
Salicylic acid alleviates growth inhibition and oxidative stress caused by zucchini yellow mosaic...
10
Physiological and Molecular Plant Pathology 69 (2006) 172–181 Salicylic acid alleviates growth inhibition and oxidative stress caused by zucchini yellow mosaic virus infection in Cucurbita pepo leaves Deya Eldeen Mohammed Radwan a,b , Khalaf Ali Fayez b , Sabry Younis Mahmoud c , Ahmed Hamad d , Guoquan Lu a, a College of Agriculture and Biotechnology, Zhejiang University, Hangzhou 310029, China b Botany Department, Faculty of Science, Sohag University, Sohag 82524, Egypt c Virology Department, Faculty of Agriculture, Sohag University, Sohag 82786, Egypt d Botany Department, Faculty of Science, Assiut University, Assiut, Egypt Accepted 18 April 2007 Abstract The effects of zucchini yellow mosaic virus (ZYMV) infection and pretreatments with salicylic acid (SA) on biomass accumulation of pumpkin (Cucurbita pepo cv. Eskandarani) were investigated. The response of photosynthesis, transpiration and the activities of antioxidant enzymes in leaves was also considered. Significant reductions in growth parameters (i.e. leaf area, biomass and shoot height), photosynthesis and chlorophyll a and b content were detected in ZYMV-infected leaves in comparison to healthy controls. Antioxidant enzyme activities were increased up to 3-fold for peroxidase (POD), 2-fold for ascorbate peroxidase (APX) and catalase (CAT) activities and 1.3-fold for SOD activity by virus infection. ZYMV infection also caused increases in H 2 O 2 and malondialdehyde (MDA) contents. These results suggest that ZYMV infection causes oxidative stress in pumpkin leaves leading to the development of epidemiological symptoms. Interestingly, spraying pumpkin leaves with SA led to recovery from the undesirable effects of ZYMV infection. Leaves treated with 100 mM SA three days before inoculation had the appearance of healthy leaves. No distinct disease symptoms were observed on the leaves treated with 100 mM SA followed by inoculation with ZYMV. In non-infected plants, SA application increased activities of POD and superoxide dismutase (SOD) and inhibited APX and CAT activities. In contrast, SA treatment followed by ZYMV inoculation stimulated SOD activity and inhibited activities of POD, APX and CAT. In addition, MDA displayed an inverse relation, indicating inhibition of lipid peroxidation in cells under SA treatment. It is suggested that the role of SA in inducing plant defense mechanisms against ZYMV infection might have occurred through the SA-antioxidant system. Such interference might occur through inhibition or activation of some antioxidant enzymes, reduction of lipid peroxidation and induction of H 2 O 2 accumulation following SA application. r 2007 Elsevier Ltd. All rights reserved. Keywords: Leaf growth; Photosynthesis rate; Pigments; Pumpkin; Salicylic acid; Zucchini yellow mosaic virus; Antioxidant enzyme activity; Gas exchange 1. Introduction Zucchini yellow mosaic virus (ZYMV) is one of the most important viruses affecting cucurbit production globally. It causes destructive diseases on a large variety of economic- ally important cucurbits including zucchini squash (Cu- curbita pepo) [1]. It is efficiently transmitted by several aphid species in a non-persistent mode. The disease can completely destroy the crop under favorable conditions leading to yield loss. Moreover, ZYMV is widely spread in many countries including Egypt and is considered a serious problem in the major cucurbit producing regions worldwide. Resistance to ZYMV has not yet been established in C. pepo germplasm collections. Therefore, ZYMV resistance has been introduced into zucchini squash ARTICLE IN PRESS www.elsevier.com/locate/pmpp 0885-5765/$ - see front matter r 2007 Elsevier Ltd. All rights reserved. doi:10.1016/j.pmpp.2007.04.004 Abbreviations: Chl, chlorophyll; POX, peroxidase; SA, salicylic acid; SA+V, treated with salicylic acid then inoculated with virus; ZYMV, Zucchini yellow mosaic virus; E, transpiration rate; Ci, intercellular CO 2 concentration; Gs, stomatal conductance; WUE, water use efficiency Corresponding author. Tel./fax: +86 571 86971117. E-mail addresses: [email protected] (D.E.M. Radwan), [email protected] (G. Lu).
Transcript of Salicylic acid alleviates growth inhibition and oxidative stress caused by zucchini yellow mosaic...

ARTICLE IN PRESS
0885-5765/$ - se
doi:10.1016/j.pm
Abbreviations
SA+V, treated
Zucchini yellow
concentration;�CorrespondE-mail addr
Physiological and Molecular Plant Pathology 69 (2006) 172–181
www.elsevier.com/locate/pmpp
Salicylic acid alleviates growth inhibition and oxidative stress caused byzucchini yellow mosaic virus infection in Cucurbita pepo leaves
Deya Eldeen Mohammed Radwana,b, Khalaf Ali Fayezb, Sabry Younis Mahmoudc,Ahmed Hamadd, Guoquan Lua,�
aCollege of Agriculture and Biotechnology, Zhejiang University, Hangzhou 310029, ChinabBotany Department, Faculty of Science, Sohag University, Sohag 82524, Egypt
cVirology Department, Faculty of Agriculture, Sohag University, Sohag 82786, EgyptdBotany Department, Faculty of Science, Assiut University, Assiut, Egypt
Accepted 18 April 2007
Abstract
The effects of zucchini yellow mosaic virus (ZYMV) infection and pretreatments with salicylic acid (SA) on biomass accumulation of
pumpkin (Cucurbita pepo cv. Eskandarani) were investigated. The response of photosynthesis, transpiration and the activities of
antioxidant enzymes in leaves was also considered. Significant reductions in growth parameters (i.e. leaf area, biomass and shoot height),
photosynthesis and chlorophyll a and b content were detected in ZYMV-infected leaves in comparison to healthy controls. Antioxidant
enzyme activities were increased up to 3-fold for peroxidase (POD), 2-fold for ascorbate peroxidase (APX) and catalase (CAT) activities
and 1.3-fold for SOD activity by virus infection. ZYMV infection also caused increases in H2O2 and malondialdehyde (MDA) contents.
These results suggest that ZYMV infection causes oxidative stress in pumpkin leaves leading to the development of epidemiological
symptoms. Interestingly, spraying pumpkin leaves with SA led to recovery from the undesirable effects of ZYMV infection. Leaves
treated with 100mM SA three days before inoculation had the appearance of healthy leaves. No distinct disease symptoms were observed
on the leaves treated with 100 mM SA followed by inoculation with ZYMV. In non-infected plants, SA application increased activities of
POD and superoxide dismutase (SOD) and inhibited APX and CAT activities.
In contrast, SA treatment followed by ZYMV inoculation stimulated SOD activity and inhibited activities of POD, APX and CAT. In
addition, MDA displayed an inverse relation, indicating inhibition of lipid peroxidation in cells under SA treatment. It is suggested that
the role of SA in inducing plant defense mechanisms against ZYMV infection might have occurred through the SA-antioxidant system.
