Spanner, D.C.: Isoelectric focusing of sieve-tube protein. Planta (Bed.) 106, 369 373 (1972)
Zee, S.-Y. : P-protein producing ceils of the primary root of Vicia faba. Austr. J. Biol. Sci. 22, 1573-1576 (1969)
Received 17 January; accepted 3 February 1976




