Such interference might occur through inhibition or activation of some antioxidant enzymes, reduction of lipid peroxidation and
induction of H2O2 accumulation following SA application.
r 2007 Elsevier Ltd. All rights reserved.
Keywords: Leaf growth; Photosynthesis rate; Pigments; Pumpkin; Salicylic acid; Zucchini yellow mosaic virus; Antioxidant enzyme activity; Gas exchange
1. Introduction
Zucchini yellow mosaic virus (ZYMV) is one of the mostimportant viruses affecting cucurbit production globally. It
e front matter r 2007 Elsevier Ltd. All rights reserved.
pp.2007.04.004
: Chl, chlorophyll; POX, peroxidase; SA, salicylic acid;
with salicylic acid then inoculated with virus; ZYMV,
mosaic virus; E, transpiration rate; Ci, intercellular CO2
Gs, stomatal conductance; WUE, water use efficiency
ing author. Tel./fax: +86 571 86971117.
esses: [email protected] (D.E.M. Radwan),
.cn (G. Lu).
causes destructive diseases on a large variety of economic-ally important cucurbits including zucchini squash (Cu-
curbita pepo) [1]. It is efficiently transmitted by severalaphid species in a non-persistent mode. The disease cancompletely destroy the crop under favorable conditionsleading to yield loss. Moreover, ZYMV is widely spread inmany countries including Egypt and is considered aserious problem in the major cucurbit producing regionsworldwide. Resistance to ZYMV has not yet beenestablished in C. pepo germplasm collections. Therefore,ZYMV resistance has been introduced into zucchini squash

ARTICLE IN PRESSD.E.M. Radwan et al. / Physiological and Molecular Plant Pathology 69 (2006) 172–181 173
commercial cultivars through crosses with Cucurbita
moschata [2].Several chemicals have been reported as resistance
inducers in plants [3]. Among these chemicals, salicylicacid (SA) is considered one of the key components ofdefense signal transduction, inducing the full set ofsystemic acquired resistance genes [4]. Moreover, exogen-ous application of SA may influence some physiologicalprocesses such as seed germination and fruit yield [5],transpiration rate [6], stomatal closure [7], membranepermeability [8], growth and photosynthesis [9–13] andantioxidant capacity [14]. Thus, SA may affect plantdefense mechanisms against harmful diseases. SA gener-ally, can control both biotic and abiotic defense programs[15]. Nowadays, the importance of SA in pathogen-induceddisease resistance is well documented [4,16]. To ourknowledge, the use of SA against viral infections inzucchini squash (C. pepo) is still limited.
Certain biochemical and physiological changes asso-ciated with virus infection include the oxidative burst [17],growth inhibition, changes in pigment content, chlorophyllsynthesis and antioxidant enzyme systems [18]. Alteredactive oxygen species (AOS) formation occurs under bioticand abiotic stress [19]. This alteration causes direct orindirect damage to plants leading to the appearance ofepidemiological symptoms.
Plants have a set of enzymes and redox metabolites thatcarry out AOS detoxification and known as antioxidantsystems. The main antioxidant enzymes are superoxidedismutase (SOD) catalyzing the dismutation of O2
� toH2O2, catalase (CAT) that dismutates H2O2 to oxygen andwater, and ascorbate peroxidase (APX) that reduces H2O2
to water by utilizing ascorbate as specific electron donor.Moreover, other enzymes involved in the ascorbate–glutathione ASC–GSH cycle [glutathione reductase (GR),monodehydroascorbate reductase (MDHAR) and dehy-droascorbate reductase (DHAR)] as well as glutathioneperoxidase (GPX) and glutathione-S-transferases, areimportant in protecting cells against oxidative stress.
These enzymes are present in different isoforms inseveral cell compartments and their expression isgenetically controlled and regulated both by develop-mental and environmental stimuli, according to thenecessity to remove AOS produced in cells [20–22]. Thebalance between SOD and APX or CAT activities in cells iscrucial for determining the steady-state level of O2
� andH2O2.
Reported data indicate that under abiotic and bioticstress conditions, which induce an over-production of AOSin cells, plant resistance is due to the capability to increasethe activity of AOS-detoxifying enzymes or the bio-synthesis or regeneration of antioxidant metabolites[19,21,23,24].
It is well established that plant viruses affect manyphysiological processes such as photosynthesis [20] bydecreasing the photosynthesis rate (depending on theinfection stage), decreasing pigment contents [18,25],
increasing or decreasing the respiration rates and alteredgas exchange relations [26,27].Although there are many reports concerning the change
in physiological properties after virus infection, there islimited information on physiological responses in plantsinfected by ZYMV. To our knowledge, no information isavailable on the effects of ZYMV infection and exogenousSA on transpiration, gaseous exchange, photosynthesisrate and antioxidant enzyme activities. Therefore, the mainobjectives of the present study are to investigate the effectsof ZYMV infection and SA treatments on plant growth,oxidative stress, the response of certain antioxidantenzymes, photosynthetic properties and biomass accumu-lation in pumpkin (C. pepo cv. Eskandarani) leaves.
2. Materials and methods
2.1. Plant materials
Seeds of pumpkin (C. pepo cv. Eskandarani) were sown ina mixture of sand and clay (1:2, v/v) in plastic pots (10 cm indiameter) in separate growth chambers under a photoperiodof 12h. Day and night temperatures were 24 and 18 1C,respectively; the relative humidity was about 70%. Theplants were kept at 100% water holding capacity.
2.2. SA treatments and virus inoculation
After 21 days of growth, plants were divided into sevengroups. Each group comprised four replications. The firstgroup was considered as a control without any treatment.Second, third and fourth groups were treated with 10, 50and 100 mM of SA respectively by spraying the leaves untilrun-off. The control was sprayed with water. To improvespread on leaves, two drops of Tween 80 were added priorto spraying. Inoculation of virus was performed 3 daysafter spraying. All leaves were mechanically inoculated.The fifth group was inoculated with ZYMV at the sametime of inoculation of the other groups. The sixth group(SA+V ino) was inoculated with a mixture of virusinoculum and 100 mM SA solution (1:1). The last groupwas sprayed with 100 mM SA without inoculation. Theinoculum was prepared from infected top leaves, groundin a mortar containing 0.1M phosphate buffer pH 7.0(1:2, w/v); the homogenate was filtrated through two layersof muslin, and the leaves of healthy plants were dusted withcarborundum, and rubbed gently with a cotton swabpreviously dipped into the suspension of virus inoculum.Two weeks after inoculation, the youngest fully developedleaves from both control and treated plants were collectedfor analysis of biochemical changes.
2.3. Chlorophyll content and photosynthetic gas exchange
analyses
Fifteen days after inoculation with virus, the leaves withsimilar age were harvested and ground in 85% acetone to

ARTICLE IN PRESSD.E.M. Radwan et al. / Physiological and Molecular Plant Pathology 69 (2006) 172–181174
extract the pigments. Contents of chlorophyll a (Chl a) andchlorophyll b (Chl b) were determined spectrophotome-trically according to Metzner et al. [28]. Chlorophyllcontents of infected leaves, SA treated and inoculatedafter treatment with SA were then compared to the control.Photosynthetic rate (Pn), stomatal conductance (Gs),intercellular CO2 concentration (Ci) and transpiration rate(E) were determined by using an Infra Red Analyzer (LI-6400 System, Li-COR Company Lincoln, NE). The valuespresented represent the means of eight replicates.
2.4. Antioxidant enzyme activity analyses
For analyses of enzyme activity, leaves were homoge-nized with a mortar and pestle under chilled conditions in abuffer solution specific for each enzyme. The homogenatewas filtered through four layers of muslin cloth andcentrifuged at 12 000 rpm for 10min at 4 1C. The super-natants were used for enzyme assays.
CAT (EC 1.11.1.6) and peroxidase (POD, EC 1.11.1.7)activities were measured according to Chandlee andScandalios [29] and Zhang [30], respectively, with somemodifications. For POD, the reaction mixture consisted of50mM potassium phosphate buffer (pH 6.1), 1% guaiacol,0.4% H2O2 and enzyme extract. Increase in the absorbancedue to oxidation of guaiacol ðE ¼ 25:5mM�1 cm�1Þ wasmeasured at 470 nm. Enzyme activity was calculated interms of mmol of guaiacol oxidized min�1 g�1 fresh weightat 2572 1C. CAT activity was measured by monitoring thedisappearance of H2O2 by measuring the decrease inabsorbance at 240 nm (E ¼ 0.036mM�1 cm�1) of areaction mixture consisting of 25mM potassium phosphatebuffer (pH 7.0), 10mMH2O2 and enzyme extract.
APX (EC 1.11.1.11) was determined according toNakano and Asada [31] and with modified procedure ofHemandez et al. [32]. The decrease in ascorbate concentra-tion was followed by decline in absorbance at 290 nm andactivity was calculated using the extinction coefficient(E ¼ 2.8mM�1 cm�1 at 290 nm) for ascorbate.
SOD (EC 1.15.1.1) activity was assayed according toBeauchamp and Fridovich [33] with some modifications.The samples (0.5 g) were homogenized in 5ml extractionbuffer consisting of 50mM phosphate, pH 7.8, 0.1% (w/v),BSA, 0.1% (w/v) ascorbate, 0.05% (w/v) b-mercaptoethanol. The assay mixture in 3ml contained 50mMphosphate buffer (pH 7.8), 9.9mM L-methionine, 57 lMNBT, 0.025% (w/v) Triton X-100 and 0.0044% (w/v)riboflavin. The photo-reduction of NBT (formation ofpurple formazan) was measured at 560 nm. One unit ofSOD activity was defined as extract volume that caused50% inhibition of the photo-reduction of NBT.
2.5. H2O2, MDA contents and antioxidant activity
H2O2 content was determined colorimetrically as de-scribed by Jana and Choudhuri [34]. H2O2 was extractedby homogenizing 0.5 g leaf tissue with 3ml phosphate
buffer (50mM, pH 6.5). The homogenate was centrifugedat 6000 rpm for 25min. To determine H2O2 level, 3mlof extracted solution was mixed with 1ml of 0.1%titanium sulfate in 20% H2SO4. The mixture was thencentrifuged at 6000 g for 15min. The intensity of the yellowcolor of the supernatant at 410 nm was measured. H2O2
level was calculated using the extinction coefficient(E ¼ 0.28 mmol cm�1).Lipid peroxidation was expressed as malondialdehyde
(MDA) content and was determined as 2thiobar-bituric acid (TBA) reactive metabolites according to Zhang[30]. Lipid peroxidation was expressed as nmol (g freshweight)�1 by using an extinction coefficient (E ¼155mMcm�1). Antioxidant activity (AOA) was calculatedaccording to Arabshahi et al. [35] and expressed aspercentage inhibition of lipid peroxidation relative to thecontrol using absorbance at 532 nm obtained from MDAanalysis according to the following equation:
AOA activity ð%Þ
¼absorbance of control� absorbance of sample
absorbance of control� 100.
2.6. Statistical analysis
All data were subjected to ANOVA test and means werecompared by two conventional methods of analysis. TheLSD values for significant mean differences at levelsPo0.05 and Po0.01 were separated. The data were alsostatistically analyzed by Duncan’s range test to estimate thedifferences among the variants [36,37]. All statistics testswere carried out using SPSS program.
3. Results
3.1. Morphological and growth parameters
Tables 1 and 2 show the effects of ZYMV infection andSA treatments with or without virus inoculation on the leafarea, plant height, biomass and water contents of pumpkinleaves (C. pepo cv. Eskandarani). Growth inhibition wasdetected in response to ZYMV infection which wasindicated by reduction of leaf area, biomass and plantheight by 81%, 28.9% and 37%, respectively of corre-sponding control values. In contrast, leaf water contentseemed to be unaffected by ZYMV infection.SA application aided recovery from the pathological
effects caused by virus infection especially with 100 mM(SA+V) level (Fig. 1). Plants treated with 100 mM SA hadleaf area and plant height more or less similar to controls.Moreover, biomass accumulation seemed to be induced bySA application regardless of viral infection. From theresults in Table 2, we can observe the maximum increase inbiomass was detected in the plants treated with 100 SAwithout virus inoculation. At the same time, plantstreated with 100 SA+V showed an increase in biomass

ARTICLE IN PRESS
Table 1
Effect of ZYMV infection and SA treatments on leaf area (cm2 plant�1)
and plant height (cm) of C. pepo (cv. Eskandarani) leaves
Treatments Leaf area (cm2 plant�1) Plant height (cm)
M7SD % M7SD %
Control 161.03a732.67 100 19.11a,b,c72.04 100
Infected 30.36**c72.53 18.86 12.01**d70.57 62.84
10 mM SA+V 63.10**c79.37 39.19 16.02c70.69 86.42
50 mM SA+V 104.04*b716.19 64.61 17.18b,c71.25 89.92
100 mM SA +V 106.36*b725.50 66.05 20.64a,b72.27 108.05
100 mM SA +V ino 127.15a,b721.95 78.96 14.01c70.62 87.50
100 mM SA 155.10a73.56 96.32 21.19a73.23 110.93
Statistical significance of differences compared to control: *Po0.05;
**Po0.01. Duncan’s composition between variants at a ¼ 0.05. Variants
with the same letters show the same reaction. The values are means (M) of
four replicates7standard deviation (SD).
Table 2
Effect of ZYMV infection and SA treatments on biomass (g plant�1) and
water content (%) of C. pepo (cv. Eskandarani) leaves
Treatments Leaf dry weight (g plant�1) Water content
(%)
M7SD % M7SD
Control 0.38b,c70.02 100 93.98a70.93
Infected 0.27**d70.004 69.83 93.86a70.83
10 mM SA+V 0.33c,d70.03 87.04 91.42b70.63
50 mM SA+V 0.38b,c70.05 97.83 92.18a,b70.07
100mM SA +V 0.41b,c70.05 106.26 90.85*b71.10
100mM SA +V ino 0.43a,b70.04 111.22 90.81*b70.85
100mM SA 0.48**a70.06 125.57 91.76b70.97
Statistical significance of differences compared to control: *, significant at
Po0.05; **, significant at Po0.01. Duncan’s composition between
variants at a ¼ 0.05. Variants with the same letters show the same
reaction. The values are means (M) of four replicates7standard deviation
(SD).
D.E.M. Radwan et al. / Physiological and Molecular Plant Pathology 69 (2006) 172–181 175
accumulation. These results indicate that ZYMV couldcause serious disease leading to growth reduction andmorphological leaf modification which could be alleviatedby exogenous application of SA.
3.2. Chlorophyll content and photosynthesis rate
Pumpkin leaves infected with ZYMV showed a sig-nificant reduction in photosynthetic rates (Pn) andchlorophyll content (see Table 3). The reduction inchlorophyll content clearly appeared as the developmentof leaf mosaics and disease symptoms which was confirmedby the chemical analysis of the chlorophyll (a+b)concentration and Pn. Chlorophyll (a+b) concentrationand Pn were reduced by 45% and 52.5%, respectively, incomparison to corresponding controls.
An alleviation of ZYMV severe infection effects wasevidenced by alteration in Pn and chlorophyll contents at50 and 100 mM (SA+V) levels. Compared with control, thevalues of Pn showed no significant difference for 50 and
100 mM (SA+V) treated leaves. Similarly, chlorophyll(a+b) concentration decreased slightly but not signifi-cantly at 50 mM (SA+V) level.In all SA treatments, chlorophyll a/b ratio was higher
than healthy controls. The increase in a/b ratio is due to thedecrease in Chl b content. So, Chl b is more sensitive toZYMV infection and SA treatments than Chl a. Moreover,leaf Pn and chlorophyll (a+b) concentration seemed to beunaffected by 100 mM SA without further virus inoculation(Table 3).
3.3. Antioxidant enzyme activities
The analyses of four antioxidant enzymes activities,SOD, CAT, POD and APX, revealed many changes due toZYMV infection and SA treatments. ZYMV infectioncaused significant increases in enzyme activities up to3-fold in the case of POD, 2-fold for APX and CATactivities and 1.3-fold greater than controls for SODactivity (Tables 4 and 5).On the other hand, the effect of SA levels on antioxidant
enzyme activities differed markedly with or without virusinoculation. For example, with increasing SA dose(10–100 mM SA+V), the activities of POD, APX andCAT decreased markedly while SOD activity increased.Interestingly, although there was a decrease in POD andAPX activities, the values of POD were still higher thancontrols, although APX was lower than the control. Incontrast, SOD activities increased gradually with increas-ing SA dose and the values were significantly higher thanrespective controls. It seems that spraying SA and virusinoculation counteracted the enhancement of SOD andinhibition of CAT activities.Without virus inoculation, spraying 100 mM SA to the
leaves increased POD and SOD activities by 141.37% and111.78%, respectively. In contrast, the APX and CAT wereinhibited in response to SA spraying and their activitieswere reduced by 36.19% and 46.25% of the control,respectively.
3.4. MDA and H2O2 contents
Due to ZYMV infection, MDA and H2O2 concentra-tions increased significantly in pumpkin (C. pepo cv.Eskandarani) leaves. This increase led to increases of54.2% and 30.08% in H2O2 and MDA contents, respec-tively, compared with the control (Table 6).Regardless of inoculation, SA dramatically increased
H2O2 while resulting in decreased MDA contents inpumpkin. After virus inoculation, SA-treated plantsshowed decreases in MDA for all SA concentrations,which were lower than respective controls. The highlysignificant decrease of MDA was noticed for 100 mM(SA+V) and 100 mM (SA+V inoculated together) treat-ments.All (SA+V) treatments caused increases in H2O2
content. For instance, 100 mM (SA+V) increased of

ARTICLE IN PRESS
Fig. 1. Effect of ZYMV infection SA and (SA+V) treatments on leaf morphology of pumpkin (C. pepo cv. Eskandarani). ZYMV-infected old leaf (a)
showing severe mosaic and young leaf (b) showing chlorosis, green blisters, size reduction and malformation. (c–e) leaves treated with 10, 50 and 100 mM(SA+V) respectively, (f) sprayed with 100mM (SA+V ino). All SA treatments caused decrease in disease severity and the symptoms totally disappeared
following treatment with 100mM (SA+V). Control leaf (g) without any treatment and (h) treated with 100mMSA.
D.E.M. Radwan et al. / Physiological and Molecular Plant Pathology 69 (2006) 172–181176
H2O2 content by 36.13% in comparison with the control.In other words, H2O2 and MDA contents were SA-leveldependent.
Without virus inoculation, SA treatment increased H2O2
by 27.73% compared to controls, while MDA content
decreased by 11.81% compared to controls (Table 6). TheAOA of leaf extracts were calculated on the basis ofrelative amounts of MDA, which indicated lipid peroxida-tion, with respect to control MDA concentration. It wasfound that AOA varied greatly among the treatment levels.

ARTICLE IN PRESS
Table 3
Effect of ZYMV infection and SA treatments on photosynthetic rate (mmolm�2 s�1) and chlorophyll content (mg g�1 FW) of C. pepo (cv. Eskandarani)
leaves
Treatments Net photosynthesis rate (Pn) (mmolm�2 s�1) Chlorophyll content (mg g�1 FW)
M7SD % M7SD % a/b ratio
Control 17.48a70.94 100 0.60a70.08 100.00 2.30
Infected 8.30**d70.73 47.50 0.27**c70.01 45.00 2.75
10 mM SA+V 12.53**c72.81 71.67 0.35**c70.03 58.30 2.52
50 mM SA+V 15.18a,b,c72.59 86.84 0.48*b70.02 80.00 3.88
100mM SA +V 16.85a,b71.26 96.42 0.54a,b70.08 90.00 2.09
100mM SA +V ino 14.35*b,c72.60 82.12 0.60a70.01 100.00 3.00
100mM SA 16.55a,b70.35 94.71 0.62a70.01 103.33 2.79
Statistical significance of differences compared to control: *, significant at Po0.05; **, significant at Po0.01. Duncan’s composition between variants at
a ¼ 0.05. Variants with the same letters show the same reaction. The values are means (M) of eight replicates7standard deviation (SD).
Table 4
Effect of ZYMV infection and SA treatments on POD and APX activities
(unit g�1 FW) of C. pepo (cv. Eskandarani) leaves
Treatments POD activity
(unit g�1 FW)
APX (unit g�1 FW)
M7SD % M7SD %
Control 41.67g78.124 100 22.38b70.18 100
Infected 128.62**d711.10 308.69 47.07**a71.10 210.34
10 mM SA+V 202.19**a77.82 485.26 11.19b,c71.46 50.00
50 mM SA+V 161.51**b71.24 387.63 7.33*b,c70.27 32.76
100 mM SA +V 143.77**c712.02 345.04 4.63*c70.27 20.69
100 mM SA +V ino 74.50**e71.90 178.79 13.89b,c71.17 62.07
100 mM SA 58.91*f79.96 141.39 8.10*b,c70.50 36.21
Statistical significance of differences compared to control: *, significant at
Po0.05; **, significant at Po0.01. Duncan’s composition between
variants at a ¼ 0.05. Variants with the same letters show the same
reaction. The values are means (M) of four replicates7standard deviation
(SD).
Table 5
Effect of ZYMV infection and SA treatments on CAT and SOD activities
(unit g�1 FW) of C. pepo (cv. Eskandarani) leaves
Treatments CAT activity
(unit g�1 FW)
SOD activity
(unit g�1 FW)
M7SD % M7SD %
Control 40.00b,c70.49 100 35.56b,c70.04 100
Infected 79.00**a72.12 197.5 47.06a,b70.40 132.33
10 mM SA+V 45.25b70.04 113.1 31.66c77.84 89.027
50 mM SA+V 35.00b,c70.80 87.5 47.99a,b78.13 134.97
100 mM SA +V 30.25b,c71.73 75.63 50.91*a74.52 143.16
100 mM SA +V ino 25.00b,c71.72 62.5 49.01*a,b71.77 137.82
100 mM SA 18.50*c70.44 46.25 39.75 a,b,c78.02 111.78
Statistical significance of differences compared to control: *, significant at
Po0.05; **, significant at Po0.01. Duncan’s composition between
variants at a ¼ 0.05. Variants with the same letters show the same
reaction. The values are means (M) of four replicates7standard deviation
(SD).
Table 6
Effect of ZYMV infection and SA treatments on H2O2 concentration
(mmol g�1 FW) and MDA content (nmol g�1 FW) of C. pepo (cv.
Eskandarani) leaves
Treatments H2O2 (nmol g�1 FW) MDA
(mmol g�1 FW)
AOA
M7SD % M7SD (%)
Control 14.28d70.52 100 12.70b70.20
Infected 22.02**a71.17 147.99 16.52**a70.09 �30.08
10 mM SA+V 16.00*c70.62 114.37 12.79b70.61 �0.71
50 mM SA+V 17.20*c70.35 119.25 11.42*c71.01 10.08
100 mM SA +V 19.44**b70.51 132.32 9.56**d70.40 24.72
100 mM SA +V ino 20.13**a70.27 140.69 10.06**d70.47 20.79
100 mM SA 18.24**b70.64 128.86 11.20*c70.08 11.81
Statistical significance of differences compared to control: *, significant at
Po0.05; **, significant at Po0.01. Duncan’s composition between
variants at a ¼ 0.05. Variants with the same letters show the same
reaction. The values are means (M) of four replicates7standard deviation
(SD).
D.E.M. Radwan et al. / Physiological and Molecular Plant Pathology 69 (2006) 172–181 177
For example, ZYMV infection showed very low negativeAOA. Moreover, with the increase of SA treatments theAOA increased and this increase was SA concentration-
dependent. Interestingly, the highest increase in AOA wasobtained from leaves treated with 100 mM SA followed byZYMV inoculation.
3.5. Gas exchange
ZYMV infection and SA treatments strongly affectedgas exchange parameters, i.e. transpiration rate (E),intercellular CO2 concentration (Ci), stomatal conductance(Gs) and water use efficiency (WUE) (Figs. 2–5). Plantsinfected with ZYMV showed a significant reduction inboth transpiration rate (E) and stomatal conductance (Gs)in comparison with healthy plants. Furthermore, inter-cellular CO2 concentration (Ci) and water use efficiency(WUE ¼ Pn/E) seemed to be unaffected by ZYMVinfection.Exogenous SA application recovered the undesirable
effects of ZYMV infection. Thus, transpiration rates (E)and stomatal conductance (Gs) were almost similar tocontrol in all (SA+V) treatments. The pattern of increaseor decrease in E and Gs was inconsistent with increasing
ARTICLE IN PRESS
0
2
4
6
8
10
12
Control Infected 10 SA+V 50 SA+V 100 SA+V 100SA+V
ino
SA only
Treatments
E(m
mol m
-1S
-1)
Transpiration rate
aa
a aa
a
b**
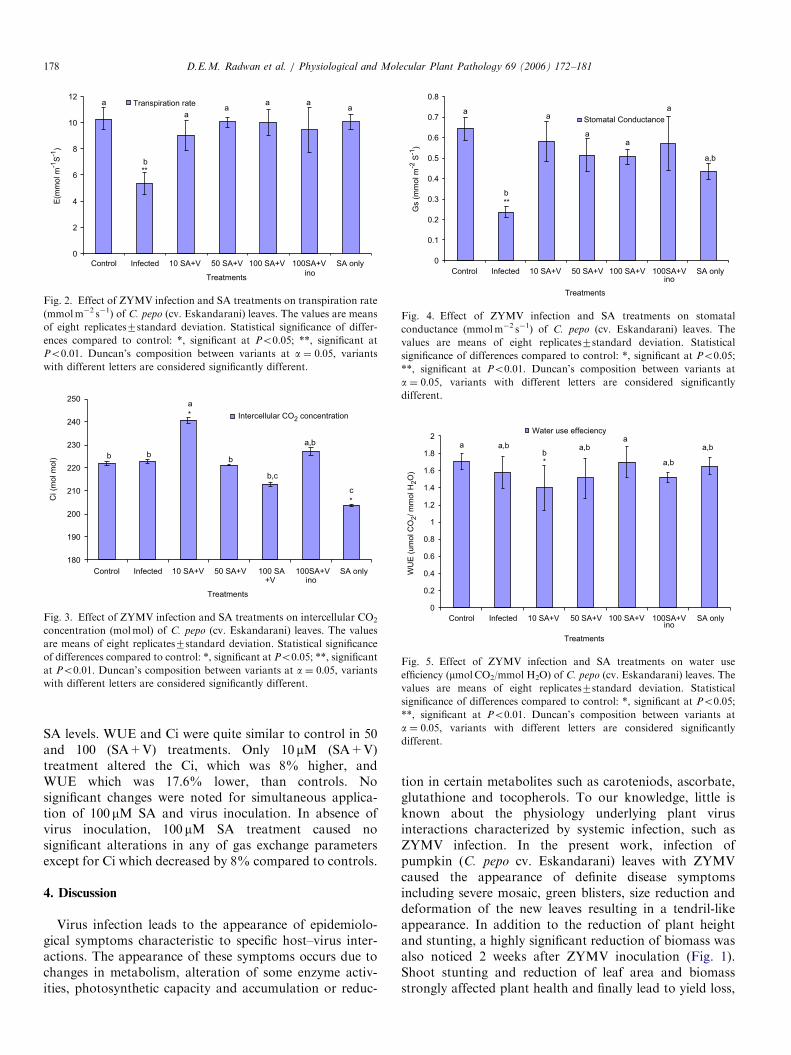
Fig. 2. Effect of ZYMV infection and SA treatments on transpiration rate
(mmolm�2 s�1) of C. pepo (cv. Eskandarani) leaves. The values are means
of eight replicates7standard deviation. Statistical significance of differ-
ences compared to control: *, significant at Po0.05; **, significant at
Po0.01. Duncan’s composition between variants at a ¼ 0:05, variants
with different letters are considered significantly different.
180
190
200
210
220
230
240
250
Control Infected 10 SA+V 50 SA+V 100 SA+V
100SA+Vino
SA only
Treatments
Ci (m
ol m
ol)
a
*
c
*
b,c
a,b
bbb
Intercellular CO2 concentration
Fig. 3. Effect of ZYMV infection and SA treatments on intercellular CO2
concentration (molmol) of C. pepo (cv. Eskandarani) leaves. The values
are means of eight replicates7standard deviation. Statistical significance
of differences compared to control: *, significant at Po0.05; **, significant
at Po0.01. Duncan’s composition between variants at a ¼ 0:05, variantswith different letters are considered significantly different.
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
Control Infected 10 SA+V 50 SA+V 100 SA+V 100SA+Vino
SA only
Treatments
Gs (
mm
ol m
-2 S
-1)
Stomatal Conductance
b
**
a,b
a
a
a
aa
Fig. 4. Effect of ZYMV infection and SA treatments on stomatal
conductance (mmolm�2 s�1) of C. pepo (cv. Eskandarani) leaves. The
values are means of eight replicates7standard deviation. Statistical
significance of differences compared to control: *, significant at Po0.05;
**, significant at Po0.01. Duncan’s composition between variants at
a ¼ 0:05, variants with different letters are considered significantly
different.
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
2
Control Infected 10 SA+V 50 SA+V 100 SA+V 100SA+Vino
SA only
Treatments
WU
E (
um
ol C
O2/ m
mol H
2O
)
a a,bb*
a,ba
a,b
a,b
Water use effeciency
Fig. 5. Effect of ZYMV infection and SA treatments on water use
efficiency (mmolCO2/mmol H2O) of C. pepo (cv. Eskandarani) leaves. The
values are means of eight replicates7standard deviation. Statistical
significance of differences compared to control: *, significant at Po0.05;
**, significant at Po0.01. Duncan’s composition between variants at
a ¼ 0:05, variants with different letters are considered significantly
different.
D.E.M. Radwan et al. / Physiological and Molecular Plant Pathology 69 (2006) 172–181178
SA levels. WUE and Ci were quite similar to control in 50and 100 (SA+V) treatments. Only 10 mM (SA+V)treatment altered the Ci, which was 8% higher, andWUE which was 17.6% lower, than controls. Nosignificant changes were noted for simultaneous applica-tion of 100 mM SA and virus inoculation. In absence ofvirus inoculation, 100 mM SA treatment caused nosignificant alterations in any of gas exchange parametersexcept for Ci which decreased by 8% compared to controls.
4. Discussion
Virus infection leads to the appearance of epidemiolo-gical symptoms characteristic to specific host–virus inter-actions. The appearance of these symptoms occurs due tochanges in metabolism, alteration of some enzyme activ-ities, photosynthetic capacity and accumulation or reduc-
tion in certain metabolites such as caroteniods, ascorbate,glutathione and tocopherols. To our knowledge, little isknown about the physiology underlying plant virusinteractions characterized by systemic infection, such asZYMV infection. In the present work, infection ofpumpkin (C. pepo cv. Eskandarani) leaves with ZYMVcaused the appearance of definite disease symptomsincluding severe mosaic, green blisters, size reduction anddeformation of the new leaves resulting in a tendril-likeappearance. In addition to the reduction of plant heightand stunting, a highly significant reduction of biomass wasalso noticed 2 weeks after ZYMV inoculation (Fig. 1).Shoot stunting and reduction of leaf area and biomassstrongly affected plant health and finally lead to yield loss,

ARTICLE IN PRESSD.E.M. Radwan et al. / Physiological and Molecular Plant Pathology 69 (2006) 172–181 179
although the leaf water content remained unaffected byZYMV infection. The observed morphological changeswere similar to those described previously with Styrianpumpkin [1] and other cucurbit plants infected withZYMV [38,39].
It is known that photosynthesis plays an important rolein plant growth and productivity. The net photosyntheticrate is positively associated with the crop yield in asparagusplants [9]. Any kind of stress, such as ZYMV disease,affecting photosynthesis rate can directly affect plantgrowth. In this experiment, the net photosynthesis rateand chlorophyll content under ZYMV stress were sig-nificantly reduced to reach half values of the control. Fromchlorophyll a/b ratios, it can be concluded that ZYMVinfection might affect chlorophyll molecules by degrada-tion or synthesis inhibition especially chlorophyll b whichwas more sensitive to disease stress. Wood [40] demon-strated that a decrease in net photosynthetic rate wasaccompanied by a decrease in chlorophyll contents invirus-infected plants. Photosynthetic properties and growthof infected plants have been shown negatively influenced byvirus infection [41]. In contrast, SA-treated plants exhibitedphotosynthesis rates and chlorophyll concentrations simi-lar to those of controls, especially with the concentration of100 mM SA with or without virus. This indicates that SAmight play a certain role in alleviating photosynthesisinhibition caused by virus infection. This role might bethrough conservation of chlorophyll molecules and pre-vention from further degradation during symptom devel-opment. In the present work, carotenoid content wasunaffected by SA application (data not shown). Thus SAmight have protected the carotenoid molecules fromdegradation.
The production of AOS in plant cells in response to virusinfection is well documented [32,42,43]. AOS metabolisminduces many changes in antioxidant enzyme systemswhich are still under discussion. Many reports describecontradictory results with both induction and inhibition ofantioxidant enzyme activities involved in free radicalscavenging [44–46]. We determined the activities of someantioxidant enzymes that decreased or increased inresponse to virus infection. Moreover, changes in theseantioxidant enzymes are known to be directly involved inthe activation of plant defense responses [32,42,46]. Thepresent results show that the activities of POD, APX, CATand SOD enzymes increased significantly (up to 2-fold orgreater) as a result of ZYMV infection. Our resultsconcerning POD and APX activities are in agreement withthose reported by Riedle-Bauer [43] who found a highlysignificant increase in POD, APX, CAT and SOD in bothCucumis sativus and C. pepo infected by ZYMV, whileHernandez et al. [32] reported no changes in antioxidantenzymes in peach plants infected with long-term plum poxvirus. Furthermore, in this work, exogenous application ofSA decreased the POD, APX and CAT activities butincreased SOD activity with increasing SA level. Suppres-sion of POD, APX and CAT activities and induction of
SOD reflects the role of SA in defense responses againstvirus infection. Formation of H2O2 in the cells duringdisease development is controlled by the activities ofantioxidant enzymes which are directly or indirectlycontrolled by hormones such as SA [45]. Because of thisphenomenon, it is convenient to analyze H2O2 level in bothZYMV-infected and SA-treated leaves. Interestingly, aninduction of H2O2 was recorded in our experiments underSA treatment and ZYMV infection. It is known that SAacts as electron donor for peroxidase isoforms, causesinhibition of the enzyme and leads to the production of SA-free radicals. Inhibition of peroxidase leads to enhancedH2O2 accumulation causing an alteration in the cellularredox state. So, application of SA induces at least one ofthe cellular protective mechanisms through potentiation ofan oxidative burst. The mode of SA was proposed toincrease H2O2 level by inhibition of CAT activity duringplant–pathogen interactions. Moreover, pretreatment withSA alone specifically inhibited CAT activity and increasedH2O2 level in rice roots. H2O2 accumulation affected by SAhas also been observed in other plant species [47,48].During pathogen attack in plants, AOS, including H2O2
accumulation, can act with at least one of three mechan-isms against pathogens. First, H2O2 can act directly bykilling the pathogen. Second, H2O2 can react withtransition metals leading to enhanced generation ofreactive hydroxyl radicals, the devastating effect on bio-molecules of which is well known. Third, H2O2 also hinderspenetration of plants by micro-organisms. It contributes tocell wall stiffening by facilitating peroxidase reactionscatalyzing intra- and inter-molecular cross-links betweenstructural components of cell walls and lignin polymeriza-tion. As a consequence, an increase in mechanical barriersslows down pathogen penetration allowing plant cells toarrange defenses which require more time to be activated.In addition, H2O2 can diffuse easily through biologicalmembranes, so it acts as intercellular signal which is able toactivate defense response against pathogen attack [42].Oxidative stress can be demonstrated by MDA concen-
tration which is considered a general indicator of lipidperoxidation [49]. Generally, high MDA contents weredetected in ZYMV-infected leaves while all SA treatmentscaused a noticeable reduction in MDA contents. Increasedlipid peroxidation can be observed both in plants withmosaic symptoms and in plants showing leaf-yellowing andmalformations. In other words, lipid peroxy-radicals areknown to oxidize several pigment molecules leading tobleaching of these pigments. This co-oxidation of pigmentmolecules might be responsible for yellowing of virus-invaded tissues. Kluge [50] reported that yellowing of virus-infected leaves of Beta vulgaris has been attributed to thisco-oxidation. Moreover, the present results suggest thatapplication of exogenous SA could increase the AOA ofleaf extracts under ZYMV infection.Many reports indicate that plant growth mainly depends
on photosynthesis capacity [9]. Stomatal closure and gasexchange parameters are defined as more or less controlling

ARTICLE IN PRESSD.E.M. Radwan et al. / Physiological and Molecular Plant Pathology 69 (2006) 172–181180
photosynthesis capacity. A noticeable reduction in E
(transpiration rate), stomatal number (data not shown)and Gs was observed in the present study in response toZYMV infection, agreeing with a previous report byRyslava et al. [41] in tobacco leaves. Although a reductionof Gs had been noticed in ZYMV-infected leaves, the Civalues remained unchanged. This may be due to decreasingphotosynthetic CO2 fixation or enhancing photorespirationwhich can keep CO2 concentration almost similar tocontrol while stomatal conductance is variable. To ourknowledge, there are few reports concerning gas exchangerelations in virus-infected plants. Guo et al. [26] reportedreductions in transpiration rate and stomatal conductancein Brassica juncea under turnip mosaic virus infection.Furthermore, we found that SA treatment could help torecover the reduction in transpiration rate occurring underZYMV infection, since SA reversed stomatal closure [7].The changes in photosynthesis after exogenous SAapplication under environmental stresses were due to eitherstomatal or non-stomatal limitations [51,52].
In conclusion, the following points can be drawn fromthe present investigation:
�
ZYMV infection negatively affected various physiologi-cal and metabolic processes of pumpkin leaves. Thesenegative effects could be reversed by exogenous SAapplication. � Plant health and normal growth, as well as the predictedyield of the plant, were affected by ZYMV infection,which in turn influenced the whole physiological net-work of pumpkin leaf cells.
� Because no apparent symptoms were noticed on theleaves, the 100mM SA concentration seemed to bethe most suitable concentration to alleviate growthand photosynthesis inhibition caused by ZYMV infection.
� The role of SA in inducing plant defense mechanismsagainst virus infection might be described through SAinterfering with the antioxidant systems. Such inter-ference occurred by either inhibition or activation ofsome antioxidant enzymes, reduction of lipid peroxida-tion or induction of H2O2 accumulation.
References
[1] Zechmann B, Muller M, Zellnig G. Cytological modifications in
zucchini yellow mosaic virus (ZYMV)-infected Styrian pumpkin
plants. Arch Virol 2003;148:1119–33.
[2] Provvidenti R. Inheritance of resistance to a strain of zucchini yellow
mosaic virus in cucumber. HortScience 1987;22:102–3.
[3] von Rad U, Mueller MJ, Durner J. Evaluation of natural and
synthetic stimulants of plant immunity by microarray technology.
New Phytol 2005;165:191–202.
[4] Maleck K, Levine A, Eulgem T, Morgan A, Schmid J, Lawton K, et
al. The transcriptome of Arabidopsis thaliana during systemic
acquired resistance. Nat Genet 2000;26:403–9.
[5] Cutt JR, Klessing DF. Salicylic acid in plants: a changing perspective.
Pharmaceut Technol 1992;16:25–34.
[6] Larque-Saavedra A. Stomatal closure in response to acetylsalicylic
acid treatment. Z Pflannzenphysiol 1979;93:371–5.
[7] Rai VK, Sharma SS, Sharma S. Reversal of ABA-induced stomatal
closure by phenolic compounds. J Exp Bot 1986;37:129–34.
[8] Barkosky RR, Einhellig FA. Effects of salicylic acid on plant water
relationship. J Chem Ecol 1993;19:237–47.
[9] Arfan M, Athar HR, Ashraf M. Does exogenous application of
salicylic acid through the rooting medium modulate growth and
photosynthetic capacity in two differently adapted spring wheat
cultivars under salt stress? J Plant Physiol 2006 [ahead of print].
[10] El-Tayeb MA. Response of barley grains to the interactive effect of
salinity and salicylic acid. Plant Growth Regul 2005;45:215–24.
[11] Gunes A, Inal A, Alpaslan M, Eraslan F, Bagci EG, Cicek N.
Salicylic acid induced changes on some physiological parameters
symptomatic for oxidative stress and mineral nutrition in maize (Zea
mays L.) grown under salinity. J Plant Physiol 2006 [ahead of print].
[12] Khan W, Prithiviraj B, Smith DL. Photosynthetic responses of corn
and soybean to foliar application of salicylates. J Plant Physiol
2003;160:485–92.
[13] Khodary SEA. Effect of salicylic acid on growth, photosynthesis and
carbohydrate metabolism in salt stressed maize plants. Int J Agric
Biol 2004;6:5–8.
[14] Ananievaa EA, Christova KN, Popova LP. Exogenerous treatment
with salicylic acid leads to increased antioxidant capacity in leaves
of barley plants exposed to Paraquat. J Plant Physiol 2004;161(3):
319–28.
[15] Borsani O, Valpuesta V, Botella MA. Evidence for a role of salicylic
acid in the oxidative damage generated by NaCl and osmotic stress in
Arabidopsis seedlings. Plant Physiol 2001;126:1024–30.
[16] Audenaert K, Pattery T, Cornelis P, Hofte M. Induction of systemic
resistance to Botrtis cinerea in tomato by Psedomonas aeruginosa
7NSK2: role of salicylic acid pyochelin, and pyocyanin. Mol Plant
Microbe Interact 2002;15:1147–56.
[17] Low PS, Merida JR. The oxidative burst in plant defense function
and signal transduction. Physiol Plant 1996;96:533–42.
[18] Milavec M, Ravnikar M, Kovac M. Peroxidases and photosynthetic
pigments in susceptible potato infected with potato virus YNTN.
Plant Physiol Biochem 2001;39:891–8.
[19] Mittler R. Oxidative stress, antioxidants and stress tolerance. Trends
Plant Sci 2002;7:405–10.
[20] Di Cagno R, Guidi L, De Gara L, Soldatini GF. Combined cadmium
and ozone treatments affects photosynthesis and ascorbate-depen-
dent defences in sunflower. New Phytol 2001;151:627–36.
[21] Noctor G, Foyer CH. Ascorbate and glutathione: keeping active
oxygen under control. Annu Rev Plant Physiol Plant Mol Biol
1998;49:249–79.
[22] Tommasi F, Paciolla C, De Pinto MC, De Gara L. A comparative
study of glutathione and ascorbate metabolism during germination of
Pinus pinea L seeds. J Exp Bot 2001;52:1647–54.
[23] Asada K. The water–water cycle in chloroplasts: scavenging of active
oxygens and dissipation of excess photons. Annu Rev Plant Physiol
Plant Mol Biol 1999;50:601–39.
[24] Smirnoff N. Ascorbic acid: metabolism and functions of a multi-
faceted molecule. Curr Opin Plant Biol 2000;3:229–35.
[25] Miteva E, Hristova D, Nenova V, Maneva S. Arsenic as a factor
affecting virus infection in tomato plants: changes in plant growth,
peroxidase activity and chloroplast pigments. Scientia Horticult
2005;105:343–58.
[26] Guo D, Guo Y, Zhao J, Liu H, Peng Y, Wang Q, et al.
Photosynthetic rate and chlorophyll fluorescence in leaves of stem
mustard (Brassica juncea var. tsatsai) after turnip mosaic virus
infection. Plant Sci 2005;168:57–63.
[27] Shalitin D, Wolf S. Cucumber mosaic virus infection affects sugar
transport in melon plants. Plant Physiol 2000;123:597–604.
[28] Metzner H, Rau H, Senger H. Untersuchuger Zue Synchronisierbar-
keit Einzelnar- pigment-mangel mutanten Von Chlorella. Planta
1965;65:186–94.
[29] Chandlee JM, Scandalios JG. Analysis of variants affecting the
catalase development program in maize scutellum. Theor Appl Genet
1984;69:71–7.

ARTICLE IN PRESSD.E.M. Radwan et al. / Physiological and Molecular Plant Pathology 69 (2006) 172–181 181
[30] Zhang XZ. In: Zhang XZ, editor. Research methodology of crop
physiology. Beijing Agricultural Press; 1992. p. 208–11.
[31] Nakano Y, Asada K. Hydrogen peroxide is scavenged by ascorbate-
specific peroxidase in spinach chloroplasts. Plant Cell Physiol
1981;22:867–80.
[32] Hernandez JA, Rubio M, Olmos E, Ros Barcelo A, Martinez-Gomez
P. Oxidative stress induced by long-term plum pox virus infection in
peach (Prunus persica). Physiol Plant 2004;122:486–95.
[33] Beauchamp C, Fridovich I. Superoxide dismutase: improved assays
and an assay applicable to acrylamide gels. Anal Biochem 1971;44:
276–87.
[34] Jana S, Choudhuri MA. Glycolate metabolism of three submerged
aquatic angiosperms during aging. Aquat Bot 1981;12:345–54.
[35] Arabshahi DS, Vishalakshi Devi D, Urooj A. Evaluation of
antioxidant activity of some plant extracts and their heat, pH and
storage stability. Food Chem 2007;100:1100–5.
[36] Duncan DB. A significance test for differences between ranked
treatments in analyses of variance. Virginia J Sci 1951;2:171–89.
[37] Sokal RR, Rohlf FJ. Biometry freeman and company. San Francisco;
1980. p. 321–64.
[38] Lisa V, Boccardo G, D’Agostino G, Dellavalle G, D’Aquilio M.
Characterization of a potyvirus that causes zucchini yellow mosaic.
Phytopathology 1981;71:667–72.
[39] Lisa V, Lecoq H. Zucchini yellow mosaic virus. CMI/AAB
description of plant viruses, No. 282. Kew, Surrey; 1984.
[40] Wood KR. Pathophysiological alternations. In: Mandahar CL,
editor. Plant viruses pathology, vol. 2. Boca Raton: CRC Press;
1990. p. 30–52.
[41] Ryslava H, Muller K, Semoradova S, Synkova H, Cerovska N.
Photosynthesis and activity of phosphoenolpyruvate carboxylase in
Nicotiana tabacum L. leaves infected by potato virus A and potato
virus Y. Photosynthetica 2003;41:357–63.
[42] De Gara L, Maria C. de Pinto, Tommasi F. The antioxidant systems
vis-a-vis reactive oxygen species during plant–pathogen interaction.
Plant Physiol Biochem 2003;41:863–70.
[43] Riedle-Bauer M. Role of reactive oxygen species and antioxidant
enzymes in systemic virus infections of plants. J Phytopathol 2000;
148:297–302.
[44] Baker CJ, Orlandi EW. Active oxygen in plant pathogenesis. Annu
Rev Phytopathol 1995;33:299–321.
[45] Clarke SF, Guy PL, Burritt DJ, Jameson PE. Changes in the
activities of antioxidant enzymes in response to virus infection and
hormone treatment. Physiol Plant 2002;114:157–64.
[46] Hernandez JA, Talavera JM, Martinez-Gomez P, Dicenta F, Sivilla
F. Response of antioxidant enzymes to plum pox virus in two apricot
cultivars. Physiol Plant 2001;111:313–21.
[47] Chen ZX, Silva H, Klessig DF. Active oxygen species in the induction
of plant systematic acquired resistance by salicylic acid. Science
1993;262:1883–6.
[48] Chen ZX, Lyer S, Caplan A, Klessig DF, Fan BF. Differential
accumulation of salicylic acid and salicylic acid-sensitive catalase in
different rice tissues. Plant Physiol 1997;114:193–201.
[49] Chaoui A, Mazhoudi S, Ghorbal MH, Ferjeni E. Cadmium and zinc
induction of lipid peroxidation and effects on antioxidant enzyme
activities in bean (Phaseoleus vulgaris L.). Plant Sci 1997;127:139–47.
[50] Kluge S. Viren und Viroide. In: Hoc B, Elstner EF, editors.
Schadwirkungen Auf Pflanzen—Lehrbuch der Pflanzentoxikologie.
Heidelberg: Spektrum Akademischer; 1995. p. 375–83.
[51] Athar HR, Ashraf M. Photosynthesis under drought stress. In:
Pessarakli M, editor. Handbook of photosynthesis. New York: CRC
Press, Taylor & Francis Group; 2005. p. 793–809.
[52] Dubey RS. Photosynthesis in plants under stressful conditions. In:
Pessarakli M, editor. Handbook of photosynthesis. 2nd ed. New
York: CRC Press, Taylor & Francis Group; 2005. p. 717–37.














